Embed Size (px)
Citation preview

De la epidemiologia molecular a la epidemiologia
genomica: Una nueva era para la investigacion de
enfermedades infecciosas en un contexto global.
V JORNADA SOBRE VIGILANCIA DE LA SALUD PUBLICA
INTEGRACION DE NUEVOS CONOCIMIENTOS Y
EXPERIENCIAS EN LA VIGILANCIA DE LA SALUD PUBLICA

Departament of Biology and Biochemistry
Universidad de Bath, Reino Unido



The Future of PulseNet: Faster, cheaper, and better foodborne disease detectionA new testCulture-independent diagnostic tests, or CIDTs, are new tests that can
detect the DNA of bacteria directly from patient samples like stool. As
their name implies, CIDTs do not need a culture (cells grown in a lab) to
identify the bacteria that caused a patient’s illness. Clinical laboratories
are increasing their use of CIDTs because they save costs and diagnose
illnesses faster.
The challenge to PulseNetFor PulseNet to work, scientists need genetic information drawn from
cultures. Because CIDTs skip the step of producing a culture, these
newer tests don’t produce DNA fingerprints to store in PulseNet’s
database. Without these data, public health scientists will not be able
to find, monitor, and prevent foodborne disease outbreaks or follow
trends to know if prevention policies are working. Many outbreaks
will go undetected or be detected later when the outbreak has grown,
and contaminated products will remain on the market longer .
Harmonizing the systemCDC is working closely with public health officials, diagnostic
laboratories, CIDT kit manufacturers, and clinicians to ensure that
PulseNet collects the DNA fingerprints it needs. PulseNet is also
developing a new way to detect outbreaks that doesn’t depend on
cultures. These innovations are in the early stages of research and
development, but we are heading toward a system that answers both
needs: rapid and less costly diagnosis for individuals and a way to
ensure essential data are still collected.
Transforming disease detection In 2013, CDC began using whole-genome sequencing (WGS)
to detect outbreaks caused by Listeria, the most deadly
foodborne pathogen. WGS reveals all the genetic material,
or the genome, of an organism (like bacteria and viruses) in
one efficient process. The Listeria project was one of the first
to receive support from CDC’s Advanced Molecular Detection
(AMD) initiative, a program that uses newer, more powerful,
pathogen detection technologies to find outbreaks sooner.
CDC is partnering with other federal agencies and state and
local health departments to analyze Listeria from human cases
and possible food sources.
Since the Listeria project began, scientists have detected
more clusters, solved outbreaks faster with fewer cases,
linked cases to likely food sources, and identified Listeria in
unexpected foods, such as caramel apples and ice cream. CDC
is quickly expanding the use of WGS in state laboratories and
has begun using WGS for investigations of other foodborne
pathogens such as Campylobacter, E. coli, and Salmonella.
WGS has been a game changer for outbreak detection,
spurring innovation and new discoveries. With WGS and
PulseNet working in tandem, we have taken a significant
leap forward in making food safer to eat.
CS263572A
100
90
80
70
60
50
40
30
20
10
0
Clusters detected
LISTERIA
Clusters detected
sooner or only by WGS
1419
21
6 60
Outbreaks solved
14
9
Cases linked to
food source
6
16
93
Num
ber of clu
sters
Detecting Listeria: Before and After WGS*
* Whole genome sequencing
Pre-WGS (Sept 2012 – Aug 2013)
WGS Year 1 (Sept 2013 – Aug 2014)
WGS Year 2 (Sept 2014 – Aug 2015)
years ofyears ofTM

Number of EU/EEA countries applying (green bar) or planning within three
years (hatched green bar) to apply WGS-based typing for (A) surveillance
application and (B) outbreak investigations, by pathogen target
ECDC roadmap for integration of molecular and genomic typing into European-level surveillance and epidemic preparedness
ECDC TECHNICAL REPORT
ECDC roadmap for integration of molecular and genomic typing into European-level surveillance and epidemic preparedness – Version 2.1, 2016–19




10Didelot et al. (2011) Recombination and Population Structure in Salmonella enterica. PLoS Genet 7(7)
Salmonella Typhimurium LT2 genome
microarray
genomes
MLST scheme




Técnicas clásicas
de typado
Genomas
completos

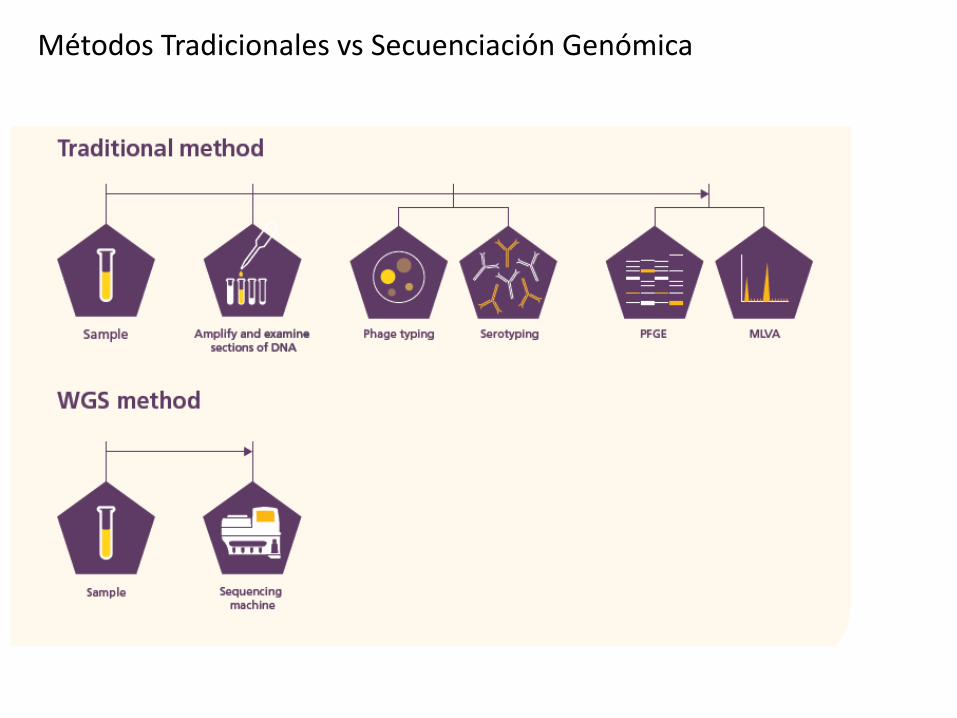
Métodos Tradicionales vs Secuenciación Genómica

Flujo de trabajo para el análisis de datos genómicos de organimos patógenos


WHY DO WE NEED TYPING?
Related isolates
Share the same strain
Unrelated isolates
Have a different strain
OUTBREAK STRAIN 1
STRAIN 2
STRAIN 3
STRAIN 4
Identification of epidemiologically linked isolates



WHY DO WE NEED TYPING?
Related isolates
Share the same strain
Unrelated isolates
Have a different strain
OUTBREAK STRAIN 1
STRAIN 2
STRAIN 3
STRAIN 4
Identification of epidemiologically linked isolates

OUTBREAK STRAIN 1 OUTBREAK STRAIN 2
Ancestro común???


Como varía el genoma de una bacteria????
Transducción
Fagos
Transformación
DNA ambienteConjugación
Plásmidos

Core genome
• Mutaciones
• Recombinación
Genoma accesorio
• Genes transmitidos
lateralmente
Filogenia
Adaptación local
Resistencia AB
Virulencia
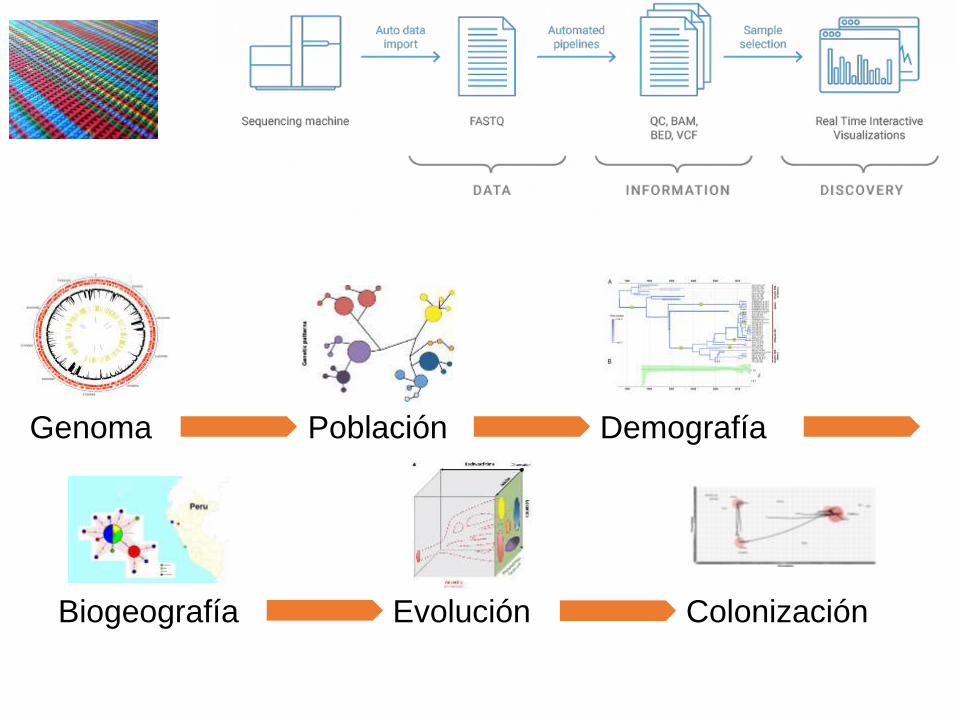
Genoma Población Demografía
Biogeografía Evolución Colonización

STRAIN 1 STRAIN 2
Ancestro común
Cuándo?
Dónde?
Fecha de emergencia de clones y variantes
En que lugar se origináron y vias de dispersión
Introducción? Vía de entrada del patógeno
Adaptado? Posibilidades de llegar a ser endémico
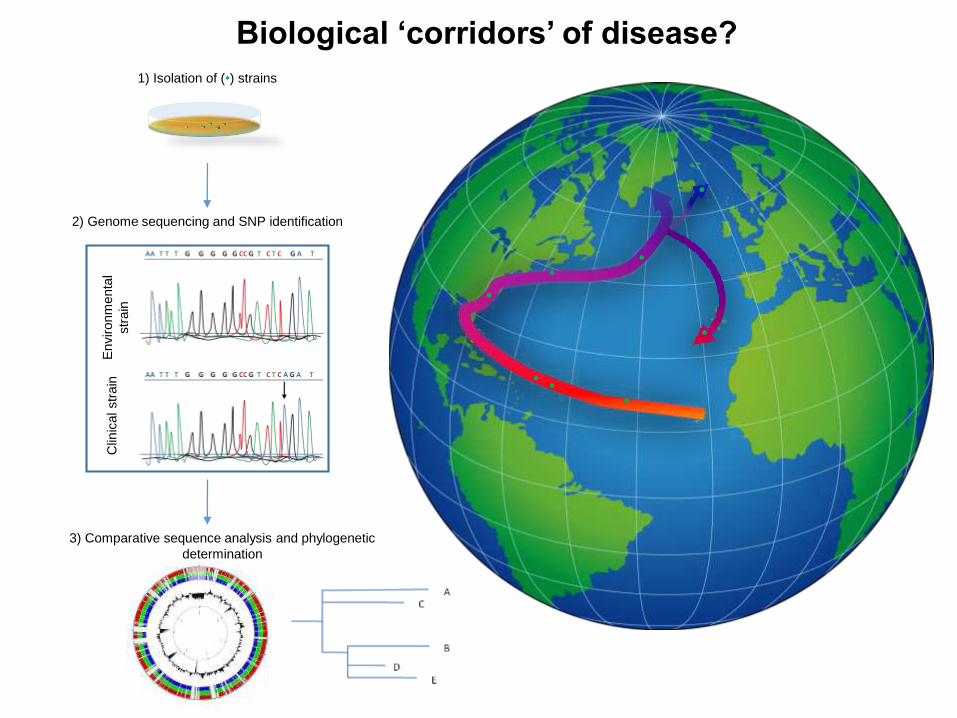
Biological ‘corridors’ of disease?
Environm
enta
l
str
ain
Clin
ical str
ain
2) Genome sequencing and SNP identification
3) Comparative sequence analysis and phylogenetic
determination
1) Isolation of (•) strains

List of software
• TRIMMINGtrimommatichttp://www.usadellab.org/cms/?page=trimmomatic
• MAPPINGBWAhttp://bio-bwa.sourceforge.net
• SNP CALLINGSAMtools (includes bcftools)http://samtools.sourceforge.netFreeBayeshttps://github.com/ekg/freebayesSnpEffhttp://snpeff.sourceforge.net/GATKhttps://www.broadinstitute.org/gatk/downloadArtemis + ACT + DNAPlotterhttp://www.sanger.ac.uk/resources/software/artemis/http://www.sanger.ac.uk/resources/software/act/#downloadshttp://www.sanger.ac.uk/resources/software/dnaplotter/
• Filtering SNPVCFToolshttp://vcftools.sourceforge.net/
• Visualization IGVhttp://www.broadinstitute.org/igv/BamViewhttp://bamview.sourceforge.net/CIRCOShttp://circos.ca/software/download/CGViewhttp://www.bioinformatics.org/cgview/
• PHYLOGENYPhyMLhttp://code.google.com/p/phyml/RAxMLhttp://sco.h-its.org/exelixis/web/software/raxml/index.htmlPUmPERhttps://github.com/fizquierdo/perpetually-updated-treesMrBayeshttp://mrbayes.sourceforge.net/download.php
• Molecular clockBEASThttp://beast.bio.ed.ac.uk/home#toc-2
• Population structureStructure (fine Structure)http://www.maths.bris.ac.uk/~madjl/finestructure/finestructure_info.html
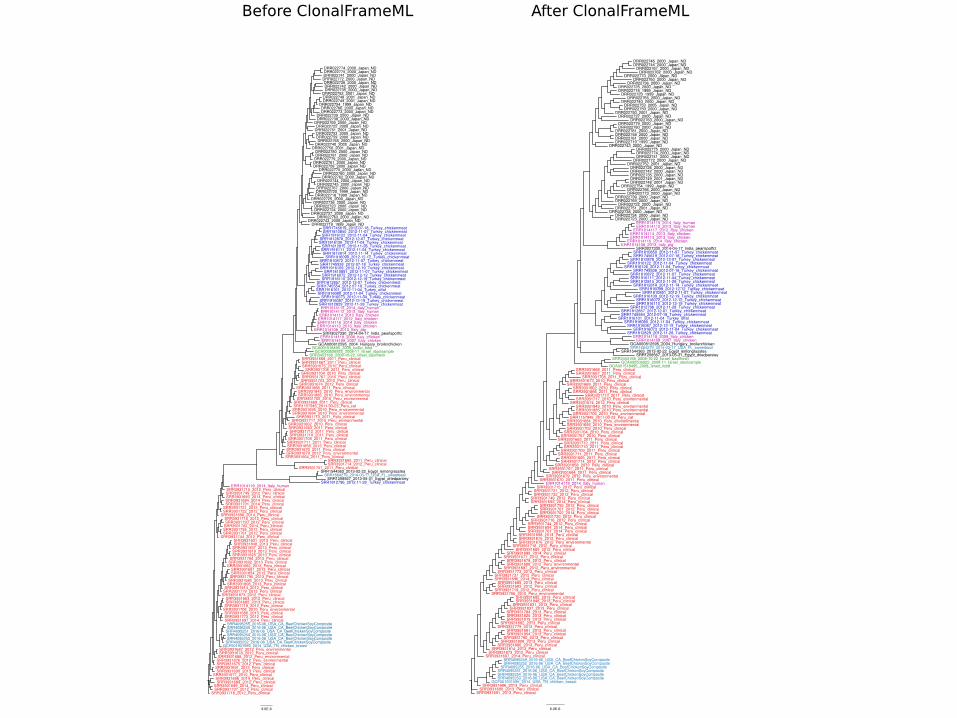
• ClonalOrigenMauvehttp://gel.ahabs.wisc.edu/mauve/Clonalframehttp://www.xavierdidelot.xtreemhost.com/clonalframe.htm

Raw sequences
Process sequences (QA, adapter removal)(Trimmomatic)
Genome assembly(A5 pipeline)
Genome alignment(Harvest tools)
Molecular clock analysis(Beast)
Recombination analysis(ClonalframeML)
Core SNPsCore genome
Phylogenetic analysis(RAxML)
Remove recombinant regions(cutseq)
Remove gaps(Trimal)
Extract SNPs in coding regions
(SnpEff)

….. más alla de secuenciación y bioinfomática




Significant global rates of movement identified by BSSVS.The color of the connections represent the relative strength by which the rates among these two locations are supported: white = weak, magenta = strong.
Richard Neher (University of Basel) Trevor
Bedford (Fred Hutchinson Cancer Research Center, Seattle)



Evolución y Diseminación Transcontinental
de Salmonella Infantis ST32
Salmonella
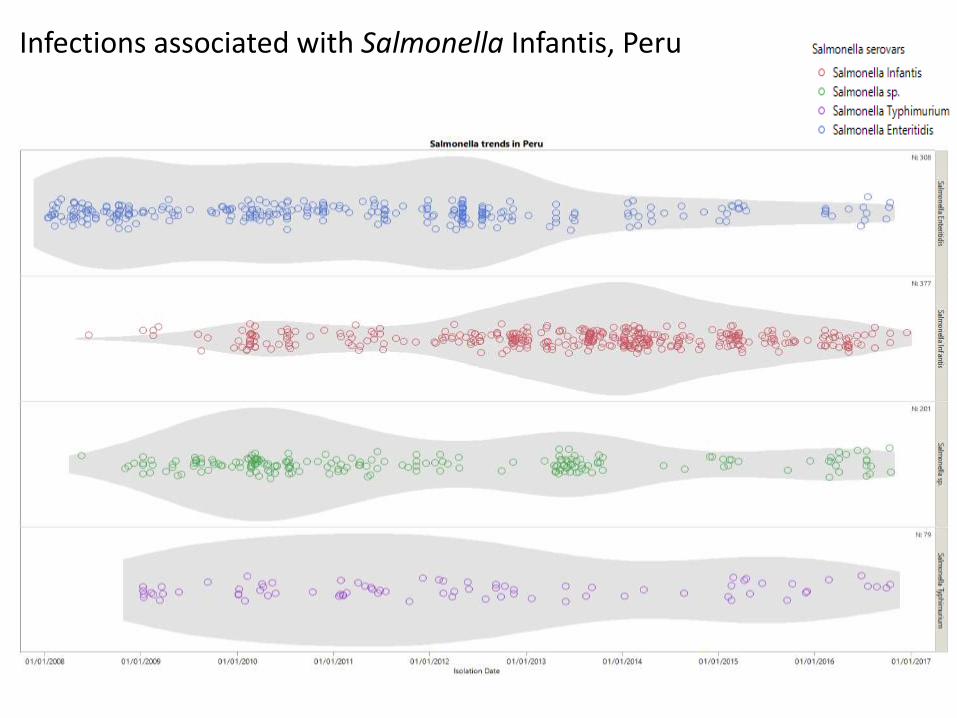
Infections associated with Salmonella Infantis, Peru

Strain Year ST Source AM C SXT CIP CTX NA N CAZ TE AMC
SRR3931661 20/4/2010 32 C S S S S S R R S R
SRR3931669 25/5/2011 32 C S S R S S R R S S
SRR3931684 8/3/2010 32 E S S R S S R R S R
SRR3931695 8/3/2010 32 E S S R S S R R S R
SRR3931858 3/1/2013 32 C R S R S S R R S R S
SRR3931714 23/4/2012 32 C R R R S R R R R R R
SRR3931679 20/2/2012 32 E S S R S S R R S R S
SRR3931664 7/5/2011 32 C S S R S S R S R S
SRR3931707 4/6/2011 32 C R S S R R R R R S
SRR3931689 12/9/2012 32 C R R R S R R R R R S
SRR3931677 1/10/2012 32 E R R R S R R R R R R
SRR3931690 10/4/2013 32 C R R R S R R R S R S
SRR3931691 10/7/2013 32 C R R R S R R R R R S
SRR3931681 29/8/2013 32 C R R R S R R R S R S
SRR3931680 27/8/2013 32 C R R R S R R R S R S
SRR3931692 10/8/2013 32 C R R R S R R R S R S
SRR3931682 27/8/2013 32 C R R R S R R R S R S
SRR3931685 10/5/2013 32 C R R R S R R R R R S
SRR3931683 10/12/2012 32 C R R R S R R R R R S
SRR3931678 8/11/2012 32 C R R R S R R R R R R
SRR3931687 19/12/2012 32 E R R R S R R R R S
SRR3931688 19/12/2012 32 E R R R S R R R R R S
SRR3931676 4/6/2012 32 E R R R S R R R R R S
SRR3931693 26/7/2014 32 C R R R S R R R R R S
SRR3931694 10/4/2014 32 C R R R S R R R R R S
SRR3931686 23/5/2013 32 C R R R S R R R R R S
ANTIBIOTICS
AM: ampicillin, C: chloroamphenicol, SXT: trimethoprim/sulfamethoxazole, CIP: ciprofloxacin, CTX: cefotaxime, NA: nalidixicacid, N: neomycin, CAZ: ceftazidime, TE: tetracycline, AMC: amoxicillin & clavulanic acid. Strains: blue is clade 1, green isclade 2 and pink clade 3 as per the phylogenetic trees.
Antibiotic resistance profile for all Peru genomes
• All Peru strains were resistant to: nalidixic acid, neomycin and tetracycline.
• All of clade 3 was resistant to: ampicillin, chloramphenicol, cefotaxime and trimethoprim/sulfamethoxazole
• SRR3931714, SRR3931677 and SRR3931678 were resistant to: all antibiotics except ciprofloxacin which was the only antibiotic all strains were sensitive to.
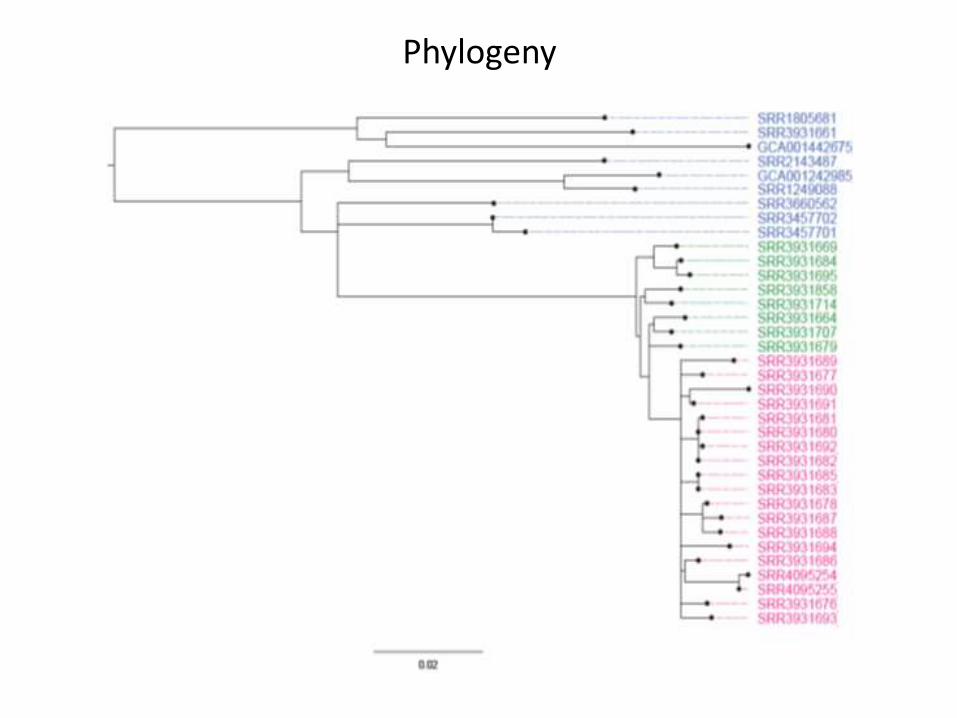
Phylogeny

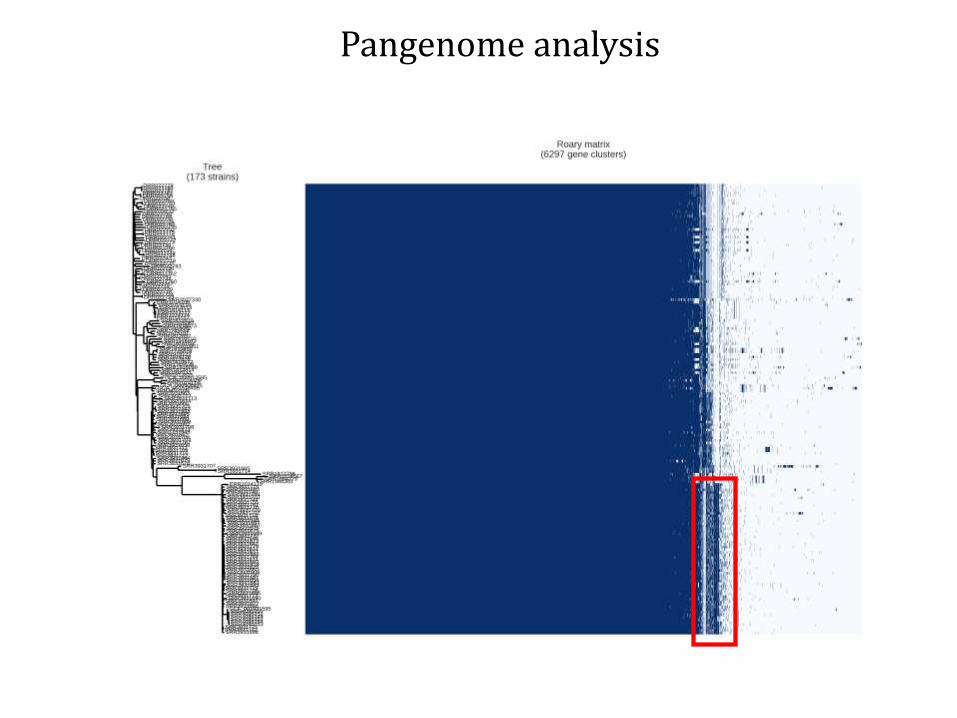
Pangenome analysis

Pangenome analysis

CVM44454_2014-05-09_USA-MA_clinical --> 316160 bpFSIS1502169 2015 USA-NC young chicken --> 323122 bpFSIS1502916_2015_USA-NJ_comminuted_chicken --> 322518 bpN55391_2014_USA-TN_chicken_breast --> 316814 bpSRR3931690_2013_Peru_clinical <-- 323100 bp (in silico)pC271 E. coli aislada en Bolivia el 2011 <-- 125315 bp
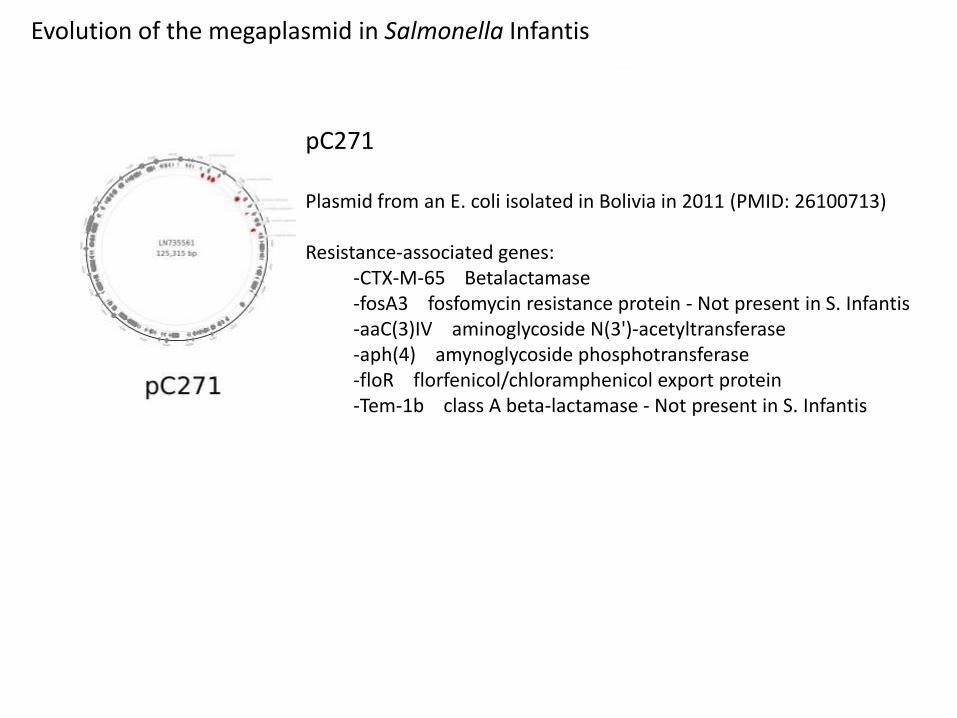
pC271
Plasmid from an E. coli isolated in Bolivia in 2011 (PMID: 26100713)
Resistance-associated genes:-CTX-M-65 Betalactamase-fosA3 fosfomycin resistance protein - Not present in S. Infantis-aaC(3)IV aminoglycoside N(3')-acetyltransferase -aph(4) amynoglycoside phosphotransferase -floR florfenicol/chloramphenicol export protein -Tem-1b class A beta-lactamase - Not present in S. Infantis
Evolution of the megaplasmid in Salmonella Infantis

Evolution of the (mega)plasmid in Salmonella Infantis
pFSIS1502169 (323122 bp, 296 CDS)
(125315 bp, 170 CDS)
• Both plasmids have 99% of sequence similirarity
• Share 116597/125315 bp (93,04%).
• 14 CDS exclusive in pC271 and 6 CDS -in the shared region- only pFSIS1502169.
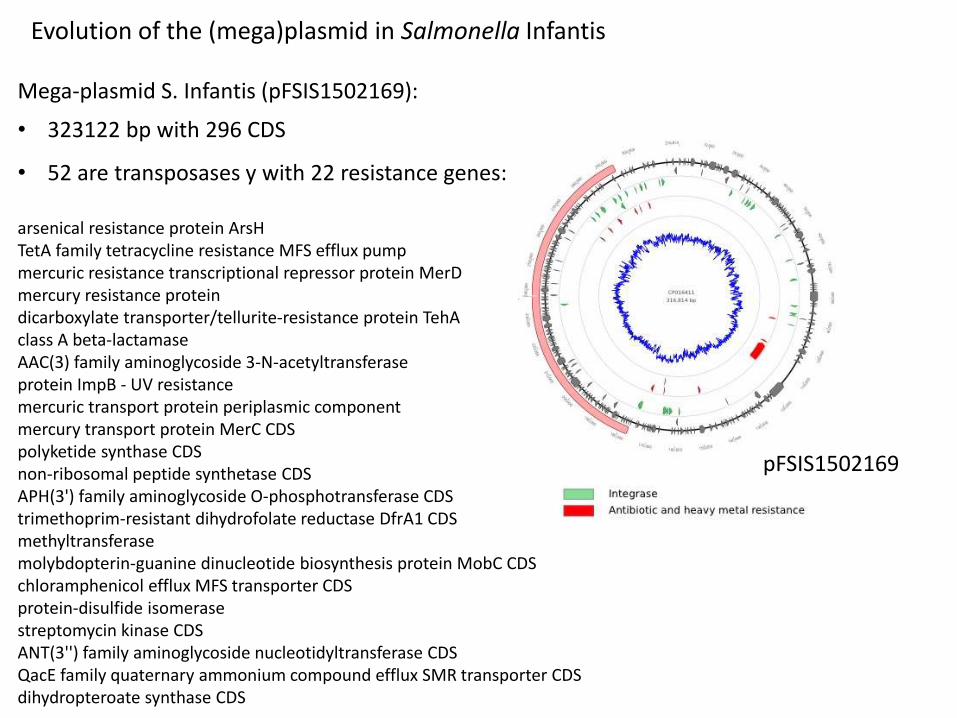
Evolution of the (mega)plasmid in Salmonella Infantis
Mega-plasmid S. Infantis (pFSIS1502169):
• 323122 bp with 296 CDS
• 52 are transposases y with 22 resistance genes:
arsenical resistance protein ArsHTetA family tetracycline resistance MFS efflux pump mercuric resistance transcriptional repressor protein MerDmercury resistance protein dicarboxylate transporter/tellurite-resistance protein TehAclass A beta-lactamase AAC(3) family aminoglycoside 3-N-acetyltransferase protein ImpB - UV resistance mercuric transport protein periplasmic component mercury transport protein MerC CDS polyketide synthase CDS non-ribosomal peptide synthetase CDS APH(3') family aminoglycoside O-phosphotransferase CDS trimethoprim-resistant dihydrofolate reductase DfrA1 CDS methyltransferase molybdopterin-guanine dinucleotide biosynthesis protein MobC CDS chloramphenicol efflux MFS transporter CDS protein-disulfide isomerase streptomycin kinase CDS ANT(3'') family aminoglycoside nucleotidyltransferase CDS QacE family quaternary ammonium compound efflux SMR transporter CDS dihydropteroate synthase CDS
pFSIS1502169
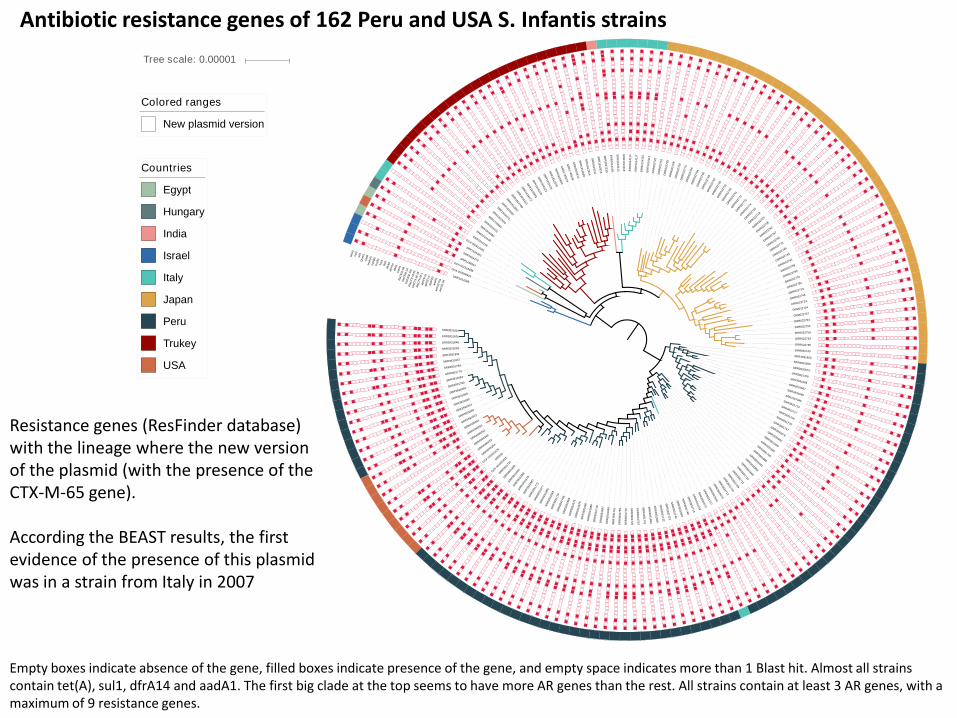
Empty boxes indicate absence of the gene, filled boxes indicate presence of the gene, and empty space indicates more than 1 Blast hit. Almost all strains contain tet(A), sul1, dfrA14 and aadA1. The first big clade at the top seems to have more AR genes than the rest. All strains contain at least 3 AR genes, with a maximum of 9 resistance genes.
Antibiotic resistance genes of 162 Peru and USA S. Infantis strains
SRR3931708
SRR3931680
SRR3931779
SR
R3
93
16
94
SRR1916101
DRR022744
SRR3931819
DRR022767
SR
R181
08
72
SR
R39
31
69
7
SRR3931790
DRR02
2720
SR
R393168
9
DRR022736
SRR3931665
SR
R3
93
16
75
DR
R0
22
72
2
ER
R1
01
411
7
DR
R02
273
5
SR
R3
93
17
20
SR
R3
931
69
6
SRR3931667
SRR3931695
SRR3931668
SRR4095252
SR
R1916122
SRR1157945
DR
R02
27
51
DR
R0
22
739
DR
R02
2741
N553
91
GC
F 001931575
DRR022
718
SR
R3931670
SR
R1812857
SRR3931692
DRR022763
DRR022719
SR
R18
10851
DR
R0
22
738
SR
R3
931
693
DR
R022752
DRR022765
SR
R1
81
28
15
DRR
022725
DRR022780
SR
R3931719
DR
R0
22
76
9
ER
R1
01
41
08
SRR4095249
SRR3931858
SR
R393
1676
SRR39
3173
7
ER
R1
01
411
4
SR
R3931707
DRR022737
SR
R1916126
ER
R1
01
411
6
SR
R3931711
GCA 000812595
SRR3931669
SR
R3
93
172
1
SRR3931684
SRR3931713
SR
R1810850
SR
R1812878
SRR4095253
DR
R0
22773
SR
R3
93
169
8
SR
R19
16110
DRR022781
SR
R393
1710
DRR022779
SRR3931681
DR
R022
726
SRR4095254
SR
R3931685
SRR3931662
DRR022723
SR
R19
16
111
SRR3931666
SR
R3
93
17
02
ERR1014109
SRR3931682
DRR022759
SR
R1745619
SR
R393
1773
DR
R022754
SR
R19
16109
SRR3931814
SRR1745554
SR
R1
745538
SRR3453168
SR
R3931664
DRR022745
SRR
3931885
DRR022740
SR
R39
3168
8
SR
R1
81
281
4
DR
R022749
SRR1812829
SR
R39
31
722
SRR4095255
DRR022761
DRR022770
GCA 001518495
SRR1916087
SRR3931843
GCF 0
01931
615
SRR3931837
SRR1544363
SRR3931717
SRR1664279
DR
R022766
SRR3931848
SR
R39
3169
0
SR
R39
3169
1
ER
R1014119
SR
R3
93
16
99
SR
R39
3171
2
SR
R3
02
73
30
SRR3931873
SRR3931673
SR
R39
316
78
SR
R3931683
SR
R39
316
86
DR
R02
2772
SRR3931674
SR
R3
93
17
55
DRR022753
ER
R1
01
411
5
SR
R3
93
17
01
DRR022750
DRR022758
SRR3931854
DRR
0227
74
SRR1916080
SRR3931703
SR
R3931715
SR
R3931677
SRR3931663
SR
R39
316
87
SRR1916073
SRR3931784
DRR022762
SR
R3
931749
SR
R3
931
71
8
SRR1258567
GCA 000506925
SRR3931825
DR
R02
2742
SR
R19
16072
ERR1014118
SRR
3931714
SR
R1
91
60
99
SR
R3931706
SR
R3
93
17
16
SRR3931808
SRR3931705
SRR3931704
DRR022760
ER
R1
01
411
2
DRR022724
ER
R1
01
411
3
SR
R3
93
17
44
SRR3931767
SR
R393
1709
SRR3931831
DR
R022748
SR
R3
93
17
61
DRR022743
SR
R1812796
SRR4095251
DR
R02
2775
SR
R393
167
9
aac(3
)-IId
aac(
3)-
IVa
aadA
1
aadA
22
aph(3
')-I
c
aph
(4)-
Ia
bla
CT
X-M
-1
bla
CT
X-M
-15
bla
CT
X-M
-65
bla
TE
M-1
16
bla
TE
M-1
A
bla
TE
M-1
B
cm
lA1
dfr
A1
dfr
A14
dfr
A8
floR
fosA
lnu(B
)
mef(
B)
mp
h(A
)
oqxB
QnrS
1
sul1
sul3
tet(
A)
Colored ranges
New plasmid version
Countries
Egypt
Hungary
India
Israel
Italy
Japan
Peru
Trukey
USA
Tree scale: 0.00001
Resistance genes (ResFinder database) with the lineage where the new version of the plasmid (with the presence of the CTX-M-65 gene).
According the BEAST results, the first evidence of the presence of this plasmid was in a strain from Italy in 2007

Molecular Clock

Phylogeography of S. Infantis ST32 1999 to 2016

Dr. Narjol Gonzalez-Escalona
Dr. Joaquin Triñanes
Dr. Ronnie Gavilán
Rebecca Girton
Michel Abanto
Dr Cristóbal ChaidezDr Juan Ramón Ibarra