Embed Size (px)
Citation preview

Gene. 42 (1986) 225-229
Elsevier
225
GENE 1557
Genomic organization and nucleotide sequences of two corn histone H4 genes *
(Recombinant DNA; gene library; 2 vector; Zea ways)
Gabriel Philipps, Nicole Chaubet, Marie-Edith Chaboute, Martine Ehling and Claude Gigot **
Luboratoire de Virologie, lnstitut de Biologie Mokculaire et Cellulaire du CNRS, 15 rue Descartes, 67084 Strasbourg CPdex
(France) Tel. 88.61.02.02
(Received November 18th, 1985)
(Revision received January 8th, 1986)
(Accepted January lOth, 1986)
____
SUMMARY
The sea urchin histone H4 gene has been used as a probe to clone two corn histone H4 genes from a
1gt WES . J.B corngenomic library. The nucleotide (nt) sequences of both genes showed that the encoded amino
acid sequences were identical to that of the H4 of pea and one variant of wheat. The nt sequences of the coding
regions showed 92% homology. 5’- and 3’-flanking regions do not show extensive nt sequence analogies.
Southern blotting of the EcoRI digested genomic DNA suggests the existence of multiple H4 genes dispersed
throughout the genome.
INTRODUCTION
The genes encoding histones have been extensive-
ly studied in a wide variety of organisms (reviews by
Hentschel and Birnstiel, 1981; Maxson et al., 1983).
* On request, the authors will supply a detailed experimental
evidence for the conclusions reached in this short presentation.
** To whom all correspondence and reprint requests should be
addressed.
Abbreviations: aa, amino acid(s); bp, base pair(s); H41C7-
In the plant kingdom the structures and genomic
organizations of the genes encoding the histones H3
(Tabata et al., 1984) and two variants of H4 (Tabata
et al., 1983; Tabata and Iwabuchi, 1984) have re-
cently been analysed in wheat. It was shown that in
this plant the majority of the multiple copies of these
genes (80 to 100 and 100 to 125 copies of H3 and H4,
respectively) are dispersed in the genome. On the
other hand, it was reported that in rice at least one
set of the genes encoding H2A, H2B and H4 were
located on a cloned 6.64-kb DNA fragment (Tho-
mas and Padayatty, 1983). In higher plants no other
histone genes have been studied UD to now. H41C14, corn genomic DNA fragments cloned in IgtWES. IB
L
DNA; H4pUC7 and H4pUCl4, DNA fragments of H41C7 and In this communication we report the molecular
H41C14, respectively, containing the coding regions subcloned cloning and nt sequences of the coding regions and
in pUC9; H4C7-H4C14, sequenced regions of H4pLJC7 and the adjacent 5’- and 3’-flanking regions of two corn
H4pUC14, respectively; nt, nucleotide(s). histone H4 genes.
0378-l I19/86/$03.50 0 1986 Elsewer Science Publishers l&V. (Biomedical Division)
226
kXPERIMENT.4L.
(a) Cloning and sequencing
A Southern blot of EcoRI-digested corn (Zea
map, INRA 258) genomic DNA, purified by a tech-
nique derived from that of Zimmerman and Gold-
berg (1977) was hybridized with the “P-labelled
coding region of the sea urchin Psammechinus miliar-
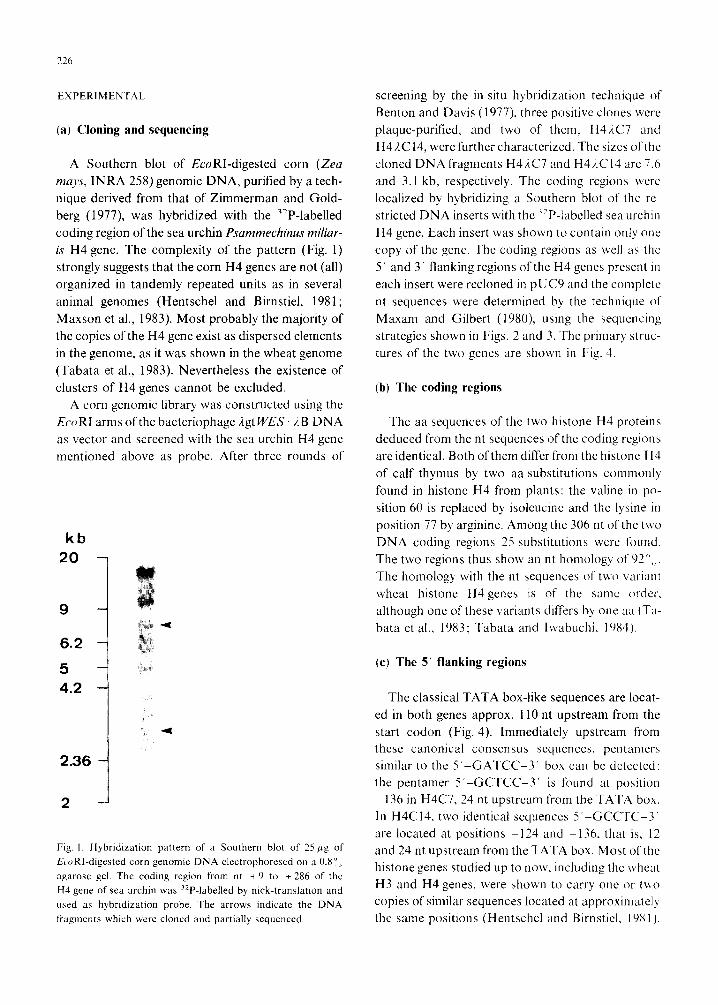
is H4 gene. The complexity of the pattern (Fig. 1)
strongly suggests that the corn H4 genes are not (all)
organized in tandemly repeated units as in several
animal genomes (Hentschel and Birnstiel, 1981;
Maxson et al., 1983). Most probably the majority of
the copies of the H4 gene exist as dispersed elements
in the genome, as it was shown in the wheat genome
(Tabata et al., 1983). Nevertheless the existence of
clusters of H4 genes cannot be excluded.
A corn genomic library was constructed using the
EcoRI arms ofthe bacteriophage Jgt WES iB DNA
as vector and screened with the sea urchin H4 gene
mentioned above as probe. After three rounds of
kb
20 -
9
6.2 -
5
4.2 -
2.36 -
2
Fig. I Hybridization pattern of a Southern blot of 25 1~9 of
&oRI-digested corn genomic DNA electrophoresed on a 0.8”,,
agarose gel. The coding region from nt + 9 to + 286 of the
H4 gene of sea urchin was “P-labelled by nick-translation and
used as hybridization probe. The arrows indicate the DNA
fragments which were cloned and partially sequenced.
screening by the in situ hybridization technique of
Benton and Davis (1977). three positive clones were
plaque-purified, and two of them, H4iLC7 and
H41.C 14, were further characterized. The sizes of the
cloned DNA fragments H4iC7 and H4iC 14 arc 7.6
and 3.1 kb, respectively. The coding regions were
localized by hybridizing a Southern blot of the rc-
stricted DNA inserts with the “P-labelled sea urchin
H4 gene. Each insert was shown to contain only one
copy of the gene. The coding regions as ucll as the
5’ and 3’ flanking regions of the H4 genes present in
each insert were recloned in pUC9 and the complctc
nt sequences were determined by the technique of
Maxam and Gilbert (1980) using the sequencing
strategies shown in Figs. 2 and 3. The primary struc-
tures of the tuo genes are shown in Fig. 4.
(b) The coding regions
The aa sequences of the two histone H4 proteins
deduced from the nt sequences of the coding regions
are identical. Both ofthem differ from the histone H4
of calf thymus by two aa substitutions commonly
found in histone H4 from plants: the valine in po-
sition 60 is replaced by isoleucine and the lysine in
position 77 by arginine. Among the 306 nt of the two
DNA coding regions 25 substitutions were found.
The two regions thus show an nt homology of 92”,,
The homology with the nt sequences of two variant
wheat histonc H4 genes is of the satnc order.
although one of these variants differs by one aa (Ta-
bata et al., 1983; Tabata and Iwabuchi, 1984).
(c) The 5’ flanking regions
The classical TATA box-like sequences are locat-
ed in both genes approx. 110 nt upstream from the
start codon (Fig. 4). Immediately upstream from
these canonical consensus sequences, pentamers
similar to the 5’-GATCC-3’ box can be detected:
the pentamer 5’-GCTCC-3’ is found at position
-136 in H4C7, 24 nt upstream from the TATA box.
In H4C14. two identical sequences 5’-GCCTC-3’
are located at positions -174 and - 136, that is, 12
and 24 nt upstream from the TATA box. Most of the
histone genes studied up to now, including the wheat
H3 and H4 genes, were shown to carry one or t\co
copies of similar sequences located at approximately
the same positions (Hentschcl and Birnsticl, 19X 1).

221
1 Kb _. ES A
Act I I I I I _
Ava I II b Bgl II I
Dde I I I I I I
Fig. 2. Restriction maps and sequencing strategy of the cloned genomic DNA fragment H42C7. (a) Restriction map of H41C7. The
2.0-kb fragment EcoRI-PsrI was further recloned in pUC9. A, AvaI; B, BarnHI; Bg, BglII; E, EcoRI; H, HindIII; P, PstI; S, SalI. The
black boxes indicate the position of the H4 coding region, and the arrow head indicates the 5’-3’ orientation. (b) Restriction maps of
H4pUC7. Only those restriction sites used in the sequencing experiments (after 32P 5’-end labelling) are shown. (c) Sequencing strategy.
The arrows indicate the direction of sequencing.
No typical CCAAT sequences are present. A
5’-CCATC-3’ motif was found in H4C7 at position
-164 and twice in H4Cl4 at nt -144 and -192.
Whether or not these sequences play the role of a
typical “CCAAT-box” remains to be determined.
Sequence homologies between the 5’ flanking
regions of the two genes are restricted to three small
stretches located between the “TATA-box” and the
initiation codon (see Fig. 4).
-- - .4 t_
c- . --
c- )___f
Fig. 3. Restriction maps and sequencing strategy of the cloned
genomic DNA fragment H41C14. (see Fig. 2 for details and
symbols). (a) Restriction map of H42C14. The 1.2-kb fragment
P.~rl-Sal1 was further recloned in pUC9; (b) restriction maps of
H4pUC14; (c) sequencing strategy.
(d) The 3’ flanking regions
No significant sequence homologies were found in
this region. In most of the animal histone genes there
are two sequences involved in the 3’ RNA process-
ing: a G + C-rich T-hyphenated dyad symmetry mo-
tif followed by a FAAAGA “core” (Birnstiel et al.,
1985). In corn H4 as well as in the wheat H3 and
H4 genes this typical inverted repeat is absent.
Nevertheless, in both corn H4 genes there exist
inverted repeats approx. 20 nt after the terminal
codon. These structures are followed by short
A + G-rich sequences: AGAAG at nt + 39 in H4C7
and AGGGGA at nt + 33 in H4C14.
ACKNOWLEDGEMENTS
We wish to thank Prof. L. Hirth for his constant
interest in this work. We also thank
Prof. M.L. Birnstiel for the gift of the plasmid
pHae 18 1 containing the coding region of the histone
H4 gene of sea urchin.

228
H4C7 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
-350
. . . . . . . . . . . . . . . . ..*.................. . . . . . . . ..GAATfCGAAGTITC'BGCCCAGCGGTCCACCC
-300 -250
CCRAA~~CACGCG~~CACACRGCA~~CCGCACCAC~ACCGCC~C~C~~CC~CG&G~CC ~AGTCrC~CGGTCG~~TCGACACACGTCCGTCGGTCCCCTCC
ACGCGGAtiGTWCCCATkACG~C:ACTTCG"c?"CT AACGGCGCAGAGTCGCAGACTGGCI'CCGCACCCCACCACACGCCG
-50
ATCCAATCCI;TCAGTACCCAATCTf TACACCAG~G~~nrcCG~A~TC~CCGCG C!TCGTCTCATCTCTKCATACGACC CCCTACCTCC~~CCCRATi’GTC~C~~C~CAGAG~CGAG~ACC
+50
An: TCT GGG CCC GGC AAG G&C GGC AAG GG;. CTG GGC AAG 6GC GGC GCG AiG CCC CAC CC;; I.. ..G . . . . . . ..A . . . . . . ..G .._ ..G . . . . . . . . . 1.. . . . . . . . . . . . T . . . ..C Met Ser Gly Arg Gly Lys Gly Gly Lys Cly Leu Gly Lys Gly Gly Ala LYS Arq His Arg
AAG GTG CiX CCC GAC RAC A+C CAG GGC AT;‘ ACC AAG CCC ;;CG ATC CGG AkG CTG GCf A& c .., . . . .‘. ,.. . . . . . . . . . . . T ..G . . . ._. . . . . . . . . . . . . .._ ..G ..A . . . . .
Lys Val Leu Arq Asp Asn lie Gin Gly Ile Thr Lys Pro Ala Ile Arg Arg Leu Ala Arg
+150
ACG GGT GGC kG AAG CGC A+C TCG GGG CTC ATC TAC GAG CAC ACC CGC GGC GTG CTC AA; . . . . . c *.. . . . . . . ,.. . . . . . . .,. I.. . . . . . . .., ..* . . . . . . . . . . . . . . . Arg Gly Gly Val Lys Arg Xle Ser Gly Leu Ile Tyr Glu Glu Thr Arg Gly Val Leu Ly+
+200
An: 'ITT CTC ;;AG AAC GTC ATC CGC GAC GCC GTC ACC TAC k(I GAG CAC CCC CGC CCC AA& . . . ..C . . . . . . ..T . . . ..T . . . .__ ..T 1.. .._ _.. ..G . . . . . . ..C . . . ..T . . . Ile Phe Leu Glu Am Val Ile Arg Asp Ala Val Thr Tyr 'Rx Glu His Ala Arg Arg Lys
+zso +300
ACC GTG ACC CCC AT(; GAC G+C GTC TAC CCC m MG CGC GAG xc cGc AEC mc TAC G& . . . ..c . . . . . . . . . . . . . . . ,.. . . . . . . . . . . . . . . G . . . . . . . . . . . . . . . . . . ..A Thr Val Thr Ale Met Asp Val Val Tyr Ala Leu Lys Ar9 Gln Gly Arg Thr Leu Tyr Gly
+350
TIC GGA GGG TAGGCTAGGCGCC~~T~CAT~G~~~AG~G~CA~~~~A~~~~ ..T ..C ..C .GAGGGGTCAGGGGTGTCCG~"IGClG~TCGn;GCAGAAGTC'MGGGTCGCGGTCACGAAA Phe Gly Gly End
+400 +450
+!ioo
+650
TCAG~CCM~A~A~A~A~M~~CGAC . . . . . . . . . . . . . ..s......................... . . . . . . . . . .
Fig. 4. Comparison of the nt sequences of the two histone H4 genes H4C7 and H4C14. The nt of the 5’-flanking region are numbered
(negatively) from the start codon (the C preceding ATG is -1). The TATA sequence is boxed and significant sequence homologies are
overlined and/or underlined. Putative GATCC-like boxes are marked with horizontal brackets. Horizontal arrows indicate inverted
DNA repeats. Dots in the coding region of H4C7 indicate nt identities with H4C14. Dotted lines in the 5’- and 3’-flanking regions indicate
that the nt sequence was not established.

229
REFERENCES
Benton, W.D. and Davis, R.W.: Screening Igt recombinant
clones by hybridization to single plaques “in situ”. Science
196 (1977) 180-182.
Birnstiel. M.L., Busslinger, M. and Strub, K.: Transcription ter-
mination and 3’ processing: the end is in site. Cell 41 (1985)
349-359.
Hentschel, C. and Birnstiel, M.L.: The organization and expres-
sion of histone gene families. Cell 25 (1981) 301-313.
Maxam, A.M. and Gilbert, W.: Sequencing end-labeled DNA
with base-specific chemical cleavage. Methods Enzymol. 65
(1980) 499-560.
Maxson, R., Cohn, R., Kedes, L. and Mohun, T.: Expression and
organization of histone genes. Annu. Rev. Genet. 17 (1983)
239-277.
Tabata, T. and Iwabuchi, M.: Molecular cloning and nucleotide
sequence of a variant wheat histone H4 gene. Gene 3 I (1984)
285-289.
Tabata, T., Sasaki, K. and Iwabuchi, M.: The structural organi-
zation and DNA sequence of a wheat histone H4 gene. Nucl.
Acids Res. I1 (1983) 5865-5875.
Tabata, T., Fukasawa, M. and Iwabuchi, M.: The structural
organization and DNA sequence of a wheat histone H4 gene.
Mol. Gen. Genet. 196 (1984) 397-400.
Thomas, G. and Padayatty, J.D.: Organization and bidirectional
transcription of H2A. H2B and H4 histone genes in rice
embryos. Nature 306 (1983) 82-84.
Zimmerman, J.L. and Goldberg. R.B.: DNA sequence organi-
zation in the genome of Nicoticrnc~ tubacum. Chromosoma 59
(1977) 227-252.
Communicated by J.-P. Lecocq