Embed Size (px)
Citation preview

1544 B. J . Howlett Elcctrophorcsrs 1997, 18. 1544-1547
Barbara J. Howlett
plant Cell Biology Research Centre,
Genome analysis of the fungal plant pathogen, Leptosphaeria rnaculans using pulsed field gel
School of Botany, The University of Melbourne, Parkville, Victoria, electrophoresis Australia
Pulsed field gel electrophoresis (PFGE), or electrophoretic karyotyping, sepa- rates chromosomal-sized pieces of DNA in agarose gels where the orientation of the electric field is periodically altered. This technique has revealed that many fungi have a high degree of chromosomal length polymorphisms. Often the only isolates with identical karyotypes are derived from a single clone, thus PFGE provides a 'genetic fingerprint' for them. The size range and number of chromosomes within isolates of a particular species are usually con- stant, hence PFGE can distinguish between morphologically similar fungi. This technique can also be used to follow inheritance of chromosomal length polymorphisms and shows that in some fungi novel-sized chromosomes are produced during meiosis. As well as resolving the nuclear (A-type) chromo- somes, it can also resolve dispensable (B-type) chromosomes and cytoplasmic genomes including mitochondrial DNA and linear plasmids. The application of this technique to Australian isolates of Leptosphaeria maculans, which causes blackleg disease of canola (Brussica napus), is discussed.
1 Pulsed field gel electrophoresis of fungi
Pulsed field gel electrophoresis (PFGE) is particularly applicable to fungi because their chromosomes are usu- ally of the size range in which optimal resolution occurs. Consequently, karyotypes of fungi that previously could not be studied using cytological methods, due to the chromosomes failing to condense or being too small to visualise, can now be determined. Furthermore, this technique allows genome size and genetic maps to be developed for fungi that are not amenable to classical linkage analysis. Chromosomal DNA of fungi is pre- pared by embedding vegetative spores or mycelial proto- plasts into low gelling-temperature agarose in a mold, and treating them with protease and detergent to digest away other cellular components. The resultant agarose plugs are then placed in the wells of an agarose gel. The most widely used PFGE apparatus is contour-clamped homogeneous electric field [l], with a hexagonal array of electrodes. As the electric field switches through 120", the DNA molecules reorientate and then move through the gel in the new direction. Small molecules reorientate and migrate more quickly than large ones. Thus long switching intervals are required to resolve large mole- cules, whilst smaller intervals separate small molecules. A linear gradient of switching intervals from about 1.5 to 60 min over a total period of 3-8 days is used to resolve chromosomes of most filamentous fungi. When PFGE was first applied to fungi in the late 1980s the degree of karyotypic variation displayed was most unex-
pected (for reviews see [2-41. Generally, this variation is manifested as chromosomal length polymorphisms, dif- ferences in ploidy levels of individual chromosomes (aneuploidy), and presence or absence of dispensable (B-type) chromosomes. Chromosomal length polymorph- isms may result from deletions, insertions, transloca- tions, or recombination between homologous chromo- somes (homologues) of unequal length [4].
2 Leptosphaeria maculans comprises two strains, each one having a high degree of karyotypic variability
The ascomycetous fungus Leptosphaeria maculans causes blackleg disease of canola (Brussica nupus) worldwide
Correspondence: Dr. B. J. Howlett, Plant Cell Biology, Research . Centre, School of Botany, The University of Melbourne, Pdrkvik, Vic-
toria 3052, Australia (Fax: +613-9347-1071; E-mail: howlett@bota ny.unimelb.edu.au)
Nonstandard abbreviations: RAPD, random amplified polymorphic DNA; TBE, Tris-borate-EDTA
Keywords: Chromosomal length polymorphisms / B-type chromo- somes / Linear plasmids / Mitochondrial DNA / Leptosphaeria macu- l a m
Figure 1. PFGE can discriminate between highly and weakly virulent strains of L. rnaculans. Electrophoresis conditions: 0.8% agarose gel (Sigma type 11); 0.5 X 89 mM Tris-borate 2 mM EDTA (TBE) buffer; switching interval, 500 s for 72 h followed by 420 s for 48 h; 100 V. Sizes (Mbp) of several S. cerevisiae and S. pombe chromosomal DNAs are marked. Highly virulent (HV) isolates have at least 6 chromosomal DNAs that are larger than 2.2 Mbp whilst weakly virulent (WV) isolates have only 1 or 2 chromosomal DNAs that are larger than 2.2 Mbp.
0 VCH Verlagsgesellschaft mbH, 69451 Weinheim, 1997 0173-0835/97/0909-1544 $17,50+.50/Il
Elerrrnphorcris 1997, 18. 1544-1547 Genome analysis of Leptoshaeria maculans 1545
and limits the expansion of the canola industry in Australia [5]. In spite of its economic importance as a pathogen worldwide, as yet little is known about L. maculans at the molecular or genetic level. We have used PFGE to examine genome organisation of Austra- lian isolates of this fungus. Firstly we discovered that there are two strains of L. maculans in Australia which are indistinguishable morphologically but can be differ- entiated by their symptoms on canola, one (‘highly virulent’) causing stem cankers (lesions that girdle the stem) and the other (‘weakly virulent’) unable to cause such cankers. In Australia the weakly virulent strain is of much lower abundance than the highly virulent one. These strains can easily be distinguished by PFGE as the isolates of the highly virulent strain have at least six chromosomal DNAs larger than 2.2 Mbp whilst those of the weakly virulent strain have only one or two in this size range (Fig. 1). This classification is also supported by RFLP and random amplified polymorphic DNA (RAPD) analysis [ 6 ] . A similar situation occurs with L. maculans isolates from Canada and Europe where the highly and weakly virulent strains can be readily classi- fied by PFGE, RAPD and RFLP analysis and are thought to be separate species [7, 81. Within each strain we observed a high degree of chromosomal length poly- morphisms between unrelated field isolates (as in [7,8]).
3 Novel-sized chromosomes are produced during meiosis in L. maculans
L. maculans is heterothallic (outcrossing) and sexual fruiting bodies of the highly virulent strain are abundant on canola stubble in the field in Australia. Sexual fruiting bodies contain asci, sacs that house eight asco-
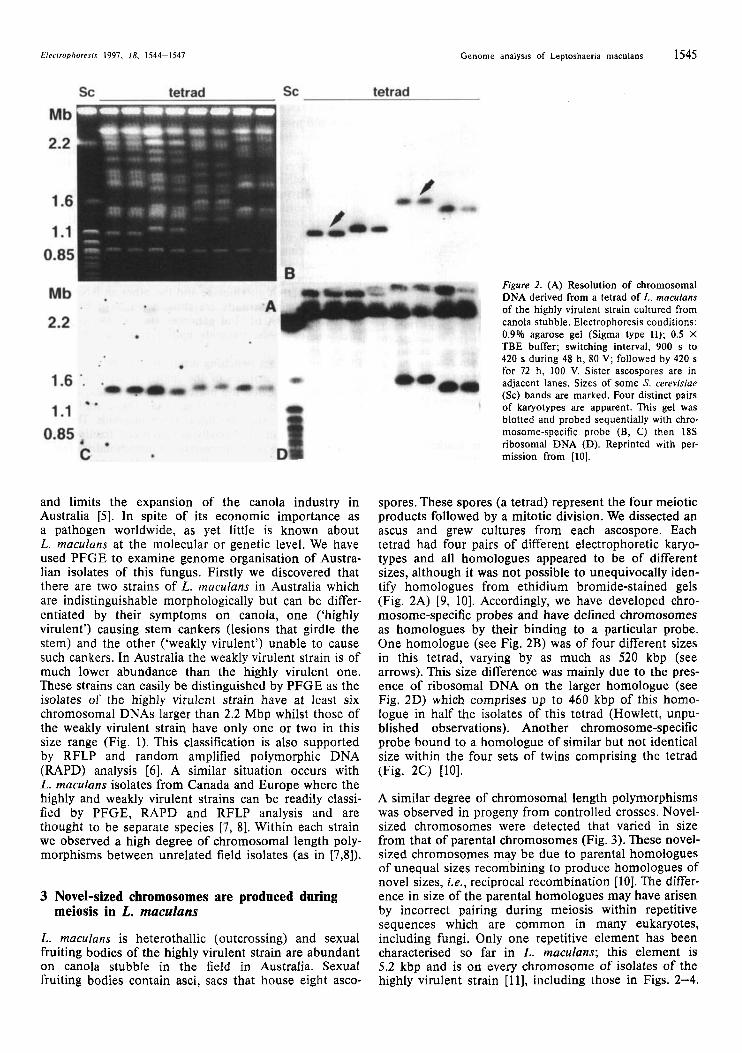
Figure 2. (A) Resolution of chromosomal DNA derived from a tetrad of L. mnculuns of the highly virulent strain cultured from canola stubble. Electrophoresis conditions: 0.9% agarose gel (Sigma type 11); 0.5 X TBE buffer; switching interval, 900 s to 420 s during 48 h, 80 V; followed by 420 s for 72 h, 100 V. Sister ascospores are in adjacent lanes. Sizes of some S. cerevisiae (Sc) bands are marked. Four distinct pairs of karyotypes are apparent. This gel was blotted and probed sequentially with chro- mosome-specific probe (B, C) then 18s ribosomal DNA (D). Reprinted with per- mission from [IO].
spores. These spores (a tetrad) represent the four meiotic products followed by a mitotic division. We dissected an ascus and grew cultures from each ascospore. Each tetrad had four pairs of different electrophoretic karyo- types and all homologues appeared to be of different sizes, although it was not possible to unequivocally iden- tify homologues from ethidium bromide-stained gels (Fig. 2A) [9, 101. Accordingly, we have developed chro- mosome-specific probes and have defined chromosomes as homologues by their binding to a particular probe. One homologue (see Fig. 2B) was of four different sizes in this tetrad, varying by as much as 520 kbp (see arrows). This size difference was mainly due to the pres- ence of ribosomal DNA on the larger homologue (see Fig. 2D) which comprises up to 460 kbp of this homo- logue in half the isolates of this tetrad (Howlett, unpu- blished observations). Another chromosome-specific probe bound to a homologue of similar but not identical size within the four sets of twins comprising the tetrad (Fig. 2C) [IO].
A similar degree of chromosomal length polymorphisms was observed in progeny from controlled crosses. Novel- sized chromosomes were detected that varied in size from that of parental chromosomes (Fig. 3). These novel- sized chromosomes may be due to parental homologues of unequal sizes recombining to produce homologues of novel sizes, i.e., reciprocal recombination [IO]. The differ- ence in size of the parental homologues may have arisen by incorrect pairing during meiosis within repetitive sequences which are common in many eukaryotes, including fungi. Only one repetitive element has been characterised so far in L. maculans; this element is 5.2 kbp and is on every chromosome of isolates of the highly virulent strain [ 111, including those in Figs. 2-4.
1546 R. J. Howlett Elecrrophorcyis 1997, 18, 1544-1517
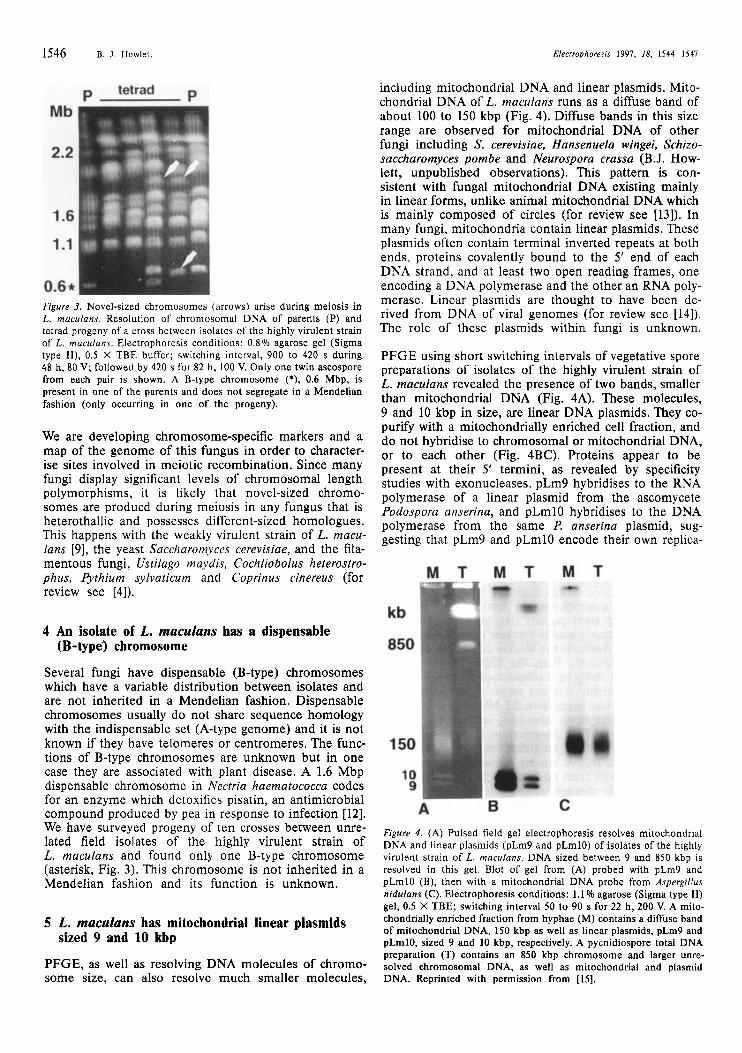
Figure 3. Novel-sized chromosomes (arrows) arise during meiosis in L. muculans. Resolution of chromosomal DNA of parents (P) and tetrad progeny of a cross between isolates of the highly virulent strain of L. maculuns. Electrophoresis conditions: 0.8% agarose gel (Sigma type II), 0.5 X TBE buffer; switching interval, 900 to 420 s during 48 h, 80 V; followed by 420 s for 82 h, 100 V. Only one twin ascospore from each pair is shown. A B-type chromosome (*), 0.6 Mbp, is present in one of the parents and does not segregate in a Mendelian fashion (only occurring in one of the progeny).
We are developing chromosome-specific markers and a map of the genome of this fungus in order to character- ise sites involved in meiotic recombination. Since many fungi display significant levels of chromosomal length polymorphisms, it is likely that novel-sized chromo- somes are producd during meiosis in any fungus that is heterothallic and possesses different-sized homologues. This happens with the weakly virulent strain of L. macu- lans [9], the yeast Saccharomyces cerevisiae, and the fila- mentous fungi, Ustilago maydis, Cochliobolus heterostro- phus, b t h i u m sylvaticum and Coprinus cinereus (for review see [4]).
4 An isolate of L. maculans has a dispensable (B-type') chromosome
Several fungi have dispensable (B-type) chromosomes which have a variable distribution between isolates and are not inherited in a Mendelian fashion. Dispensable chromosomes usually do not share sequence homology with the indispensable set (A-type genome) and it is not known if they have telomeres or centromeres. The func- tions of B-type chromosomes are unknown but in one case they are associated with plant disease. A 1.6 Mbp dispensable chromosome in Nectria haernatococca codes for an enzyme which detoxifies pisatin, an antimicrobial compound produced by pea in response to infection [12]. We have surveyed progeny of ten crosses between unre- lated field isolates of the highly virulent strain of L. maculans and found only one B-type chromosome (asterisk, Fig. 3). This chromosome is not inherited in a Mendelian fashion and its function is unknown.
5 L. maculans has mitochondrial linear plasmids
PFGE, as well as resolving DNA molecules of chromo- some size, can also resolve much smaller molecules,
sized 9 and 10 kbp
including mitochondrial DNA and linear plasmids. Mito- chondrial DNA of L. maculans runs as a diffuse band of about 100 to 150 kbp (Fig. 4). Diffuse bands in this size range are observed for mitochondrial DNA of other fungi including S. cerevisiae, Hansenuela wingei, Schizo- saccharomyces pombe and Neurospora crassa (B.J. How- lett, unpublished observations). This pattern is con- sistent with fungal mitochondrial DNA existing mainly in linear forms, unlike animal mitochondrial DNA which is mainly composed of circles (for review see 1131). In many fungi, mitochondria contain linear plasmids. These plasmids often contain terminal inverted repeats at both ends, proteins covalently bound to the 5' end of each DNA strand, and at least two open reading frames, one encoding a DNA polymerase and the other an RNA poly- merase. Linear plasmids are thought to have been de- rived from DNA of viral genomes (for review see [14]). The role of these plasmids within fungi is unknown.
PFGE using short switching intervals of vegetative spore preparations of isolates of the highly virulent strain of L. maculans revealed the presence of two bands, smaller than mitochondrial DNA (Fig. 4A). These molecules, 9 and 10 kbp in size, are linear DNA plasmids. They co- purify with a mitochondrially enriched cell fraction, and do not hybridise to chromosomal or mitochondrial DNA, or to each other (Fig. 4BC). Proteins appear to be present at their 5' termini, as revealed by specificity studies with exonucleases. pLm9 hybridises to the RNA polymerase of a linear plasmid from the ascomycete Podospora anserina, and pLmlO hybridises to the DNA polymerase from the same P. anserina plasmid, sug- gesting that pLm9 and pLmlO encode their own replica-
Figure 4. (A) Pulsed field gel electrophoresis resolves mitochondrial DNA and linear plasmids (pLm9 and pLmlO) of isolates of the highly virulent strain of L. maculans. DNA sized between 9 and 850 kbp is resolved in this gel. Blot of gel from (A) probed with pLm9 and pLmlO (B), then with a mitochondrial DNA probe from Aspergillus nidulans (C). Electrophoresis conditions: 1.1 % agarose (Sigrna type 11) gel, 0.5 X TBE; switching interval 50 to 90 s for 22 h, 200 V. A mito- chondrially enriched fraction from hyphae (M) contains a diffuse band of rnitochondrial DNA, 150 kbp as well as linear plasmids, pLm9 and pLrnlO, sized 9 and 10 kbp, respectively. A pycnidiospore total DNA preparation (T) contains an 850 kbp chromosome and larger unre- solved chromosomal DNA, as well as mitochondrial and plasmid DNA. Reprinted with permission from [15].

Electrophoresis 1997, 18, 1544-1547 Genome analysis of Leptoshaeria maculans 1547
tion and transcription enzymes [15]. pLm9 and pLmIO co-purify with mitochondria and share no sequence homology to chromosomal or mitochondrial DNA, sug- gesting that these plasmids are not derived from DNA of these organelles of L. maculans. These plasmids have not been found in isolates of the weakly virulent strain of L. maculans, but there is no evidence that they have any direct role in virulence.
6 Concluding remarks
Like many other filamentous fungi, L. rnaculans shows a high degree of karyotypic variability. Australian isolates of L. maculans comprise two strains (highly and weakly virulent) which differ in their symptoms on canola and can be readily distinguished by PFGE, RFLP and RAPD analysis. The profound differences between the genomes of these two strains suggest that they are different spe- cies. Novel-sized PFGE chromosomes are generated during meiosis in both strains and may be due to recip- rocal recombination between parental chromosomes of unequal sizes. The highly virulent strain of L. maculans possesses linear plasmids sized 9 and 10 kbp, and a B-type chromosome occurs in one isolate. The unique- ness of karyotypes of L. maculans and other fungi is valuable for population studies of pathogens as it pro- vides a fingerprint for clonal populations. Hence PFGE can provide knowledge of population structure and genome organisation within fungi. Such knowledge can assist plant breeders to design disease resistance strate- gies.
I ihank Professor Adrienne Clarke for her constant encoura- gement and for making the facilities of the Plant Cell Biol- ogy Research Centre available. This project is funded by the Australian Research Council and the Australian Grains Research and Development Corporation.
Received December 20, 1996
7 References
[l] Chu, G., Vollrath, D., Davis, R. W., Science 1986, 234, 1582-1585. (21 Mills, D., McCluskey, K., MO/. Plant-Microbe Inter. 1990, 3,
[3] Kistler, H. C., Miao, V. P. W., Annu. Rev. Phyiopath. 1992, 30,
[4] Zolan, M. E., Microbial. Rev. 1995, 59, 686-698. [5] Salisbury, P. A., Ballinger, D. J., Wratten, N., Plummer, K. M.,
[6] Plummer, K. M., Dunse, K., Howlett, B. J., Aust. J. Bat. 1994, 42,
[7] Taylor, J. L., Borgmann, I. E., Seguin-Swartz, G . , Curr. Genet.
[8] Morales, V. M., Seguin-Swartz, G. , Taylor, J. L., Phytopath. 1993,
[9] Plummer, K. M., Howlett, B. J., Curr. Genet. 1993, 24, 107-113. [ l O ] Plummer, K. M., Howlett, B. J., Mol. Gen. Genet. 1995, 247,
[ l l ] Taylor, J. L., Borgmann, 1. E., Mol. Plant-Microbe Inter. 1994, 7,
[12] Miao, V. P., Covert, S. F., VanEtten, H. D., Science 1991, 254,
[13] Bendich, A., Curr. Genet. 1993, 24, 279-290. [14] Meinhardt, F., Kempken, F., Kamper, J., Esser, K., Curr. Genet.
[15] Lim, L., Howlett, B. J. (1994), Curr. Genet. 1994, 26, 276-280.
35 1-357.
3 13-352.
Howlett, B. J., Aust. J. Exp. Agr. 1995, 35, 665-674.
1-8.
1991, 19, 273-277.
83, 503-509.
41 6-422.
181-188.
1773- 1776.
1990, 17, 89-95.





























