Embed Size (px)
Citation preview

2/13/2012
1
Genetics Lecture 7 More Mendelian Genetics Continued
Novel Phenotypes• Other cases of gene interaction yield novel, or new, phenotypes in
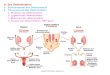
the F2 generation, in addition to producing modified dihybrid ratios. • An example, the inheritance of fruit shape in the summer squash
Cucurbita papa. • When plants with disc‐shaped fruit (AABB) are crossed with plants
with long fruit (aabb), the F1 generation all have disc fruit. • However, in the F2 progeny, fruit with a novel shape— sphere—
appear, as well as fruit exhibiting the parental phenotypes. • The F2 generation, with a modified 9:6:1 ratio, is generated as
f ffollows: In this example of gene interaction, both gene pairs influence fruit shape equally A dominant allele at either locus ensures a sphere—shaped fruit.
• In the absence of dominant alleles, the fruit is long. •
2
3

2/13/2012
2
4
Other Modified Dihybrid Ratios
• First, we need not violate the principles of segregation and independent assortment to explain the inheritance pattern
• The added complexity of inheritance in these examples does not detract from the validity of Mendel’s
l iconclusions. • Second, the F2 phenotypic ratio in each example has been expressed in sixteenths.
• When sixteenths are seen in the ratios of crosses where the inheritance pattern is unknown, they suggest to geneticists that two gene pairs are controlling the observed phenotypes.
5
Complementation Analysis Can Determine lf Two Mutations Causing a Similar Phenotype Are Alleles of
the Same Gene• An interesting situation arises when two mutations that both
produce a similar phenotype are isolated independently. • Suppose that two investigators independently isolate and establish
a true‐breeding strain of wingless Drosophila and demonstrate that each mutant phenotype is due to a recessive mutation.
• We might assume that both strains contain mutations in the same gene.
• However, since we know that many genes are involved in the formation of wings, we must consider the possibility that mutations in any one of them might inhibit wing formation during development.
• An analytical procedure called complementation analysis allows us to determine whether two independently isolated mutations are in the same gene‐—that is, whether they are alleles—or whether they represent mutations in separate genes.
6

2/13/2012
3
• Our analysis seeks to answer this simple question; Are two mutations that yield similar phenotypes present in the same gene or in two different genes?two different genes?
• To find the answer, we cross the two mutant strains and analyze the F1 generation.
7
• Case 1. All offspring develop normal wings. • Interpretation: The two recessive mutations are in separate
genes and are not alleles of one another. • Following the cross, all F1 flies are heterozygous for both
genes. Si h i i i d h fl i• Since each mutation is in a separate gene and each F1 fly is heterozygous at both loci, the normal products of both genes are produced (by the one normal copy of each gene), and wings develop.
• Under such circumstances, the genes complement one another in restoration of the wild‐type phenotype, and complementation is said to occur because the two mutations are in different genes.
8
• Case 2. All offspring fail to develop wings. Interpretation: The two mutations affect the same gene and are alleles of one another.
• Complementation does not occur. • Since the two mutations affect the same gene, the F1 flies are homozygous for the two mutant alleles (the ma allele and the mb allele).
••
9

2/13/2012
4
10
Expression of a Single Gene May Have Multiple Effects
• While the previous sections have focused on the effects of two or more genes on a single characteristic, the converse situation, where expression of a single gene has multipleexpression of a single gene has multiple phenotypic effects, is also quite common.
• This phenomenon, which often becomes apparent when phenotypes are examined carefully, is referred to as pleiotropy.
11
• The first disorder is Marfan syndrome, a human malady resulting from an autosomal dominant mutation in the gene encoding the connective tissue protein fibrillin.
• Because this protein is widespread in many tissues in the body, one would expect multiple effects of such a defect.
• In fact, fibrillin is important to the structural integrity of the lens of the eye to the lining of vessels such asof the lens of the eye, to the lining of vessels such as the aorta, and to bones, among other tissues.
• As a result, the phenotype associated with Marfan syndrome includes lens dislocation, increased risk of aortic aneurysm, and lengthened long bones in limbs.
• This disorder is of historical interest in that speculation abounds that Abraham Lincoln was afflicted.
12

2/13/2012
5
X‐Linkage Describes Genes on the X Chromosome
• ln many animals and some plant species, one of the sexes contains a pair of unlike chromosomes that are involved in sex determination.
• In many cases, these are designated as X and Y • For example, in both Drosophila and humans, males contain an X and a Y
chromosome, whereas females contain two X chromosomes. • The Y chromosome must contain a region of pairing homology with the X
chromosome if the two are to synapse and segregate during meiosis but achromosome if the two are to synapse and segregate during meiosis, but a major portion of the Y chromosome in humans as well as other species is considered to be relatively inert genetically.
• While we now recognize a number of male—specific genes on the human Y chromosome, it lacks copies of most genes present on the X chromosome.
••
13
X‐Linkage in Humans
• In humans, many genes and the respective traits controlled by them are recognized as being linked to the X chromosome.
• These X—linked traits can be easily identified in a pedigree because of the crisscross pattern of inheritance.
• The mother in generation 1 passes the trait to all her sons• The mother in generation 1 passes the trait to all her sons but to none of her daughters. If the offspring in generation 2 marry normal individuals, the colorblind sons will produce all normal male and female offspring; the normal‐visioned daughters will produce normal‐visioned female offspring, as well as color‐blind and normal‐visioned male offspring.
14
15

2/13/2012
6
16
• The way in which X‐linked genes are transmitted causes unusual circumstances associated with recessive X‐linked disorders, in comparison to recessive autosomal disorders.
• For example, if an X‐linked disorder debilitates or is lethal to the affected individual prior to reproductive maturation, the disorder occurs exclusively in males.
• This is so because the only sources of the lethal allele in the population are in heterozygous females who are “carriers” and do not express the disorder.
• They pass the allele to one‐half of their sons, who develop the disorder because they are hemizygous but rarely, if ever, reproduce.
• Heterozygous females also pass the allele to one‐half of their daughters, who become carriers but do not develop the disorder.
• An example of such an X‐ linked disorder is Duchenne muscular dystrophy.
• The disease has an onset prior to age 6 and is often lethal around age 20.
• It normally occurs only in males.
17
Sex‐Limited and Sex‐Influenced Inheritance, an Individual's Sex Influences the Phenotype
• In contrast to X‐linked inheritance, patterns of gene expression may be affected by the sex of an individual even when the genes are not on the X chromosome.
• ln numerous examples in different organisms, the sex of the individual plays a determining role in the
i f h texpression of a phenotype. • In some cases, the expression of a specific phenotype is absolutely limited to one sex; in others, the sex of an individual influences the expression of a phenotype that is not limited to one sex or the other.
••
18

2/13/2012
7
• In both types of inheritance, autosomal genes are responsible for the existence of contrasting phenotypes, but the expression of these genes is dependent on the hormone constitution of the individual.
• Thus, the heterozygous genotype may exhibit one phenotype in males and the contrasting one in females.
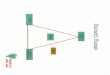
• In domestic fowl, for example, tail and neck plumage is often distinctly different in males and females, demonstrating sex‐limited inheritance.
• Cock feathering is longer, more curved, and pointed, whereas hen feathering is shorter and less curvedfeathering is shorter and less curved.
• Inheritance of these feather phenotypes is controlled by a single pair of autosomal alleles whose expression is modified by the individual’s sex hormones.
• As shown in the following chart, hen feathering is due to a dominant allele, H, but regardless of the homozygous presence of the recessive h allele, all females remain hen‐feathered.
• Only in males does the hh genotype result in cock feathering.
19
20
pattern baldness in humans• Cases of sex influenced inheritance include pattern
baldness in humans, horn formation in certain breeds of sheep, and certain coat patterns in cattle.
• In such cases, autosomal genes are responsible for the contrasting phenotypes, and while the trait may be displayed by both males and females, the expression of h i d d h h i i fthese genes is dependent on the hormone constitution of the individual.
• Thus, the heterozygous genotype exhibits one phenotype in one sex and the contrasting one in the other.
• Females can display pattern baldness, but this phenotype is much more prevalent in males. – When females do inherit the BB genotype, the phenotype is less
pronounced than in males and is expressed later in life.
21

2/13/2012
8
22
Genetic Background and the Environment May Alter Phenotypic Expression
• We assumed that the genotype of an organism is always directly expressed in its phenotype.
• For example, pea plants homozygous for the recessive d allele (dd) will always be dwarf.
• We discussed gene expression as though the genes operate in a closed system in which the presence or absence of functional products directly determines the collective phenotype of an individualindividual.
• The situation is actually much more complex. • Most gene products function within the internal milieu of the cell,
and cells interact with one another in various ways. • Furthermore, the organism exists under diverse environmental
influences. ••
23
Penetrance and Expressivity
• Some mutant genotypes are always expressed as a distinct phenotype, whereas others produce a proportion of individuals whose phenotypes cannot be distinguished from normal (wild type).
• The degree of expression of a particular trait canThe degree of expression of a particular trait can be studied quantitatively by determining the penetrance and expressivity of the genotype under investigation.
• The percentage of individuals that show at least some degree of expression of a mutant genotype defines the penetrance of the mutation.
24

2/13/2012
9
• For example, the phenotypic expression of many of the mutant alleles found in Drosophila can overlap with wild‐type expression.
• lf 15 percent of flies with a given mutant genotype show the wild‐type appearance, the mutant gene is id t h t f 85 tsaid to have a penetrance of 85 percent.
• By contrast, expressivity reflects the range of expression of the mutant genotype.
• Flies homozygous for the recessive mutant gene eyeless exhibit phenotypes that range from the presence of normal eyes to a partial reduction in size to the complete absence of one or both eyes
25
26
• Although the average reduction of eye size is one‐fourth to one—half, expressivity ranges from complete loss of both eyes to completely normal eyes.
• Examples such as the expression of the eyeless gene have provided the basis for experiments to determine the causes of phenotypic variation.
• lf the laboratory environment is held constant and extensive variation is still observed, other genes may be influencing or modifying the phenotype. g y g p yp
• On the other hand, if the genetic background is not the cause of the phenotypic variation, environmental factors such as temperature, humidity, and nutrition may be involved.
• In the case of the eyeless phenotype, experiments have shown that both genetic background and environmental factors influence its expression.
27

2/13/2012
10
Genetic Background: Position Effects
• Although it is difficult to assess the specific effect of the genetic background and the expression of a gene responsible for determining a potential phenotype, one effect of genetic background has been well characterized, called the position effect.
• In such instances, the physical location of a gene in relation to other genetic material may influence its expressionto other genetic material may influence its expression.
• For example, if a region of a chromosome is relocated or rearranged (called a translocation or inversion event), normal expression of genes in that chromosomal region may be modified.
••
28
• An example of a position effect involves female Drosophila heterozygous for the X‐linked recessive eye color mutant White (W).
• The w+/w genotype normally results in a wild—type brick—red eye colorbrick red eye color.
• However, if the region of the X chromosome containing the wild‐type allele is translocated so that it is close to a heterochromatic region, expression of the w+ allele is modified.
• Instead of having a red color, the eyes are variegated, or mottled with red and white patches.
29
30

2/13/2012
11
• Therefore, following translocation, the dominant effect of the normal w+ allele is intermittent.
• A similar position effect is produced if a heterochromatic region is relocated next to the white locus on the X chromosome.
• Apparently, heterochromatic regions inhibit the expression of adjacent genes.
•••
31
Temperature Effects‐An Introduction to Conditional Mutations
• Chemical activity depends on the kinetic energy of the reacting substances, which in turn depends on the surrounding temperature.
• We can thus expect temperature to influence phenotypes.
• An example is seen in the evening primrose, which produces red flowers when grown at 23°C and white flowers when grown at l8°C.
• An even more striking example is seen in Siamese cats and Himalayan rabbits, which exhibit dark fur in certain regions where their body temperature is slightly cooler, particularly the nose, ears, and paws.
32
33

2/13/2012
12
• In these cases, it appears that the enzyme normally responsible for pigment production is functional only at the lower temperatures present in the extremities, but it loses its catalytic function at the slightly higher temperatures found throughout the rest of the body.
• Mutations whose expression is affected by temperature, called temperature‐sensitive mutations, are examples of conditional mutations, whereby phenotypic expression is determined by environmental conditionsdetermined by environmental conditions.
• Examples of temperature‐sensitive mutations are known in viruses and a variety of organisms, including bacteria, fungi, and Drosophila.
• In extreme cases, an organism carrying a mutant allele may express a mutant phenotype when grown at one temperature but express the wild‐type phenotype when reared at another temperature.
34
Nutritional Effects
• Another category of phenotypes that are not always a direct reflection of the organisms genotype consists of nutritional mutations.
• In microorganisms, mutations that prevent synthesis of nutrient molecules are quite common, such as when an enzyme essential to a biosynthetic pathway becomes inactive. A i i b i h i i ll d h• A microorganism bearing such a mutation is called an auxotroph.
• lf the end product of a biochemical pathway can no longer be synthesized, and if that molecule is essential to normal growth and development, the mutation prevents growth and may be lethal.
• For example, if the bread mold Neurospora can no longer synthesize the amino acid leucine, proteins cannot be synthesized.
•
35
• A slightly different set of circumstances exists in humans. • The ingestion of certain dietary substances that normal
individuals may consume without harm can adversely affect individuals with abnormal genetic constitutions.
• Often, a mutation may prevent an individual from metabolizing some substance commonly found in normal diets.
• For example, those afflicted with the genetic disorder phenylketonuria cannot metabolize the amino acidphenylketonuria cannot metabolize the amino acid phenylalanine.
• Those with galactosemia cannot metabolize galactose. • Those with lactose intolerance cannot metabolize lactose.
However, if the dietary intake of the involved molecule is drastically reduced or eliminated, the associated phenotype may be ameliorated.
36

2/13/2012
13
Onset of Genetic Expression• Not all genetic traits become apparent at the same time during an
organism’s life span. • In most cases, the age at which a mutant gene exerts a noticeable
phenotype depends on events during the normal sequence of growth and development.
• In humans, the prenatal, infant, preadult, and adult phases require different genetic information. g
• As a result, many severe inherited disorders are not manifested until after birth.
• For example, Tay‐Sachs disease, inherited as an autosomal recessive, is a lethal lipid metabolism disease involving an abnormal enzyme, hexosaminidase A.
• Newborns appear to be phenotypically normal for the first few months. ••
37
• Perhaps the most delayed and highly variable age of onset for a genetic disorder in humans is seen in Huntington disease.
• Inherited as an autosomal dominant disorder, Huntington disease affects the frontal lobes of the cerebral cortex, where progressive cell death occurs over a period of more than a decade.
• Brain deterioration is accompanied by spastic uncontrolled movements, intellectual deterioration, and ultimately death.
• While onset of these symptoms has been reported at all ages, they are most often initially observed between ages 30 and 50, with a mean onset age of 38 years.
• These examples support the concept that gene products may play more essential roles at certain times during the life cycle of an organism.
• One may be able to tolerate the impact of a mutant gene for a considerable period of time without noticeable effect. At some point, however, a mutant phenotype is manifested.
38
Genetic Anticipation
• Interest in studying the genetic onset of phenotypic expression has intensified with the discovery of heritable disorders that exhibit a progressively earlier age of onset and an increased severity of the disorder in each successive generation.
• This phenomenon is referred to as genetic anticipation. • Myotonic dystrophy (DM), the most common type of adult
l d h l l ill i i i imuscular dystrophy clearly illustrates genetic anticipation.• Individuals afflicted with this autosomal dominant disorder exhibit
extreme variation in the severity of symptoms. • Mildly affected individuals develop cataracts as adults, but have
little or no muscular weakness. • Severely affected individuals demonstrate more extensive
weakness, as well as myotonia (muscle hyperexcitability) and in some cases mental retardation.
• In its most extreme form, the disease is fatal just after birth.
39

2/13/2012
14
• A great deal of excitement was generated in 1989, when C. I. Howeler and colleagues confirmed the correlation of increased severity and earlier onset with successive generations of inheritance.
• The researchers studied 61 parent‐child pairs, and in 60 of the cases, age of onset was earlier and more severe in the child than in his or her affected parent.
• A three‐nucleotide DNA sequence of the DM gene is repeated a variable number of times and is unstable.
• Normal individuals have about 5 to 35 copies of this sequence; p q ;affected individuals have between 80 and >2500 copies.
• Those with a greater number of repeats are more severely affected.• The most remarkable observation was that, in successive
generations of DM individuals, the size of the repeated segment increases.
• We now know that the RNA transcribed from mutant DM genes is the culprit in the disorder and alters the expression of still other genes.
40
Genomic (Parental) Imprinting and Gene Silencing
•
• Examples involve cases where genes or regions of a chromosome are imprinted on one homolog but not the other.
• The impact of silencing depends on the parental origin of the genes or regions that are involved.
• Such silencing leads to the direct phenotypic expression of the allele(s) on• Such silencing leads to the direct phenotypic expression of the allele(s) on the homolog that is not silenced.
• Thus, the imprinting step, the critical issue in understanding this phenomenon, is thought to occur before or during gamete formation, leading to differentially marked genes (or chromosome regions) in sperm‐forming versus egg‐forming tissues.
• The first example of genomic imprinting was discovered in 1991, in three specific mouse genes.
• One is the gene encoding insulin‐like growth factor H (Igf2).
41
• A mouse that carries two normal alleles of this gene is normal in size, whereas a mouse that carries two mutant alleles lacks the growth factor and is dwarf.
•
• The mouse is normal in size if the normal allele comes from the father, but it is dwarf if the normal allele came from the mother.
• From this, we can deduce that the normal 1gf2 gene is imprinted and thus silenced during egg production, but it functions normally when it has passed through sperm producing tissue in maleswhen it has passed through sperm producing tissue in males.
• The imprint is inherited in the sense that the Igf2 gene in all progeny cells formed during development remain silenced.
• Imprinting in the next generation then depends on whether the gene passes through sperm producing or egg‐forming tissue.
42