Embed Size (px)
Citation preview

Genetic differentiation and distribution of Pyropia acanthophora (Bangiales,Rhodophyta) in the PhilippinesRichard V. Dumilaga,b,c and Zae-Zae A. Aguinaldoa,c
aDepartment of Biological Sciences, Institute of Arts and Sciences, Far Eastern University, Nicanor Reyes Sr. St., Sampaloc, Manila,Philippines 1015; bInstitute of Biology, College of Science, University of the Philippines, Diliman, Quezon City, Philippines 1101;cMarine Genomics and Molecular Genetics Laboratory, The Marine Science Institute, College of Science, University of thePhilippines, Diliman, Quezon City, Philippines 1101
ABSTRACTPyropia acanthophora is a foliose Bangiales with widely known endemic populations in Indo-Pacific region. This alga hasexpanded its range recently as a consequence of introduction. In an attempt to explore the genetic diversity of Py.acanthophora within the Philippines and the impact of the introduction of the species from elsewhere, an examination ofmolecular differentiation and distribution was undertaken using the mitochondrial COI-5P and plastid rbcL genesequences. The results revealed that the populations of Py. acanthophora in the Philippines exhibited high haplotypic andgenetic diversities, and were found to be distinct from those previously reported as conspecific populations found inTaiwan, India, Japan, Hawaii, and from those introduced populations from Brazil. The network analyses as inferred fromrbcL and from the combined COI-5P and rbcL genes showed evidence that the Philippine populations of Py. acanthophoraexhibited a chaotic patchiness pattern characterized by a population with highly site-exclusive haplotypes, wide geneticvariability and lack of local geographic patterns. The distribution of Py. acanthophora within the Philippines was also foundto be greater than what was previously known, ranging from the extreme northwest to extreme northeast mainland Luzoncoasts, including Camiguin Is., Cagayan. Understanding the genetic diversity and distribution of Py. acanthophora in thePhilippines provides valuable information in relation to the conservation and effective resource management of nativepopulations of Py. acanthophora in the tropical Asian region.
ARTICLE HISTORY Received 25 January 2016; Accepted 26 July 2016
KEYWORDS Bangiales; chaotic patchiness; COI-5P; conservation; haplotype; species introductions; Philippines; Pyropia; Pyropiaacanthophora; rbcL
Introduction
The red algal genus Pyropia J. Agardh, Bangiales,includes species of considerable systematic and bio-geographic interest due to widespread distribution(Sutherland et al., 2011) and economic value of itscircumscribed taxa (Hanisak, 1998; Mumford &Miura, 1998). Notwithstanding the greater speciesdiversity of Pyropia across boreal to cold temperateregions (Brodie et al., 1998, 2007; Broom et al., 1999;Kunimoto et al., 1999; Jones et al., 2004; Kucera &Saunders, 2012; Mols-Mortensen et al., 2012, 2014),the distribution of the genus extends to some Asiantropical areas, albeit with fewer species (Tanaka &Pham-Hoàng, 1962; Cordero, 1974; Soe-Htun & ZawZaw Pe, 1986; Istini et al., 1998; Lewmanomont, 1998;Ogawa, 2001; Kavale et al., 2015a, 2015b). The genus isthe most speciose of the Bangiales, which currentlyincludes 64 taxonomically accepted species (Guiry &Guiry, 2016). The group is notoriously difficult interms of species discrimination, largely because ofwide convergence in almost any available taxonomicdiagnostics observed among its members (Sutherlandet al., 2011). It has been suggested that the total
number of taxa or even the number of genera withinthe Pyropia lineage might be much higher than cur-rently recognized (Sutherland et al., 2011).
Molecular genetic data have revealed importantinsights into the diversity of taxa whose informativephenotypic characters are insufficient for speciesidentification. Especially among members of thegenus Pyropia, integrative studies incorporating mor-phological, molecular and biogeographic data haverecently addressed key hypotheses on genetic diver-sity and distribution (Sutherland et al., 2011).Comparative analysis of variable sites of the plastidribulose-1,5-bisphosphate carboxylase/oxygenaselarge subunit gene (rbcL) and the 5’ end region ofthe mitochondrial cytochrome c oxidase I gene (COI-5P) sequences have overcome the limit of morpholo-gical identification among the members of Pyropiaand have resolved issues on the extent of speciesdistribution (Kucera & Saunders, 2012; Guilleminet al., 2016).
Although a significant number of Pyropia speciesare distributed endemically (Yoshida et al., 1990;Griffin et al., 1999; Kucera & Saunders, 2012;
CONTACT Richard V. Dumilag [email protected], [email protected]
EUROPEAN JOURNAL OF PHYCOLOGY, 2017VOL. 52, NO. 1, 104–115http://dx.doi.org/10.1080/09670262.2016.1230786
© 2017 British Phycological Society

Mateo-Cid et al., 2012; Nelson & D’Archino, 2014;Ramírez et al., 2014; Lindstrom et al., 2015; Hardenet al., 2016), there are a few taxa that have beenintroduced into new areas, which have either nothistorically been part of their native ranges (Yarishet al., 1998; Watson et al., 1998, 1999; West et al.,2005; Verlaque, 2001; Neefus et al., 2008; Vergéset al., 2013a, 2013b; Sánchez et al., 2014; Nelsonet al., 2014) or have long been considered as part oftheir local floras (Brodie et al., 1998; Milstein et al.,2015). Most of these taxa have been introduced toAtlantic and Mediterranean coasts from the Pacificregion and had occurred at an earlier time via globalanthropogenic activities (Table 1) but went unrec-orded due to difficulties in species identification(Brodie et al., 1998; Neefus et al., 2008; Milsteinet al., 2015). The establishment of the specific sourcesof the introduced species has also been a challengedue to the lack of information on their correspondingconspecific strains (or identical haplotypes), whichrepresent their assumed native ranges (Neefus et al.,2008; Dumilag et al., 2016).
Pyropia acanthophora (E. C. Oliveira & Coll) M. C.Oliveira, D. Milstein & E. C. Oliveira was originallydescribed from Ubatuba, São Paulo, Brazil asPorphyra acanthophora (Oliveira Filho & Coll,1975). The current range of distribution of this spe-cies has been reported along the warm temperateregion of Atlantic Ocean (John et al., 2004; Milstein& Oliveira, 2005; Creed et al., 2010; Sutherland et al.,2011; Milstein et al., 2012, 2015) and Indo-Pacificregion (Yoshida, 1997, 1998; Lee & Kang, 2001;Kavale et al., 2015a; Sherwood et al., 2010; Rivera,2014; Xie et al., 2015). For over 40 years, Py. acantho-phora was regarded as part of the indigenous flora inBrazil (Oliveira Filho & Coll, 1975), but recent mole-cular analysis has suggested that this species wasintroduced (Milstein et al., 2012, 2015). Species iden-tification for Py. acanthophora can be difficult with-out molecular data (Milstein et al., 2015; Xie et al.,2015). The presence of Py. acanthophora in thePhilippines was recently confirmed using COI-5Pgene sequences based on the specimens collectedfrom Cagayan (Rivera, 2014). Those specimens mayhave been described previously as Pyropia denticulata(Levring) J.A. Philips & J.E. Sutherland (Quisumbing,1951) or Pyropia suborbiculata (Kjellman) J.E.Sutherland, H.G. Choi, M.S. Hwang & W.A. Nelson(Cordero, 1974; Masuda et al., 1991) due to theirmorphological similarities based on the range of mor-phology in different habitats, especially on the shapeof the thallus and on the presence of a dentate marginof the blade (Levring, 1953; Broom et al., 2002; Xieet al., 2015).
On the basis of phylogenetic analysis, the commonancestry and native lineages of Py. acanthophora areconfirmed to occur across much of its known current Table
1:Su
mmaryof
thecurrently
know
nintrod
uced
Pyropiaspecies.
Species
Prob
able
vector
ofintrod
uctio
nOrigin
Extent
(Reference)
Py.acanthophora(E.C
.Oliveira
&Coll)M.C
.Oliveira,
D.M
ilstein
&E.
C.O
liveira
ballastwater,
shipping
,aquacultu
re
Indo
-Pacific
Brazil(Milstein
etal.,2015)
Py.k
atadae
(A.M
iura)M.S.H
wang,
H.G.C
hoi,N.
Kikuchi
&M.M
iyata
shipping
Northwestern
Pacific:
prob
ably
Japanor
China
Atlantic
UnitedStates:N
ewEn
gland(N
eefuset
al.,2008)
Py.leucosticta
(Thu
ret)C.D.N
eefus&
J.Brodie
shipping
Pacific
North
Atlantic
Ocean:G
reat
Britain
andIreland(Brodieet
al.,1998)
Py.koreana
(M.S.H
wang&I.K.L
ee)M.S.H
wang,H.
G.C
hoi,Y.S
.Oh&
I.K.L
eeno
data
Northwestern
Pacific
BalearicIs.,Spain(V
ergéset
al.,2013bas
Py.o
livii),N
ewZealand
(Nelsonet
al.,2014)
Py.sub
orbiculata
(Kjellm
an)J.E
.Sutherland,
H.G.C
hoi,
M.S.H
wang&
W.A.N
elson
shipping
Northwestern
Pacific:Japan,
China
Atlantic
UnitedStates:C
onnecticut,D
elaw
are,North
Carolina,
CanaryIsland
s,Mexico,
New
Zealand
(Hum
m,1979as
Po.carolinensisJ.Coll&
J.Cox;B
room
etal.,2002;A
guilar-Rosas
&Aguilar-Rosas,2003;
Neefuset
al.,2008),Brazil(Torrano
-Silvaet
al.,2010;M
ilstein
etal.,2015),andEu
rope:N
ortheastern
Atlantic&Mediterraneancoasts(H
arou
netal.,2002,asPo
.carolinensis;Milstein
etal.,2012;V
ergésetal.,
2013a;Sànchezet
al.,2014)
Py.tan
egashimensis(Shinm
ura)
N.K
ikuchi
&E.
Fijiyoshi
ballastwater,
shipping
,aquacultu
re
Indo
-Pacific
Brazil(Milstein
etal.2
015)
Py.vietnam
ensis(Tak.T
anaka&
Pham
-Hoàng
Ho)
J.E.
Sutherland
&Mon
otilla
ballastwater,
shipping
,aquacultu
re
Indo
-Pacific
Brazil(Milstein
etal.2
015)
Py.y
ezoensis(U
eda)
M.S.H
wang&
H.G
.Cho
i(f.
yezoensis&
f.na
rawaensis)
shellfish
aquacultu
re,
shipping
North
Asiaand/or
EastAsia
Pacific:Japan
MediterraneanSea:ThauLagoon
(Verlaqu
e,2001),NortheasternAmerica(Yarishet
al.,1998):Maine
(Watsonet
al.,1998,1
999),N
ewHam
pshire
(Westet
al.,2005),New
England(N
eefuset
al.,2008)
EUROPEAN JOURNAL OF PHYCOLOGY 105

distributional ranges in the Indo-Pacific region(Milstein et al., 2015). While populations of Py.acanthophora from India and Taiwan have been pre-viously examined using COI-5P and rbcL sequences(Kavale et al., 2015a; Xie et al., 2015), the geneticdiversity and distribution of Py. acanthophora withinthe Philippines are still virtually unknown.
The aim of this study is to describe the geneticdifferentiation and haplotype distribution of COI-5Pand rbcL sequences of Py. acanthophora from thePhilippines, including also available sequences of Py.acanthophora from other locations with emphasis onthe introduced Py. acanthophora strains.
Materials and methods
Fifty specimens of Py. acanthophora were collectedfrom 13 sites in the northern Philippines (Table 2).Upon collection, a fragment of each specimen waspreserved in absolute ethanol in a 1.5 ml microcen-trifuge tube. Voucher specimens were prepared fol-lowing Trono & Ganzon-Fortes (1988) and weredeposited in Jose Vera Santos Memorial Herbarium(PUH) with replicates in Far Eastern UniversityHerbarium (FEUH) and in Gregorio T. VelasquezPhycological Herbarium, Marine Science Institute,University of the Philippines, Diliman, Quezon City,Philippines (MSI). Herbarium abbreviations followedThiers (2016).
DNA extraction was carried out using QiagenPlant MiniKit (Valencia, CA, USA) following themanufacturer’s protocol. The rbcL primers used inthis study included Kito-F1 (Kito et al., the primersequence published only from GenBank entryAB118578) and JrSR (Broom et al., 2010). Four pri-mer combinations were also used to amplify the rbcLgene for samples that did not amplify successfullyusing the former primers: F67 and R502, F461 andR901, F870 and R1312, and 5rc and rbcspc (Teasdaleet al., 2002, Vergés et al., 2013a); the amplificationprofiles followed that of Teasdale et al. (2002) andVergés et al. (2013). For COI-5P gene amplification,the primers GazF1 and GazR1 (Saunders, 2005) wereused following the profiles used by Hebert et al.(2003). All PCR reactions were prepared in volumesof 30 µl with 1X Vivantis™ Buffer A, 0.1mM dNTPs,1.5 mM MgCl2 and 0.5 U Taq polymerase. Ampliconswere sent to First Base Co. (Selangor, Malaysia) forsequencing. Viewing of chromatograms and contig-uous sequence assembly were carried out using DNABaser version 3.5 (Heracle Biosoft). Generatedsequences together with other related sequencesfrom GenBank were aligned using ClustalW(Thompson et al., 1994) via the software MEGA ver.6 (Tamura et al., 2011). The COI-5P and rbcL regionsof Bangia fuscopurpurea (Dillwyn) Lyngbye isolateGWS001869 (Kucera & Saunders, 2012) were used
as outgroup sequences for the construction of phylo-grams. For sequence divergence, percent Kimura-2parameter (K2P) distances were computed usingMEGA ver. 6. The aligned sequences for rbcL(n=70) and COI-5P (n=63) were trimmed to finallength of 1317 bp and 642 bp, respectively. A hier-archical-likelihood ratio test was computed inModeltest 3.04 (Posada, 2008) to determine themodel of evolution that best fit the sequence data.
Bayesian inference (BI) was estimated using theprogram MrBayes v3.1.2 (Ronquist & Huelsenbeck,2003). The posterior probabilities were computedusing a Metropolis-coupled Markov chain MonteCarlo sampling with 10 million generations. Onehundred thousand trees were produced from savinga single tree in every 100 generations. Standard devia-tion of the split frequency between two simultaneousruns and the Potential Scale Reduction Factor (PSRF)diagnostics was examined to confirm convergence.The first 25% of the trees generated from the burn-in period was discarded and the remaining trees wereused to obtain the majority-rule consensus tree. Theresulting phylogenetic trees were visualized usingFigTree (available http://tree.bio.ed.ac.uk/software).
A statistical parsimony analysis of haplotypes wasperformed using TCS ver. 1.21 (Clement et al., 2000)with a 95% parsimony connection limit and treat-ment of gaps as missing states. Departure from neu-trality on the basis of mismatch distribution (Tajima’sD and Fu & Li’s D* tests) and genetic diversitystatistics including haplotype diversity (h), nucleotidediversity (π) and average sequence divergence (k)were computed using DNAsp ver.5 (Librado &Rozas, 2009). Haplotype diversity and nucleotidediversity values range from 0 to 1. We consideredany population with high h and π diversities if theestimates are greater than 0.5. All figures presented inthis study were edited using Adobe® Illustrator® CS5.
Results
Species identification based on interspecificvariation and phylogenetic analysis
The intraspecific Kimura-2 parameter (K2P) diver-gence for the COI-5P of Py. acanthophora rangedfrom 0–0.7% while its rbcL sequences ranged from0–0.9%. These divergence ranges were consistentwith intraspecific variation reported withinPyropia species (Lindstrom, 2008; Kucera &Saunders, 2012; Milstein et al., 2015 and referencestherein) and thus the respective sequences could beassigned conspecifically.
Species identification of our materials was con-firmed based on inferred phylogenetic trees usingrbcL and COI-5P aligned matrices. The resultingphylogram based on COI-5P dataset showed a
106 R. V. DUMILAG AND Z.-Z. A. AGUINALDO
distinct lineage for Py. acanthophora with polyto-mous internodes, representing conspecific samplesfrom the Philippines, Taiwan, Hong Kong, India,Hawaii and Brazil (Fig. 1). All Philippine Py.acanthophora specimens (n = 50) nested withinconspecific materials from Brazil, India, Japan,Taiwan and Hong Kong with strong support.Although most of the specimens of Philippine Py.acanthophora formed a clade with those conspecificmaterials collected from different regions, therewere clades that included only Philippine specimens(C11 and C12). The inferred phylogenetic treebased on rbcL dataset showed several lineages forPhilippine Py. acanthophora specimens (Fig. 2).Contrary to the result of the COI-5P dataset, therbcL phylogram indicated that those Philippine Py.
acanthophora materials did not group with any ofthose conspecific specimens collected from otherlocations.
Haplotype analysis
TCS resolved well-defined networks based on similardatasets used in phylogenetic analysis excludingrelated Pyropia and outgroup species (Bangia fusco-purpurea). Twelve COI-5P haplotypes (n=84) weredefined in a star-like network and lacked apparentgeographic clustering. There were five haplotypesobserved that represented the Philippine specimens(Fig. 3). The central haplotype C8, found in theintroduced Py. acanthophora from Brazil, wasdetected in 27 samples from the Philippines and
Table 2. Summary of sampling locations and haplotype information of Philippine Py. acanthophora specimens used in thisstudy.
Site Abbreviation Collecting site (Coordinates) Isolate
Haplotype Genbank Accession Number
rbcL COI-5P rbcL COI-5P
APA1 Brgy. San Antonio, Aparri, Cagayan(18°35’97” N, 121°64’12” E)
APA1.1 R5 C11 KU234722 KX036865APA1.2 R6 C11 KU234723 KX036866APA1.3 R10 C11 KU234724 KX036867APA1.4 R7 C11 KU234725 KX036868APA1.5 R20 C8 KU234726 KX036869APA1.6 R10 C11 KU234727 KX036870
APA2 Brgy. San Antonio-Punta Boundary,Aparri, Cagayan (18°36’08” N,121°63’65” E)
APA2.1 R10 C11 KU234728 KX036871APA2.2 R10 C11 KU234729 KX036872APA2.3 R10 C11 KU234730 KX036873
APA3 Brgy. Punta (Port Aparri,) Aparri,Cagayan (18°36’29’’ N, 121°62’86”E)
APA3.1 R22 C11 KU234731 KX036874APA3.2 R10 C11 KU234732 KX036875APA3.3 R10 C11 KU234733 KX036876
BUR Brgy. Saoit, Burgos, Ilocos Norte(18°53’82” N, 120°65’74’’ E)
BUR1 R11 C8 KU234734 KX036877BUR2 R11 C8 KU234735 KX036878BUR3 R11 C8 KU234736 KX036879
CAM Sitio Minabel, Camiguin Is.,Cagayan (19°00’23” N, 121°94’48”E)
CAM1 R8 C8 KU234737 KX036880CAM2 R5 C8 KU234738 KX036881CAM3 R5 C8 KU234739 KX036882CAM4 R5 C8 KU234740 KX036883CAM5 R5 C8 KU234741 KX036884CAM6 R5 C8 KU234742 KX036885
DLE De Leaño, Claveria, Cagayan (18°62’16” N, 121°09’34” E)
DLE1 R9 C4 KU234743 KX036886DLE2 R9 C4 KU234744 KX036887
MIN Mingay, Brgy. San Juan, Sta.Praxedes, Cagayan (18°59’86” N,120°99’09” E)
MIN1 R21 C12 KU234745 KX036888MIN2 R19 C4 KU234746 KX036889MIN3 R19 C4 KU234747 KX036890
NAN Sitio Nanaplaan, Brgy. San Juan, Sta.Praxedes, Cagayan (18°59’92” N,120°99’00” E)
NAN1 R19 C4 KU234748 KX036891NAN2 R19 C4 KU234749 KX036892NAN3 R19 C4 KU234750 KX036893
PAM Pamoctan Is., Camiguin, Cagayan(18°89’98” N, 121°83’82” E)
PAM1 R16 C8 KU234751 KX036894PAM2 R16 C3 KU234752 KX036895PAM3 R16 C8 KU234753 KX036896PAM4 R16 C8 KU234754 KX036897PAM5 R17 C8 KU234755 KX036898PAM6 R17 C8 KU234756 KX036899PAM7 R16 C8 KU234757 KX036900PAM8 R16 C8 KU234758 KX036901PAM9 R18 C8 KU234759 KX036902
PAG Pagudpud, Ilocos Norte (18°61’46”N, 120°86’96’’ E)
PAG1 R11 C8 KU234760 KX036903PAG2 R11 C8 KU234761 KX036904PAG3 R11 C8 KU234762 KX036905
PAT Sitio Malolog, Patunungan, Sta. Ana,Cagayan (18°44’41” N, 122°25’92”E)
PAT1 R14 C8 KU234763 KX036906PAT2 R14 C8 KU234764 KX036907PAT3 R15 C8 KU234765 KX036908
PEA Pata East, Claveria, Cagayan (18°62’52” N, 121°09’66” E)
PEA1 R13 C8 KU234766 KX036909PEA2 R13 C8 KU234767 KX036910PEA3 R23 C8 KU234768 KX036911
TAG Taggat, Claveria, Cagayan (18°61’35”N, 121°04’79” E)
TAG1 R12 C4 KU234769 KX036912TAG2 R12 C4 KU234770 KX036913TAG3 R5 C4 KU234771 KX036914
EUROPEAN JOURNAL OF PHYCOLOGY 107

from those previously collected materials fromTaiwan, Hong Kong and Hawaii. Haplotypes C3and C4 were found from the Philippines andTaiwan. The haplotype C12 was private, representedby a single specimen from Mingay, Sta. Praxedes,Cagayan, Philippines. Private haplotypes were alsorecognized from the specimens collected from Brazil(haplotype C7, presumably an introduced strain) andIndia (haplotype C10).
The minimum spanning tree of rbcL haplotypesshowed a total number of 23 Py. acanthophora
haplotypes from 68 conspecific sequences (Fig. 4).The 50 specimens collected from the Philippinesconstituted 19 haplotypes with 15 private haplo-types representing site-specific distribution. Theintroduced Py. acanthophora rbcL haplotype R1from Brazil was undetected from the Philippinesamples but recognized from materials previouslysequenced from Japan and Taiwan. One privatehaplotype each was observed from Taiwan (haplo-type R2), Hong Kong (haplotype R3) and India(haplotype R4).
Fig. 1. Phylogenetic tree based on Bayesian Inference (BI) analysis using COI-5P gene sequences. C1-12 represents thehaplotype clades. The numbers at the nodes indicate posterior probabilities support values.
108 R. V. DUMILAG AND Z.-Z. A. AGUINALDO

The haplotype network for the combined Py.acanthophora COI-5P-rbcL (referred to herein ascombined organellar genes) dataset among endemicpopulations in Indo-Pacific region revealed 31 dis-tinct sequences including 22 haplotypes for thePhilippines, seven for Taiwan and a single haplotypeeach for Hong Kong and India (Fig. 5). Py. acantho-phora populations from India (haplotype I1) andHong Kong (haplotype H1) were closer to thosefrom Taiwanese Py. acanthophora haplotypes T6
and T1 by a single to two mutation step distances.The Philippine haplotype P1 was separated fromTaiwanese haplotype T7 by two mutational steps.
The distribution of haplotypes for the combinedorganellar gene dataset is shown in Figs 6 and 7.Although most of the Philippine Py. acanthophorapopulations appeared specific to particular site, thedistribution of private haplotypes within the northernPhilippine region was also evident albeit observedonly in limited areas. Despite the high number of
Fig. 2. Phylogenetic tree based on Bayesian Inference (BI) analysis using rbcL gene sequences. R1-20 represents thehaplotype clades. The numbers at the nodes indicate posterior probabilities support values.
EUROPEAN JOURNAL OF PHYCOLOGY 109

private haplotypes in the combined organellar genes,no geographic structure within the Philippines wasobserved.
Genetic differentiation and neutrality test
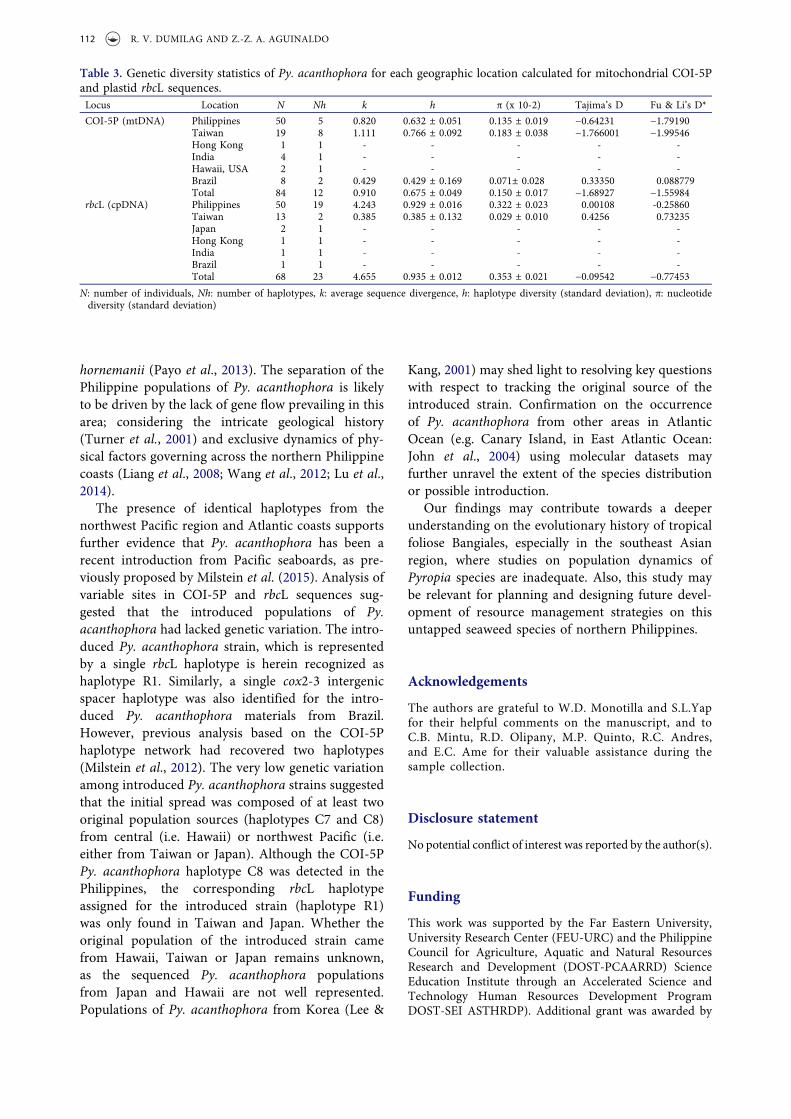
To analyse genetic diversity of Philippine Py.acanthophora, computation for haplotype diversity(h), nucleotide diversity (π) and average sequencedivergence (k) were performed using DNAsp ver.5(Table 3). For all the datasets, the Philippine Py.acanthophora populations showed high haplotype(h) but low nucleotide (π) diversities. The averagesequence divergence (k) was relatively higheramong the Philippine samples than any otherpopulation. As expected, the introduced popula-tions of Py. acanthophora in Brazil representedrelatively the lowest h, π and k values. Over all,the rbcL dataset had higher h, π, and k values thanthe COI-5P dataset. All values computed forTajima’s D and Fu & Li’s D* neutrality tests wereinsignificant.
Discussion
We confirmed a significant increase in the number ofknown populations and distribution ranges of Py.acanthophora in the Philippines, occurring acrossnorthwestern (Ilocos Norte) to northeastern main-land Luzon (Sta. Ana, Cagayan including Pamoctanand Camiguin Islands), based on COI-5P and rbcLsequences. While the COI-5P Py. acanthophora hap-lotypes from the Philippines were also dominantlyfound in Taiwan, the rbcL and the combined orga-nellar haplotype network analyses showed clearer dis-crimination of Philippine Py. acanthophorapopulations from those conspecific populationsfrom other known geographic areas (e.g. Brazil,Hong Kong, India, Japan and Taiwan).
Surprisingly, the rbcL sequences of Philippine Py.acanthophora specimens showed higher variabilitythan their corresponding COI-5P sequences.Although the common range of rbcL divergence infoliose Bangiales is up to 0.4% (Nelson & Broom,2010; Mols-Mortensen et al., 2012; Kucera &Saunders, 2012; Lindstrom et al., 2015), higher levelssuch as 0.7% (Guillemin et al., 2016) and 1%(Lindstrom, 2008) have also been reported. The intras-pecific sequence divergences of COI-5P gene in manyrhodophytes are commonly higher than rbcL gene (e.g.Geraldino et al., 2006; Yang et al., 2008; Freshwateret al., 2010; Tan et al., 2012). However, greater rbcLintraspecies variability (up to 2.1%) might be evidentlydetected especially on ‘centre of diversity’ sites(McIvor et al., 2001). Therefore, the presence of richergenetic diversity among endemic populations of Py.acanthophora in the Philippines, and in Taiwan (Xieet al., 2005) may also suggest that the west Pacificseaboard might be the centre of Py. acanthophoradiversity rather than other Indo-Pacific areas.
In all the datasets we used, the haplotype analysisand those computed values that do not significantlydiffer from zero based on neutrality tests indicatedthat there exist distinct and demographically stablepopulations of Py. acanthophora in the Philippines.The greater number of site-specific haplotypes bear-ing high genetic variability and the absence of geo-graphic patterns observed in Philippine Py.acanthophora populations are typical characteristicsof a ‘chaotic patchiness’ pattern (Johnson & Black1982). Although populations that demonstrate chao-tic patchiness are more common among marineinvertebrates (e.g. Johnson & Black, 1982; Arnaud-Haond et al., 2008; Muths et al., 2009; Fernándezet al., 2014), some indications of this type of geneticstructure especially at small geographic scales, havealso been observed for other algal species: Lessonianigrescens (Martínez et al., 2003), Mazaella laminar-ioides (Faugeron et al., 2001) and Portieria
Figs 3–4. Statistical parsimony analysis representing theentire Py. acanthophora haplotypes inferred from Fig. 3COI-5P and Fig. 4 rbcL using TCS. The size of the circlesrepresents the number of individuals; black dots correspondto missing haplotypes.
110 R. V. DUMILAG AND Z.-Z. A. AGUINALDO
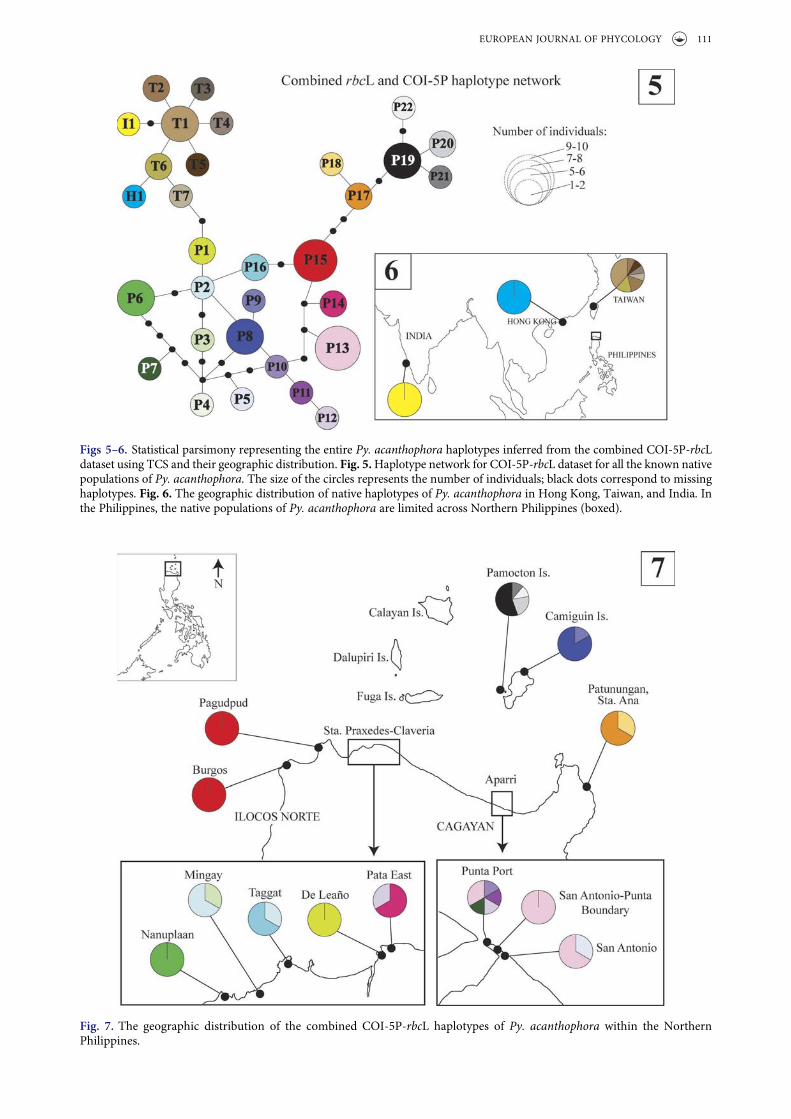
Figs 5 –6. Statistical parsimony representing the entire Py. acanthophora haplotypes inferred from the combined COI-5P-rbcLdataset using TCS and their geographic distribution. Fig. 5.Haplotype network for COI-5P-rbcL dataset for all the known nativepopulations of Py. acanthophora. The size of the circles represents the number of individuals; black dots correspond to missinghaplotypes. Fig. 6. The geographic distribution of native haplotypes of Py. acanthophora in Hong Kong, Taiwan, and India. Inthe Philippines, the native populations of Py. acanthophora are limited across Northern Philippines (boxed).
Fig. 7. The geographic distribution of the combined COI-5P-rbcL haplotypes of Py. acanthophora within the NorthernPhilippines.
EUROPEAN JOURNAL OF PHYCOLOGY 111
hornemanii (Payo et al., 2013). The separation of thePhilippine populations of Py. acanthophora is likelyto be driven by the lack of gene flow prevailing in thisarea; considering the intricate geological history(Turner et al., 2001) and exclusive dynamics of phy-sical factors governing across the northern Philippinecoasts (Liang et al., 2008; Wang et al., 2012; Lu et al.,2014).
The presence of identical haplotypes from thenorthwest Pacific region and Atlantic coasts supportsfurther evidence that Py. acanthophora has been arecent introduction from Pacific seaboards, as pre-viously proposed by Milstein et al. (2015). Analysis ofvariable sites in COI-5P and rbcL sequences sug-gested that the introduced populations of Py.acanthophora had lacked genetic variation. The intro-duced Py. acanthophora strain, which is representedby a single rbcL haplotype is herein recognized ashaplotype R1. Similarly, a single cox2-3 intergenicspacer haplotype was also identified for the intro-duced Py. acanthophora materials from Brazil.However, previous analysis based on the COI-5Phaplotype network had recovered two haplotypes(Milstein et al., 2012). The very low genetic variationamong introduced Py. acanthophora strains suggestedthat the initial spread was composed of at least twooriginal population sources (haplotypes C7 and C8)from central (i.e. Hawaii) or northwest Pacific (i.e.either from Taiwan or Japan). Although the COI-5PPy. acanthophora haplotype C8 was detected in thePhilippines, the corresponding rbcL haplotypeassigned for the introduced strain (haplotype R1)was only found in Taiwan and Japan. Whether theoriginal population of the introduced strain camefrom Hawaii, Taiwan or Japan remains unknown,as the sequenced Py. acanthophora populationsfrom Japan and Hawaii are not well represented.Populations of Py. acanthophora from Korea (Lee &
Kang, 2001) may shed light to resolving key questionswith respect to tracking the original source of theintroduced strain. Confirmation on the occurrenceof Py. acanthophora from other areas in AtlanticOcean (e.g. Canary Island, in East Atlantic Ocean:John et al., 2004) using molecular datasets mayfurther unravel the extent of the species distributionor possible introduction.
Our findings may contribute towards a deeperunderstanding on the evolutionary history of tropicalfoliose Bangiales, especially in the southeast Asianregion, where studies on population dynamics ofPyropia species are inadequate. Also, this study maybe relevant for planning and designing future devel-opment of resource management strategies on thisuntapped seaweed species of northern Philippines.
Acknowledgements
The authors are grateful to W.D. Monotilla and S.L.Yapfor their helpful comments on the manuscript, and toC.B. Mintu, R.D. Olipany, M.P. Quinto, R.C. Andres,and E.C. Ame for their valuable assistance during thesample collection.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Funding
This work was supported by the Far Eastern University,University Research Center (FEU-URC) and the PhilippineCouncil for Agriculture, Aquatic and Natural ResourcesResearch and Development (DOST-PCAARRD) ScienceEducation Institute through an Accelerated Science andTechnology Human Resources Development ProgramDOST-SEI ASTHRDP). Additional grant was awarded by
Table 3. Genetic diversity statistics of Py. acanthophora for each geographic location calculated for mitochondrial COI-5Pand plastid rbcL sequences.Locus Location N Nh k h π (x 10-2) Tajima’s D Fu & Li’s D*
COI-5P (mtDNA) Philippines 50 5 0.820 0.632 ± 0.051 0.135 ± 0.019 −0.64231 −1.79190Taiwan 19 8 1.111 0.766 ± 0.092 0.183 ± 0.038 −1.766001 −1.99546Hong Kong 1 1 - - - - -India 4 1 - - - - -Hawaii, USA 2 1 - - - - -Brazil 8 2 0.429 0.429 ± 0.169 0.071± 0.028 0.33350 0.088779Total 84 12 0.910 0.675 ± 0.049 0.150 ± 0.017 −1.68927 −1.55984
rbcL (cpDNA) Philippines 50 19 4.243 0.929 ± 0.016 0.322 ± 0.023 0.00108 -0.25860Taiwan 13 2 0.385 0.385 ± 0.132 0.029 ± 0.010 0.4256 0.73235Japan 2 1 - - - - -Hong Kong 1 1 - - - - -India 1 1 - - - - -Brazil 1 1 - - - - -Total 68 23 4.655 0.935 ± 0.012 0.353 ± 0.021 −0.09542 −0.77453
N: number of individuals, Nh: number of haplotypes, k: average sequence divergence, h: haplotype diversity (standard deviation), π: nucleotidediversity (standard deviation)
112 R. V. DUMILAG AND Z.-Z. A. AGUINALDO

the International Association of Plant Taxonomy (IAPT)Research Grants Program in Plant Systematics to RVD.
Author Contributions
R.V. Dumilag: original concept, field work, laboratoryexperiments, molecular analysis, drafting and editingmanuscript; Z.A. Aguinaldo: field work, laboratory experi-ments, editing manuscript.
References
Aguilar-Rosas, R. & Aguilar-Rosas, L.E. (2003). El géneroPorphyra (Bangiales, Rhodophyta) en la costa Pacifico diMéxico. I. Porphyra suborbiculata Kjellman.Hydrobiologia, 13: 51–56.
Arnaud-Haond, S., Vonau, V., Rouxel, C., Bonhomme, F.,Prou, J., Goyard, E. & Boudry, P. (2008). Genetic struc-ture at different spatial scales in the pearl oyster(Pinctada margaritifera cumingii) in French Polynesianlagoons: beware of sampling strategy and genetic patchi-ness. Marine Biology, 155: 147–157.
Brodie, J., Hayes, P.K., Barker, G.L., Irvine, L.M. & Bartsch,I. (1998). A reappraisal of Porphyra and Bangia(Bangiales, Rhodophyta) in the northeast Atlanticbased on the rbcL-rbcS intergenic spacer. Journal ofPhycology, 34: 1069–1074.
Brodie, J., Bartsch, I., Neefus, C., Orfinidis, S., Bray, T. &Mathieson, A. (2007). New insights into the cryptic diversityof the North Atlantic-Mediterranean ‘Porphyra leucosticta’complex: P. olivii sp. nov. and P. rosengurttii (Bangiales,Rhodophyta). European Journal of Phycology, 42: 3–28.
Broom, J.R., Jones, W.A., Hill, D.F., Knight, G.A. & Nelson,W.A. (1999). Species recognition in New ZealandPorphyra using 18S rDNA sequencing. Journal ofApplied Phycology, 11: 421–428.
Broom, J.E., Nelson, W.A., Yarish, C., Jones, W.A., Aguilar-Rosas, R. & Aguilar-Rosas, L.E. (2002). A reassessment ofthe taxonomic status of Porphyra suborbiculata, Porphyracarolinensis and Porphyra lilliputiana (Bangiales,Rhodophyta) based on molecular and morphologicaldata. European Journal of Phycology, 37: 227–235.
Broom, J.E., Nelson, W.A., Farr, T.J., Phillips, L.E. &Clayton M. (2010). Relationships of the Porphyra(Bangiales, Rhodophyta) flora of the Falkland Islands: amolecular survey using rbcL and nSSU sequence data.Australian Systematic Botany, 23: 27–37.
Clement, M., Posada, D. & Crandall, K.A. (2000). TCS: acomputer program to estimate gene genealogies.Molecular Ecology, 9: 1657–1659.
Cordero, Jr., P.A. (1974). Phycological observations I:Genus Porphyra of the Philippines, its species and theiroccurrences. Bulletin of Japanese Society of Phycology, 22:134–142.
Creed, M., Fujii, M.T., Barreto, M.B. de B., Guimarães, S.M.P. de B., Cassano, V., Pereira, S.M.B., Carvalho, M. deF., de O. & Khader, S. (2010). Rhodophyceae. InCatálogo de plantas e fungos do Brasil Vol. 1 (Forzza,R.C., editor), 416–436. Andrea Jakobsson Estúdio,Instituto de Pesquisas Jardim Botânico do Rio deJaneiro, Rio de Janeiro.
Dumilag, R.V., Aguinaldo, Z-Z., Mintu, C.B., Quinto, M.P.,Ame, E.C., Andres, R.C., Monotilla, W. D. & Yap, S.L.(2016). Morphological and molecular confirmation ofthe occurrence of Pyropia tanegashimensis (Bangiales,
Rhodophyta) from Palaui Is., Sta. Ana, Cagayan,Philippines. Phytotaxa, 255: 83–90.
Faugeron, S., Valero, M., Destombe, C., Martínez, E.A. &Correa J.A. (2001). Hierarchical spatial structure anddiscriminant analysis of genetic diversity in the redalga Mazzaella laminarioides (Gigartinales,Rhodophyta). Journal of Phycology, 37: 705–716.
Fernández, R., Lemer, S., McIntyre, E. & Giribet, G. (2014).Comparative phylogeography and population geneticstructure of three widespread mollusc species in theMediterranean and near Atlantic. Marine Ecology, 36:701–715.
Freshwater, W.D., Tudor, K., O’Shaughnessy, K. &Wysor, B.(2010). DNA barcoding in the red algal order Gelidiales:comparison of COI with rbcL and verification of the “bar-coding gap”. Cryptogamie, Algologie, 31: 435–449.
Geraldino, P.J.L., Yang, E.C. & Boo, S.M. (2006). Morphologyand molecular phylogeny of Hypnea flexicaulis(Gigartinales, Rhodophyta) from Korea.Algae, 21: 417–423.
Griffin, N.J., Bolton, J.J. & Anderson, R.J. (1999). Porphyraaeodis sp. nov. (Bangiales, Rhodophyta), an epiphyte ofAeodes orbitosa from South Africa. European Journal ofPhycology, 34: 505–512.
Guillemin, M., Contreras-Porcia, L. Ramírez, M.E.,Macaya, E.C., Contador, C.B., Woods, H., Wyatt, C. &Brodie, J. (2016). The bladed Bangiales (Rhodophyta) ofthe South Eastern Pacific: molecular species delimitationreveals extensive diversity. Molecular Phylogenetics andEvolution, 94: 814–826.
Guiry, M.D. & Guiry, G.M. (2016). AlgaeBase. World-wideelectronic publication, National University of Ireland,Galway. http://www.algaebase.org; searched on 25 May2016.
Hanisak, M.D. (1998). Seaweed cultivation: global trends.World Aquaculture Magazine, 29: 18–21.
Harden, L.K., Morales, K.M. & Hughey, J.R. (2016).Identification of a new marine algal species Pyropianitida sp. nov. (Bangiales: Rhodophyta) fromMonterey, California. Mitochondrial DNA Part A: DNAMapping, Sequencing, and Analysis, 4: 3058–3062.
Haroun, R.J., Gil-Rodriguez, M.C., Díaz De Castro J. &Prud’Homme van Reine, W.F. (2002). A checklist ofthe marine plants from Canary Islands (Central EasternAtlantic Ocean). Botanica Marina, 45: 139–169.
Hebert, P.D.N., Cywinska, A., Ball, S.L. & deWaard, J.R.(2003). Biological identifications through DNA bar-codes. Proceedings of the Royal Society B: BiologicalSciences, 270: 313–322.
Humm, H. J. (1979). The Marine Algae of Virginia, SpecialPapers in Marine Science No. 3. University of Virginia,Charlottesville.
Istini, S., Zatnika, A. & Sujatmiko, W. (1998). The seaweedresources of Indonesia. In Seaweed Resources of theWorld (Critchley, A.T. & Ohno, M., editors), 92–98.Kanagawa International Fisheries Training Centre,Japan International Agency.
John, D.M., Prud’homme van Reine, W.F., Lawson, G.W.,Kostermans, T.B. & Price, J.H. (2004). A taxonomic andgeographical catalogue of the seaweeds of the westerncoast of Africa and adjacent islands. Beihefte zur NovaHedwigia, 127: 1–339.
Johnson, M.S. & Black, R. (1982). Chaotic genetic patchi-ness in an inter-tidal limpet, Siphonaria sp. MarineBiology, 70: 157–164.
Jones, W.A., Griffin, N.J., Jones, D.T., Nelson, W.A., Farr,T.J. & Broom, J.E. (2004). Phylogenetic diversity inSouth African Porphyra (Bangiales, Rhodophyta)
EUROPEAN JOURNAL OF PHYCOLOGY 113

determined by nuclear SSU sequence analysis. EuropeanJournal of Phycology, 39: 197–211.
Kavale, M.G., Kazi, M.A. & Sreenadhan, N. (2015a). Pyropiaacanthophora var. robusta (Bangiales, Rhodophyta) fromGoa, India. Indian Journal of Geo-Marine Sciences, 44: 1–8.
Kavale, M.G., Kazi, M.A., Sreenadhan, N. & Singh, V.V.(2015b).Morphological, ecological andmolecular character-ization of Pyropia vietnamensis (Bangiales, Rhodophyta)from the Konkan region, India. Phytotaxa, 224: 45–58.
Kucera, H. & Saunders, G.W. (2012). A survey of Bangiales(Rhodophyta) based on multiple molecular markersreveals cryptic diversity. Journal of Phycology, 48: 869–882.
Kunimoto, M., Kito, H., Kaminishi, Y., Mizukami, Y. &Murase, N. (1999). Molecular divergence of the SSUrRNA gene and internal transcribed spacer 1 inPorphyra yezoensis (Rhodophyta). Journal of AppliedPhycology, 11: 211–216.
Lee, Y. & Kang, S. (2001). A catalogue of the seaweeds inKorea. Cheju National University Press, Jeju, Korea.
Levring, T. (1953). The marine algae of Australia. I.Rhodophyta: Goniotrichales, Bangiales andNemalionales. Arkiv för Botanik, Uppsala, 2: 457–530.
Lewmanomont, K. (1998). The seaweed resources ofThailand. In Seaweed Resources of the World (Critchley,A.T. & Ohno, M., editors), 70–78. Kanagawa InternationalFisheries Training Centre, Japan International Agency.
Liang, W.-D., Yang, Y.J., Tang, T.Y. & Chuang, W.-S.(2008). Kuroshio in the Luzon Strait. Journal ofGeophysical Research-Oceans, 113: C08048, 1–19.
Librado, P. & Rozas, J. (2009). DnaSP v5: a software forcomprehensive analysis of DNA polymorphism data.Bioinformatics, 25: 1451–1452.
Lindstrom, S.C. (2008). Cryptic diversity, biogeographyand genetic variation in Northeast Pacific species ofPorphyra sensu lato (Bangiales, Rhodophyta). Journalof Applied Phycology, 20: 951–962.
Lindstrom, S.C., Hughey, J.R. & Aguilar Rosas, L.E. (2015).Four new species of Pyropia (Bangiales, Rhodophyta)from the west coasts of North America: the Pyropialanceolata species complex updated. PhytoKeys, 52: 1–22.
Lu, W., Yan, X.-H. & Jiang, Y. (2014). Winter bloom andassociated upwelling northwest of the Luzon Island: acoupled physical-biological modeling approach. Journalof Geophysical Research-Oceans, 120: 533–546.
Martínez, E.A., Cárdenas, L. & Pinto, R. (2003). Recoveryand genetic diversity of the intertidal kelp Lessonianigrescens (Phaeophyceae) 20 years after El Niño 1982/83. Journal of Phycology, 39: 504–508
Masuda, M., Ohno, M. & Trono, G.C. Jr. (1991). A taxo-nomic assessment of Porphyra suborbiculata Kjellman, afood species from the Philippines. Japan Journal ofPhycology, 39: 375–380.
Mateo-Cid, L.E., Mendoza-Gonzalez, A.C., Díaz-Larrea, J.,Sentíes, A., Pedroche, P.F. & Sánchez Heredia, J.D.(2012). A new species of Pyropia (Rhodophyta,Bangiaceae), from the Pacific coast of Mexico, based onmorphological and molecular evidence. Phytotaxa, 54:1–12.
McIvor, L., Maggs, C.A., Provan, J. & Stanhope, M.J.(2001). rbcL sequences reveal multiple cryptic introduc-tions of the Japanese red alga Polysiphonia harveyi.Molecular Ecology, 10: 911–919.
Milstein, D. & Oliveira, M.C. (2005). Molecular phylogenyof Bangiales (Rhodophyta) based on small subunitrDNA sequencing: emphasis on Brazilian Porphyra spe-cies. Phycologia, 44: 212–221.
Milstein, D., Medeiros, A., Oliveira, E.C. & Oliveira, M.C.(2012). Will a DNA barcoding approach be useful toidentify Porphyra species (Bangiales, Rhodophyta)? Acase study with Brazilian taxa. Journal of AppliedPhycology, 24: 837–845.
Milstein, D., Madeiros, A.S., Oliveira, E.C. & Oliveira, M.C.(2015). Native or introduced? A re-evaluation of Pyropiaspecies (Bangiales, Rhodophyta) from Brazil based onmolecular analysis. European Journal of Phycology, 50:37–45.
Mols-Mortensen A., Neefus, C.D., Nielsen, R., GunnarssonK., Egilsdóttir, Pedersen P.M. & Brodie J. (2012). Newinsights into the biodiversity and genetic relationships offoliose Bangiales (Rhodophyta) in Iceland and the FaroeIslands. European Journal of Phycology, 47: 146–159.
Mols-Mortensen A., Neefus, C.D., Pedersen P.M. & Brodie,J. (2014). Diversity and distribution of foliose Bangiales(Rhodophyta) in West Greenland: a link between NorthAtlantic and North Pacific. European Journal ofPhycology, 49: 1–10.
Mumford, T.F. Jr. & Miura, A. (1998). Porphyra as food:cultivation and economics. In Algae and Human Affairs(Lembi, C.A. & Waaland, J.R., editors), 87–117.Cambridge University Press, Cambridge.
Muths, D., Jollivet, D., Gentil, F. & Davoult, D. (2009).Large-scale genetic patchiness among NE Atlantic popu-lations of the brittle star Ophiothrix fragilis. AquaticBiology, 5: 117–132.
Neefus, C.D., Mathieson, A.C., Bray, T.L. & Yarish, C.(2008). The distribution, morphology, and ecology ofthree introduced Asiatic Porphyra (Bangiales,Rhodophyta) in the northwestern Atlantic. Journal ofPhycology, 44: 1399–1414.
Nelson, W.A. & Broom, J.E.S. (2010). The identity ofPorphyra columbina (Bangiales, Rhodophyta) originallydescribed from the New Zealand subantarctic islands.Australian Systematic Botany, 23: 16–26.
Nelson, W.A. & D’Archino, R. (2014). Three new macro-algae from the Three Kings Islands New Zealand includ-ing the first southern Pacific Ocean record of theFurcellariaceae (Rhodophyta). Phycologia, 53: 602–613.
Nelson, W.A., Sutherland, J.E., Hwang, M.S. & Choi, H.-G.(2014). New distributional record for Pyropia koreana:confirmed to occur on the South Island, New Zealand.Algae, 29: 177–181.
Ogawa, H. (2001). Porphyra in South Asian Waters. InProceedings of the 11th JSPS Joint Seminar on MarineScience (Taira, M.A., Uematsu, M., Michida, Y. &Kaneko, T., editors), 94–101. Center for InternationalCooperation, Ocean Research Institute, University ofTokyo, Tokyo, Japan.
Oliveira Filho, E.C. de & Coll, J. (1975). The genusPorphyra C. Ag. (Rhodophyta, Bangiales) in theAmerican south Atlantic I. Brazilian species. BotanicaMarina, 18: 191–197.
Payo, D.A., Leliaert, F., Verbruggen, H., D’hondt, S. &Calumpong, H.P. (2013). Extensive cryptic species diver-sity and fine-scale endemism in the marine red algaPortieria in the Philippines. Proceedings of the RoyalSociety B, 280: 20122660.
Posada, D. (2008). jModelTest: phylogenetic model aver-aging. Molecular Biology and Evolution, 25: 1253–1256.
Quisumbing, E. (1951). Medicinal plants of the Philippines.Republic of the Philippines, Department of Agricultureand Natural Resources, Technical Bulletin, 16: 1–1234.
Ramírez, M.E., Contreras Porcia, L., Guillemin, M.-L.,Brodie, J., Valdivia, C., Flores Molina, M.R., Núñez, A.,
114 R. V. DUMILAG AND Z.-Z. A. AGUINALDO

Bulboa Contador, C. & Lovazzano, C. (2014). Pyropiaorbicularis sp. nov. (Rhodophyta, Bangiaceae) based on apopulation previously known as Porphyra columbinafrom the central coast of Chile. Phytotaxa, 158: 133–153.
Rivera, R.A. (2014). Physicochemical properties and qualityevaluation of Porphyra acanthophora in the Philippines.MSc. thesis. Pukyong National University.
Ronquist, F. & Huelsenbeck, J.P. (2003). MrBayes 3:Bayesian phylogenetic inference under mixed models.Bioinformatics, 19: 1572–1574.
Sánchez, N., Vergès, A., Peteiro, C., Sutherland, J. &Brodie, J. (2014). Diversity of bladed Bangiales(Rhodophyta) in western Mediterranean: recognition ofthe genus Themis and descriptions of T. ballesterosii sp.nov., T. iberica sp. nov., and Pyropia parva sp. nov.Journal of Phycology, 50: 908–929.
Saunders, G.W. (2005). Applying DNA barcoding to redmacroalgae: a preliminary appraisal holds promise forfuture applications. Philosophical Transactions of theRoyal Society London Biology, 360: 1879–1888.
Sherwood, A.R., Kurihara, A., Conklin, K.Y., Sauvage, T. &Presting, G.G. (2010). The Hawaiian Rhodophyta biodi-versity survey (2006–2010): a summary of principal find-ings. BMC Plant Biology, 10: 258.
Soe-Htun, U. & Zaw Zaw Pe, U. (1986). Studies on thecultivation of conchocelis filaments in Porphyra crispataKjellman. Burma Research Congress.
Sutherland, J.E., Lindstrom, S.C., Nelson, W.A., Brodie, J.,Lynch, M.D.J., Hwang, M.S., Choi, H., Miyata, M.,Kikuchi, N., Oliveira, M.C., Farr, T., Neefus, C., Mols-Mortensen, A., Milstein, D. & Müller, K.M. (2011). A newlook at an ancient order: generic revision of the Bangiales(Rhodophyta). Journal of Phycology, 47: 1131–1151.
Tamura, K., Peterson. D., Peterson. N., Stecher, G., Nei. M.& Kumar. S. (2011). MEGA.5: molecular evolutionarygenetics analysis using maximum likelihood, evolution-ary distance, and maximum parsimony methods.Molecular Biology and Evolution, 28: 2731–2739.
Tan, J., Lim, P.-E., Phang, S.-M., Hong, D.D., Sunarpi, H.& Hurtado, A.Q. (2012). Assessment of four molecularmarkers as potential DNA barcodes for red algaeKappaphycus Doty and Eucheuma J. Agardh(Solieriaceae, Rhodophyta). PLoS ONE, 7: e52905.
Tanaka, T. & Pham-Hoàng, H. (1962). Notes on somemarine algae from Viet-Nam-I. Memoirs of the Facultyof Fisheries, Kagoshima University, 11: 24–40.
Teasdale, B., West, A., Taylor, H. & Klein, A. (2002). Asimple restriction fragment length polymorphism(RFLP) assay to discriminate common Porphyra(Bangiophyceae, Rhodophyta) taxa from NorthwestAtlantic. Journal of Applied Phycology, 14: 293–298.
Thiers, B. (2016). Index Herbariorum: A global directory ofpublic herbaria and associated staff. New York BotanicalGarden’s Virtual Herbarium. http://sweetgum.nybg.org/science/ih/.
Thompson, J.D., Higgins, D.J. & Gibson, T.J. (1994).CLUSTALW: Improving the sensitivity of progressivemultiple sequence alignment through sequence weight-ing, position specific gap penalties and weight matrixchoice. Nucleic Acids Research, 22: 4673–4680.
Torrano-Silva, B.N., Amancio, C.E. & Oliviera, E.C. (2010).Exotic marine macroalgae on Brazilian coast: a revision.Oecologia Australis, 14: 403–414.
Trono, G.C. Jr. & Ganzon-Fortes, E.T. (1988). PhilippineSeaweeds. National Book Store, Manila.
Turner, H., Hovenkamp, P. & van Welzen, P.C. (2001).Biogeography of Southeast Asia and the West Pacific.Journal of Biogeography, 28: 217–230.
Vergés, A., Sánchez, N., Peteiro, C., Polo, L. & Brodie, J.(2013a). Pyropia suborbiculata (Bangiales, Rhodophyta):first records from the northeastern Atlantic andMediterranean of this North Pacific species. Phycologia, 52:121–129.
Vergés, A., Comalada, N., Sánchez, N. & Brodie, J. (2013b).A reassessment of the foliose Bangiales (Rhodophyta) inthe Balearic Islands including the proposed synonymy ofPyropia olivii with Pyropia koreana. Botanica Marina,56: 229–240.
Verlaque, M. (2001). Checklist of macroalgae of ThauLagoon (Hérault, France), a hot spot of marine speciesintroduction in Europe. Oceanologica Acta, 24: 291–249.
Wang, G., Wang, D. & Zhou, T. (2012). Upper layercirculation in the Luzon Strait. Aquatic EcosystemHealth and Management, 15: 39–45.
Watson, K., Cheney, D. & Levine, I. (1998). Can the aqua-cultured non-indigenous red alga Porphyra yezoensisrecruit in Eastport, Maine? Journal of Phycology, 34(Suppl.): 62.
Watson, K., Levine, I. & Cheney, D. (1999). Biomonitoringof an aquacultured introduced seaweed, Porphyra yezoen-sis (Rhodophyta, Bangiophycidae) in Cobscook Bay,Maine, USA. In Marine Invasions (Pederson, J., editor),260–264. Proceedings of the First National Conference,January 24–27, 1999, Massachusetts Institute ofTechnology Sea Grant College Program, Cambridge, MA.
West, A.L., Mathieson, A.C., Klein, A.S., Neefus, C.D. &Bray, T.L. (2005). Molecular ecological studies of NewEngland species of Porphyra (Rhodophyta, Bangiales).Beihefte zur Nova Hedwigia, 80: 1–24.
Wynne, M.J. (2005). A check-list of benthic marine algae ofthe tropical and subtropical western Atlantic: secondrevision. Beihefte zur Nova Hedwigia, 129: 1–152.
Xie, Z., Lin, S., Liu, L., Ang, P.O. Jr & Shyu, J. (2015).Genetic diversity and taxonomy of foliose Bangiales(Rhodophyta) from Taiwan based on rbcL and cox1sequences. Botanica Marina, 58: 189–2012.
Yang, E.C., Kim, M.S., Geraldino, P.J., Sahoo, D.,Shin, J.-A. & Boo, S.M. (2008). Mitochondrialcox1 and rbcL genes of Gracilaria vermiculophylla(Gracilariaceae, Rhodophyta). Journal of AppliedPhycology, 20: 161–168.
Yarish, C.R., Wilkes, R., Chopin, T., Fei, X.G.,Mathieson, A.C., Klein, A.S., Neefus, C.D., Mitman,G.G. & Levine, I. (1998). Domestication of indigen-ous Porphyra (nori) species for commercial cultiva-tion in Northeastern America. World Aquaculture,29: 26–29.
Yoshida, T. (1997). Japanese marine algae: new combina-tions, new names and new species. PhycologicalResearch, 45: 163–167.
Yoshida, T. (1998). Marine Algae of Japan. UchidaRokakuho Publishing Co. Ltd, Tokyo.
Yoshida, T., Nakajima, Y. & Nakata, Y. (1990). Check-listof marine algae of Japan (revised in 1990). JapaneseJournal of Phycology, 38: 269–320.
EUROPEAN JOURNAL OF PHYCOLOGY 115
View publication statsView publication stats





























