Embed Size (px)
Citation preview

1
GENETIC AND MOLECULAR ANALYSIS OF PATHOGENICITY GENES IN Xanthomonas citri subsp. citri
By
JOSÉ FRANCISCO LISSONI FIGUEIREDO
A DISSERTATION PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT
OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY
UNIVERSITY OF FLORIDA
2009

2
© 2009 José Francisco Lissoni Figueiredo

3
To my wife, Tania and my kids, João Pedro and Antonio, for the love and support allowing me to accomplish my dreams

4
ACKNOWLEDGMENTS
I would like extend my gratitude to Dr. Jeffrey B. Jones, who served as my co-advisor. I
am really grateful to him to accept as student in his laboratory. I appreciate all the unconditional
support and encouragement during my graduate studies. I am also grateful to Dr. James H.
Graham, who served as my committee chair and for providing the financial support for all these
years. I really thank Dr. Frank F. White for invited me to work with him at Kansas State
University and to give me the opportunity to start my Ph.D. studies in the U.S.A. I also
appreciate the valuable help from the members of my committee; Dr. Robert E. Stall for provide
plant material and support with the plant assays, and Dr. James Preston for reviewing the
manuscript. I would like to thank Jerry Minsavage for all the time spent to teach me new
techniques, discussing all the problems with this study and for his unconditional help and support
with my research. Without him, this research would not have been completed.
I thank the faculty and staff of the Department of Plant Pathology; they were very
supportive since I started my program here. For all my friends at the plant pathology departments
at University of Florida and Kansas State University that helped me with valuable discussions
and friendship, I would like to give a hearty "Muito Obrigado".
I especially thank my wife for her patience and courage in all these years. This dissertation
is also dedicated to my kids, who were always waiting for me with a smile on their face. I am
thank to my precious family. My father, mother, sister, brother, nephews and my loved uncle for
all the love along these years. Even as I am so far away, they never doubted my capacity.
Last but not least, I thank all my fellow Brasileiros who live here in U.S.A. or in Brazil for
their sympathy, support and friendship.

5
TABLE OF CONTENTS
page
ACKNOWLEDGMENTS.................................................................................................................... 4
LIST OF TABLES................................................................................................................................ 8
LIST OF FIGURES .............................................................................................................................. 9
ABSTRACT ........................................................................................................................................ 11
CHAPTER
1 THE WEAPONRY INVOLVED ON PLANT-PATHOGEN INTERACTION .................... 13
Introduction ................................................................................................................................. 13 Plant Pathogen Translocon Pathways ................................................................................. 14
Type I secretion system ............................................................................................... 14 Type II secretion system .............................................................................................. 15 Type III secretion system............................................................................................. 16 Type IV secretion system ............................................................................................ 18
Hypersensitive Response and Pathogenicity (hrp) Genes ................................................ 19 Harpins a HR elicitor protein ...................................................................................... 22
Type Three Secretion System-Dependent Effectors ......................................................... 23 Project Goals and Objectives...................................................................................................... 25
2 MUTATIONAL ANALYSIS OF TYPE III EFFECTORS GENES FROM Xanthomonas citri subsp. citri.................................................................................................... 27
Introduction ................................................................................................................................. 27 Materials and Methods ................................................................................................................ 28
Bacterial Strains and Plasmids ............................................................................................ 28 Media and Growth Conditions ............................................................................................ 28 Recombinant DNA Techniques .......................................................................................... 29 DNA Amplification ............................................................................................................. 30 Mutation of Candidate Genes by Homologous Recombination ....................................... 30 Complementation of ΩhrpG and ΩhrpX Deficient Mutants of Xcc Strain 306 .............. 31 Plant Material and Inoculations .......................................................................................... 32
Results .......................................................................................................................................... 33 Analysis of HrpX-Regulon Candidate Genes .................................................................... 33 Mutations of the regulatory and structural genes of Xcc 306 hrp pathway cause loss
of pathogenicity ................................................................................................................ 33 Phenotypic Analysis of Nineteen Candidate Effector and Hrp-Regulon Associated
Genes. ............................................................................................................................... 34 Discussion .................................................................................................................................... 34

6
3 IDENTIFICATION OF A HRP-INDEPENDENT HR ELICITOR FROM Xanthomonas citri subsp. citri............................................................................................................................ 50
Introduction ................................................................................................................................. 50 Materials and Methods ................................................................................................................ 52
Bacterial Strains and Plasmids ............................................................................................ 52 Media and Growth Conditions ............................................................................................ 53 Recombinant DNA Techniques .......................................................................................... 53 DNA Amplification ............................................................................................................. 54 Mutagenesis of hrpG and hrpX in X. citri subsp. citri 306 ............................................... 54 Subcloning of HR-Elicitor Candidate from Xaa Library .................................................. 55 Plant Material and Plant Inoculations................................................................................. 57 Measurement of Electrolyte Leakage ................................................................................. 58 Protein Thermal Stability .................................................................................................... 58
Results .......................................................................................................................................... 58 Unusual Incompatible Responses of Mutants 306::ΩhrpG and 306::ΩhrpX in
Tomato .............................................................................................................................. 58 Subclone of X. f. pv. aurantifolli Confers the Ablilty to Trigger an HR in Tomato ....... 59 Sequencing Analysis Revealed Novel Genes Controlling Nonhost HR Elicitor ............ 60
Discussion .................................................................................................................................... 62
4 GENETIC CHARACTERIZATION OF HRPW OF Xanthomonas citri subsp. citri ............ 76
Introduction ................................................................................................................................. 76 Material and Methods ................................................................................................................. 78
Bacterial Strains and Plasmids ............................................................................................ 78 Media and Growth Conditions ............................................................................................ 78 Recombinant DNA Techniques .......................................................................................... 78 DNA Sequence Alignment and Phylogenetic Analysis of hrpW Gene............................ 79 Generation of hrpW Deletion Mutants ............................................................................... 80 Construction of hrpW-Harpin Domain ............................................................................... 81 Complementation Tests ....................................................................................................... 82 Plant Material ....................................................................................................................... 83 Plant Inoculations ................................................................................................................ 83 Bacterial Populations ........................................................................................................... 84 Translocation Activity in 306::∆harpin Mutant ................................................................. 84 Construction of hrpW::avBs2 Fusion ................................................................................. 85
Results .......................................................................................................................................... 85 Xanthomonas citri subsp. citri hrpW Phylogeny ............................................................... 85 Total Deletion of hrpW Gene Is Irrelevant for Xcc Pathogenicity and HR in Non-
Host Plants ........................................................................................................................ 86 Harpin Domain in hrpW Gene is Required for Pathogenicity .......................................... 87 Complementation of HrpW∆harpin Mutant In-Cis ........................................................... 88 Translocon Machinery, T3SS, Is Not Affected by ∆harpin Mutant in Xcc ..................... 88 HprW is not Translocated into the Mesophyll of Plant Cell ............................................. 89
Discussion .................................................................................................................................... 89

7
5 CHARACTERIZATION OF avrGf1 FROM Xanthomonas citri subsp. citri STRAIN AW .............................................................................................................................................. 117
Introduction ............................................................................................................................... 117 Materials and Methods .............................................................................................................. 118
Bacterial Strains and Plasmids .......................................................................................... 118 Media and Growth Conditions .......................................................................................... 118 Recombinant DNA Techniques ........................................................................................ 118 Plant Material and Plant Inoculations............................................................................... 119 Agrobacterium-mediated expression of avrGf1 . ............................................................ 120 Bacterial Expression of avrGf1......................................................................................... 122
Results ........................................................................................................................................ 123 Induction of Hypersensitive Response by Transient Expression of avrGf1 Within
Citrus ............................................................................................................................... 123 The N-terminal and C-terminal Are Required for HR Elicitation .................................. 124 Xanthomonas citri subsp. citri Harboring avrGf1 Induces a Hypersensitive
Reaction in Grapefruit ................................................................................................... 124 AvrGf1 Is Translocated Inside the Host Plant Cell ......................................................... 125
Discussion .................................................................................................................................. 125
6 OVERALL SUMMARY AND DISCUSSION ...................................................................... 135
APPENDIX: SEQUENCE AND ALIGNMENT .......................................................................... 139
LIST OF REFERENCES ................................................................................................................. 142
BIOGRAPHICAL SKETCH ........................................................................................................... 163

8
LIST OF TABLES
Table page 2-1 Xanthomonas citri pv. citri genes used in the study. ........................................................... 38
2-2 Bacterial strains and plasmids used in the study. ................................................................. 39
2-3 Oligonucleotides sequence used in this study. ..................................................................... 42
2-4 Candidate PIP box (-like) and -10 box-like sequences of the proposed hrpX regulons and putative effector/avirulence genes of Xcc. .................................................................... 44
2-5 Xanthomonas citri pv. citri mutants pathogenicity phenotype in grapefruit leaveas and HR elicitation in tomato leaflets..................................................................................... 45
3-1 Bacterial strains and plasmids used in the study. ................................................................. 64
4-1 Bacterial strains and plasmids used in the study. ................................................................. 93
4-2 Oligonucleotides sequence used in this study. ..................................................................... 95
5-1 Bacterial strains and plasmids used in the study. ............................................................... 128
5-2 Oligonucleotides sequence used in this study. ................................................................... 130

9
LIST OF FIGURES
Figure page 2-1 Genetic organization of the hrp genes of X. citri subsp. citri.............................................. 46
2-2 Proposed model for hrp gene regulation cascade in X. citri subsp. citri ............................ 47
2-3 Comparison of the bacterial pathogenicity and HR elicitor in grapefruit leaves and tomato leaflets, respectively .................................................................................................. 48
2-4 Integration by site-specific recombination using one single target sited ............................ 49
3-1 Phenotype of grapefruit leaves were used to distinguish among the strains ...................... 66
3-2 Phenotype of 306 wild type, 306::ΩhrpG and 306:: ΩhrpX in leaflet of tomato cv Bonny Best.............................................................................................................................. 67
3-3 Hypersensitive response assay in tomato cv. Bonny Best and pepper cv. ECW-20R ....... 68
3-4 Electrolyte leakage in tomato plant is changed in WT::pLAFR3, compatible interaction, and WT:: 450, incompatible interaction ........................................................... 69
3-5 Subclone, B38, triggers hypersensitive response (HR) independent of T3SS in X. perforans in non-host plants, tomato cv. Bonny Best and pepper cv. ECW20R ............... 70
3-6 Protein alignment of XAC3857 gene and Xanthomonas spp .............................................. 71
3-7 Protein alignment of XAC3858 gene and Xanthomonas spp .............................................. 72
3-8 Protein alignment of XAC3859 gene and Xanthomonas spp .............................................. 73
3-9 Transcriptional organization of the ORFs XAC3857 to XAC3859 region in Xanthomonas citri subsp. citri............................................................................................... 75
4-1 Neighbor joining analyses using full nucleotide sequence of hrpW gene from X. c. subsp. citri strain 306 ............................................................................................................. 96
4-2 Sequence alignment of Xcc strains collected world-wide ................................................... 97
4-3 Sequence alignment of Xcc strain 306 – wild type (W1); W::∆Harpin mutant (W2); and 306::∆hrpW HrpW (W3) .............................................................................................. 112
4-4 Phenotype assay in grapefruit leaves .................................................................................. 113
4-5 Populations of Xcc 306 (Xcc306) ....................................................................................... 114
4-6 Canker symptoms on abaxial leaf surface of grapefruit .................................................... 115

10
4-7 PCR amplification of genomic DNA of Xcc 306 .............................................................. 116
5-1 Diagram of AvrGf1 and truncated versions used to test regions of the protein for elicitor activity ...................................................................................................................... 131
5-2 Agrobacterium-mediated expression of avrGf1 in grapefruit leaf tissue ......................... 132
5-3 Hypersensitive reaction induced by X. citri subsp. citri 306 expressing avrGf1 ............. 133
5-4 Phenotypes elicited on pepper plants cv. ECW20R 24 h after inoculation ...................... 134
A-1 hrpW nucleotide sequence and deduced amino acid sequence. ........................................ 139
A-2 Nucleotide sequence of AvrGf11-106::AvrBs262-574 fused protein ..................................... 140
A-3 Nucleotide sequence of HrpW1-109::AvrBs262-574 fused protein ........................................ 141

11
Abstract of Dissertation Presented to the Graduate School of the University of Florida in Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy
GENETIC AND MOLECULAR ANALYSIS OF PATHOGENICITY GENES IN Xanthomonas
citri subsp. citri
By
José Francisco Lissoni Figueiredo
May 2009 Chair: James H. Graham Cochair: Jeffrey B. Jones Major: Plant Pathology
Xanthomonas citri subsp. citri (Xcc), which causes citrus canker, relies on a type III
secretion system (T3SS) to successfully develop disease. Various plant pathogenic bacteria
carrying a T3SS inject more than 40 different effector proteins (T3-effectors) into the plant cells
via this apparatus. In Xanthomonas, the genes for the T3SS are regulated by HrpX, which is an
AraC-type transcriptional regulator. The possible roles that nineteen candidate T3-effector genes
play in the ability of Xcc to cause citrus canker were investigated through the use of site-directed
mutagenesis. The candidate genes were selected on the basis of having promoter features similar
to gene regulated by HrpX or by sequence similarity to known T3-effectors in other plant
pathogenic bacteria. Inoculation in grapefruit revealed that none of these mutants were visually
impaired for disease development. Unlike the hrpW null mutant, deletion in the harpin domain
from hrpW resulted in the loss of Xcc pathogenicity symptoms, while not affecting the ability of
of the bacteria to multiply in the plant. The mutants were also assessed for the ability to elicit
hypersensitive response (HR) in non-host plant tomato. The Xcc hrp- mutants, in contrast to
many pathogenic xanthomonads, retained the ability to trigger HR in the nonhost. Using
subcloning procedures and homology search, three candidates open reading frames, XAC3857,

12
XAC3858 and XAC3859, were identified in Xcc that might be responsible for this T3SS-
independent HR elicitation. Experiments were also performed to further characterize a new
avirulence gene, avrGf1, isolated from Xcc strain Aw, which induces HR in grapefruit. The
avrGf1 gene was demonstrated to encode a protein that is translocated into plant cells via the
T3SS. Additionally, a transient expression on grapefruit leaves was devised using an
Agrobacterium-mediated delivery system. Different avrGf1 deletion mutants in the N- and C-
terminal coding regions were tested using the system, and the results showed that the first 116
amino acids in the N-terminal and the last 83 amino acids in the C-terminal were crucial for HR
elicitation in grapefruit. In summary, this study reported the presence of candidate T3-effector
genes that did not affect the disease progress under the experimental conditions and a harpin
domain of hrpW gene is essential for symptom development. Additionally, the development of
an efficient transient expression in grapefruit leaves revealed specific regions of avrGf1 are
required for defense activation.

13
CHAPTER 1 THE WEAPONRY INVOLVED ON PLANT-PATHOGEN INTERACTION
Introduction
Many complex steps are involved in the interaction of plants and their pathogens.
Although plants are exposed to attack by many bacteria, disease development is a relatively rare
event. Usually, plant defenses retard or stop bacterial ingress without the development of disease
symptoms. The term innate immunity, which is observed in both plants and animals, is often
used to refer to the defenses after the physical barriers are broached by microbes (Menezes et al.,
2000). Microbe invasion commonly induces a series of biochemical-changes that may lead to
resistance to disease development, such as production of the salicylic acid (SA), deposition of
lignin and callose in the cell wall, transcription of pathogenesis-related (PR) genes, and
production of antimicrobial compounds (Loake and Grant, 2007). The plant may also recognize
specific pathogen effectors that interact, directly or indirectly, with resistance (R) proteins,
leading to the activation of a rapid defense response and accompanying hypersensitive reaction
(HR), which is also called effector-triggered immunity (ETI) (Jones and Dangl, 2006). Basal
plant defenses are also triggered by the recognition of common microbial molecules called
pathogen associated molecular patterns (PAMPs). This recognition, which is called PAMP-
triggered immunity (PTI), involves recognition of PAMPs by pattern recognition receptors
(PRRs), which are receptor-like kinases (Jones and Dangl, 2006; Block et al., 2008). The major
known elicitors of PTI include flagellin peptide (flg22) (Zipfel et al., 2004), lipopolysacharides
(LPS) derived from pathogen cell walls (Keshavarzi et al., 2004), peptidoglycans, microbial cell
wall fragments, phospholipids, proteins, double stranded RNA and methylated DNA (Ingle et al.,
2006; Iriti and Faoro, 2007). Activation of basal defense responses by PTI results in rapid
increases of the Ca2+ flux, nitric oxide and reactive oxygen species (Nurnberger et al., 2004).

14
Phytopathogenic proteobacteria (Gram-negative bacteria) rely on different weapons to
neutralize the host defenses and promote the colonization of the host tissue. The weapons include
as several protein secretions systems, T3- effector proteins, toxins, small molecule suppressors,
cell wall degradation enzymes, and genes involved in flagellar and biofilm biosynthesis
(Shiraishi et al., 1997; Gross et al., 1999; Alfano et al., 2002; Wolpert et al., 2002; Moissiard
and Voinnet, 2004; Hommais et al., 2008). Pathogens use this diversity of weapons to undermine
core components of plant defenses, such as the HR, cell wall-based defenses, jasmonic acid
signaling, systemic acquired and induced resistance and expression of defense genes
(Abromovitch and Martin, 2004). Most phytopahogenic bacteria carry four secretion systems,
although new studies have demonstrated the presence of up to six secretion systems (Yahr,
2006). Four of the secretion systems have been well characterized in plant pathogenic bacteria
and are fundamental for the ingress, colonization, suppression of host defenses and disease
development. Approximately 20% of all polypeptides synthesized by bacteria are located
partially or completely outside of cytoplasm, and this arsenal of proteins makes use of these
systems for secretion and translocation into the apoplast and into plant cells (Pugsley, 1993;
Salmond and Reeves, 1993). In general, pathogenicity in proteobacteria is dependent upon
secretion machinery that mediates the transport and injection of toxic molecules into target
tissues or cells. These secretion systems are classified into six types (I to VI) and a summary of
the four major types are presented here.
Plant Pathogen Translocon Pathways
Type I secretion system
The route that each protein will be translocated through bacterial membranes is dependent
on the substrate. Translocation of most proteins, in general, is catalyzed by the secretory (Sec)
apparatus although some exported proteins are translocated independently of the core Sec-

15
translocon, named sec-independent (Danese and Silhavy, 1998; Cristobal et al., 1998). This
system, which is sec-independent and ABC-transporter dependent, is specific for a particular
protein or a family of closely related proteins (Higgins, 1995). The proteins are exported by ABC
(ATP-binding cassette) transporters, which are involved in the import or export of a great variety
of substrates such as different small peptide signaling molecules, bacteriocins and amino acids,
through the outer membrane (Gray et al., 1989; Wandersman, 1992; Higgins, 1995; Young and
Holland, 1999). Basically, the type I translocator is assembled on two dedicated inner membrane
proteins, first, the ABC transporter (provide energy and it is the initial channel across the inner
membrane), second, the membrane fusion protein (MFP), which starts in the periplasm and form
a channel to the surface with an third protein, TolC, in the outer membrane protein (Holland et
al., 2005). The proteins that are translocated by this system, usually, carry the secretion signal at
the C-terminal and is not removed during transport (Holland et al., 2005). The type I translocon
system plays an important role in both animal and plant bacteria pathogens. A type I mutant in
Actinobacillus pleuropneumoniae, which has hemolytic activity, is defective in protein secretion
and the strain is non-hemolytic (Kuhnert et al., 2005). The importance of the type I system is
demonstrated in plants as well. Rice plants that carry the Xa21 resistance gene, which mediates
recognition of bacterial strains expressing AvrXa21 activity, are resistant to Xanthomonas oryzae
pv oryzae although plants lacking Xa21 are susceptible (Gomez-Gomez and Boller, 2002).
Mutational analysis of X. oryzae pv oryzae genes involved in type I secretion revealed that the
strain is no longer recognized by Xa21 (da Silva et al., 2003).
Type II secretion system
Of the six secretion systems reported in phytopathogenic bacteria, the most prolific
pathway is the type II secretion system from which most proteins are secreted, including elastase,
lipase, phospholipases and exotoxins (Filloux et al., 1998; Ball et al., 2002). For these proteins to

16
reach their final destination, they may be delivered through two routes that are via sec machinery
or Tat export pathways (Douglas et al., 1987; Voulhoux et al., 2001). The proteins translocated
through the type II-dependent system are first translocated across the cytoplasmic membrane via
the sec machinery. The next step in the secretion process involves the mature periplasmic protein
being transported across outer membrane, which requires the function of at least 12 gene
products of a specialized apparatus called the secreton (Pugsley, 1993; Sandkvist, 2001).
Depending on the species, between 12 and 15 genes (named by letters A through O and S) are
essential for type II secretion and minor differences in genetic organization of this cluster in
closely related species have been observed (Pugsley, 1997; Sandkvist, 2001). Studies on
secretion genes in Erwinia chrysanthemi and Erwinia carotovora, which cause soft rot diseases
on various plants, showed that one of the genes, outN, is missing in E. chrysanthemi whereas its
presence in E. carotovora is required for functioning of the system (Lindeberg et al., 1996). da
Silva et al. (2002) describe the presence of two type II secretion systems in Xanthomonas citri
subsp citri (Xcc) and Xanthomonas campestris pv campestris, , that are involved in the secretion
of degradative enzymes and toxins. Although HrpX is known as type III secretion system
regulator in Ralstonia solanacearum (Genin et al., 1992) and Xanthomonas spp (Wengelnik and
Bonas, 1996), it also regulates some type II secretory proteins in Xanthomonas oryzae pv oryzae,
causal agent of bacterial leaf blight (Furutani et al., 2004). This broad regulation is also observed
in X. citri subsp citri where HrpG, which regulates HrpA and HrpX (Brito et al., 1999;
Wengelnik et al., 1996b), also regulates 11 type II secretion system-related proteins (Yamazaki
et al., 2008).
Type III secretion system
A major breakthrough in the plant pathology field was the elucidation of T3SS, which is
used for most Gram-negative pathogenic bacteria and, it is conserved among animal and plant

17
bacteria (Cornelis and Van Gijsegem, 2000). Species in the genera Yersinia, Shigella flexneri,
Salmonella typhimurium, Salmonella typhimurium, enteropathogenic E. coli, Erwinia amylovora,
Pseudomonas syringae, Pseudomonas solanacearum and Xanthomonas spp rely on the T3SS for
successful colonization of their respective hosts (Hueck, 1998; Cornelis and Van Gijsegem,
2000). Phytopathogenic bacteria such as Pseudmonas, Rasltonia, Xanthomonas, Erwinia, and
Pantoea invade their hosts by using natural openings, such as stomata, or wounds. When bacteria
ingress the host they promote the secretion and translocation a diverse group of bacterial
virulence factors through T3SS, refered to here as T3-effectors (Cornelis and Van Gijsegem,
2000). The first demonstration of the translocation of T3-effectors was observed with Yersinia
effector proteins (Rosqvist et al., 1994). Once these T3-effectors are inside the host cell, it is
expected to modulate plant physiology providing an environment beneficial to sustain the growth
of the pathogen outside the cell and suppress host defenses (Mudgett, 2005). The identification
of the P. syringae hypersensitive response and pathogenicity (hrp) gene cluster suggest that the
expression of these genes results in the formation of a T3SS structure or hrp pilus, which creates
a bridge between pathogen and host cell for protein delivery (Aizawa et al., 1998; Kimbrough
and Miller, 2000). All proteins secreted and/or translocated from T3S system are delivered from
the bacterial cytoplasm to the host cell interior passing between the bacterial and host
membranes/cell wall through this channel (Whittam et al., 2004). The remarkable achievement
related with T3S system in mammalian and plant bacterial pathogenesis is the finding that a
defect in this system often leads to complete loss of bacterial pathogenicity (Lindgren et al.,
1986; Zischeck et al., 1987). Despite the obvious importance of the T3SS, which functions in
delivering the type three effectors (T3-effectors) into plant cells (Galan & Collmer, 1999;
Kjemtrup et al., 2000), not all plant pathogens bacteria rely on this particular system, including

18
one of major pathogens of citrus trees in Brazil, Xylella fastidiosa and the causal agent of citrus
variegated chlorosis (CVC), (Dow and Daniels, 2000).
Type IV secretion system
Type IV secretion (T4SS) systems mediate the transport of toxic molecules to host cells
and DNA or protein-DNA complexes across the cell envelopes of Gram-negative and Gram-
positive bacteria (Cascales and Christie, 2003; Grohman et al., 2003). This system is involved in
the bacterial conjugation process, where two bacteria exchange genetic material, transmission of
DNA among diverse species of bacteria, and also able to deliver DNA to fungal, plant, and
mammalian cells (Bundock et al., 1995; Baron et al., 2002). Consequently, it plays crucial roles
in the spread of antibiotic resistance genes and other fitness traits among different species
(Christie et al., 2005). Studies have demonstrated that T4SSs are able to deliver not only DNA
but also effector proteins into the cytosols of eukaryotic cells, which in turn induce physiological
changes to aid in establishment of pathogenic or symbiotic relationships (Fisher et al., 2002;
Ding et al., 2003).
Several phytopathogenic bacteria carry the T4SS system but the significance of this system
for the plant pathology is that T4SS in Agrobacterium tumefaciens holds the ability to transfer
DNA to different kingdoms. T4SS system, in Gram-negative, is a multisubunit cell-envelope-
spanning structure composed of a transenvelope channel or pilus or other surface filament that
spans both membranes, the periplasm, and the cell wall (Christie, 2004; Lawley et al., 2003).
T4SS systems have been classified as type IVA, IVB, or other (Christie and Vogel, 2000).
Basically, the composition of type IVA shows similarity of the archetypic A. tumefaciens
VirB/D4 system, which is responsible for the synthesis this secretion machinery (Christie, 2004).
In addition, a subunit of genes termed Mpf proteins is required for assembly of the channel or
pilus for protein secretion (Haase et al., 1995). A T4SS system also found in Xcc is involved in

19
the adhesion of this pathogen to plant cells through the fimbriae, but the fimbrillin genes, which
are responsible for encoding the proteins that build the fimbriae structure, are different from
other Xanthomonas species (Cao et al., 2001; da Silva et al., 2002).
Hypersensitive Response and Pathogenicity (hrp) Genes
The T3SS is crucial in the process of colonization and establishment of favorable
environment for plant pathogenic bacteria in the host, thus, failure or malfunction in the system
severely impairs disease development. The first clue about the existence of hypersensitive
reaction and pathogenicity (hrp) genes was their requirement for elicitation of hypersensitive
reaction (HR) in nonhost plants and disease stimulation in susceptible plants (Lindgren et al.
1986). The correlation between hrp genes and secretion of bacterial proteins was first observed
some time later (Collmer et al., 1993; Rosqvist et al., 1994,; Wei and Beer, 1994). The first hrp
genes described were in P. s. pv. phaseolicola and have more recently been identified in most
gram-negative phytopathogenic bacteria, including P. syringae (Cuppels, 1986; Lindgren, 1997;
Huang et al., 1988), Xanthomonas campestris (Bonas et al., 1991), X. oryzae pv. oryzae (Kamdar
et al., 1993), Ralstonia solanacearum (Boucher et al.,1987), Erwinia amylovara (Barny et al.,
1990), E. chrysanthemi (Bauer et al., 1994), E. carotovora (Cui et al., 1996) and Pantoea
stewartii (Frederick et al., 1991). Usually the hrp cluster is composed of ~ 20 genes of which
nine genes have been shown to share high-level of homology among species, and as a result,
those nine genes have been named hrc (for hrp gene conserved; Bogdanove et al., 1996; Hueck,
1998). Moreover, these genes may be clustered in the chromosome as part of a large
pathogenicity island (PAI), which are characterized by having a G + C content different from the
rest of the chromosome and/or flanked by remnants of mobile DNA elements (Hacker and
Kaper, 2000). The Xcc strain 306 hrp cluster is composed of 26 genes extending from hpa2 to
hrpF (da Silva et al., 2002). As a result of structural studies, the structure or T3SS pilus consists

20
of two outer rings that interact with the outer membrane, two inner rings that interact with the
cytoplasmic membrane, and an extracellular needle-like extension that is ~8 nm in diameter and
80 nm in length in animal pathogenic bacteria (Aizawa et al., 1998; Kimbrough and Miller,
2000). In plant pathogens, the T3SS pilus is much longer (several µm) but has the same diameter
(Roine et al., 1997). Although this system is conserved among proteobacteria, the genes
responsible for the Hrp pilus forT3SS pilin subunits are hypervariable in their sequence, even
within pathovars. For example, the major subunit of the T3SS pilus of P.s. pv. tomato is the 11-
kDa HrpA protein, which shares only 30% identity with the HrpA protein of P. s. pv. syringae
(Deng et al., 1998). Mutational analyses on hrp genes have demonstrated that disruption in core
genes in this system leads to failure to cause disease or multiply in susceptible plants, as well in
the ability to trigger HR on nonhost plants (Lindgren et al., 1986).
T3SS gene expression in phytobacteria is induced in the plant apoplast, in close contact
with host cells, or some formulations of minimal media, (Schulte and Bonas, 1992; Galan and
Collmer, 1999; Alfano et al., 2000). The T3SS of Xanthomonas spp. is regulated by two major
regulators, HrpG and HrpX, which are transcribed from a divergent promoter region (Oku et al.,
1995; Wengelnik et al., 1996; Wengelnik and Bonas, 1996). HrpG is a member of the OmpR
subclass of two-component regulatory systems (Wengelnik et al., 1999). HrpX is another hrp-
regulatory gene, which belongs to the AraC family of regulatory proteins, and responsible for the
transcriptional activation of genes in the HrpA to HrpF operons. The hypothesis behind the hrp
system regulation is that HrpG is phosphorylated by sensing an unknown environmental signal,
the phosphorylated HrpG regulates expression of the hrpX and hrpA, and HrpX subsequently
activates transcription of the remaining hrp genes and some genes encoding T3-effector proteins
(Wengelnik et al., 1996; Wengelnik and Bonas, 1996). For HrpX-dependent activation, the

21
presence of a PIP-box (plant-inducible promoter box, consensus TTCGC-N15-TTCGC) in the
promoter region of each HrpX-regulated gene is required (Fenselau and Bonas, 1995; Wengelnik
and Bonas, 1996). Additionally, the HrpX inducibility requires a cis-acting element (YANNRT),
which is present in the – 10 promoter region (Ciesiolka et al., 1999; Cunnac et al., 2004).
The hrp-regulatory genes in P. syringae are dependent of the activator HrpL which is a
member of the extra cytoplasmic functions (ECF) family of response regulators (Xiao et al.,
1992; Morett and Segovia, 1993; Xiao et al., 1994). hrpL expression is controlled by HrpR and
HrpS, which are related to the NtrC class of response regulators and, synergistically, activate
expression of hrpL in response to plant signals (Albright et al., 1989; X. Tang et al., 2006).. The
mechanism of regulation of the hrp genes in R. solanacearum is similar to Xanthomonas. A
signal is sensed by an outer membrane receptor PrhA and the signal is transferred by the putative
transmembrane protein PrhR and the ECF sigma factor PrhI. Thus, the activated PrhI controls
expression of the regulators PrhJ, HrpG and HrpB. HrpB is 40% identical and 58% similar to
HrpX in X. c. pv. vesicatoria and functions as a positive regulatory protein that activates the
expression of hrp transcriptional units in R. solanacearum (Genin et al., 1992; Marenda et al.,
1998; Gurlebeck et al., 2006).
HrpA in P. syringae pv. tomato is the major structural component of a pilus-like structure,
and consequently, inactivation of hrpA leads to the termination of T3-effectors flux out of the
bacteria, and disease progress (Roine et al., 1997). In Xanthomonas, HrpE is the structural
component of the Hrp pilus and shows no sequence similarity to other T3SS pilin genes. Models
for X. c. pv. vesicatoria HrpF protein proposed that the protein produces a pore in the plant cell
membrane and facilitates protein translocation into plant cells (Rossier et al., 2000; Buttner et

22
al., 2002). However, HrpF plays a role in protein translocation and pathogenicity in vivo, it is
dispensable for type III mediated protein secretion in vitro (Buttner et al., 2002).
Harpins a HR elicitor protein
The hrp cluster of phytopathogenic bacteria carry genes that encode proteins able to elicit
HR and have been named harpins. Harpin or harpin-like proteins, in general, are glycine-rich,
heat-stable proteins, that are secreted through T3SS system and trigger the elicitation of HR in
plant tissues. However, harpins do not contain an amino-terminal signal peptide similar to T3-
effectors (Wei et al., 1992; Alfano and Collmer, 1997; Zhu et al., 2000; Zou et al., 2006). In
addition, evidence suggests that HR elicitation by harpins does not always result in a visible HR,
it may trigger other plant defense mechanism, such as systemic acquired resistance (SAR) or
expression of defense-related proteins (Peng et al., 2004; Liu et al., 2006; Wang et al., 2008).
The first harpin isolated was HrpN from E. amylovora Ea321 (Wei et al., 1992). Purified HrpN
was able to elicit a HR on incompatible tobacco plants, and the E. amylovora hrpN mutant was
no longer able to induce HR on tobacco plants or cause disease on susceptible apple plants (Wei
et al., 1992). Based on homology searches, hrpN homologs were found in other Erwinia species
including E. chrysanthemi (Bauer et al., 1995) and E. carotovora showing the same features (Cui
et al., 1996). In 1998, a second Harpin protein was isolated from E. amylovora (Kim and Beer,
1998). The novel harpin, named HrpW, shows a C-terminal domain that is homologous to fungal
pectate lyases (Pels) and an N-terminal similar to harpin protein. Contrasting with hrpN, hrpW is
not required for HR and pathogenicity although it still elicits an HR in its purified form and
enhances the HR activity of a hrpN mutant (Kim and Beer, 1998).
Harpins are not exclusive to Erwinia species. An homologous protein, HrpZ, has been
isolated form P. syringae pathovars (Lindgren, 1997; He et al., 1993). Like HrpN, purified HrpZ
induces a HR in tobacco and tomato plants, and hrpZ mutants show slight difference in virulence

23
when compared with the wild type strain (Preston et al., 1996). HrpW was also isolated in P. s.
pv. tomato DC3000. Like harpins, the protein is acidic and glycine rich lacks cysteine and is
deficient in aromatic amino acids (Charkowski et al., 1998). In agreement with the finding by
Kim and Beer (1998), the protein sequence revealed two distinct domains, an N-terminal harpin-
like domain and a C-terminal pectic lyase (Pel) domain similar to Nectria haematococca mating
type IV protein (Fusarium solani f. sp. pisi), bacterial Pels in E. carotovora, E. chrysanthemi,
and Bacillus subtilis (Liu et al., 1994; Guo et al., 1995; Shevchik et al., 1997; Charkowski et al.,
1998). Additionally, the gene is preceded by a consensus hrp promoter, PIB box, followed by a
rho-independent transcription terminator, which is characteristic of a monocistronic operon
(Charkowski et al., 1998). More recently, genome sequencing projects have identified hrpW
homologs in X. citri subsp citri (strain 306) and X. campestris pv. campestris (strain 33913) (da
Silva et al., 2002).
Type Three Secretion System-Dependent Effectors
The mechanism that controls secretion of the T3-effectors to plants is still unclear.
However, the secretion signals are conserved among plant and animal pathogenic bacteria. X. c.
pv. vesicatoria is able to secrete T3SS heterelogous proteins such as PopA from R.
solanacearum, AvrB from P. syringae and YopE from Yersinia pseudotuberculosis inside of
host cells, showing that the secretion signal is conserved among the T3-effectors (Rossier et al.,
1999). Studies demonstrated that the secretion signal for T3-effectors is in the N-terminus, e. g.
the first 11-17 amino acids of Yersinia outer proteins (Yops) are sufficient to induce its secretion
through T3SS (Sory et al.,1995; Lloyd et al., 2001). The secretion signal of AvrBs2 protein from
X. c. pv. vesicatoria is present in the first 58 residues (Mudgett et al., 2002). In addition, the N-
terminal sequences of the T3-effectors show some similarity such as presence of hydrophilic
amino acids, absence of acidic residues in the first 12 amino acids residues and high content of

24
serine residues (Guttman et al., 2002; Petnicki-Ocwieja et al., 2002; Gurlebeck et al., 2006). The
serine content in the first 25 amino acids of X. c. pv. vesicatoria T3SS substrates also present
certain level of variation (Gurlebeck et al., 2006). Studies have been reported that some animal
and plant pathogenic T3-effectors require specific chaperones for secretion (Bennett and Hughes,
2000; Gaudriault et al., 2002). In X. c. pv. vesicatoria, the protein HpaB, a so-called hrp
associated protein, is a T3SS chaperon that interacts with more than one effector and inhibits the
translocation of non-effector ptoteins such as HrpF. A hpaB mutant was shown to strongly
reduce secretion and translocation of effector proteins and enable the translocation of HrpF
(Buttner et al., 2004).
The T3-effectors inhibit different defense mechanisms in host cells in various ways. One
mechanism is the suppression of plant innate immunity by removing or inactivating a defense
signalling component. A high number of T3-effectors are cysteine proteases. For example,
AvrXv4 from X. c. pv. vesicatoria and a member of the AvrRxv/YopJ family, is a small
ubiquitin-related modifier (SUMO) protease that blocks ubiquitination and subsequent
proteolysis (Orth et al., 2000; Roden et al., 2004). AvrPphB from P. syringae is a papain-like
cysteine protease and cleaves the Arabidopsis protein kinase PBS1, which forms a complex with
RPS5 (resistance protein). The role that the protease plays in defense suppression is unknown in
this case. However, cleavage of this complex initiates ETI (Shao et al., 2003; Ade et al., 2007);
AvrRpt2, another cysteine protease from P. syringae cleaves RIN4, an interactor of the matching
RPS2 resistance protein and a negative regulator of PTI (Kim et al., 2005; Kim et al., 2005b);
AvrPtoB, in P. syringae, is a ubiquitin E3 ligase and participates in the degradation of the tomato
protein kinase Fen, which promotes the ETI activation by the interaction between Fen and

25
AvrPtoB. AvrPtoB also suppresses PTI-induced by MAPK pathways in Arabidopsis (He et al.,
2006; Rosebrock et al., 2007).
Another strategy of phytopathogenic bacteria to overcome host defense mechanisms is to
deliver T3-effectors that induce alterations in host transcription that results in increase in host
susceptibility (Block et al., 2008). Part of that group is AvrBs3 protein family, which in Xcc is
represented by the PthA protein that plays a role in pathogenicity by promoting cell hypertrophy,
division and, finally, death (Swarup et al., 1991, Duan et al., 1999). Three other AvrBs3-like
members from X. o. pv. oryzae, PthXo1, PthXo6 and PthXo7, that are transcription activator-like
T3-effectors increased the expression of rice genes Os8N3, OsTFX1 (a bZIP transcription factor)
and OsTFIIAλ1 (a small subunit of the transcription factor IIA), which increase disease
susceptibility (Yang et al, 2006; Sugio et al., 2007). AvrBs3 was recently shown to bind to
upa20 promoter, which encodes a transcription factor that regulates cell size. Thus, induces gene
expression and causes hypertrophy of plant mesophyll cells by mimicking eukaryotic
transcription factors (Kay et al., 2007). Today more than 40 effectors have been analyzed and
characterized although the complete understanding of the complex interaction of plant-pathogen
still has gaps.
Project Goals and Objectives
The aim of this project was to screen putative Xcc T3-effector genes that may be involved
with Xcc pathogenicity and virulence by mutagenesis. Additionally, a transient expression assay
in citrus leaf tissue was developed to better characterize bacterial proteins recognized by citrus
plants and which activate defense mechanisms. The objectives were to (I) use a molecular
mutagenesis tools to screen for candidate HrpX-regulon genes; (II) identify an unknown T3SS-
independent HR elicitor in Xcc; (III) characterize hrpW, and its role in pathogenicity; and (IV)

26
identify core regions of avrGf1, which is linked with hypersensitive reaction in grapefruit plants,
using a transient expression procedure.

27
CHAPTER 2 MUTATIONAL ANALYSIS OF TYPE III EFFECTORS GENES FROM Xanthomonas citri
subsp. citri
Introduction
Most phytopathogenic bacteria rely on complex gene networks to penetrate the defenses of
their host plants (Nurnberger and Lipka, 2005). Xanthomonas citri subsp. citri (Xcc) is the causal
agent of the citrus bacterial canker and gains access into the host through wounds or natural
openings (Brunings and Gabriel, 2003). Once inside, the bacterium injects a collection of
proteins, called type III effectors (T3-effectors) into the plant cells. T3-effectors are delivered by
a conserved type III secretion system (T3SS), and are indispensable for disease in most situation
and condition the host cell environment in favor of bacterial growth (Buttner and Bonas, 2002;
Alfano and Collmer, 2004). The T3SS system is encoded by hypersensitive reaction and
pathogenicity (hrp) genes (Cornelis and Van Gijsegem, 2000; Alfano and Collmer, 2004;
Collmer et al., 1993). Little is understood about the activities and targets within the host cells of
many T3-effectors but undobutely more than one T3-effector is expressed by a given pathogenic
strain (Cunnac et al., 2004; Schechter et al., 2004; Abramovitch et al., 2006). On the host side,
many plant species have evolved mechanisms to recognize specific T3-effectors, and the genes
that control recognition specificity are known as resistance (R) genes (Flor, 1971). This specific
recognition is called effector-triggered immunity (ETI), which generally culminates in the
induction of a form of rapid cell death known as the hypersensitive reaction or HR (Greenberg
and Yao, 2004; Block et al., 2008)
According to the genome sequence of Xcc strain 306, the hrp gene cluster is composed of
six hrp operons (hrpA, hrpB, hrpC, hrpD, hrpE, and hrpF), six hpa (hpa2, hpa1, hpaP, hpaA,
hpaB, and hpaF) genes and nine hrp conserved genes – hrc (hrcC, hrcT, hrcN, hrcJ, hrcU, hrcV,
hrcQ, hrcR, and hrcS) (Figure 2-1; da Silva et al., 2002). Expression of the hrp genes is up-

28
regulated during growth in the host. Induction is believed to be initiated upon phosphorylation of
the hrpG gene, which is promoted by a yet unidentified environmental stimulus. Phosphorylated
HrpG, which is a member of the response regulator of two-component regulatory systems, in
turn, activates the transcription of hrpX and hrpA (Wengelnik and Bonas, 1996). Consequently,
HrpX, which is a member of the AraC family of transcription factors, activates the expression of
hrpB to hrpF operons (Figure 2-2). The genes in the hrpB, hrpC, hrpD, hrpE and hrpF operons
carry a promoter element known, in xanthomonads as a PIP (plant inducible promoter)-box,
which is represented by the consensus sequence TTCGC-N15-TTCGC and the binding site for
the HrpX protein (Wengelnik and Bonas, 1996; Wengelnik et al., 1996a; Wengelnik et al.,
1996b). Additionally, another motif known as the - 10 box-like motif (YANNNT; Y: C/T; N:
A/T/C/G) is localized 30 – 32 base pairs downstream of the PIP box and has been proposed to be
necessary for HrpX-mediated regulation (Cunnac et al., 2004).
Our laboratory is interested in elucidating the function of proteins secreted through the hrp
system in the X. citri supbs. citri citrus interaction. The genome sequence analysis of Xcc strain
306 revealed greater than 30 candidate effector genes based on sequence relatedness to known
effectors and promoter elements (Table 2-1; da Silva et al., 2002). Therefore, a targeted
mutational analysis of the HrpX-regulon genes was undertaken.
Materials and Methods
Bacterial Strains and Plasmids
The bacterial strains and plasmids used in this study and their sources are described in
Table 2-2.
Media and Growth Conditions
Escherichia coli DH5α and C2110 were cultivated in Luria-Bertani broth (LB; 1% bacto
tryptone, 0.5% bacto-yeast extract, 1.0% sodium chloride) or on LB agar (2% agar) plates at

29
37oC with the appropriate antibiotics. For solid media BD Bacto agar was added at 15 g l-1.
Strains of Xanthomonas were routinely grown on nutrient agar (NA) medium or on tryptone
sucrose agar at 28oC (Zhu et al., 2000). All bacterial strains used in this study were stored in
20% glycerol in sterile tap water and maintained at -80oC. Triparental conjugations were
performed on NYG agar (Daniels et al., 1984). Antibiotics were used at the following
concentrations: ampicillin (Amp) 100 µg ml-1; chloramphenicol (Cm) 100 µg ml-1; kanamycin
(Kn) 50 µg ml-1; spectinomycin (Sp) 100 µg ml-1; rifampicin (Rif) µg ml-1; and tetracycline (Tc)
12.5 µg ml-1.
Recombinant DNA Techniques
Genomic and plasmid DNA were collected as described by Sambrook et al. (1989).
Restriction enzymes and T4 DNA ligase were obtained from, Promega (Madison, WI, USA) and
used following the manufacturer’s recommendations. Chemicals were purchased from Sigma-
Aldrich (St. Louis, MO, USA), and Fisher Scientific (Hampton, NH, USA). Polymerase chain
reaction (PCR) was performed with GoTaq® Flexi DNA Polymerase (Promega, Madison, WI,
US) and the PCR Express Thermal Cycler (ThermoHybaid, Ashford, UK).PCR products were
cloned with a TA cloning kit (Invitrogen Corp., Carlsbad, CA, USA). Individual constructs were
transfected into E. coli DH5α cells as described by Sambrook et al. (1989). Generation of Xcc
mutants was made using single homologous recombination methodology as described Sugio et
al. (2005). The selected plasmids were introduced into Xcc strain 306 by electroporation of Xcc
competent cells using a BioRad GenePulser II Electroporator. The conditions applied were 200
OHMs (resistance), 25 µFD (capacitance) and 2.5 Volts. After the pulse, cells were diluted by
addition of 250 µL of NB media. The cells were incubated for 1 h at 28oC with constant shaking
and plated on NA containing kanamycin at 50 µg ml-1. Plates were incubated at 28oC for 3 days.

30
Generation of Xcc 306 mutants was made using single homologous recombination methodology
as described by Sugio et al. (2005). Triparental mating, as described by Daniels et al. (1984),
was also used as a technique to introduce plasmids into Xcc 306. DNA sequencing on both
strands of DNA fragments was completed at DNA Sequencing Core Laboratory of the
Interdisciplinary Center for Biotechnology Research, University of Florida – Gainesville, FL
(UF-ICBR).
DNA Amplification
Individual sets of oligonucleotide primers were selected from the nucleotide sequence of
each gene of Xcc strain 306. The oligonucleotide primer sequences used in this experiment are
shown in Table 2-3. F1 and R1 primers were used to amplify a partial fragment from each gene.
A set of upstream primers (Up primers) were applied to confirm the insertional mutation in each
gene. DNA was amplified in a total volume of 25 µL. The reaction mixture contained 5 µL of 5x
buffer, 0.7 mM MgCl2, 0.2 mM of each deoxynucleotide triphosphate (Promega), 2 pmol of each
primer, and 0.04 U of Taq polymerase. The amount of template DNA added was 100 ng of
purified total bacterial DNA or 25 ng of a plasmid preparation, unless otherwise stated. A total of
30 amplification cycles were performed in an automated thermocycler. Each cycle consisted of 1
min of denaturation at 95oC, 1 min of annealing at XoC (X = the annealing temperature was used
in accord with the primer), and 1 min per kb of extension at 72oC . The last extension step was
extended to 10 min. Amplified DNAs were detected by electrophoresis in 1.0% agarose gels in
TAE buffer. The gel was stained with 0.5 µg of ethidium bromide per ml and then photographed
over a UV transilluminator.
Mutation of Candidate Genes by Homologous Recombination
A partial fragment, amplified by PCR and having an average size between 300-900 bp,
from Xcc genomic DNA was cloned in pCR2.1-TOPO, containing kanamycin and ampicillin

31
resistance genes. The plasmids containing the amplified fragment were used for transformation
of E. coli DH5α cells. The cells were plated onto LB agar plates containing both kanamycin and
ampicillin. The plasmid DNA extracted from DH5α transformants were restricted with EcoR I,
under conditions specified by the manufacturer, for insertion confirmation, and M13 forward and
M13 reverse primers were used to sequence the inserts in pCR2.1. The resultant plasmids with
partial sequence were introduced into Xcc 306 cells by electroportation as described above. The
pCR2.1 plasmid cannot replicate in Xcc strain 306, and cells containing the plasmids integrated
into the Xcc strain 306 genome, putatively by a single homologous recombination event, were
selected on NA plates containing kanamycin. After 3 days of culture, a number of the
transformed colonies were tested by PCR, using a primer flanking the upstream region of the
target gene combined with a pCR2.1 internal primer, to confirm an insertion event ocurred the
chromosomal gene of interest. Because of the presence of the linearized plasmid in the gene, the
insertional mutation was deemed to cause a disruption of the transcription of inserted gene
(Figure 2-3). A sequence analysis of the PCR fragment was also performed on each transformant
for validation of the insertion (Altschul et al., 1997). The insertional mutants were then tested for
pathogenicity and induction of the HR on appropriate plant cultivars and species.
Complementation of ΩhrpG and ΩhrpX Deficient Mutants of Xcc Strain 306
For the complementation of ΩhrpG and ΩhrpX mutants a pLAFR3 clone pL22, from X.
citri pv. citri strain Aw (XccAw) containing hrpG and hrpX, was used to complement both
mutants (Rybak et al., 2009). pL22 was transferred to the two Xcc mutants, 306::ΩhrpG and
306::ΩhrpX by biparental mating (Daniels et al., 1984). The transconjugants were then used for
phenotype analysis.

32
Plant Material and Inoculations
Grapefruit cv. Duncan (Citrus paradisi) plants were grown from seed in 15 cm plastic pots
in Terra-Lite® agricultural mix (Scott Sierra Horticultural Products Co. Marysville, OH). The
plants were kept in the glasshouse of the University of Florida in Gainesville, Florida at
temperatures ranging from 25-30oC. Tomato plants (Solanum lycopersicum) cvs. Bonny Best
were planted from seeds in Plugmix (W. R. Grace & Co., Cambridge, MA, USA). After two
weeks, the seedlings were transferred to Metromix 300 (W. R. Grace & Co) in 10 cm plastics
pots. The plants were kept in the glasshouse of the University of Florida in Gainesville, Florida
at temperatures ranging from 25-30oC. For preparation of bacterial suspensions, 18 h cultures
were harvested from the NA plates and suspended in sterile tap water, and standardized to an
optical density at 600 nm (OD600) = 0.3 (5 x 108 CFU/ml) with a Spectronic 20 Genesys
spectrophotometer (Spectronic-UNICAM, Rochester, NY, USA). For disease symptom assays,
bacterial suspensions of the strains used in this study were infiltrated at 5 x 108 CFU/ml into
abaxial surface of citrus leaves by using hypodermic syringe and needle, and the symptoms were
assessed up to 10 days after inoculation. Following inoculation, plant responses were evaluated
after 3-4 days for water-soaking and 6-7 days for pathogenicity. For HR tests, bacterial
suspensions adjusted to 5 x 108 CFU/ml in sterile tap water were infiltrated into leaflets of
tomato cv. Bonny Best with a hypodermic syringe and needle. The inoculated plants were
incubated in a growth room at 24-28oC and assessed for HR elicitation 24 h after infiltration for
HR induction. Disease scores are the means of those for three leaves. All the experiments were
repeated at least three times.

33
Results
Analysis of HrpX-Regulon Candidate Genes
HrpX controls expression of the operons hrpB to hrpF by binding to the PIP-box of the hrp
genes (Wengelnik and Bonas, 1996). In this study, DNA sequence analysis revealed 19 putative
HrpX-regulon genes in Xcc strain 306 based on the presence of the motif TTCGC-N15-TTCGC.
Six genes were present with a perfect PIP-box sequence (TTCGC-N15/16-TTCGC). Eleven genes
had imperfect PIP-box elements (mostly with one base change in the third base - TTCGN-N15-
TTCGN). Several genes were chosen simply on the basis of homology to other T3-effectors
despite the absence of the PIP box element such as avrPphE2 and xopX (Table 2-4). The
promoter regions were also analyzed for the presence of the -10 box-like motif YANNNT (A and
T are conserved, Y represents C or T). In total, 11 putative -10 box-like sequence motifs were
found among the 19 genes (Table 2-4).
Mutations of the regulatory and structural genes of Xcc 306 hrp pathway cause loss of pathogenicity
Mutations were constructed in hrp mutants, 306::ΩhrpG, 306:: ΩhrpX and 306:: ΩhrpA in
order to compare the range of host response from wild type to nonpathognenic as well as to
determine the effectiveness of the single recombination mutagenesis strategy in Xcc306.
Mutations were generated by single recombination in the regulatory genes hrpG and hrpX and
the structural gene hrpA (hrcC). The virulence of the mutants was determined by observing the
response in inoculated grapefruit leaves at high bacterial concentration (5 x 108 CFU/ml) in
comparison to the response of the wild-type strain Xcc 306. All three mutants were
nonpathogenic (Figure 2-3). Complementation assays of 306::ΩhrpG and 306:: ΩhrpX mutants
were performed using pL22 cosmid containing hrpG and hrpX genes from XccAw , which was
introduced, individually, into each mutant by conjugation. Consequently, full recovery of the Hrp

34
phenotype was observed with pL22 (Figure 2-3). The hrp mutants were also tested for their
ability to elicit a nonhost HR on tomato. Mutants 306::ΩhrpG and 306:: ΩhrpX did not lose the
ability to trigger HR in non-host tomato plants at a 5 x 108 CFU/ml inoculum concentration. In
addition, the HR induction observed for 306::ΩhrpG and 306:: ΩhrpX mutants was with similar
intensity and speed compared with the HR triggered by the Xcc wild type strain (Figure 2-3).
One possible explanation for the observed phenotypes was that a T3SS-independent protein is
recognized by a tomato resistance protein.
Phenotypic Analysis of Nineteen Candidate Effector and Hrp-Regulon Associated Genes.
To evaluate the role in pathogenicity of nineteen Xcc T3-effector candidate genes, we
mutated each gene, individually, using insertional mutagenesis procedure (Figure 2-4). The
insertion event on each mutant was analyzed by PCR and the resulting fragment was sequenced
to confirm the mutagenic plasmid has inserted in the proper position in the Xcc 306 genome. The
virulence assay in grapefruit leaves with all generated Xcc 306 mutants showed that all nineteen
mutants retained the ability to produce disease symptoms.
Discussion
In an attempt to isolate and characterize new pathogenicity genes from X. citri subsp. citri
strain 306, we have analyzed, using mutagenesis tools, 19 putative HrpX-regulated and/or
candidate effector genes, a hrpA gene that is expressed independently of the HrpX-regulon, and
two hrp-regulatory genes, hrpX and hrpG, as revealed by genomic sequence data (da Silva et al.,
2002). Not all of the Xcc HrpX-regulon candidate genes carry the consensus PIP-box motif
upstream of the start codon and the associated – 10 box-like sequence (Table 2-4). This motif has
been proposed to function as another cis element of HrpX regulon, and has been identified in
several HrpX-dependent expression genes (Noel et al., 2001; Furutani et al., 2006, Koebnik et

35
al., 2006; Furutani et al., 2009). Such motifs are found in most of the HrpX regulated genes in
Xanthomonas spp. (Wengelnik and Bonas, 1996; Wengelnik et al., 1996a; Wengelnik et al.,
1996b; Cunnac et al., 2004). Additionally, both motifs have been reported to be required for
HrpX-dependent expression, and base substitution in either motif can affect the expression of the
genes (Tsuge et al., 2005; Furatani et al., 2006; Furatani et al., 2009). And here we identified 11
genes preceded by both perfect/imperfect PIP-box and – 10 box-like motifs in each promoter
region, six genes only with perfect/imperfect PIP-box motif and two genes, avrPphE2 and xopX,
with the absence of both motifs (Table 2-4). The absence of PIP-box region in avrPphE2 has
been reported (da Silva et al., 2002) and the xopX gene has been identified in X. oryzae pv.
oryzae and X. campestris pv. vesicatoria, which possess a imperfect PIP-box and – 10 box-like
region (TTCTT-N15-TTCGC-N30-CAAATG) (Koebnik et al., 2006; Salzberg et al., 2008).
Recently studies have demonstrated that a perfect/imperfect PIP-boxes and – 10 like-box motifs
may be not sufficient for HrpX-dependent expression of genes and in xanthomonads, example,
xopB and hrpF genes lacking the PIP-box are expressed under HrpX-regulatory protein (Furutani
et al., 2004; Furutani et al., 2006). Moreover, base substitution in the conserved region may
reduce the promoter activity but when substitution affects several bases such as TTCGB-N15-
TTCGB, TRCGB-N15-TTCGB, TTCTB-N15-TTCGB, TTCGB-N15-VTCGB, TTCGB-N15-
TRCGB, TTCGB-N15-TTCHB (B: C/T; H: A/C/T; R: A/G; V: A/C/G) it can have a negative
effect on gene activities (Furutani et al., 2006).
Pathogenicity experiments to assess the influence of each gene for disease development
were carried out. Surprisingly, none of the 19 mutants generated in this study was observed to
affect the ability to incite disease symptoms in susceptible grapefruit leaf. Neither mutated
homologous genes, which have been reported to be required for pathogenicity, such as XAC0661

36
(endopolygalacturonase) and XAC2374 (polygalacturonase) that are similar to E. carotovora
subsp. carotovora PehA- polygalacturonase, a cell wall-degrading enzyme and HR elicitor in
Arabidopsis (Kariola et al., 2003). However, X. o. pv. oryzae T3-effectors suppress a plant basal
defense response elicited by T2-secreted plant cell-wall-degrading enzymes (Jha et al., 2007). In
addition, Yamazaki et al. (2008) suggested that Xcc may suppress the polygalacturonase-
mediated plant defense by coregulating polygalacturonase isozymes.
The pathogenicity assay with 306::Ωhpa1 and 306::ΩhpaF mutants did not show any
macroscopic change in the symptoms compared with the Xcc wild type strain contrasting with
the results reported in X. o. pv. oryzae and X. a. pv. glycines, where hpa1 and hpaF are required
for full virulence in (Zhu et al., 2000; Kim et al., 2003; Cho et al., 2007).. In the class of
Xanthomonas outer proteins, two mutants were assessed 306::ΩxopX (carrying an insertion in
xopX gene) and 306::ΩxopQ (carrying an insertion in xopQ gene) for pathogenicity, neither one
demonstrated canker symptoms. However, xopQ and xopX have been reported to be highly
conserved among Xanthomonas spp. (Roden et al., 2004; Furutani et al., 2009). In Xcv, XopX,
contributes to the virulence on host pepper and tomato plants and is T3SS-dependent (Metz et
al., 2005).
When the avirulence genes, 306avrXacE1, avrXacE2, avrXacE3 and avBs2 were mutated
in Xcc strain they produced typical canker symptoms on grapefruit leaves. Similar results were
reported previously with avrXacE1 and avrXacE3 homologs in Xcv ∆xopE1 and ∆xopE2,
respectively, mutants when tested in susceptible pepper plants – ECW (Thieme et al., 2007).
However, avrBs2 phenotype observed here contrasts with the phenotype demonstrated with
avrBs2 mutant from X. c. pv. vesicatoria, which is required for full virulence (Swords et al.,

37
1996). In conclusion these avirulence genes do not appear to have a pronounced effect on Xcc
fitness in disease development in grapefruit.
We demonstrated above that a putative pathogenicity Xcc genes tested are not required for
disease development in greenhouse conditions. These genes were identified, based on sequence
analysis, by da Silva et al. (2002) and they are considered genes that are regulated by HrpX. Our
results indicate that the putative hrpX-regulon and effector/avirulence genes do not have any
direct role in disease development (Table 2-5). However, we can not conclude that they are not
important for bacterial survival.
In xanthomonads hrp activation cascade, the HrpG up-regulates expression of hrpA and
hrpX genes, and then HrpX activates the transcription the hrpB to hrpF operons (Wengelnik and
Bonas, 1996; Wengelnik et al., 1996a; Wengelnik et al., 1996b). In our experiments, the
disruption of either hrpG or hrpX severely disrupts the induction of any symptoms in grapefruit
leaves but, unexpectedly, both mutants retained the ability to trigger HR in non-host tomato
plants. Similar results were reported by Marutani et al. (2005), where a P. syringae pv. tabaci
hrp-regulatory ∆hrcC mutant also was able to trigger HR in non-host tomato plants in the hrp-
independent manner (Chapter 3). Additionally, a hrp X. perforans mutant harboring a X. fuscans
pv. aurantifolli clone was able to trigger HR in similar manner. Based on these results, we
speculate that an unknown T3SS-independent gene in Xcc is a HR-elicitor. To deepen our
understanding the role of the HrpX-regulon genes for pathogenicity and the model of action of
T3SS-independent HR-elicitor proteins, more detailed mutagenesis and protein secretion analysis
are required.

38
Table 2-1. Xanthomonas citri pv. citri genes used in the study ORF Number Relevant characteristics* Hrp Gene Cluster XAC0416 hpa1 XAC0415 hrpA (hrcC) XAC1265 hrpG XAC1266 hrpX Extended Hrp Conserved Regulon XAC0277 Conserved hypothetical protein XAC0661 endopolygalacturonase XAC1706 Alkanal monooxygenase XAC1886 β-K-adipate enol-lactone hydrolase XAC2374 Polygalacturonase XAC2534 Conserved hypothetical protein XAC3309 aminopeptidase Additional Hrp Regulon Candidate XAC2370 Endopeptidase XAC2922 hprW XAC3230 Actin-ADP-ribosylating toxin domain Putative Effector/Avirulence XAC0286 avrPphE1 XAC3224 avrPphE2 XACb0011 avrPphE3 XAC0393 hpaF XAC3090 Leucine rich protein XAC0076 avrBs2 Xanthomonas outer protein XAC0543 xopX XAC4330 xopQ
* All the genes shown in this table were identified by da Silva et al. (2002)
39
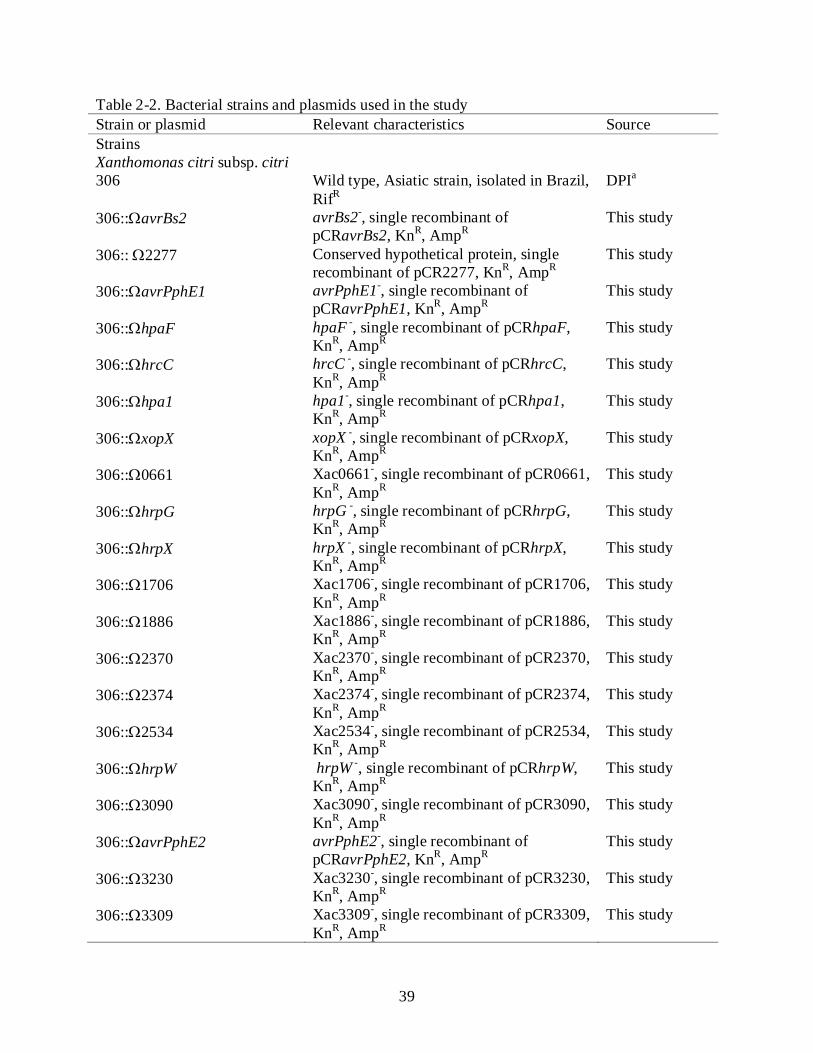
Table 2-2. Bacterial strains and plasmids used in the study Strain or plasmid Relevant characteristics Source Strains Xanthomonas citri subsp. citri 306 Wild type, Asiatic strain, isolated in Brazil,
RifR DPIa
306::ΩavrBs2 avrBs2-, single recombinant of pCRavrBs2, KnR, AmpR
This study
306:: Ω2277 Conserved hypothetical protein, single recombinant of pCR2277, KnR, AmpR
This study
306::ΩavrPphE1 avrPphE1-, single recombinant of pCRavrPphE1, KnR, AmpR
This study
306::ΩhpaF hpaF -, single recombinant of pCRhpaF, KnR, AmpR
This study
306::ΩhrcC hrcC -, single recombinant of pCRhrcC, KnR, AmpR
This study
306::Ωhpa1 hpa1-, single recombinant of pCRhpa1, KnR, AmpR
This study
306::ΩxopX xopX -, single recombinant of pCRxopX, KnR, AmpR
This study
306::Ω0661 Xac0661-, single recombinant of pCR0661, KnR, AmpR
This study
306::ΩhrpG hrpG -, single recombinant of pCRhrpG, KnR, AmpR
This study
306::ΩhrpX hrpX -, single recombinant of pCRhrpX, KnR, AmpR
This study
306::Ω1706 Xac1706-, single recombinant of pCR1706, KnR, AmpR
This study
306::Ω1886 Xac1886-, single recombinant of pCR1886, KnR, AmpR
This study
306::Ω2370 Xac2370-, single recombinant of pCR2370, KnR, AmpR
This study
306::Ω2374 Xac2374-, single recombinant of pCR2374, KnR, AmpR
This study
306::Ω2534 Xac2534-, single recombinant of pCR2534, KnR, AmpR
This study
306::ΩhrpW hrpW -, single recombinant of pCRhrpW, KnR, AmpR
This study
306::Ω3090 Xac3090-, single recombinant of pCR3090, KnR, AmpR
This study
306::ΩavrPphE2 avrPphE2-, single recombinant of pCRavrPphE2, KnR, AmpR
This study
306::Ω3230 Xac3230-, single recombinant of pCR3230, KnR, AmpR
This study
306::Ω3309 Xac3309-, single recombinant of pCR3309, KnR, AmpR
This study
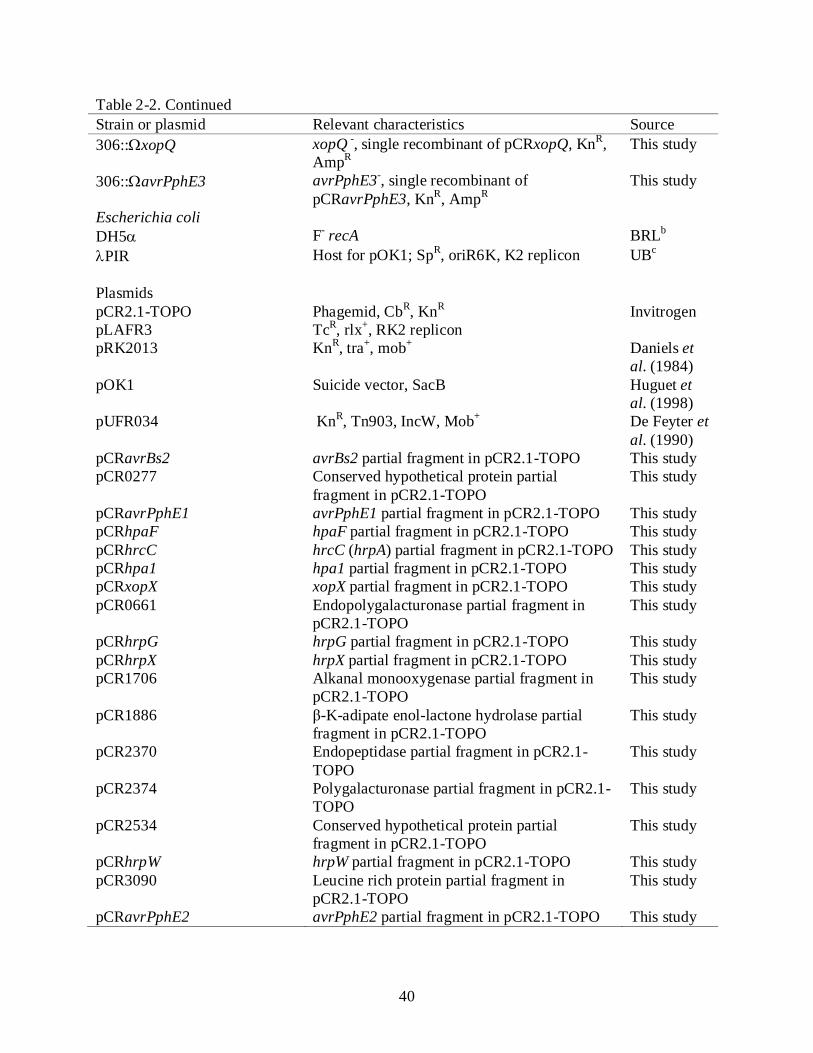
40
Table 2-2. Continued Strain or plasmid Relevant characteristics Source 306::ΩxopQ xopQ -, single recombinant of pCRxopQ, KnR,
AmpR This study
306::ΩavrPphE3 avrPphE3-, single recombinant of pCRavrPphE3, KnR, AmpR
This study
Escherichia coli DH5α F- recA BRLb
λPIR Host for pOK1; SpR, oriR6K, K2 replicon UBc
Plasmids pCR2.1-TOPO Phagemid, CbR, KnR Invitrogen pLAFR3 TcR, rlx+, RK2 replicon pRK2013 KnR, tra+, mob+ Daniels et
al. (1984) pOK1 Suicide vector, SacB Huguet et
al. (1998) pUFR034 KnR, Tn903, IncW, Mob+ De Feyter et
al. (1990) pCRavrBs2 avrBs2 partial fragment in pCR2.1-TOPO This study pCR0277 Conserved hypothetical protein partial
fragment in pCR2.1-TOPO This study
pCRavrPphE1 avrPphE1 partial fragment in pCR2.1-TOPO This study pCRhpaF hpaF partial fragment in pCR2.1-TOPO This study pCRhrcC hrcC (hrpA) partial fragment in pCR2.1-TOPO This study pCRhpa1 hpa1 partial fragment in pCR2.1-TOPO This study pCRxopX xopX partial fragment in pCR2.1-TOPO This study pCR0661 Endopolygalacturonase partial fragment in
pCR2.1-TOPO This study
pCRhrpG hrpG partial fragment in pCR2.1-TOPO This study pCRhrpX hrpX partial fragment in pCR2.1-TOPO This study pCR1706 Alkanal monooxygenase partial fragment in
pCR2.1-TOPO This study
pCR1886 β-K-adipate enol-lactone hydrolase partial fragment in pCR2.1-TOPO
This study
pCR2370 Endopeptidase partial fragment in pCR2.1-TOPO
This study
pCR2374 Polygalacturonase partial fragment in pCR2.1-TOPO
This study
pCR2534 Conserved hypothetical protein partial fragment in pCR2.1-TOPO
This study
pCRhrpW hrpW partial fragment in pCR2.1-TOPO This study pCR3090 Leucine rich protein partial fragment in
pCR2.1-TOPO This study
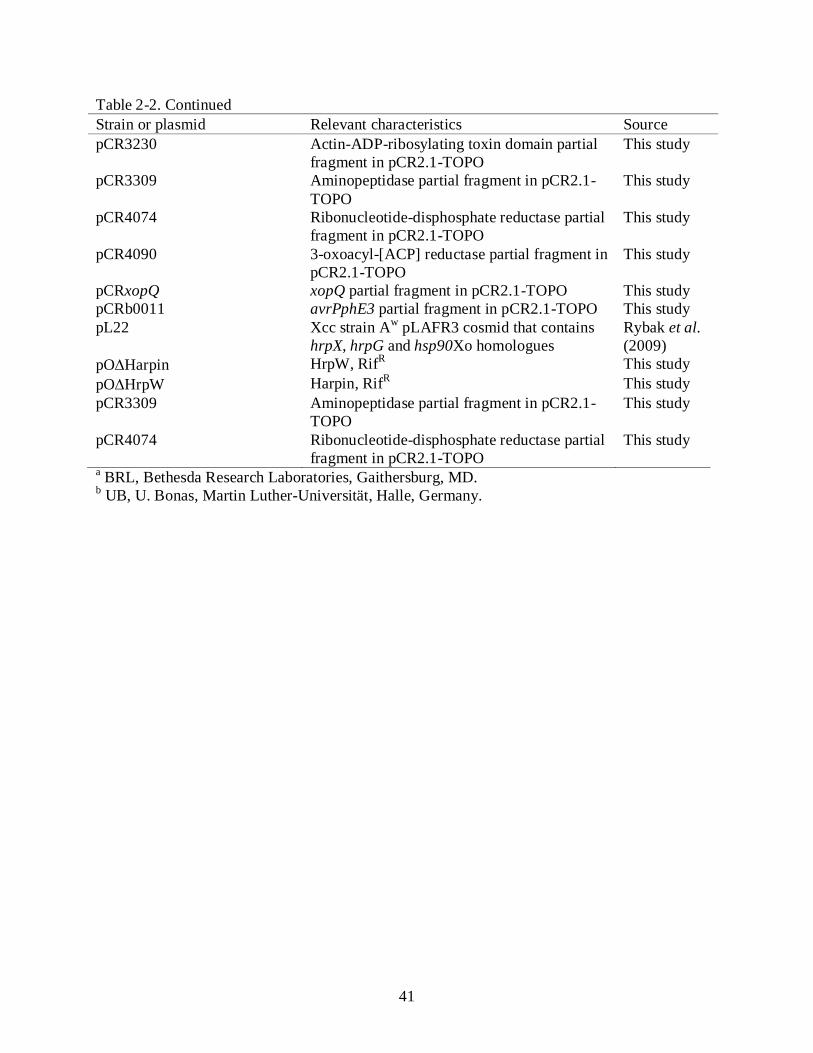
pCRavrPphE2 avrPphE2 partial fragment in pCR2.1-TOPO This study
41
Table 2-2. Continued Strain or plasmid Relevant characteristics Source pCR3230 Actin-ADP-ribosylating toxin domain partial
fragment in pCR2.1-TOPO This study
pCR3309 Aminopeptidase partial fragment in pCR2.1-TOPO
This study
pCR4074 Ribonucleotide-disphosphate reductase partial fragment in pCR2.1-TOPO
This study
pCR4090 3-oxoacyl-[ACP] reductase partial fragment in pCR2.1-TOPO
This study
pCRxopQ xopQ partial fragment in pCR2.1-TOPO This study pCRb0011 avrPphE3 partial fragment in pCR2.1-TOPO This study pL22 Xcc strain Aw pLAFR3 cosmid that contains
hrpX, hrpG and hsp90Xo homologues Rybak et al. (2009)
pO∆Harpin HrpW, RifR This study pO∆HrpW Harpin, RifR This study pCR3309 Aminopeptidase partial fragment in pCR2.1-
TOPO This study
pCR4074 Ribonucleotide-disphosphate reductase partial fragment in pCR2.1-TOPO
This study
a BRL, Bethesda Research Laboratories, Gaithersburg, MD. b UB, U. Bonas, Martin Luther-Universität, Halle, Germany.
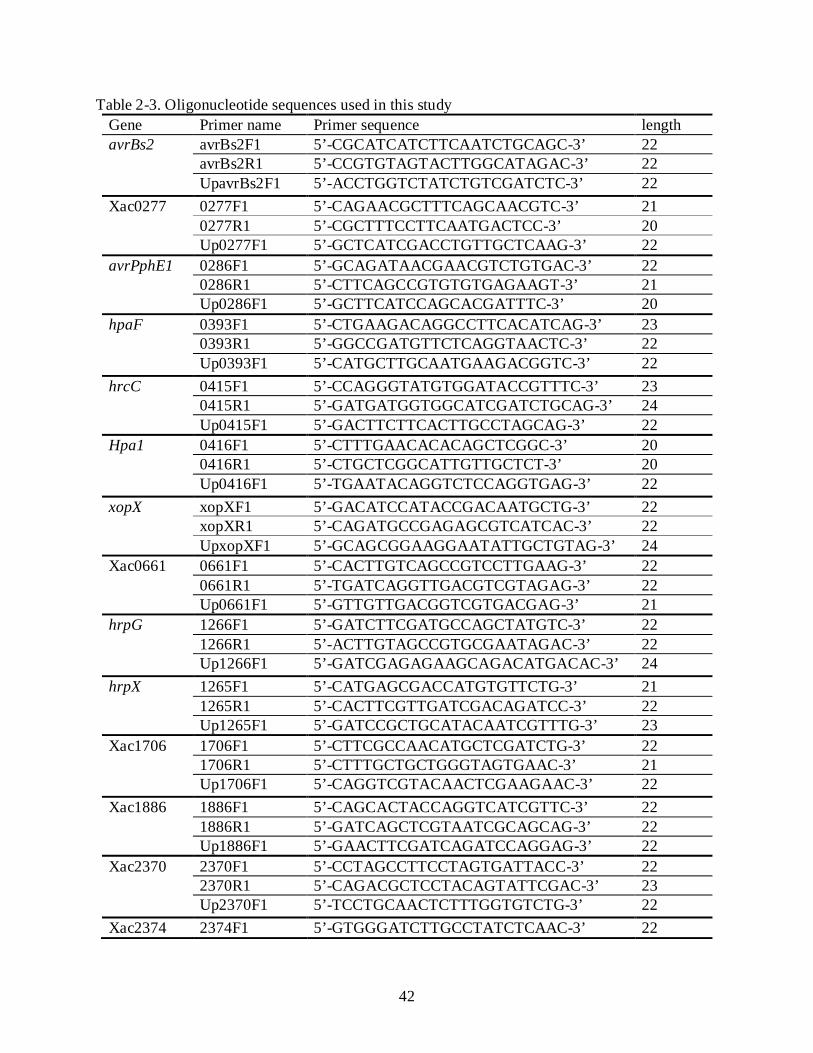
42
Table 2-3. Oligonucleotide sequences used in this study Gene Primer name Primer sequence length avrBs2 avrBs2F1 5’-CGCATCATCTTCAATCTGCAGC-3’ 22
avrBs2R1 5’-CCGTGTAGTACTTGGCATAGAC-3’ 22 UpavrBs2F1 5’-ACCTGGTCTATCTGTCGATCTC-3’ 22
Xac0277 0277F1 5’-CAGAACGCTTTCAGCAACGTC-3’ 21 0277R1 5’-CGCTTTCCTTCAATGACTCC-3’ 20 Up0277F1 5’-GCTCATCGACCTGTTGCTCAAG-3’ 22
avrPphE1 0286F1 5’-GCAGATAACGAACGTCTGTGAC-3’ 22 0286R1 5’-CTTCAGCCGTGTGTGAGAAGT-3’ 21 Up0286F1 5’-GCTTCATCCAGCACGATTTC-3’ 20
hpaF 0393F1 5’-CTGAAGACAGGCCTTCACATCAG-3’ 23 0393R1 5’-GGCCGATGTTCTCAGGTAACTC-3’ 22 Up0393F1 5’-CATGCTTGCAATGAAGACGGTC-3’ 22
hrcC 0415F1 5’-CCAGGGTATGTGGATACCGTTTC-3’ 23 0415R1 5’-GATGATGGTGGCATCGATCTGCAG-3’ 24 Up0415F1 5’-GACTTCTTCACTTGCCTAGCAG-3’ 22
Hpa1 0416F1 5’-CTTTGAACACACAGCTCGGC-3’ 20 0416R1 5’-CTGCTCGGCATTGTTGCTCT-3’ 20 Up0416F1 5’-TGAATACAGGTCTCCAGGTGAG-3’ 22
xopX xopXF1 5’-GACATCCATACCGACAATGCTG-3’ 22 xopXR1 5’-CAGATGCCGAGAGCGTCATCAC-3’ 22 UpxopXF1 5’-GCAGCGGAAGGAATATTGCTGTAG-3’ 24
Xac0661 0661F1 5’-CACTTGTCAGCCGTCCTTGAAG-3’ 22 0661R1 5’-TGATCAGGTTGACGTCGTAGAG-3’ 22 Up0661F1 5’-GTTGTTGACGGTCGTGACGAG-3’ 21
hrpG 1266F1 5’-GATCTTCGATGCCAGCTATGTC-3’ 22 1266R1 5’-ACTTGTAGCCGTGCGAATAGAC-3’ 22 Up1266F1 5’-GATCGAGAGAAGCAGACATGACAC-3’ 24
hrpX 1265F1 5’-CATGAGCGACCATGTGTTCTG-3’ 21 1265R1 5’-CACTTCGTTGATCGACAGATCC-3’ 22 Up1265F1 5’-GATCCGCTGCATACAATCGTTTG-3’ 23
Xac1706 1706F1 5’-CTTCGCCAACATGCTCGATCTG-3’ 22 1706R1 5’-CTTTGCTGCTGGGTAGTGAAC-3’ 21 Up1706F1 5’-CAGGTCGTACAACTCGAAGAAC-3’ 22
Xac1886 1886F1 5’-CAGCACTACCAGGTCATCGTTC-3’ 22 1886R1 5’-GATCAGCTCGTAATCGCAGCAG-3’ 22 Up1886F1 5’-GAACTTCGATCAGATCCAGGAG-3’ 22
Xac2370 2370F1 5’-CCTAGCCTTCCTAGTGATTACC-3’ 22 2370R1 5’-CAGACGCTCCTACAGTATTCGAC-3’ 23 Up2370F1 5’-TCCTGCAACTCTTTGGTGTCTG-3’ 22
Xac2374 2374F1 5’-GTGGGATCTTGCCTATCTCAAC-3’ 22
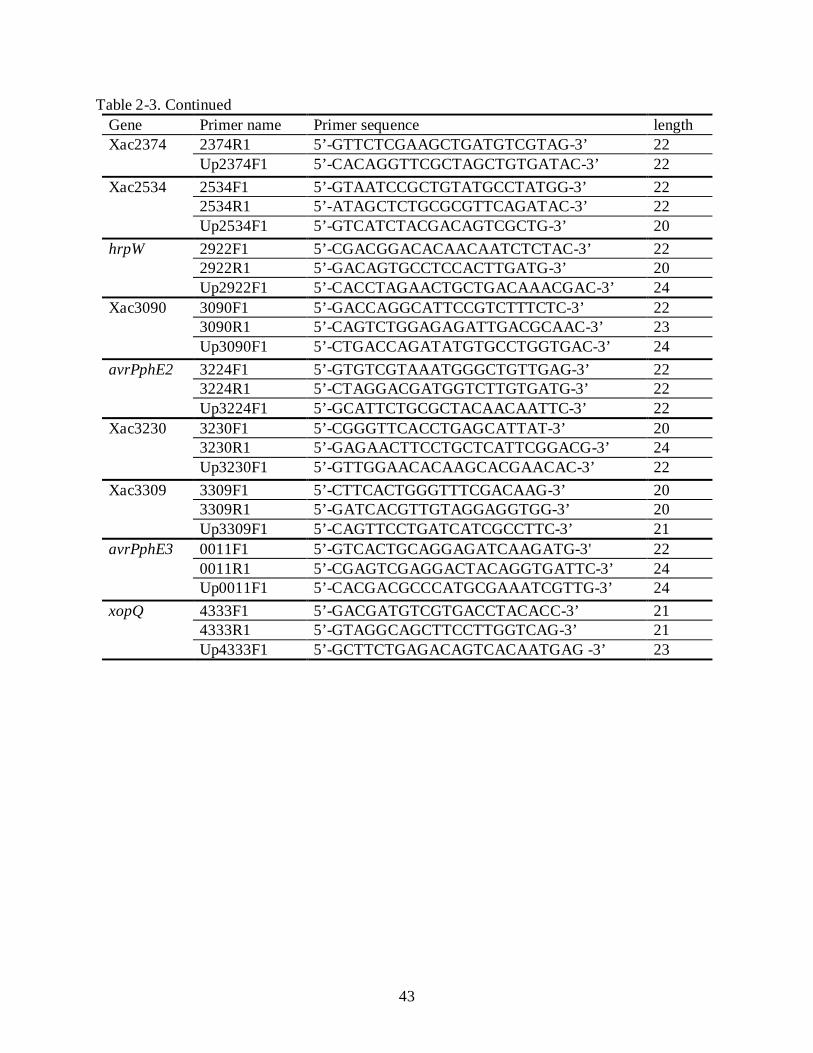
43
Table 2-3. Continued Gene Primer name Primer sequence length Xac2374 2374R1 5’-GTTCTCGAAGCTGATGTCGTAG-3’ 22
Up2374F1 5’-CACAGGTTCGCTAGCTGTGATAC-3’ 22 Xac2534 2534F1 5’-GTAATCCGCTGTATGCCTATGG-3’ 22
2534R1 5’-ATAGCTCTGCGCGTTCAGATAC-3’ 22 Up2534F1 5’-GTCATCTACGACAGTCGCTG-3’ 20
hrpW 2922F1 5’-CGACGGACACAACAATCTCTAC-3’ 22 2922R1 5’-GACAGTGCCTCCACTTGATG-3’ 20 Up2922F1 5’-CACCTAGAACTGCTGACAAACGAC-3’ 24
Xac3090 3090F1 5’-GACCAGGCATTCCGTCTTTCTC-3’ 22 3090R1 5’-CAGTCTGGAGAGATTGACGCAAC-3’ 23 Up3090F1 5’-CTGACCAGATATGTGCCTGGTGAC-3’ 24
avrPphE2 3224F1 5’-GTGTCGTAAATGGGCTGTTGAG-3’ 22 3224R1 5’-CTAGGACGATGGTCTTGTGATG-3’ 22 Up3224F1 5’-GCATTCTGCGCTACAACAATTC-3’ 22
Xac3230 3230F1 5’-CGGGTTCACCTGAGCATTAT-3’ 20 3230R1 5’-GAGAACTTCCTGCTCATTCGGACG-3’ 24 Up3230F1 5’-GTTGGAACACAAGCACGAACAC-3’ 22
Xac3309 3309F1 5’-CTTCACTGGGTTTCGACAAG-3’ 20 3309R1 5’-GATCACGTTGTAGGAGGTGG-3’ 20 Up3309F1 5’-CAGTTCCTGATCATCGCCTTC-3’ 21
avrPphE3 0011F1 5’-GTCACTGCAGGAGATCAAGATG-3' 22 0011R1 5’-CGAGTCGAGGACTACAGGTGATTC-3’ 24 Up0011F1 5’-CACGACGCCCATGCGAAATCGTTG-3’ 24
xopQ 4333F1 5’-GACGATGTCGTGACCTACACC-3’ 21 4333R1 5’-GTAGGCAGCTTCCTTGGTCAG-3’ 21 Up4333F1 5’-GCTTCTGAGACAGTCACAATGAG -3’ 23
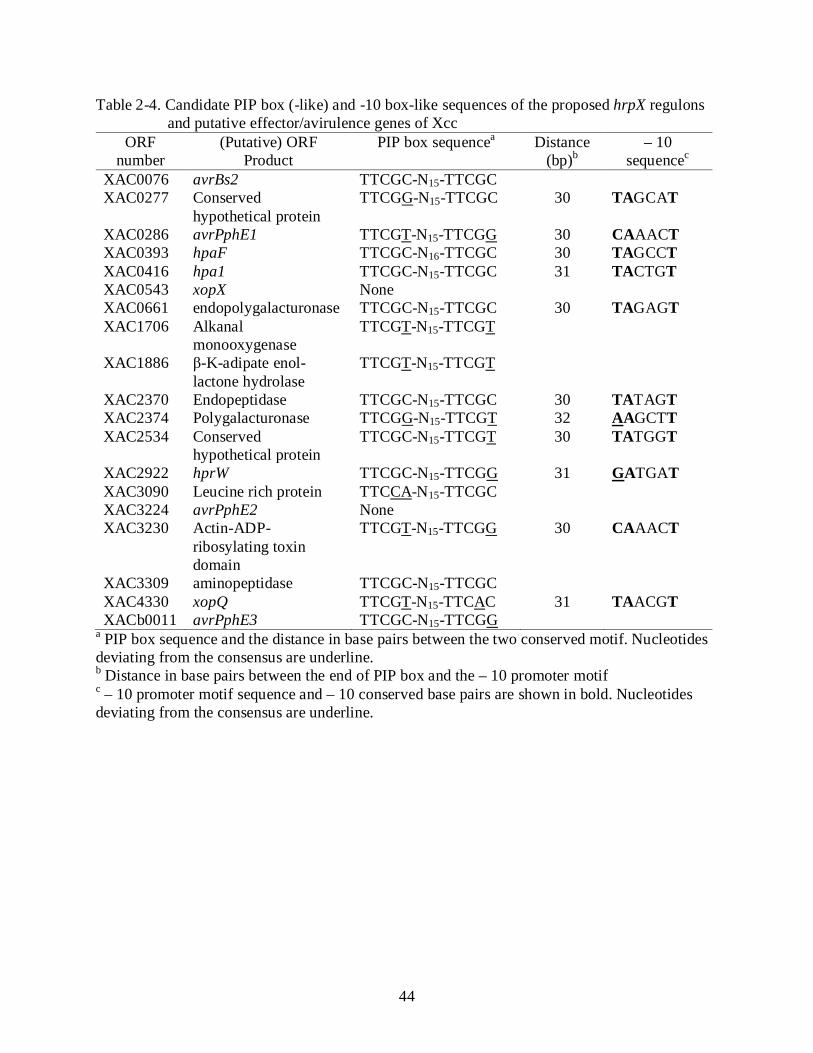
44
Table 2-4. Candidate PIP box (-like) and -10 box-like sequences of the proposed hrpX regulons and putative effector/avirulence genes of Xcc
ORF number
(Putative) ORF Product
PIP box sequencea
Distance (bp)b
– 10 sequencec
XAC0076 avrBs2 TTCGC-N15-TTCGC XAC0277 Conserved
hypothetical protein TTCGG-N15-TTCGC 30 TAGCAT
XAC0286 avrPphE1 TTCGT-N15-TTCGG 30 CAAACT XAC0393 hpaF TTCGC-N16-TTCGC 30 TAGCCT XAC0416 hpa1 TTCGC-N15-TTCGC 31 TACTGT XAC0543 xopX None XAC0661 endopolygalacturonase TTCGC-N15-TTCGC 30 TAGAGT XAC1706 Alkanal
monooxygenase TTCGT-N15-TTCGT
XAC1886 β-K-adipate enol-lactone hydrolase
TTCGT-N15-TTCGT
XAC2370 Endopeptidase TTCGC-N15-TTCGC 30 TATAGT XAC2374 Polygalacturonase TTCGG-N15-TTCGT 32 AAGCTT XAC2534 Conserved
hypothetical protein TTCGC-N15-TTCGT 30 TATGGT
XAC2922 hprW TTCGC-N15-TTCGG 31 GATGAT XAC3090 Leucine rich protein TTCCA-N15-TTCGC XAC3224 avrPphE2 None XAC3230 Actin-ADP-
ribosylating toxin domain
TTCGT-N15-TTCGG 30 CAAACT
XAC3309 aminopeptidase TTCGC-N15-TTCGC XAC4330 xopQ TTCGT-N15-TTCAC 31 TAACGT XACb0011 avrPphE3 TTCGC-N15-TTCGG
a PIP box sequence and the distance in base pairs between the two conserved motif. Nucleotides deviating from the consensus are underline. b Distance in base pairs between the end of PIP box and the – 10 promoter motif c – 10 promoter motif sequence and – 10 conserved base pairs are shown in bold. Nucleotides deviating from the consensus are underline.
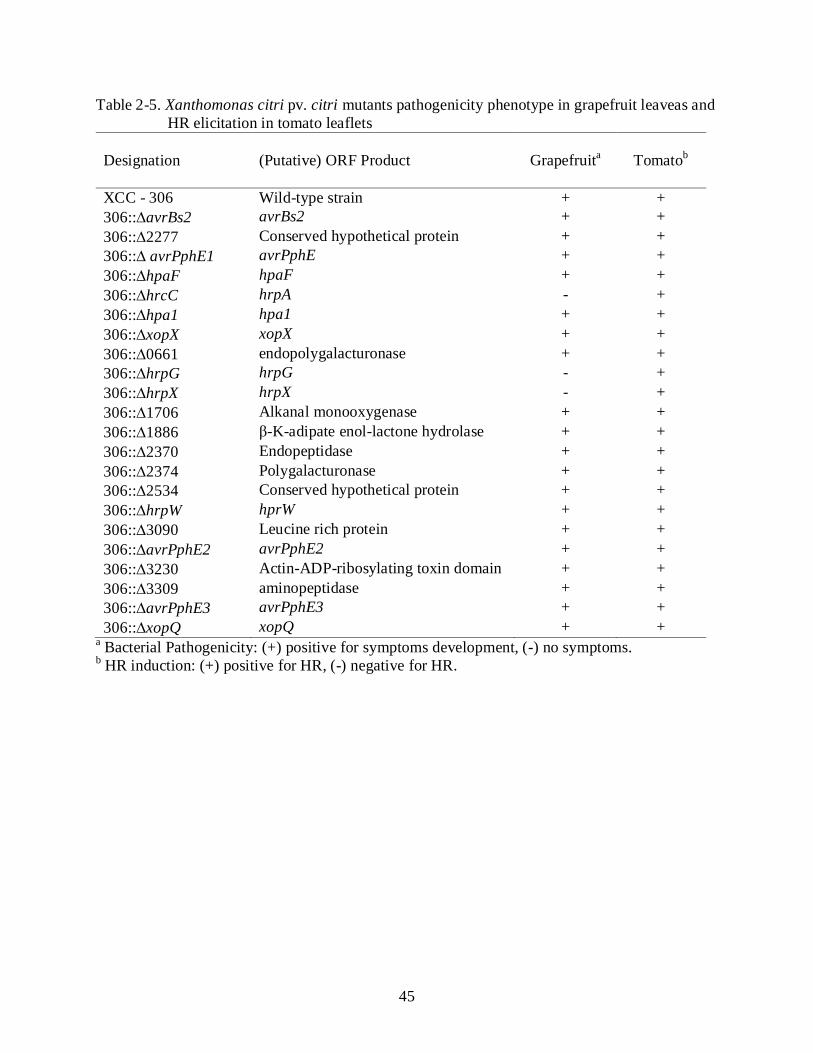
45
Table 2-5. Xanthomonas citri pv. citri mutants pathogenicity phenotype in grapefruit leaveas and HR elicitation in tomato leaflets
Designation (Putative) ORF Product Grapefruita
Tomatob
XCC - 306 Wild-type strain + + 306::∆avrBs2 avrBs2 + + 306::∆2277 Conserved hypothetical protein + + 306::∆ avrPphE1 avrPphE + + 306::∆hpaF hpaF + + 306::∆hrcC hrpA - + 306::∆hpa1 hpa1 + + 306::∆xopX xopX + + 306::∆0661 endopolygalacturonase + + 306::∆hrpG hrpG - + 306::∆hrpX hrpX - + 306::∆1706 Alkanal monooxygenase + + 306::∆1886 β-K-adipate enol-lactone hydrolase + + 306::∆2370 Endopeptidase + + 306::∆2374 Polygalacturonase + + 306::∆2534 Conserved hypothetical protein + + 306::∆hrpW hprW + + 306::∆3090 Leucine rich protein + + 306::∆avrPphE2 avrPphE2 + + 306::∆3230 Actin-ADP-ribosylating toxin domain + + 306::∆3309 aminopeptidase + + 306::∆avrPphE3 avrPphE3 + + 306::∆xopQ xopQ + +
a Bacterial Pathogenicity: (+) positive for symptoms development, (-) no symptoms. b HR induction: (+) positive for HR, (-) negative for HR.

46
Figure 2-1. Genetic organization of the hrp genes of X. citri subsp. citri. The arrows indicate the
orientation of the six hrp operons, hrpA to hrpF. The boxes correspond to open reading frames (ORFs). hrc genes encode proteins conserved among type III secretion system; hrp and hpa genes encode non-conserved proteins (da Silva et al., 2002; Dunger et al., 2005).
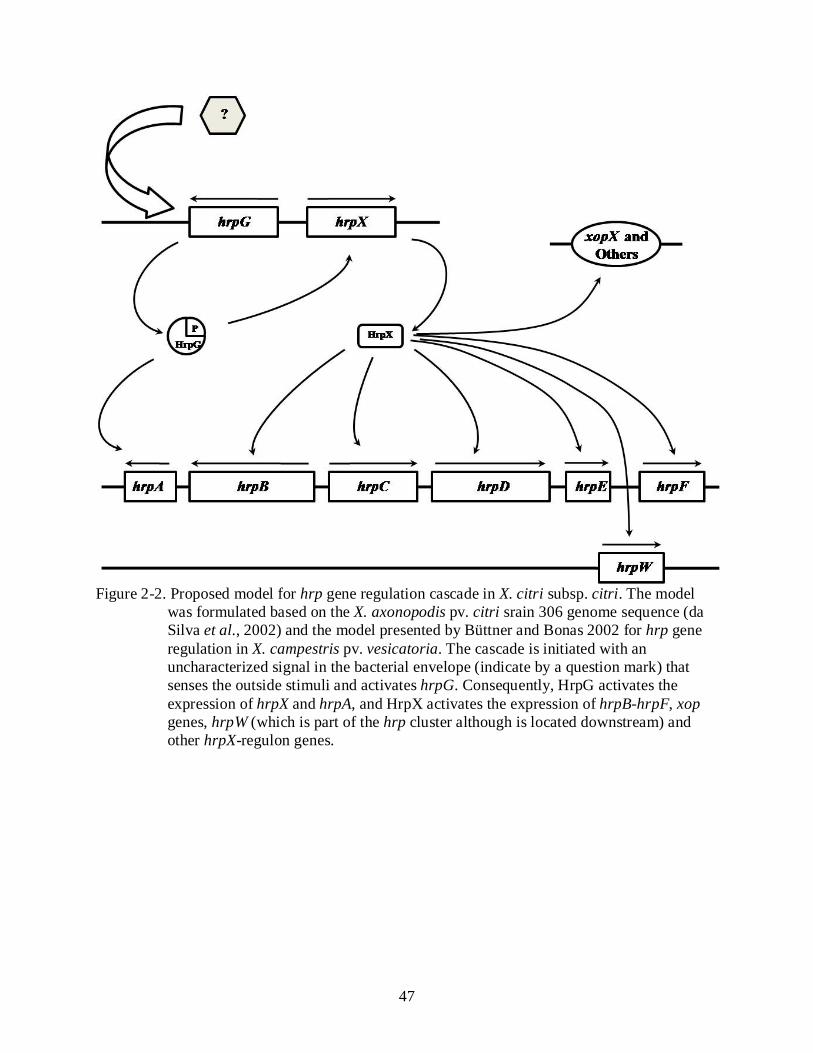
47
Figure 2-2. Proposed model for hrp gene regulation cascade in X. citri subsp. citri. The model
was formulated based on the X. axonopodis pv. citri srain 306 genome sequence (da Silva et al., 2002) and the model presented by Büttner and Bonas 2002 for hrp gene regulation in X. campestris pv. vesicatoria. The cascade is initiated with an uncharacterized signal in the bacterial envelope (indicate by a question mark) that senses the outside stimuli and activates hrpG. Consequently, HrpG activates the expression of hrpX and hrpA, and HrpX activates the expression of hrpB-hrpF, xop genes, hrpW (which is part of the hrp cluster although is located downstream) and other hrpX-regulon genes.

48
Figure 2-3. Comparison of the bacterial pathogenicity and HR elicitor in grapefruit leaves and tomato leaflets, respectively. The strains were infiltrated into the leaves at concentrations of 5 x 108 CFU/mL. Labeling: (1) 306::ΩhrpG-comlemented; (2) 306:: ΩhrpG; (3) Xcc strain 306 – wild type; (4) 306:: ΩhrpX; (5) 306:: ΩhrpX-comlemented; (6) 306:: ΩhrpA; (7) 306:: ΩhrpG. The grapefruit leaves were photographed 7 days after infiltration and the tomato leaf (right) was photographed 24 h after infiltration.
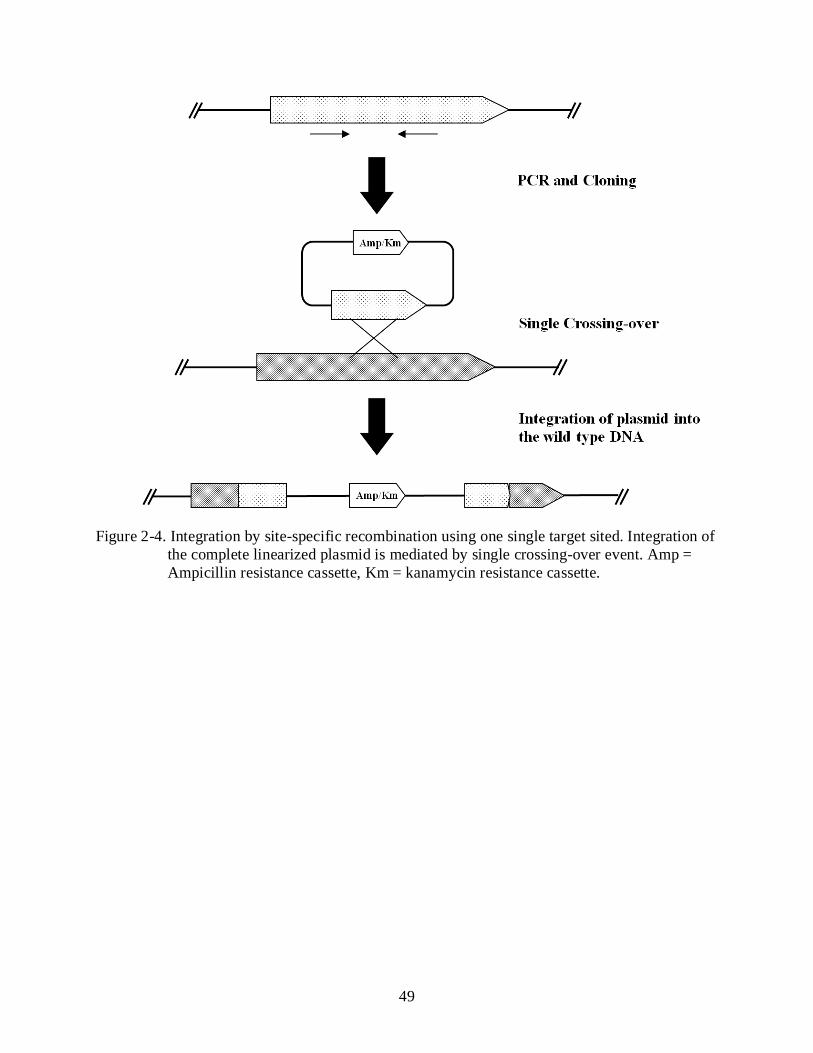
49
Figure 2-4. Integration by site-specific recombination using one single target sited. Integration of the complete linearized plasmid is mediated by single crossing-over event. Amp = Ampicillin resistance cassette, Km = kanamycin resistance cassette.

50
CHAPTER 3 IDENTIFICATION OF A HRP-INDEPENDENT HR ELICITOR FROM Xanthomonas citri
subsp. citri
Introduction
Plants are frequently under attack by pathogens. Plants are resistant to most pathogens, and
the responses to pathogen attack involve different innate immunity responses. Innate immunity
responses can be initiated by a local infection where the defense response extends to unaffected
parts, a response called induced systemic resistance (ISR) or systemic acquired resistance (SAR)
(Hunt et al., 1996; Hammerschimidt, 1999). The elicitation of plant innate immunity by bacteria
is sometimes divided, conceptually, into two type of elicitation. PAMP triggered immunity
(PTI) involves the induction of defense response by a variety of common molecular components
of bacteria known collectively as pathogen associate molecular patterns. Effector triggered
immunity (ETI) involves the triggering of defense by effector proteins of the type III secretion
pathway, which are present in many plant and animal pathogenic proteobacteria. Innate
immunity involves the activation of mitogen-activated protein kinase (MAPK) cascades and
synthesis of hormones such as salicylic acid (SA) or jasmonic acid (JA) (Block et al., 2008). The
bacterial pathogens also trigger the production of an oxidative burst, which, ultimately, limits
pathogen spread by induction of host cell death (Baker and Orlandi, 1995; Gozzo, 2003; Block et
al., 2008) and leads to changes in the cell wall composition and the de novo synthesis of
antimicrobial compounds such as phytoalexins and pathogen-related (PR) proteins (Bestwick et
al., 1998; Hammerschimidt, 1999b; Innes, 2001). Other responses include closure of stomata in
response to bacterial attack (Melotto et al., 2006); and the hypersensitive reaction (HR) around
the initial infection sites isolating the pathogen in dead cells, consequently, preventing the spread
throughout the plant (Alfano and Collmer, 2004).

51
Type III secretion systems (T3SS) function to translocate a collection of effectors (T3-
effectors) into the plant cells, where the T3-effectors modulate the cell environment and enhance
the conditions for bacterial multiplication (Cornelis and van Gijsegem, 2000). The genes
encoding most of the components for the T3SS are clustered into the hypersensitive reaction and
pathogenicity (hrp) gene regions (Büttner and Bonas, 2003). Basically, hrp gene clusters in plant
pathogenic bacteria, including Xanthomonas species, are regulated and dependent on the host
interaction. In Xanthomonas species, two key regulatory proteins are HrpG and HrpX
(Wengelnik and Bonas, 1996). In R. solanacearum, the hrp system is regulated through a HrpG
homolog and, downstream, a HrpX homolog originally named HrpB. HrpB and HrpX belong to
to the AraC family of transcriptional regulators (Wengelnik and Bonas, 1996; Brito et al., 1999).
P. syringae possesses three regulatory genes hrpR, hrpS and hrpL. The products of two genes,
HprR and HrpS, positively regulate the expression of hrpL, which encodes an alternative sigma
factor responsible for transcriptional activation of other hrp genes (Xiao et al., 1994). In
addition, homologs of hrpS and hrpL also occur in Erwinia amylovora (Sneath et al., 1990; Wei
et al., 1992).
In 1971, Flor demonstrated that a component from the bacteria, avirulence (avr) gene,
when recognized by the host triggered a defense response. Later, studies showed that T3SS was
the system used by the bacteria to inject the avr genes into the host cells (T3-effectors) and the
recognition was performed by host resistance protein. A malfunction of the T3SS generally
results in the complete loss of disease development and the loss of ETI or Hrp-dependent
elicitation of resistance responses due to the inability to transport the avr protein into the host
(Jones and Dangl, 2006; Block et al., 2008). The first avirulence gene, avrA, was isolated from
P. s. pv. phaseolicola in 1984 (Staskawicz et al., 1984). More than 40 Avr proteins have since

52
been identified. The first direct interaction between an Avr protein and the cognate R protein was
demonstrated with the tomato bacterial speck bacterium P. s. pv. tomato harboring the gene
avrPto, whose product is recognized by the tomato resistance protein Pto (Tang et al., 1996;
Mucyn et al., 2006). Additionally, Pto also recognizes another distinct Avr protein from P. s. pv.
tomato – known as AvrPtoB (Abramovitch et al., 2003,Wu et al., 2004). AvrXa27 in X. oryzae
pv. oryzae also induces resistance in rice plants carrying the cognate R gene Xa27 but the
resistance is induced only in the presence of the pathogen (Gu et al., 2005). PTI, on the other
hand, is the recognition of PAMPs by receptor-like kinases, which are localized in the
extracellular matrix (Jones and Dangl, 2006). PTI response is not generally considered to be a
race-specific defense. Therefore, ETI requires the T3-effectors and is essentially T3SS-
dependent, and PTI is T3SS-independent and is generally not accompanied by an HR.
In chapter 2 was demonstrated that hrpG and hrpX mutants of Xanthomonas citri subsp.
citri strain 306, the causal agent of citrus canker (Stall and Civerolo, 1991), retained the
capability to trigger an HR in the non-host plant tomato, indicating that the HR was T3SS-
independent. Similarly, Xanthomonas fuscans subsp. aurantifolli (Xfa) strain C, the causal agent
of citrus canker in Key lime (Schubert et al., 2001; Schaad et al., 2005), also triggered an HR in
tomato in a T3SS-independent manner as (data unpublished). In order to identify the elicitor of
HR, a cloning strategy using restriction enzymes was initiated to identify the genes controlling
the T3SS-independent HR response in tomato.
Materials and Methods
Bacterial Strains and Plasmids
Bacterial strains and plasmids used in this study are listed in Table 3-1.

53
Media and Growth Conditions
Escherichia coli was grown in Luria Bertani (LB) medium (Sambrook et al., 1989) at 37oC
with appropriate antibiotics. For solid media BD Bacto agar was added at 15 g l-1. Strains of
Xanthomonas were cultured on nutrient agar (NA) medium and on tryptone sucrose agar at 28oC
(Zhu et al., 2000). All bacterial strains used in this study were stored in 20% glycerol in sterile
tap water and maintained at -80oC. Triparental matings were performed on nutrient-yeast extract-
glycerol (NYG) agar (Daniels et al., 1984). Antibiotics were used in the following
concentrations: ampicillin (Amp) 100 µg ml-1; kanamycin (Kn) 50 µg ml-1; rifampicin (Rif) 100
µg ml-1; and tetracycline (Tc) 12.5 µg ml-1.
Recombinant DNA Techniques
Standard DNA manipulations were performed as described by Sambrook et al. (1989).
Restriction enzymes, T4 DNA ligase, and GoTaq® Flexi DNA Polymerase were used according
the recommendations of the manufacturer (Promega, Madison, WI, USA). Chemicals were
purchased from Sigma-Aldrich (St. Louis, MO, USA), and Fishers Scientific (Hampton, NH,
USA). The polymerase chain reaction (PCR) was performed in a PCR Express Thermal Cycler
(ThermoHybaid, Ashford, UK). The Topo TA cloning kit was used for cloning of PCR products
(Invitrogen, Carlsbad, CA, USA). All the enzymes and kits were used according to specifications
of the manufacturers. Constructs were transformed into competent Escherichia coli DH5α cells
described by Sambrook et al. (1989). Generation of Xcc 306 mutants was made using single
homologous recombination methodology as described by Sugio et al. (2005). The selected
plasmids were introduced into Xcc 306 thorough electroporation. Aliquots of Xcc competent
cells were mixed with plasmid DNA and electroporated using a BioRad GenePulser II
Electroporator instrument. The conditions applied were 200 OHMs (resistance), 25 µFD
(capacitance) and 2.5 Volts. After the pulse, the cells were immediately diluted by addition of

54
250 µL of NB medium. The cells were incubated for 1 h at 28oC with constant shaking, and
plated on NA containing kanamycin at 50 µg ml-1;, then incubated at 28oC for 2 days. Triparental
mating, as described by Daniels et al. (1984), was used as a technique to introduce plasmids into
Xcc 306. Sequencing was performed at DNA Sequencing Core Laboratory of the
Interdisciplinary Center for Biotechnology Research, University of Florida – Gainesville, FL
(UF-ICBR).
DNA Amplification
For mutagenesis analysis of hprG and hprX genes in Xcc 306, the custom primer pairs of
1266F1 5’-GATCTTCGATGCCAGCTATGTC-3 and 1266R1 5’-ACTTGTAGCCGTGCGAAT
AGAC-3' and 1265F1 5’-CATGAGCGACCATGTGTTCTG-3’ and1265F1 5’-CATGAGCGAC
CATGTGTTCTG-3’ and 1265R1 5’-GATCG-AGAGAAGCAGACATGACAC-3’ were
designed to amplify a partial fragment of hprG and hprX genes, respectively, from Xcc 306
genomic DNA. For subsequent validation of gene insertion, the primers Up1266F1 5’-GATCGA
AGAGAAGCAGACATGACAC-3’ Up1265F1 5’-GATCCGCTGCATACAATCGTTTG-3’,
were designed upstream start codon of hrpG and hrpX, respectively, and were used in
combination with the standard flanking vector M13FOR and M13REV primers of the TOPO
vector pCR2.1. The parameters used for PCR were as follows: Step 1. 90oC for 5 min; step 2.
95oC for 1 min; step 3. annealing for 1 min (the annealing temperature was calculated in
according to the C/G content of the primer); step 4. 72oC for 1 min per Kb, 30 cycles from step 2
to step 4; step 5. 72oC for 10 min.
Mutagenesis of hrpG and hrpX in X. citri subsp. citri 306
A partial fragment of each gene was amplified by PCR using Xcc genomic DNA as
template and the appropriate primer pair. The PCR fragments were purified using QIAquick PCR
Purification Kit purification system (QIAGEN) and cloned into pCR2.1. The constructs were

55
transformed into E. coli DH5α and the transformants containing the single homologous
recombination were selected by plating onto LB media containing kanamycin 50 µg ml-1.
Transformants were analyzed by restriction enzyme digestion profiles and sequencing. The
derivative plasmids with either a portion of hrpG or hrpX, respectively, were introduced into Xcc
306 by electroporation. Treated strains were spread on NA plates containing Kanamycin 100 µg
ml-1 and incubated at 28oC for 2-3 days. PCR amplification with upstream primers and the
standard flanking vector M13FOR and M13REV primers were used to confirm if colonies
contained a mutation in the appropriate gene as a result of a single crossover and insertion of the
pCR2.1 vector into the wild type gene. The appropriate wild type gene in pLAFR3, pVX9, was
re-introduced into the respective mutant (306::ΩhrpG and 306::ΩhrpX) and tested for
complementation of the mutant phenotype. Wild type, mutant, and complemented strains were
then tested for pathogenicity on citrus and their ability to elicit an HR in the non host plants
tomato and pepper.
Subcloning of HR-Elicitor Candidate from Xaa Library
The genomic library used in this study was made using genomic DNA from X. a. pv.
aurantifolli C strain. The DNA was partially digested with Sau3A and fragments in the 25- to
30-kilobase (Kb) range were isolated from the gel following gel electrophoresis. DNA was
ligated with BamHI-digested pLAFR3, and transformed and maintained in E. coli DH5α
(Staskawicz et al., 1984). The cosmids were introduced into a hrp- mutant of X. perforans strain
91-118 (91::Ωhrp) by triparental mating. Mixing 91::Ωhrp mutant cells as recipient with cosmid
as donor and with pRK2073 as the helper plasmid on NYG agar. The ratio of recipient to donor
to helper was 2:1:1 (vol/vol/vol). After 24 h of incubation at 28oC, the mating mixtures were
resuspended in sterile tap water, and aliquots were spread onto NA plates containing rifamycin

56
and tetracycline (100 µg ml-1 and 12.5 µg ml-1, respectively) for selection of transconjugants.
Individual colonies were selected and subsequently inoculated in tomato plants to screen for HR
induction after 24 h (Gerald V. Minsavage - personal communication).
Sequencing analysis of the pL450 clone, which was identified as HR positive in the tomato
screen, was performed using custon primers, NP5 (5’-CCCTTCACCAAGTTCGACGACA-3’)
and NP3 (5’-GCGGGTGCCGTGCTCGTGTT-3’), used to sequence both ends of the 450 clone.
By sequencing the ends of the cosmid we generated a restriction map to facilitate subcloning the
HR-elicitor. The subcloning of the 450 clone was performed by restriction digestion of the
original 450 clone with BamHI enzymes and purification of the excised fragments from an
agarose gel by using the QIAquick Gel Extraction Kit purification system (QIAGEN). The
fragments were ligated individually into BamHI digested pUFR034 using T4 DNA ligase
according to the manufacturer’s instructions. Ligation products were transformed into competent
cells of E. coli DH5α produced by the calcium chloride procedure as described by Sambrook et
al. (1989). Triparental matings were performed with Xcv (wild type), 91::Ωhrp mutant,
opportunistic xanthomonads T55 and INA42 as recipients with plasmid as donor and with
pRK2073 as the conjugational helper on NYG agar. Individual transconjugant colonies were
injected into tomato leaflets to detect subclones carrying the HR-elicitor genes.
A 4.5 Kb HR positive subclone, B38, was digested with BamHI and EcoRI enzymes, and
purification was performed as described above. Two fragments, 3.0 kb and 1.5 kb, were ligated
into BamHI and EcoRI digested phagemid vector pBluescript II/KS (Stratagene, La Jolla, CA)
using T4 DNA ligase used according to the manufacturer’s instructions. Ligation and triparental
matings in X. perforans (wild type) and 91::Ωhrp mutant were performed as described above.
Individual transconjugant colonies were injected into tomato leaflets to detect which fragment

57
was carrying the HR-elicitor gene (s). For sequence analysis, sequencing of the 3.0 kb fragment
subclone (B38.3) was performed using the standard flanking vector R24 and T7 primers. DNA
sequencing was conducted at the ICBR sequencing facility (University of Florida, Gainesville,
FL) with the Applied Biosystems model 373 system (Foster City, CA).
Plant Material and Plant Inoculations
Grapefruit cv. Duncan (Citrus paradisi) plants were grown from seed in 15 cm plastic pots
in Terra-Lite® agricultural mix (Scott Sierra Horticultural Products Co. Marysville, OH). The
plants were kept in the glasshouse of the University of Florida in Gainesville, Florida at
temperatures ranging from 25-30oC. Tomato plants (Solanum lycopersicum ) cv. Bonny Best
and pepper plants (Capsicum annuum) cv. ECW and its NIL ECW20R containing the Bs2
resistance gene were planted from seeds in Plugmix (W. R. Grace & Co., Cambridge, MA,
USA). After two weeks, the seedlings were transferred to Metromix 300 (W. R. Grace & Co) in
10 cm plastics pots. The plants were kept in the glasshouse of the University of Florida in
Gainesville, Florida at temperature ranging from 25-30oC. For preparation of bacterial
suspensions, 18 h cultures were harvested from the NA plates and suspended in sterile tap water,
and standardized to an optical density at 600 nm (OD600) = 0.3 (5 x 108 CFU/ml) with a
Spectronic 20 Genesys spectrophotometer (Spectronic-UNICAM, Rochester, NY, USA). For
disease symptom assays, bacterial suspensions of the strains used in this study were infiltrated at
5 x 108 CFU/ml into abaxial surface of citrus leaves by using hypodermic syringe and needle,
and the symptoms were assessed up to 10 days after inoculation. Plant responses were evaluated
after 3-4 days for water-soaking and 6-7 days for pathogenicity. For HR tests, bacterial
suspensions adjusted to 5 x 108 CFU/ml in sterile tap water were infiltrated into leaflets of
tomato cvs. Bonny Best and pepper plants cv. ECW and ECW20R with a hypodermic syringe
and needle. The inoculated plants were incubated in a growth room at 24-28oC and assessed for

58
HR elicitation 24 h after infiltration for HR induction. Disease scores are the means of those for
three leaves. All the experiments were repeated at least three times.
Measurement of Electrolyte Leakage
Electrolyte leakage assessments were determined for different strains to evaluate disease
progress and amount of tissue damage after inoculation. Bacterial suspension, adjusted to 3 x 108
CFU/ml in sterile tap water, was injected into tomato leaflets. Inoculated plants were maintained
at 25-28oC in the growth room. Following inoculation and 24 and 48 h after inoculation, 3-cm2
of inoculated leaf tissue were transferred to 3 ml of de-ionized water in 16 x 100-mm test tubes
and the initial conductivity reading. The samples were then placed under vacuum for 1 min,
shaken for 1 h at 28oC, and then vortexed briefly before a second (final) electrolyte reading was
made. Leakage was reported as the difference in the two readings for each sample as micro-
omhs. The readings were recorded for each sample using a Model 31 conductivity bridge (YSI
Instrument Co., Inc., Yellow Springs, OH, USA).
Protein Thermal Stability
To estimate thermal stability of the elicitor activity due to the B38 subclone in 91-118,
sterile tap-water bacterial suspensions of 91::B38 and 91::Ωhrp mutant harboring B38 clone
(91Ωhrp::B38) at 5 x 108 CFU/ml were boiled in hot water for 10 min. The boiled bacterial
suspensions were infiltrated into tomato plants with hypodermic syringe and needle. For control,
part of the bacterial suspensions listed above was left untreated and infiltrated into tomato leaves
at the same time. The plants were kept in the growth room for 24 to 72 hr at 28oC.
Results
Unusual Incompatible Responses of Mutants 306::ΩhrpG and 306::ΩhrpX in Tomato
The wild-type Xcc 306 and the mutants 306::Ω hrpG and 306::ΩhrpX were infiltrated at 5
x 108 CFU/ml in grapefruit leaves. Water soaked lesions were observed in the leaves inoculated

59
with Xcc 306 wild-type 3 – 4 days after inoculation (DAI), while after 6 – 8 days raised lesions
typical of a compatible interaction were observed (Figure 3-1). In contrast, the infiltrated areas
where the 306::Ω hrpG and 306::ΩhrpX were injected were without any sign of citrus canker
symptoms (Figure 3-1). Additionally, the ability to incite citrus canker symptoms was
complemented by the complementation with the wild type genes. The 306::ΩhrpG and 306::
ΩhrpX mutants were inoculated at 5 x 108 CFU/ml into tomato cultivar Bonny Best leaves and
pepper cultivars ECW and ECW20R leaves. Neither mutant lost the ability to trigger HR in
tomato (Figure 3-2) 24 h after inoculation. The HR induced by the mutants was compared with
the HR generated by the Xcc wild-type and no macroscopic difference was observed (Figure 3-
2).
Subclone of X. f. pv. aurantifolli Confers the Ablilty to Trigger an HR in Tomato
In order to investigate genes involved in non-host defense activation we constructed a
genomic library of the wild type X. f. pv. aurantifoli and it was conjugated into Xanthomonas
perforans strain 91-118. All the transconjugants were inoculated on tomato to find HR positive
clones. Subsequently, the HR positive clones were then conjugated into X. perforans strain 91-
118 hrp (91::Ωhrp) mutant to identify genes that were able to trigger HR in the T3SS-
independent manner. Clone pL450 when expressed in 91::Ωhrp mutant retained the ability to
induce cell death in tomato plants. The 91::Ωhrp mutant was selected for use in this experiment
because the mutant was non-pathogenic on tomato cv. Bonny Best due to core deletions in the
hrp cluster. After screening for HR positive transconjugants in tomato plants, one clone
(designed pL450) was selected and used as a model to isolate the HR-elicitor in Xcc for further
experiments. The 91::Ωhrp mutant harboring pL450 clone (91Ωhrp::450) was infiltrated into
tomato plants at concentration of 5 x 108 CFU/ml. On Bonny Best plants a typical HR was noted

60
for the 91Ωhrp::450 mutant 24 h after infiltration; as expected, the 91::Ωhrp mutant did not
show any signs of compatible or incompatible interaction (Figure 3-3). When pepper plants
(cultivars ECW and ECW-20R) were challenged with 91Ωhrp::450 mutant at the same bacterial
concentration used above, neither, ECW or ECW20R infiltrated leaves developed HR 24 h after
inoculation (Figure 3-3).
Electrolyte leakage in tomato plants was performed to measure, quantitatively, the amount
of tissue damage caused by each strain. In the first experiment, we compared the 91-118 wild-
type (WT) by itself and carrying pLAFR3 and pL450. The electrolyte leakage patterns for
91::pLAFR3 and the WT were similar but the 91-118 carrying pL450 showed high electrolyte
leakage 48 h after infiltration compared with the controls (Figure 3-4a). We analyzed the second
group, 91Ωhrp; 91Ωhrp::pLAFR3; 91Ωhrp:: 450; 91Ωhrp::XV9, Ωhrp mutant complemented
with pXV9 clone. The electrolyte leakage patterns of 91Ωhrp::XV9, although higher, and
91Ωhrp::pL450 were similar and characteristic of an HR, while the pattern for 91Ωhrp and
91Ωhrp::pLAFR3 were similar to a compatible interaction (Figure 3-4b).
Sequencing Analysis Revealed Novel Genes Controlling Nonhost HR Elicitor
The pL450 clone was estimated in length at ~31 kb based on multiple restriction enzyme
digestion analyses (data not shown). Digestion of pL450 clone with BamH I generated many
different size fragments, which were cloned into pUFR034 and transformed by triparental mating
into X. perforans strain 91-118 wild-type and a Ωhrp- mutant. The transconjugants were
infiltrated into tomato leaflets to assess their ability to direct a nonhost HR. The 91-118 wild type
and Ωhrp- mutant transconjugants with subclone pB38 containing a 4.5 kb insert elicit an HR in
tomato plants but not in pepper plants (Figure 3-5). Additionally, pB38 clone was transformed by
triparental mating into Xanthomonas strain INA42, which is an opportunistic bacterium isolated

61
from orange and not pathogenic on tomato; furthermore, the strain does not induce a nonhost HR
(Jones et al., 1993). The transconjugant of INA42::B38, which contained the clone pB38,
induced an HR in tomato plants but not in pepper (Figure 3-5). A restriction enzyme analysis
with BamHI and EcoRI of the pB38 subclone revealed two fragments of 1.5 Kb and 3.0 Kb,
respectively. The fragments were further subcloned into pUFR034 resulting in pB38-1 (1.5 Kb)
and pB38-3 (3.0 Kb). The clones were introduced by into the X. perforans Ωhrp mutant and the
transconjugants were tested for the nonhost HR phenotype. The results showed that only the
91Ωhrp::B38-3, carrying the pB38-3 clone, triggered an HR on tomato.
The ends of the pB38-3 plasmid were sequenced, and a comparison to the genome of Xcc
306 revealed that 5’ end was 93% identical with ORF XAC3855 and another end showed 94%
identity with XAC3859. Due to the fact that the homology in the XAC3855 started in the middle
of the gene, we chose to work with the following ORFs, XAC3857, XAC3858 and XAC3859. A
sequence homology search of all three HR-elicitor predicted proteins was carried out with the
NCBI-Blastp program (http://blast.ncbi.nlm.nih.gov/Blast.cgi). The output of the search
indicated that XAC3857 showed high homology to many other Xanthomonas spp. conserved
hypothetical proteins (CHP), including XCV3975 (96% identity and 98% similarity) from Xcv
strain 85-10; CHP (90% identity and 95% similarity) from X. o. pv. oryzicola strain BLS256,
CHP (89% identity and 93% similarity) from Xoo strain POX99A; and CHP (84% identity and
94% similarity) from X. campestris pv. campestris strain ATCC33913. To better visualize the
homology, we used the Clustal W alignment program to demonstrate similarity. In addition, the
protein sequence translated from the nucleotide sequence of pCR3857 clone was used to search
for homology in other Xanthomonas spp. (Figure 3-6). XAC3858 showed very similar high
homology with different Xanthomonas spp. conserved hypothetical proteins (Figure 3-7). ORF

62
XAC3859 exihibited high homology to D-alanyl-D-alanine dipeptidase precursor, including to
D-alanyl-D-alanine dipeptidase precursor (92% identity and 95% similarity) from Xcv strain 85-
10; D-alanyl-D-alanine dipeptidase (82% identity and 89% similarity) from Xoo strain POX99A;
D-alanyl-D-alanine dipeptidase (84% identity and 91% similarity) from X. o. pv. oryzicola strain
BLS256; and vanX (72% identity and 79% similarity) from X. c. pv. campestris strain B100.
Additionally, this gene is part of VanY superfamily, which is related to vancomycin resistance
(Evers and Courvalin, 1996; Bussiere et al,. 1998; Lee et al., 2000). We also search for the
presence plant-inducible promoter box (PIP box) sequence upstream of the XAC3859, which
was not found. Moreover, the translated protein sequence showed certain level of homology with
other Xanthomonas spp. (Figure 3-8). Based on the DNA sequence of B38-3 subclone, where the
tree genes described above are presented, and the transcriptional organization (Figure 3-9), we
can speculate that these genes could be part of an operon due the fact that all the genes are the
same direction.
Discussion
Besides the fact that the hrp-regulators share a degree of homology among some important
plant pathogenic bacteria, most hrp-regulatory mutants are totally impaired with disease
symptoms, bacterial growth and HR elicitation (Wei et al., 1992; Xiao et al., 1994; Wengelnik et
al., 1995; Wengelnik and Bonas, 1996; Brito et al., 1999; Sneath et al., 1990). In this study, the
ability of Xcc to induce symptoms on highly susceptible grapefruit was dependent on a
functional T3SS, which corroborates with the previous studies mentioned above. However, the
ability of 306::ΩhrpG and 306::ΩhrpX mutants to induce an HR on non-host plants is unusual
with respect to hrp mutants. The infiltration of 306::ΩhrpG and 306::ΩhrpX mutants into non-
host tomato plants induced strong hypersensitive responses. This phenotype was reported with a

63
hrcC mutant on P. syringae pv. tabaci, which completely lost pathogenicity on tobacco leaves,
but still induce a strong HR in non-host tomato plants. The HR-elicitor for the hrcC mutant was
determined to be flagellin protein (Marutani et al., 2004).
A parallel study in this laboratory demonstrated that a clone, pXfa450, from X. fuscans pv.
aurantifolli (Xfa) genomic library elicited an HR in tomato plants in the T3SS-independent
manner. Speculating that the genome sequence between X. citri subsp. citri and X. f. pv.
aurantifolli are similar, we used the 450 clone to identify the unknown T3SS-independent HR-
elicitor from X. f. pv. aurantolli. In the search for the Xcc T3SS-independent HR clone, a series
of subclones from the 450 cosmid were isolated into X. perforans strain 91-118 hrp mutant
(91Ωhrp). A 3.0 Kb subclone was identified and called B38-3, which was able to elicit the
nonhost HR phenotype. The sequence analysis of the B38-3 demonstrated that two ORFs,
XAC3857 and XAC3858, are homologous to conserved hypothetical proteins in other
Xanthomonas spp. but have no association with known HR-elicitor was observed. The third
gene, XAC3859 contains a conserved domain, which is part of the VanY superfamily and
involved in resistance to antibiotic, vancomycin (Evers and Courvalin, 1996; Bussiere et al,.
1998; Lee et al., 2000); and it is highly conserved in other Xanthomonas spp. This implies that
this phenotype, probably, is not connected with flagella, or flagella-related proteins. Moreover,
the transcriptional region where the genes are located showed that all 10 genes possess the same
orientation. This may indicate that these genes are part of an operon and the phenotype is linked
to two or more genes, instead of a single gene. However, in order to confirm this hypothesis
mutational analysis, creating a polar affect, must be performed.

64
Table 3-1. Bacterial strains and plasmids used in the study Strain or plasmid Relevant characteristics Source Strains Xanthomonas citri subsp. citri 306 Wild type, Asiatic strain, isolated in
Brazil, RifR DPIa
306::ΩhrpG hrpG , single recombinant of pCRhrpG, KnR, AmpR
This study
306::ΩhrpX hrpX , single recombinant of pCRhrpX, KnR, AmpR
This study
ΩhrpG::XV9 hrpG , complemented with pXV9, KnR, AmpR, TcR
This study
ΩhrpX::XV9 hrpX , complemented with pLHrpG/X, KnR, AmpR, TcR
This study
Xanthomonas perforans 91-118 Wild type, pathogenic to tomato, RifR Jones et al.
(2004) 91::450 91-118 carrying the 450 cosmid This study 91Ωhrp::450 91-118 defective T3SS carrying the 450
cosmid This study
91Ωhrp::Bs3 91-118 defective T3SS carrying a AvrBs3 homolog
This study
91::B38 91-118 carrying the B38 plasmid This study 91:: Ωhrp 91-118 defective T3SS This study 91Ωhrp::B38 91-118 defective T3SS carrying the B38
plasmid This study
91::B38-3 91-118 carrying the B38-3 plasmid This study X. fuscans pv. aurantifolli Xfa-C 5979 Xc 70, isolated in Brazil DPI Opportunistic xanthomononad INA42 Isolated in orange R. E. Stallb
INA42::B38 4.5 Kb Xaa fragment cloned into BamH I pUF034
R. E. Stall
Escherichia coli DH5α FrecAϕ80dlacZ∆M15 Invitrogen
Plasmids
pCR2.1-TOPO Phagemid, CbR, KnR Invitrogen pBluescript-KS+/- Phagemid, pUC derivative; AmpR Stratagene pRK2013 KnR, tra+, mob+ Daniels et al.
(1984) pLAFR3 TcR, rlx+, RK2 replicon BJSc
pUFR034 KnR, Tn903, IncW, Mob+ De Feyter et al. (1990)
pCRhrpG hrpG partial fragment in pCR2.1-TOPO This study pCRhrpX hrpX partial fragment in pCR2.1-TOPO This study pCR3857 3857 ORF and endogenous promoter in This study

65
Table 3-1. Continued pCR2.1-TOPO pCR3859 3859 ORF and endogenous promoter in
pCR2.1-TOPO This study
pXV9 ~25kb of hrp cluster of X. c. pv. vesicatoria 75-3 cloned in pLAFR3
Bonas et al. (1991)
pXfa450 32kb Xaa fragment cloned into pLAFR3 G. Minsavaged
B38 4.5 Kb Xaa fragment cloned into BamH I pUF034
This study
B38.3 3.0 Kb Xaa fragment cloned into BamH I and EcoR I pUF034
This study
B38.1 1.5 Kb Xaa fragment cloned into BamH I and EcoR I pUF034
This study
a DPI, Division of Plant Industry of the Florida Department of Agriculture and Consumer Services, Gainesville, FL, USA. b R.E. Stall, Robert E. Stall, University of Florida, Gainesville, FL. c BJS, B. J. Staskawicz, University of California, Berkley, CA. d G. Minsavage, Gerald Minsavage, University of Florida, Gainesville, FL.

66
Figure 3-1. Phenotype of grapefruit leaves were used to distinguish among the strains. Strains were infiltrated into the leaves at concentrations of 5 x 108 CFU/mL. Labeling: (1) ΩhrpG::XV9; (2) 306::ΩhrpG; (3) Xcc strain 306 – wild type; (4) 306::ΩhrpX; (5) ΩhrpX::XV9. The grapefruit leaves were photographed 7 days after infiltration.
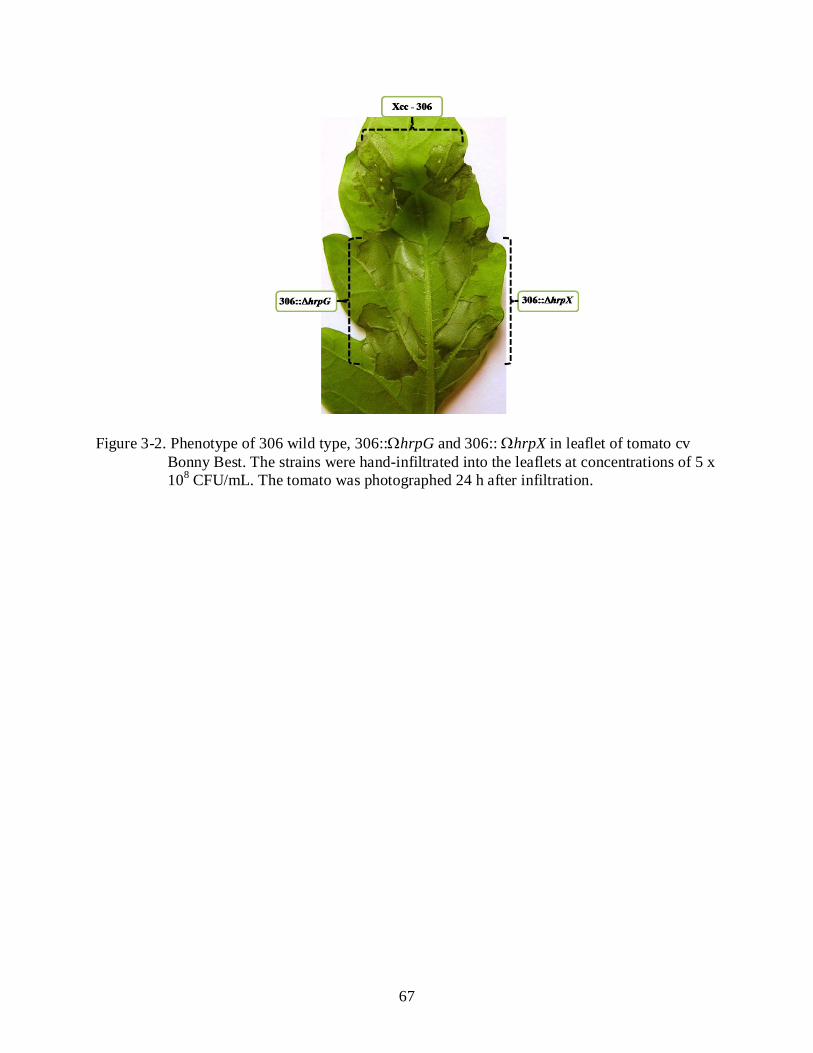
67
Figure 3-2. Phenotype of 306 wild type, 306::ΩhrpG and 306:: ΩhrpX in leaflet of tomato cv Bonny Best. The strains were hand-infiltrated into the leaflets at concentrations of 5 x 108 CFU/mL. The tomato was photographed 24 h after infiltration.

68
Figure 3-3. Hypersensitive response assay in tomato cv. Bonny Best and pepper cv. ECW-20R (A) left: 91Ωhrp::450; right: 91::450. (B) left: 91Ωhrp::450; right-down: 91-118 – wild-type; right-up: 91Ωhrp. The strains were infiltrated into the leaves at concentrations of 5 x 108 CFU/mL. The tomato and pepper leaves were photographed 24 h after infiltration.
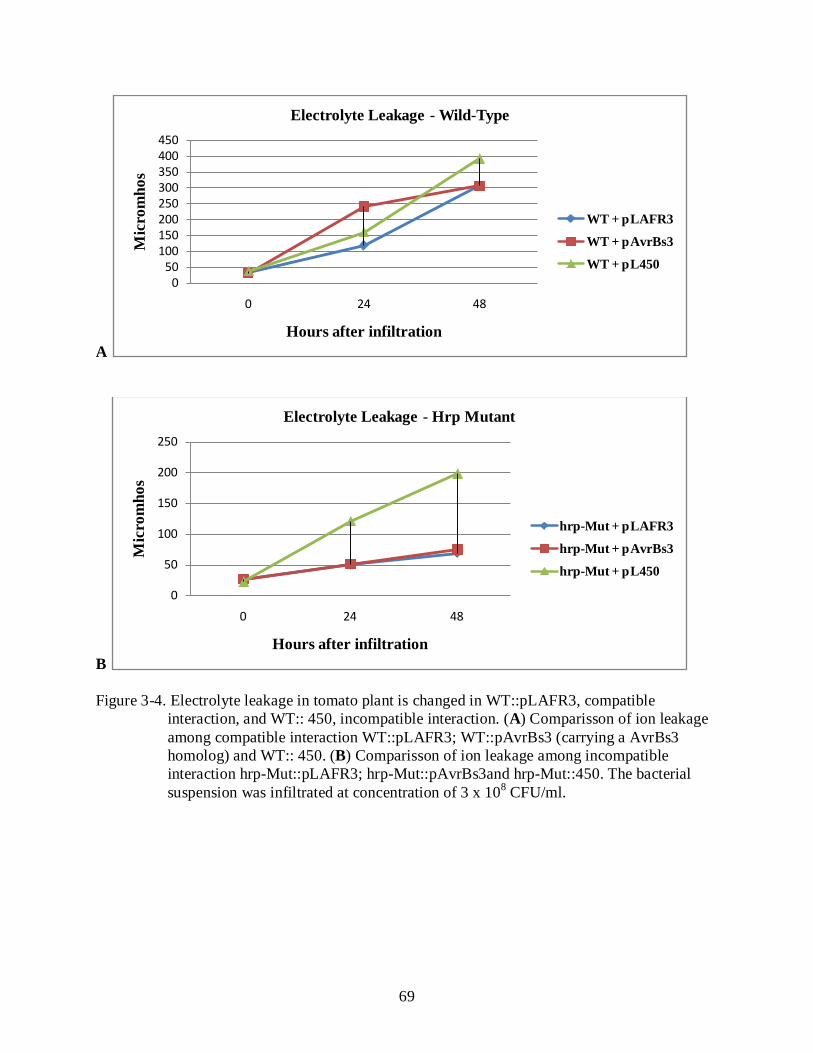
69
A
B Figure 3-4. Electrolyte leakage in tomato plant is changed in WT::pLAFR3, compatible
interaction, and WT:: 450, incompatible interaction. (A) Comparisson of ion leakage among compatible interaction WT::pLAFR3; WT::pAvrBs3 (carrying a AvrBs3 homolog) and WT:: 450. (B) Comparisson of ion leakage among incompatible interaction hrp-Mut::pLAFR3; hrp-Mut::pAvrBs3and hrp-Mut::450. The bacterial suspension was infiltrated at concentration of 3 x 108 CFU/ml.
050
100150200250300350400450
0 24 48
Mic
rom
hos
Hours after infiltration
Electrolyte Leakage - Wild-Type
WT + pLAFR3
WT + pAvrBs3
WT + pL450
0
50
100
150
200
250
0 24 48
Mic
rom
hos
Hours after infiltration
Electrolyte Leakage - Hrp Mutant
hrp-Mut + pLAFR3
hrp-Mut + pAvrBs3
hrp-Mut + pL450

70
Figure 3-5. Subclone, B38, triggers hypersensitive response (HR) independent of T3SS in X. perforans in non-host plants, tomato cv. Bonny Best and pepper cv. ECW20R. (A) left: 91::38 (open arrow); right: 91-118 wild-type (open arrow); left: 91Ωhrp::B38 (close arrow); right: 91::Ωhrp (close arrow). (B) left: 91Ωhrp::B38 (open arrow); right: 91::Ωhrp (open arrow); left: OPP::B38 (close arrow) ; right: OPP-INA42 (close arrow). (C) 1. 91-118 wild-type; 2. 91::B38; 3. 91::Ωhrp; 4. 91Ωhrp::B38; 5. OPP-INA42; 6. OPP::B38. The strains were infiltrated into the leaves at concentrations of 5 x 108 CFU/mL. The tomato and pepper leaves were photographed 24 h after infiltration.

71
XAC3857 (A) MNLPLHFGLLGSLEAGLIALILGVVVFAAVERGGRRLQFNHGHTLGIACLLAVAIGAGYDIWHLVYTSIV RLESPLYARLALARIHDPNELGSRVVLEVAGALAGVVLGWKLFSSGGWENDPPSA (B) XOOBLS MNLPLHFGLLGSLEAGLIALVLGVLVFAAVERIGRRLQFNHGQTLGIACLLTVAIGAGYD 60 XOO99 MNLPLHFGLLGSLEAGLIALILGVLVFAAVGRIGRRLQFNHGQMLGIACLLTVAIGAGYD 60 XCC306 MNLPLHFGLLGSLEAGLIALILGVVVFAAVERGGRRLQFNHGHTLGIACLLAVAIGAGYD 60 3857 MNLPLHFGLLGSLEAGLIALILGVVVFAAVERGGRRLQFNHGHTLGIACLLAVAIGAGYD 60 XCV85-10 MNLPLHFGLLGSLEAGLIALVLGVVVFAAVERGGRRLQFNHGHTLGIACLLAVAIGAGYD 60 XCC33913 MNLPLHFGLLGSLEAGLIALVIGVLLFAGVEHLGRRLQFTHGHTLGIACLLAVAIGAGYD 60 ********************::**::**.* : ******.**: *******:******** XOOBLS IWNLVYTSIVRLESPLYARLALAKIHDPNELGSRVVLDVAGALAGVVLGWKLFSSGSWDK 120 XOO99 IWNLVYTSIVRLESPLYARLALAKIHDPNELGSRVVLDVAGALAGVVLGWKLFSSGSWDK 120 XCC306 IWHLVYTSIVRLESPLYARLALARIHDPNELGSRVVLEVAGALAGVVLGWKLFSSGGWEN 120 3857 IWHLVYTSIVRLESPLYARLALARIHDPNELGSRVVLEVAGALAGVVLGWKLFSSGGWEN 120 XCV85-10 IWHLVYTSIVRLESPLYAHLALAKIHDPNELGSRVVLEVAGALAGVVLGWKLFSSGSWEN 120 XCC33913 MWHLVYTSVVRLESPLYARVALAKIHDPNELGSRVVLEVAGAIAGVVLGWRLFSSATWDD 120 :*:*****:*********::***:*************:****:*******:****. *:. XOOBLS DAPSA 125 XOO99 DAPSA 125 XCC306 DPPSA 125 3857 DPPSA 125 XCV85-10 DPPSA 125 XCC33913 DAPAA 125 *.*:*
Figure 3-6. Protein alignment of XAC3857 gene and Xanthomonas spp. (A) Original protein sequence of the ORF XAC3857 from X. citri subsp. citri strain 306; (B) Protein alignment among X. citri subsp. citri strain 306 (XCC306), XAC3857 clone (3857), X. campestris pv. vesicatoria strain 85-10 (XCV85-10), X. oryzae subsp. oryzicola strain BLS256 (XOOBLS), X. oryzae subsp. oryzae strain POX99A (XOO99), and X. campestris subsp. campestris strain 33913 (XCC333913). Asterisks indicate that the amino acids in that column are identical in all sequences. Colon shows that conserved amino acids substitutions have been observed. Dot means that semi-conserved substitutions are observed.

72
XAC3858 (A) MSDPERFVDIACPYCGEWITLALDLTGGDQHYIEDCQVCCKPIAVSVRWDEEGEAQVSARGQDDA (B) XCC306 MSDPERFVDIACPYCGEWITLALDLTGGDQHYIEDCQVCCKPIAVSVRWDEEGEAQVSAR 60 XCV85-10 MSDPERFVDIACPYCGEWITLALDLTGGDQHYIEDCQVCCKPIAVSVRWDEEGEAQVTAR 60 XOO99 MSDPERFVDIACPYCGEWITLALDLTGGDQHYIEDCQVCCKPISVRVRWDEEGEAQVSAR 60 XOOBLS MSDPERFVDIACPYCGEWITLALDLTGGDQHYIEDCQVCCKPISVRVRWDEEGEAQVSAR 60 XCC8004 MSDPERFVDIACPYCGEWITLALDLSGGDQHYIEDCQVCCKPISVSVQWDEEGEAQVSAR 60 XCC33913 MSDPERFVDIACPYCGEWITLVLDLSGGDQHYIEDCQVCCKPISVSVQWDEEGEAQVSAR 60 *********************.***:*****************:* *:*********:** XCC306 GQDDA 65 XCV85-10 GQDDA 65 XOO99 GQDDA 65 XOOBLS GQDDA 65 XCC8004 GQDDA 65 XCC33913 GQDDA 65 ***** Figure 3-7. Protein alignment of XAC3858 gene and Xanthomonas spp. (A) Original protein
sequence of the ORF XAC3858 from X. citri subsp. citri strain 306; (B) Protein alignment among X. citri subsp. citri strain 306 (XCC306), X. campestris pv. vesicatoria strain 85-10 (XCV85-10), X. oryzae subsp. oryzae strain POX99A (XOO99), X. oryzae subsp. oryzicola strain BLS256 (XOOBLS), X. campestris subsp. campestris strain 8004 (XCC8004), and X. campestris subsp. campestris strain 33913 (XCC333913). Asterisks indicate that the amino acids in that column are identical in all sequences. Colon shows that conserved amino acids substitutions have been observed. Dot means that semi-conserved substitutions are observed.
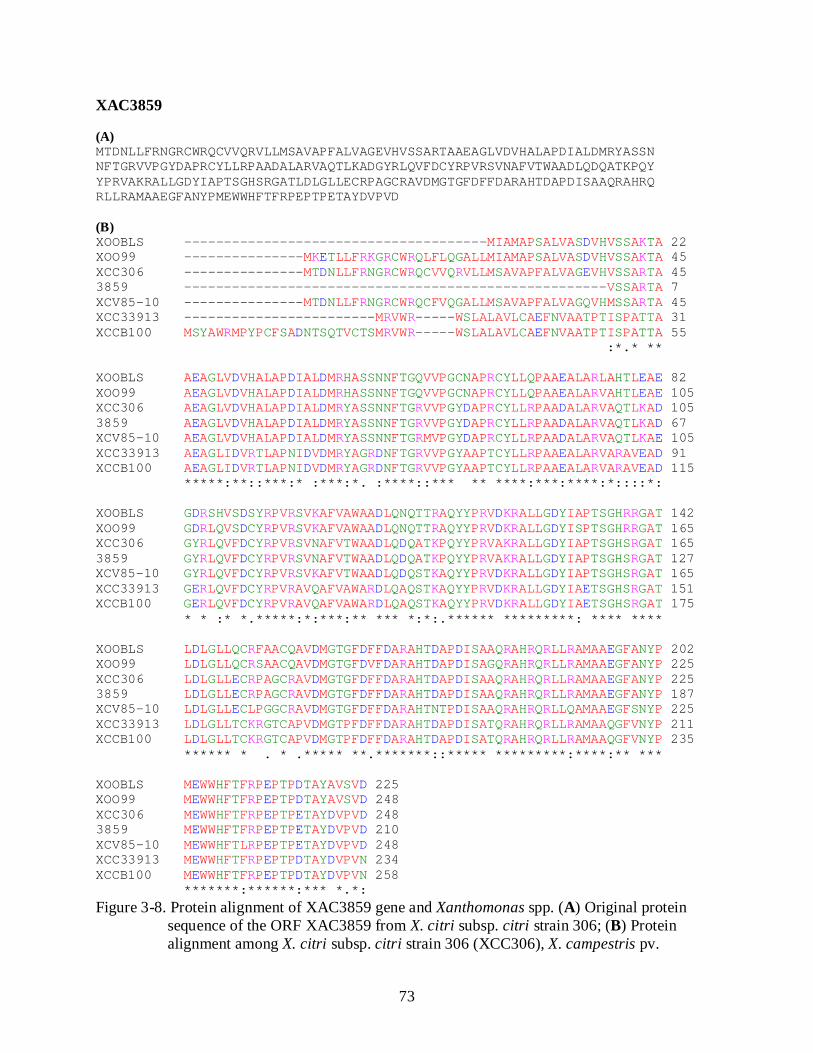
73
XAC3859 (A) MTDNLLFRNGRCWRQCVVQRVLLMSAVAPFALVAGEVHVSSARTAAEAGLVDVHALAPDIALDMRYASSN NFTGRVVPGYDAPRCYLLRPAADALARVAQTLKADGYRLQVFDCYRPVRSVNAFVTWAADLQDQATKPQY YPRVAKRALLGDYIAPTSGHSRGATLDLGLLECRPAGCRAVDMGTGFDFFDARAHTDAPDISAAQRAHRQ RLLRAMAAEGFANYPMEWWHFTFRPEPTPETAYDVPVD (B) XOOBLS --------------------------------------MIAMAPSALVASDVHVSSAKTA 22 XOO99 ---------------MKETLLFRKGRCWRQLFLQGALLMIAMAPSALVASDVHVSSAKTA 45 XCC306 ---------------MTDNLLFRNGRCWRQCVVQRVLLMSAVAPFALVAGEVHVSSARTA 45 3859 -----------------------------------------------------VSSARTA 7 XCV85-10 ---------------MTDNLLFRNGRCWRQCFVQGALLMSAVAPFALVAGQVHMSSARTA 45 XCC33913 ------------------------MRVWR-----WSLALAVLCAEFNVAATPTISPATTA 31 XCCB100 MSYAWRMPYPCFSADNTSQTVCTSMRVWR-----WSLALAVLCAEFNVAATPTISPATTA 55 :*.* ** XOOBLS AEAGLVDVHALAPDIALDMRHASSNNFTGQVVPGCNAPRCYLLQPAAEALARLAHTLEAE 82 XOO99 AEAGLVDVHALAPDIALDMRHASSNNFTGQVVPGCNAPRCYLLQPAAEALARVAHTLEAE 105 XCC306 AEAGLVDVHALAPDIALDMRYASSNNFTGRVVPGYDAPRCYLLRPAADALARVAQTLKAD 105 3859 AEAGLVDVHALAPDIALDMRYASSNNFTGRVVPGYDAPRCYLLRPAADALARVAQTLKAD 67 XCV85-10 AEAGLVDVHALAPDIALDMRYASSNNFTGRMVPGYDAPRCYLLRPAADALARVAQTLKAE 105 XCC33913 AEAGLIDVRTLAPNIDVDMRYAGRDNFTGRVVPGYAAPTCYLLRPAAEALARVARAVEAD 91 XCCB100 AEAGLIDVRTLAPNIDVDMRYAGRDNFTGRVVPGYAAPTCYLLRPAAEALARVARAVEAD 115 *****:**::***:* :***:*. :****::*** ** ****:***:****:*::::*: XOOBLS GDRSHVSDSYRPVRSVKAFVAWAADLQNQTTRAQYYPRVDKRALLGDYIAPTSGHRRGAT 142 XOO99 GDRLQVSDCYRPVRSVKAFVAWAADLQNQTTRAQYYPRVDKRALLGDYISPTSGHRRGAT 165 XCC306 GYRLQVFDCYRPVRSVNAFVTWAADLQDQATKPQYYPRVAKRALLGDYIAPTSGHSRGAT 165 3859 GYRLQVFDCYRPVRSVNAFVTWAADLQDQATKPQYYPRVAKRALLGDYIAPTSGHSRGAT 127 XCV85-10 GYRLQVFDCYRPVRSVKAFVTWAADLQDQSTKAQYYPRVDKRALLGDYIAPTSGHSRGAT 165 XCC33913 GERLQVFDCYRPVRAVQAFVAWARDLQAQSTKAQYYPRVDKRALLGDYIAETSGHSRGAT 151 XCCB100 GERLQVFDCYRPVRAVQAFVAWARDLQAQSTKAQYYPRVDKRALLGDYIAETSGHSRGAT 175 * * :* *.*****:*:***:** *** *:*:.****** *********: **** **** XOOBLS LDLGLLQCRFAACQAVDMGTGFDFFDARAHTDAPDISAAQRAHRQRLLRAMAAEGFANYP 202 XOO99 LDLGLLQCRSAACQAVDMGTGFDVFDARAHTDAPDISAGQRAHRQRLLRAMAAEGFANYP 225 XCC306 LDLGLLECRPAGCRAVDMGTGFDFFDARAHTDAPDISAAQRAHRQRLLRAMAAEGFANYP 225 3859 LDLGLLECRPAGCRAVDMGTGFDFFDARAHTDAPDISAAQRAHRQRLLRAMAAEGFANYP 187 XCV85-10 LDLGLLECLPGGCRAVDMGTGFDFFDARAHTNTPDISAAQRAHRQRLLQAMAAEGFSNYP 225 XCC33913 LDLGLLTCKRGTCAPVDMGTPFDFFDARAHTDAPDISATQRAHRQRLLRAMAAQGFVNYP 211 XCCB100 LDLGLLTCKRGTCAPVDMGTPFDFFDARAHTDAPDISATQRAHRQRLLRAMAAQGFVNYP 235 ****** * . * .***** **.*******::***** *********:****:** *** XOOBLS MEWWHFTFRPEPTPDTAYAVSVD 225 XOO99 MEWWHFTFRPEPTPDTAYAVSVD 248 XCC306 MEWWHFTFRPEPTPETAYDVPVD 248 3859 MEWWHFTFRPEPTPETAYDVPVD 210 XCV85-10 MEWWHFTLRPEPTPETAYDVPVD 248 XCC33913 MEWWHFTFRPEPTPDTAYDVPVN 234 XCCB100 MEWWHFTFRPEPTPDTAYDVPVN 258 *******:******:*** *.*: Figure 3-8. Protein alignment of XAC3859 gene and Xanthomonas spp. (A) Original protein
sequence of the ORF XAC3859 from X. citri subsp. citri strain 306; (B) Protein alignment among X. citri subsp. citri strain 306 (XCC306), X. campestris pv.

74
Figure 3-8.Continued vesicatoria strain 85-10 (XCV85-10), X. oryzae subsp. oryzae strain POX99A (XOO99), X. oryzae subsp. oryzicola strain BLS256 (XOOBLS), X. campestris subsp. campestris strain B100 (XCCB100), and X. campestris subsp. campestris strain 33913 (XCC333913). Asterisks indicate that the amino acids in that column are identical in all sequences. Colon shows that conserved amino acids substitutions have been observed. Dot means that semi-conserved substitutions are observed.

75
Figure 3-9. Transcriptional organization of the ORFs XAC3857 to XAC3859 region in
Xanthomonas citri subsp. citri. The region contains11 genes. The arrowheads indicate direction of transcription.

76
CHAPTER 4 GENETIC CHARACTERIZATION OF HRPW OF Xanthomonas citri subsp. citri
Introduction
The Gram-negative bacterium, Xanthomonas citri pv. citri (Xcc), is a plant pathogen that
causes citrus canker diseases in most citrus cultivars (Stall and Civerolo, 1991; Schaad et al.,
2006). Many pathogenic proteobacteria rely on the type III secretion system (T3SS), encoded by
hypersensitive reaction and pathogenicity (hrp) genes, to secrete bacterial effector proteins into
the extracellular milieu and translocate them into the eukaryotic host cell cytosol and
consequently modulate host physiology (Chang et al., 2004; Tang et al., 2006). To deliver these
T3-effector proteins into host cells three biological barriers must be crossed, the membranes in
the host and the inner and outer bacterial. This process involves the T3SS pore-forming
translocon complex (Cornelis, 2006). In Xanthomonas spp. the hrpF is a typical translocon
component involved in pore-formation in the planar lipid bilayers but it is not required for
secretion (Rossier et al., 2000; Buttner et al., 2002). Moreover, pathogens carrying a defective
translocon are impaired in T3-effectors translocation, thus, inhibiting disease progress (Rossier et
al., 2000; Sugio et al., 2005; Cornelis, 2006)
Lorenz et al. (2008) classified the T3S substrates in two groups: (I) extracellular
components of the secretion apparatus, and (II) effector proteins that are translocated into the
host cell modifying cells metabolism (He et al., 2004; Mudgett, 2005). One class of proteins that
crosses the Hrp type III secretion apparatus is composed of the harpin proteins, which are T3SS-
dependent and secreted in the extracellular milieu during interaction (Perino et al., 1999;
Tampakaki and Panopoulos, 2000). Harpin proteins are unique to phytopathogens and do not
show homology to any other known protein, heat stable, glycine rich and lack of cysteine
residues. Importantly, harpins trigger hypersensitive response (HR) cell death in non-host plants

77
and may contribute to effector translocation (Wei et al., 1992; He et al., 1993; Arlat et al., 1994;
Charkowski et al., 1998; Kim et al., 2004; Kvitko et al., 2007). Additionally, in recent studies
the harpin from Erwinia amylovora (HrpN) is weakly translocated into tobacco cells (Bocsanczy
et al., 2008); purified harpin modulates the accumulation of salicylic acid (Clarke et al., 2005);
induces activation of mitogen-activated protein kinases (MAPKs) (Desikan et al., 2001),
elevation of cytosolic Ca2+ (Cessna et al., 2001), activation of active oxygen species (AOS) and
ion flux modulation (Reboutier et al., 2007).
Since the first harpin, HrpN, was isolated from E. amylovora (Wei et al., 1992), many
harpin-like proteins or genes have been identified from diverse plant pathogenic bacteria,
including Hpa1 from X. oryzae pv. oryzae (Zhu et al., 2000), HpaG from X. a. pv. glycines and
X. c. pv. vesicatoria (Kim et al., 2003; Thieme et al., 2005), and PopA1 from R. solanacearum
(Arlat et al., 1994), One of the most widely distributed members of the harpin-like protein group
is HrpW. HrpW was first described in E. amylovora, and subsequently identified in P. s. pv.
tomato and Xcc (Charkowski et al., 1998; Kim and Beer 1998; da Silva et al., 2002). HrpW
contains a C-terminal domain homologous to class III pectate lyase (PEL) and an N-terminal
harpin-like domain. While located within the respective hrp gene clusters of E. amylovora and P.
syringae, the hrpW gene of Xcc strain 306 is located downstream of the hrp cluster (da Silva et
al., 2002). The role of harpin-like proteins, including HrpW, play in bacterial pathogenisis or
whether or not HrpW is even required for pathogenesis in Xcc is unknown. Therefore, a genetic
and biochemical analysis of the hrpW gene of Xcc was undertaken. Additionally, because it was
thought that hrpW was important, was hypothesized that hrpW is conserved among diverse X.
citri strains. To address this hypothesis hrpW sequences of X. citri subsp. citri were produced
and compared along with similar sequences derived from different X. citri strains.

78
Material and Methods
Bacterial Strains and Plasmids
Bacterial strains and plasmids used in this study are listed in Table 4-1.
Media and Growth Conditions
Escherichia coli cells were grown in at 37oC in Luria-Bertani (LB) medium (1% (w/v)
tryptone, 0.5% (w/v) yeast extract, 1% (w/v) sodium chloride, pH 7.5). For solid media BD
Bacto agar was added at 15 g l-1. Xanthomonas strains were cultivated at 28oC on nutrient agar
(NA) medium and on tryptone sucrose agar (Zhu et al., 2000). All bacterial strains used in this
study were stored in 20% glycerol in sterile tap water and maintained at -80oC. Triparental
matings were performed on nutritent-yeast extract-glycerol (NYG) agar (Daniels et al., 1984).
Antibiotics were used in the following concentrations: ampicillin (Amp) 100 µg ml-1; kanamycin
(Kn) 50 µg ml-1; rifampicin (Rif) 100 µg ml-1; spectinomycin (Spc) 100 µg ml-1; and tetracycline
(Tc) 12.5 µg ml-1.
Recombinant DNA Techniques
Standard methods were used for DNA cloning, restriction mapping and gel electrophoresis
as described by Sambrook et al. (1989) Restriction enzymes, T4 DNA ligase, and GoTaq® Flexi
DNA Polymerase were used following the manufacturer’s recommendations (Promega -
Madison, WI, USA). Chemicals were purchased from Sigma-Aldrich (St. Louis, MO, USA), and
Fisher Scientific (Hampton, NH, USA). PCR was performed in a PCR Express Thermal Cycler
(ThermoHybaid, Ashford, UK). Topo TA cloning kit was purchased from Invitrogen (Carlsbad,
CA, USA). Plasmids were introduced into Escherichia coli DH5α competent cells as described
by Sambrook et al. (1989) and into Xanthomonas spp. by electroporation and conjugation. The
conjugation was performed using pRK2013 as helper plasmid in triparental matings (Daniels et
al., 1984), and the electroporation using aliquots of Xcc competent cells were mixed with

79
plasmid DNA and electroporated using a Bio-Rad electroporation instrument. The conditions
applied were 200 OHMs (resistance), 25 µFD (capacitance) and 2.5 Volts. After the pulse, the
cells were immediately diluted by adding 250 µL of NA medium. The cells were incubate for 1 h
at 28oC with constant shaking, and plated on NA containing kanamycin, then incubated at 28oC
for 2 days (Sun et al., 2003). Insertion mutants in X. c. subsp. citri were generated using single
homologous recombination methodology as described by Sugio et al. (2005) and the deletion
mutants were generated using the suicide vector pOK1 as described by Huguet et al. (1998). The
oligonucleotides used in this study are listed in Table 4-2. All oligonucleotides were designed by
using the Primer3 (v. 0.4.0) program (http://frodo.wi.mit.edu) to minimize secondary structure
and dimer formation and were chemically synthesized by Genomechanix (Gainesville, FL USA).
DNA sequence data were analyzed by DNA Sequencing Core Laboratory of the Interdisciplinary
Center for Biotechnology Research, University of Florida – Gainesville, FL (UF-ICBR).
DNA Sequence Alignment and Phylogenetic Analysis of hrpW Gene
hrpW gene from X. c. subsp. citri and different Xcc strains were sequenced using forward
and reverse, UpHrpWF1 and DHrpWR1, respectively, primers based on the sequence of hrpW
from Xcc genome sequence (da Silva et al., 2002), and designed to cover the entire gene. The
amplified fragments were purified using QIAquick PCR purification Kit (Qiagen). The purified
PCR products were sequenced in both directions at the University of Florida ICBR DNA
Sequencing Core. Nucleotides sequence alignments were done using the Clustal W program.
For phylogenetic studies, we performed a homology search using NCBI-BLAST to
identify hrpW homologs in other bacterium strains besides Xcc. Based on protein level
similarity, the DNA sequence of X. c. pv. campestris strain 33913, R. solanacearum strain
UW551, Acidovorax avenuae subsp. citrulli strain AAC00-1, Pseudomonas viridiflava, Erwina
amylovora, and Erwinia tasmaniensis strain Et1/99 were taken from GenBank accession

80
numbers NP_636593.2, ZP_00943832.1, YP_969269.1, CAA74158.1, and YP_001906489.1,
respectively. The phylogenetic trees were built by first aligning the nucleotide sequence
automatically using Clustal W program and then performing manually alignment using
MacClade (version 4.0, Sinauer Associates). PAUP* was used for analysis and creating
parsimony consensus trees, parsimony bootstrap trees, maximum likelihood (ML) trees, ML
bootstrap trees, and Bayesian consensus trees. Parsimonious trees were created using the default
settings and performing a heuristic search that resulted in one tree. Bootstrap analysis for the
parsimony tree was conducted with 1,000 bootstrapping replicates. For ML analysis, PAUP*
performed a heuristic search under the default ML settings, which resulted in initial tree and the
base likelihood score. Next, the data were analyzed by assessing models for nucleotide
substitution using the ML approach as implemented in the program Modeltest 3.7 (Posada and
Crandall, 1998). A best-fit nucleotide substitution model selection was selected for our set of
aligned sequences, and model selection was conducted on the basis of hierarchal likelihood ratios
tests (hLRT). The best-fit moder from the likelihood settings was the (GTR+G) selected by
Akaike’s information criterion (AIC). ML bootstrap analysis was performed with 1,000
bootstrapping replicates. Bayesian analysis was performed by using Mr. Bayes 3.1 (Huelsenbeck
et al., 2001). Settings for the analysis were set at 1,030,000 generations, sampled at every 1000
generations. The burn-in was set at sample 250 and the consensus trees were created by PAUP*.
Generation of hrpW Deletion Mutants
In-frame deletion derivative of hrpW306∆hrpW was generated by using pOK1 suicide
vector. To generate a full-length deletion of hrpW, divergent primers POWF1 and POWR1 were
used to amplify up- and downstream regions of DNA flanking hrpW from pCR2.1-TOPO clone
pCRHrpW-EP. Each 25 µl PCR contained 5x buffer, 0.7 mM MgCl2, 0.2 mM each

81
deoxynucleotide triphosphate (Promega), 2 pmol of each primer, and 0.04 U of Taq polymerase.
The reaction mixtures were heated for 4 min at 95oC and amplified over 1 cycle of 1 min at
95oC, 1 min at 53oC, and 7 min at 72oC, followed by 30 cycles of 45 sec at 95oC, 30 sec at 60oC,
and 5 min at 72oC. Through the amplification process were added a unique Sal I restriction site,
used to recircularize the construct. The PCR product was purified using QIAquick PCR
Purification Kit system (QIAGEN), and then, directly cloned into pCR2.1-TOPO containing
kanamycin and ampicillin resistance genes following manufacturer’s protocol (Invitrogen). To
construct the pO∆hrpW suicide vector, the deleted region was excised with BamH I and Xba I,
which flanking the pCR2.1 polylinker. The deleted region, a 1.0 Kb - fragment, was cloned into
the suicide vector pOK1 linearized with the same two enzymes. The mutated sequence was then
introduced into the Xcc genome by homologous recombination in two steps described by Huguet
et al. (1998). The recombination events were analyzed by PCR using UpHrpWF1 and
DHrpWR1, which cover the deleted region. In addition, the PCR fragment generated here was
sequenced and the sequence generated was analyzed a multiple alignment tool Clustal-W
(http://www.ebi.ac.uk/Tools/clustalw2/index.html) to confirm the in-frame deletion in hrpW
gene.
Construction of hrpW-Harpin Domain
For the generation of hrpW/harpin domainHrpW∆harpin mutant a 1.2-kb region flanking
hrpW gene was PCR-amplified from Xcc – 306 genomic DNA. The fragment was TOPO-cloned
into pCR2.1 and transformed in E. coli following the manufacturer’s procedure (Invitrogen).
Derivative plasmids carrying the insert were digested with Aat II restriction enzyme, which
deleted 87-bp from the harpin domain, creating the pCR∆harpin. To create the pO∆harpin
suicide vector, the pCR∆harpin was double-digested with BamH I and Xba I, which flank the

82
pCR2.1 polylinker. The deleted fragment was cloned into the suicide vector pOK1 linearized
with the same two enzymes. The mutated sequence was introduced into the Xcc genome by
homologous recombination in two steps described by Huguet et al. (1998), creating
HrpW∆harpin mutant. To confirm the recombination event and the harpin mutation, we PCR-
amplified the deleted region using UpHrpWF1 and DHrpWR1 primers and sequenced the
product. The nucleotide sequences were analyzed by Clustal-W program
(http://www.ebi.ac.uk/Tools/clustalw2/index.html) to identify the deleted region. Blast program
was used for homology searches (Altshul et al., 1997).
Complementation Tests
The hrpW gene carrying the native promoter was PCR amplified using UpHprWF1 and
DoHprWR1 primers. Cloning into pCR2.1-TOPO, creating pCRHrpW-EP, and DH5α
transformation were performed was described above. The digested HrpW-EP fragment was
subcloned from pCR2.1-TOPO by restriction with BamH I and Xba I, and ligated into the BamH
I and Xba I-linearized pOK1 suicide vector to create pOHrpW-EP. E. coli λPIR was transformed
with pOHrpW-EP. The transfomants were introduced in a 306::∆harpin mutant by triparental
mating. Mixing 306::∆harpin mutant cells as recipient with pOHrpW-EP as donor and with
pRK2073 as the conjugational helper on NYG agar. The ratio of recipient to donor to helper was
2:1:1 (vol/vol/vol). After 24 h of incubation at 28oC, the mating mixtures were resuspended in
sterile tap water, and aliquots were spread onto NA plates containing rifamycin and
spectinomycin (100 µg ml-1 and 100 µg ml-1, respectively) for selection of transconjugants.
Transconjugant colonies were subsequently inoculated in grapefruit leaves to screen for
complementation of the pathogenicity.

83
Plant Material
Grapefruit cv. Duncan (Citrus paradisi) plants were grown from seed in 15 cm plastic pots
in Terra-Lite® agricultural mix (Scott Sierra Horticultural Products Co. Marysville, OH). The
plants were kept in the glasshouse of the University of Florida in Gainesville, Florida at
temperature ranging from 25-30oC.
Pepper plants (Capsicum annuum) cv. ECW and the near isogenic line ECW20R
containing the Bs2 resistance gene were planted from seeds in Plugmix (W. R. Grace & Co.,
Cambridge, MA, USA). After two weeks, the seedlings were transferred to Metromix 300 (W. R.
Grace & Co) in 10 cm plastics pots. The plants were kept in the glasshouse of the University of
Florida in Gainesville, Florida at temperature ranging from 25-30oC.
Plant Inoculations
All the Xanthomonas spp. and mutants were cultured on nutrient agar plates for 18 h at
28oC. For preparation of bacterial suspensions, 18 h cultures were harvested from the NA plates
and suspended in sterile tap water, and standardized to an optical density at 600 nm (OD600) =
0.3 (5 x 108 CFU/ml) with a Spectronic 20 Genesys spectrophotometer (Spectronic-UNICAM,
Rochester, NY, USA).
For pathogenicity tests, bacterial suspensions of the wild-type, mutant and complemented
Xcc were infiltrated at 5 x 108 CFU/ml into abaxial surface of citrus leaves by using hypodermic
syringe and needle. Plant responses were evaluated after 3-4 days for water-soaking and 6-7 days
for citrus canker symptoms. The inoculated plants were kept in a growth room at 28-30oC up to
10 days after inoculation.
The sensitivity of the Xcc wild-type and its ∆hrpW mutants was evaluated with pin-prick
inoculation in grapefruit leaves. Bacteria were suspended in sterile tap water at a concentration
of 5 x 108 CFU/ml. A drop of the suspension was placed on the adaxial surface of young

84
grapefruit leaf. Immediately after a needle was pierced through the bacterial suspension and
through the leaves, the inoculum drops were wiped off with sterile cotton. The inoculated plants
were then grown in a glasshouse up to 30 days. The diameter of circular lesions, including cork
tissue and water-soaked margin, was measured after inoculation.
Elicitation of HR was tested by infiltrating bacterial suspensions, which were prepared
with sterile tap water at concentration of 5 x 108 CFU/ml, into leaflets of tomato cv. Bonny Best
and pepper plants cv. ECW and ECW20R with a hypodermic syringe and needle. The inoculated
plants were incubated in a growth room at 24-28oC and assessed 24 h after infiltration for HR
induction. Score are the means of those for three leaves. All the experiments were repeated at
least three times.
Bacterial Populations
Bacterial suspensions with 5 x 108 CFU/mL were infiltrated in grapefruit leaves with a
syringe and 27-gauge needle (Klement, 1963). A 0.5 cm2 of inoculated area from each leaf, each
sample was infiltrated in three leaves, was excised and macerated in 1 mL of sterile tap water, 50
µL were spread onto NA plates. The plates were incubated at 28oC for 2 – 3 days, thus, the
colonies were counted.
Translocation Activity in 306::∆harpin Mutant
A pL799 clone, carrying avrGf1 gene from Xcc strain Aw that triggers HR in grapefruit
(Rybak et al., 2009), was conjugated by triparental mating into Xcc wild-type and HrpW∆harpin
mutant to investigate if the mutation on the harpin domain has any affect in the T3SS
functioning. The transconjugant 306::799 and ∆harpin::799 were inoculated at concentration of 5
x 108 CFU/mL in grapefruit and kept at growth room at 28oC. The plants were assessed for HR
induction up to 5 days after infiltration.

85
Construction of hrpW::avBs2 Fusion
For translocation studies, pepper plants cv. ECW and ECW-20R were infiltrated with X.
campestris pv. vesicatoria strain XV1922 (wild type), 1922::WBs (pUWBs) leading to the
expression of the first 109 amino-acids from hrpW gene fused with avrBs2 C-terminal with 62 –
574 amino acids. The strains were resuspended to an OD600 of 0.3 and infiltrated into the
mesophyll of pepper leaves. 24 hr after infiltration, the inoculated pepper plants were assessed
for HR elicitation.
The plasmid was constructed as follows. First, 327 bp fragment was amplified using the
primers NTW-F1 and NTW-R1 introducing a Bgl II site in both ends. The PCR fragment was
excised from the agarose gel 1% and purified with QIAquick Gel Purification Kit system
(QIAGEN). The digested PCR fragment was cloned into pBS1 (BglII::avrBs262-574::HA) plasmid
resulting in pBWBs. For construction of pUWBs plasmid, pBWBs was digested using BamH I
and Kpn I and ligated into the expression vector pUFR034 digested with the same restriction
enzymes. The resulting plasmid pUWBs was used to transform X. c. pv. vesicatoria strain
XV1922 by triparental matings. Furthermore, the selected transconjugants were analyzed for
induction of HR in pepper leaves.
Results
Xanthomonas citri subsp. citri hrpW Phylogeny
Comparison of phylogenies from sequences of conserved chromosomal genes is an
effective method to evaluate the evolution among different pathovars in the same genus. In this
study, we analyzed the full sequence of hrpW gene from Xcc strain 306 with strains collected
world-wide and Florida State, and plant pathogenic bacteria that carry hrpW homologous
sequences. The neighbor joining (NJ) tree obtained via analysis of the hrpW sequence (Figure 4-
1) shows that the genomospecies of X. citri, in general, cluster together very well, with the

86
exception of two X. citri strains XS2003-0004 and XS1997-00018 (represented in the tree as
XC04 and XC018, respectively). However, sequence alignment analysis revealed that these two
strains, XS2003-0004 and XS1997-00018, show a certain level of nucleotide similarity (Figure
4-2). The close similarity between hrpW from Xcc strain 306 and X. c. pv. campestris strain
33913 and E. amylovora has already been demonstrated by da Silva et al. (2002). Meanwhile,
the phylogenies of R. solanacearum strain UW551 (UW551), Acidovorax avenae subsp. citrulli
strain AAC00-1 (AAC00), Pseudomonas viridiflava (PV), and Erwinia tasmaniensis strain
Et1/99 indicate that hrpW sequence is also conserved in these strains (Figure 4-1).
The maximum parsimony (MP) analysis of the hrpW sequence set yielded one
parsimonious tree, which was virtually identical to NJ tree, except for the positions of taxa
within two strains, R. solanacearum strain UW551/ Acidovorax avenae subsp. citrulli strain
AAC00-1 (AAC00). The maximum likelihood (ML) tree was identical to the NJ tree, with the
exception that the positions of Acidovorax avenae subsp. citrulli strain AAC00-1 (AAC00) and
Pseudomonas viridiflava (PV) to E. amylovora and Erwinia tasmaniensis strain Et1/99. The
hrpW phylogeny data in the NJ tree were supported by the bootstrap values of 100 in both the
ML analysis and the MP analysis.
Total Deletion of hrpW Gene Is Irrelevant for Xcc Pathogenicity and HR in Non-Host Plants
To determine the relevance of hrpW gene for the pathogenicity for Xcc strain 306 in a
compatible and incompatible reaction, deletion in the 0.9 kb coding sequence of hrpW was
generated using Aat II restriction enzyme. One clone, 306::∆hrpW, representing a hrpW full
length deletion was selected for further characterization. Using sequence alignment we showed
the precise location of the mutations in the HrpW (Figure 4-3). Grapefruit leaves were infiltrated
with bacterial suspensions of 5 x 108 CFU/ml and the generation of symptoms was assessed 7

87
days after inoculation. 306::∆hrpW mutant induced typical canker symptoms on grapefruit at 7
days postinoculation, additionally, the symptoms incited by the mutant are macroscopically
identical with the Xcc wild type (Figure 4-4). Xcc strain 306 induced hypersensitive response
(HR) in tomato plants. In order determine if the mutation in hrpW affect Xcc HR activity, we
inoculated tomato (Bonny Best) and pepper (ECW and ECW-20R) leaves with bacterial
suspensions of 5 x 108 CFU/ml and observations of macroscopic tissue collapse typical of HR
were made 24 h postinoculation. At this concentration, both strains Xcc wild type and
306::∆hrpW induced an HR. Moreover, no difference was noted in the bacterial population
among the strains tested in this experiment (Figure 4-5).
Harpin Domain in hrpW Gene is Required for Pathogenicity
In order to explore the effect of a deletion in the harpin domain of hrpW gene may have on
pathogenicity and HR elicitor activity of Xcc, we used deletion mutagenesis to generate harpin
mutant, W∆harpin, which 87 bp were deleted (Figure 4-3). The HR activity of the W∆harpin
mutant was the same as that of wild type hrpW. Surprisingly, the W::∆harpin mutant showed a
severe reduction on the ability to induce symptoms compared with the Xcc-306 (Figure 4-4). The
W::∆harpin mutant phenotype was restored when we replaced the deleted harpin portion, using
pOHrpW-EP, by the harpin wild type region in cis. Considering the fact that 306::∆hrpW mutant
did not affect the pathogenicity and W::∆harpin blocked the progress of symptom development
in grapefruit, we may propose that this phenotype is given by the expression of a defective HrpW
protein, which may destabilize some protein interaction necessary for the disease progress.
Although the W::∆harpin mutant is affecting the phenotype in a compatible reaction, no change
in the W::∆harpin mutation population was observed compared with the Xcc wild type or the
306::∆hrpW mutant (Figure 4-5).

88
Complementation of HrpW::∆harpin Mutant In-Cis
The first attempts in recovery the W::∆harpin mutant in trans using pUHrpW-EP, which
carry the hrpW gene drive by it native promoter, were unsuccessful. In order to demonstrate a
non-polar mutation effect in W∆harpin mutant, we complemented the W∆harpin mutant using a
0.9 kb PCR fragment from Xcc containing the full length hrpW gene, and cloned into the pOK1
suicide vector, pOHrpW-EP. Inoculations performed in grapefruit leaf showed very similar
symptoms with ∆harpin::harpin complemented and Xcc wild type (Figure 4-4). The results
reported lead us to speculate that this incapability to restore the pathogenicity in trans is due to a
dominant-negative effect of the defective protein, in cis, exerted in the wild type protein
expressed in trans. To determine if our theory is correct, we conjugated the pU∆harpin plasmid
in Xcc strain 306, creating 306::∆harpin(T) transconjugant, that expresses the defective harpin
domain in trans. Therefore, if the expressed defective ∆harpin protein is negatively dominant in
cis, this dominance should be maintained when the defective protein is expressed in trans.
Grapefruit leaves were inoculated by pin-prick method with Xcc wild type, W::∆harpin,
306::∆harpin(T), ∆harpin::harpin and 306::∆hrpW. All strains tested showed typical canker
lesion formation 28 days postinoculation (Figure 4-6). The mutations and complemented mutant
were confirmed by size PCR-fragment (Figure 4-7). The results demonstrated that the loss of
pathogenicity by the W∆harpin mutant it is not caused by negative dominance effect. Therefore,
more experiments need to be performed to explain the role of harpin domain for Xcc virulence.
Translocon Machinery, T3SS, Is Not Affected by ∆harpin Mutant in Xcc
The effect delete of W::∆harpin mutant may have on the ability of Xcc strain 306 to deliver
protein effectors through the T3SS was investigated using pL799 clone, which contain the
avirulence gene, avrGf1, that triggers HR in grapefruit and it is T3SS-dependent (Rybak et al.,

89
2009). The W::∆harpin transconjugant, W∆harpin::799, was infiltrated in grapefruit at 5 x 108
CFU/mL and compared with Xcc wild type strain with avrGf1 clone (pL799). The HR elicitation
was observed in both strains with the same intensity five days after inoculation.
HprW is not Translocated into the Mesophyll of Plant Cell
To determine the targeting of HrpW protein to host cells a fused protein was constructed
with HrpW N-terminal and C-terminal from AvrBs2 of X. c. pv. vesicatoria. The fused protein
was expressed in X. c. pv. vesicatoria strain XV1922, a avrBs2 defective strain, to test HrpW
translocation into pepper leaves. As a positive control we used the XV1922-1 clone, which
contains the avrGf11-106::avrBs262-574 and gives HR in pepper cv ECW-20R. The inoculation
assay shows no HR induction after 24 hours, which means that HrpW protein is not translocated
into the plant cells.
Discussion
HrpW was first identified in E. amylovora (Kim and Beer, 1998). Thereafter, hrpW was
reported in many phytopathogenic bacteria (Gaudriault et al., 1998; Charkowski et al., 1998; da
Silva et al., 2002; Shrestha et al., 2004) including xanthomonads. Therefore, we used the hrpW
sequence from Xcc strain 306 (da Silva et al., 2002), to establish the diversity among several
Xcc strains collected world-wide (Table 4-1). The phylogram of the Xcc strains indicates very
high level of sequence conservation among all Xcc strains analyzed in this study and that it is,
somewhat, less conserved among the other phytopathogenic bacteria (Figure 4-6). With the
exception of Xcc strains XS2003-0004 and XS1997-00018, which diverge from the group
showing high similarity to hrpW sequence from Xcc strain 306. Although the dendrogram
demonstrated that these two strains branched differently from the other strains, the sequence
alignment showed a considerable level of similarity between these two specific strains (Figure 4-
2). The slight diversity reported in the XS2003-0004 and XS1997-00018 strains may be

90
explained due to a flexible element that could affect DNA similarity among pathovars in the
same species, e. g. transposable elements, which are often variable within strains of species
(Parkinson et al., 2007).
Taken together that hrpW is highly conserved among Xanthomonas citri strains and it
downstream location outside of hrp cluster reported by da Silva et al. (2002), we hypothesized
that hrpW might be involved in the Xcc strain 306 pathogenicity. Here, we show that the full
sequence of HrpW is not required for the Xcc pathogenicity and virulence, and neither, for HR
induction in non-host tomato plants. Similar results were reported with hrpW mutants from E.
amylovora and P. s. pv. tomato (Kim and Beer, 1998; Charkowski et al., 1998). Moreover,
previous reports demonstrated that HrpW of E. amylovora inhibits HR induced by the HrpN
(Reboutier et al., 2007), and the expression of hrpW in trans also blocked the HR induction by P.
fluorescens (Charkowski et al., 1998). Based on these reports, we can speculate that the lack of
HrpW in Xcc eliminates an unknown antagonistic interaction with other pathogenic factors.
Studies comparing HrpW protein from Xcc and X. c. pv. campestris with HrpW harpins of
Pseudomonas and Erwinia species, showed that the HrpW protein from both Xanthomonas did
not elicit HR in non-host plants. Surprisingly, the deletion of a few amino acids in the N-
terminal, which encodes the putative harpin domain, totally shutdown the Xcc virulence in
grapefruit plants but does not affect the bacterial population in planta and the ability to trigger
HR in non-host tomato plants. Although it is well known that harpins are HR-elicitors, their role
in bacteria pathogenicity is still unclear. Some of the harpins identified are directly linked with
pathogenicity such as, HrpN in E. amylovora and E. chrysanthemi (Wei et al., 1992), HrpN also
is involved in the translocation of T3-effector DspA/E (Bocsanczy et al., 2008) while HrpZ from

91
P. s. pv. phaseolicola is associated with pore-forming activity that might be involved in nutrient
release and/or entry for bacterial effectors (Lee et al., 2001).
Intriguingly, the absence of the entire hrpW gene did not compromise the Xcc
pathogenicity in a compatible reaction in contrast what was observed with the deletion on the
harpin domain. We therefore suggest that the protein-protein interaction complex that HrpW
form with other hrp genes is disrupted due to change in the protein conformation expressed by
the ∆harpin mutant. Alegria et al. (2004) demonstrated that HrpW forms a protein complex with
three other Xcc proteins, HrpD6 and HrpB1, which are cytoplasmic proteins that shown
homology with proteins in X. c. pv. vesicatoria that are essential for pathogenicity and conserved
hypothetical proteins, respectively (Rossier et al., 2000) and HrpB4, although the interaction of
this third proteins is not physically demonstrated. Xcv HrpB4 protein is essential for
pathogenicity and T3-effectors translocation and not secreted or translocated, and shows
association with signal sensors (Rossier et al., 2002; Alegria et al., 2004). These findings support
our suggestion that a defective hrpW protein encoded by harpin mutant destabilizes this complex,
consequently, reducing the HrpB4 interaction or the sense of a specific signal. However, the
translocation of T3-effector, AvrGf1, into the host cell was not terminated in the harpin mutant.
Considering this fact, we may speculate that this phenotype was caused by the interruption or
reduction of the translocation of a specific crucial factor for pathogenicity. Our hypothesis is also
supported by recently studies showing that phytopathogenic bacteria carrying a defective harpin
protein impaired the translocation of several type III secretion system effectors (Kvitko et al.,
2007; Bocsanczy et al., 2008).
In addition, we have characterized the stability of the T3SS in the ∆harpin mutant and
translocation of HrpW from Xcv strain XV1922 to pepper plants. We show that the deletion on

92
the harpin domain does not affect the T3-effector translocation inside of plant cells. Furthermore,
the use of AvrBs2 C-terminal, a Xcv T3-effector protein, as a reporter demonstrated that the
HrpW is not translocated into the mesophyll of host cells. However, using CyaA reporter system
demonstrated that HrpN, a harpin protein from E. amylovora, is slightly translocated into tobacco
plant cells (Bocsanczy et al., 2008). The lack of translocation reported here may be due the fact
that harpin domain in Xcc is smaller and has highly homology with the Pel domains of the
Pseudomonas and Erwinia than with harpin domains (Kim et al., 2004).

93
Table 4-1. Bacterial strains and plasmids used in the study Strain or plasmid Relevant characteristics Source Strains X. citri subsp. citri 290 Isolated from Saudi Arabia R. E. Stalla 112 Isolated from China R. E. Stall 131 Isolated from Maldive Islands R. E. Stall 106 Isolated from Australia R. E. Stall 46 Isolated from India R. E. Stall 257-2 Isolated from Thailans R. E. Stall 62 Isolated from Japan R. E. Stall 126 Isolated from Korea R. E. Stall 101 Isolated from Guam R. E. Stall XI2000-00194 Isolated from Florida DPIb
XS1999-0038 Isolated from Florida DPI X1999-12815 Isolated from Florida DPI XN03-2912 Isolated from Florida DPI XI3001-00098 Isolated from Florida DPI X2000-12878 Isolated from Florida DPI XI1999-00112 Isolated from Florida DPI XN03-11#3 Isolated from Florida DPI XS2003-0004 Isolated from Florida DPI XS1997-00018 Isolated from Florida DPI 306 Wild type, Asiatic strain, isolated in Brazil,
RifR DPI
306::∆ hrpW hrpW -, 855 bp deleted, RifR This study W::∆harpin Harpin -, 87 bp deleted, RifR This study 306::∆hrpG hrpG -, single recombinant of pCRhrpG, KnR,
AmpR, RifR This study
∆harpin::Harpin Harpin – complemented with pOHrpW-EP, RifR
This study
306::∆harpin(T) 306 carrying pU∆Harpin, KnR, RifR This study 306::799 306 carrying pL799, TcR, RifR This study X. c. pv. campestris XV1922 Pepper race 6, RifR Jones, J. B.c 1922::WBs strain XV1922 carrying pUWBs, KnR, RifR This study ∆harpin::799 Harpin – carrying pL799, TcR, RifR This study Escherichia coli DH5α FrecAϕ80dlacZ∆M15 Invitrogen
λPIR Host for pOK1; SpR oriR6K replicon UBd Plasmids
pCR2.1-TOPO Phagemid, CbR, KnR Invitrogen pBluescript-KS+/- Phagemid, pUC derivative; AmpR Stratagene pRK2013 KnR, tra+, mob+, SmR Daniels et al. (1984) pOK1 Suicide vector, pKNG101 derivative,
SmR/SucS Huguet et al. (1998)

94
Table 4-1. Continued pLAFR3 TcR, rlx+, RK2 replicon BJSe
pUFR034 Tn903, IncW, Mob+, KnR, De Feyter et al. (1990) pCRhrpW hrpW partial fragment in pCR2.1-TOPO This study pCR∆harpin Harpin -, 87 bp deleted with Aat II, KnR,
AmpR
pCRHrpW-EP pCR2.1 with1.8-kb hrpW plus native promoter, KnR, AmpR
This study
pOHrpW-EP pOK1 clone containing hpW plus native promoter, SmR
This study
pO∆Harpin/PEL pOK1 with HrpW-, SmR This study pO∆Harpin pOK1 with Harpin-, SmR This study pL22 Xcc strain Aw pLAFR3 cosmid that contains
hrpX, hrpG and hsp90Xo homologues, TcR Rybak et al. (2009)
pL799 pLAFR3 with DNA fragment from Xcc-Aw that contains avrGf1, TcR
Rybak et al. (2009)
pBS1 BglII::avrBs262-574::HA of pBluescript-KS+/-, AmpR Mudgett, M. B.f
pBWBs hrpW1-109 cloned in pBS1 This study
pUWBs hrpW1-109 fused to avrBs262-574 cloned pUFR034 This study
pUHrpW-EP Xcc pUFR034 clone containing hpW plus native promoter, KnR
This study
pU∆Harpin Xcc pUFR034 clone containing harpin domain deleted, KnR
This study
a R.E. Stall, Robert E. Stall, University of Florida, Gainesville, FL. b DPI, Division of Plant Industry of the Florida Department of Agriculture and Consumer Services, Gainesville, FL, USA. c Jones, J. B., University of Florida, Gainesville, FL, USA. d UB, Ulla Bonas, Martin-Luther-Universität, Halle, Germany. e BJS, B. J. Staskawicz, University of California, Berkley, CA f Mudgett, M. B., Stanford University, Stanford, CA.

95
Table 4-2. Oligonucleotides sequence used in this study Gene Primer name Primer sequence length hrpW POWF1 5’-ACGCGTCGACGTCAGCCGAAAGGAATATACAG-3’ 32
POWR1 5’-CGCGTCGACCAACCGCTGTAGCAATTCCGAC-3’ 31 UpHprWF1 5’-CCTATATCAGTGCTAACCCACT-3’ 22 DoHprWR1 5’-CCTATATCAGTGCTAACCCACT-3’ 22 NTW-F1 5’-GGAAGATCTGATGTCTTTGGCCAACACACTGC-3’ 32 NTW-R1 5’-GGAAGATCTGCCGACGTAGAGATTGTTGTGTCC-3’ 33
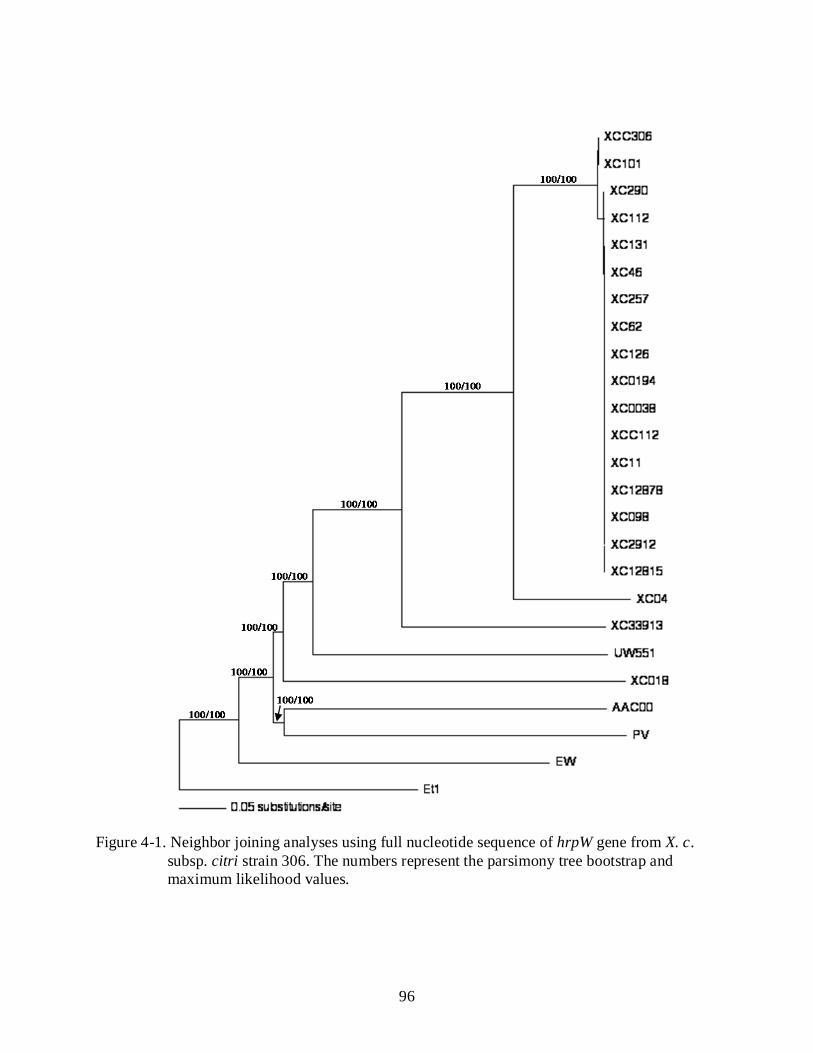
96
Figure 4-1. Neighbor joining analyses using full nucleotide sequence of hrpW gene from X. c. subsp. citri strain 306. The numbers represent the parsimony tree bootstrap and maximum likelihood values.

97
XC04 ------------------------------------------------------------ XC018 ------------------------------------------------------------ XCC306 ------------------------------------------------------------ XCC112 ------------------------------------------------------------ XC290 ------------------------------------------------------------ XC112 ------------------------------------------------------------ XC131 ------------------------------------------------------------ XC46 ------------------------------------------------------------ XC257-2 ------------------------------------------------------------ XC62 ------------------------------------------------------------ XC126 ------------------------------------------------------------ XC101 ------------------------------------------------------------ XC0194 ------------------------------------------------------------ XC0038 ------------------------------------------------------------ XC12815 ------------------------------------------------------------ XC2912 ------------------------------------------------------------ XC098 ------------------------------------------------------------ XC12878 ------------------------------------------------------------ XCC11 ------------------------------------------------------------ XC33913 ------------------------------------------------------------ UW551 ------------------------------------------------------------ AAC00 ------------------------------------------------------------ EW -------------ATGTCAATTCTTACGCTTAACAACAATACCTC----GTCCTCGCCGG 43 ET1 -------------ATGTCAGTTCTTACGCTTAACATCACTATCCC----ATCCGTGCAGG 43 PV GGTCAGGGCGGACAAAGCGATCTGGACTCGCTGCTGCAGTCGCTCCAGGGCAACGGTCAG 300 XC04 ------------------------------------------------------------ XC018 ------------------------------------------------------------ XCC306 ------------------------------------------------------------ XCC112 ------------------------------------------------------------ XC290 ------------------------------------------------------------ XC112 ------------------------------------------------------------ XC131 ------------------------------------------------------------ XC46 ------------------------------------------------------------ XC257-2 ------------------------------------------------------------ XC62 ------------------------------------------------------------ XC126 ------------------------------------------------------------ XC101 ------------------------------------------------------------ XC0194 ------------------------------------------------------------ XC0038 ------------------------------------------------------------ XC12815 ------------------------------------------------------------ XC2912 ------------------------------------------------------------ XC098 ------------------------------------------------------------ XC12878 ------------------------------------------------------------ XCC11 ------------------------------------------------------------ XC33913 ------------------------------------------------------------ UW551 ------------------------------------------------------------ AAC00 ------------------------------------------------------------ EW GTCTGTTCCAGTCCGGG-GGGGACAACGGGCTTGGT---GGTCAT--AATGCAAATTCTG 97 ET1 GGCTGATGCAGCCCGGC-GCGGATAGCGGGTTTTCCACGGGAAAC--AACGCCAACGCCG 100 PV GGCGGTGACAAGCAGACCAAGGATTCCGGCGCTGCCGGCGGCGGCGAAAAGGAACTGCTG 360
Figure 4-2. Sequence alignment of Xcc strains collected world-wide and sequences from X. c. pv. campestris strain 33913, R. solanacearum strain UW551, Acidovorax avenuae subsp. citrulli strain AAC00-1, Pseudomonas viridiflava, Erwina amylovora, and Erwinia tasmaniensis strain Et1/99.

98
XC04 ------------------------------------------------------------ XC018 ------------------------------------------------------------ XCC306 ------------------------------------------------------------ XCC112 ------------------------------------------------------------ XC290 ------------------------------------------------------------ XC112 ------------------------------------------------------------ XC131 ------------------------------------------------------------ XC46 ------------------------------------------------------------ XC257-2 ------------------------------------------------------------ XC62 ------------------------------------------------------------ XC126 ------------------------------------------------------------ XC101 ------------------------------------------------------------ XC0194 ------------------------------------------------------------ XC0038 ------------------------------------------------------------ XC12815 ------------------------------------------------------------ XC2912 ------------------------------------------------------------ XC098 ------------------------------------------------------------ XC12878 ------------------------------------------------------------ XCC11 ------------------------------------------------------------ XC33913 ------------------------------------------------------------ UW551 ------------------------------------------------------------ AAC00 ------------------------------------------------------------ EW CGTTGGGGCAACAACCCATCGATCGGCAAACCATTGAGCAAAT-GGCTCAATTATTGGCG 156 ET1 GGTTGGGGCAACAGCCCTTCGACAAACAAACCATTGAGCAAAT-GGCGCAGTTATTGGGA 159 PV ACTCAGGTCTTGATGGCCCTGTTCGAGAAGATCCTTGGCGGCTCGGATTCATCATC--CG 418 XC04 ------------------------------------------------------------ XC018 ------------------------------------------------------------ XCC306 ------------------------------------------------------------ XCC112 ------------------------------------------------------------ XC290 ------------------------------------------------------------ XC112 ------------------------------------------------------------ XC131 ------------------------------------------------------------ XC46 ------------------------------------------------------------ XC257-2 ------------------------------------------------------------ XC62 ------------------------------------------------------------ XC126 ------------------------------------------------------------ XC101 ------------------------------------------------------------ XC0194 ------------------------------------------------------------ XC0038 ------------------------------------------------------------ XC12815 ------------------------------------------------------------ XC2912 ------------------------------------------------------------ XC098 ------------------------------------------------------------ XC12878 ------------------------------------------------------------ XCC11 ------------------------------------------------------------ XC33913 ------------------------------------------------------------ UW551 ------------------------------------------------------------ AAC00 ------------------------------------------------------------ EW GAACTGTTAAAGTCACTGCTATCGCCACAATCAGGTAATGCGGCAACCGGAGCCGGTGGC 216 ET1 GAGCTGTTAAAGCCGCTGCTATCGCCACAGGCAGGCAATGCGGCAACGGGCGCTAACGGC 219 PV GCGCGGGCAACGGCTCTGGCGGTGGTTCGGGCGGCGCCAGTGGCA-TTGGCGGTGGCGGA 477
Figure 4-2.Continued

99
XC04 ------------------------------------------------------------ XC018 ------------------------------------------------------------ XCC306 ------------------------------------------------------------ XCC112 ------------------------------------------------------------ XC290 ------------------------------------------------------------ XC112 ------------------------------------------------------------ XC131 ------------------------------------------------------------ XC46 ------------------------------------------------------------ XC257-2 ------------------------------------------------------------ XC62 ------------------------------------------------------------ XC126 ------------------------------------------------------------ XC101 ------------------------------------------------------------ XC0194 ------------------------------------------------------------ XC0038 ------------------------------------------------------------ XC12815 ------------------------------------------------------------ XC2912 ------------------------------------------------------------ XC098 ------------------------------------------------------------ XC12878 ------------------------------------------------------------ XCC11 ------------------------------------------------------------ XC33913 ------------------------------------------------------------ UW551 ---ATGCTACGCGCCTCATCCGAGGGGCGCGGGCTCCCACAACCTTTCTTTGTCGAGACC 57 AAC00 ------------------------------------------------------------ EW AATGACCAGACTACAGGAGTTGGTAACGCTGGCGGCCTGAACGGACGAAAAGGCACAGCA 276 ET1 GAGGGCCAGCTGGCCGGCGTGGGGAACGCCGGCGGTCCGTCAGAGCAGAATGGTGCCCTG 279 PV ACCGGCGCCAGCAAAGATGCCGGTGCTCTCGGTCAAGGGCAGGGACTCGGCGGCACTCAG 537 XC04 ------------------------------------------------------------ XC018 ------------------------------------------------------------ XCC306 ------------------------------------------------------------ XCC112 ------------------------------------------------------------ XC290 ------------------------------------------------------------ XC112 ------------------------------------------------------------ XC131 ------------------------------------------------------------ XC46 ------------------------------------------------------------ XC257-2 ------------------------------------------------------------ XC62 ------------------------------------------------------------ XC126 ------------------------------------------------------------ XC101 ------------------------------------------------------------ XC0194 ------------------------------------------------------------ XC0038 ------------------------------------------------------------ XC12815 ------------------------------------------------------------ XC2912 ------------------------------------------------------------ XC098 ------------------------------------------------------------ XC12878 ------------------------------------------------------------ XCC11 ------------------------------------------------------------ XC33913 ------------------------------------------------------------ UW551 GCCATGCCCATCCAGGTAGATCGCCCGAGTGACCATTTCCATGTGCCCTCCACGTGGAAT 117 AAC00 ------------------------------------------------------------ EW GGAACCACT------CCGCAGTCTGACAGTCAG---AACATGCTGAGTGAGA---TGGGC 324 ET1 GGGGCAACG------CCACAGAATAACGGCCAG---AACGCGTTAAGTGAGA---TGGGC 327 PV GGCGCTACGGGCGGATCGAGCACTGAGGATCTGGTCAACACGCTGATGCAGAAACTGGGT 597
Figure 4-2.Continued

100
XC04 ------------------------------------------------------------ XC018 ------------------------------------------------------------ XCC306 ------------------------------------------------------------ XCC112 ------------------------------------------------------------ XC290 ------------------------------------------------------------ XC112 ------------------------------------------------------------ XC131 ------------------------------------------------------------ XC46 ------------------------------------------------------------ XC257-2 ------------------------------------------------------------ XC62 ------------------------------------------------------------ XC126 ------------------------------------------------------------ XC101 ------------------------------------------------------------ XC0194 ------------------------------------------------------------ XC0038 ------------------------------------------------------------ XC12815 ------------------------------------------------------------ XC2912 ------------------------------------------------------------ XC098 ------------------------------------------------------------ XC12878 ------------------------------------------------------------ XCC11 ------------------------------------------------------------ XC33913 ------------------------------------------------------------ UW551 CAAGACACTGGTTCCAAGATCGATACCAGCCAACTTCAGCATGCCGTACAACTGCTCCAG 177 AAC00 -----ATGCGACGCCGTGCATCGGGCATGGCGTACCGGCGCTTCGGCGCCGCACTTTTCG 55 EW AACAACGGGCTGGATCAGGCCATCACGCCCGATGGCCAGGGCGGCGGGCAGATCGGCGAT 384 ET1 AACAACGGGCTGGATCAGGCCCTGACCCCCGATGGCCAGGGTGGAGGGCAAATCAGCGAT 387 PV GGTGGATCGCTGGATAACTCCATTCAGCCGACTGCCGATGGCGGCGGTGAGGTGTCCCAG 657 XC04 ------------------------GTGGCCTCCTACACACGGGCGGGTGTGGGGATATAC 36 XC018 ------------------------------------------------------------ XCC306 ------------------------------------------------------------ XCC112 ------------------------------------------------------------ XC290 ------------------------------------------------------------ XC112 ------------------------------------------------------------ XC131 ------------------------------------------------------------ XC46 ------------------------------------------------------------ XC257-2 ------------------------------------------------------------ XC62 ------------------------------------------------------------ XC126 ------------------------------------------------------------ XC101 ------------------------------------------------------------ XC0194 ------------------------------------------------------------ XC0038 ------------------------------------------------------------ XC12815 ------------------------------------------------------------ XC2912 ------------------------------------------------------------ XC098 ------------------------------------------------------------ XC12878 ------------------------------------------------------------ XCC11 ------------------------------------------------------------ XC33913 ------------------------------------------------ATGCAACGCATG 12 UW551 CAGGTCTTGCAGCAGGTCCAAGCGAACAAGCTGTTCGGGAACGTGCTGAACCAGCCGGAA 237 AAC00 CGAGATCCATGAACCCCATCGCCTTTCTCTCGTCTTCCCTGACCGCACGGCATGCCGCGG 115 EW AATCCTTTACTGAAAGCCATGCTGAAGCTTATTGCACGCATGATGGACGGCCAAAGCGAT 444 ET1 AATCCGTTGCTGAAAGCCCTGCTGAAACTGATCGCCCGAATGATGGACGGGCAGAGCGAC 447 PV AACGGCAAGCTCAAAGAGCTGCTGGAAATGATCGCTCAGTTCATGGACAGCCATCCGGAG 717
Figure 4-2.Continued
101
XC04 CATAGATCCTCCCACTATTGCCCAGACCTCTCTCCCAAAAGAGCACCG-TGCGCGCAATA 95 XC018 ------------------------------------------------------------ XCC306 ----------------------------ATGCTGATGTCGGAATTGCT-ACAGCGGTTGA 31 XCC112 ----------------------------ATGCTGATGTCGGAATTGCT-ACAGCGGTTGA 31 XC290 ----------------------------ATGCTGATGTCGGAATTGCT-ACAGCGGTTGA 31 XC112 ----------------------------ATGCTGATGTCGGAATTGCT-ACAGCGGTTGA 31 XC131 ----------------------------ATGCTGATGTCGGAATTGCT-ACAGCGGTTGA 31 XC46 ----------------------------ATGCTGATGTCGGAATTGCT-ACAGCGGTTGA 31 XC257-2 ----------------------------ATGCTGATGTCGGAATTGCT-ACAGCGGTTGA 31 XC62 ----------------------------ATGCTGATGTCGGAATTGCT-ACAGCGGTTGA 31 XC126 ----------------------------ATGCTGATGTCGGAATTGCT-ACAGCGGTTGA 31 XC101 ----------------------------ATGCTGATGTCGGAATTGCT-ACAGCGGTTGA 31 XC0194 ----------------------------ATGCTGATGTCGGAATTGCT-ACAGCGGTTGA 31 XC0038 ----------------------------ATGCTGATGTCGGAATTGCT-ACAGCGGTTGA 31 XC12815 ----------------------------ATGCTGATGTCGGAATTGCT-ACAGCGGTTGA 31 XC2912 ----------------------------ATGCTGATGTCGGAATTGCT-ACAGCGGTTGA 31 XC098 ----------------------------ATGCTGATGTCGGAATTGCT-ACAGCGGTTGA 31 XC12878 ----------------------------ATGCTGATGTCGGAATTGCT-ACAGCGGTTGA 31 XCC11 ----------------------------ATGCTGATGTCGGAATTGCT-ACAGCGGTTGA 31 XC33913 CTCAGGGACATGCTGCAGCCACAGGATCAGCCATTCAGCGACGGTCAG-CCACCGACCAA 71 UW551 TTCGGCAACTCCGGTCAGGGGCATGGCGGCAGCCACGGTGGCGGCCATCACGGCGGCTCG 297 AAC00 CATCGGACCAGGCCGCCTCCGCGGCCCGGCACAACGGCAGCCAGGCACTGCAGCAATCCC 175 EW CAGTTTGGCCAACCT------------GGTACGGGCAACAACAGTGCCTCTTCCGGTACT 492 ET1 CCGCTCGGCCAGCAG------------GGCGCTGGCAGTCACAATGCGTCTTCCGGCACG 495 PV ACTTTCAATCAGCCGTCTGATGCTGCGGGCAAAGGTGGTGGCGGTGGCGGTGGTGGAACA 777 XC04 TAAGGGATTTGGGAAAGCG-CCTTTGCT-------GGCCTTTCCCCAATGGGGACCTGCC 147 XC018 ------------------------------------------------------------ XCC306 TACAGACCCAATTCCAACC-TCAAAACA-------CGTCGCAATACGATTGGAACCCTTC 83 XCC112 TACAGACCCAATTCCAACC-TCAAAACA-------CGTCGCAATACGATTGGAACCCTTC 83 XC290 TACAGACCCAATTCCAACC-TCAAAACA-------CGTCGCAATACGATTGGAACCCTTC 83 XC112 TACAGACCCAATTCCAACC-TCAAAACA-------CGTCGCAATACGATTGGAACCCTTC 83 XC131 TACAGACCCAATTCCAACC-TCAAAACA-------CGTCGCAATACGATTGGAACCCTTC 83 XC46 TACAGACCCAATTCCAACC-TCAAAACA-------CGTCGCAATACGATTGGAACCCTTC 83 XC257-2 TACAGACCCAATTCCAACC-TCAAAACA-------CGTCGCAATACGATTGGAACCCTTC 83 XC62 TACAGACCCAATTCCAACC-TCAAAACA-------CGTCGCAATACGATTGGAACCCTTC 83 XC126 TACAGACCCAATTCCAACC-TCAAAACA-------CGTCGCAATACGATTGGAACCCTTC 83 XC101 TACAGACCCAATTCCAACC-TCAAAACA-------CGTCGCAATACGATTGGAACCCTTC 83 XC0194 TACAGACCCAATTCCAACC-TCAAAACA-------CGTCGCAATACGATTGGAACCCTTC 83 XC0038 TACAGACCCAATTCCAACC-TCAAAACA-------CGTCGCAATACGATTGGAACCCTTC 83 XC12815 TACAGACCCAATTCCAACC-TCAAAACA-------CGTCGCAATACGATTGGAACCCTTC 83 XC2912 TACAGACCCAATTCCAACC-TCAAAACA-------CGTCGCAATACGATTGGAACCCTTC 83 XC098 TACAGACCCAATTCCAACC-TCAAAACA-------CGTCGCAATACGATTGGAACCCTTC 83 XC12878 TACAGACCCAATTCCAACC-TCAAAACA-------CGTCGCAATACGATTGGAACCCTTC 83 XCC11 TACAGACCCAATTCCAACC-TCAAAACA-------CGTCGCAATACGATTGGAACCCTTC 83 XC33913 CCCGAATCCGACACCACCG-ATGCCGCC-------CTCGCGCCCGCCCTGTTCCACCAAC 123 UW551 ACCGGGTTCGGCGAAAACGGTCGATTCG-------GTTTGCCGTACGCCAAGCCGCCTGC 350 AAC00 TCGCCACCCTGCTCGAAGG-CCTCCTGA-------AGCGCATGCGCGGCGGAGCCGCCCC 227 EW TCTTCATCTGGCGGTTCCCCTTTTAACG-------ATCTATCA-----GGGGGGAAGGCC 540 ET1 TCCTCAACGGGGAGCTCCCCTTCCAACG-------ATCTATCA-----GGCAACGGCTTC 543 PV CCTTCCGTAGGCGGTGGCGGTGGCGGTGGCGGTACACCTTCCGTGGGCGGTGGCGGAGAT 837
Figure 4-2.Continued
102
XC04 CACATTGGCAAAGGGGGA--GGGGCAAACCGAA-GCAAGTGCCGGGTCGGTAA------- 197 XC018 ------------------------------------------------------------ XCC306 GCCTTCCCATGGGACGTC--CAATGGCCAAGGG-GAGGGCAACGAGCATGGCG------- 133 XCC112 GCCTTCCCATGGGACGTC--CAATGGCCAAGGG-GAGGGCAACGAGCATGGCG------- 133 XC290 GCCTTCCCATGGGACGTC--CAATGGCCAAGGG-GAGGGCAACGAGCATGGCG------- 133 XC112 GCCTTCCCATGGGACGTC--CAATGGCCAAGGG-GAGGGCAACGAGCATGGCG------- 133 XC131 GCCTTCCCATGGGACGTC--CAATGGCCAAGGG-GAGGGCAACGAGCATGGCG------- 133 XC46 GCCTTCCCATGGGACGTC--CAATGGCCAAGGG-GAGGGCAACGAGCATGGCG------- 133 XC257-2 GCCTTCCCATGGGACGTC--CAATGGCCAAGGG-GAGGGCAACGAGCATGGCG------- 133 XC62 GCCTTCCCATGGGACGTC--CAATGGCCAAGGG-GAGGGCAACGAGCATGGCG------- 133 XC126 GCCTTCCCATGGGACGTC--CAATGGCCAAGGG-GAGGGCAACGAGCATGGCG------- 133 XC101 GCCTTCCCATGGGACGTC--CAATGGCCAAGGG-GAGGGCAACGAGCATGGCG------- 133 XC0194 GCCTTCCCATGGGACGTC--CAATGGCCAAGGG-GAGGGCAACGAGCATGGCG------- 133 XC0038 GCCTTCCCATGGGACGTC--CAATGGCCAAGGG-GAGGGCAACGAGCATGGCG------- 133 XC12815 GCCTTCCCATGGGACGTC--CAATGGCCAAGGG-GAGGGCAACGAGCATGGCG------- 133 XC2912 GCCTTCCCATGGGACGTC--CAATGGCCAAGGG-GAGGGCAACGAGCATGGCG------- 133 XC098 GCCTTCCCATGGGACGTC--CAATGGCCAAGGG-GAGGGCAACGAGCATGGCG------- 133 XC12878 GCCTTCCCATGGGACGTC--CAATGGCCAAGGG-GAGGGCAACGAGCATGGCG------- 133 XCC11 GCCTTCCCATGGGACGTC--CAATGGCCAAGGG-GAGGGCAACGAGCATGGCG------- 133 XC33913 CCCGGCGGACAACACGGTAGCGGTACTGGCGAT-GGCAGCGGCGGCAATGGCG------- 175 UW551 GCAGCCGGACATTGAGCTGCCGGCCAACAAGCCCAACCCCGGCAAGCACAACAC---TTC 407 AAC00 GTCACCCCGCTCTGGCGG---AAGCGCCGCGGG-GCATGCCGCCGGGACGGCG------- 276 EW CCTTCCGGCAACTCCCCTTCCGGCAACTACTCTCCCGTCAGTACCTTCTCACCC-CCATC 599 ET1 CCTTCCGG---CCCTTCTTCCGGCGGCACAGCGCCAACCCATTCTGACTCACCG-CCGTC 599 PV GGTGGCGGAACACCTTCCGTAGGCGGCGGAGGCGGAGGTGGTGGCGGAACACCTTCCGTA 897 XC04 -GGAAAAGGAACGCGGGCGGGTGATAAAATTTTCTTTGGCCAACCACGTCTCGAG-GACG 255 XC018 ------------------------------------------------------------ XCC306 -GTCGCAGGAACGGACGCGGCGGTGACAACTTCCTCG---CCACCACGCCC-----GAGA 184 XCC112 -GTCGCAGGAACGGACGCGGCGGTGACAACTTCCTCG---CCACCACGCCC-----GAGA 184 XC290 -GTCGCAGGAACGGACGCGGCGGTGACAACTTCCTCG---CCACCACGCCC-----GAGA 184 XC112 -GTCGCAGGAACGGACGCGGCGGTGACAACTTCCTCG---CCACCACGCCC-----GAGA 184 XC131 -GTCGCAGGAACGGACGCGGCGGTGACAACTTCCTCG---CCACCACGCCC-----GAGA 184 XC46 -GTCGCAGGAACGGACGCGGCGGTGACAACTTCCTCG---CCACCACGCCC-----GAGA 184 XC257-2 -GTCGCAGGAACGGACGCGGCGGTGACAACTTCCTCG---CCACCACGCCC-----GAGA 184 XC62 -GTCGCAGGAACGGACGCGGCGGTGACAACTTCCTCG---CCACCACGCCC-----GAGA 184 XC126 -GTCGCAGGAACGGACGCGGCGGTGACAACTTCCTCG---CCACCACGCCC-----GAGA 184 XC101 -GTCGCAGGAACGGACGCGGCGGTGACAACTTCCTCG---CCACCACGCCC-----GAGA 184 XC0194 -GTCGCAGGAACGGACGCGGCGGTGACAACTTCCTCG---CCACCACGCCC-----GAGA 184 XC0038 -GTCGCAGGAACGGACGCGGCGGTGACAACTTCCTCG---CCACCACGCCC-----GAGA 184 XC12815 -GTCGCAGGAACGGACGCGGCGGTGACAACTTCCTCG---CCACCACGCCC-----GAGA 184 XC2912 -GTCGCAGGAACGGACGCGGCGGTGACAACTTCCTCG---CCACCACGCCC-----GAGA 184 XC098 -GTCGCAGGAACGGACGCGGCGGTGACAACTTCCTCG---CCACCACGCCC-----GAGA 184 XC12878 -GTCGCAGGAACGGACGCGGCGGTGACAACTTCCTCG---CCACCACGCCC-----GAGA 184 XCC11 -GTCGCAGGAACGGACGCGGCGGTGACAACTTCCTCG---CCACCACGCCC-----GAGA 184 XC33913 -GCAACCACAACAGAGGCAACGGCGGCACCTGCGGAACGGGAGCCGATCACCGC--GCAT 232 UW551 CGCGCCGACCACGGAGACGACCCCGCCTGCTGTTTCGCCCAAGCCCGATAC---G-TCTC 463 AAC00 -CCTCCGGCCTCGCACACCACGCAACGTGCCGTGGCGC--ACACCGATGCCGGCC-GCGC 332 EW CACGCCAACGTCCCCTACCTCACCGCTTGATTTCCCTTCTTCTCCCACCAAAGCA-GCCG 658 ET1 CACGCCAACCTCCCCCACCTCACCGCTTGATTTCCCGTCTTCCCCCACCGGCGGA-GCCG 658 PV GGCGGCGGTGGCGGGGGTGGTGGCGGTACACCTTCCATAGGCGGTGGCGGTGGTACGCCT 957
Figure 4-2.Continued

103
XC04 TTCTTGATCATTTCGAAAAAA---TTCGGTTCAATTCGAACCGTT-ACCGGGGAAGCGGG 311 XC018 -----------------------------------------------------CAACGGC 7 XCC306 CGTCCGACCATTCCGACAAA-----TCCGCTCACTC--GACCGTC-ACCGG--CAGCGGC 234 XCC112 CGTCCGACCATTCCGACAAA-----TCCGCTCACTC--GACCGTC-ACCGG--CAGCGGC 234 XC290 CGTCCGACCATTCCGACAAA-----TCCGCTCACTC--GACCGTC-ACCGG--CAGCGGC 234 XC112 CGTCCGACCATTCCGACAAA-----TCCGCTCACTC--GACCGTC-ACCGG--CAGCGGC 234 XC131 CGTCCGACCATTCCGACAAA-----TCCGCTCACTC--GACCGTC-ACCGG--CAGCGGC 234 XC46 CGTCCGACCATTCCGACAAA-----TCCGCTCACTC--GACCGTC-ACCGG--CAGCGGC 234 XC257-2 CGTCCGACCATTCCGACAAA-----TCCGCTCACTC--GACCGTC-ACCGG--CAGCGGC 234 XC62 CGTCCGACCATTCCGACAAA-----TCCGCTCACTC--GACCGTC-ACCGG--CAGCGGC 234 XC126 CGTCCGACCATTCCGACAAA-----TCCGCTCACTC--GACCGTC-ACCGG--CAGCGGC 234 XC101 CGTCCGACCATTCCGACAAA-----TCCGCTCACTC--GACCGTC-ACCGG--CAGCGGC 234 XC0194 CGTCCGACCATTCCGACAAA-----TCCGCTCACTC--GACCGTC-ACCGG--CAGCGGC 234 XC0038 CGTCCGACCATTCCGACAAA-----TCCGCTCACTC--GACCGTC-ACCGG--CAGCGGC 234 XC12815 CGTCCGACCATTCCGACAAA-----TCCGCTCACTC--GACCGTC-ACCGG--CAGCGGC 234 XC2912 CGTCCGACCATTCCGACAAA-----TCCGCTCACTC--GACCGTC-ACCGG--CAGCGGC 234 XC098 CGTCCGACCATTCCGACAAA-----TCCGCTCACTC--GACCGTC-ACCGG--CAGCGGC 234 XC12878 CGTCCGACCATTCCGACAAA-----TCCGCTCACTC--GACCGTC-ACCGG--CAGCGGC 234 XCC11 CGTCCGACCATTCCGACAAA-----TCCGCTCACTC--GACCGTC-ACCGG--CAGCGGC 234 XC33913 CGCCCACCAACCCGGGTCAGGCGGTGAGGCCCTGCCCCGATGGTCCGCAGGCCCCACTTC 292 UW551 CGGCCTCCAAGCCTGACACGTC--CTCGACCTCCACATCCGCGACGGAAGGCAAGGTGAC 521 AAC00 CGGCACGGCGCACGCAGCGCGCGGCACGGCCACCACGGCCCATGCCGCGAAGCCTCCCGC 392 EW G----GGGCAGCACGCCGGTAACCGATCATCCTGACCCTGTTGGTAGCGCGGGCATCGGG 714 ET1 G----GGGCAGCACGCCGGTAACCGATCGTCCCGATCCTGTTGGCAGTTCAGGCGTAGGG 714 PV GCACCGACCGGCCCGACGGGAACACCTTCACCCACCGGTCCGACCGGCACAGGCACCAGC 1017 XC04 C-------CCCGTGGGGGAGCGGGTTTGCACGGAT---------CAACGTCCAA--CAGT 353 XC018 C-------CCGGGTGGGGAGCGG-TCGGCACCGATT--------CAACGTCCA---CAGT 48 XCC306 C-------CCG--TGGGGAGCGG--TCGCAC-GAT---------CAACGTCAA---CAGT 270 XCC112 C-------CCG--TGGGGAGCGG--TCGCAC-GAT---------CAACGTCAA---CAGT 270 XC290 C-------CCG--TGGGGAGCGG--TCGCAC-GAT---------CAACGTCAA---CAGT 270 XC112 C-------CCG--TGGGGAGCGG--TCGCAC-GAT---------CAACGTCAA---CAGT 270 XC131 C-------CCG--TGGGGAGCGG--TCGCAC-GAT---------CAACGTCAA---CAGT 270 XC46 C-------CCG--TGGGGAGCGG--TCGCAC-GAT---------CAACGTCAA---CAGT 270 XC257-2 C-------CCG--TGGGGAGCGG--TCGCAC-GAT---------CAACGTCAA---CAGT 270 XC62 C-------CCG--TGGGGAGCGG--TCGCAC-GAT---------CAACGTCAA---CAGT 270 XC126 C-------CCG--TGGGGAGCGG--TCGCAC-GAT---------CAACGTCAA---CAGT 270 XC101 C-------CCG--TGGGGAGCGG--TCGCAC-GAT---------CAACGTCAA---CAGT 270 XC0194 C-------CCG--TGGGGAGCGG--TCGCAC-GAT---------CAACGTCAA---CAGT 270 XC0038 C-------CCG--TGGGGAGCGG--TCGCAC-GAT---------CAACGTCAA---CAGT 270 XC12815 C-------CCG--TGGGGAGCGG--TCGCAC-GAT---------CAACGTCAA---CAGT 270 XC2912 C-------CCG--TGGGGAGCGG--TCGCAC-GAT---------CAACGTCAA---CAGT 270 XC098 C-------CCG--TGGGGAGCGG--TCGCAC-GAT---------CAACGTCAA---CAGT 270 XC12878 C-------CCG--TGGGGAGCGG--TCGCAC-GAT---------CAACGTCAA---CAGT 270 XCC11 C-------CCG--TGGGGAGCGG--TCGCAC-GAT---------CAACGTCAA---CAGT 270 XC33913 C-------TCCCGGCAAGGTCGTCACCGCGC-CACC--------CGGCCTCCAGAACAAG 336 UW551 A-------TACGGCGTGAAGCCG-CCCGAGCCGACCGGCGTGGTCGACGTCAA---CAAG 570 AAC00 C-------AAC-ATGCGGGTCGG---CGCGCCGACGGGCACGGTGGAGGTGAA---CAAA 438 EW G--CCGGAAATTCGGTGGCCTTCACCAGCGCCGGCGC-TAATCAGACGGTGCTGCATGAC 771 ET1 G--CCGGGCAGGCCGTTGATTTCCCTGGCGCCAGCGC-CAATCCCACGGTGGTGCATGAC 771 PV GGCTCGGCCACTCCGGTCTCTTTCCCGACGGCCTCGGGCACGCCGACCGTCGTCAACGAG 1077 *
Figure 4-2.Continued
104
XC04 CCGGATCGTGGTTTACAAAGGGGGAAGGTGTTTG-ACGGACACAACAAATCTTTAGTCGG 412 XC018 CCCGATCGTGGTC-ACAA---GGCGAGTTGTTCGTACGGACACAACATTCTCTACGTCGG 104 XCC306 CC-GATCGTGGTTCACAA--GGGCGAGGTGTTCG-ACGGACACAACAATCTCTACGTCGG 326 XCC112 CC-GATCGTGGTTCACAA--GGGCGAGGTGTTCG-ACGGACACAACAATCTCTACGTCGG 326 XC290 CC-GATCGTGGTTCACAA--GGGCGAGGTGTTCG-ACGGACACAACAATCTCTACGTCGG 326 XC112 CC-GATCGTGGTTCACAA--GGGCGAGGTGTTCG-ACGGACACAACAATCTCTACGTCGG 326 XC131 CC-GATCGTGGTTCACAA--GGGCGAGGTGTTCG-ACGGACACAACAATCTCTACGTCGG 326 XC46 CC-GATCGTGGTTCACAA--GGGCGAGGTGTTCG-ACGGACACAACAATCTCTACGTCGG 326 XC257-2 CC-GATCGTGGTTCACAA--GGGCGAGGTGTTCG-ACGGACACAACAATCTCTACGTCGG 326 XC62 CC-GATCGTGGTTCACAA--GGGCGAGGTGTTCG-ACGGACACAACAATCTCTACGTCGG 326 XC126 CC-GATCGTGGTTCACAA--GGGCGAGGTGTTCG-ACGGACACAACAATCTCTACGTCGG 326 XC101 CC-GATCGTGGTTCACAA--GGGCGAGGTGTTCG-ACGGACACAACAATCTCTACGTCGG 326 XC0194 CC-GATCGTGGTTCACAA--GGGCGAGGTGTTCG-ACGGACACAACAATCTCTACGTCGG 326 XC0038 CC-GATCGTGGTTCACAA--GGGCGAGGTGTTCG-ACGGACACAACAATCTCTACGTCGG 326 XC12815 CC-GATCGTGGTTCACAA--GGGCGAGGTGTTCG-ACGGACACAACAATCTCTACGTCGG 326 XC2912 CC-GATCGTGGTTCACAA--GGGCGAGGTGTTCG-ACGGACACAACAATCTCTACGTCGG 326 XC098 CC-GATCGTGGTTCACAA--GGGCGAGGTGTTCG-ACGGACACAACAATCTCTACGTCGG 326 XC12878 CC-GATCGTGGTTCACAA--GGGCGAGGTGTTCG-ACGGACACAACAATCTCTACGTCGG 326 XCC11 CC-GATCGTGGTTCACAA--GGGCGAGGTGTTCG-ACGGACACAACAATCTCTACGTCGG 326 XC33913 AC-CATCGTGGTGCATAA--GGGCGAGGTATTCG-ACGGCAAGGGTCAGACCTACATCGG 392 UW551 CC-GATCGTCGTCAAGGC--GGGTGAGACGTTCA-ATGGCGAGGGCAAGTATTACCGCC- 625 AAC00 CC-CATCGTCGTGAAGGC--CGGCCAGACCTTCG-ACGGCCAGGGCCGGCTCTTCAGCGC 494 EW AC-CATTACCGTGAAAGC--GGGTCAGGTGTTTG-ATGGCAAAGGACAAACCTTCACCG- 826 ET1 AC-CATTACCGTAAAAGC--GGGCCAGGTGTTTG-ATGGTAAAGGGCAAACCTTTACCG- 826 PV AC-CATCAAGGTGGGGCC--TGGCGAGACGTTCG-ACGGCGGCGGCAAGACCTTCACCG- 1132 * ** ** ** ** ** * ** * * XC04 CGGGTCGGGC-ATCGGCGATGG-TTGCAGTCCGAGCACCAGCAACCGATGTTTGTCGT-G 469 XC018 CGGGTTCGGC-ATCG-CGATGGATCGCAGTTCGAGCAACAGCAACCGATGTTCGTCGTCG 162 XCC306 CGGGTCCGGC-ATCGGCGATGGTTCGCAGTCCGAGCACCAGCAACCGATGTTCGTCGTCG 385 XCC112 CGGGTCCGGC-ATCGGCGATGGTTCGCAGTCCGAGCACCAGCAACCGATGTTCGTCGTCG 385 XC290 CGGGTCCGGC-ATCGGCGATGGTTCGCAGTCCGAGCACCAGCAACCGATGTTCGTCGTCG 385 XC112 CGGGTCCGGC-ATCGGCGATGGTTCGCAGTCCGAGCACCAGCAACCGATGTTCGTCGTCG 385 XC131 CGGGTCCGGC-ATCGGCGATGGTTCGCAGTCCGAGCACCAGCAACCGATGTTCGTCGTCG 385 XC46 CGGGTCCGGC-ATCGGCGATGGTTCGCAGTCCGAGCACCAGCAACCGATGTTCGTCGTCG 385 XC257-2 CGGGTCCGGC-ATCGGCGATGGTTCGCAGTCCGAGCACCAGCAACCGATGTTCGTCGTCG 385 XC62 CGGGTCCGGC-ATCGGCGATGGTTCGCAGTCCGAGCACCAGCAACCGATGTTCGTCGTCG 385 XC126 CGGGTCCGGC-ATCGGCGATGGTTCGCAGTCCGAGCACCAGCAACCGATGTTCGTCGTCG 385 XC101 CGGGTCCGGC-ATCGGCGATGGTTCGCAGTCCGAGCACCAGCAACCGATGTTCGTCGTCG 385 XC0194 CGGGTCCGGC-ATCGGCGATGGTTCGCAGTCCGAGCACCAGCAACCGATGTTCGTCGTCG 385 XC0038 CGGGTCCGGC-ATCGGCGATGGTTCGCAGTCCGAGCACCAGCAACCGATGTTCGTCGTCG 385 XC12815 CGGGTCCGGC-ATCGGCGATGGTTCGCAGTCCGAGCACCAGCAACCGATGTTCGTCGTCG 385 XC2912 CGGGTCCGGC-ATCGGCGATGGTTCGCAGTCCGAGCACCAGCAACCGATGTTCGTCGTCG 385 XC098 CGGGTCCGGC-ATCGGCGATGGTTCGCAGTCCGAGCACCAGCAACCGATGTTCGTCGTCG 385 XC12878 CGGGTCCGGC-ATCGGCGATGGTTCGCAGTCCGAGCACCAGCAACCGATGTTCGTCGTCG 385 XCC11 CGGGTCCGGC-ATCGGCGATGGTTCGCAGTCCGAGCACCAGCAACCGATGTTCGTCGTCG 385 XC33913 TGGCCCTGGT-CTGGGCGATGGCTCCCAGAACGAACACCAGCAGCCGCTGTTCGTCGTCG 451 UW551 CGACCAAGGCGATGGGCGACAACTCGCAGAACGAGCACCAGCAGGCCGTGTTCATCCTGG 685 AAC00 CGGGCCCGGG-CTCAACGGCGGCGGCACGGCGGAGACGCATCTGCCGGTCTTCATCCTGG 553 EW CCGGTTCAGAATTAGGCGATGGCGGCCAGTCTGAAAACCAGAAACCGCTGTTTATACTGG 886 ET1 CCGGTTCGGAATTAGGCGATGGTGGCCAGTCGGAAAGCCAGAAACCGCTGTTCAAACTGG 886 PV CAGGCAAGGCTCTGGGAGATGGCGGACAGGGCGAGGGACAGAAGCCGATGTTCGAACTGG 1192 * * * * ** ** * * ** * *
Figure 4-2.Continued

105
XC04 AACAAGGG-GGCACC--T-CCAAAACGTGCGGATGAGCGGGGGCGGCGACGGCA-CCATC 524 XC018 GACAAAGGCGGCACCCTT-CCAAAACGTGCGGATGAGCGGGGGCGGCGACGGCATCCATC 221 XCC306 AACAA-GGCGGCACCCTC--CAAAACGTGCGGATGAGCGGGGGCGGCGACGGCATCCATC 442 XCC112 AACAA-GGTGTCGACGTCACCGAAAGGAATATACAGGTTGA------------------- 425 XC290 AACAA-GGCGGCACCCTC--CAAAACGTGCGGATGAGCGGGGGCGGCGACGGCATCCATC 442 XC112 AACAA-GGCGGCACCCTC--CAAAACGTGCGGATGAGCGGGGGCGGCGACGGCATCCATC 442 XC131 AACAA-GGCGGCACCCTC--CAAAACGTGCGGATGAGCGGGGGCGGCGACGGCATCCATC 442 XC46 AACAA-GGCGGCACCCTC--CAAAACGTGCGGATGAGCGGGGGCGGCGACGGCATCCATC 442 XC257-2 AACAA-GGCGGCACCCTC--CAAAACGTGCGGATGAGCGGGGGCGGCGACGGCATCCATC 442 XC62 AACAA-GGCGGCACCCTC--CAAAACGTGCGGATGAGCGGGGGCGGCGACGGCATCCATC 442 XC126 AACAA-GGCGGCACCCTC--CAAAACGTGCGGATGAGCGGGGGCGGCGACGGCATCCATC 442 XC101 AACAA-GGCGGCACCCTC--CAAAACGTGCGGATGAGCGGGGGCGGCGACGGCATCCATC 442 XC0194 AACAA-GGCGGCACCCTC--CAAAACGTGCGGATGAGCGGGGGCGGCGACGGCATCCATC 442 XC0038 AACAA-GGCGGCACCCTC--CAAAACGTGCGGATGAGCGGGGGCGGCGACGGCATCCATC 442 XC12815 AACAA-GGCGGCACCCTC--CAAAACGTGCGGATGAGCGGGGGCGGCGACGGCATCCATC 442 XC2912 AACAA-GGCGGCACCCTC--CAAAACGTGCGGATGAGCGGGGGCGGCGACGGCATCCATC 442 XC098 AACAA-GGCGGCACCCTC--CAAAACGTGCGGATGAGCGGGGGCGGCGACGGCATCCATC 442 XC12878 AACAA-GGCGGCACCCTC--CAAAACGTGCGGATGAGCGGGGGCGGCGACGGCATCCATC 442 XCC11 AACAA-GGCGGCACCCTC--CAAAACGTGCGGATGAGCGGGGGCGGCGACGGCATCCATC 442 XC33913 AGGAC-GGCGGTAGCCTG--TGCAACGTCAACATGAAGGGCGGCGGCGACGGCGTGCATT 508 UW551 AGCCC-GGCGCGAAGCTG--GAGAACGTGCA---GTATTCCGGCGCCGATGGCATCCACC 739 AAC00 AGCCG-GGCGCGTCCGTG--AAGAACCTGCA---GTTCAAGGGCGGCGACGGCATCCACC 607 EW AAGAC-GGTGCCAGCCTG--AAAAACGTCACCATGGGCGACGACGGGGCGGATGGTATTC 943 ET1 AGGAC-GGGGCCAGCCTG--AAAAACGTCACTATCGGCAATAACGGCGCGGATGGCATTC 943 PV CTGAG-GGCGCCACCCTG--AAAAACGTGGTGTTGGGTGACAACGCTGCCGATGGTGTGC 1249 ** * ** XC04 TGTTGGGGACGCC-ACGTTGAAGGAA-TCCGCAACC-GAACGTCACAGAGGATGCGATGA 581 XC018 TGCTCGGCGACGC-ACGCTGAAGGAAGTCAGCACCCTGAACGTCAGCGAGGATGCGATGA 280 XCC306 TGCTCGGCGACGCCACGCTGAAGAACGTCCACAACCTGAACGTCAGCGAGGATGCGATGA 502 XCC112 ------------------------------------------------------------ XC290 TGCTCGGCGACGCCACGCTGAAGAACGTCCACAACCTGAACGTCAGCGAGGATGCGATGA 502 XC112 TGCTCGGCGACGCCACGCTGAAGAACGTCCACAACCTGAACGTCAGCGAGGATGCGATGA 502 XC131 TGCTCGGCGACGCCACGCTGAAGAACGTCCACAACCTGAACGTCAGCGAGGATGCGATGA 502 XC46 TGCTCGGCGACGCCACGCTGAAGAACGTCCACAACCTGAACGTCAGCGAGGATGCGATGA 502 XC257-2 TGCTCGGCGACGCCACGCTGAAGAACGTCCACAACCTGAACGTCAGCGAGGATGCGATGA 502 XC62 TGCTCGGCGACGCCACGCTGAAGAACGTCCACAACCTGAACGTCAGCGAGGATGCGATGA 502 XC126 TGCTCGGCGACGCCACGCTGAAGAACGTCCACAACCTGAACGTCAGCGAGGATGCGATGA 502 XC101 TGCTCGGCGACGCCACGCTGAAGAACGTCCACAACCTGAACGTCAGCGAGGATGCGATGA 502 XC0194 TGCTCGGCGACGCCACGCTGAAGAACGTCCACAACCTGAACGTCAGCGAGGATGCGATGA 502 XC0038 TGCTCGGCGACGCCACGCTGAAGAACGTCCACAACCTGAACGTCAGCGAGGATGCGATGA 502 XC12815 TGCTCGGCGACGCCACGCTGAAGAACGTCCACAACCTGAACGTCAGCGAGGATGCGATGA 502 XC2912 TGCTCGGCGACGCCACGCTGAAGAACGTCCACAACCTGAACGTCAGCGAGGATGCGATGA 502 XC098 TGCTCGGCGACGCCACGCTGAAGAACGTCCACAACCTGAACGTCAGCGAGGATGCGATGA 502 XC12878 TGCTCGGCGACGCCACGCTGAAGAACGTCCACAACCTGAACGTCAGCGAGGATGCGATGA 502 XCC11 TGCTCGGCGACGCCACGCTGAAGAACGTCCACAACCTGAACGTCAGCGAGGATGCGATGA 502 XC33913 TCATGGGCAGCGGCAAGATGGTCAACTGCGTCAACGACGATGTCAGCGAGGATGCGGTCA 568 UW551 TGCTGGGCAGCGCCACGCTGGACCGCGTCGTCAACCGCCAGGTGGGCGAAGATGCTTTCA 799 AAC00 TGCTGGGCGACGCCAGGCTCGACAACGTGCACGGCCTGCAGACGGACGACGACTTCATCA 667 EW ATCTTTACGGTGAT--GCCAAAA-------------TAGAC---AATCTGCACGTCACCA 985 ET1 ATCTCTATGGCGAT--GCGAAAA-------------TAGAT---AACCTGCACGTCACTA 985 PV ATGTCCGTGCTGCCA-GCGAAAAAGC--CGTGAACGTGGAT---AACGTGCATTGGACCA 1303
Figure 4-2.Continued

106
XC04 C-ATAGAGGGAGT---GCAAC-GGGAACAT-ACTCGCGC-TTGCCGGAAC--GCCTGCCA 632 XC018 CCATCGACTGGTCCTGGCAACCGGGAACATGACTCGCGCATTGCCGGAAC--GCCTGCCA 338 XCC306 CCATCGAC-GGCCCTGGCAACCGGGAACATGACTCGCGCATTGCCGGAAC--GCCTGCCA 559 XCC112 ------------------------------------------------------------ XC290 CCATCGAC-GGCCCTGGCAACCGGGAACATGACTCGCGCATTGCCGGAAC--GCCTGCCA 559 XC112 CCATCGAC-GGCCCTGGCAACCGGGAACATGACTCGCGCATTGCCGGAAC--GCCTGCCA 559 XC131 CCATCGAC-GGCCCTGGCAACCGGGAACATGACTCGCGCATTGCCGGAAC--GCCTGCCA 559 XC46 CCATCGAC-GGCCCTGGCAACCGGGAACATGACTCGCGCATTGCCGGAAC--GCCTGCCA 559 XC257-2 CCATCGAC-GGCCCTGGCAACCGGGAACATGACTCGCGCATTGCCGGAAC--GCCTGCCA 559 XC62 CCATCGAC-GGCCCTGGCAACCGGGAACATGACTCGCGCATTGCCGGAAC--GCCTGCCA 559 XC126 CCATCGAC-GGCCCTGGCAACCGGGAACATGACTCGCGCATTGCCGGAAC--GCCTGCCA 559 XC101 CCATCGAC-GGCCCTGGCAACCGGGAACATGACTCGCGCATTGCCGGAAC--GCCTGCCA 559 XC0194 CCATCGAC-GGCCCTGGCAACCGGGAACATGACTCGCGCATTGCCGGAAC--GCCTGCCA 559 XC0038 CCATCGAC-GGCCCTGGCAACCGGGAACATGACTCGCGCATTGCCGGAAC--GCCTGCCA 559 XC12815 CCATCGAC-GGCCCTGGCAACCGGGAACATGACTCGCGCATTGCCGGAAC--GCCTGCCA 559 XC2912 CCATCGAC-GGCCCTGGCAACCGGGAACATGACTCGCGCATTGCCGGAAC--GCCTGCCA 559 XC098 CCATCGAC-GGCCCTGGCAACCGGGAACATGACTCGCGCATTGCCGGAAC--GCCTGCCA 559 XC12878 CCATCGAC-GGCCCTGGCAACCGGGAACATGACTCGCGCATTGCCGGAAC--GCCTGCCA 559 XCC11 CCATCGAC-GGCCCTGGCAACCGGGAACATGACTCGCGCATTGCCGGAAC--GCCTGCCA 559 XC33913 CCATCGAT-GGCCAGGGCAACCGCGAGCATGATGCGCGCATTGCCGGT-T--GTCGCCCG 624 UW551 CCATCGAC-GGTGTCAAGAACCGCCTGCACGATGCCAAGATCGCCGGGATCGACCCGGCA 858 AAC00 CCGTCGAT-GGCGCGGAGAACCGGGCCGTGGATGCGCAGCGCGCCGGCTATTCCGCCAAG 726 EW ACGTGGGT-------------------GAGGACGCGA---TTACCGTTAA--GCCAAACA 1021 ET1 ACGTGGGG-------------------GAGGACGCGA---TCACCGTGAA--GCCAAACG 1021 PV ACGTCGGC-------------------GAGGACGCGC---TGACCGTGAA--GGGTGAGG 1339 XC04 GCCGCC-GCCGGCGCGT-CCAAAGATCGAAGTGCTCGA--ATTCCTTCGACAATGCCTCC 688 XC018 GCGGCCTGCTCGCGCGT-CCAAAGATCGAAATCCTCGACAGTTTCTTCGACAATGCCTCC 397 XCC306 GCGGCCTGCCGGCGCGT-CCAAAGATCGAAATCCTCGACAGTTCCTTCGACAATGCCTCC 618 XCC112 ------------------------------------------------------------ XC290 GCGGCCTGCCGGCGCGT-CCAAAGATCGAAATCCTCGACAGTTCCTTCGACAATGCCTCC 618 XC112 GCGGCCTGCCGGCGCGT-CCAAAGATCGAAATCCTCGACAGTTCCTTCGACAATGCCTCC 618 XC131 GCGGCCTGCCGGCGCGT-CCAAAGATCGAAATCCTCGACAGTTCCTTCGACAATGCCTCC 618 XC46 GCGGCCTGCCGGCGCGT-CCAAAGATCGAAATCCTCGACAGTTCCTTCGACAATGCCTCC 618 XC257-2 GCGGCCTGCCGGCGCGT-CCAAAGATCGAAATCCTCGACAGTTCCTTCGACAATGCCTCC 618 XC62 GCGGCCTGCCGGCGCGT-CCAAAGATCGAAATCCTCGACAGTTCCTTCGACAATGCCTCC 618 XC126 GCGGCCTGCCGGCGCGT-CCAAAGATCGAAATCCTCGACAGTTCCTTCGACAATGCCTCC 618 XC101 GCGGCCTGCCGGCGCGT-CCAAAGATCGAAATCCTCGACAGTTCCTTCGACAATGCCTCC 618 XC0194 GCGGCCTGCCGGCGCGT-CCAAAGATCGAAATCCTCGACAGTTCCTTCGACAATGCCTCC 618 XC0038 GCGGCCTGCCGGCGCGT-CCAAAGATCGAAATCCTCGACAGTTCCTTCGACAATGCCTCC 618 XC12815 GCGGCCTGCCGGCGCGT-CCAAAGATCGAAATCCTCGACAGTTCCTTCGACAATGCCTCC 618 XC2912 GCGGCCTGCCGGCGCGT-CCAAAGATCGAAATCCTCGACAGTTCCTTCGACAATGCCTCC 618 XC098 GCGGCCTGCCGGCGCGT-CCAAAGATCGAAATCCTCGACAGTTCCTTCGACAATGCCTCC 618 XC12878 GCGGCCTGCCGGCGCGT-CCAAAGATCGAAATCCTCGACAGTTCCTTCGACAATGCCTCC 618 XCC11 GCGGCCTGCCGGCGCGT-CCAAAGATCGAAATCCTCGACAGTTCCTTCGACAATGCCTCC 618 XC33913 GAGGTCAGC--GGCCGG-CCGAAGGTCGACATCATCAATTGCACGTTCAAGAATGCCGCA 681 UW551 TCGATCCCCGGGGGCACGCCCAAGGTGGAGATCATCAACAGCGCGTTCTACGGCGCCAAG 918 AAC00 AGCATCCCTCCGGGCCCGGCCCACGTGGAGATCACGAACTCGTCGTTCCAGAACAGCCAC 786 EW GCGCG--GGCAAAAAAT--CCCACGTTGAAATCACTAACAGTTCCTTCGAGCACGCCTCT 1077 ET1 GCGCG--GGCAAAAAAT--CTCACGTTGAAATCACCAACAGCACTTTCCAGAACGCCTCT 1077 PV GCGGT--GCCAAGGTCA--CCAACCTGAACATCACCAACAGCAGTGCCCAGGGTGCCAAT 1395
Figure 4-2.Continued

107
XC04 CACAAGGTGATCCAGGACAT-CAGGCGGCAGATGTCT--GTAGGTA-CTTCATCGTGAAC 744 XC018 GACAAGGTGATCCAGGACAAGCACGCGGCAGATGTGCTGGTGCGTAACTTCAGCGTGAAC 457 XCC306 GACAAGGTGATCCAGGACAACCAGGCGGCAGATGTGCTGTTGCGTAACGTCAGCGTGAAC 678 XCC112 ------------------------------------------------------------ XC290 GACAAGGTGATCCAGGACAACCAGGCGGCAGATGTGCTGTTGCGTAACGTCAGCGTGAAC 678 XC112 GACAAGGTGATCCAGGACAACCAGGCGGCAGATGTGCTGTTGCGTAACGTCAGCGTGAAC 678 XC131 GACAAGGTGATCCAGGACAACCAGGCGGCAGATGTGCTGTTGCGTAACGTCAGCGTGAAC 678 XC46 GACAAGGTGATCCAGGACAACCAGGCGGCAGATGTGCTGTTGCGTAACGTCAGCGTGAAC 678 XC257-2 GACAAGGTGATCCAGGACAACCAGGCGGCAGATGTGCTGTTGCGTAACGTCAGCGTGAAC 678 XC62 GACAAGGTGATCCAGGACAACCAGGCGGCAGATGTGCTGTTGCGTAACGTCAGCGTGAAC 678 XC126 GACAAGGTGATCCAGGACAACCAGGCGGCAGATGTGCTGTTGCGTAACGTCAGCGTGAAC 678 XC101 GACAAGGTGATCCAGGACAACCAGGCGGCAGATGTGCTGTTGCGTAACGTCAGCGTGAAC 678 XC0194 GACAAGGTGATCCAGGACAACCAGGCGGCAGATGTGCTGTTGCGTAACGTCAGCGTGAAC 678 XC0038 GACAAGGTGATCCAGGACAACCAGGCGGCAGATGTGCTGTTGCGTAACGTCAGCGTGAAC 678 XC12815 GACAAGGTGATCCAGGACAACCAGGCGGCAGATGTGCTGTTGCGTAACGTCAGCGTGAAC 678 XC2912 GACAAGGTGATCCAGGACAACCAGGCGGCAGATGTGCTGTTGCGTAACGTCAGCGTGAAC 678 XC098 GACAAGGTGATCCAGGACAACCAGGCGGCAGATGTGCTGTTGCGTAACGTCAGCGTGAAC 678 XC12878 GACAAGGTGATCCAGGACAACCAGGCGGCAGATGTGCTGTTGCGTAACGTCAGCGTGAAC 678 XCC11 GACAAGGTGATCCAGGACAACCAGGCGGCAGATGTGCTGTTGCGTAACGTCAGCGTGAAC 678 XC33913 GACAAGGTCGTGCAGGACAATGGTGCTGCCGACGTGGTGATGTCGGGTTGCACCGTCAAC 741 UW551 GACAAAGTCGGTCAGATCAACGGCGACGTAGACCTGCAGGTGCGGGGCATGTACGTCAAT 978 AAC00 GACAAGGCGATCCAGATCAACGGCGACGTGGACCTGAAGCTGCGCGGCATCTATGCGAAC 846 EW GACAAGATCCTGCAGCTGAATGCCGATACTAACCTGAGCGTTGACAACGTGAAGGCCAAA 1137 ET1 GACAAGATCCTACAGCTGAATGCCGATACCAGCCTGACCGTTGATAACGTGAAGGCCAAA 1137 PV GACAAGGTGTTCCAGCTCAACGCCGATGCGAACGTGAACGTGGACAACTTCAAGGTCAAG 1455 XC04 GGTCCCGGCAAGGTGTTACGCAC-AACGGCGT-CACACCGACATGGGT-CACACGTCTCG 801 XC018 GGAGGCGGCAAGGTGTTCCGCGCCAACGGCGGACACACCGACATCGATTCACACGTCACG 517 XCC306 GGAGCCGGCAAGGTGTTCCGCACCAACGGCGGACACACCGACATCGATTCACACGTCACG 738 XCC112 ------------------------------------------------------------ XC290 GGAGCCGGCAAGGTGTTCCGCACCAACGGCGGACACACCGACATCGATTCACACGTCACG 738 XC112 GGAGCCGGCAAGGTGTTCCGCACCAACGGCGGACACACCGACATCGATTCACACGTCACG 738 XC131 GGAGCCGGCAAGGTGTTCCGCACCAACGGCGGACACACCGACATCGATTCACACGTCACG 738 XC46 GGAGCCGGCAAGGTGTTCCGCACCAACGGCGGACACACCGACATCGATTCACACGTCACG 738 XC257-2 GGAGCCGGCAAGGTGTTCCGCACCAACGGCGGACACACCGACATCGATTCACACGTCACG 738 XC62 GGAGCCGGCAAGGTGTTCCGCACCAACGGCGGACACACCGACATCGATTCACACGTCACG 738 XC126 GGAGCCGGCAAGGTGTTCCGCACCAACGGCGGACACACCGACATCGATTCACACGTCACG 738 XC101 GGAGCCGGCAAGGTGTTCCGCACCAACGGCGGACACACCGACATCGATTCACACGTCACG 738 XC0194 GGAGCCGGCAAGGTGTTCCGCACCAACGGCGGACACACCGACATCGATTCACACGTCACG 738 XC0038 GGAGCCGGCAAGGTGTTCCGCACCAACGGCGGACACACCGACATCGATTCACACGTCACG 738 XC12815 GGAGCCGGCAAGGTGTTCCGCACCAACGGCGGACACACCGACATCGATTCACACGTCACG 738 XC2912 GGAGCCGGCAAGGTGTTCCGCACCAACGGCGGACACACCGACATCGATTCACACGTCACG 738 XC098 GGAGCCGGCAAGGTGTTCCGCACCAACGGCGGACACACCGACATCGATTCACACGTCACG 738 XC12878 GGAGCCGGCAAGGTGTTCCGCACCAACGGCGGACACACCGACATCGATTCACACGTCACG 738 XCC11 GGAGCCGGCAAGGTGTTCCGCACCAACGGCGGACACACCGACATCGATTCACACGTCACG 738 XC33913 GGCGCCGGCAAGGTCTTTCGTACCAACGGTGGACACGCTGATATCGACACGCATCTTCAG 801 UW551 GGTGCCGGCAAGGTCTTCCGTACCAACGGCGGCGACGAGCAGATCAAGGCGAATGTCGCG 1038 AAC00 CAGATCGGGCAACTGGCCGTCACGCGCGGCGGCTACCCGATCACCG---CGCATGTGGAC 903 EW GACTTTGGTACTTTTGTACGCACTAACGGCGGTCAACAGGGTAACTG---GGATCTGAAT 1194 ET1 GACTTTGGCACTTTCGTGCGCACCAACGGCGGCCAGCAGGGCAACTG---GGATCTGAAT 1194 PV GATTTCGGCACCTTCATGCGCACCAATGGCGGGCAGCAGGGTGACTG---GAACCTGGAC 1512
Figure 4-2.Continued

108
XC04 G-CGAAGACTCCACCCTGAAAAGCATCAAGGAACCAGTATTCCGCCCCGATGCCCCCGGC 860 XC018 GTCGAAGACTCCACCCTTAAAGGCATCAAGGAAGCAGTATTCCGCAGCTATGCGCGCGGC 577 XCC306 GTCGAAGACTCCACCCTGAAAGGCATCAAGGAAGCAGTATTCCGCACCGATGCCCCCGGC 798 XCC112 ------------------------------------------------------------ XC290 GTCGAAGACTCCACCCTGAAAGGCATCAAGGAAGCAGTATTCCGCACCGATGCCCCCGGC 798 XC112 GTCGAAGACTCCACCCTGAAAGGCATCAAGGAAGCAGTATTCCGCACCGATGCCCCCGGC 798 XC131 GTCGAAGACTCCACCCTGAAAGGCATCAAGGAAGCAGTATTCCGCACCGATGCCCCCGGC 798 XC46 GTCGAAGACTCCACCCTGAAAGGCATCAAGGAAGCAGTATTCCGCACCGATGCCCCCGGC 798 XC257-2 GTCGAAGACTCCACCCTGAAAGGCATCAAGGAAGCAGTATTCCGCACCGATGCCCCCGGC 798 XC62 GTCGAAGACTCCACCCTGAAAGGCATCAAGGAAGCAGTATTCCGCACCGATGCCCCCGGC 798 XC126 GTCGAAGACTCCACCCTGAAAGGCATCAAGGAAGCAGTATTCCGCACCGATGCCCCCGGC 798 XC101 GTCGAAGACTCCACCCTGAAAGGCATCAAGGAAGCAGTATTCCGCACCGATGCCCCCGGC 798 XC0194 GTCGAAGACTCCACCCTGAAAGGCATCAAGGAAGCAGTATTCCGCACCGATGCCCCCGGC 798 XC0038 GTCGAAGACTCCACCCTGAAAGGCATCAAGGAAGCAGTATTCCGCACCGATGCCCCCGGC 798 XC12815 GTCGAAGACTCCACCCTGAAAGGCATCAAGGAAGCAGTATTCCGCACCGATGCCCCCGGC 798 XC2912 GTCGAAGACTCCACCCTGAAAGGCATCAAGGAAGCAGTATTCCGCACCGATGCCCCCGGC 798 XC098 GTCGAAGACTCCACCCTGAAAGGCATCAAGGAAGCAGTATTCCGCACCGATGCCCCCGGC 798 XC12878 GTCGAAGACTCCACCCTGAAAGGCATCAAGGAAGCAGTATTCCGCACCGATGCCCCCGGC 798 XCC11 GTCGAAGACTCCACCCTGAAAGGCATCAAGGAAGCAGTATTCCGCACCGATGCCCCCGGC 798 XC33913 GTCGAAAATTGCGATTTCCAACAGGTCAAGGAAGCGGTGTTCCGTACCGATGCGCCGGGC 861 UW551 GTCCGGGACTCGAATTTCCAGAACGTGGCGGAGGCGGTCTTCCGTACCGATTCCAAATAC 1098 AAC00 ATCGCCGACTCCACCGCGCAGAACCTGAAATCCTTCCTCTTCCGCTTCGACTCCCGGCAG 963 EW CTGAGCCATATCAGCGCAGAAGACGGTAAGTTCTCGTTCGTTAAAAGCGATAGCGAGGGG 1254 ET1 CTGAGTCATATCAGCGCGCAGAACGGTAAGTTCTCATTCGTGAAAAGCGACAGCGAGGGA 1254 PV CTGAAAAACATCAGCGCTGAAGACGGCAAGTTCTCCTTCGTGAAGAGCGACAGCGAAGGC 1572 XC04 GCACATG--TCAGGGT----GCA-TGTCCTCGCGAGGAT---GCTCCCCTTCAAGTGGAG 910 XC018 GCACATG--TCAGTCT----GCAGTGCGTCCGCGACGAT---GCTAGCCATCAAGTGGAG 628 XCC306 GCACATG--TCAGTCT----GCATGGCGTCCGCGACGAT---GCTCCCCATCAAGTGGAG 849 XCC112 ------------------------------------------------------------ XC290 GCACATG--TCAGTCT----GCATGGCGTCCGCGACGAT---GCTCCCCATCAAGTGGAG 849 XC112 GCACATG--TCAGTCT----GCATGGCGTCCGCGACGAT---GCTCCCCATCAAGTGGAG 849 XC131 GCACATG--TCAGTCT----GCATGGCGTCCGCGACGAT---GCTCCCCATCAAGTGGAG 849 XC46 GCACATG--TCAGTCT----GCATGGCGTCCGCGACGAT---GCTCCCCATCAAGTGGAG 849 XC257-2 GCACATG--TCAGTCT----GCATGGCGTCCGCGACGAT---GCTCCCCATCAAGTGGAG 849 XC62 GCACATG--TCAGTCT----GCATGGCGTCCGCGACGAT---GCTCCCCATCAAGTGGAG 849 XC126 GCACATG--TCAGTCT----GCATGGCGTCCGCGACGAT---GCTCCCCATCAAGTGGAG 849 XC101 GCACATG--TCAGTCT----GCATGGCGTCCGCGACGAT---GCTCCCCATCAAGTGGAG 849 XC0194 GCACATG--TCAGTCT----GCATGGCGTCCGCGACGAT---GCTCCCCATCAAGTGGAG 849 XC0038 GCACATG--TCAGTCT----GCATGGCGTCCGCGACGAT---GCTCCCCATCAAGTGGAG 849 XC12815 GCACATG--TCAGTCT----GCATGGCGTCCGCGACGAT---GCTCCCCATCAAGTGGAG 849 XC2912 GCACATG--TCAGTCT----GCATGGCGTCCGCGACGAT---GCTCCCCATCAAGTGGAG 849 XC098 GCACATG--TCAGTCT----GCATGGCGTCCGCGACGAT---GCTCCCCATCAAGTGGAG 849 XC12878 GCACATG--TCAGTCT----GCATGGCGTCCGCGACGAT---GCTCCCCATCAAGTGGAG 849 XCC11 GCACATG--TCAGTCT----GCATGGCGTCCGCGACGAT---GCTCCCCATCAAGTGGAG 849 XC33913 GCGACGG--TGAAGCT----GGCAAACCTGAAGACCGAT---GCGCCCGACGAGGTGATT 912 UW551 TCGACTGCGTCGTTCTC---GGACGATGTGAAGTCGGAT---GCGCCCTTCGATGGGCTG 1152 AAC00 TCCACCGTGCGCATCGCCAACACCGACGTGGACGGAGGCCGCACGCCGGTGAACGTGATG 1023 EW CTAAACGTCAATACCAGTGATATCTCACTGGGTGATGTTGAAAACCACTACA---AAGTG 1311 ET1 TTAAACGTCAATACCCAGGATATCTCCCTGGATAATGTGCAAAACCACTACA---AAGTT 1311 PV CTGAACCTGACCACCAGCGGCATTGATCTGAAGAATGTCGAGAACGCCTACAGCAAGCTG 1632
Figure 4-2.Continued

109
XC04 GCCCTGTACCCCAGTATGTCAGATG----CGCCGAGCTCAGCCGAAAGGAATA-GCAGGT 965 XC018 GCCATGTACACCAGTCAGGCAGATGGGGGCATCAGCGTCAGCCGAAAGGAATATACAGGT 688 XCC306 GCACTGTCCCCCAGTCAGGCAGATGGCGCACGCAGCGTCAGCCGAAAGGAATATACAGGT 909 XCC112 ------------------------------------------------------------ XC290 GCACTGTCCCCCAGTCAGGCAGATGGCGCACGCAGCGTCAGCCGAAAGGAATATACAGGT 909 XC112 GCACTGTCCCCCAGTCAGGCAGATGGCGCACGCAGCGTCAGCCGAAAGGAATATACAGGT 909 XC131 GCACTGTCCCCCAGTCAGGCAGATGGCGCACGCAGCGTCAGCCGAAAGGAATATACAGGT 909 XC46 GCACTGTCCCCCAGTCAGGCAGATGGCGCACGCAGCGTCAGCCGAAAGGAATATACAGGT 909 XC257-2 GCACTGTCCCCCAGTCAGGCAGATGGCGCACGCAGCGTCAGCCGAAAGGAATATACAGGT 909 XC62 GCACTGTCCCCCAGTCAGGCAGATGGCGCACGCAGCGTCAGCCGAAAGGAATATACAGGT 909 XC126 GCACTGTCCCCCAGTCAGGCAGATGGCGCACGCAGCGTCAGCCGAAAGGAATATACAGGT 909 XC101 GCACTGTCCCCCAGTCAGGCAGATGGCGCACGCAGCGTCAGCCGAAAGGAATATACAGGT 909 XC0194 GCACTGTCCCCCAGTCAGGCAGATGGCGCACGCAGCGTCAGCCGAAAGGAATATACAGGT 909 XC0038 GCACTGTCCCCCAGTCAGGCAGATGGCGCACGCAGCGTCAGCCGAAAGGAATATACAGGT 909 XC12815 GCACTGTCCCCCAGTCAGGCAGATGGCGCACGCAGCGTCAGCCGAAAGGAATATACAGGT 909 XC2912 GCACTGTCCCCCAGTCAGGCAGATGGCGCACGCAGCGTCAGCCGAAAGGAATATACAGGT 909 XC098 GCACTGTCCCCCAGTCAGGCAGATGGCGCACGCAGCGTCAGCCGAAAGGAATATACAGGT 909 XC12878 GCACTGTCCCCCAGTCAGGCAGATGGCGCACGCAGCGTCAGCCGAAAGGAATATACAGGT 909 XCC11 GCACTGTCCCCCAGTCAGGCAGATGGCGCACGCAGCGTCAGCCGAAAGGAATATACAGGT 909 XC33913 GCCCCCAGCGCTTTCCAGGCCACCGGAGCGACCCGAATCGGGCATCAGCCCTACAGCGGC 972 UW551 GCTCCCAACAAGAGCCAGGTGACGGGCACCAACAAGGTGAGCTACAAGGCCTACTCGGGC 1212 AAC00 GCCGGCAACCCCGCCAACGTGCACGGCACGCGCAGCGTGCGCCAGTCGATCCAGACGGTC 1083 EW CCGATGTCCGCCAACCTGAAGGTGGCTGAATGA--------------------------- 1344 ET1 CCCGCTTCCGCTAACCTGAAGGTGGCCGAATGA--------------------------- 1344 PV CCAGGCTCGACCAACCACAAGGAGGCTTGA------------------------------ 1662 XC04 TTCAGGCTGTAGTATTGCAG-ATGACG-CTGAGTGGCCGCTCGTCCTTGCAGCG-AGCGG 1022 XC018 TTCAAACACTAGTCCTGCACCATGACGACTGGGTGGCCGCTCATCCTTGGAGCGGAGCGG 748 XCC306 TGA--------------------------------------------------------- 912 XCC112 ------------------------------------------------------------ XC290 TGA--------------------------------------------------------- 912 XC112 TGA--------------------------------------------------------- 912 XC131 TGA--------------------------------------------------------- 912 XC46 TGA--------------------------------------------------------- 912 XC257-2 TGA--------------------------------------------------------- 912 XC62 TGA--------------------------------------------------------- 912 XC126 TGA--------------------------------------------------------- 912 XC101 TGA--------------------------------------------------------- 912 XC0194 TGA--------------------------------------------------------- 912 XC0038 TGA--------------------------------------------------------- 912 XC12815 TGA--------------------------------------------------------- 912 XC2912 TGA--------------------------------------------------------- 912 XC098 TGA--------------------------------------------------------- 912 XC12878 TGA--------------------------------------------------------- 912 XCC11 TGA--------------------------------------------------------- 912 XC33913 TGA--------------------------------------------------------- 975 UW551 TGA--------------------------------------------------------- 1215 AAC00 TGA--------------------------------------------------------- 1086 EW ------------------------------------------------------------ ET1 ------------------------------------------------------------ PV ------------------------------------------------------------
Figure 4-2.Continued

110
XC04 GCAACCGACTCACCGGCGCGGCCCCGCGCGACTCAACGCACCTCGGAAATTTCCACCCCG 1082 XC018 GCAACTGACTCACCGGCTCGACCGCGGGCGACTCAGCGCACTTCGGAAATTTCCACGCCG 808 XCC306 ------------------------------------------------------------ XCC112 ------------------------------------------------------------ XC290 ------------------------------------------------------------ XC112 ------------------------------------------------------------ XC131 ------------------------------------------------------------ XC46 ------------------------------------------------------------ XC257-2 ------------------------------------------------------------ XC62 ------------------------------------------------------------ XC126 ------------------------------------------------------------ XC101 ------------------------------------------------------------ XC0194 ------------------------------------------------------------ XC0038 ------------------------------------------------------------ XC12815 ------------------------------------------------------------ XC2912 ------------------------------------------------------------ XC098 ------------------------------------------------------------ XC12878 ------------------------------------------------------------ XCC11 ------------------------------------------------------------ XC33913 ------------------------------------------------------------ UW551 ------------------------------------------------------------ AAC00 ------------------------------------------------------------ EW ------------------------------------------------------------ ET1 ------------------------------------------------------------ PV ------------------------------------------------------------ XC04 -CCAGCCCCATCGACAAGTTCTAGGCATCGCCGACCTGCCAGAGCTTGATGCGCAGCCGG 1141 XC018 TCCAGCCCCTTCACCAAGGTCTAGGCATTGCCGACCTGCCAGAGCTTGATGTCCATCCGG 868 XCC306 ------------------------------------------------------------ XCC112 ------------------------------------------------------------ XC290 ------------------------------------------------------------ XC112 ------------------------------------------------------------ XC131 ------------------------------------------------------------ XC46 ------------------------------------------------------------ XC257-2 ------------------------------------------------------------ XC62 ------------------------------------------------------------ XC126 ------------------------------------------------------------ XC101 ------------------------------------------------------------ XC0194 ------------------------------------------------------------ XC0038 ------------------------------------------------------------ XC12815 ------------------------------------------------------------ XC2912 ------------------------------------------------------------ XC098 ------------------------------------------------------------ XC12878 ------------------------------------------------------------ XCC11 ------------------------------------------------------------ XC33913 ------------------------------------------------------------ UW551 ------------------------------------------------------------ AAC00 ------------------------------------------------------------ EW ------------------------------------------------------------ ET1 ------------------------------------------------------------ PV ------------------------------------------------------------
Figure 4-2.Continued

111
XC04 ACCTCGTTCTGCGAGTCGGCGCAACGCAACCGCGTCTTCGTAACATGATTTCGTCCCCGC 1201 XC018 ACCTCGTTCTGCGAGTCGGTGCAACGCAAC-GCGTCTTCGTAACAGATTATCGTCCCCGC 927 XCC306 ------------------------------------------------------------ XCC112 ------------------------------------------------------------ XC290 ------------------------------------------------------------ XC112 ------------------------------------------------------------ XC131 ------------------------------------------------------------ XC46 ------------------------------------------------------------ XC257-2 ------------------------------------------------------------ XC62 ------------------------------------------------------------ XC126 ------------------------------------------------------------ XC101 ------------------------------------------------------------ XC0194 ------------------------------------------------------------ XC0038 ------------------------------------------------------------ XC12815 ------------------------------------------------------------ XC2912 ------------------------------------------------------------ XC098 ------------------------------------------------------------ XC12878 ------------------------------------------------------------ XCC11 ------------------------------------------------------------ XC33913 ------------------------------------------------------------ UW551 ------------------------------------------------------------ AAC00 ------------------------------------------------------------ EW ------------------------------------------------------------ ET1 ------------------------------------------------------------ PV ------------------------------------------------------------ XC04 CGTGG-GTATG 1211 XC018 CGTTGTAAATA 938 XCC306 ----------- XCC112 ----------- XC290 ----------- XC112 ----------- XC131 ----------- XC46 ----------- XC257-2 ----------- XC62 ----------- XC126 ----------- XC101 ----------- XC0194 ----------- XC0038 ----------- XC12815 ----------- XC2912 ----------- XC098 ----------- XC12878 ----------- XCC11 ----------- XC33913 ----------- UW551 ----------- AAC00 ----------- EW ----------- ET1 ----------- PV -----------
Figure 4-2.Continued

112
W1 MSELLQRLIQTQFQPQNTSQYDWNPSPSHGTSNGQGEGNEHGGRRNGRGGDNFLATTPET 60 W2 MSELLQRLIQTQFQPQNTSQYDWNPSPSHGTS---------------------------- 32 W3 MSELLQRL---------------------------------------------------- 8 ************************************************************ W1 SDHSDKSAHSTVTGSGPVGSGRTINVNSPIVVHKGEVFDGHNNLYVGGSGIGDGSQSEHQ 120 W2 -DHSDKSAHSTVTGSGPVGSGRTINVNSPIVVHKGEVFDGHNNLYVGGSGIGDGSQSEHQ 91 W3 ------------------------------------------------------------ ************************************************************ W1 QPMFVVEQGGTLQNVRMSGGGDGIHLLGDATLKNVHNLNVSEDAMTIDGPGNREHDSRIA 180 W2 QPMFVVEQGGTLQNVRMSGGGDGIHLLGDATLKNVHNLNVSEDAMTIDGPGNREHDSRIA 151 W3 ------------------------------------------------------------ ************************************************************ W1 GTPASGLPARPKIEILDSSFDNASDKVIQDNQAADVLLRNVSVNGAGKVFRTNGGHTDID 240 W2 GTPASGLPARPKIEILDSSFDNASDKVIQDNQAADVLLRNVSVNGAGKVFRTNGGHTDID 211 W3 ------------------------------------------------------------ ************************************************************ W1 SHVTVEDSTLKGIKEAVFRTDAPGAHVSLHGVRDDAPHQVEALSPSQADGARSVSRKEYT 300 W2 SHVTVEDSTLKGIKEAVFRTDAPGAHVSLHGVRDDAPHQVEALSPSQADGARSVSRKEYT 271 W3 ---------------------------------------------------RSVSRKEYT 17 ************************************************************ W1 G 301 W2 G 272 W3 G 18
Figure 4-3. Sequence alignment of Xcc strain 306 – wild type (W1); W::∆Harpin mutant (W2); and 306::∆hrpW HrpW (W3). Dots show parts of hrpW gene that were deleted.

113
Figure 4-4. Phenotype assay in grapefruit leaves. A. Xcc strain 306 wild type (1), W::∆harpin
mutant (circle) and ∆harpin::Harpin complemented (2); B. Xcc strain 306 wild type (1) and 306::∆hrpW mutant. The bacterial suspension at 5 x 108 CFU/mL were syringe and needle infiltrated and photographed 7 days later.
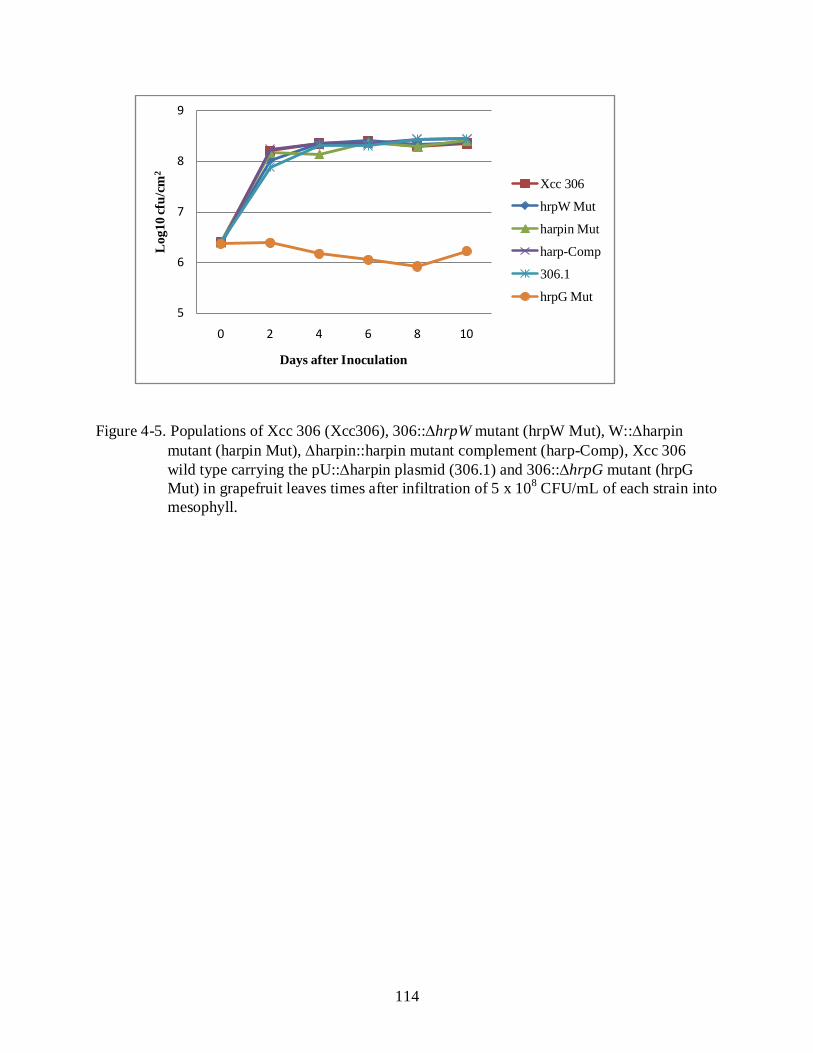
114
Figure 4-5. Populations of Xcc 306 (Xcc306), 306::∆hrpW mutant (hrpW Mut), W::∆harpin
mutant (harpin Mut), ∆harpin::harpin mutant complement (harp-Comp), Xcc 306 wild type carrying the pU::∆harpin plasmid (306.1) and 306::∆hrpG mutant (hrpG Mut) in grapefruit leaves times after infiltration of 5 x 108 CFU/mL of each strain into mesophyll.
5
6
7
8
9
0 2 4 6 8 10
Log
10 cf
u/cm
2
Days after Inoculation
Xcc 306
hrpW Mut
harpin Mut
harp-Comp
306.1
hrpG Mut

115
Figure 4-6. Canker symptoms on abaxial leaf surface of grapefruit. Symptoms developed 28 days after pin-prick inoculation with Xcc 306 (all circled), 306::∆hrpW mutant mutant (A), W::∆harpin mutant (B), ∆harpin::harpin mutant complement (C), Xcc 306 wild type carrying the pU::∆harpin plasmid (D), and 306::∆hrpG mutant (E).
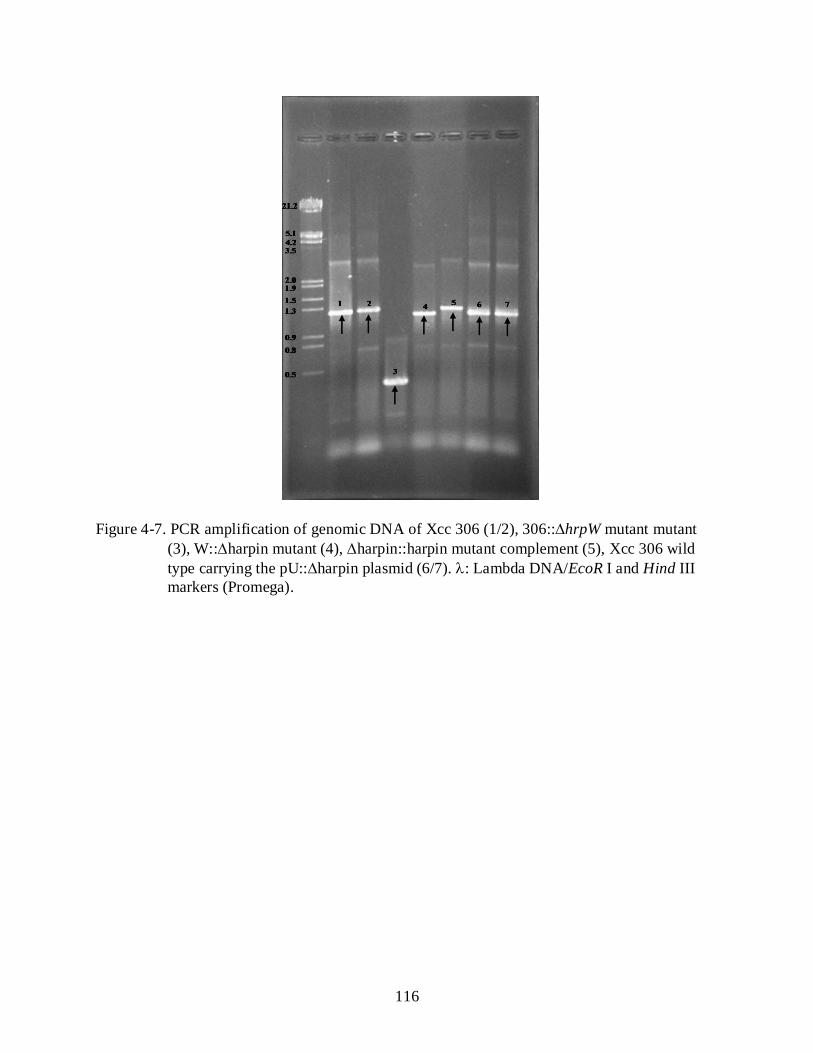
116
Figure 4-7. PCR amplification of genomic DNA of Xcc 306 (1/2), 306::∆hrpW mutant mutant (3), W::∆harpin mutant (4), ∆harpin::harpin mutant complement (5), Xcc 306 wild type carrying the pU::∆harpin plasmid (6/7). λ: Lambda DNA/EcoR I and Hind III markers (Promega).

117
CHAPTER 5 CHARACTERIZATION OF AVRGF1 FROM XANTHOMONAS CITRI SUBSP. CITRI
STRAIN AW
Introduction
Host resistance is often linked to the recognition of specific elicitors encoded by the
pathogen avirulence (avr) genes by a single resistance (R) gene in the host (Hammond-Kosack
and Jones, 1997). This recognition is now commonly referred to as effector-triggered immunity
(ETI) (Flor, 1971; Jones and Dangl, 2006). Phytopathogenic proteobacteria, such as
Xanthomonas, employ a type III secretion system (T3SS) in the initial infection phase to inject
T3-effector proteins through this apparatus (Bonas et al., 1991; Fenselau et al., 1992). The
activity of T3-effectors is required collectively to modulate the plant cell environment to provide
optimum conditions for bacterial colonization (Cornelis and van Gijsegem, 2000). While the
general function of T3-effectors is to suppress plant defenses, some T3-effectors trigger a rapid
and localized programmed cell death known as the hypersensitive reaction (HR) (Lindgren et al.,
1986; Staskawicz et al., 1995;.Block et al., 2008). The list of T3-effectors with dual activity
include avrRpm1 from Pseudomonas syringae pv. maculicola (Ritter and Dangl, 1995), avrXa7
in X. oryzae pv. oryzae (Yang et al., 2004); avrPto of P. s. pv. tomato (Chang et al., 2000) and
avrPtoB in P. s. pv. tomato DC300 (Scofield et al., 1996; Tang et al., 1999; Kim et al., 2002).
Xanthomonas axonopodis pv. citri, recently renamed to Xanthomonas citri subsp. citri –
Xcc (Schaad et al., 2005), is the causal agent of citrus canker, which is one of the most
destructive citrus diseases world-wide. An Xcc strain that attacks Key lime (Citrus aurantifolia)
and alemow (C. macrophylla) plants but not grapefruit (C. paradisi) and orange (C. sinensis)
was isolated and named as Xcc Aw (Schubert et al., 1996; Sun et al., 2004). Rybak et al. (2009)
investigated the factors involved in limiting the ability of XccAw to cause disease in grapefruit
and isolated a single avirulence gene, termed avrGf1 that elicits an HR in grapefruit. The

118
predicted product of avrGf1, AvrGf1 has sequence similarity to HopG1 in P. s. pv. phaseolicola
and P. s. pv. tomato, and a putative T3-effector protein in R. solanacearum (Rybak et al., 2009).
A number of methods have been used to characterize avr proteins including reporter gene
fusions, gene bombardment and Agrobacterium-mediated gene transfer (Duan et al., 1999;
Roden et al., 2004). In order to understand the structural requirements for AvrGf1 function as an
elicitor, transient expression and bacterial reporter gene fusion systems for AvrGf1 were devised.
Materials and Methods
Bacterial Strains and Plasmids
The strains of Escherichia coli, X. citri subsp citri and Agrobacterium strains, and plasmids
used in this study are listed in Table 5-1.
Media and Growth Conditions
Escherichia coli DH5α was used as the cloning host throughout this study and cultured in
Luria Bertani (LB) medium (Sambrook et al., 1989) at 37oC with appropriate antibiotics. For
solid medium, 1.5% BD Bacto agar was added. The strains of Xanthomonas were cultured on
nutrient agar (NA) medium and tryptone sucrose agar at 28oC (Zhu et al., 2000). Agrobacterium
tumefaciens strains were grown in YEP medium (An, 1987) and Murashige and Skoog medium
without plant growth regulators (MSO) (Murashige and Skoog, 1962). Triparental matings were
performed on nutrient-yeast extract-glycerol (NYG) agar (Daniels et al., 1984). All bacterial
strains were stored in 20% glycerol in sterile tap water and maintained at -80oC. Antibiotic
selection used ampicillin (Amp) at 100 µg ml-1; kanamycin (Kn), 50 µg ml-1; rifampicin (Rif),
100 µg ml-1; and spectinomycin (Spc), 100 µg ml-1.
Recombinant DNA Techniques
All DNA manipulations, including isolation of total DNA, the alkaline method of plasmid
purification, ligation and gel electrophoresis were performed as described previously (Sambrook

119
et al., 1989). Restriction enzymes, T4 DNA ligase, and GoTaq® Flexi DNA Polymerase were
used following the manufacturer’s recommendations (Promega, Madison, WI, USA). Chemicals
were purchased from Sigma-Aldrich (St. Louis, MO, USA), and Fisher Scientific (Hampton,
NH, USA). The polymerase chain reaction (PCR) was performed in a PCR Express Thermal
Cycler (ThermoHybaid, Ashford, UK). The Topo TA cloning kit was used for cloning PCR
products (Invitrogen, Carlsbad, CA, USA). All the enzymes and kits were used according to
specifications of the manufacturers. Constructs were transformed into competent Escherichia
coli DH5α cells described by Sambrook et al. (1989). The selected plasmids were introduced
into Xcc 306 by triparental conjugations (Daniels et al., 1984), and into A. tumefaciens GV3101
strains by electroporation (Van Larebeke et al., 1974). Aliquots of competent cells were mixed
with plasmid DNA and electroporated using a BioRad GenePulser II Electroporator instrument.
The conditions applied were 200 OHMS (resistance), 25 µFD (capacitance) and 2.5 Volts. After
the pulse, the cells were immediately diluted by addition of 250 µL of NB medium. The cells
were incubated for 1 h at 28oC with constant shaking, and plated on NA and YEP containing
kanamycin at 50 µg ml-1 and rifampicin at 100 µg ml-1, then incubated at 28oC for 2 days.
Sequencing was completed at Interdisciplinary Center for Biotechnology (ICBR), University of
Florida – Gainesville, FL, USA with an Applied Biosystem model 373 system (Foster City, CA,
USA).
Plant Material and Plant Inoculations
Grapefruit cv. Duncan plants were grown from seed in 15 cm plastic pots in Terra-Lite®
agricultural mix (Scott Sierra Horticultural Products Co., Marysville, OH). The plants were kept
in the glasshouse of the University of Florida in Gainesville at temperatures ranging from 25-
30oC. A scale to standardize the citrus leaves was adopted in this study and consisted of young

120
leaves, (three weeks-old leaves after pruning back the branches), intermediate aged leaves (three
to five weeks-old leaves after the pruning), and old leaves (five or more weeks-old leaves after
pruning). Pepper plants (Capsicum annuum) cv. ECW and its NIL ECW20R containing the Bs2
resistance gene were planted from seeds in Plugmix (W. R. Grace & Co., Cambridge, MA,
USA). After two weeks, the seedlings were transferred to Metromix 300 (W. R. Grace & Co) in
10 cm plastics pots. The plants were kept in the glasshouse at temperatures ranging from 25-
30oC. For preparation of bacterial suspensions, 18 h cultures were harvested from the NA plates,
suspended in sterile tap water, and standardized to an optical density at 600 nm (OD600) = 0.3 (5
x 108 CFU/ml) with a Spectronic 20 Genesys spectrophotometer (Spectronic-UNICAM,
Rochester, NY, USA). Bacterial suspensions of the strains used in this study, except
Agrobacterium strains, were infiltrated at 5 x 108 CFU/ml into the abaxial surface of citrus leaves
by using a hypodermic syringe and needle, and the symptoms were assessed up to 10 days after
inoculation. Plant responses were evaluated 3-4 days after inoculation for water-soaking or
chlorosi, and after 6-7 days for pathogenicity.
Agrobacterium-mediated expression of avrGf1 .
The full-length avrGf1 coding sequence, 1.6 kb fragment, was amplified by PCR using a
forward primer 01-Fwd and reverse primer 02-Rev (Table 5-2). To construct the entry vector
pEAvrGf1, the avrGf1 PCR product was cloned into Gateway entry vector pENTR/D-TOPO
following the manufacturer’s instructions (Invitrogen). The avrGf1 sequence was then inserted
in-frame with the GFP sequence from the GATEWAY vector pGWB5 (Tsuyoshi Nakagawa,
Research Institute for Molecular Genetics, Shimane University, Japan) by performing an LR
reaction between the entry clone and pGWB5, creating pGGf1B5. Additionally, the pEAvrGf1
entry clone was used to construct the binary plasmid pGGf1B2 with the GATEWAY LR Clonase
Mix (Invitrogen) and the destination vector pGWB2 (Tsuyoshi Nakagawa, Research Institute for

121
Molecular Genetics, Shimane University, Japan). The pGWB2, which does not carry GFP
protein, was used as a control to test the possibility that GFP is responsible for the observed
phenotypes. All pGWBs contain NPTII gene for kanamycin selection driven by the NOS
promoter and terminator sequences. The green fluorescent protein (GFP) in the pGWB5 is driven
by the CaMV 35S promoter and terminator.
Ti-plasmid carrying deletions on the N-term and C-term of avrGf1 were constructed by
PCR-amplification, using custom primers (Table 5-2) from the pL799 plasmid DNA. N-terminal
deletions were constructed introducing an ATG before AvrGf1 codon 14 and 117, creating
Gf1::∆N13 and Gf1::∆N116 mutants. We also deleted 7 and 83 amino acids of the C-terminal
region of AvrGf1, creating Gf1::∆C7 and Gf1::∆C83 mutants, respectively, to identify the region
required for the AvrGf1 HR induction in Bs2 pepper plants. Thus, all avrGf1-PCR fragments
were cloned into Gateway entry vector pENTR/D-TOPO following the manufacturer’s
instructions (Invitrogen). The selected entry vector carrying the avrGf1 was then LR-
recombined, where the cloning process involves the transfer of the avrGf1 into the expression
vector (pGWB2 and 5) through a simple recombination reaction that maintains protein reading
frame , using Invitrogen Gateway® manipulations, with pGWB2 and pGWB5 binary vectors
(Table 5-1). Each resulting construct in pGWB2 and pGWB5 binary vectors were conjugated
into A. tumefaciens GV3101 by electroporation and the resulting transconjugants were selected
on YEP medium containing Kn 50 µg ml-1 to use for plant transient expression in citrus.
Young leaves of Grapefruit cv. Duncan were used as a source of tissue for transient
expression assay. The empty vectors pGWB2 and pGWB5 served as negative control plasmids.
Bacteria were cultured overnight at 28oC in YEP liquid medium containing 50 µg ml-1 of
kanamycin with 200rpm shaking. The optical density (OD) of the bacterial cells was measured in

122
the spectrophotometer and adjusted to an OD600 of 0.3 with YEP medium. Three milliters of cells
were collected by centrifugation for 5 minutes at 3500 rpm, and gently resuspended with 0.9 mL
of MSO medium. Acetosyringone was added to a final concentration of 0.01mM. The
agrobacterium suspensions were infiltrated into the mesophyll of young grapefruit leaves.
Bacterial Expression of avrGf1.
The plasmids used in the expression avrGf1 in Xanthomonas included pCR799, a pCR2.1
TOPO-cloning vector. The avrGf1 gene and adjacent region were PCR-amplified with HRF1 and
HRR1 primers from clone pL799 (Table 5-2; Rybak et al., 2009). The parameters used for PCR
were as follows: Step 1. 90oC for 5 min; step 2. 95oC for 1 min; step 3. 55oC for 1 min; step 4.
72oC for 1 min per kb, 30 cycles from step 2 to step 4; step 5. 72oC for 10 min. The PCR
fragments were purified using QIAquick PCR Purification Kit system (QIAGEN). Purified
fragments were directly cloned into pCR2.1-TOPO, containing kanamycin and ampicillin
resistance genes following manufacturer’s protocol. Additionally, the LA plates were augmented
with X-gal (80 µg/mL) and Isopropyl β-D-1-thiogalactopyranoside (IPTG – 0.5mM) for
blue/white screening of the lacZ promoter. To construct pHAvrGf1, the pCR799 and pHM1 were
digested with Xho I and Sal I, respectively, linearizing both plasmids. The digested plasmids
were ligated and transformed into E. coli DH5α. Cloning was confirmed by PCR with primers
flanking the avrGf1 regions, HRF1 and HRR1, as well as sequencing of the PCR fragment with
the same set of primers. The search for nucleotide and amino acid sequence homology was
conducted with the BLAST 2.0 algorithm (http://blast.ncbi.nlm.nih.gov/Blast.cgi) to confirm the
insertion (Altschul et al., 1997). The resulting construct pHAvrGf1 was conjugated into
Xanthomonas citri subsp. citri strain 306 by electroporation, and the resulting transconjugants
were selected on NA medium containing Spc 100 µg ml-1. A construct encoding AvrGf11-

123
106::AvrBs262-574 was generated for translocation assay in pepper cvs. ECW and ECW20R. For
making pBSNGf1 plasmid, a PCR fragment encoding the N-term for AvrGf1 was amplified from
Xcc 306 genomic DNA using NGf1-F1 and NGf1-R1 primers, and cloned into the Bgl II
restriction site in the pBS(BglII::avrBs262-574::HA) plasmid. The BamH I – Kpn I fragment
containing AvrGf11-106::AvrBs262-574 region was excised from the resulting plasmid and ligated
into the pUFR034 vector previously digested by the same enzymes, creating pUF12. The
resulting plasmid was introduced into X. campestris pv. vesicatoria strain XC1922 by triparental
mating. Xcv carrying the construct was then tested for the ability to elicit an AvrBs2-HR in the
two pepper genotypes.
Results
Induction of Hypersensitive Response by Transient Expression of avrGf1 Within Citrus
To investigate whether the induced HR in grapefruit leaves is triggered by AvrGf1 alone or
in combination with other proteins from Xanthomonas, we expressed the full length of avrGf1
under control of the constitutive cauliflower mosaic virus 35S promoter in grapefruit leaves
using Agrobacterium tumefaciens-mediated gene delivery. The amplified avrGf1 fragment was
cloned into pGWB5 and pGWB2 binary vector (Figure 5-1) using Gateway cloning system and
Agrobacterium transformed with pGGf1B5 and pGGf1B5 (both carrying the avrGf1), and pGWB5 and
pGWB2 (empty vectors) were inoculated in grapefruit leaves. Transient expression of avrGf1 under
the control of the 35S Cauliflower mosaic virus promoter in young grapefruit leaves resulted in a
typical HR 4 – 5 days after inoculation (Figure 5-2). The A. tumefaciens strain GV3101 by itself
or carrying pGWB2 or pGWB5, empty vectors, did not show any visible sign of necrosis (Figure
5-2), implying that the HR is induced by the avrGf1 and not due to an Agrobacterium protein, or
the green fluorescent protein (GFP) or other protein expressed by T-DNA genes delivered by the
empty vectors..

124
The N-terminal and C-terminal Are Required for HR Elicitation
Deletion mutants were constructed affecting both ends of avrGf1 (Figure 5-1a). Two
different deletions were performed in each terminal. The plasmids carrying the N- and C-
terminal coding sequence deletions, p5∆N13 and p5∆N116, p5∆C7 and p5∆C83, respectively,
were electroporated into the A. tumefaciens strain GV3101. The resulting strains were tested for
their HR elicitation in grapefruit leaf tissue. The transient expression of the deletion mutants
Gf1::∆N13 and Gf1::∆C7, with 13 deleted amino acids after the start codon and 7 deleted amino
acids before the stop codon, produced an HR comparable to that of A. tumefaciens carrying the
full length avrGf1, 31::Gf1B5 and 31::Gf1B2 (Figure 5-1). However, the HR induction of the
Gf1::∆N116 and Gf1::∆C83, which carry 116 deleted amino acids after the start codon and 83
deleted amino acids before the stop codon, respectively, showed complete absence of a
macroscopical HR induction (Figure 5-1).Thus, both N- and C-terminal are required for HR
elicitation.
Xanthomonas citri subsp. citri Harboring avrGf1 Induces a Hypersensitive Reaction in Grapefruit
To determine if the Xcc, which induces a compatible reaction, was able to delivery AvrGf1
into the plant host cell of grapefruit leaf tissue, and then, trigger an incompatible reaction we
transformed Xcc with pHArGf1, which carries the avrGf1 driver by its own native promoter. The
transconjugants Xcc 306 expressing in-trans the avrGf1 gene (306::avrGf1) were infiltrated into
grapefruit and Valencia leaf tissue at 5 x 108 CFU/ml, and leaves were observed after 5 days for
the development of visible HR (Figure 5-3). The Xcc 306 by itself showed water-soaking in the
edges of the infiltrated area, which is a classical symptom of disease development. The HR in
planta of grapefruit elicited by 306::avrGf1 did not differ among the three repeats.

125
AvrGf1 is Translocated Inside the Host Plant Cell
Rybak et al. (2009) reported that avrGf1 requires a functional T3SS and triggers a HR in
grapefruit plants. To further test if the AvrGf1 is translocated into host plant cells or only
secreted into the apoplast, the N-terminal coding region of AvrGf1 (codons 1 – 106) was fused to
the C-terminal coding region of AvrBs2 (codons 62 – 574), a X. campestris pv. vesicatoria type
III effector (Kearney and Staskawicz, 1990; Bonas, 1991). We inoculated Xcv XV1922
expressing the AvrGf11-106::AvrBs262-574 fusion protein into pepper leaves. We chose Xcv
XV1922 to express the fusion protein because it does not trigger HR in pepper cv. ECW20R due
a natural mutation at avrBs2 gene. The bacterial suspension was infiltrated at 5 x 108 CFU/ml
into the leaf mesophyll. Resistant Bs2 pepper plants specifically recognized AvrBs2 truncated
protein leading to HR 24 h after inoculation (Figure 5-4). Furthermore, no HR was observed in
pepper plants without Bs2 resistant gene (Figure 5-4). This data showed that the first 106 amino
acids were sufficient to translocate AvrGf11-106::AvrBs262-574 fused protein into Bs2 pepper
plants.
Discussion
AvrGf1 was characterized by molecular analysis to understand the role of this protein in
XccAw HR elicitation in grapefruit as originally identified by Rybak et al. (2009) They
determined that AvrGf1 is a homolog of several T3-effectors and it is T3SS-dependent. T3SS
facilitates the translocation of a collection of proteins into plant cells (Buttner and Bonas, 2002).
The translocation signal in the avrBs2 gene of X. c. pv. vesicatoria is located between 1 - 58
amino acid and the region of 62 – 714 amino acids activate Bs2 disease resistance (Mudgett et
al., 2000). In this work, we used AvrBs2 as a sensitive reporter (Guttman et al., 2002; Roden et
al., 2004) to identify if the AvrGf1 is translocated inside the host cell in Bs2 resistance pepper
plants from Xcv strain XV1922 transfomed with the AvrGf11-106::AvrBs262-574 fused protein. The

126
AvrBs2/Bs2 report system has been used to identify new translocation effectors (Roden et al.,
2004). The HR induced for the XV1922, expressing the fusion protein, showed that the
translocation signal is present in the first 106 amino acids in the N-terminus. The fact that the
translocation signal is present in the N-terminus is not surprising, since most T3-effectors carry
the translocation signal in the N-terminal, corroborating the results reported by previous studies
(Leach and White, 1996; Mudgett et al., 2002; Roden et al., 2004). Additionally, Rybak et al.
(2009) demonstrated that AvrGf1 is translocated into grapefruit cells in a T3SS-dependent
manner.
The deletion mutagenesis analysis per the N-terminus showed that deletion of 116 amino
acids affect HR activity in pepper plant cv. ECW20. Gathering the data from the
AvrGf1::AvrBs2 translocation study indicate that the first 116 amino acids on the N-terminal
definitely play important roles in protein translocation and and HR induction. In contrast to the
HopG1 in P. s. pv. tomato DC3000, which is a homolog, that does not elicit cell death in
Nicotiana benthamiana (Wei et al., 2007). Additionally, the deletion of only 83 amino acids in
the C-terminal, directly upstream of the stop codon, was shown to be enough to block cell death
elicitation. While, Xcv AvrBs2 protein required, at least, the deletion of 297 amino acids
upstream of the stop codon to lose the ability to trigger HR in Bs2 pepper plants (Mudgett et al.,
2000).
In this work, an Agrobacterium-mediated transient expression protocol was established for
grapefruit to analyze the gene-for-gene interaction between AvrGf1 and the putative R gene
product controlling the HR response in grapefruit. An Agrobacterium-mediated transient
expression in citrus leaves has been reported, which followed the procedure described by Kapila
et al. (1997) and uses vacuum to infiltrate citrus seedling (Duan et al., 1999). However, the aim

127
of that experiment was to show that the expression of pthA pathogenicity gene from Xcc alone
can induce citrus canker symptoms in the absence of the pathogen and no HR. The symptoms
showed by the transient expression are much smaller than the lesions induced by Xcc, thus, this
difference may be because of the relatively inefficient gene transfer (Duan et al., 1999). In
contrast, in the procedure developed here we exclude the necessity of orange seedlings that are
time-consuming and the vacuum chamber. Furthermore, the procedure here demonstrated the
same level of HR elicitation compared with the Xcc wild type strain. Infiltrated plants showed
stable expression of avrGf1 through the HR in young tissue but the same results were not
observed in intermediate and old leaves. Previous attempts to perform Agrobacterium-mediated
transient expression in mature leaves have failed. The inefficiency of this method to function in
old leaves may be because, in general, plant cells have hard and thick cell walls that reduce
access to the bacterium. In addition, this procedure excludes the need to regenerate transformed
cells that are used commonly for functional analysis of gene regulation. The circumvention of
tissue culture is particularly valuable in the case of gene expression of woody trees such as
citrus, which are difficult to regenerate (Ghorbel et al., 1999; Chávez-Bárcenas et al., 2000;
Tucker et al., 2002).

128
Table 5-1. Bacterial strains and plasmids used in the study Desigantion Relevant characteristics Source Strains X. citri subsp. citri 306 Wild type, Asiatic strain, isolated in Brazil,
RifR DPIa
306::avrGf1 306 carrying pHavrGf1, AmpR, KnR, SpcR This study X. campestris pv. campestris XV1922 Pepper race 6, RifR This study
XV1922-1 1922 carrying pUF12, KnR, RifR This study Agrobacterium tumefaciens GV3101 RifR Lahaye, T.b
31::WB5 GV3101 carrying pGWB5, AmpR, RifR This study 31::WB2 GV3101 carrying pGWB2, AmpR, RifR This study 31::Gf1B5 GV3101 carrying pGGf1B5, AmpR, RifR This study 31::Gf1B2 GV3101 carrying pGGf1B2, AmpR, RifR This study Gf1::∆N13 GV3101 carrying p5∆N13, KnR, RifR This study Gf1::5∆N116 GV3101 carrying p5∆N116, KnR, RifR This study Gf1::5∆C7 GV3101 carrying p5∆C7, KnR, RifR This study Gf1::5∆C83 GV3101 carrying p5∆C83, KnR, RifR This study Gf1::2∆N13 GV3101 carrying p2∆N13, KnR, RifR This study Gf1::2∆N116 GV3101 carrying p2∆N116, KnR, RifR This study Gf1::2∆C7 GV3101 carrying p2∆C7, KnR, RifR This study Gf1::2∆C83 GV3101 carrying p2∆C83, KnR, RifR This study Escherichia coli DH5α FrecAϕ80dlacZ∆M15 Invitrogen
Plasmids
pCR2.1-TOPO Phagemid, AmpR, KnR Invitrogen pENTR-D Entry vector Invitrogen pBluescript-KS+/- Phagemid, pUC derivative; AmpR Stratagene pRK2013 KnR, tra+, mob+ Daniels et
al. (1984) pHM1 Broad-host range vector with pUC19
polylinker, SpR Hopkins et al. (1992)
pLAFR3 TcR, rlx+, RK2 replicon BJSc
pUFR034 KnR, Tn903, IncW, Mob+ De Feyter et al. (1990)
pGWB2 Binary gfp expression vector, contains 35S promoter upstream of attR1-CmR-ccdB-attR2-sgfp
Lahaye, T.
pGWB5 Binary vector Lahaye, T. pBS1 BglII::avrBs262-574::HA of Bluescript-KS+/-,
AmpR Mudgettd
pCR799 avrGf1 total sequence in pCR2.1-TOPO, AmpR, KnR
This study
pHAvrGf1 avrGf1 (pCR799) ligated with pHM1 This study
129
Table 5-1. Continued Desigantion Relevant characteristics Source pBSNGf1 avrGf11-106 fused to avrBs262-574 of pBS1 This study PUF12 avrGf11-106::avrBs262-574 fusion in pUFR034 This study pEavrGf1 avrGf1 total sequence in pENTR-D, AmpR,
KnR This study
pE∆N13 avrGf1 deleted 13 codon after ATG in pENTR-D, AmpR, KnR
This study
pE∆N116 avrGf1 deleted 116 codon after ATG in pENTR-D, AmpR, KnR
This study
pE∆C7 avrGf1 deleted 7 codon before stop codon in pENTR-D, AmpR, KnR
This study
pE∆C83 avrGf1 deleted 83 codon before stop codon in pENTR-D, AmpR, KnR
This study
pGGf1B5 avrGf1 total sequence in pGWB5, RifR, KnR This study pGGf1B2 avrGf1 total sequence in pGWB2, RifR, KnR This study p5∆N13 avrGf1 deleted 13 codon after ATG in
pGWB5, AmpR, KnR This study
p5∆N116 avrGf1 deleted 116 codon after ATG in pGWB5, AmpR, KnR
This study
p5∆C7 avrGf1 deleted 7 codon before stop codon in pGWB5, AmpR, KnR
This study
p5∆C83 avrGf1 deleted 83 codon before stop codon in pGWB5, AmpR, KnR
This study
p2∆N13 avrGf1 deleted 13 codon after ATG in pGWB2, AmpR, KnR
This study
p2∆N116 avrGf1 deleted 116 codon after ATG in pGWB2, AmpR, KnR
This study
p2∆C7 avrGf1 deleted 7 codon before stop codon in pGWB2, AmpR, KnR
This study
p2∆C83 avrGf1 deleted 83 codon before stop codon in pGWB2, AmpR, KnR
This study
a DPI, Division of Plant Industry of the Florida Department of Agriculture and Consumer Services, Gainesville, FL, USA. b T. Lahaye, Martin-Luther-Universität, Halle, Germany. c BJS, B. J. Staskawicz, University of California, Berkley, CA. d M. B. Mudgett, Stanford University, Stanford, CA.
130
Table 5-2. Oligonucleotides sequence used in this study Gene Primer Name Primer sequence Length avrGf1 HRF1 5’-CGCCAGGAAGGGCCTGCCATG-3’ 21
HRR1 5’-CCTCAACCGGGCGCCAAGTTGCGT-3’ 24 01-Fwd 5’-CACCATGGCTCCGSGCSYGCATTCGGC-3’ 27 02-Rev 5’-GTCGCTGCTGGTCATTGACTTTCCTGC-3’ 27 NGF1-F1 5’-GGAAGATCTATTCGAGATGCAGACAGCTCAG-3’ 31 NGF1-R1 5’-GGAAGATCTGCCTTCTTTTTCGCCGGACTTC-3’ 31
GF1::∆N13 N13-F1 5’-CACCATGCTGCACTTGAGAGATACATCCAT -3’ 30 HRR1 5’-CCTCAACCGGGCGCCAAGTTGCGT-3’ 24
GF1::∆N116 N116-F1 5’-CACCATGTTCGTTCTGTCCACGACATCGC -3’ 29 HRR1 5’-CCTCAACCGGGCGCCAAGTTGCGT-3’ 24
GF1::∆C7 HRF1 5’-CGCCAGGAAGGGCCTGCCATG-3’ 21 C07-R1 5’-TCCTGCATAGAATGCCCGTGCAGC-3’ 24
GF1::∆C83 HRF1 5’-CGCCAGGAAGGGCCTGCCATG-3’ 21 C83-R1 5’-TGCAACGGCATTTGCACTAGTAAACG-3’ 26
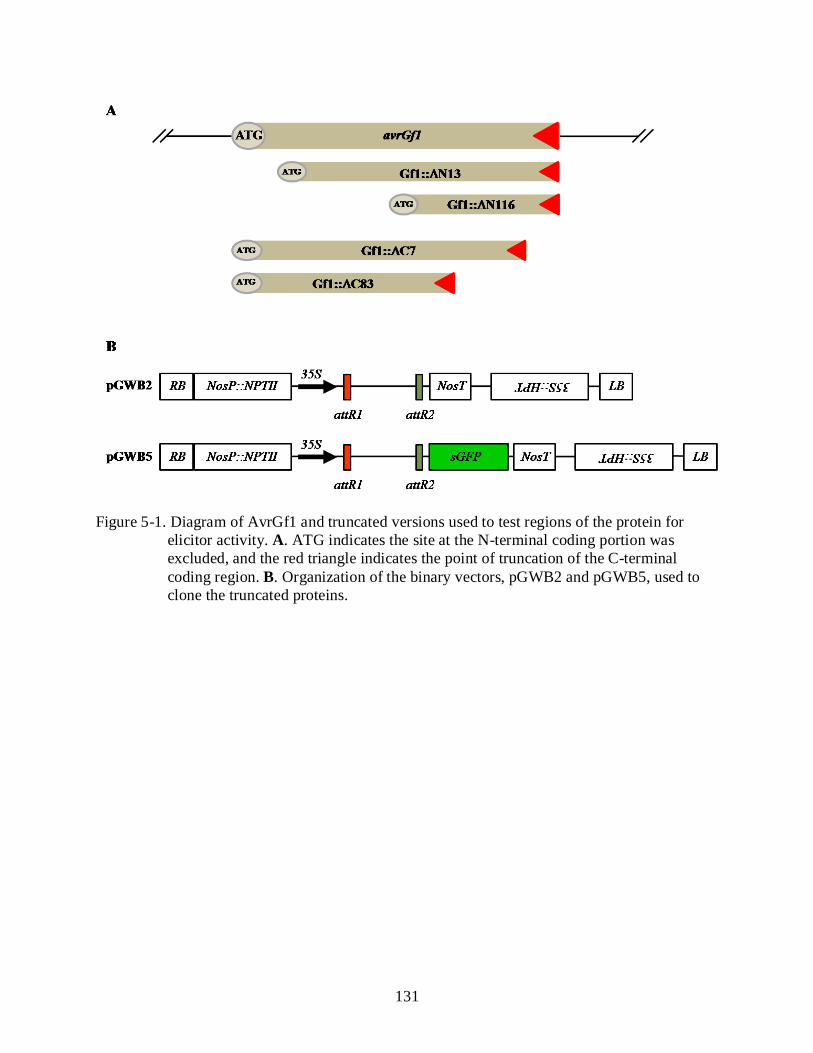
131
Figure 5-1. Diagram of AvrGf1 and truncated versions used to test regions of the protein for elicitor activity. A. ATG indicates the site at the N-terminal coding portion was excluded, and the red triangle indicates the point of truncation of the C-terminal coding region. B. Organization of the binary vectors, pGWB2 and pGWB5, used to clone the truncated proteins.

132
Figure 5-2. Agrobacterium-mediated expression of avrGf1 in grapefruit leaf tissue. A Phenotype elicited by Agrobacterium, harboring 31::Gf1B2 plasmid, grown overnight in YEP medium and directly infiltrated into grapefruit leaves (indicate by circle and arrow) and leaf infiltrated with Agrobacterium, carrying the 31::Gf1B2 plasmid, grown under the conditions developed in the section 5 of materials and methods (indicate arrow), 31::Gf1B5 without cell treatment (indicate by circle) and 31::Gf1B5 treated cells; B. Lower panel: Agrobacterim carrying pGWB5 empty vector, Up panel: Agrobacterium tumefaciens strain GV3101; C. Phenotype of the N-terminal mutants Gf1::∆N13 (low) and Gf1::∆N116 (up); D. Upper panel: HR induced by strain containing 31::Gf1B5; lower panel: C-terminal mutant containing Gf1::∆C7. Bacterial suspensions were prepared following the transient expression protocol developed in this study (see Methods and Materials). The plants were photographed 5 days after infiltration.
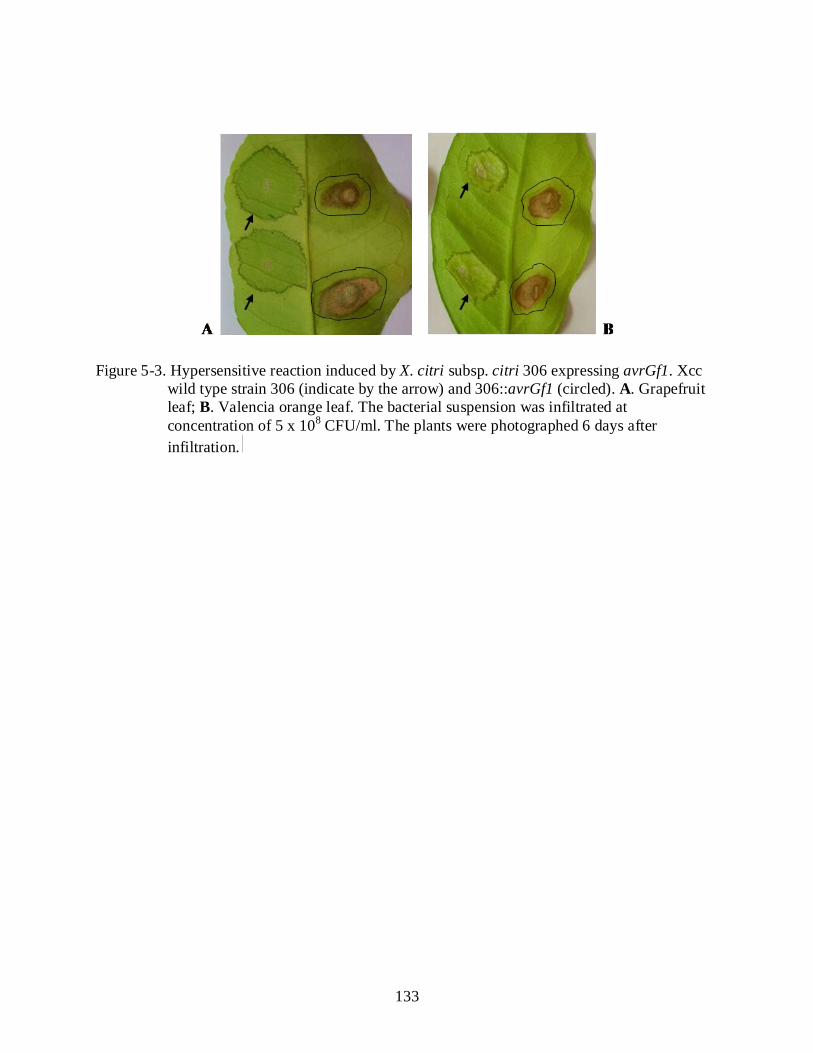
133
Figure 5-3. Hypersensitive reaction induced by X. citri subsp. citri 306 expressing avrGf1. Xcc wild type strain 306 (indicate by the arrow) and 306::avrGf1 (circled). A. Grapefruit leaf; B. Valencia orange leaf. The bacterial suspension was infiltrated at concentration of 5 x 108 CFU/ml. The plants were photographed 6 days after infiltration.

134
Figure 5-4. Phenotypes elicited on pepper plants cv. ECW20R 24 h after inoculation. A. Xcv: X. campestris pv. vesicatoria wild type strain XV1922; Xcv::A, Xcv::B and Xcv::C: different X. c. pv. vesicatoria transconjugants carrying the pUF12. B. Close-up of the X. c. pv. vesicatoria transconjugants, Xcv::A and Xcv::C.

135
CHAPTER 6 OVERALL SUMMARY AND DISCUSSION
The most aggressive form of Asiatic citrus canker disease is caused by Xanthomonas citri
subsp. citri (Xcc) and designated as an A-strain. Another strain, Xanthomonas fuscans pv. citri
(Xfc) and known as a C-strain, is known also to incite citrus canker-like symptoms in
Key/Mexican lime (Citrus aurantiifolia) and hpersensitive response (HR) in grapefruit (C.
paradisi) (Stall and Civerolo, 1991; Rybak et al., 2009). The pathogenicity and virulence of Xcc
rely on the highly specialized type three secretion system (T3SS) delivery system, which
transports a collection of effector proteins directly inside host cells and thereby amending the
host cell environment for bacterial colonization (Bonas et al., 1991; Fenselau et al., 1992;
Cornelis and van Gijsegem, 2000). Hypersensitive response and pathogenicity (hrp) genes
encode the T3SS system, and they are also chaperones for some effectors in the translocation
process (Chang et al., 2004; Tang et al., 2006). The aim of this study was to characterize the
putative T3-effectors involved in the pathogenicity of Xcc. To accomplish this aim and to further
characterize some of the genes that were identified in the process, the study was separated into
four objectives: I. Identification and characterization of candidate T3-effector genes in Xcc based
on genomic sequencing data by da Silva et al. (2002) using mutagenesis analysis; II. Genetic
characterization of a T3SS-independent HR elicitor activity in Xcc; III. Characterization of hrpW
and the domains, harpin and pectate lyase, in the Xcc pathogenicity; and, IV. Molecular
characterization of the Xfc avrGf1 gene and development of avrGf1 as an efficient transient
expression system in citrus leaves.
The candidate effector genes in this study were selected from genes that were scanned for
the presence of the plant inducible promoter (PIP) Box (TTCGC-N15-TTCGC), which is the
transcriptional signal for HrpX regulator (Wengelnik and Bonas, 1996; Wengelnik et al., 1996a;

136
Wengelnik et al., 1996b), another motif called the - 10 box-like motif (YANNNT; Y: C/T; N:
A/T/C/G), which is localized 30 – 32 bases pair from the PIP box that has been proposed to be
necessary for HrpX regulon, and for genes with sequence similarity to known T3-effectors
(Cunnac et al., 2004). Only two candidate T3-effector genes with homology to known T3-
effectors were found without the PIP Box, avrPphE2, which was previously reported by da Silva
et al. (2002) and xopX, which was identified in another Xanthomonas on the basis of an analysis
of T3SS-dependent extracellular protein secretion (Koebnik et al., 2006). Insertional
mutagenesis was used as the primary research approach in this study to evaluate the contribution
of candidate T3-effector genes of Xcc in pathogenicity. Pathogenicity assays in grapefruit
showed that none of the mutants created in this study had visible effects on citrus canker
phenotype, nor had visible effects on the HR elicitation activity on the non-host species tomato.
At the same time, insertions into hrp genes previously known to control T3SS function (hrpA,
hrpG, and hrpX) resulted in loss of pathogenicity. However, mutations in hrpA, hrpG and hrpX
also failed to result in the loss of the nonhost HR on tomato. Concluding, most of the genes
studied here and shown to be conserved in other Xanthomonas were not found to contribute to
the visible canker symptooms. The presence of the PIP-box also does not define the direct
interaction with pathogencity or T3SS, such as avrPphE or xopX which may be pseudogenes and
which may not be involved in pathogenicity. Therefore, the lack of PIP does not mean PIP-box
does not define direct involvement. Further pathogenicity tests, possibly involving bacterial cell
counts, will be conducted to determine what role, if any, the candidate genes play in citrus
canker.
The insertional and deletion mutagenesis performed in Xcc hrpW domains, harpin and
pectate lyase (PEL), demonstrated that the deletion of harpin plays crucial function on Xcc

137
compatible interaction in grapefruit. The hrpW null mutation is not impaired with the
pathogenicity ability. Probably the phenotype observed with harpin mutant is caused by a change
in protein conformation, thus altering the formation of a protein complex that HrpW and the
other proteins (Alegria et al., 2004). Additionally, attempts to complement the harpin mutant
with full length hrpW genes driven by the native promoter in trans failed several times, although,
when the mutation was replaced in the chromosome, the pathogenicity was recovered. This
inability to complement the harpin mutant might be a dominant negative effect of the defective
protein expressed by the mutant.
The Xcc hrp regulator mutants, 306::ΩhrpX and 306::ΩhrpG, preserved the ability to
induce HR in non-host tomato plant. Thus, we speculate that an unknown T3SS-independent
gene, or genes, controls the biosynthesis of the elicitor. Previously, a study by our group and
involving the characterization of Xfc genes that are involved in incompatible host reactions
revealed a clone designated 450, which, when expressed in hrp- mutant of the tomato pathogen
X. perforans, demonstrated same phenotype in non-host tomato plants as Xcc and Xfc. To
identify the possible candidate(s) for the T3-independent HR phenotype, clone 450 was
subcloned, and a screen was performed in tomato plants, revealing a subclone of 3.0 kb that
triggered an HR when expressed in trans in the X. perforans hrp- mutant. Sequence analysis of
the flanking ends of this subclone revealed sequence similarity for the ends of a region
containing three genes in Xcc, namely, XAC3857, XAC3858 and XAC3859. The search for
homology with these three ORFs did not show any relation with known HR elicitor. BLAST
analysis of the NCBI Genbank indicated that none of the ORF had relatedness to any knonw HR-
related proteins. Further characterization of the three genes will reveal whether one or more of
the genes control the T3SS- independent HR phenotype of Xcc and Xfa.

138
Further experiments are in progess to develop an efficient Agrobacterium-mediated
transient expression procedure in citrus leaf tissue. This system will be used to characterize the
critical domains of avrGf1 gene that are required for the HR in grapefruit and as a reporter
system for candidate T3-effector genes of Xcc and related pathogens. In this study, a deletion
mutagenesis analysis made in the N-terminus and C-terminus revealed that the region between
13 and 116 amino acids (aa) in the N-terminus are required for HR elicitation, and also, the
region 7to 83 aa before the stop codon was required. Moreover, a translocation assay using a
AvrGf11-106::AvrBs262-574 fused protein in Bs2 pepper plants, revealed that the secretion signal
resided within the first 106 aa of AvrGf1.

139
APPENDIX A SEQUENCE AND ALIGNMENT
1 ATGTCGGAATTGCTACAGCGGTTGATACAGACCCAATTCCAACCTCAAAACACGTCGCAA
61 TACGATTGGAACCCTTCGCCTTCCCATGGGACGTCCAATGGCCAAGGGGAGGGCAACGAG
121 CATGGCGGTCGCAGGAACGGACGCGGCGGTGACAACTTCCTCGCCACCACGCCCGAGACG
181 TCCGACCATTCCGACAAATCCGCTCACTCGACCGTCACCGGCAGCGGCCCCGTGGGGAGC
241 GGTCGCACGATCAACGTCAACAGTCCGATCGTGGTTCACAAGGGCGAGGTGTTCGACGGA
301 CACAACAATCTCTACGTCGGCGGGTCCGGCATCGGCGATGGTTCGCAGTCCGAGCACCAG
361 CAACCGATGTTCGTCGTCGAACAAGGCGGCACCCTCCAAAACGTGCGGATGAGCGGGGGC
421 GGCGACGGCATCCATCTGCTCGGCGACGCCACGCTGAAGAACGTCCACAACCTGAACGTC
481 AGCGAGGATGCGATGACCATCGACGGCCCTGGCAACCGGGAACATGACTCGCGCATTGCC
541 GGAACGCCTGCCAGCGGCCTGCCGGCGCGTCCAAAGATCGAAATCCTCGACAGTTCCTTC
601 GACAATGCCTCCGACAAGGTGATCCAGGACAACCAGGCGGCAGATGTGCTGTTGCGTAAC
661 GTCAGCGTGAACGGAGCCGGCAAGGTGTTCCGCACCAACGGCGGACACACCGACATCGAT
721 TCACACGTCACGGTCGAAGACTCCACCCTGAAAGGCATCAAGGAAGCAGTATTCCGCACC
781 GATGCCCCCGGCGCACATGTCAGTCTGCATGGCGTCCGCGACGATGCTCCCCATCAAGTG
841 GAGGCACTGTCCCCCAGTCAGGCAGATGGCGCACGCAGCGTCAGCCGAAAGGAATATACA
901 GGTTGA
MSELLQRLIQTQFQPQNTSQYDWNPSPSHGTSNGQGEGNEHGGRRNGRGGDNFLATTPETSDHSDKSAHS
TVTGSGPVGSGRTINVNSPIVVHKGEVFDGHNNLYVGGSGIGDGSQSEHQQPMFVVEQGGTLQNVRMSGG
GDGIHLLGDATLKNVHNLNVSEDAMTIDGPGNREHDSRIAGTPASGLPARPKIEILDSSFDNASDKVIQD
NQAADVLLRNVSVNGAGKVFRTNGGHTDIDSHVTVEDSTLKGIKEAVFRTDAPGAHVSLHGVRDDAPHQV
EALSPSQADGARSVSRKEYTG
Figure A-1. hrpW nucleotide sequence and deduced amino acid sequence.

140
Bgl II
AGATCTATTCGAGATGCAGACAGCTCAGGCCTTCAGGAAAAATAGGTCGTCAATCCGGTAGCAGTGAAATACACGGC
GTTATATGCACATTTACGTCTTGTCATCCCGATGGCTTGTCGCTCCGGATCGCTGGTAACTCACCACGGTCGATGAG
TGCCGCCAGCCCAGGCACGCACAGTCAACACGGCCTCGCCGCGGACTGATAGCAATCTTCCATGTGATACCCGCAGA
GTGGATCGATATGTCGTCGGTAGCGCGCGGCCATGCTTTTCATAGTCCCTATTCCATTCGTCCATCAGCACTTCGCC
TAGTCAACAAGCTCCGATGGCTCCGAGCATGCATTCGGCGGCGTCGCCGGTTTCTGTCCTGCACTTGAGAGATACAT
CCATGGCTCCGAGCATGCATTCGGCGGCGTCGCCGGTTTCTGTCCTGCACTTGAGAGATACATCCATGCGCACCAAA
GCCCAACTCCCATTGACTGCCATTCAACGGTTTCTTGCCCATGATGCAGCGTCAACGCAGGCCCCCTCTGCATCGGC
ATCCACATCGCTCCACAAAAATGAGACCGCAGGCTTGCTGGCAGCCTTGCCAGCGCGAAACGCCAGGCAAGGAGCGC
Bgl II AGAGGAAGTCCGGCGAAAAAGAAGGCAGATCTATGACCGGCAAGCCGGCCCTGGTGGCGCTCGACAGCGAATTCTCC
GAGCAGCGTCTGGCCGAAGTGCAGGCGCGCCAGATCACCGTGCAGACCCTGCAAGGCAAGCTTGCCACGCATCTTGC
GCAGGCCGGCACGGCGCTCAAACCCGACAGCATTGCGGCACGCTTTGCTGCCGGCACACTGGAGCCGGTGTATCTGG
ATACCGCCGCCTTCAATGCCATGTCGCGCGGGCTGCCCGCACGCGCACGTGCGGCCGCAGGCCCGGTGCTGATCGAT
GCACAACAAGGTCGCATCATCTTCAATCTGCAGCGCGCGTTTGCGCCTGGCGACACCTTCAGCGACGCGGCGCTTGC
CGCGCTTGGCAAGCAGTTGAATCTTTCCGGCCACGGGCTGGCAACGCCGAACTGGCTGCAGCCTGCCGCAGGCACGC
CGGGGCGGCGCAAGCTGCAGCAAGCCGCGCGCTATCACGGCCACGAGGTGCCGGCCCGCGACGGTGGCGCCGGGTTC
TTCAAGGCCAACGACCATCGCCTGCTGGAAGGCAAGCAAGTGCTGTTGCGCAATCATCAAAAGTCGCTCGTGCACAA
CCACTACTTCGAAGCACCCAGCACGCGTGCGTTCGGAAAGGACGTCATGGTGCATCGCGGGCTGTTCGATAATCACG
CCGGCATTCCGGAAAACTCGCTGGCGTCCATCGATCATGCCTACGAGCAGGGCTACCGCAATCTGGAGCTGGACGTC
GAAGTCAGTTCCGATGGCGTGCCGGTGTTGATGCACGATTTCAGCATCGGCCGCATGGCAGGCGACCCGCAGAACCG
GTTGGTGTCGCAGGTGCCGTTTGCCGAGCTGCGTGAAATGCCGTTGGTGATTCGCAACCCGTCTGACGGAAACTACG
TCAAGACCGACCAGACCATCGCCGGTGTGGAGCAGATGCTGGAGCACGTGCTCAAAAAGCCCGAGCCGATGTCGGTG
GCGCTGGACTGCAAGGAAAACACCGGCGAAGCAGTGGCGATGCTGCTGATGCGCCGGCCGGACCTGCGCAAGGCTGC
GGCGATCAAGGTCTATGCCAAGTACTACACGGGCGGCTTTGACCAATTCCTGTCCAATTTGTACAAGCACTACCAGA
TCAACCCGTTGCACTCGCAGGATGCGCCGCGCCGCGCCGCGCTGGATCGCTTGCTGGCCAAGATCAACGTGGTGCCG
GTCTTGAGCCAGGGCATGTTGAACGACGAGCGCTTGCGCGGCTTCTTTCGAAGCAATGAGCAGGGCGCCGCGGGGCT
CGCAGACACCGCAATGCAGTGGCTGGACAGCTGGACCAAGATGCGCCCGGTGATCGTGGAGGCGGTGGCCACCGACG
ACAGCGATGCCGGCAAGGCCATGGAAATGGCTCGGACGCGGATGCGCCAGCCGGACTCGGCCTACGCGAAGGCCGCG
TATTCGGTGAGCTACCGGTATGAGGACTTTTCCGTGCCGCGCGCCAATCACGACAAGGACTACTACGTTTACCGCAA
CTTCGGTGAACTGCAAAAGCTCACCAACGAG
Figure A-2. Nucleotide sequence of AvrGf11-106::AvrBs262-574 fused protein. The restriction sites and the start codon of each gene is underline.

141
Bgl II
AGATCTGATGTCTTTGGCCAACACACTGCGCACTGGGGCGGACATACCGCGCCAGACGCGCTCGTTTCGCTACACGC
AACCGCACTTCGGCTTGGCATCGATCAGCACAATCAGCACGCACGATGATCGTCGAATAGCCGACCACGATGGGAGA
GCCTCAGATGATTTCCTCAATCAACCCCTATATCAGTGCTAACCCACTCCTCAACCACACGCTTCAGACGAAGCAGC
CCGCATTTTCCGACACGATCATTATGCTGATGTCGGAATTGCTACAGCGGTTGATACAGACCCAATTCCAACCTCAA
AACACGTCGCAATACGATTGGAACCCTTCGCCTTCCCATGGGACGTCCAATGGCCAAGGGGAGGGCAACGAGCATGG
CGGTCGCAGGAACGGACGCGGCGGTGACAACTTCCTCGCCACCACGCCCGAGACGTCCGACCATTCCGACAAATCCG
CTCACTCGACCGTCACCGGCAGCGGCCCCGTGGGGAGCGGTCGCACGATCAACGTCAACAGTCCGATCGTGGTTCAC
Bgl II
AAGGGCGAGGTGTTCGACGGACACAACAATCTCTACGTCGGCAGATCTATGACCGGCAAGCCGGCCCTGGTGGCGCT
CGACAGCGAATTCTCCGAGCAGCGTCTGGCCGAAGTGCAGGCGCGCCAGATCACCGTGCAGACCCTGCAAGGCAAGC
TTGCCACGCATCTTGCGCAGGCCGGCACGGCGCTCAAACCCGACAGCATTGCGGCACGCTTTGCTGCCGGCACACTG
GAGCCGGTGTATCTGGATACCGCCGCCTTCAATGCCATGTCGCGCGGGCTGCCCGCACGCGCACGTGCGGCCGCAGG
CCCGGTGCTGATCGATGCACAACAAGGTCGCATCATCTTCAATCTGCAGCGCGCGTTTGCGCCTGGCGACACCTTCA
GCGACGCGGCGCTTGCCGCGCTTGGCAAGCAGTTGAATCTTTCCGGCCACGGGCTGGCAACGCCGAACTGGCTGCAG
CCTGCCGCAGGCACGCCGGGGCGGCGCAAGCTGCAGCAAGCCGCGCGCTATCACGGCCACGAGGTGCCGGCCCGCGA
CGGTGGCGCCGGGTTCTTCAAGGCCAACGACCATCGCCTGCTGGAAGGCAAGCAAGTGCTGTTGCGCAATCATCAAA
AGTCGCTCGTGCACAACCACTACTTCGAAGCACCCAGCACGCGTGCGTTCGGAAAGGACGTCATGGTGCATCGCGGG
CTGTTCGATAATCACGCCGGCATTCCGGAAAACTCGCTGGCGTCCATCGATCATGCCTACGAGCAGGGCTACCGCAA
TCTGGAGCTGGACGTCGAAGTCAGTTCCGATGGCGTGCCGGTGTTGATGCACGATTTCAGCATCGGCCGCATGGCAG
GCGACCCGCAGAACCGGTTGGTGTCGCAGGTGCCGTTTGCCGAGCTGCGTGAAATGCCGTTGGTGATTCGCAACCCG
TCTGACGGAAACTACGTCAAGACCGACCAGACCATCGCCGGTGTGGAGCAGATGCTGGAGCACGTGCTCAAAAAGCC
CGAGCCGATGTCGGTGGCGCTGGACTGCAAGGAAAACACCGGCGAAGCAGTGGCGATGCTGCTGATGCGCCGGCCGG
ACCTGCGCAAGGCTGCGGCGATCAAGGTCTATGCCAAGTACTACACGGGCGGCTTTGACCAATTCCTGTCCAATTTG
TACAAGCACTACCAGATCAACCCGTTGCACTCGCAGGATGCGCCGCGCCGCGCCGCGCTGGATCGCTTGCTGGCCAA
GATCAACGTGGTGCCGGTCTTGAGCCAGGGCATGTTGAACGACGAGCGCTTGCGCGGCTTCTTTCGAAGCAATGAGC
AGGGCGCCGCGGGGCTCGCAGACACCGCAATGCAGTGGCTGGACAGCTGGACCAAGATGCGCCCGGTGATCGTGGAG
GCGGTGGCCACCGACGACAGCGATGCCGGCAAGGCCATGGAAATGGCTCGGACGCGGATGCGCCAGCCGGACTCGGC
CTACGCGAAGGCCGCGTATTCGGTGAGCTACCGGTATGAGGACTTTTCCGTGCCGCGCGCCAATCACGACAAGGACT
ACTACGTTTACCGCAACTTCGGTGAACTGCAAAAGCTCACCAACGAG
Figure A-3. Nucleotide sequence of HrpW1-109::AvrBs262-574 fused protein. The restriction sites and the start codon of each gene is underline.

142
LIST OF REFERENCES
Abramovitch, R.B., Kim, Y.J., Chen, S., Dickman, M.B., and Martin, G.B. (2003) Pseudomonas type III effector AvrPtoB induces plant disease susceptibility by inhibition of host programmed cell death. EMBO J. 1, 60-69.
Abramovitch R.B., Anderson J.C., and Martin G.B. (2006) Bacterial elicitation and evasion
of plant innate immunity. Nat Rev Mol Cell Biol 7: 601-611. Abramovitch, R.B., and Martin, G.B. (2004) Strategies used by bacterial pathogens to
suppress plant defenses. Curr. Opin. Plant. Biol. 74, 356-364. Ade J., DeYoung B.J., Golstein C., and Innes R.W. (2007) Indirect activation of a plant
nucleotide binding site-leucine-rich repeat protein by a bacterial protease. Proc Natl Acad Sci U S A 104: 2531-2536.
Agrios, G.N. (1997) Plant Pathology. San Diego: 4th ed. Academic Press. Albright, L.M., Huala, E., and Ausubel, F.M. (1989) Prokaryotic signal transduction mediated
by sensor and regulator protein pairs. Annu. Rev. Genet. 23, 311-336. Alegria, M.C., Docena, C., Khater, L., Ramos, C.H., da Silva, A.C., and Farah, C.S. (2004)
New protein-protein interactions identified for the regulatory and structural components and substrates of the type III Secretion system of the phytopathogen Xanthomonas axonopodis Pathovar citri. J. Bacteriol. 18, 6186-6197.
Alfano, J.R., and Collmer, A. (1997) The type III (Hrp) secretion pathway of plant pathogenic
bacteria: trafficking harpins, Avr proteins, and death. J. Bacteriol. 18, 5655-5662. Alfano, J.R., and Collmer, A. (2004) Type III secretion system effector proteins: double agents
in bacterial disease and plant defense. Annu. Rev. Phytopathol. 42, 385-414. Alfano J.R., Charkowski A.O., Deng W.L., Badel J.L., Petnicki-Ocwieja T., van Dijk K.,
and Collmer A. (2000) The pseudomonas syringae hrp pathogenicity island has a tripartite mosaic structure composed of a cluster of type III secretion genes bounded by exchangeable effector and conserved effector loci that contribute to parasitic fitness and pathogenicity in plants. Proc Natl Acad Sci U S A 97: 4856-4861.
Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang, Z., Miller, W., and Lipman,
D.J. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic. Acids Res. 17, 3389-3402.
An, G. (1987) Binary Ti vectors for plant transformation and promotor analysis. Methods
Enzymol. 153, 292-305.

143
Arlat, M., Van Gijsegem, F., Huet, J.C., Pernollet, J.C., and Boucher, C.A. (1994) PopA1, a protein which induces a hypersensitivity-like response on specific Petunia genotypes, is secreted via the Hrp pathway of Pseudomonas solanacearum. EMBO J. 3, 543-553.
Astua-Monge, G., Minsavage, G.V., Stall, R.E., Vallejos, C.E., Davis, M.J., and Jones, J.B.
(2000) Xv4-vrxv4: a new gene-for-gene interaction identified between Xanthomonas campestris pv. vesicatoria race T3 and wild tomato relative Lycopersicon pennellii. Mol. Plant-Microbe Interact. 12, 1346-1355.
Axtell, M.J., Chisholm, S.T., Dahlbeck, D., and Staskawicz, B.J. (2003) Genetic and
molecular evidence that the Pseudomonas syringae type III effector protein AvrRpt2 is a cysteine protease. Mol. Microbiol. 6, 1537-1546.
Baker, C.J., and Orlandi, E.W. (1995) Active oxygen in plant pathogenesis. Annu. Rev.
Phytopathol. 33, 299-321. Ball, G., Durand, E., Lazdunski, A., and Filloux, A. (2002) A novel type II secretion system in
Pseudomonas aeruginosa. Mol. Microbiol. 2, 475-485. Barny, M.A. (1995) Erwinia amylovora hrpN mutants, blocked in harpin synthesis, express a
reduced virulence on host plants and elicit variable hypersensitive reactions on tobacco. Eur. J. Plant. Pathol. 101, 333-340.
Barny, M.A., Guinebretiere, M.H., Marcais, B., Coissac, E., Paulin, J.P., and Laurent, J.
(1990) Cloning of a large gene cluster involved in Erwinia amylovora CFBP1430 virulence. Mol. Microbiol. 5, 777-786.
Baron, C.O., Callaghan, D., and Lanka, E. (2002) Bacterial secrets of secretion:
EuroConference on the biology of type IV secretion processes. Mol. Microbiol. 5, 1359-1365. Bauer, D.W., Bogdanove, A.J., Beer, S.V., and Collmer, A. (1994) Erwinia chrysanthemi hrp
genes and their involvement in soft rot pathogenesis and elicitation of the hypersensitive response. Mol. Plant-Microbe Interact. 5, 573-581.
Bauer, D.W., Wei, Z.M., Beer, S.V., and Collmer, A. (1995) Erwinia chrysanthemi harpinEch:
an elicitor of the hypersensitive response that contributes to soft-rot pathogenesis. Mol. Plant-Microbe Interact. 4, 484-491.
Bender, C.L., Alarcon-Chaidez, F., and Gross, D.C. (1999) Pseudomonas syringae
phytotoxins: mode of action, regulation, and biosynthesis by peptide and polyketide synthetases. Microbiol. Mol. Biol. Rev. 2, 266-292.
Bennett J.C., and Hughes C. (2000) From flagellum assembly to virulence: The extended
family of type III export chaperones. Trends Microbiol 8: 202-204.

144
Bestwick, C.S., Bennett, M.H., and Mansfield, J.W. (1995) Hrp Mutant of Pseudomonas syringae pv. phaseolicola Induces Cell Wall Alterations but Not Membrane Damage Leading to the Hypersensitive Reaction in Lettuce. Plant Physiol. 2, 503-516.
Block, A., Li, G., Fu, Z.Q., and Alfano, J.R. (2008) Phytopathogen type III effector weaponry
and their plant targets. Curr. Opin. Plant. Biol. 4, 396-403. Bocsanczy, A.M., Nissinen, R.M., Oh, C.S., and Beer, S.V. (2008) HrpN of Erwinia
amylovora functions in the translocation of DspA/E into plant cells. Mol. Plant. Pathol. 4, 425-434.
Bogdanove, A.J., Beer, S.V., Bonas, U., Boucher, C.A., Collmer, A., Coplin, D.L., Cornelis,
G.R., Huang, H.C., Hutcheson, S.W., Panopoulos, N.J., and Van Gijsegem, F. (1996) Unified nomenclature for broadly conserved hrp genes of phytopathogenic bacteria. Mol. Microbiol. 3, 681-683.
Bonas, U., Schulte, R., Fenselau, S., Minsavage, G.V., Staskawicz, B.J., and Stall, R.E.
(1991) Isolation of a gene cluster from Xanthomonas campestris pv. vesicatoria that determines pathogenicity and the hypersensitive response on pepper and tomato. Mol. Plant-Microbe Interact. 4, 81-88.
Bonas, U., and Van den Ackerveken, G. (1999) Gene-for-gene interactions: bacterial
avirulence proteins specify plant disease resistance. Curr. Opin. Microbiol. 1, 94-98. Boucher, C.A., Van Gijsegem, F., Barberis, P.A., Arlat, M., and Zischek, C. (1987)
Pseudomonas solanacearum genes controlling both pathogenicity on tomato and hypersensitivity on tobacco are clustered. J. Bacteriol. 12, 5626-5632.
Brito, B., Marenda, M., Barberis, P., Boucher, C., and Genin, S. (1999) prhJ and hrpG, two
new components of the plant signal-dependent regulatory cascade controlled by PrhA in Ralstonia solanacearum. Mol. Microbiol. 1, 237-251.
Bundock, P., den Dulk-Ras, A., Beijersbergen, A., and Hooykaas, P.J. (1995) Trans-kingdom
T-DNA transfer from Agrobacterium tumefaciens to Saccharomyces cerevisiae. EMBO J. 13, 3206-3214.
Bussiere, D.E., Pratt, S.D., Katz, L., Severin, J.M., Holzman, T., and Park, C.H. (1998) The
structure of VanX reveals a novel amino-dipeptidase involved in mediating transposon-based vancomycin resistance. Mol. Cell, 1, 75-84.
Buttner, D., and Bonas, U. (2002) Port of entry the type III secretion translocon. Trends
Microbiol. 4, 186-192. Buttner. D., and Bonas, U. (2003) Common infection strategies of plant and animal pathogenic
bacteria. Curr. Opin. Plant Biol. 4, 312-319.

145
Buttner, D., Gurlebeck, D., Noel, L.D., and Bonas, U. (2004) HpaB from Xanthomonas campestris pv. vesicatoria acts as an exit control protein in type III-dependent protein secretion. Mol. Microbiol. 3, 755-768.
Buttner, D., Nennstiel, D., Klusener, B., and Bonas, U. (2002) Functional analysis of HrpF, a
putative type III translocon protein from Xanthomonas campestris pv. vesicatoria. J. Bacteriol. 9, 2389-2398.
Cao, H., Baldini, R.L., and Rahme, L.G. (2001) Common mechanisms for pathogens of plants
and animals. Annu. Rev. Phytopathol. 39, 259-284. Cascales, E., and Christie, P.J. (2003) The versatile bacterial type IV secretion systems. Nat.
Rev. Microbiol. 2, 137-149. Casper-Lindley, C., Dahlbeck, D., Clark, E.T., and Staskawicz, B.J. (2002) Direct
biochemical evidence for type III secretion-dependent translocation of the AvrBs2 effector protein into plant cells. Proc. Natl. Acad. Sci. USA, 99, 8336-8341.
Chang, J.H., Goel, A.K., Grant, S.R., and Dangl, J.L. (2004) Wake of the flood: ascribing
functions to the wave of type III effector proteins of phytopathogenic bacteria. Curr. Opin. Microbiol. 1, 11-18.
Chang, J.H., Rathjen, J.P., Bernal, A.J., Staskawicz, B.J., and Michelmore, R.W. (2000)
avrPto enhances growth and necrosis caused by Pseudomonas syringae pv. tomato in tomato lines lacking either Pto or Prf. Mol. Plant-Microbe Interact. 5, 568-571.
Charkowski, A.O., Alfano, J.R., Preston, G., Yuan, J., He, S.Y., and Collmer, A. (1998) The
Pseudomonas syringae pv. tomato HrpW protein has domains similar to harpins and pectate lyases and can elicit the plant hypersensitive response and bind to pectate. J. Bacteriol. 19, 5211-5217.
Chavez-Barcenas, A.T., Valdez-Alarcon, J.J., Martinez-Trujillo, M., Chen, L., Xoconostle-
Cazares, B., Lucas, W.J., and Herrera-Estrella, L. (2000) Tissue-specific and developmental pattern of expression of the rice sps1 gene. Plant Physiol. 2, 641-654.
Chen, G., Zhang, X., and Wang, J. (2000) Characterization of hrp mutants and their conjugates
with hrp gene clones of Xanthomonas oryzae pv. oryzicola. J. Agric. Biotechnol. 8, 117-122. Cho, H.J., Park, Y.J., Noh, T.H., Kim, Y.T., Kim, J.G., Song, E.S., Lee, D.H., and Lee, B.M.
(2008) Molecular analysis of the hrp gene cluster in Xanthomonas oryzae pathovar oryzae KACC10859. Microb. Pathogenesis, 6, 473-483.
Christie, P.J. (2004) Type IV secretion: the Agrobacterium VirB/D4 and related conjugation
systems. Biochim. Biophys. Acta, 1, 219-234.

146
Christie, P.J., Atmakuri, K., Krishnamoorthy, V., Jakubowski, S., and Cascales, E. (2005) Biogenesis, architecture, and function of bacterial type IV secretion systems. Annu. Rev. Microbiol. 59, 451-485.
Christie, P.J., and Vogel, J.P. (2000) Bacterial type IV secretion: conjugation systems adapted
to deliver effector molecules to host cells. Trends Microbiol. 8, 354-360. Ciesiolka, L.D., Hwin, T., Gearlds, J.D., Minsavage, G.V., Saenz, R., and Bravo, M. (1999)
Regulation of expression of avirulence gene avrRxv and identification of a family of host interaction factors by sequence analysis of avrBsT. Mol Plant Microbe Interact 12: 35-44.
Collmer, A., and Keen, N.T. (1986) The role of pectic enzymes in plant pathogenesis. Annu.
Rev. Phytopathol. 24, 383-409. Collmer, A., Lindeberg, M., Petnicki-Ocwieja, T., Schneider, D.J., and Alfano, J.R. (2002)
Genomic mining type III secretion system effectors in Pseudomonas syringae yields new picks for all TTSS prospectors. Trends Microbiol. 10, 462-469.
Cornelis, G.R. (2006) The type III secretion injectisome. Nat Rev Microbiol 4: 811-825. Cornelis, G.R., and Van Gijsegem, F. (2000) Assembly and function of type III secretory
systems. Annu. Rev. Microbiol. 54, 735-774. Cristobal, S., de Gier, J.W., Nielsen, H., and von Heijne, G. (1999) Competition between Sec-
and TAT-dependent protein translocation in Escherichia coli. EMBO J. 11, 2982-2990. Cui, Y., Madi, L., Mukherjee, A., Korsi, D.C., and Chatterjee, A.K. (1996) The Rsm¡
mutants of Erwinia carotovora subsp. carotovora strain Ecc71 overexpress hrpNEcc and elicit a hypersensitive reaction-like response in tobacco leaves. Mol. Plant-Microbe Interact. 9, 565-573.
Cunnac, S., Occhialini, A., Barberis, P., Boucher, C., and Genin, S. (2004) Inventory and
functional analysis of the large Hrp regulon in Ralstonia solanacearum: identification of novel effector proteins translocated to plant host cells through the type III secretion system. Mol. Microbiol. 1, 115-128.
Cuppels, D.A. (1986) Generation and Characterization of Tn5 Insertion Mutations in
Pseudomonas syringae pv. tomato. Appl. Environ. Microbiol. 2, 323-327.

147
da Silva, A.C., Ferro, J.A., Reinach, F.C., Farah, C.S., Furlan, L.R., Quaggio, R.B., Monteiro-Vitorello, C.B., Van Sluys, M.A., Almeida, N.F., Alves, L.M., do Amaral, A.M., Bertolini, M.C., Camargo, L.E., Camarotte, G., Cannavan, F., Cardozo, J., Chambergo, F., Ciapina, L.P., Cicarelli, R.M., Coutinho, L.L., Cursino-Santos, J.R., El-Dorry, H., Faria, J.B., Ferreira, A.J., Ferreira, R.C., Ferro, M.I., Formighieri, E.F., Franco, M.C., Greggio, C.C., Gruber, A., Katsuyama, A.M., Kishi, L.T., Leite, R.P., Lemos, E.G., Lemos, M.V., Locali, E.C., Machado, M.A., Madeira, A.M., Martinez-Rossi, N.M., Martins, E.C., Meidanis, J., Menck, C.F., Miyaki, C.Y., Moon, D.H., Moreira, L.M., Novo, M.T., Okura, V.K., Oliveira, M.C., Oliveira, V.R., Pereira, H.A., Rossi, A., Sena, J.A., Silva, C., de Souza, R.F., Spinola, L.A., Takita, M.A., Tamura, R.E., Teixeira, E.C., Tezza, R.I., Trindade dos Santos, M., Truffi, D., Tsai, S.M., White, F.F., Setubal, J.C, and Kitajima, J.P. (2002) Comparison of the genomes of two Xanthomonas pathogens with differing host specificities. Nature, 417, 459-463.
da Silva, F.G., Shen, Y., Dardick, C., Burdman, S., Yadav, R.C., de Leon, A.L., and
Ronald, P.C. (2004) Bacterial genes involved in type I secretion and sulfation are required to elicit the rice Xa21-mediated innate immune response. Mol. Plant-Microbe Interact. 6, 593-601.
Danese, P.N., and Silhavy, T.J. (1998) Targeting and assembly of periplasmic and outer-
membrane proteins in Escherichia coli. Annu. Rev. Genet. 32, 59-94. Daniels, M. J., Barber, C. E., Turner, D. C., Cleary, W. G., and Sawage, M. (1984) Isolation
of mutants of Xanthomonas campestris pv. campestris showing altered pathogenicity. J. Gen. Microbiol. 130, 2447-2455.
Daniels, M.J., Barber, C.E., Turner, P.C., Sawczyc, M.K., Byrde, R.J., and Fielding, A.H.
(1984) Cloning of genes involved in pathogenicity of Xanthomonas campestris pv. campestris using the broad host range cosmid pLAFR1. EMBO J. 13, 3323-3328.
De Feyter, R., Kado, C.I., and Gabriel, D.W. (1990) Small, stable shuttle vectors for use in
Xanthomonas. Gene, 1, 65-72. De Feyter, R., Yang, Y., and Gabriel, D.W. (1993) Gene-for-genes interactions between cotton
R genes and Xanthomonas campestris pv. malvacearum avr genes. Mol Plant-Microbe Interact. 2, 225-237.
de Oliveira, M.L., Febres, V.J., Costa, M.G., Moore, G.A., and Otoni, W.C. (2008) High-
efficiency Agrobacterium-mediated transformation of citrus via sonication and vacuum infiltration. Plant Cell Rep. 28, 387-395.
Deng, W.L. Preston, G., Collmer, A., Chang, C.J., and Huang, H.C. (1998) Characterization
of the hrpC and hrpRS operons of Pseudomonas syringae pathovars syringae, tomato, and glycinea and analysis of the ability of hrpF, hrpG, hrcC, hrpT, and hrpV mutants to elicit the hypersensitive response and disease in plants. J. Bacteriol. 17, 4523-4531.

148
Deslandes, L., Olivier, J., Peeters, N., Feng, D.X., Khounlotham, M., Boucher, C., Somssich, I., Genin, S., and Marco, Y. (2003) Physical interaction between RRS1-R, a protein conferring resistance to bacterial wilt, and PopP2, a type III effector targeted to the plant nucleus. Proc. Natl. Acad. Sci. USA, 100, 8024-8029.
Ding, Z., Atmakuri, K., and Christie, P.J. (2003) The outs and ins of bacterial type IV
secretion substrates. Trends Microbiol. 11, 527-535. Douglas, C.M., Guidi-Rontani, C., and Collier, R.J. (1987) Exotoxin A of Pseudomonas
aeruginosa: active, cloned toxin is secreted into the periplasmic space of Escherichia coli. J. Bacteriol. 11, 4962-4966.
Dow, J.M., and Daniels, M.J. (2000) Xylella genomics and bacterial pathogenicity to plants.
Yeast, 4, 263-271. Duan, Y.P., Castañeda, A., Zhao, G., Erdos, G., and Gabriel, D.W. (1999) Expression of a
Single, Host-Specific, Bacterial Pathogenicity Gene in Plant Cells Elicits Division, Enlargement, and Cell Death. Mol. Plant-Microbe Interact. 6, 556-560.
Dunger, G., Arabolaza, A. L., Gottig, N., Orellano, E. G., and Ottado, J. (2005) Participation
of Xanthomonas axonopodis pv. citri hrp cluster in citrus canker and nonhost plant responses. Plant Pathol. 54, 781:788.
Evers, S., and Courvalin, P. (1996) Regulation of VanB-type vancomycin resistance gene
expression by the VanS(B)-VanR (B) two-component regulatory system in Enterococcus faecalis V583. J. Bacteriol. 5, 1302-1309.
Felix, G., and Boller, T. (2003) Molecular sensing of bacteria in plants. The highly conserved
RNA-binding motif RNP-1 of bacterial cold shock proteins is recognized as an elicitor signal in tobacco. J. Biol. Chem. 8, 6201-6208.
Felix, G., Duran, J.D., Volko, S., and Boller, T. (1999) Plants have a sensitive perception
system for the most conserved domain of bacterial flagellin. Plant J. 3, 265-276. Fenselau, S., and Bonas, U. (1995) Sequence and expression analysis of the HrpB pathogenicity
operon of Xanthomonas campestris pv. vesicatoria which encodes eight proteins with similarity to components of the Hrp, Ysc, Spa, and Fli secretion systems. Mol. Plant-Microbe Interact. 6, 845-854.
Filloux, A., Michel, G., and Bally, M. (1998) GSP-dependent protein secretion in gram-
negative bacteria: the Xcp system of Pseudomonas aeruginosa. FEMS Microbiol. Rev. 3, 177-198.
Fischer, W., Haas, R., and Odenbreit, S. (2002) Type IV secretion systems in pathogenic
bacteria. Int. J. Med. Microbiol. 292, 159-168.

149
Flor, H.H. (1971) Current status of the gene-for-gene concept. Annu. Rev. Phytopathol. 9, 275-296.
Frederick, R.D., Majerczak, D.R., and Coplin, D.L. (1991) Characterization of wts genes
from Erwinia stewartii and their homology with the hrp gene cluster from Pseudomonas syringae pv. phaseolicola. Phytopathology, 81, 1187.
Fu, Z.Q., Guo, M., Jeong, B.R., Tian, F., Elthon, T.E., Cerny, R.L., Staiger, D., and Alfano,
J.R. (2007) A type III effector ADP-ribosylates RNA-binding proteins and quells plant immunity. Nature, 447, 284-288.
Furutani, A., Nakayama, T., Ochiai, H., Kaku, H., Kubo, Y., and Tsuge, S. (2006)
Identification of novel HrpXo regulons preceded by two cis-acting elements, a plant-inducible promoter box and a -10 box-like sequence, from the genome database of Xanthomonas oryzae pv. oryzae. FEMS Microbiol. Lett. 1, 133-141.
Furutani, A., Takaoka, M., Sanada, H., Noguchi, Y., Oku, T., Tsuno, K., Ochiai, H., and
Tsuge, S. (2009) Identification of novel type III secretion effectors in Xanthomonas oryzae pv. oryzae. Mol. Plant-Microbe Interact. 1, 96-106.
Furutani, A., Tsuge, S., Ohnishi, K., Hikichi, Y., Oku, T., Tsuno, K., Inoue, Y., Ochiai, H.,
Kaku, H., and Kubo, Y. (2004) Evidence for HrpXo-dependent expression of type II secretory proteins in Xanthomonas oryzae pv. oryzae. J. Bacteriol. 5, 1374-1380.
Gabriel, D.W. (1999) Why do pathogens carry avirulence genes? Physiol. Mol. Plant Pathol.
55, 205-214. Galan, J.E., and Collmer, A. (1999) Type III secretion machines: bacterial devices for protein
delivery into host cells. Science, 284, 1322-1328. Gaudriault, S., Brisset, M.N., and Barny, M.A. (1998) HrpW of Erwinia amylovora, a new
Hrp-secreted protein. FEBS Lett. 3, 224-228. Gaudriault, S., Paulin, J.P., and Barny, M.A. (2002) The DspB/F protein of erwinia
amylovora is a type III secretion chaperone ensuring efficient secretion of the DspA/E essential pathogenicity factor. Mol Plant Pathol 5: 313-313.
Genin, S., Gough, C.L., Zischek, C., and Boucher, C.A. (1992) Evidence that the hrpB gene
encodes a positive regulator of pathogenicity genes from Pseudomonas solanacearum. Mol. Microbiol. 20, 3065-3076.
Ghorbel, R., Juarez, J., Navarro, L., and Pena, L. (1999) Green fluorescent protein as a
screenable marker to increase the efficiency of generating transgenic woody fruit plants. Theor. Appl. Genet. 99, 350-358.

150
Gomez-Gomez, L., and Boller, T. (2002) Flagellin perception: a paradigm for innate immunity. Trends Plant Sci. 6, 251-256.
Gozzo, F. (2003) Systemic acquired resistance in crop protection: from nature to a chemical
approach. J. Agric. Food Chem. 16, 4487-4503. Greenberg, J.T., and Yao, N. (2004) The role and regulation of programmed cell death in
plant-pathogen interactions. Cell Microbiol 6: 201-211. Gray, L., Baker, K., Kenny, B., Mackman, N., Haigh, R., and Holland, I.B. (1989) A novel
C-terminal signal sequence targets Escherichia coli haemolysin directly to the medium. J. Cell Sci. Suppl. 11, 45-47.
Grohmann, E., Muth, G., and Espinosa, M. (2003) Conjugative plasmid transfer in gram-
positive bacteria. Microbiol. Mol. Biol. Rev. 2, 277-301. Gu, K., Yang, B., Tian, D., Wu, L., Wang, D., Sreekala, C., Yang, F., Chu, Z., Wang, G.L.,
White, F.F., and Yin, Z. (2005) R gene expression induced by a type-III effector triggers disease resistance in rice. Nature, 435, 1122-1125.
Guo, W., Gonzalez-Candelas, L., and Kolattukudy, P.E. (1995) Cloning of a new pectate
lyase gene pelC from Fusarium solani f. sp. pisi (Nectria haematococca, mating type VI) and characterization of the gene product expressed in Pichia pastoris. Arch. Biochem. Biophys. 323, 352-360.
Gurlebeck, D., Thieme, F., and Bonas, U. (2006) Type III effector proteins from the plant
pathogen Xanthomonas and their role in the interaction with the host plant. J. Plant Physiol. 3, 233-255.
Guttman, D.S., and Greenberg, J.T. (2001) Functional analysis of the type III effectors
AvrRpt2 and AvrRpm1 of Pseudomonas syringae with the use of a single-copy genomic integration system. Mol. Plant-Microbe Interact. 2, 145-155.
Guttman, D.S., Vinatzer, B.A., Sarkar, S.F., Ranall, M.V., Kettler, G., and Greenberg, J.T.
(2002) A functional screen for the type III (Hrp) secretome of the plant pathogen Pseudomonas syringae. Science, 295, 1722-1726.
Haase, J., Lurz, R., Grahn, A.M., Bamford, D.H., and Lanka, E. (1995) Bacterial
conjugation mediated by plasmid RP4: RSF1010 mobilization, donor-specific phage propagation, and pilus production require the same Tra2 core components of a proposed DNA transport complex. J. Bacteriol. 16, 4779-4791.
Hacker, J., and Kaper, J.B. (2000) Pathogenicity islands and the evolution of microbes. Annu.
Rev. Microbiol. 54, 641-679. Hammerschmidt, R. (1999a) Induced disease resistance: how to induced plants stop pathogens?
Physiol. Mol. Plant Pathol. 55, 77-84.

151
Hammerschmidt, R. (1999b) Phytoalexins: What Have We Learned After 60 Years? Annu. Rev.
Phytopathol. 37, 285-306. Hammond-Kosack, K.E., and Jones, J.D. (1997) Plant Disease Resistance Genes. Annu. Rev.
Plant Physiol. 48, 575-607. He, P., Shan, L., Lin, N.C., Martin, G.B., Kemmerling, B., Nurnberger, T., and Sheen, J.
(2006) Specific bacterial suppressors of MAMP signaling upstream of MAPKKK in arabidopsis innate immunity. Cell 125: 563-575.
He, S.Y., Huang, H.C., and Collmer, A. (1993) Pseudomonas syringae pv. syringae harpinPss:
a protein that is secreted via the Hrp pathway and elicits the hypersensitive response in plants. Cell, 7, 1255-1266.
He, S.Y., Nomura, K., and Whittam, T.S. (2004) Type III protein secretion mechanism in
mammalian and plant pathogens. Biochim. Biophys. Acta, 1694, 181-206. Heath, M.C. (2000) Nonhost resistance and nonspecific plant defenses. Curr. Opin. Plant Biol.
4, 315-319. Higgins, C.F. (1995) The ABC of channel regulation. Cell, 5, 693-696. Hommais, F., Oger-Desfeux, C., Van Gijsegem, F., Castang, S., Ligori, S., Expert, D.,
Nasser, W., and Reverchon, S. (2008) PecS is a global regulator of the symptomatic phase in the phytopathogenic bacterium Erwinia chrysanthemi 3937. J. Bacteriol. 190, 7508-7522.
Hopkins, C.M., White, F.F., Choi, S.H., Guo, A., and Leach, J.E. (1992) Identification of a
family of avirulence genes from Xanthomonas oryzae pv. oryzae. Mol. Plant-Microbe Interact. 6, 451-459.
Huang, H.C., Schuurink, R., Denny, T.P., Atkinson, M.M., Baker, C.J., Yucel, I.,
Hutcheson, S.W., and Collmer, A. (1988) Molecular cloning of a Pseudomonas syringae pv. syringae gene cluster that enables Pseudomonas fluorescens to elicit the hypersensitive response in tobacco plants. J. Bacteriol. 10, 4748-4756.
Huelsenbeck, J.P., Ronquist, F., Nielsen, R., and Bollback, J.P. (2001) Bayesian inference of
phylogeny and its impact on evolutionary biology. Science 294: 2310-2314. Hueck, C.J. (1998) Type III protein secretion systems in bacterial pathogens of animals and
plants. Microbiol. Mol. Biol. Rev. 2, 379-433. Huguet, E., Hahn, K., Wengelnik, K., and Bonas, U. (1998) hpaA mutants of Xanthomonas
campestris pv. vesicatoria are affected in pathogenicity but retain the ability to induce host-specific hypersensitive reaction. Mol. Microbiol. 6, 1379-1390.

152
Hunt, M.D., Neuenschwander, U.H., Delaney, T.P., Weymann, K.B., Friedrich, L.B., Lawton, K.A., Steiner, H.Y., and Ryals, J.A. (1996) Recent advances in systemic acquired resistance research a review. Gene, 1, 89-95.
Innes, R.W. (2004) Targeting the targets of type III effector proteins secreted by
phytopathogenic bacteria. Mol. Plant Pathol. 2, 109-113. Innes, R.W., Bent, A.F., Kunkel, B.N., Bisgrove, S.R., and Staskawicz, B.J. (1993) Molecular
analysis of avirulence gene avrRpt2 and identification of a putative regulatory sequence common to all known Pseudomonas syringae avirulence genes. J. Bacteriol. 15, 4859-4869.
Jha, G., Rajeshwari, R., and Sonti, R.V. (2007) Functional interplay between two
Xanthomonas oryzae pv. oryzae secretion systems in modulating virulence on rice. Mol. Plant-Microbe Interact. 1, 31-40.
Jin, Q., Thilmony, R., Zwiesler-Vollick, J., and He, S.Y. (2003) Type III protein secretion in
Pseudomonas syringae. Microbes Infect. 4, 301-310. Jones, J.B., Minsavage, G.V., Stall, R.E., Kelly, R.O., and Bouzar, H. (1993) Genetic
Analysis of a DNA Involved in Expression of Two Epitopes Associated with Lipopolysaccharide in Xanthomonas campestris pv. vesicatoria. Mol. Plant Pathol. 83, 551-556.
Jones, J.D., and Dangl, J.L. (2006) The plant immune system. Nature, 444, 323-329. Kaewnum, S., Prathuangwong, S., and Burr, T.J. (2006) A Pectate Lyase Homolog, xagP, in
Xanthomonas axonopodis pv. glycines Is Associated with Hypersensitive Response Induction on Tobacco. Phytopathology, 11, 1230-1236.
Kajava, A.V. (1998) Structural diversity of leucine-rich repeat proteins. J. Mol. Biol. 3, 519-527. Kamdar, H.V., Kamoun, S., and Kado, C.I. (1993) Restoration of pathogenicity of avirulent
Xanthomonas oryzae pv. oryzae and X. campestris pathovars by reciprocal complementation with the hrpXo and hrpXc genes and identification of HrpX function by sequence analyses. J. Bacteriol. 7, 2017-2025.
Kapila, J., DeRycke, R., Van Montagu, M., and Angenon, G. (1997) An Agrobacterium-
mediated transient gene expression system for intact leaves. Plant Sci. 122, 101-108. Kariola, T., Palimaki, T.A., Brader, G., and Palva, E.T. (2003) Erwinia carotovora subsp.
carotovora and Erwinia-derived elicitors HrpN and PehA trigger distinct but interacting defense responses and cell death in Arabidopsis. Mol. Plant-Microbe Interact. 16, 179-187.
Kay, S., Hahn, S., Marois, E., Hause, G., and Bonas, U. (2007) A bacterial effector acts as a
plant transcription factor and induces a cell size regulator. Science, 318, 648-651.

153
Kearney, B., and Staskawicz, B.J. (1990) Widespread distribution and fitness contribution of Xanthomonas campestris avirulence gene avrBs2. Nature, 346, 385-386.
Keshavarzi, M., Soylu, S., Brown, I., Bonas, U., Nicole, M., Rossiter, J., and Mansfield, J.
(2004) Basal defenses induced in pepper by lipopolysaccharides are suppressed by Xanthomonas campestris pv. vesicatoria. Mol. Plant-Microbe Interact. 7, 805-815.
Kim, H.S., Desveaux, D., Singer, A.U., Patel, P., Sondek, J., and Dangl, J.L. (2005) The
pseudomonas syringae effector AvrRpt2 cleaves its C-terminally acylated target, RIN4, from arabidopsis membranes to block RPM1 activation. Proc Natl Acad Sci U S A 102: 6496-6501.
Kim, J.F., and Beer, S.V. (1998) HrpW of Erwinia amylovora, a new harpin that contains a
domain homologous to pectate lyases of a distinct class. J. Bacteriol. 19, 5203-5210. Kim, J.G., Park, B.K., Yoo, C.H., Jeon, E., Oh, J., and Hwang, I. (2003) Characterization of
the Xanthomonas axonopodis pv. glycines Hrp pathogenicity island. J. Bacteriol. 10, 3155-3166.
Kim, M.G., da Cunha, L., McFall, A.J., Belkhadir, Y., DebRoy, S., Dangl, J.L., and
Mackey, D. (2005) Two pseudomonas syringae type III effectors inhibit RIN4-regulated basal defense in arabidopsis. Cell 121: 749-759.
Kim, Y.J., Lin, N.C., and Martin, G.B. (2002) Two distinct pseudomonas effector proteins
interact with the pto kinase and activate plant immunity. Cell 109: 589-598. Kimbrough, T.G., and Miller, S.I. (2000) Contribution of Salmonella typhimurium type III
secretion components to needle complex formation. Proc. Natl. Acad. Sci. USA. 97, 11008-11013.
Kjemtrup, S., Nimchuk, Z., and Dangl, J.L. (2000) Effector proteins of phytopathogenic
bacteria: bifunctional signals in virulence and host recognition. Curr. Opin. Microbiol. 1, 73-78.
Klement, Z. (1963) Rapid Detection of the Pathogenicity of Phytopathogenic Pseudomonads.
Nature, 199, 299-300. Koebnik, R., Kruger, A., Thieme, F., Urban, A., and Bonas, U. (2006) Specific binding of the
Xanthomonas campestris pv. vesicatoria AraC-type transcriptional activator HrpX to plant-inducible promoter boxes. J. Bacteriol. 21, 7652-7660.
Kubori, T., Matsushima, Y., Nakamura, D., Uralil, J., Lara-Tejero, M., Sukhan, A., Galan,
J.E., and Aizawa, S.I. (1998) Supramolecular structure of the Salmonella typhimurium type III protein secretion system. Science, 280, 602-605.
Kuhnert, P., Schlatter, Y., and Frey, J. (2005) Characterization of the type I secretion system
of the RTX toxin ApxII in "Actinobacillus porcitonsillarum". Vet. Microbiol. 107, 225-232.

154
Kvitko, B.H., Ramos, A.R., Morello, J.E., Oh, H.S., and Collmer, A. (2007) Identification of
harpins in Pseudomonas syringae pv. tomato DC3000, which are functionally similar to HrpK1 in promoting translocation of type III secretion system effectors. J. Bacteriol. 22, 8059-8072.
Lahaye, T., and Bonas, U. (2001) Molecular secrets of bacterial type III effector proteins.
Trends Plant Sci. 10, 479-485. Lawley, T.D., Klimke, W.A., Gubbins, M.J., and Frost, L.S. (2003) F factor conjugation is a
true type IV secretion system. FEMS Microbiol. Lett. 1, 1-15. Leach, J.E., and White, F.F. (1996) Bacterial avirulence genes. Annu. Rev. Phytopathol. 34,
153-179. Leach, J.E., Vera Cruz, C.M., Bai, J., and Leung, H. (2001) Pathogen fitness penalty as a
predictor of durability of disease resistance genes. Annu Rev Phytopathol 39: 187-224. Lee, J., Klusener, B., Tsiamis, G., Stevens, C., Neyt, C., Tampakaki, A.P., Panopoulos, N.J.,
Noller, J., Weiler, E.W., Cornelis, G.R., Mansfield, J.W., and Nurnberger, T. (2001) HrpZ (Psph) from the plant pathogen Pseudomonas syringae pv. phaseolicola binds to lipid bilayers and forms an ion-conducting pore in vitro. Proc. Natl. Acad. Sci. USA, 98, 289-294.
Lee, S.W., Han, S.W., Bartley, L.E., and Ronald, P.C. (2006) From the Academy:
Colloquium review. Unique characteristics of Xanthomonas oryzae pv. oryzae AvrXa21 and implications for plant innate immunity. Proc. Natl. Acad. Sci. USA, 103, 18395-18400.
Lee, W., McDonough, M.A., Kotra, L., Li, Z.H., Silvaggi, N.R., Takeda, Y., Kelly, J.A., and
Mobashery, S. (2001) A 1.2-A snapshot of the final step of bacterial cell wall biosynthesis. Proc. Natl. Acad. Sci. USA, 98, 1427-1431.
Li, H., Xu, H., Zhou, Y., Zhang, J., Long, C., Li, S., Chen, S., Zhou, J.M., and Shao, F.
(2007) The phosphothreonine lyase activity of a bacterial type III effector family. Science, 315, 1000-1003.
Lindgren, P.B. (1997) The role of hrp genes during plant-bacterial interactions. Annu. Rev.
Phytopathol. 35, 129-152. Lindgren, P.B., Peet, R.C., and Panopoulos, N.J. (1986) Gene cluster of Pseudomonas
syringae pv. "phaseolicola" controls pathogenicity of bean plants and hypersensitivity of nonhost plants. J. Bacteriol. 2, 512-522.
Liu, A.X., Li, D.C., Dong, H.S., Wang, J.S. (2006) Harpin Xoo induced hypersensitive cell
death: H2O2 accumulation and related gene expression. Acta Phytopathol. Sin. 36, 163-168.

155
Liu, Y., Chatterjee, A., and Chatterjee, A. K. (1994) Nucleotide sequence and expression of a novel pectate lyase gene (pel-3) and a closely linked endopolygalacturonase gene (peh-1) of Erwinia caratovora subsp. carotovora 71. Appl. Environ. Microbiol. 60, 2545-2552.
Lloyd, S.A., Norman, M., Rosqvist, R., and Wolf-Watz, H. (2001) Yersinia YopE is targeted
for type III secretion by N-terminal, not mRNA, signals. Mol Microbiol 39: 520-531. Loake, G., and Grant, M. (2007) Salicylic acid in plant defence--the players and protagonists.
Curr Opin Plant Biol 10: 466-472. Lonetto, M.A., Brown, K.L., Rudd, K.E., and Buttner, M.J. (1994) Analysis of the
Streptomyces coelicolor sigE gene reveals the existence of a subfamily of eubacterial RNApolymerase F factors involved in the regulation of extracytoplasmic functions. Proc. Natl. Acad. Sci. USA, 91, 7573-7577.
Lorenz, C., Kirchner, O., Egler, M., Stuttmann, J., Bonas, U., and Buttner, D. (2008) HpaA
from xanthomonas is a regulator of type III secretion. Mol Microbiol 69: 344-360. Mansfield, J., Jenner, C., Hockenhull, R., Bennett, M.A., and Stewart, R. (1994)
Characterization of avrPphE, a gene for cultivar-specific avirulence from Pseudmonas syringae pv. phaseolicola which is physically linked to hrpY, a new hrp gene identified in the halo-blight bacterium. Mol. Plant-Microbe Interact. 6, 726-739.
Marenda, M., Brito, B., Callard, D., Genin, S., Barberis, P., Boucher, C., and Arlat, M.
(1998) PrhA controls a novel regulatory pathway required for the specific induction of ralstonia solanacearum hrp genes in the presence of plant cells. Mol Microbiol 27: 437-453.
Marutani, M., Taguchi, F., Shimizu, R., Inagaki, Y., Toyoda, K., Shiraishi, T., and
Ichinose, Y. (2004) Flagellin from Pseudomonas syringae pv. tabaci induced hrp-independent HR in tomato. J. Gen. Plant Pathol. 71, 289-295.
Meindl, T., Boller, T., and Felix, G. (2000) The bacterial elicitor flagellin activates its receptor
in tomato cells according to the address-message concept. Plant Cell, 9, 1783-1794. Melotto, M., Underwood, W., Koczan, J., Nomura, K., and He, S.Y. (2006) Plant stomata
function in innate immunity against bacterial invasion. Cell, 5, 969-980. Metz, M., Dahlbeck, D., Morales, C.Q., Al Sady, B., Clark, E.T., and Staskawicz, B.J.
(2005) The conserved Xanthomonas campestris pv. vesicatoria effector protein XopX is a virulence factor and suppresses host defense in Nicotiana benthamiana. Plant J. 6, 801-814.
Minsavage, G.V., Mudgett, M.B., Stall, R.E., and Jones, J.B. (2004) Importance of opgHXcv
of Xanthomonas campestris pv. vesicatoria in host-parasite interactions. Mol. Plant-Microbe Interact. 2, 152-161.

156
Moissiard, G., and Voinnet, O. (2004) Viral suppression of RNA silencing in plants. Mol. Plant Pathol. 5, 71-82.
Morett, E., and Segovia, L. (1993) The sigma 54 bacterial enhancer-binding protein family:
mechanism of action and phylogenetic relationship of their functional domains. J. Bacteriol. 19, 6067-6074.
Mucyn, T.S., Clemente, A., Andriotis, V.M., Balmuth, A.L., Oldroyd, G.E., Staskawicz,
B.J., and Rathjen, J.P. (2006) The tomato NBARC-LRR protein Prf interacts with Pto kinase in vivo to regulate specific plant immunity. Plant Cell, 10, 2792-2806.
Mudgett, M.B. (2005) New insights to the function of phytopathogenic bacterial type III
effectors in plants. Annu. Rev. Plant Biol. 56, 509-531. Mudgett, M.B., Chesnokova, O., Dahlbeck, D., Clark, E.T., Rossier, O., Bonas, U., and
Staskawicz, B.J. (2000) Molecular signals required for type III secretion and translocation of the Xanthomonas campestris AvrBs2 protein to pepper plants. Proc. Natl. Acad. Sci. USA, 97, 13324-13329.
Murashige, T., and Skoog, F. (1962) A revised medium for rapid growth and bioassays with
tobacco tissue cultures. Physiol. Plant, 15, 473-497. Noel, L., Thieme, F., Gabler, J., Buttner, D., and Bonas, U. (2003) XopC and XopJ, two
novel type III effector proteins from Xanthomonas campestris pv. vesicatoria. J. Bacteriol. 24, 7092-7102.
Nurnberger, T., and Lipka, V. (2005) Non-host resistance in plants: New insights into an old
phenomenon. Mol Plant Pathol 6: 335-345. Oh, C.S., Kim, J.F., and Beer, S.V. (2005) The Hrp pathogenicity island of Erwinia amylovora
and identification of three novel genes required for systemic infection. Mol. Plant Pathol. 2, 125-135.
Oku, T., Alvarez, A.M., and Kado, C.I. (1995) Conservation of the hypersensitivity-
pathogenicity regulatory gene hrpX of xanthomonas campestris and X. oryzae. DNA Seq 5: 245-249.
Orth, K. (2002) Function of the Yersinia effector YopJ. Curr. Opin. Microbiol. 1, 38-43. Park, D.S., Hyun, J.W., Park, Y.J., Kim, J.S., Kang, H.W., Hahn, J.H., and Go, S.J. (2006)
Sensitive and specific detection of xanthomonas axonopodis pv. citri by PCR using pathovar specific primers based on hrpW gene sequences. Microbiol Res 161: 145-149.
Parkinson, N., Aritua, V., Heeney, J., Cowie, C., Bew, J., and Stead, D. (2007) Phylogenetic
analysis of xanthomonas species by comparison of partial gyrase B gene sequences. Int J Syst Evol Microbiol 57: 2881-2887.

157
Peng, J., Bao, Z., Ren, H., Wang, J., Dong, H. (2004) Expression of harpinXoo in transgenic tobacco induces pathogen defense in the absence of hypersensitive response. Phytopathology, 94, 1048-1055.
Perino, C., Gaudriault, S., Vian, B., and Barny, M.A. (1999) Visualization of harpin secretion
in planta during infection of apple seedlings by erwinia amylovora. Cell Microbiol 1: 131-141.
Petnicki-Ocwieja, T., Schneider, D.J., Tam, V.C., Chancey, S.T., Shan, L., and Jamir, Y.
(2002) Genomewide identification of proteins secreted by the hrp type III protein secretion system of pseudomonas syringae pv. tomato DC3000. Proc Natl Acad Sci U S A. 99: 7652-7657.
Posada, D., and Crandall, K.A. (1998) MODELTEST: Testing the model of DNA substitution.
Bioinformatics 14: 817-818. Preston, G., Huang, H.C., He, S.Y., and Collmer, A. (1995) The HrpZ proteins of
Pseudomonas syringae pvs. syringae, glycinea, and tomato are encoded by an operon containing Yersinia ysc homologs and elicit the hypersensitive response in tomato but not soybean. Mol. Plant-Microbe Interact. 8, 717-732.
Pugsley, A.P. (1993) The complete general secretory pathway in gram-negative bacteria.
Microbiol. Rev. 1, 50-108. Pugsley, A.P., Francetic, O., Possot, O.M., Sauvonnet, N., and Hardie, K.R. (1997) Recent
progress and future directions in studies of the main terminal branch of the general secretory pathway in Gram-negative bacteria--a review. Gene, 1, 13-19.
Ritter, C., and Dangl, J.L. (1995) The avrRpm1 gene of Pseudomonas syringae pv. maculicola
is required for virulence on Arabidopsis. Mol. Plant-Microbe Interact. 3, 444-453. Roden, J.A., Belt, B., Ross, J.B., Tachibana, T., Vargas, J., and Mudgett, M.B. (2004) A
genetic screen to isolate type III effectors translocated into pepper cells during Xanthomonas infection. Proc. Natl. Acad. Sci. USA, 101, 16624-16629.
Roden, J., Eardley, L., Hotson, A., Cao, Y., and Mudgett, M.B. (2004) Characterization of
the Xanthomonas AvrXv4 effector, a SUMO protease translocated into plant cells. Mol. Plant-Microbe Interact. 6, 633-643.
Roine, E., Wei, W., Yuan, J., Nurmiaho-Lassila, E.L., Kalkkinen, N., Romantschuk, M.,
and He, S.Y. (1997) Hrp pilus: an hrp-dependent bacterial surface appendage produced by Pseudomonas syringae pv. tomato DC3000. Proc. Natl. Acad. Sci. USA, 94, 3459-3464.
Rosebrock, T.R., Zeng, L., Brady, J.J., Abramovitch, R.B., Xiao, F., and Martin, G.B.
(2007) A bacterial E3 ubiquitin ligase targets a host protein kinase to disrupt plant immunity. Nature 448: 370-374.

158
Rosqvist, R., Magnusson, K.E., and Wolf-Watz, H. (1994) Target cell contact triggers expression and polarized transfer of Yersinia YopE cytotoxin into mammalian cells. EMBO J. 4, 964-972.
Rossier, O., Van den Ackerveken, G., and Bonas, U. (2000) HrpB2 and HrpF from Xanthomonas are type III-secreted proteins and essential for pathogenicity and recognition by the host plant. Mol. Microbiol. 4, 828-838.
Rybak, M., Minsavage, G.V., Stall, R.E., and Jones, J.B. (2009) Identification of
Xanthomonas citri ssp. citri host specificity genes in a heterologous expression host. Mol. Plant-Microbe Interact. 2, 249-262.
Salmond, G.P., and Reeves, P.J. (1993) Membrane traffic wardens and protein secretion in
gram-negative bacteria. Trends Biochem. Sci. 1, 7-12. Sambrook, J., Fristsch, E., and Maniatis, T. (1989) Molecular cloning: a laboratory manual.
Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press. Sandkvist, M. (2001) Type II secretion and pathogenesis. Infect. Immun. 6, 3523-3535. Schubert, T.S., Rizvi, S.A., Sun, X., Gottwald, J.H., Graham, J.H., and Dixon, W.N. (2001)
Meeting the challenge of eradicating citrus canker in Florida-again. Plant Dis. 85, 340-356. Schulte, R., and Bonas, U. (1992) Expression of the Xanthomonas campestris pv. vesicatoria
hrp gene cluster, which determines pathogenicity and hypersensitivity on pepper and tomato, is plant inducible. J. Bacteriol. 3, 815-823.
Scofield, S.R., Tobias, C.M., Rathjen, J.P., Chang, J.H., Lavelle, D.T., Michelmore, R.W.,
and Staskawicz, B.J. (1996) Molecular Basis of Gene-for-Gene Specificity in Bacterial Speck Disease of Tomato. Science, 274, 2063-2065.
Shao, F., Golstein, C., Ade J., Stoutemyer, M., Dixon, J.E., and Innes, R.W. (2003) Cleavage
of arabidopsis PBS1 by a bacterial type III effector. Science 301: 1230-1233. Shrestha, R., Tsuchiya, K., Baek, S.J., Bae, H.N., Hwang, J. H., and and Lim, C.K. (2004)
Identification of dspEF, hrpW, and hrpN loci and characterization of the hrpNep gene in erwinia pyrifoliae. Journal of General Plant Pathology 71: 211-220.
Shevchik, V.E., and Condemine, G. (1998) Functional characterization of the Erwinia
chrysanthemi OutS protein, an element of a type II secretion system. Microbiology, 11, 3219-3228.
Shevchik, V.E., Robert-Baudouy, J., and Hugouvieux-Cotte-Pattat, N. (1997) Pectate lyase
PelI of Erwinia chrysanthemi 3937 belongs to a new family. J. Bacteriol. 23, 7321-7330.

159
Shiraishi, T., Yamada, T., Ichinose, Y., Kiba, A., and Toyoda, K. (1997) The role of suppressors in determining host–parasite specificities in plant cells. Int. Rev. Cytol. 172, 55-93.
Sneath, B.J., Howson, J.M., and Beer, S.V. (1990) A pathogenicity gene from Erwinia
amylovora encodes a predicted protein product homologous a family of procaryotic response regulators. Phytopathology, 80, 1032.
Song, W.Y., Wang, G.L., Chen, L.L., Kim, H.S., Pi, L.Y., Holsten, T., Gardner, J., Wang,
B., Zhai, W.X., Zhu, L.H., Fauquet, C., and Ronald, P. (1995) A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science, 270, 1804-1806.
Sory, M.P., Boland, A., Lambermont, I., and Cornelis, G.R. (1995) Identification of the
YopE and YopH domains required for secretion and internalization into the cytosol of macrophages, using the cyaA gene fusion approach. Proc Natl Acad Sci U S A. 92: 11998-12002.
Stall, R.E., and Civerolo, E.L. (1991) Research relating to the recent outbreak of citrus canker
in Florida. Annu. Rev. Phytopathol. 29, 399-420. Staskawicz, B.J., Ausubel, F.M., Baker, B.J., Ellis, J.G., and Jones, J.D. (1995) Molecular
genetics of plant disease resistance. Science 268: 661-667. Staskawicz, B., Dahlbeck, D., Keen, N., and Napoli, C. (1987) Molecular characterization of
cloned avirulence genes from race 0 and race 1 of Pseudomonas syringae pv. glycinea. J. Bacteriol. 12, 5789-5794.
Stavrinides, J., McCann, H.C., and Guttman, D.S. (2008) Host-pathogen interplay and the
evolution of bacterial effectors. Cell Microbiol. 2, 285-292. Sugio, A., Yang, B., and White, F.F. (2005) Characterization of the hrpF pathogenicity
peninsula of Xanthomonas oryzae pv. oryzae. Mol. Plant-Microbe Interact. 6, 546-554. Sugio, A., Yang, B., Zhu, T., and White, F.F. (2007) Two type III effector genes of
xanthomonas oryzae pv. oryzae control the induction of the host genes OsTFIIAgamma1 and OsTFX1 during bacterial blight of rice. Proc Natl Acad Sci U S A 104: 10720-10725.
Sun, Q., Wu, W., Qian, W., Hu, J., Fang, R., and He, C. (2003) High-quality mutant libraries
of Xanthomonas oryzae pv. oryzae and X. campestris pv. campestris generated by an efficient transposon mutagenesis system. FEMS Microbiol. Lett. 1, 145-150.
Swarup, S., De Feyter, R., Brlansky, R.H., and Gabriel, D.W. (1991) A pathogenicity locus
from xanthomonas citri enables strains from several pathovars of X. campestris to elicit cankerlike lesions on citrus. Phytopathology 81: 802-809.

160
Swarup, S., Yang,Y., Kingsley, M.T., and Gabriel, D.W. (1992) An Xanthomonas citri pathogenicity gene, pthA, pleiotropically encodes gratuitous avirulence on nonhosts. Mol. Plant-Microbe Interact. 3, 204-213.
Swords, K.M., Dahlbeck, D., Kearney, B., Roy, M., and Staskawicz, B.J. (1996) Spontaneous
and induced mutations in a single open reading frame alter both virulence and avirulence in Xanthomonas campestris pv. vesicatoria avrBs2. J. Bacteriol. 15, 4661-4669.
Tampakaki, A. P., and Panopoulos, N. (2000) Elicitation of hypersensitive cell death by
extracellularly target HrpZpsph produced in planta. Mol Plant Microbe Interact 13: 1366-1374. Tang, X., Frederick, R.D., Zhou, J., Halterman, D.A., Jia, Y., and Martin, G.B. (1996)
Initiation of Plant Disease Resistance by Physical Interaction of AvrPto and Pto Kinase. Science, 274, 2060-2063.
Tang, X., Xiao, Y., and Zhou, J.M. (2006) Regulation of the type III secretion system in
phytopathogenic bacteria. Mol. Plant-Microbe Interact. 11, 1159-1166. Thieme, F., Szczesny, R., Urban, A., Kirchner, O., Hause, G., and Bonas, U. (2007) New
type III effectors from Xanthomonas campestris pv. vesicatoria trigger plant reactions dependent on a conserved N-myristoylation motif. Mol. Plant-Microbe Interact. 10, 1250-1261.
Tsuge, S., Terashima, S., Furutani, A., Ochiai, H., Oku, T., and Tsuno, K. (2005) Effects on
promoter activity of base substitutions in the cis-acting regulatory element of HrpXo regulons in xanthomonas oryzae pv. oryzae. J Bacteriol 187: 2308-2314.
Tucker, M.L., Whitelaw, C.A., Lyssenko, N.N., and Nath, P. (2002) Functional analysis of
regulatory elements in the gene promoter for an abscission-specific cellulase from bean and isolation, expression, and binding affinity of three TGA-type basic leucine zipper transcription factors. Plant Physiol. 3, 1487-1496.
Van Larebeke, N., Engler, G., Holsters, M., Van den Elsacker, S., Zaenen, I., Schilperoort,
R.A., and Schell, J. (1974) Large plasmid in Agrobacterium tumefaciens essential for crown gall-inducing ability. Nature, 252, 169-170.
Voulhoux, R., Ball, G., Ize, B., Vasil, M.L., Lazdunski, A., Wu, L.F., and Filloux, A. (2001)
Involvement of the twin-arginine translocation system in protein secretion via the type II pathway. EMBO J. 23, 6735-6741.
Wandersman, C. (1992) Secretion across the bacterial outer membrane. Trends Genet. 8, 317-
322. Wang, X., Li, M., Zhang, J., Zhang, Y., Zhang, G., Wang, J. (2007) Identification of a key
functional region in harpins from Xanthomonas that suppresses protein aggregation and mediates harpin expression in E. coli. Mol. Biol. Rep. 34, 189-198.

161
Wang, X.Y., Song, C.F., Miao, W.G., Ji, Z.L., Wang, X., Zhang, Y., Zhang, J.H., Hu, J.S., Borth, W., and Wang, J.S. (2008) Mutations in the N-terminal coding region of the harpin protein Hpa1 from Xanthomonas oryzae cause loss of hypersensitive reaction induction in tobacco. Appl. Microbiol. Biotechnol. 81, 359-369.
Weber, E., Ojanen-Reuhs, T., Huguet, E., Hause, G., Romantschuk, M., and Korhonen,
T.K. (2005) The type III-dependent hrp pilus is required for productive interaction of xanthomonas campestris pv. vesicatoria with pepper host plants. J Bacteriol 187: 2458-2468.
Wei, C.F., Kvitko, B.H., Shimizu, R., Crabill, E., Alfano, J.R., Lin, N.C., Martin, G.B.,
Huang, H.C., and Collmer, A. (2007) A Pseudomonas syringae pv. tomato DC3000 mutant lacking the type III effector HopQ1-1 is able to cause disease in the model plant Nicotiana benthamiana. Plant J. 1, 32-46.
Wei, Z.M., and Beer, S.V. (1993) HrpI of Erwinia amylovora functions in secretion of harpin
and is a member of a new protein family. J. Bacteriol. 24, 7958-7967. Wei, Z.M., and Beer, S.V. (1995) hrpL activates Erwinia amylovora hrp gene transcription and
is a member of the ECF subfamily of sigma factors. J. Bacteriol. 21, 6201-6210. Wei, Z.M., Laby, R.J., Zumoff, C.H., Bauer, D.W., He, S.Y., Collmer, A., and Beer, S.V.
(1992) Harpin, elicitor of the hypersensitive response produced by the plant pathogen Erwinia amylovora. Science, 257, 85-88.
Wei, Z.M., Sneath, B.J., and Beer, S.V. (1992) Expression of Erwinia amylovora hrp genes in
response to environmental stimuli. J. Bacteriol. 6, 1875-1882. Wengelnik, K., and Bonas, U. (1996) HrpXv, an AraC-type regulator, activates expression of
five of the six loci in the hrp cluster of Xanthomonas campestris pv. vesicatoria. J. Bacteriol. 12, 3462-3469.
Wengelnik, K., Marie, C., Russel, M., and Bonas, U. (1996a) Expression and localization of
HrpA1, a protein of Xanthomonas campestris pv. vesicatoria essential for pathogenicity and induction of the hypersensitive reaction. J. Bacteriol. 4, 1061-1069.
Wengelnik, K., Rossier, O., and Bonas, U. (1999) Mutations in the regulatory gene hrpG of
xanthomonas campestris pv. vesicatoria result in constitutive expression of all hrp genes. J Bacteriol 181: 6828-6831.
Wengelnik, K., Van den Ackerveken, G., and Bonas, U. (1996b) HrpG, a key hrp regulatory
protein of Xanthomonas campestris pv. vesicatoria is homologous to two-component response regulators. Mol. Plant-Microbe Interact. 8, 704-712.
Wolpert, T.J., Dunkle, L.D., and Ciuffetti, L.M. (2002) Host-selective toxins and avirulence
determinants: what's in a name? Annu. Rev. Phytopathol. 40, 251-285.

162
Wu, A.J., Andriotis, V.M., Durrant, M.C., and Rathjen, J.P. (2004) A patch of surface-exposed residues mediates negative regulation of immune signaling by tomato Pto kinase. Plant Cell, 10, 2809-2821.
Xiao, Y., and Hutcheson, S.W. (1994) A single promoter sequence recognized by a newly
identified alternate sigma factor directs expression of pathogenicity and host range determinants in Pseudomonas syringae. J. Bacteriol. 10, 3089-3091.
Xiao, Y., Heu, S., Yi, J., Lu, Y., and Hutcheson, S.W. (1994) Identification of a putative
alternate sigma factor and characterization of a multicomponent regulatory cascade controlling the expression of Pseudomonas syringae pv. syringae Pss61 hrp and hrmA genes. J. Bacteriol. 4, 1025-1036.
Xiao, Y., Lu, Y., Heu, S., and Hutcheson, S.W. (1992) Organization and environmental
regulation of the pseudomonas syringae pv. syringae 61 hrp cluster. J Bacteriol 174: 1734-1741.
Yamazaki, A., Hirata, H., and Tsuyumu, S. (2008) HprG regulates type II secretory proteins in
Xanthomonas axonopodis pv. citri. J. Gen. Plant Pathol. 74, 138-150. Yahr, T. L. (2006) A Critical New Pathway for Toxin Secretion? New England Journal of
Medicine. 355, 1171-1172. Young, J., and Holland, I.B. (1999) ABC transporters: bacterial exporters-revisited five years
on. Biochim. Biophys. Acta, 2, 177-200. Zhu, W., MaGbanua, M.M., and White, F.F. (2000) Identification of two novel hrp-associated
genes in the hrp gene cluster of Xanthomonas oryzae pv. oryzae. J. Bacteriol. 7, 1844-1853. Zou, L., Wang, X., Xiang, Y., Zhang, B., Li, Y., Xiao, Y., Wang, J., Walmsley, A.R., Chen,
G. (2006) Elucidation of the hrp clusters of Xanthomonas oryzae pv. oryzicola that control the hypersensitive response in non-host tobacco and pathogenicity in susceptible host rice. Appl. Environ. Microbiol. 72, 6212-6224.

163
BIOGRAPHICAL SKETCH
José Francisco Lissoni Figueiredo was born in the city of Cosmorama, Brazil. During his
undergraduate studies in biomedicine at the University of Barão de Mauá, he started a trainee
program in the sequencing genome project of the Xanthomonas citri subsp. citri and
Xanthomonas campestris pv. campestris. In 2001, he started the Master of Science program at
University of State of São Paulo in the genetics and plant improvement program. After
graduating with his master’s, he was invited by Dr. Frank F. White to work with Xanthomonas
campestris pv. campestris in the Plant Pathology Department at Kansas State University. In
2004, he was admitted in the Plant Pathology Department Ph.D. program at Kansas State
University and transferred to the Plant Pathology Department at University of Florida where he
received his Ph.D. in 2009 conducting a research project on screen pathogenic factors involved
in Xanthomonas citri subsp. citri disease progress.





























