Embed Size (px)
Citation preview

General mechanism for modulating immunoglobulineffector functionPeter Sondermanna,1, Andrew Pinceticb,1, Jad Maamaryb, Katja Lammensc, and Jeffrey V. Ravetchb,2
aSuppreMol GmbH, 82152 Martinsried, Germany; bLaboratory of Molecular Genetics and Immunology, The Rockefeller University, New York, NY 10021;and cLudwig-Maximilians-Universität Gene Center, 81377 Munich, Germany
Contributed by Jeffrey V. Ravetch, April 26, 2013 (sent for review April 5, 2013)
Immunoglobulins recognize and clear microbial pathogens andtoxins through the coupling of variable region specificity toFc-triggered cellular activation. These proinflammatory activities areregulated, thus avoiding the pathogenic sequelae of uncontrolledinflammation bymodulating the composition of the Fc-linked glycan.Upon sialylation, the affinities for Fcγ receptors are reduced, whereasthose for alternative cellular receptors, such as dendritic cell-specific intercellular adhesion molecule-3-grabbing nonintegrin(DC-SIGN)/CD23, are increased. We demonstrate that sialylationinduces significant structural alterations in the Cγ2 domain andpropose a model that explains the observed changes in ligandspecificity and biological activity. By analogy to related complexesformed by IgE and its evolutionarily related Fc receptors, weconclude that this mechanism is general for the modulation ofantibody-triggered immune responses, characterized by a shiftbetween an “open” activating conformation and a “closed” anti-inflammatory state of antibody Fc fragments. This common mech-anism has been targeted by pathogens to avoid host defense andoffers targets for therapeutic intervention in allergic and auto-immune disorders.
conformational change | sialylated IgG Fc
IgG and IgE mediate their proinflammatory properties throughthe crosslinking of the 1:1 complex of the Fc receptor (FcR)
monomer in the Fc dimer cleft (1, 2). By contrast, both IgG andIgE can engage a second class of receptors, the evolutionarilyrelated, C-type lectins dendritic cell-specific intercellular adhe-sion molecule-3-grabbing nonintegrin (DC-SIGN) (3) and CD23(4), respectively, resulting in anti-inflammatory and immuno-suppressive responses (5, 6). The structural basis for the abilityof IgE to interact either with one or the other of these twodisparate classes of receptors has recently been defined (7). Theintrinsic flexibility of the IgE Ce3 domain results in both openand closed conformations of the IgE Fc, resulting in the bindingof either FceRI or CD23, respectively. Binding of either receptorinduces an allosteric change in the IgE Fc to the alternativeconformation, thus precluding the interaction with the otherreceptor (7). Binding of IgE to the type II, C-type lectin CD23is neither carbohydrate- nor calcium-dependent, mediated exclu-sively through protein–protein interactions, generating a 2:1complex of CD23 with the Ce3–Ce4 interface (7). DC-SIGN isa structurally homologous, calcium-dependent, carbohydrate-binding, type II lectin, tightly linked to CD23 on chromosome19 (8), displaying ligand specificity for mannose-containingglycoconjugates and fucose-containing Lewis antigens. Bindingof DC-SIGN to IgG requires that the complex, biantennaryglycan, attached to the evolutionarily conserved glycosylationsite Asn-297 and enclosed within the cavity formed by the Cγ2domains of the A and B chains of the Fc dimer, be processed tothe α2,6 sialylated form (9, 10). Importantly, no evidence has beenfound for DC-SIGN binding to sialylated glycans or glyco-conjugates (11), suggesting that the binding interaction betweensialylated Fc and DC-SIGN may not involve the canonical glycaninteractions previously defined for this lectin and bind to sialylatedFc in a manner analogous to CD23 binding to IgE.
Results and DiscussionBecause the Cγ2 domain of IgG lacks the intrinsic flexibility ofCe3 (12, 13), we hypothesized that sialylation may induce thisflexibility, allowing it to engage DC-SIGN and limiting its bind-ing to FcγRs. We therefore investigated the effect of sialylationon the Fc structure by circular dichroism, thermal and chemicaldenaturation, and anilinonaphthalene sulfonates (ANS) bind-ing (Fig. 1). The CD spectrum of neuraminidase-treated andtherefore asialylated human IgG1 Fc (NAse Fc, G2F glycoform)yields the classical spectral pattern associated with β-sheetstructure with a minimum at 216 nm and a peak near 203 nm.Deglycosylation of the Fc induces dramatic shifts in the CDspectrum consistent with the structural changes observed incrystal structures of aglycosyl Fc (14, 15). Upon sialylation(A2F form), however, we observe a small shift in the spectra forsFc (Fig. 1A). Sialylation may alter the β-sheet content in the Fcbecause this spectral shift includes a ∼14% decrease in the el-lipticity (Δe) value at 216 nm compared with NAse Fc. Degly-cosylation of the Fc abrogated the CD differences between thesetwo glycoforms (Fig. S1), indicating that the observed spectraldifferences were the result of a change in the Fc structure in-duced by sialylation.We next measured the thermal stability of these Fc glycoforms
to evaluate their thermodynamic properties. Ellipticity values(Δe) for Fcs were recorded at 206.5 nm as a function of tem-perature (16). As shown in Fig. 1B, we observe a transition phaseas the Cγ2 domain denatures above 65 °C, as described pre-viously (17). Sialylated Fc has a lower melting temperature (TM)than asialylated Fc, differing by 0.9 °C. Fully deglycosylated Fc isfurther reduced in its TM by 3.1 °C, as previously reported (17).A second transition phase occurs above 80 °C, which correspondsto the denaturation of the Cγ3 domain (17, 18). The decrease inTM
Cγ2 correlates with the CD spectral shifts illustrated in Fig.1A. This suggests that structural changes induced by sialylationoccur in the Cγ2 domain. As with TM
Cγ2, the change in van’tHoff enthalpy, ΔHV°, and the free energy of folding, ΔG°, alsodecreased with sialylation and deglycosylation relative to NAseFc (Fig. S2), which is indicative of decreased protein stability(Fig. S2).GnHCl-induced denaturation (Fig. 1C and Fig. S3) also
revealed a decrease in ΔGH2O° values for sFc relative to NAse Fc(−6.4 kcal/mol and −7.125 kcal/mol, respectively). However, incontrast to thermal denaturation, GnHCl-induced denaturationrevealed similar ΔGH2O° values between deglycosylated andNAse Fc −7.27 and −7.125 kcal/mol, respectively. M values
Author contributions: P.S., A.P., J.M., and J.V.R. designed research; A.P., J.M., and J.V.R.designed the experiments and interpreted the results; P.S. and K.L. performed molecularmodeling; A.P. and J.M. performed the experiments; P.S., A.P., J.M., and K.L. performedresearch; P.S., A.P., J.M., K.L., and J.V.R. analyzed data; and P.S., A.P., and J.V.R. wrotethe paper.
The authors declare no conflict of interest.1P.S. and A.P. contributed equally to this work.2To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1307864110/-/DCSupplemental.
9868–9872 | PNAS | June 11, 2013 | vol. 110 | no. 24 www.pnas.org/cgi/doi/10.1073/pnas.1307864110
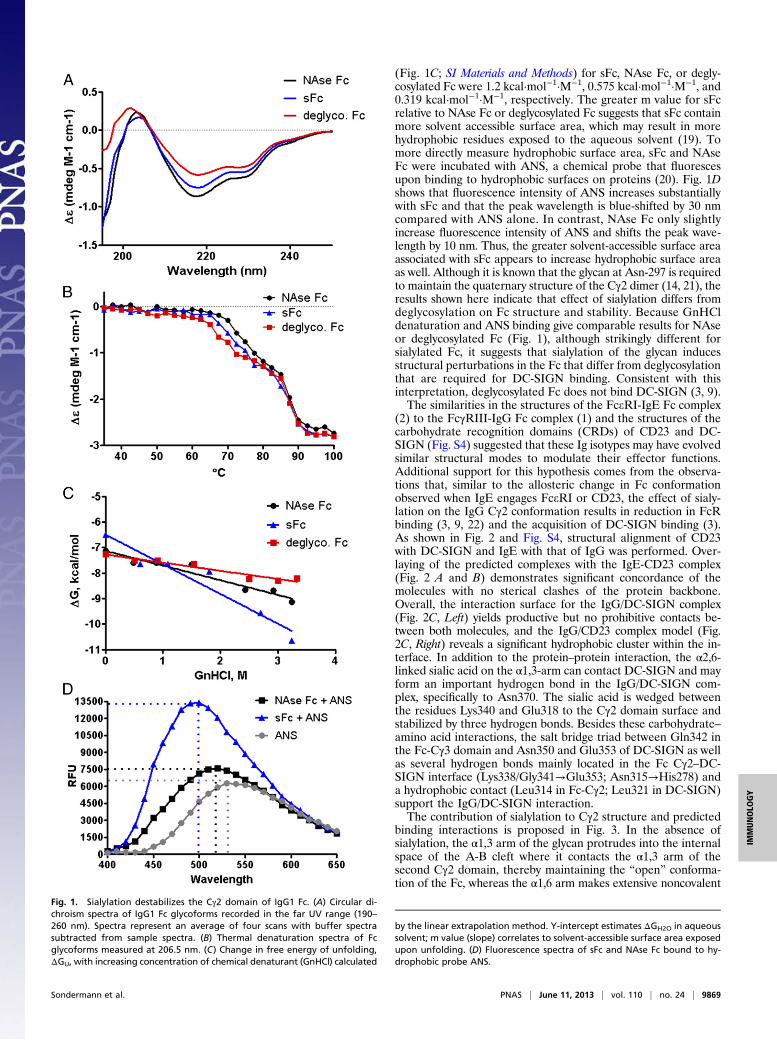
(Fig. 1C; SI Materials and Methods) for sFc, NAse Fc, or degly-cosylated Fc were 1.2 kcal·mol−1·M−1, 0.575 kcal·mol−1·M−1, and0.319 kcal·mol−1·M−1, respectively. The greater m value for sFcrelative to NAse Fc or deglycosylated Fc suggests that sFc containmore solvent accessible surface area, which may result in morehydrophobic residues exposed to the aqueous solvent (19). Tomore directly measure hydrophobic surface area, sFc and NAseFc were incubated with ANS, a chemical probe that fluorescesupon binding to hydrophobic surfaces on proteins (20). Fig. 1Dshows that fluorescence intensity of ANS increases substantiallywith sFc and that the peak wavelength is blue-shifted by 30 nmcompared with ANS alone. In contrast, NAse Fc only slightlyincrease fluorescence intensity of ANS and shifts the peak wave-length by 10 nm. Thus, the greater solvent-accessible surface areaassociated with sFc appears to increase hydrophobic surface areaas well. Although it is known that the glycan at Asn-297 is requiredto maintain the quaternary structure of the Cγ2 dimer (14, 21), theresults shown here indicate that effect of sialylation differs fromdeglycosylation on Fc structure and stability. Because GnHCldenaturation and ANS binding give comparable results for NAseor deglycosylated Fc (Fig. 1), although strikingly different forsialylated Fc, it suggests that sialylation of the glycan inducesstructural perturbations in the Fc that differ from deglycosylationthat are required for DC-SIGN binding. Consistent with thisinterpretation, deglycosylated Fc does not bind DC-SIGN (3, 9).The similarities in the structures of the FceRI-IgE Fc complex
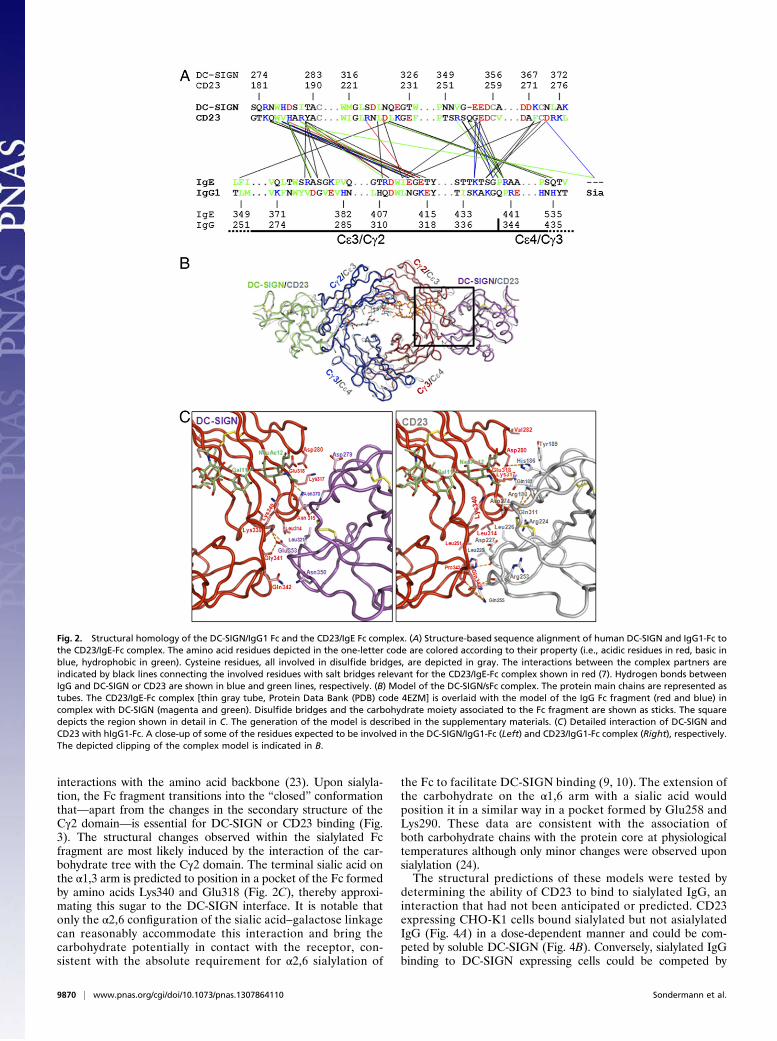
(2) to the FcγRIII-IgG Fc complex (1) and the structures of thecarbohydrate recognition domains (CRDs) of CD23 and DC-SIGN (Fig. S4) suggested that these Ig isotypes may have evolvedsimilar structural modes to modulate their effector functions.Additional support for this hypothesis comes from the observa-tions that, similar to the allosteric change in Fc conformationobserved when IgE engages FceRI or CD23, the effect of sialy-lation on the IgG Cγ2 conformation results in reduction in FcRbinding (3, 9, 22) and the acquisition of DC-SIGN binding (3).As shown in Fig. 2 and Fig. S4, structural alignment of CD23with DC-SIGN and IgE with that of IgG was performed. Over-laying of the predicted complexes with the IgE-CD23 complex(Fig. 2 A and B) demonstrates significant concordance of themolecules with no sterical clashes of the protein backbone.Overall, the interaction surface for the IgG/DC-SIGN complex(Fig. 2C, Left) yields productive but no prohibitive contacts be-tween both molecules, and the IgG/CD23 complex model (Fig.2C, Right) reveals a significant hydrophobic cluster within the in-terface. In addition to the protein–protein interaction, the α2,6-linked sialic acid on the α1,3-arm can contact DC-SIGN and mayform an important hydrogen bond in the IgG/DC-SIGN com-plex, specifically to Asn370. The sialic acid is wedged betweenthe residues Lys340 and Glu318 to the Cγ2 domain surface andstabilized by three hydrogen bonds. Besides these carbohydrate–amino acid interactions, the salt bridge triad between Gln342 inthe Fc-Cγ3 domain and Asn350 and Glu353 of DC-SIGN as wellas several hydrogen bonds mainly located in the Fc Cγ2–DC-SIGN interface (Lys338/Gly341→Glu353; Asn315→His278) anda hydrophobic contact (Leu314 in Fc-Cγ2; Leu321 in DC-SIGN)support the IgG/DC-SIGN interaction.The contribution of sialylation to Cγ2 structure and predicted
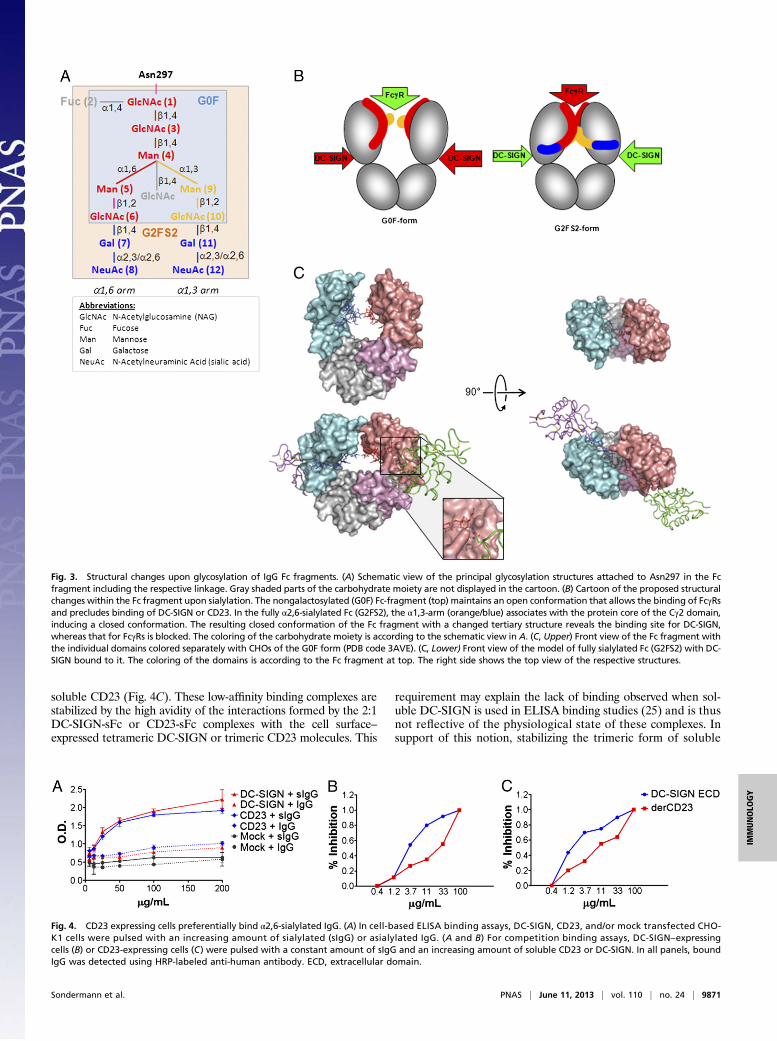
binding interactions is proposed in Fig. 3. In the absence ofsialylation, the α1,3 arm of the glycan protrudes into the internalspace of the A-B cleft where it contacts the α1,3 arm of thesecond Cγ2 domain, thereby maintaining the “open” conforma-tion of the Fc, whereas the α1,6 arm makes extensive noncovalent
Fig. 1. Sialylation destabilizes the Cγ2 domain of IgG1 Fc. (A) Circular di-chroism spectra of IgG1 Fc glycoforms recorded in the far UV range (190–260 nm). Spectra represent an average of four scans with buffer spectrasubtracted from sample spectra. (B) Thermal denaturation spectra of Fcglycoforms measured at 206.5 nm. (C) Change in free energy of unfolding,ΔGU, with increasing concentration of chemical denaturant (GnHCl) calculated
by the linear extrapolation method. Y-intercept estimates ΔGH2O in aqueoussolvent; m value (slope) correlates to solvent-accessible surface area exposedupon unfolding. (D) Fluorescence spectra of sFc and NAse Fc bound to hy-drophobic probe ANS.
Sondermann et al. PNAS | June 11, 2013 | vol. 110 | no. 24 | 9869
IMMUNOLO
GY
interactions with the amino acid backbone (23). Upon sialyla-tion, the Fc fragment transitions into the “closed” conformationthat—apart from the changes in the secondary structure of theCγ2 domain—is essential for DC-SIGN or CD23 binding (Fig.3). The structural changes observed within the sialylated Fcfragment are most likely induced by the interaction of the car-bohydrate tree with the Cγ2 domain. The terminal sialic acid onthe α1,3 arm is predicted to position in a pocket of the Fc formedby amino acids Lys340 and Glu318 (Fig. 2C), thereby approxi-mating this sugar to the DC-SIGN interface. It is notable thatonly the α2,6 configuration of the sialic acid–galactose linkagecan reasonably accommodate this interaction and bring thecarbohydrate potentially in contact with the receptor, con-sistent with the absolute requirement for α2,6 sialylation of
the Fc to facilitate DC-SIGN binding (9, 10). The extension ofthe carbohydrate on the α1,6 arm with a sialic acid wouldposition it in a similar way in a pocket formed by Glu258 andLys290. These data are consistent with the association ofboth carbohydrate chains with the protein core at physiologicaltemperatures although only minor changes were observed uponsialylation (24).The structural predictions of these models were tested by
determining the ability of CD23 to bind to sialylated IgG, aninteraction that had not been anticipated or predicted. CD23expressing CHO-K1 cells bound sialylated but not asialylatedIgG (Fig. 4A) in a dose-dependent manner and could be com-peted by soluble DC-SIGN (Fig. 4B). Conversely, sialylated IgGbinding to DC-SIGN expressing cells could be competed by
Fig. 2. Structural homology of the DC-SIGN/IgG1 Fc and the CD23/IgE Fc complex. (A) Structure-based sequence alignment of human DC-SIGN and IgG1-Fc tothe CD23/IgE-Fc complex. The amino acid residues depicted in the one-letter code are colored according to their property (i.e., acidic residues in red, basic inblue, hydrophobic in green). Cysteine residues, all involved in disulfide bridges, are depicted in gray. The interactions between the complex partners areindicated by black lines connecting the involved residues with salt bridges relevant for the CD23/IgE-Fc complex shown in red (7). Hydrogen bonds betweenIgG and DC-SIGN or CD23 are shown in blue and green lines, respectively. (B) Model of the DC-SIGN/sFc complex. The protein main chains are represented astubes. The CD23/IgE-Fc complex [thin gray tube, Protein Data Bank (PDB) code 4EZM] is overlaid with the model of the IgG Fc fragment (red and blue) incomplex with DC-SIGN (magenta and green). Disulfide bridges and the carbohydrate moiety associated to the Fc fragment are shown as sticks. The squaredepicts the region shown in detail in C. The generation of the model is described in the supplementary materials. (C) Detailed interaction of DC-SIGN andCD23 with hIgG1-Fc. A close-up of some of the residues expected to be involved in the DC-SIGN/IgG1-Fc (Left) and CD23/IgG1-Fc complex (Right), respectively.The depicted clipping of the complex model is indicated in B.
9870 | www.pnas.org/cgi/doi/10.1073/pnas.1307864110 Sondermann et al.
soluble CD23 (Fig. 4C). These low-affinity binding complexes arestabilized by the high avidity of the interactions formed by the 2:1DC-SIGN-sFc or CD23-sFc complexes with the cell surface–expressed tetrameric DC-SIGN or trimeric CD23 molecules. This
requirement may explain the lack of binding observed when sol-uble DC-SIGN is used in ELISA binding studies (25) and is thusnot reflective of the physiological state of these complexes. Insupport of this notion, stabilizing the trimeric form of soluble
Fig. 3. Structural changes upon glycosylation of IgG Fc fragments. (A) Schematic view of the principal glycosylation structures attached to Asn297 in the Fcfragment including the respective linkage. Gray shaded parts of the carbohydrate moiety are not displayed in the cartoon. (B) Cartoon of the proposed structuralchanges within the Fc fragment upon sialylation. The nongalactosylated (G0F) Fc-fragment (top) maintains an open conformation that allows the binding of FcγRsand precludes binding of DC-SIGN or CD23. In the fully α2,6-sialylated Fc (G2FS2), the α1,3-arm (orange/blue) associates with the protein core of the Cγ2 domain,inducing a closed conformation. The resulting closed conformation of the Fc fragment with a changed tertiary structure reveals the binding site for DC-SIGN,whereas that for FcγRs is blocked. The coloring of the carbohydrate moiety is according to the schematic view in A. (C, Upper) Front view of the Fc fragment withthe individual domains colored separately with CHOs of the G0F form (PDB code 3AVE). (C, Lower) Front view of the model of fully sialylated Fc (G2FS2) with DC-SIGN bound to it. The coloring of the domains is according to the Fc fragment at top. The right side shows the top view of the respective structures.
Fig. 4. CD23 expressing cells preferentially bind α2,6-sialylated IgG. (A) In cell-based ELISA binding assays, DC-SIGN, CD23, and/or mock transfected CHO-K1 cells were pulsed with an increasing amount of sialylated (sIgG) or asialylated IgG. (A and B) For competition binding assays, DC-SIGN–expressingcells (B) or CD23-expressing cells (C) were pulsed with a constant amount of sIgG and an increasing amount of soluble CD23 or DC-SIGN. In all panels, boundIgG was detected using HRP-labeled anti-human antibody. ECD, extracellular domain.
Sondermann et al. PNAS | June 11, 2013 | vol. 110 | no. 24 | 9871
IMMUNOLO
GY

CD23 through a leucine zipper motif linked to the N-terminusof the receptor (lz-CD23) allows for enhanced detection of sia-lylated IgG binding in a solid-phase ELISA format at physio-logical temperature relative to the extracellular region of CD23alone (Fig. S5), as has been reported for IgE (26). However, incontrast to IgE, binding of sialylated IgG to trimeric CD23 is notenhanced at 4 °C.The structural model and data reported here support the
conclusion that a common mechanism for regulating the ef-fector activity of immunoglobulins is accomplished through thealternation of Fc conformations between open and closedstates, thereby regulating Fc binding to FcRs or SIGN/CD23,respectively. Regulation of these conformations may be in-trinsic, as observed for IgE, resulting from the disordered Ce3“molten globular” domain, or extrinsic, the result of modifi-cation of the IgG Asn-297 N-linked glycan. Pathogens haveexploited this common mechanism to avoid host defense byshifting the equilibrium of the Fc conformation to the closedstate, either by modulating the glycan composition (27) or
stabilizing the closed conformation (28). Sialylation of the N-linked glycan of IgG induces a conformation that will also bindCD23 in addition to DC-SIGN, thereby providing a mecha-nism for suppressing follicular B-cell activation by regulatingIgG sialylation to maintain homeostasis through feedback regu-lation by IgG of its synthesis.
Materials and MethodsMonoclonal human IgG1 preparations were used for biophysical character-ization of Fc glycoforms. Sialylated Fc produced in two-step in vitro reactionwith β1,4-GalT and ST6Gal. Binding assays performed in cell-based ELISAformat with CHO-K1 cells transiently expressing full-length DC-SIGN or CD23.Detailed experimental and analytical procedures are presented in SI Mate-rials and Methods.
ACKNOWLEDGMENTS. We thank Frederik Wermeling for his insightfulcomments and suggestions and all the members of the J.V.R. laboratoryfor technical assistance and helpful discussions. This work was supported, inpart, by grants from the National Institutes of Health and with the generoussupport of Eileen Greenland.
1. Sondermann P, Huber R, Oosthuizen V, Jacob U (2000) The 3.2-A crystal structure ofthe human IgG1 Fc fragment-Fc gammaRIII complex. Nature 406(6793):267–273.
2. Garman SC, Wurzburg BA, Tarchevskaya SS, Kinet JP, Jardetzky TS (2000) Structure ofthe Fc fragment of human IgE bound to its high-affinity receptor Fc epsilonRI alpha.Nature 406(6793):259–266.
3. Anthony RM, Wermeling F, Karlsson MC, Ravetch JV (2008) Identification of a re-ceptor required for the anti-inflammatory activity of IVIG. Proc Natl Acad Sci USA105(50):19571–19578.
4. Hibbert RG, et al. (2005) The structure of human CD23 and its interactions with IgEand CD21. J Exp Med 202(6):751–760.
5. Anthony RM, Kobayashi T, Wermeling F, Ravetch JV (2011) Intravenous gammaglo-bulin suppresses inflammation through a novel T(H)2 pathway. Nature 475(7354):110–113.
6. Acharya M, et al. (2010) CD23/FceRII: Molecular multi-tasking. Clin Exp Immunol162(1):12–23.
7. Dhaliwal B, et al. (2012) Crystal structure of IgE bound to its B-cell receptor CD23reveals a mechanism of reciprocal allosteric inhibition with high affinity receptorFceRI. Proc Natl Acad Sci USA 109(31):12686–12691.
8. Soilleux EJ, Barten R, Trowsdale J (2000) DC-SIGN; a related gene, DC-SIGNR; and CD23form a cluster on 19p13. J Immunol 165(6):2937–2942.
9. Kaneko Y, Nimmerjahn F, Ravetch JV (2006) Anti-inflammatory activity of immuno-globulin G resulting from Fc sialylation. Science 313(5787):670–673.
10. Anthony RM, et al. (2008) Recapitulation of IVIG anti-inflammatory activity witha recombinant IgG Fc. Science 320(5874):373–376.
11. Holla A, Skerra A (2011) Comparative analysis reveals selective recognition of glycansby the dendritic cell receptors DC-SIGN and Langerin. Protein Eng Des Sel 24(9):659–669.
12. Price NE, Price NC, Kelly SM, McDonnell JM (2005) The key role of protein flexibility inmodulating IgE interactions. J Biol Chem 280(3):2324–2330.
13. Borthakur S, Andrejeva G, McDonnell JM (2011) Basis of the intrinsic flexibility of theCe3 domain of IgE. Biochemistry 50(21):4608–4614.
14. Borrok MJ, Jung ST, Kang TH, Monzingo AF, Georgiou G (2012) Revisiting the role ofglycosylation in the structure of human IgG Fc. ACS Chem Biol 7(9):1596–1602.
15. Feige MJ, et al. (2009) Structure of the murine unglycosylated IgG1 Fc fragment. J MolBiol 391(3):599–608.
16. Greenfield NJ (2006) Using circular dichroism collected as a function of temperatureto determine the thermodynamics of protein unfolding and binding interactions. NatProtoc 1(6):2527–2535.
17. Mimura Y, et al. (2001) Role of oligosaccharide residues of IgG1-Fc in Fc γ RIIb binding.J Biol Chem 276(49):45539–45547.
18. Tischenko VM, Abramov VM, Zav’yalov VP (1998) Investigation of the cooperativestructure of Fc fragments from myeloma immunoglobulin G. Biochemistry 37(16):5576–5581.
19. Myers JK, Pace CN, Scholtz JM (1995) Denaturant m values and heat capacity changes:Relation to changes in accessible surface areas of protein unfolding. Protein Sci 4(10):2138–2148.
20. Hawe A, Sutter M, Jiskoot W (2008) Extrinsic fluorescent dyes as tools for proteincharacterization. Pharm Res 25(7):1487–1499.
21. Krapp S, Mimura Y, Jefferis R, Huber R, Sondermann P (2003) Structural analysis ofhuman IgG-Fc glycoforms reveals a correlation between glycosylation and structuralintegrity. J Mol Biol 325(5):979–989.
22. Scallon BJ, Tam SH, McCarthy SG, Cai AN, Raju TS (2007) Higher levels of sialylated Fcglycans in immunoglobulin G molecules can adversely impact functionality. MolImmunol 44(7):1524–1534.
23. Wormald MR, et al. (1997) Variations in oligosaccharide-protein interactions in im-munoglobulin G determine the site-specific glycosylation profiles and modulate thedynamic motion of the Fc oligosaccharides. Biochemistry 36(6):1370–1380.
24. Barb AW, et al. (2012) NMR characterization of immunoglobulin G Fc glycan motionon enzymatic sialylation. Biochemistry 51(22):4618–4626.
25. Yu X, Vasiljevic S, Mitchell DA, Crispin M, Scanlan CN (2013) Dissecting the molecularmechanism of IVIg therapy: The interaction between serum IgG and DC-SIGN is in-dependent of antibody glycoform or Fc domain. J Mol Biol 425(8):1253–1258.
26. Chen BH, et al. (2003) Temperature effect on IgE binding to CD23 versus Fc epsilon RI.J Immunol 170(4):1839–1845.
27. Albert H, Collin M, Dudziak D, Ravetch JV, Nimmerjahn F (2008) In vivo enzymaticmodulation of IgG glycosylation inhibits autoimmune disease in an IgG subclass-dependent manner. Proc Natl Acad Sci USA 105(39):15005–15009.
28. Borthakur S, et al. (2012) Mapping of the CD23 binding site on immunoglobulin E(IgE) and allosteric control of the IgE-Fc epsilonRI interaction. J Biol Chem 287(37):31457–31461.
9872 | www.pnas.org/cgi/doi/10.1073/pnas.1307864110 Sondermann et al.





























