Embed Size (px)
Citation preview

Molecular & Biochemical Parasitology 136 (2004) 157–164
Gene synteny and chloroquine resistance inPlasmodium chabaudiPaul Hunta,∗, Axel Martinellia, Richard Fawcetta,Jane Carltonb, Richard Cartera, David Wallikera
a Institute of Cell, Animal and Population Biology, University of Edinburgh, West Mains Road, Edinburgh EH9 3JT, Scotland, UKb The Institute for Genomics Research, 9712 Medical Center Drive, Rockville, MD 20850, USA
Received 29 February 2004; accepted 3 March 2004
Available online 24 April 2004
Abstract
Chloroquine resistance in the rodent malaria parasitePlasmodium chabaudihas been shown to be caused by a gene on chromosome11, and is not linked to orthologues of thePlasmodium falciparumchloroquine resistance transporter (pfcrt) or Pgh-1 (pfmdr1) genes.In the current work, the progeny of crosses between chloroquine-resistant and sensitive clones ofP. chabaudihave been analysed for theinheritance of 658 AFLP markers. Markers linked to the chloroquine responses of the progeny, including two which are completely linked,have been genetically mapped, sequenced and their homologues, or closely linked loci, identified inP. falciparum. The chromosome 11markers most closely linked to chloroquine resistance inP. chabaudimap to loci which are also closely linked inP. falciparum, although intwo linkage groups on chromosomes 6 and 13 of this species. TheP. falciparumorthologue of the gene conferring chloroquine resistancein P. chabaudiis predicted to lie within a 250 kb region ofP. falciparumchromosome 6, containing approximately 50 genes. The geneticorder of the markers inP. chabaudiis co-linear with the physical linkage represented in theP. falciparumgenome database. The findingsprovide evidence for extensive conservation of synteny between the two species.© 2004 Elsevier B.V. All rights reserved.
Keywords: Plasmodium chabaudi; Plasmodium falciparum; Chloroquine; Genetic linkage; Amplified fragment length polymorphism (AFLP); Synteny
1. Introduction
Chloroquine resistance (CQR) in the human malaria par-asitePlasmodium falciparumis influenced by at least twogenes,pfcrtandpfmdr1. Pfcrtwas identified by linkage anal-ysis in a genetic cross between a chloroquine resistant and achloroquine sensitive clone ofP. falciparum[1] and its pri-mary role in resistance has subsequently been confirmed infield isolates[2–6] and in transfection studies[7]. Polymor-phisms inpfmdr1 appear to be involved in the generationof higher level resistance in certain countries, e.g. Sudan[2]. Transfection studies support this conclusion[8]. Despitethese major advances in our understanding of the geneticsof chloroquine resistance, there is general agreement that anumber of other genes may be involved in influencing thelevel of resistance seen in natural infections of this parasite.
Abbreviations:AFLP, amplified fragment length polymorphism; DNA,deoxyribonucleic acid; EDTA, ethylenediamine tetra acetic acid; PBS,phosphate buffered saline; PCR, polymerase chain reaction; RFLP, re-striction fragment length polymorphism; SDS, sodium dodecyl sulphate
∗ Corresponding author. Tel.:+44-131-650-8661;fax: +44-131-650-6564.
E-mail address:[email protected] (P. Hunt).
The rodent malaria parasite,Plasmodium chabaudiisan excellent model system for genetic studies such as theidentification of genes involved in drug resistance[9]. Thisis partly because it is relatively easy to perform geneticcrosses, generating recombinant progeny which can begenotyped and phenotyped. Also drug-resistant mutantscan be generated easily by growing drug-sensitive clones inthe presence of drugs. The effect of mutations upon drugresponses can then be studied in parasites with the samegenetic background. The roles of the orthologues ofpfcrtand pfmdr1 in P. chabaudi(pccg10and pcmdr1, respec-tively) have been investigated by analysing recombinantprogeny clones of a cross between the chloroquine-resistantparasite, AS(3CQ), and the chloroquine-sensitive par-asite, AJ. Neither of these genes (located on chromo-somes 12 and 14, respectively) were found to be linkedto chloroquine sensitivity[10]. Indeed, previous work[11] had identified RFLP markers on chromosome 11which were linked to chloroquine resistance. The recentidentification of numerous AFLP markers which dis-tinguish the parental clones AS(3CQ) and AJ[12] hasnow enabled us to investigate this locus at higher resolu-tion.
0166-6851/$ – see front matter © 2004 Elsevier B.V. All rights reserved.doi:10.1016/j.molbiopara.2004.03.008

158 P. Hunt et al. / Molecular & Biochemical Parasitology 136 (2004) 157–164
Previous studies have demonstrated considerable genesynteny betweenPlasmodiumspecies[13–15], in which thephysical linkage of genes in one species is conserved in an-other. This is of considerable significance when attemptingto identify closely linked genes inP. chabaudi. For instance,depending upon the extent of gene synteny, it is now possibleto use the complete sequence ofP. falciparum[16], or theless complete but extensive sequence data from the rodentmalaria speciesP. yoelii [15], as ‘scaffolds’ upon which toorganise the less extensive genome data currently availablefor P. chabaudi.
We describe here the characteristics of recombinantclones from two genetic crosses between drug-resistantand drug-sensitive clones ofP. chabaudiusing 658 AFLPmarkers. We identify markers linked to the chloroquineresistance phenotype and identify their positions within agenetic linkage map[17]. Sequence analysis of these mark-ers has enabled the identification of homologous sequencesin P. falciparum. In this way, we have produced a map of theP. chabaudiCQR locus at considerably higher resolutionthan available previously, and show that homologues of ge-netically linked markers inP. chabaudiare also physicallylinked in P. falciparum.
2. Materials and methods
2.1. Parasite clones and genetic crosses
The work was carried out with clones derived from twoclonedP. chabaudilines denoted AS and AJ, which differfrom one another by numerous AFLP markers[12]. The ori-gins of theP. chabaudiparasite clones used here are shownin Table 1. Procedures for genetic crosses and the determi-nation of chloroquine responses of independent recombi-nant parasites have been described previously[11,18–20].Briefly, a low-level chloroquine-resistant clone AS(3CQ)was generated by growing the chloroquine sensitive (butpyrimethamine resistant) clone AS(0CQ) in the presenceof low concentrations of chloroquine. After cloning andpassage through mosquitoes, this parasite was crossed witha genetically distinct chloroquine-sensitive clone, AJ. Theprogeny were cloned and the responses of independentrecombinant clones to chloroquine were determined[11].Twenty of these clones are included in this analysis.
Table 1P. chabaudiclones used in this study. r, resistant, hr, high resistance, vhr,very high resistance
Clone Selected from Chloroquineresponse
Crosswith AJ
Reference
AS Sensitive [18]AS(0CQ) AS Sensitive Yes [18]AS(3CQ) AS(0CQ) Resistant (r) Yes [11,19]AS(15CQ) AS(3CQ) Resistant (hr) No [20]AS(30CQ) AS(15CQ) Resistant (vhr) Yes [20]AJ Sensitive [11,18–20]
Eight clones from another genetic cross, between AJand a highly chloroquine-resistant clone AS(30CQ)[20],were also characterised for chloroquine response and AFLPmarkers.
2.2. Tests for chloroquine sensitivity
Parent and cross progeny clones of the AS(3CQ)× AJcross were tested for chloroquine sensitivity in CBA mice,using daily oral doses of 3 mg kg−1 chloroquine for 8 days,as described by Carlton et al.[11]. Resistance levels wereclassified as ‘lr’ (low resistance), or ‘r’ (resistance similarto AS(3CQ)), depending on their growth patterns followingtreatment[11]. Briefly, clones which were eliminated by thedrug until at least day 12 after commencement of drug treat-ment were classified as sensitive. Clones which were ini-tially eliminated by the drug, but re-appeared in the bloodat very low parasitaemia from day 9 and grew slowly there-after, were classified as having low resistance (lr). Cloneswhich exhibited high parasitaemias immediately after drugtreatment, and continued to grow well on subsequent days,were defined as resistant (r).
Progeny of the AS(30CQ)× AJ cross were tested simi-larly for chloroquine response, using the method of Padua[20] in which doses of chloroquine of either 5, 10, or30 mg kg−1 were administered orally for 6 days. Cloneswere classified according to their pattern of growth aftertreatment as having very high resistance to chloroquine(vhr), if growth was similar to AS(30CQ), resistance (r),low resistance (lr) or sensitivity (s).
2.3. DNA extraction
DNA was prepared from individual recombinant progenyclones for AFLP analysis as previously described[12].Parasitised red cells were harvested into citrate saline andpassed twice through a column of fibrous cellulose pow-der (CF11, Whatman)[21] and subsequently twice throughPlasmodipurTM filters (Euro-Diagnostica), in order to re-move mouse leucocytes. The parasitised cells were washedtwice in PBS and the parasites then released by saponinlysis [22]. The parasite pellet was resuspended in lysissolution (10 mM Tris, pH 8.0, 50 mM EDTA, 0.1% SDS,Proteinase K 1 mg ml−1) and incubated overnight at 42◦C.After phenol extraction, DNA was first precipitated bypropan-2-ol and 4 M ammonium acetate, and then treatedusing standard procedures[23].
2.4. AFLP markers
AFLP analysis of parental and recombinant clones fromthe crosses was performed as previously described[12].Briefly, genomic DNA was digested withEcoRI andMseI.Pairs of complementary primers (adaptors) specific for eachrestriction site were annealed and ligated to the cohesiveends of the digested DNA. DNA was pre-amplified using

P. Hunt et al. / Molecular & Biochemical Parasitology 136 (2004) 157–164 159
pairs of primers complementary to each of the adaptor pairs.Further amplification of subsets of these products was per-formed using radioactively labelledEcoRI primers and unla-belledMseI primers, each containing two additional (selec-tive) bases at their 3′ termini. After electrophoresis of am-plified products and autoradiography, bands unique to eachparental clone, AS and AJ (AFLP markers), were scoredfor presence or absence in all of the progeny clones. 28 re-combinant clones were scored in this way for each of 658markers.
2.5. Linkage analysis
Linkage between the inheritance of AJ and AS parentalAFLP markers with chloroquine sensitivity or resistanceamong the progeny clones was calculated and expressed as alinkage ratio[24]. The linkage ratio is the number of progenyclones showing linkage between the parental allele at a spe-cific locus and drug response divided by the total numberof progeny characterised. Those markers showing completelinkage with the chloroquine response have a linkage ratioof 1. Markers showing no linkage have linkage ratios dis-tributed around 0.5.
2.6. Genetic mapping of linked markers
Markers showing a linkage ratio greater than 0.75 wereidentified and organised into a linkage map in two ways,as follows. First, the pattern of inheritance of these mark-ers amongst the 20 recombinant clones was examined andused to order the markers manually into identical linkagegroups. These groups were then organised into an orderwhich gave the fewest predicted recombination events anddouble crossovers. Secondly, the same markers were mappedusing Mapmaker[25]. Any ambiguities between the re-sults of the two methods were identified and resolved byre-inspection of AFLP gels.
2.7. Characterisation of linked markers
A number of AFLP markers which were linked to chloro-quine resistance were characterised as follows. The AFLPband was cut from the gel and soaked overnight in TE buffer(10 mM Tris–Cl, 1 mM EDTA, pH 8.0). DNA was ampli-fied using appropriate adaptor primers with selective bases.PCR products were either sequenced directly in both di-rections using appropriate primers, or cloned using TOPOTA Cloning® (InvitrogenTM) prior to sequencing of plas-mid DNA. After cloning, a number of isolated transformantswere used to generate plasmid DNA using QIAprep SpinMiniprep Kits (Qiagen®). Sequencing used ABI Prism®
BigDyeTM Terminator (Applied Biosystems) cycle sequenc-ing ready reaction kits. Sequences were analysed, and com-plementary sequences aligned using GCG (University ofWisconsin Genetics Computer Group).
2.8. Identification of orthologous loci in P. falciparum andP. yoelii
TheP. chabaudigenomic database at The Sanger Centre(http://www.sanger.ac.uk/Projects/Pchabaudi/blastserver.shtml) was searched with AFLP sequences. Identical indi-vidual sequences or contigs were identified and used to in-terrogate theP. falciparumor P. yoelii genome databases atwww.ncbi.nlm.nih.gov/projects/Malaria/plasmodiumbl.html.
Sequence data forP. falciparum chromosomes 6 and13 were obtained from The Sanger Institute website athttp://www.sanger.ac.uk/Projects/Pfalciparum/. Sequenc-ing of P. falciparumchromosomes 6 and 13 has been ac-complished as part of the Malaria Genome Project with sup-port by The Wellcome Trust. Preliminary sequence and/orpreliminary annotated sequence data from thePlasmodiumyoelii genome was obtained from The Institute for GenomicResearch website (www.tigr.org). This sequencing programis carried out in collaboration with the Naval Medical Re-search Center and is supported by the US Department ofDefense.
If no significantly similar putativeP. falciparumortho-logue was identified,P. chabaudisequences were used toidentify orthologues in theP. yoelii database. Componentsequences fromP. yoelii ‘hits’ were then used to interro-gate theP. falciparumdatabase again. Occasionally, whenrequired, overlappingP. chabaudisequences or paired readswere used to identify sequences in either theP. yoeliior theP. falciparumdatabases.
2.9. Identification of new markers in P. chabaudi
The P. chabaudigenome database was used to identifynew polymorphic markers.P. falciparum gene sequencesat loci of interest were used to identifyP. chabaudior-thologues. These sequences were used to design primerpairs for amplification of the homologous sequence fromboth the AJ and the AS(3CQ) parent clones. These prod-ucts were sequenced, and sequence differences between theclones identified. These polymorphisms were then used todetermine from which parent each allele had been inher-ited in the recombinant progeny clones, using PCR-RFLP,mutation-specific PCR or DNA sequencing.
3. Results
3.1. Inheritance and linkage of AFLP markers
28 independent recombinant progeny clones from twogenetic crosses between the chloroquine-sensitive cloneAJ and the chloroquine-resistant clones AS(3CQ) andAS(30CQ) were analysed for their inheritance of 658 AFLPmarkers. Linkage ratios between the inheritance of eachmarker and drug response phenotype were calculated foreach marker. 613 markers gave a linkage ratio of between

160 P. Hunt et al. / Molecular & Biochemical Parasitology 136 (2004) 157–164
0.25 and 0.75. 45 markers gave a linkage ratio of greater than0.75.
The patterns of inheritance of these 45 markers in theprogeny clones were examined in order to map the mark-ers within linkage groups. 41 could be placed within sevenlinkage groups, each of which showed an identical patternof inheritance among the progeny clones (except for rareoccasions where data could not be collected). These link-age groups were ordered manually in such a way as to re-duce the number of required recombination events amongthe progeny clones. This order was confirmed using Map-maker. The predicted arrangement of the markers in link-age groups is shown inTable 2. The predicted order of thelinkage groups defines the positions of recombination events(crossovers) in progeny clones, which are also indicated inTable 2.
One linkage group (group ‘0’) included three markers(asat03ga, astc01caand ajat01aa) which showed perfectlinkage to the chloroquine resistance phenotype. Three fur-ther linkage groups (2W, 4W and 5W) are predicted to lie toone side (west—arbitrary direction) of the CQR locus. Theycontain five, six and two AFLP markers, respectively. Three
Table 2Linkage of AFLP markers with chloroquine resistance
AFLP markers (asag01tato ajat03ct) are arranged in linkage groups (5W–4E) from left to right. AJ markers shown in white, AS(3CQ) and AS(30CG)markers shown in black. Linkage group ‘0’ indicates markers which show complete linkage with chloroquine resistance. Chloroquine (CQ) responsesshown as black include all levels of resistance (lr, r and vhr). Grey boxes and ‘nd’ indicate not done.
linkage groups (1E, 2E and 4E) lie to the other side (east).They contain nine, seven and nine AFLP markers, respec-tively. This analysis therefore predicts a local genetic mapof AFLP markers in the vicinity of the CQR locus.
3.2. Sequencing of AFLP markers and characterisation ofpolymorphisms
27 of the AFLP markers linked to chloroquine-responsewere selected for sequencing. 21 of these, nine AS and 12 AJmarkers, gave unambiguous sequence data. These sequenceswere then used to search for identical or highly similar se-quences in theP. chabaudi(AS) genome database. Identi-cal (for AS markers) or near-identical (for AJ markers) se-quences were identified in four and eight cases, respectively.
Sequence differences between the eight AFLP bands spe-cific to AJ and highly similar sequences in theP. chabaudigenome database (from AS) are expected to identify the pre-cise sequence differences which give rise to the AS/AJ AFLPbands. Such polymorphisms may arise in one of five ways:(i) a mutation may occur in anEcoRI recognition sequence,(ii) a mutation may occur in anMseI recognition sequence,

P. Hunt et al. / Molecular & Biochemical Parasitology 136 (2004) 157–164 161
Table 3AFLP polymorphisms
AJ AFLP markera Linkage group Database sequence identifier Full namea AS marker predicteda Type of mutation
ajtt04ta 2W pch0172a05.p1c ajtt62ta astt48aa MseI recognition siteajga02tt 2W pch0029b02.p1c ajga170tt asca170tt EcoRI selective baseajca02gt 2W pch0981h11.q1c ajca185gt asca194gg MseI recognition siteajcc04tt 1E pch1052b01.q1c ajcc68tt – EcoRI recognition siteajcg02at 1E pch0874b01.q1c ajcg105at ascg101ta MseI recognition siteajac01ca 2E pch.0.002497 ajac129ca asac61tt MseI recognition siteajtt03at 2E pch0051d02.p1c ajac85ca - EcoRI recognition siteajtt03ca 2E pch0861f02.p1c ajtt52ca astt52ta MseI selective base
a AFLP markers are initially named according to the clone in which the band appears (letter 1–2), the two selective bases at theEcoRI site (letter3–4), numbered serially according to size (number 5–6) and the two selective bases at theMseI site (letter 7–8). In the full name, the serial numbersare replaced by the length of the fragment in base pairs.
(iii) a mutation may occur in one of the two selective basesimmediately adjacent to theEcoRI site, (iv) a mutation mayoccur in one of the two selective bases immediately adja-cent to theMseI site or, (iv) an insertion or deletion mayoccur within the fragment. Our analysis also enabled us topropose the existence, size, and selective bases of the corre-sponding AS allele (‘alternative allele’) for a number of theAJ AFLP markers.
Table 3shows a summary of the types of polymorphismspredicted for the eight AJ AFLP markers for which highlysimilar sequences appear in the AS database. Two markersshowed mutations in theEcoRI recognition sequence, fourshowed mutations in theMseI recognition sequence, oneshowed a mutation in one of theEcoRI selective bases andone showed a mutation in theMseI selective base. Nonegave an insertion or deletion (‘indel’) mutation. One ASmarker,ascg02ta(ascg101ta) was identical to the predictedalternative allele ofajcg02at.
3.3. Syntenic relationships of the AFLP markers
Having established the likely genetic linkage of our AFLPmarkers, we wished to investigate their physical linkage.In the absence of complete genome sequence data forP.chabaudi, this could most easily be attempted by identify-ing homologues of the AFLP marker sequences in theP.falciparumsequence database by BLAST searches.
Sequences of linked AFLP markers or corresponding se-quence from theP. chabaudidatabase were used to inter-rogateP. falciparumsequence databases. In the absence ofa direct ‘hit’ in theP. falciparumdatabase, more complexsearch strategies were employed. These included (i) usingpaired reads from theP. chabaudidatabase as the query ob-ject and (ii) interrogating theP. yoelii sequence databaseand then usingP. yoelii homologues as an ‘intermediate’ tosearch theP. falciparumdatabase. In total, 17 of the 21 mark-ers sequenced were successfully mapped toP. falciparum,including two (astc01caand asat03ga) of the three show-ing perfect linkage to chloroquine resistance. Nine markerswere mapped directly from AFLP sequence orP. chabaudidatabase sequence, six were mapped by usingP. yoelii se-
quence as an intermediate, one was mapped by using aP.chabaudipaired read, and one by using both aP. chabaudipaired read andP. yoeliias an intermediate. We were unableto find suitable homologues of the four remaining markersequences in eitherP. falciparumor P. yoelii.
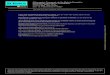
The chromosomal locations of homologues of marker se-quences (or sequences closely linked physically) inP. falci-parumare shown inTable 4. They are all situated on eitherchromosome 6 or 13. Homologues of three RFLP mark-ers previously located onP. chabaudichromosome 11 andshown to be most closely linked to the CQR locus[11] alsomap toP. falciparumchromosomes 6 (H2A) or 13 (EF-1αand PCNA). The physical locations of linked AFLP andRFLP markers inP. falciparumand the map of the geneticlinkage groups are shown inFig. 1. This shows that the lo-cation of markers in the genetic linkage map forP. chabaudiis co-linear with the physical locations ofP. falciparumhomologues. This confirms the existence of significant genesynteny betweenP. falciparumand P. chabaudi[13]. The
Table 4Physical locations of AFLP markers
AFLP marker Linkage group Mapped position inP. falciparum
ajtt04ta 2W pf6-998ajct01tc 2W pf6-1034ajga02tt 2W pf6-1073ajca02gt 2W pf6-1087asat03ga 0 pf6-1310astc01ca 0 pf6-1314ajat03aa 1E pf6-1334ajcc04tt 1E pf13-2052ajcg02at 1E pf13-2246ascg02ta 1E pf13-2246astt02aa 2E pf13-2370ajtt02ga 2E pf13-2383ajac01ca 2E pf13-2405ajtt03at 2E pf13-2537asat03aa 4E pf13-2583asca01ga 4E pf13-2603asag02ca 4E pf13-2656
AFLP markers which have been mapped toP. falciparumchromosomes.The map position is given by the chromosome number (before hyphen)and the position in kilobases along this chromosome (after hyphen).

162 P. Hunt et al. / Molecular & Biochemical Parasitology 136 (2004) 157–164
P. falciparum chromosome 13
P. chabaudi chromosome 11
100 kb
CQR locus
3W
H2A
2W
ajtt04taajct01tcajga02ttajca02gt
0
asat03gaastc01ca
1E
ajat03aaajcc04ttEF-1Aajcg02atascg02ta
2E
astt02aaajtt02gaPCNAajac01caajtt03at
4E
asat03aaasca01gaasag02ca
ajcc
04tt
EF-1
A
ajcg
02at
/asc
g02t
a
asat
03aa
asca
01ga
asag
02ca
astt0
2aa
ajtt0
2ga
PCN
Aaj
ac01
caaj
tt03a
t
ajat0
3aaaj
tt04t
aaj
ct01t
caj
ga02
ttaj
ca02
gt
asat
03ga
astc0
1ca
H2A
pf6-
1100
pf6-
1167
Linkage group
P. falciparum chromosome 6
pf13
-104
3
pf13
-157
8
pf13
-191
0pf
13-1
994
Fig. 1. Gene synteny and chloroquine resistance locus in P. chabaudi. Markers genetically linked to the CQR locus on P. chabaudi chromosome 11(bottom) are arranged by colour into linkage groups (named 3W (west)–4E (east)), each of which contains markers with identical patterns of inheritancein 28 recombinant clones from a genetic cross between the chloroquine sensitive clone AJ, and the resistant clones AS(3CQ) and AS(30CQ) (see Table 2).The order of the groups (3W (red)–4E (blue)) is according to the predicted number of recombination events (crossovers) between the markers and thechloroquine resistance locus. They have been physically mapped (genome database) to P. falciparum chromosomes 6 and 13 (top). The possible (genetic)extent of the CQR locus in P. chabaudi (below) and the physical extent of its probable homologue in P. falciparum (above) is shown as a black bar. Thephysical scale (100 kb) for P. falciparum is also indicated.
P. chabaudi chromosome 11 syntenic blocks described hereare in excess of 600 kb for P. falciparum chromosome 13and 540 kb for P. falciparum chromosome 6.
3.4. Locus determining chloroquine-resistance in P.chabaudi
The CQR locus is predicted to lie on P. chabaudi chromo-some 11 between AFLP marker ajca02gt (the most easterlyof the 2W linkage group) and ajat03aa (the most westerlyof the 1E linkage group). The homologous locus in P. falci-parum is predicted to extend 250 kb on chromosome 6. Sincethe distance between AFLP markers ajca02gt and asat03gawas quite large (210 kb), we wished to define the locus moreclosely by generating more markers in this interval and usethese to evaluate the linkage with chloroquine resistance inthe cross progeny.
The P. chabaudi orthologues of two P. falciparum genesin this region (a putative oxido-reductase (NP703919);6-pyruvoyl tetrahydropterin synthase (NP703938)) were
identified in the P. chabaudi database. These loci are calledpf6-1100 and pf6-1167, respectively and their positionsare indicated in Fig. 1. Products were amplified from P.chabaudi AS and AJ and sequenced. Both gave polymor-phisms between AS and AJ (data not shown). The inher-itance of the oxido-reductase was identified in progenyclones of the two genetic crosses, using mutation-specificPCR. That of the 6-pyruvoyl tetrahydropterin synthase wasidentified using a TaqI RFLP of the amplified product. Bothloci showed an inheritance pattern which was perfectlylinked to chloroquine resistance.
3.5. Characterisation of syntenic breakpoint in P.falciparum
In order to characterise further the syntenic breakpoint in-volving P. falciparum chromosomes 6 and 13, we identifiedfour P. falciparum loci lying upstream of a locus homol-ogous to marker ajcc04tt on chromosome 13, denoted re-spectively pf13-1043, pf13-1578, pf13-1910 and pf13-1994

P. Hunt et al. / Molecular & Biochemical Parasitology 136 (2004) 157–164 163
(Fig. 1). These markers are in genes encoding dihydrofolatesynthase (located at position 1043 kb), an ABC transportermolecule (position 1578 kb), a hypothetical protein (position1910 kb) and another ABC transporter molecule (1994 kb).Orthologues of these genes were identified in P. chabaudiand polymorphisms of each identified between AS(3CQ)and AJ. None of them were linked to the chloroquine resis-tance phenotype in the progeny of the cross, showing thatthese loci were not located on P. chabaudi chromosome 11,and were therefore part of a different syntenic group in thisspecies. A syntenic breakpoint in P. falciparum chromosome13 is therefore predicted to lie between pf13-1994 and a lo-cus homologous to ajcc04tt at position 2052 kb.
We chose not to characterise the syntenic breakpoint onP. falciparum chromosome 6, because the P. falciparum ho-mologue of AFLP marker ajat03aa lies very close to oneend of this chromosome (position 1334 kb).
4. Discussion
Previous work has shown that a chloroquine resistance lo-cus in P. chabaudi is not linked to either pccg10 or pcmdr1,the orthologues of the P. falciparum genes, pfcrt and pfmdr1,respectively [10]. Instead a locus determining resistance wasfound to be located on chromosome 11 [11]. In the latterstudy, the most closely linked RFLP markers, H2A, EF-1α
and PCNA, CRK2 and OPL-04 were predicted to lie withinabout 20 cM of the CQR locus. Here, we have confirmedand extended the analysis of recombinant progeny of geneticcrosses between chloroquine-resistant and sensitive clonesof P. chabaudi using a large number of AFLP markers. Thesemarkers have provided much more resolution than was pos-sible with the RFLP markers previously reported [11], threebeing perfectly linked with chloroquine resistance. Finer res-olution of the resistance locus is limited by the number ofcrossover events involving this region of chromosome 11among the progeny clones examined here. The inheritanceof the complete set of AFLP markers in the progeny of thegenetic crosses has enabled us to construct a genome-widegenetic map, to evaluate the distribution of AFLP markersacross the genome and to estimate the recombination rate.
It is not possible to order AFLP markers within a linkagegroup without data on their physical linkage (or further ge-netic data from additional recombinant progeny). Sequenceanalysis and BLAST searching of markers linked to thechloroquine resistance locus have identified the location oftheir orthologues to P. falciparum chromosomes 6 and 13.The physical linkage of the orthologues in P. falciparum isco-linear with the predicted genetic linkage in P. chabaudi.This confirms accumulating evidence of considerable genesynteny between Plasmodium species [13–15]. In this case,the two sequence blocks in P. chabaudi chromosome 11which are syntenic with P. falciparum chromosome 6 and13 are likely to be at least 540 and 600 kb, respectively.We believe that this is well within the range reported for P.
berghei and P. falciparum where P. berghei chromosomesshow between 1 and 5 syntenic blocks which range in sizebetween 80 kb and a whole chromosome (T. Kooij, personalcommunication).
We have been able in these ways to define the CQR locusto within an approximately 250 kb region of chromosome11 of P. chabaudi which is syntenic with chromosome 6 ofP. falciparum. Since we have been able to generate geneticmarkers at any position within this region using the P. fal-ciparum and P. chabaudi sequence databases, the possibleboundaries of the CQR locus are limited by the number ofrecombinant clones rather than the density of markers per se.
Although it is possible that some of the P. falciparumgenes in this region may not have orthologues in P. chabaudi,and that conversely there may be some genes in this regionof P. chabaudi chromosome 11 which do not have ortho-logues in P. falciparum, we are currently exploring a num-ber of strategies for identifying the gene determining thechloroquine response. Firstly, AS(3CQ) was derived directlyfrom AS(0CQ). Therefore, any gene sequence differencesbetween these clones are likely to be extremely good can-didates for mutations responsible for the chloroquine resis-tance phenotype, especially if they are located within the re-gion described here. So far, we have not found any genes inthis region encoding proteins which are obvious candidatesfor a chloroquine resistance gene, such as an ABC trans-porter. Other strategies include further cloning of recombi-nant progeny in order to obtain a clone with recombinationbetween the parental genomes within the limits of the locusdefined here.
Our data illustrate how genes encoding selectable pheno-types such as drug-resistance may be precisely mapped us-ing the genetic markers, cross progeny characterisation andthe bioinformatic strategies used here. The AFLP markershave proved particularly valuable for this approach, for thefollowing reasons. First, polymorphisms are easy to identifyas presence or absence of bands on a gel. Second, their inher-itance patterns among cross progeny clones can be assessedrapidly because many independent AFLP markers can be vi-sualised on a single gel. Third, individual markers are easy tosequence. Finally, the existence of genomic databases for P.chabaudi, P. yoelii and P. falciparum has enabled the phys-ical mapping of many of the AFLPs on to chromosomes.This approach has demonstrated that the AFLP markers usedin this work are remarkably well distributed, at least in theparts of the genome studied here. Along with previouslydescribed gene synteny, it also demonstrates how the com-plete P. falciparum database can be used as a ‘scaffold’ onwhich to structure the incomplete data from the P. chabaudigenome sequence project.
Acknowledgements
We wish to thank Ronnie Mooney and Les Steven forexcellent technical assistance, as well as the referees of this

164 P. Hunt et al. / Molecular & Biochemical Parasitology 136 (2004) 157–164
paper for their constructive criticisms. We are grateful tothe Medical Research Council and the Wellcome Trust whosupported this research.
References
[1] Fidock DA, Nomura T, Talley AT, et al. Mutations in the P. falci-parum digestive vacuole transmembrane protein Pfcrt and evidencefor their role in chloroquine resistance. Mol Cell 2000;6:861–71.
[2] Babiker HA, Pringle SJ, Abdel-Mushin A, Mackinnon M, Hunt P,Walliker D. High-level chloroquine resistance in Sudanese isolates ofPlasmodium falciparum is associated with mutations in the chloro-quine resistance transporter gene pfcrt and the multidrug-resistancegene pfmdr1. J Infect Dis 2001;183:1535–8.
[3] Chen N, Russell B, Staley J, Kotecka B, Nasveld P, Cheng Q.Sequence polymorphisms in pfcrt are strongly associated withchloroquine resistance in Plasmodium falciparum. J Infect Dis2001;183:1543–5.
[4] Djimde A, Doumbo OK, Steketee RW, Plowe CV. Application of amolecular marker for surveillance of chloroquine-resistant falciparummalaria. Lancet 2001;358:890–1.
[5] Dorsey G, Kamya MR, Singh A, Rosenthal PJ. Polymorphisms in thePlasmodium falciparum pfcrt and pfmdr1 genes and clinical responseto chloroquine in Kampala, Uganda. J Infect Dis 2001;183:1417–20.
[6] Mayor AG, Gomez-Olive X, Aponte JJ, et al. Prevalence of theK76T mutation in the putative Plasmodium falciparum chloroquineresistance transporter (pfcrt) gene and its relation to chloroquineresistance in Mozambique. J Infect Dis 2001;183:1413–6.
[7] Sidhu AB, Verdier-Pinard D, Fidock D. Chloroquine resistance inPlasmodium falciparum malaria conferred by pfcrt mutations. Sci-ence 2002;298:210–3.
[8] Reed MB, Saliba KJ, Caruana SR, Kirk K, Cowman AF. Pgh1 mod-ulates sensitivity and resistance to multiple antimalarials in Plas-modium falciparum. Nature 2000;403:906–9.
[9] Carlton JM, Hayton K, Cravo PV, Walliker D. Of mice and malariamutants: unravelling the genetics of drug-resistance using rodentmalaria models. Trends Parasitol 2001;17:236–42.
[10] Hunt P, Cravo PVL, Donleavy P, Carlton JM-R, Walliker D. Chloro-quine resistance in Plasmodium chabaudi: are chloroquine resistance
transporter (crt) and multi-drug resistance (mdr1) orthologues in-volved? Mol Biochem Parasitol 2004;133:27–35.
[11] Carlton J, Mackinnon M, Walliker D. A chloroquine resistance locusin the rodent malaria parasite Plasmodium chabaudi. Mol BiochemParasitol 1998;93:57–72.
[12] Grech K, Martinelli A, Pathirana S, Walliker D, Hunt P, Carter R.Numerous, robust genetic markers for Plasmodium chabaudi by themethod of amplified fragment length polymorphism. Mol BiochemParasitol 2002;123:95–104.
[13] Carlton JMR, Vinkenoog R, Waters A, Walliker D. Gene synteny inspecies of Plasmodium. Mol Biochem Parasitol 1998;93:285–94.
[14] Carlton JMR, Galinski MR, Barnwell JW, Dame JB. Karyotypeand synteny among the chromosomes of all four species of humanmalaria parasite. Mol Biochem Parasitol 1999;101:23–32.
[15] Carlton JM, Angiuoli SV, Suh BB, et al. Genome sequence and com-parative analysis of the model rodent malaria parasite Plasmodiumyoelii yoelii. Nature 2002;419:512–9.
[16] Gardner MJ, Hall H, Fung E, et al. Genome sequence of the humanmalaria parasite Plasmodium falciparum. Nature 2002;419:498–511.
[17] Martinelli A. Ph.D. thesis, Strain-specific immunity in malaria: amolecular and genetic approach. University of Edinburgh; 2003.
[18] Walliker D, Carter R, Sanderson A. Genetic studies on Plasmod-ium chabaudi: recombination between enzyme markers. Parasitology1975;66:309–20.
[19] Rosario VE. Genetics of chloroquine resistance in malaria parasites.Nature 1976;261:585–6.
[20] Padua RA. Plasmodium chabaudi: genetics of resistance to chloro-quine. Exp Parasitol 1981;52:419–26.
[21] Homewood CA, Neame KD. A comparison of methods used for theremoval of white cells from malaria-infected blood. Ann Trop MedParasitol 1976;70:249–51.
[22] Carter R. Studies on enzyme variation in the murine malaria parasitesPlasmodium berghei, P. yoelii, P. vinckei and P. chabaudi by starchgel electrophoresis. Parasitology 1978;76:241–67.
[23] Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a laboratorymanual, second ed. New York: Cold Spring Harbor Laboratory; 1989.
[24] Wellems TE, Walker-Jonah A, Panton LJ. Genetic mapping of thechloroquine resistance locus on Plasmodium falciparum chromosome7. Proc Natl Acad Sci USA 1991;88:3382–6.
[25] Manly KF, Cudmore Jr RH, Meer JM. Map Manager QTX,cross-platform software for genetic mapping. Mamm Genome2001;12:930–2.