Embed Size (px)
Citation preview

A r t i c l e
GENE EXPRESSION PROFILINGDURING EARLY RESPONSETO INJURY AND MICROBIALCHALLENGES IN THE SILKWORM,Bombyx mori
Fei Liu, Erjun Ling, and Shan WuResearch Center for Insect Science, Institute of Plant Physiology andEcology, Shanghai Institutes for Biological Sciences, The ChineseAcademy of Sciences Graduate School, Shanghai, People’s Republic ofChina
To identify Bombyx mori genes involved in the early response to injuryand microbial challenge, we performed genome-wide gene expression-profiling experiments using oligonucleotide DNA microarrays. Ofapproximately 23,000 genes examined, 465 displayed changes in mRNAexpression levels. Of these, 306 were induced and 159 were repressed inresponse to injury (injection with phosphate buffer saline) or challengesby Gram-negative (Serratia marcescens), Gram-positive bacteria(Staphylococcus aureus), or fungus (Beauveria bassiana). Many of thesedifferentially expressed genes can be assigned to specific functionalgroups of the innate immune response, including recognition, signaling,melanization and coagulation, and antimicrobial peptides. Seventeenpercent of differentially expressed genes encode proteins with no obvioussimilarity to known functional domains. Of particular interest is amember of the juvenile hormone-binding protein family, which was highlyinduced by both injury and microbial challenges. The possible role ofjuvenile hormone in innate immunity is discussed. �C 2009 WileyPeriodicals, Inc.
Keywords: insect immunity; Serratia marcescens; Staphylococcus aureus;Beauveria bassiana; Bombyx mori; gene expression; real-time PCR
Grant sponsor: National Hi-Tech Research and Development Program of China; Grant number: 2006AA10A119;Grant sponsor: National Basic Research Program of China; Grant number: 2007CB513107.Correspondence to: Shan Wu, Center for Insect Science, Institute of Plant Physiology and Ecology, ShanghaiInstitutes for Biological Sciences, The Chinese Academy of Sciences Graduate School, 300 Feng Lin Road,Shanghai 200032, People’s Republic of China. E-mail: [email protected]
ARCHIVES OF INSECT BIOCHEMISTRY AND PHYSIOLOGY, Vol. 72, No. 1, 16–33 (2009)
Published online in Wiley InterScience (www.interscience.wiley.com).
& 2009 Wiley Periodicals, Inc. DOI: 10.1002/arch.20320

INTRODUCTION
Insects, being the most diverse order of animals on the earth, rely on their efficientinnate immune systems to defend against microbial challenges. Cellular aspects ofinnate immunity involve phagocytosis, nodule formation, and encapsulation byhaemocytes. Humoral immunity involves the prophenoloxidase (PPO)-activatingcascade and the synthesis of a battery of anti-microbial peptides (Gandhe et al.,2006; Kanost et al., 2004).
Insect immunity is well characterized in the dipterans (Ferrandon et al., 2007) anda significant body of data is also available for some lepidopterans (Barat-Houari et al.,2006; Eum et al., 2007; Gandhe et al., 2006). Silkworm (Bombyx mori), a model systemfor lepidopteran insects and also an economically important insect, has contributedenormously to the study of insect immunology especially in humoral immunity.Despite its economic importance, little is known about the molecular mechanism ofimmune response in the silkworm. Although many immune-related proteins involvedin pattern recognition, signal transduction, and transcription regulation have beenidentified, the mechanisms of B. mori for recognizing microbes are largely unknown(Cheng et al., 2008). The completion of the genome sequencing of B. mori (Mita et al.,2004; Xia et al., 2004) provides an opportunity to analyze genome-wide informationon microbe-induced immune genes. In this study, we analyzed immune transcriptomeof the larval fat bodies of silkworms following microbial challenges or physical injurywith oligonucleotide DNA microarrays. Of approximately 23,000 genes tested, 465displayed changes in mRNA expression regardless of challenge, 306 were up-regulated, and 159 were down-regulated. The up-regulation of a selected subset ofgenes from the immune transcriptome was also verified by real-time RT-PCR.Unexpectedly, a gene with juvenile hormone-binding protein domain was found to behighly induced by physical injury and microbial challenges.
MATERIALS AND METHODS
Insects and Microbes
Bombyx mori (Nistari) was reared on mulberry leaves at 251C with 12-h light and 12-hdark cycle. The microbes used are Serratia marcescens Bizio (Strain Number: AS 1.203,Committee on Type Culture Collection; Chinese Academy of Sciences), Staphylococcusaureus subsp. aureus (Strain Number: CICC 10201, China Center of Industrial CultureCollection), and Beauveria bassiana 13 (a gift from Dr. Chengshu Wang).
Microbial Challenge and Sample Preparation
Three-day-old fifth-instar larvae were injected separately with 5ml sterilized PhosphateBuffered Saline (PBS, pH 7.2), heat-killed S. marcescens (1� 104 cells/larva), S. aureus(1� 104 cells/larva), and conidium of B. bassiana (1� 104 cells/larva) suspended in thesame volume of PBS buffer. S. marcescens and S. aureus were cultured overnight in50 ml of SM medium (peptone 10 g, beef extract 3 g, NaCl 5 g, distilled water 1,000 ml,pH 7.0) and SA medium (peptone 5 g, beef extract 3 g, NaCl 5 g, distilled water1,000 ml, pH 7.0) at 371C in a rotating incubator, respectively. B. bassiana werecultured 7 days on PDA plate at 251C in an incubator. Before injection, the bacterial
Gene Expression Profiling in B. mori � 17
Archives of Insect Biochemistry and Physiology

and fungal samples were killed by boiling in water for 5 or 20 min, respectively. PBSinjection was a control for injection wounding (Wang et al., 2004). Untreated larvaewere used as naı̈ve controls. Fat bodies for each experimental group were collectedfrom at least three silkworms. Silkworms were harvested at 6 h after injection(Engstrom et al., 2004). Isolated fat body was washed three times in PBS, andthen pulverized with liquid nitrogen and stored at �701C in Trizol (Invitrogen,Carlsbad, CA).
Oligonucleotide Microarray
The microarray, obtained from the CapitalBio Corporation (Beijing, China), contains23,022 probes, each 70 nucleotides (70-mer) in length, corresponding to theapproximately 23,000 known and predicted B. mori genes (Xia et al., 2007).
RNA isolation, amplification, labeling, hybridization, and microarray imaging anddata analysis have been described in previous studies (Xia et al., 2007). Total RNA wasisolated from each sample using Trizol reagent (Invitrogen, Carlsbad, CA) according tothe manufacturer’s instructions. Total RNA (5 mg) was used to prepare the fluorescentdye–labeled cDNA using cRNA Amplification and Labeling Kit (CapitalBio Corp.,Beijing, China). The labeled cDNAs were dissolved in 80ml of hybridization solution(3� SSC, 0.2% SDS, 5� Denhardt’s solution, 25% formamide), then the hybridiza-tions were performed in a hybridization chamber (BioMixerTM; CapitalBio Corp.)overnight at 421C. After hybridization, slides were washed with washing solution 1 and2 (0.2% SDS, 2� SSC and 2� SSC, respectively) at 421C for 5 min. Arrays werescanned with a confocal LuxScanTM scanner and the images obtained were thenanalyzed using LuxScanTM 3.0 software (both from CapitalBio Corp.). Eachexperimental group was repeated three times. Data were normalized by the LOWESSmethod. The filtered data were further examined to find genes that are differentiallyexpressed between experiment and control sample using SAM software (Tusher et al.,2001). Significance was determined with q-value (%) set at 5%, and ratio of at leasttwofold for the signal intensity between experimental sample and control.
Real-Time Reverse-Transcription Coupled With Polymerase Chain Reactions (Real-TimeRT-PCR)
RNA samples were treated with RNase-free DNase I (Promega, Madison, WI) andreverse-transcribed using M-MLV reverse transcriptase (Invitrogen, Carlsbad, CA).The real-time RT-PCR reactions were performed using the Lightcycler-Faststart DNAmaster SYBR green I PCR kit in a Lightcycler 1.2 real-time PCR System (Roche, Basel,Switzerland) according to the operator’s manual. Sequence-specific oligonucleotideprimers designed for a member of the takeout/Juvenile hormone–binding proteinfamily gene (e96h-0303 described in Saito et al., 2006, and shortened as JHBP0303),and three AMPs were used to yield approximately 100–250 base pair (bp) sequences.All primers used in this study were designed with the Primer Premier 5.0 software(Premier, Canada) and synthesized at Invitrogen. To check the specificity of theamplified product, we performed melting curve analyses at 751 to 951C and gelelectrophoresis. As a control, total RNA samples that were prepared the same way,except that no reverse transcriptase was added, were processed to demonstrateabsence of genomic DNA contamination.
The comparative threshold cycle (CT) method was used for the calculation ofamplification fold (Tichopad et al., 2003). The eukaryotic initiation factor 4A (eIF4A,
18 � Archives of Insect Biochemistry and Physiology, September 2009
Archives of Insect Biochemistry and Physiology

sw22934) gene was used for normalization of cDNA templates. All the real-time PCRexperiments were repeated three times using independent biological samples. Primersequences are presented in Table 1.
RESULTS
General Description
The genome-wide 70-mer oligonucleotide microarray was used to monitor B. mori fatbody gene expression response to microbial challenges. The slides were hybridizedwith silkworm fat body collected 6 h (Irving et al., 2001) after injection with heat-killedGram-negative (S. marcescens) or Gram-positive (S. aureus) bacteria, or conidium offungus (B. bassiana), or PBS.
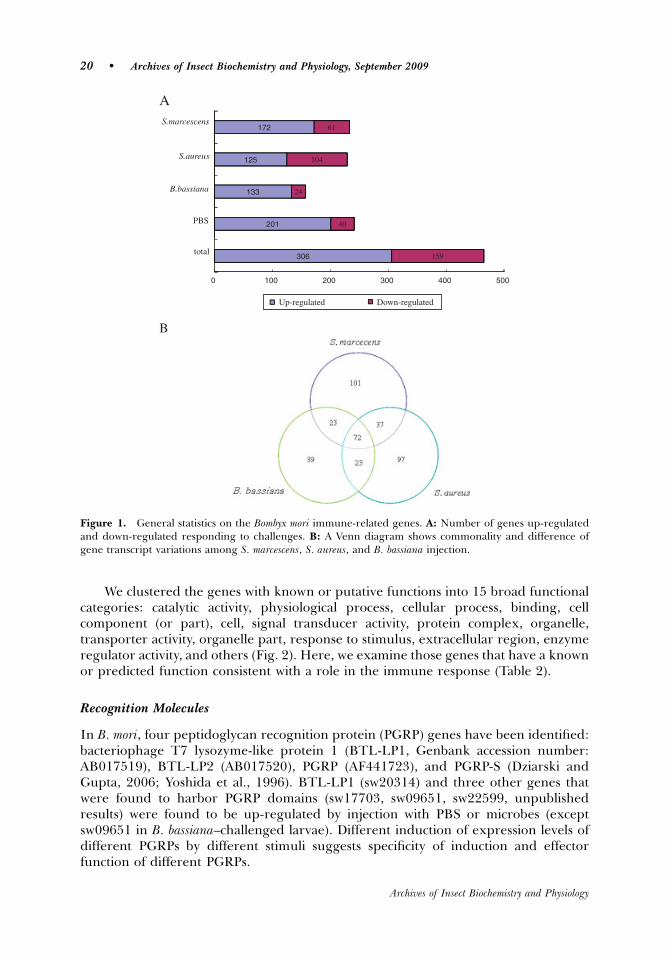
Among the 23,022 probes on the microarray, 465 displayed changes in fluorescentintensity, of which 306 exhibited elevated and 159 repressed levels in injected larvae(Fig. 1A). Of the 465 genes, 17% encodes proteins with no obvious similarity to knownfunctional domains (see Table 3).
S. marcescens, S. aureus, and PBS injection resulted in expression level changes of233, 229, and 241 genes, respectively, whereas B. bassiana injection led to a lowernumber of genes (157 genes). Of these, between 125 and 201 genes were up-regulatedand 24 to 104 genes were down-regulated by at least 2-fold (Fig. 1A).
Many significantly up- and down-regulated genes responded to more than onestimulus. Expression pattern for 50 genes were changed for all four challenges (datanot shown), and 72 in all three microbial challenges (Fig. 1B). These genes were notspecific to the pathogen type, and could be assumed to be involved in housekeeping,wound healing, and common immune-related processes (Eum et al., 2007). In contrast,the non-overlapping ones would be taken as pathogen-specific genes (Fig. 1B).
PBS injection triggers a widespread response compared to other challenges(Table 2). Surprisingly, microbial challenges yielded weaker responses from thesilkworm larvae. Thirty-nine (39) immunity-related genes that exhibited alteredexpression profile by microbial injection were found to be among those identified byinjection with PBS alone (Table 2). However, nine immunity related genes were foundspecific to pathogen challenges. Further, the change in gene expression is generallyless marked after B. bassiana injection compared with bacterial injuries (Table 2). Thisfinding is in agreement with the previous observation showing that fungal infection ismore specific than bacteria in Drosophila (Lemaitre et al., 1997).
Table 1. Primer Pairs for Real-Time RT-PCR
Probe IDIdentity/homology
Forward primer(primer sequence 50-30)
Reverse primer(primer sequence 50-30)
sw15815 Moricin 2 GTAGGAAAGGCAGTCGGTAAAG ATCCATAAATTCACGTTGACATCTsw17805 Attacin ACAGTGAACTCGGATGGAACC AAGTTGACCTTGCCTGCGACsw08844 Lebocin 3 TGTTCTTTGCTCAGGCTTCG GGATGGTCTGCGGTACTTGGsw20826 JHBP0303 AACCGTGATTTGGCGAAGAC CACCTTATCGAACAGTTTGAATGsw22934 EIF4A GGAATGGACCCTGGGACACT ACTGGGCTTGAGCGATAACA
Gene Expression Profiling in B. mori � 19
Archives of Insect Biochemistry and Physiology
We clustered the genes with known or putative functions into 15 broad functionalcategories: catalytic activity, physiological process, cellular process, binding, cellcomponent (or part), cell, signal transducer activity, protein complex, organelle,transporter activity, organelle part, response to stimulus, extracellular region, enzymeregulator activity, and others (Fig. 2). Here, we examine those genes that have a knownor predicted function consistent with a role in the immune response (Table 2).
Recognition Molecules
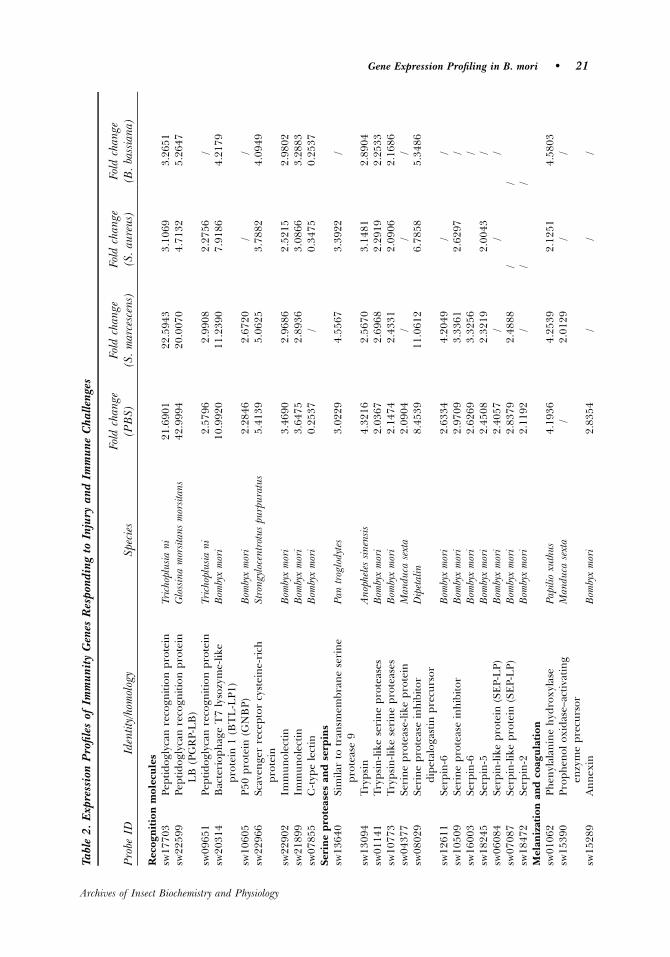
In B. mori, four peptidoglycan recognition protein (PGRP) genes have been identified:bacteriophage T7 lysozyme-like protein 1 (BTL-LP1, Genbank accession number:AB017519), BTL-LP2 (AB017520), PGRP (AF441723), and PGRP-S (Dziarski andGupta, 2006; Yoshida et al., 1996). BTL-LP1 (sw20314) and three other genes thatwere found to harbor PGRP domains (sw17703, sw09651, sw22599, unpublishedresults) were found to be up-regulated by injection with PBS or microbes (exceptsw09651 in B. bassiana–challenged larvae). Different induction of expression levels ofdifferent PGRPs by different stimuli suggests specificity of induction and effectorfunction of different PGRPs.
125
172
104
61
S.aureus
S.marcescens
201
133
40
24B.bassiana
PBS
306 159
0 100 200 300 400 500
total
Up-regulated Down-regulated
B
A
Figure 1. General statistics on the Bombyx mori immune-related genes. A: Number of genes up-regulatedand down-regulated responding to challenges. B: A Venn diagram shows commonality and difference ofgene transcript variations among S. marcescens, S. aureus, and B. bassiana injection.
20 � Archives of Insect Biochemistry and Physiology, September 2009
Archives of Insect Biochemistry and Physiology
Tabl
e2
.E
xpre
ssio
nP
rofi
les
ofIm
mu
nit
yG
enes
Res
pon
din
gto
Inju
rya
nd
Imm
un
eC
ha
llen
ges�
Pro
beID
Iden
tity
/hom
olog
yS
peci
esFo
ldch
ange
(PB
S)
Fold
chan
ge(S
.m
arce
scen
s)Fo
ldch
ange
(S.
aure
us)
Fold
chan
ge(B
.ba
ssia
na)
Rec
og
nit
ion
mo
lecu
les
sw1
77
03
Pep
tid
og
lyca
nre
cog
nit
ion
pro
tein
Tri
chop
lusi
an
i2
1.6
90
12
2.5
94
33
.10
69
3.2
65
1sw
22
59
9P
epti
do
gly
can
reco
gn
itio
np
rote
inL
B(P
GR
P-L
B)
Glo
ssin
am
orsi
tan
sm
orsi
tan
s4
2.9
99
42
0.0
07
04
.71
32
5.2
64
7
sw0
96
51
Pep
tid
og
lyca
nre
cog
nit
ion
pro
tein
Tri
chop
lusi
an
i2
.57
96
2.9
90
82
.27
56
/sw
20
31
4B
acte
rio
ph
age
T7
lyso
zym
e-lik
ep
rote
in1
(BT
L-L
P1
)B
omby
xm
ori
10
.99
20
11
.23
90
7.9
18
64
.21
79
sw1
06
05
P5
0p
rote
in(G
NB
P)
Bom
byx
mor
i2
.28
46
2.6
72
0/
/sw
22
96
6S
cave
ng
erre
cep
tor
cyst
ein
e-ri
chp
rote
inS
tron
gylo
cen
trot
us
purp
ura
tus
5.4
13
95
.06
25
3.7
88
24
.09
49
sw2
29
02
Imm
un
ole
ctin
Bom
byx
mor
i3
.46
90
2.9
68
62
.52
15
2.9
80
2sw
21
89
9Im
mu
no
lect
inB
omby
xm
ori
3.6
47
52
.89
36
3.0
86
63
.28
83
sw0
78
55
C-t
ype
lect
inB
omby
xm
ori
0.2
53
7/
0.3
47
50
.25
37
Ser
ine
pro
teas
esan
dse
rpin
ssw
13
64
0S
imilar
totr
ansm
emb
ran
ese
rin
ep
rote
ase
9P
antr
oglo
dyte
s3
.02
29
4.5
56
73
.39
22
/
sw1
30
94
Try
psi
nA
nop
hele
ssi
nen
sis
4.3
21
62
.56
70
3.1
48
12
.89
04
sw0
11
41
Try
psi
n-l
ike
seri
ne
pro
teas
esB
omby
xm
ori
2.0
36
72
.69
68
2.2
91
92
.25
33
sw1
07
73
Try
psi
n-l
ike
seri
ne
pro
teas
esB
omby
xm
ori
2.1
47
42
.43
31
2.0
90
62
.16
86
sw0
43
77
Ser
ine
pro
teas
e-lik
ep
rote
inM
andu
case
xta
2.0
90
4/
//
sw0
80
29
Ser
ine
pro
teas
ein
hib
ito
rd
ipet
alo
gas
tin
pre
curs
or
Dip
etal
in8
.45
39
11
.06
12
6.7
85
85
.34
86
sw1
26
11
Ser
pin
-6B
omby
xm
ori
2.6
33
44
.20
49
//
sw1
05
09
Ser
ine
pro
teas
ein
hib
ito
rB
omby
xm
ori
2.9
70
93
.33
61
2.6
29
7/
sw1
60
03
Ser
pin
-6B
omby
xm
ori
2.6
26
93
.32
56
/sw
18
24
5S
erp
in-5
Bom
byx
mor
i2
.45
08
2.3
21
92
.00
43
/sw
06
08
4S
erp
in-l
ike
pro
tein
(SE
P-L
P)
Bom
byx
mor
i2
.40
57
//
/sw
07
08
7S
erp
in-l
ike
pro
tein
(SE
P-L
P)
Bom
byx
mor
i2
.83
79
2.4
88
8/
/sw
18
47
2S
erp
in-2
Bom
byx
mor
i2
.11
92
//
/M
elan
izat
ion
and
coag
ula
tio
nsw
01
06
2P
hen
ylal
anin
eh
ydro
xyl
ase
Pap
ilio
xuth
us
4.1
93
64
.25
39
2.1
25
14
.58
03
sw1
53
90
Pro
ph
eno
lo
xid
ase–
acti
vati
ng
enzy
me
pre
curs
or
Man
duca
sext
a/
2.0
12
9/
/
sw1
52
89
An
nex
inB
omby
xm
ori
2.8
35
4/
//
Gene Expression Profiling in B. mori � 21
Archives of Insect Biochemistry and Physiology
Tabl
e2
.C
onti
nu
ed
Pro
beID
Iden
tity
/hom
olog
yS
peci
esFo
ldch
ange
(PB
S)
Fold
chan
ge(S
.m
arce
scen
s)Fo
ldch
ange
(S.
aure
us)
Fold
chan
ge(B
.ba
ssia
na)
An
tim
icro
bia
lp
epti
des
sw1
58
15
Mo
rici
n2
Bom
byx
mor
i3
3.4
86
23
4.5
82
62
3.1
02
81
7.5
49
7sw
11
37
2C
ecro
pin
EB
omby
xm
ori
/1
1.0
90
1/
/sw
17
80
5A
ttac
inB
omby
xm
ori
6.1
12
84
.99
26
2.3
96
42
.00
30
sw0
88
44
Leb
oci
n3
Bom
byx
mor
i4
.09
38
4.2
79
32
.81
82
2.6
30
9sw
01
77
3G
love
rin
1B
omby
xm
ori
5.8
55
17
.69
05
2.1
26
72
.25
91
sw1
25
23
Glo
veri
n2
Bom
byx
mor
i3
.90
49
5.3
55
62
.13
50
2.3
43
0sw
22
04
3G
love
rin
3B
omby
xm
ori
3.3
29
23
.28
70
/2
.33
40
sw1
38
47
Lys
ozy
me
Bom
byx
mor
i/
2.2
23
8/
2.0
60
8O
ther
ssw
16
33
2S
imilar
toca
rbo
xyl
este
rase
Api
sm
ellife
ra5
.64
99
8.4
23
77
.92
58
4.7
30
9sw
01
79
1T
riac
ylg
lyce
rol
lip
ase
Rab
bit
6.7
87
67
.06
57
7.3
49
94
.71
24
sw0
07
34
Tri
acyl
gly
cero
llip
ase
Bom
byx
mor
i/
2.3
81
1/
/sw
08
44
2E
ster
ase-
lik
ep
rote
in(E
SR
-LP
)B
omby
xm
ori
/2
.19
61
//
sw1
40
35
Juve
nile
ho
rmo
ne
este
rase
(Jh
e)M
andu
case
xta
2.1
87
3/
3.1
16
9sw
06
23
7E
ster
ase-
lik
ep
rote
in(E
SR
-LP
)S
esam
ian
onag
rioi
des
/2
.94
27
//
sw1
50
11
Est
eras
eD
Su
ssc
rofa
0.4
93
8/
//
sw1
92
72
Sim
ilar
toca
mp
-sp
ecifi
c3
,5-c
yclic
ph
osp
ho
die
ster
ase
Aed
esae
gypt
i0
.28
03
0.2
12
80
.35
28
0.4
89
9
sw1
18
73
Cac
tus
gen
ep
rod
uct
Dro
soph
ila
mel
anog
aste
r2
.25
89
2.3
69
2/
2.5
01
1sw
17
21
4S
imilar
toto
ll-i
nte
ract
ing
pro
tein
Api
sm
ellife
ra/
/0
.41
92
/sw
11
95
6S
imilar
tod
eath
-ass
oci
ated
pro
tein
Api
sm
ellife
ra/
0.4
92
5/
/sw
05
02
5L
yso
som
alb
eta-
gal
acto
sid
ase
Felis
catu
s2
.90
94
2.2
34
92
.14
92
/sw
04
27
6Im
mu
ne-
rela
ted
Hd
d1
Hyp
han
tria
cun
ea5
.05
04
8.2
66
15
.02
77
4.8
63
0sw
13
77
5P
uta
tive
alco
ho
ld
ehyd
rog
enas
e(A
dh
)B
omby
xm
ori
/0
.46
88
//
sw2
08
26
Juve
nile
ho
rmo
ne–
bin
din
gp
rote
in(J
HB
P)
Bom
byx
mor
i4
3.9
02
33
2.9
09
61
6.5
34
71
9.2
15
6
� Th
efo
ldch
ang
es6
haf
ter
S.
mar
cesc
ens,
S.au
reu
s,B
.ba
ssia
na,
and
PB
Sin
ject
ion
com
par
edw
ith
un
trea
ted
silk
wo
rms
for
imm
un
ity-
rela
ted
gen
esis
sho
wn
.T
he
rati
ow
asse
tat
leas
ttw
ofo
ldfo
rth
esi
gn
alin
ten
sity
bet
wee
nth
eex
per
imen
tal
sam
ple
and
con
tro
l.‘‘/
’’re
pre
sen
tsth
efo
ldch
ang
eso
fth
eg
ene
tran
scri
pt
leve
lsaf
ter
inju
ryo
rim
mu
ne
chal
len
ges
wer
ele
ssth
antw
o.
22 � Archives of Insect Biochemistry and Physiology, September 2009
Archives of Insect Biochemistry and Physiology

Table 3. Expression Profiles of Genes That Were Functionally Unknown�
Probe IDFold change
(PBS)Fold change
(S. marcescens)Fold change(S. aureus)
Fold change(B. bassiana)
sw17247 18.2905 21.1574 12.8344 6.5693sw20604 24.6307 17.6081 18.6700 23.1409sw00797 7.5806 6.4949 5.2014 5.3631sw02678 6.7923 9.5067 6.8036 /sw17275 6.3687 / / /sw08603 / / 4.6625 /sw11765 / / 5.4251 /sw02678 / 9.5067 / /sw00079 5.3453 5.7962 3.3129 2.7468sw02632 5.0048 / / /sw09761 4.0608 4.9885 6.4613 3.1818sw12794 3.2926 4.7914 / 2.9664sw18026 / / / 2.8456sw12493 / / / 2.0096sw03912 / / / 2.4305sw16005 3.9895 / / /sw10431 3.2988 / 2.6194 2.5324sw10148 / 4.6814 2.1957 /sw08642 4.7511 4.1974 2.8815 /sw07963 2.7883 4.0492 2.3753 2.0757sw11074 / / 2.5546 /sw19717 5.6353 4.0446 2.7095 2.7844sw13648 / / 2.1540 /sw17602 7.1853 4.0106 4.1153 5.8004sw12408 / / 3.8821 /sw03498 3.0889 3.9433 2.2826 2.3855sw14454 / 3.4944 3.1226 /sw07317 3.6575 3.4408 2.9814 2.5423sw02618 2.5332 3.2908 / /sw06897 / / 3.5085 6.1501sw20640 / / 2.2202 /sw02142 / 3.2709 / /sw07302 / 3.1618 / /sw08563 2.4355 3.0687 / /sw14629 / 2.6215 / /sw10288 / / 2.2098 2.4630sw20738 2.1487 2.5698 / /sw10594 2.3622 2.5739 / /sw07051 2.1345 2.4302 / /sw11139 2.3327 2.3656 / 2.3907sw05429 / 2.2573 / /sw19907 2.4985 2.2103 / 2.3861sw09807 / 2.1557 / /sw03561 2.0175 2.1371 / /sw21641 / 2.1265 / /sw18617 / 2.1137 / /sw08925 / 2.1072 / /sw16073 2.1093 / / /sw09222 2.0850 / / /sw02788 2.0583 / / /sw12577 2.0365 / / /sw10744 / 2.0662 / /sw19704 2.3315 / / /
Gene Expression Profiling in B. mori � 23
Archives of Insect Biochemistry and Physiology

The Gram-negative-binding protein (GNBP) was first identified in the hemolymph ofimmune-challenged silkworm B. mori (Ochiai and Ashida, 1988; Yoshida et al., 1986).GNBPs have two domains that bind microbial motifs. The C-terminal part of B. mori GNBPhas a strong specificity and high affinity for b-1, 3-glucan, and LPS (Lee et al., 1996). Inmost cases, GNBP genes are expressed in immune tissues such as the fat body andhemocytes and their transcription is upregulated when challenged (Ma and Kanost, 2000).Increased levels of GNBP mRNAs are observed in Anopheles gambiae 30 h after ingestion ofa Plasmodium berghei–infected blood-meal (Dimopoulos et al., 1998). In our study, GNBPwas detected up-regulated more than 2-fold 6 h after PBS and S. marcescens injection.
A third class of putative recognition proteins, the lectins, has been implicated inpathogen recognition in vertebrates and invertebrates (Vasta et al., 1999). C-typelectins, a superfamily of calcium-dependent carbohydrate-binding proteins, functionin pathogen recognition, cellular interactions, and innate immunity in mammals. Twoimmulectins (sw22902 and sw21899) were induced after injection with or without
Table 3. Continued
Probe IDFold change
(PBS)Fold change
(S. marcescens)Fold change(S. aureus)
Fold change(B. bassiana)
sw17282 2.3105 / / /sw08305 2.2801 / / /sw01970 2.6133 / 2.5131 /sw13773 / / 2.4041 /sw10598 2.7434 / 2.4774 /sw20308 / / / 2.0114sw18102 / 0.4875 / /sw05363 0.2528 0.4248 / 0.3462sw15096 0.4679 0.4184 0.4788 0.3339sw16197 / 0.4168 / /sw20163 0.4936 0.2981 / /sw15650 0.4358 / / 0.4453sw03180 / / 0.4964 /sw13072 / / 0.4958 /sw09738 / / 0.4907 /sw14628 / / 0.4951 /sw16440 / / / 2.2166sw17278 / / 0.4708 /sw09405 / / 0.4700 /sw11189 / / 0.4591 /sw12756 / / 0.4512 /sw22766 / / 0.4009 2.3853sw02400 / / 0.3913 /sw14088 / / 0.3642 /sw16976 / / 0.3580 /sw20009 / / 0.3490 /sw00839 / / 0.3409 /sw07211 / / 0.2079 /sw05363 / / 0.1946 /sw07781 / / 0.1533 /
�The fold changes 6 h after S. marcescens, S. aureus, B. bassiana, and PBS injection compared with untreatedsilkworms for genes with unknown functions is shown. The ratio was set at least twofold for the signal intensitybetween the experimental sample and control. ‘‘/’’ represents the fold changes of the gene transcript levels afterinjury or immune challenges were less than two.
24 � Archives of Insect Biochemistry and Physiology, September 2009
Archives of Insect Biochemistry and Physiology

microbes, while one C-type lectin (sw07855) was significantly down-regulated afterS. aureus, B. bassiana, and PBS injection.
In the present study, scavenger receptor as a microbial recognition was up-regulatedby all the treatments, including the fungus challenge. However, in Drosophila, scavengerreceptor was only capable of recognizing bacteria, but not fungi (Ramet et al., 2001).
Serine Proteases and Serpins
The serine proteases and serine protease inhibitors (serpins) constitute the mostabundant group of genes (13 genes) that were triggered by the stimulations (Table 2),
A extracellular region
catalytic activitycell part
protein complexorganelle part
organelle
g
others
response to stimulus
cell
binding
signal transducer activity
transporter activity
enzyme regulator activity
cellular process
physiological process
B S.marcescens
S.aureus
B b iB.b assiana
PBS
0 50 100 150 200 250
catalytic activity binding signal transducer activity transporter activitysignal
enzyme regulator activity physiological process cellular process response to stimulus
cell cell part protein complex organelle part
organelle extracellular region others
Figure 2. Functional distribution of differentially expressed genes by GeneOntology. A: Genes whoseexpression levels changed at least 2-fold regardless of challenge, excluding genes of unknown function. B:Number of genes whose expression changed 2-fold or more according to microbial or PBS injection,excluding genes of unknown function. Genes were assigned to functional categories according to the website(http://www.capitalbio.com).
Gene Expression Profiling in B. mori � 25
Archives of Insect Biochemistry and Physiology

including five serine proteases and eight serpins, indicating that they play animportant role in B. mori defense. PBS injection elicited more serine proteases andserpins than the microbial injections (Table 2). One serine protease (sw04377) and twoserpins (sw06084 and sw18472) were only induced by PBS injection.
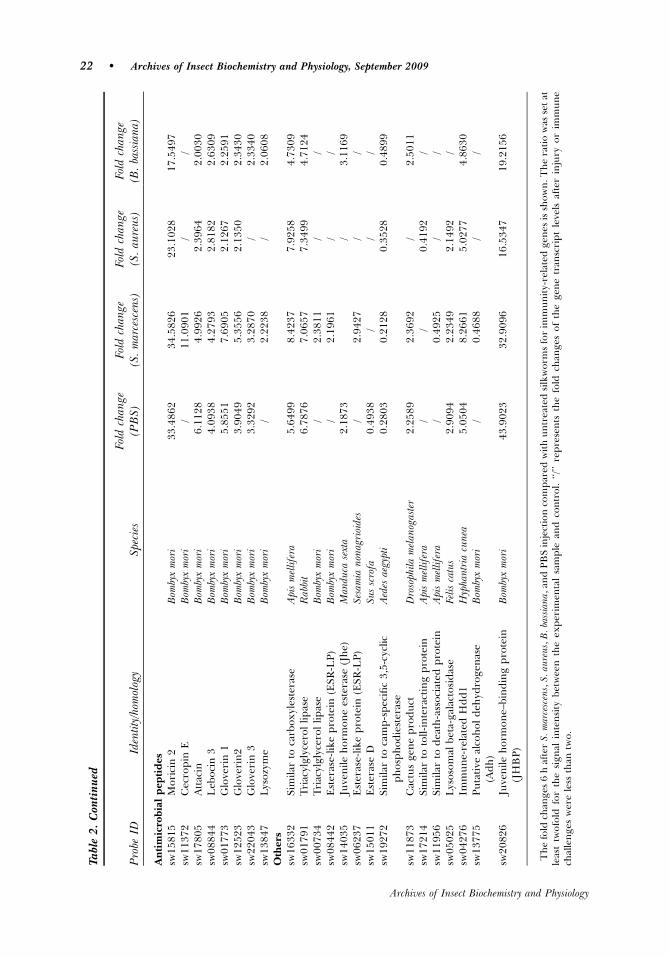
Melanization and Coagulation
Enzymes involved in melanization, including prophenoloxidase-activating enzyme(PPAE) and phenylalanine hydroxylase (PAH), are represented by probes sw15390and sw01062, respectively. PPAE was only induced after immune response toS. marcescens. PAH showed up-regulation of transcript abundance in the four injectionstimulations. PAH is involved in hydroxylation of pheylalanine to produce tyrosine,which is the rate-limiting substrate for melanin production (Johnson et al., 2003).Another induced gene up-regulated after PBS challenge was annexin, which encodes aprotein with homology with Aedes aegypti annexin X, and is involved in melanizationand coagulation (Bartholomay et al., 2004).
Antimicrobial Peptides
Many antimicrobial peptides have been isolated from various insects and at least sixtypes (cecropin, attacin, lebocin, gloverin, enbocin, and moricin) have been identifiedin B. mori (Cheng et al., 2006); five types were found to be induced in our study.Cecropin E showed strong inducement by S. marcescen injection. Gloverin 1, 2, attacin,and lebocin 3 were up-regulated by all the four injections. Gloverin 3 showedinducement by PBS, S. marcescen, and B. bassiana injections. Interestingly, moricin 2was induced much stronger by all four injuries than any other AMPs, suggesting thatmoricin 2 has a very important role in B. mori defense. Besides the AMPs discussedabove, one B. mori lysozyme (sw13847) was induced by S. marcescen and B. bassianainjections.
Other Genes
In addition to known or predicted immune response proteins, some putative genesinvolved in the immune system were identified in this study (Table 2). These genesinclude a series of lipase or esterase, which were putatively involved in detoxificationand energy metabolism (Mouches et al., 1987). A putative homologous of DrosophilaCactus and other two genes encoding toll-interacting protein and death-associatedprotein, all of which may be involved in Toll pathway, were induced and repressed,respectively. Lysosomal enzymes (lysosomal beta-galactosidase) that might play a rolein phagocytosis of invading microorganisms were up-regulated (Koo et al., 2008). Thealcohol dehydrogenase, also engaged in energy metabolism (Eum et al., 2007), whichwas defined as immune-related protein in Drosophila melanogaster and Plutella xylostellabecause of the remarkable increase after infection (Eum et al., 2007), was significantlydepressed after being challenged with S. marcescens in the present study. Thetranscripts of immune-related Hdd1 were up-regulated in each treatment. Up-regulation of Hdd1 previously was detected by PCR-based differential display analysisof fall webworm inoculated with bacteria (Shin et al., 1998).
A member of the takeout/juvenile hormone binding protein family (sw20826) wasfound to be up-regulated by a great extent in response to all the four injections.Juvenile hormone esterase (JHE) precursor was also up-regulated after B. bassiana andPBS challenges.
26 � Archives of Insect Biochemistry and Physiology, September 2009
Archives of Insect Biochemistry and Physiology

In order to confirm our microarray results, we chose three AMPs and JHBP0303for confirmatory real-time RT-PCR. Results shown in Figure 3 demonstrate goodcorrelations between microarray and real-time PCR results.
A
B
1416
els
moricin 2
2468
101214
lativ
e m
RN
A le
ve
02
Rel
PBS S.marcescens S.aureus B.bassiana
C attacin
3
4
5
6
0
1
2
3
A le
vels
Rel
ativ
e m
RN
A
D
PBS S.marcescens S.aureus B.bassiana
JHBP0303
20
30
40
0
10
20RN
A le
vels
Rel
ativ
e m
R
PBS S S B b iPBS S.marcescens S.aureus B.bassiana
lebocin 35
1
2
3
4
5
0
1
Rel
ativ
e m
RN
A le
vels
B.bassianaPBS S.aureusS.marcescens
Figure 3. Confirmatory RT-PCR results. Variations in gene transcript levels in response to microbial andPBS injection measured by real-time RT-PCR. A: Lebocin 3. B: Moricin 2. C: Attacin. D: JHBP0303.
Gene Expression Profiling in B. mori � 27
Archives of Insect Biochemistry and Physiology

DISCUSSION
Silkworm is one of the most important economic insects in the world, and is one of thebest-characterized models for biochemical, molecular genetic, developmental biology,and pathological studies of lepidoptera, because of its large size, complex metabolism,and the abundance of mutants (Xia et al., 2007). Studies on the silkworm immune-related genes will help us understand not only the molecular mechanisms of silkwormimmune system, but also the diversity of insect immunity.
We used high-density oligonucleotide arrays to catalog the genes whose mRNAlevels changed at least twofold during the B. mori response to immune challenges. Weobserved that microbial injection and physical injury induce dramatic changes in geneexpression in the larval silkworm. Among the 23,022 probes on the microarray, 465displayed significant changes in fluorescent intensity, of which 306 were up-regulatedand 159 down-regulated in injected larvae. Of the 465 genes displaying significantchanges in mRNA levels in response to injection, we identified many genes highlyhomologous to the known immune genes, but some genes were not previously knownto be immune-responsive (Table 2). We also found a large number of genes withunknown function but they may be involved in the control and execution of theimmune response (Table 3).
Pathogen recognition by the innate immune system is believed to rely on interactionbetween conserved microbial determinants on the surface of pathogens and host-recognition proteins. Peptidoglycan recognition proteins (PGRPs) are innate immunitymolecules found in many species including insects and mammals (De Gregorio et al.,2001). PGRPs have at least one carboxy-terminal PGRP domain (approximately 165amino acids long), which is homologous to bacteriophage and bacterial type 2 amidases.Insects have up to 19 PGRPs, classified into short (S) and long (L) forms (Dziarski andGupta, 2006). Peptidoglycan is a polymer of (1-4)-linked N-acetylglucosamine and N-acetylmuramic acid, cross-linked by short peptides containing alternating L- and D-aminoacids. The four putative PGRPs in our study (sw17703, sw09651, sw22599, and sw09651)all belong to the N-acetylmuramoyl-L-alanine amidase-like superfamily. The four putativePGRPs may hydrolyze the amide bond between N-acetylmuramic acid and L-alanine andthus remove stem peptides from peptidoglycan. Stem peptides are the four to five aminoacids directly bound to N-acetylmuramic acid. Digestion of peptidoglycan with amidasereduces or eliminates the ability of polymeric peptidoglycan to stimulate insect cells, andthus the function of amidase PGRPs in vivo may be to prevent excessive activation of theimmune system by bacteria (Dziarski and Gupta, 2006).
The serine proteases and serpins constitute the most abundant group of genes (13genes) that were influenced by injections (Table 2). When injected with Micrococcusluteus and Escherichia coli, P. xylostella also mounted the most abundant and varied genesof proteases and protease inhibitors against the challenges (Eum et al., 2007). Serineproteases and serpins play an important role in the insect immune response. Bothmelanotic encapsulation and the production of antimicrobial peptides are regulated byserine protease and serpins (Gorman et al., 2000). When challenged by injury ormicrobial infection, the cascades of serine proteases activate the coagulation andmelanization. However, unregulated blood clotting and complement can be harmful tothe insect. For this reason, it becomes necessary that the proteases in the system shouldbe tightly regulated by serpins (Kanost, 1999). Both proteases and the inhibitors areengaged in this precisely regulated system, and maybe that is why so many serineproteases and serpins were triggered by the stimulations.
28 � Archives of Insect Biochemistry and Physiology, September 2009
Archives of Insect Biochemistry and Physiology

More serine proteases and serpins were up-regulated by PBS injection than otherchallenges (Table 2). Injury itself is a minimal and sufficient single signal to cause somelevel of gene induction or prime the innate immune response (Markus et al., 2005).Compared with a simple mechanical injury, injections with pathogens may trigger amore complex resistant system, and maybe involve some differentially expressedgenes that are still functionally unknown (Table 3). Any increase in immunecompetence always comes at the expense of other fitness-determining traits (McKeanet al., 2008). A likely explanation for the above observation is that the silkwormsinjected with pathogens may deploy less serine proteases and serpins in order toconserve energy and other resources to deal with the pathogens.
Melanin biosynthesis is a hemocyte-mediated immune response that involves acomplex yet well-characterized cascade of reactions beginning with tyrosine andending with the polymerization of a capsule that surrounds an invading parasite (Zhaoet al., 1995; Beerntsen et al., 2000), eventually killing the parasite (Nappi andOttaviani, 2000). Ultrastructural studies demonstrate that this response is also rapidlydeployed against bacteria (Hillyer et al., 2003). Melanization-related enzymes,including PPAE and PAH, were detected in our study, but no other enzymes, suchas PPO, PPO-2 (Taft et al., 2001), PPO-3 (Genbank accession number: AF310673),PPO-4 (AF327409), dopa decarboxylase (DDC) (Ferdig et al., 1996), and dopachromeconversion enzyme (DCE) (Johnson et al., 2003) were detected. Similar results alsoappeared in microarray anaylsis of haemocytes of mosquito, A. aegypti, inoculated withbacteria, in which PAH is the only melanization-related immune gene that responds tothe challenge (Bartholomay et al., 2007). In mosquito, no change in transcriptabundance of phenoloxidase (PO) was detected in the whole body adult A. aegyptifollowing exposure to bacteria (Taft et al., 2001). Because DDC is primarily detected inovaries and DCE is constitutively expressed in adult mosquitoes (Johnson et al., 2001),it was not surprising that no change in the transcript abundance was detected. Besidesplaying a role in defense against pathogen infection and wound healing, themelanization-related genes, such as PPO, DDC, and DCE, also have diverse biologicalactivities in cuticle sclerotization, neurotransmission, reproduction, and a variety oftanning reactions required for metamorphosis (Ferdig et al., 2000). These genes wouldhave a relatively high expression level when compared to PAH, which participates inthe rate-limiting step (Johnson et al., 2003), and a further increase of the concentrationbecame unnecessary (Zou et al., 2005).
AMPs are an important factor in insect immunity to combat invadingmicroorganisms (Cheng et al., 2006). Cecropin is an alkaline peptide resistingGram-negative and Gram-positive bacteria and fungi, but acts on Gram-negativebacteria most effectively (Cheng et al., 2006). Moricin has antibacterial activity againstseveral Gram-negative and -positive bacteria, including S. aureus. Attacin and lebocinshow activity against Gram-negative bacteria. Gloverin 1, 2, and 3 genes were activatedby both Gram-positive and Gram-negative bacteria (Kaneko et al., 2007). Thus, ingeneral, expression levels of these genes in this study correspond well with the type ofbacterium used in an immune challenge. Unlike previously published results, attacin,and lebocin 3 showed weaker inducement by Gram-positive bacteria and fungus,gloverin was also induced by fungus, and gloverin 3 was not up-regulated by S. aureusinjection. One lysozyme (sw13847) was up-regulated by S. marcescen and B. bassianachallenges. The lysozyme gene is constitutively expressed at a low level in the fat body,hemocytes, and epidermal cells, and is strongly expressed upon bacterial infection(Yamakawa and Tanaka, 1999). In addition, lysozyme is considered to contribute
Gene Expression Profiling in B. mori � 29
Archives of Insect Biochemistry and Physiology

significantly to the production of soluble peptidoglycan, a strong inducer ofantibacterial proteins (Dunn et al., 1985).
Interestingly, JHBP0303 was found to be greatly induced after all four injections.This gene, originally named as e96h-0303 of the family, was found in the expressedsequence tags generated from eggs of B. mori (Saito et al., 2006). The expressionpattern of this gene at other developmental stages and/or tissues is not known. It is notclear whether its product is capable of binding to juvenile hormone, although itspredicted products contain the two conserved cysteine residues essential for binding.Exposure to stress leads to increased levels of the juvenile hormone (JH), which istransported to target tissue by a high-affinity binding protein, hemolymph JH bindingprotein (hJHBP) (Tauchman et al., 2007). JH was found to be a negative regulator ofA. aegypti transferrin, which is characterized as an immune protein due to up-regulation after immune challenges in mosquitoes, Drosophila, moths, termites, andhoneybees (Harizanova et al., 2005). In this study, the high expression of the JHBPfamily gene may reflect the need to transport JH to target tissues, or to reduce thenegative effects on immunity caused by the excessive unbound JH (Orth et al., 2003).In the same way, in order to counter the JH effects (Liu et al., 2008), JHE precursorwas induced after B. bassiana and PBS injections.
In conclusion, our work narrowed down the immune genes to a limited number,and this will lead to a better understanding of the immune response in the silkworm.Although a lot of work has been done on insect immunity, the functions of the genes,significantly up- or down-regulated in response to a foreign invader, still requirefurther investigations.
ACKNOWLEDGMENTS
We thank Chenglong Wang for advice on the experiment, Pingsheng Cao for thesupply of silkworms, Yongqi Sang for the data analysis, and Drs. Liangbiao Zheng,Chengshu Wang, and Yongping Huang for comments and discussions on thiswork.
LITERATURE CITED
Barat-Houari M, Hilliou F, Jousset FX, Sofer L, Deleury E, Rocher J, Ravallec M, Galibert L,Delobel P, Feyereisen R, Fournier P, Volkoh AN. 2006. Gene expression profiling ofSpodoptera frugiperda hemocytes and fat body using cDNA microarray reveals polydnavirus-associated variations in lepidopteran host genes transcript levels. BMC Genomics 7:160.
Bartholomay LC, Cho WL, Rocheleau TA, Boyle JP, Beck ET, Fuchs JF, Liss P, Rusch M,Butler KM, Wu RC, Lin SP, Kuo HY, Tsao IY, Huang CY, Liu TT, Hsiao KJ, Tsai SF, YangUC, Nappi AJ, Perna NT, Chen CC, Christensen BM. 2004. Description of thetranscriptomes of immune response-activated hemocytes from the mosquito vectors Aedesaegypti and Armigeres subalbatus. Infect Immun 72:4114–4126.
Bartholomay LC, Mayhew GF, Fuchs JF, Rocheleau TA, Erickson SM, Aliota MT, Christensen BM.2007. Profiling infection responses in the haemocytes of the mosquito, Aedes aegypti. Insect MolBiol 16:761–776.
Beerntsen BT, James AA, Christensen BM. 2000. Genetics of mosquito vector competence.Microbiol Mol Biol Rev 64:115–137.
30 � Archives of Insect Biochemistry and Physiology, September 2009
Archives of Insect Biochemistry and Physiology

Cheng T, Zhao P, Liu C, Xu P, Gao Z, Xia Q, Xiang Z. 2006. Structures, regulatory regions, andinductive expression patterns of antimicrobial peptide genes in the silkworm Bombyx mori.Genomics 87:356–365.
Cheng TC, Zhang YL, Liu C, Xu PZ, Gao ZH, Xia QY, Xiang ZH. 2008. Identification andanalysis of Toll-related genes in the domesticated silkworm, Bombyx mori. Dev CompImmunol 32:464–475.
De Gregorio E, Spellman PT, Rubin GM, Lemaitre B. 2001. Genome-wide analysis of theDrosophila immune response by using oligonucleotide microarrays. Proc Natl Acad Sci USA98:12590–12595.
Dimopoulos G, Seeley D, Wolf A, Kafatos FC. 1998. Malaria infection of the mosquito Anophelesgambiae activates immune-responsive genes during critical transition stages of the parasitelife cycle. Embo J 17:6115–6123.
Dunn PE, Dai W, Kanost MR, Geng CX. 1985. Soluble peptidoglycan fragments stimulateantibacterial protein synthesis by fat body from larvae of Manduca sexta. Dev CompImmunol 9:559–568.
Dziarski R, Gupta D. 2006. The peptidoglycan recognition proteins (PGRPs). Genome Biol7:232.
Engstrom Y, Loseva O, Theopold U. 2004. Proteomics of the Drosophila immune response.Trends Biotechnol 22:600–605.
Eum JH, Seo YR, Yoe SM, Kang SW, Han SS. 2007. Analysis of the immune-inducible genes ofPlutella xylostella using expressed sequence tags and cDNA microarray. Dev Comp Immunol31:1107–1120.
Ferdig MT, Li J, Severson DW, Christensen BM. 1996. Mosquito dopa decarboxylasecDNA characterization and blood-meal-induced ovarian expression. Insect Mol Biol5:119–126.
Ferdig MT, Taft AS, Smartt CT, Lowenberger CA, Li J, Zhang J, Christensen BM. 2000.Aedes aegypti dopa decarboxylase: gene structure and regulation. Insect Mol Biol 9:231–239.
Ferrandon D, Imler JL, Hetru C, Hoffmann JA. 2007. The Drosophila systemic immuneresponse: sensing and signalling during bacterial and fungal infections. Nat Rev Immunol7:862–874.
Gandhe AS, Arunkumar KP, John SH, Nagaraju J. 2006. Analysis of bacteria-challenged wildsilkmoth, Antheraea mylitta (lepidoptera) transcriptome reveals potential immune genes.BMC Genomics 7:184.
Gorman MJ, Andreeva OV, Paskewitz SM. 2000. Molecular characterization of five serine proteasegenes cloned from Anopheles gambiae hemolymph. Insect Biochem Mol Biol 30:35–46.
Harizanova N, Georgieva T, Dunkov BC, Yoshiga T, Law JH. 2005. Aedes aegypti transferrin.Gene structure, expression pattern, and regulation. Insect Mol Biol 14:79–88.
Hillyer JF, Schmidt SL, Christensen BM. 2003. Rapid phagocytosis and melanization of bacteriaand Plasmodium sporozoites by hemocytes of the mosquito Aedes aegypti. J Parasitol89:62–69.
Irving P, Troxler L, Heuer TS, Belvin M, Kopczynski C, Reichhart JM, Hoffmann JA, Hetru C.2001. A genome-wide analysis of immune responses in Drosophila. Proc Natl Acad Sci USA98:15119–15124.
Johnson JK, Li J, Christensen BM. 2001. Cloning and characterization of a dopachromeconversion enzyme from the yellow fever mosquito, Aedes aegypti. Insect Biochem Mol Biol31:1125–1135.
Johnson JK, Rocheleau TA, Hillyer JF, Chen CC, Li J, Christensen BM. 2003. A potential rolefor phenylalanine hydroxylase in mosquito immune responses. Insect Biochem Mol Biol33:345–354.
Gene Expression Profiling in B. mori � 31
Archives of Insect Biochemistry and Physiology

Kaneko Y, Furukawa S, Tanaka H, Yamakawa M. 2007. Expression of antimicrobial peptidegenes encoding Enbocin and Gloverin isoforms in the silkworm, Bombyx mori. BiosciBiotechnol Biochem 71:2233–2241.
Kanost MR. 1999. Serine proteinase inhibitors in arthropod immunity. Dev Comp Immunol23:291–301.
Kanost MR, Jiang H, Yu XQ. 2004. Innate immune responses of a lepidopteran insect, Manducasexta. Immunol Rev 198:97–105.
Koo IC, Ohol YM, Wu P, Morisaki JH, Cox JS, Brown EJ. 2008. Role for lysosomal enzyme beta-hexosaminidase in the control of mycobacteria infection. Proc Natl Acad Sci USA 105:710–715.
Lee WJ, Lee JD, Kravchenko VV, Ulevitch RJ, Brey PT. 1996. Purification and molecularcloning of an inducible gram-negative bacteria-binding protein from the silkworm, Bombyxmori. Proc Natl Acad Sci USA 93:7888–7893.
Lemaitre B, Reichhart JM, Hoffmann JA. 1997. Drosophila host defense: differential induction ofantimicrobial peptide genes after infection by various classes of microorganisms. Proc NatlAcad Sci USA 94:14614–14619.
Liu Z, Li X, Prasifka JR, Jurenka R, Bonning BC. 2008. Overexpression of Drosophila juvenilehormone esterase binding protein results in anti-JH effects and reduced pheromoneabundance. Gen Comp Endocrinol 156:164–172.
Ma C, Kanost MR. 2000. A beta1,3-glucan recognition protein from an insect, Manduca sexta,agglutinates microorganisms and activates the phenoloxidase cascade. J Biol Chem275:7505–7514.
Markus R, Kurucz E, Rus F, Ando I. 2005. Sterile wounding is a minimal and sufficienttrigger for a cellular immune response in Drosophila melanogaster. Immunol Lett101:108–111.
McKean KA, Yourth CP, Lazzaro BP, Clark AG. 2008. The evolutionary costs of immunologicalmaintenance and deployment. BMC Evol Biol 8:76.
Mita K, Kasahara M, Sasaki S, Nagayasu Y, Yamada T, Kanamori H, Namiki N, Kitagawa M,Yamashita H, Yasukochi Y, Kadono-Okuda K, Yamamoto K, Ajimura M, Ravikumar G,Shimomura M, Nagamura Y, Shin-I T, Abe H, Shimada T, Morishita S, Sasaki T. 2004. Thegenome sequence of silkworm, Bombyx mori. DNA Res 11:27–35.
Mouches C, Magnin M, Berge JB, de Silvestri M, Beyssat V, Pasteur N, Georghiou GP. 1987.Overproduction of detoxifying esterases in organophosphate-resistant Culex mosquitoesand their presence in other insects. Proc Natl Acad Sci USA 84:2113–2116.
Nappi AJ, Ottaviani E. 2000. Cytotoxicity and cytotoxic molecules in invertebrates. Bioessays22:469–480.
Ochiai M, Ashida M. 1988. Purification of a beta-1,3-glucan recognition protein in theprophenoloxidase activating system from hemolymph of the silkworm, Bombyx mori. J BiolChem 263:12056–12062.
Orth AP, Tauchman SJ, Doll SC, Goodman WG. 2003. Embryonic expression of juvenilehormone binding protein and its relationship to the toxic effects of juvenile hormone inManduca sexta. Insect Biochem Mol Biol 33:1275–1284.
Ramet M, Pearson A, Manfruelli P, Li X, Koziel H, Gobel V, Chung E, Krieger M, Ezekowitz RA.2001. Drosophila scavenger receptor CI is a pattern recognition receptor for bacteria.Immunity 15:1027–1038.
Saito K, Su ZH, Emi A, Mita K, Takeda M, Fujiwara Y. 2006. Cloning and expression analysis oftakeout/JHBP family genes of silkworm, Bombyx mori. Insect Mol Biol 15:245–251.
Shin SW, Park SS, Park DS, Kim MG, Kim SC, Brey PT, Park HY. 1998. Isolation andcharacterization of immune-related genes from the fall webworm, Hyphantria cunea, usingPCR-based differential display and subtractive cloning. Insect Biochem Mol Biol28:827–837.
32 � Archives of Insect Biochemistry and Physiology, September 2009
Archives of Insect Biochemistry and Physiology

Taft AS, Chen CC, Li J, Christensen BM. 2001. Molecular cloning of two prophenoloxidasegenes from the mosquito Aedes aegypti. Insect Mol Biol 10:97–103.
Tauchman SJ, Lorch JM, Orth AP, Goodman WG. 2007. Effects of stress on the hemolymphjuvenile hormone binding protein titers of Manduca sexta. Insect Biochem Mol Biol37:847–854.
Tichopad A, Dilger M, Schwarz G, Pfaffl MW. 2003. Standardized determination of real-timePCR efficiency from a single reaction set-up. Nucleic Acids Res 31:e122.
Tusher VG, Tibshirani R, Chu G. 2001. Significance analysis of microarrays applied to theionizing radiation response. Proc Natl Acad Sci USA 98:5116–5121.
Vasta GR, Quesenberry M, Ahmed H, O’Leary N. 1999. C-type lectins and galectins mediateinnate and adaptive immune functions: their roles in the complement activation pathway.Dev Comp Immunol 23:401–420.
Wang Y, Zhang P, Fujii H, Banno Y, Yamamoto K, Aso Y. 2004. Proteomic studies oflipopolysaccharide-induced polypeptides in the silkworm, Bombyx mori. Biosci BiotechnolBiochem 68:1821–1823.
Xia Q, Cheng D, Duan J, Wang G, Cheng T, Zha X, Liu C, Zhao P, Dai F, Zhang Z, He N, ZhangL, Xiang Z. 2007. Microarray-based gene expression profiles in multiple tissues of thedomesticated silkworm, Bombyx mori. Genome Biol 8:R162.
Xia Q, Zhou Z, Lu C, Cheng D, Dai F, Li B, Zhao P, Zha X, Cheng T, Chai C, Pan G, Xu J, Liu C,Lin Y, Qian J, Hou Y, Wu Z, Li G, Pan M, Li C, Shen Y, Lan X, Yuan L, Li T, Xu H, Yang G,Wan Y, Zhu Y, Yu M, Shen W, Wu D, Xiang Z, Yu J, Wang J, Li R, Shi J, Li H, Li G, Su J,Wang X, Li G, Zhang Z, Wu Q, Li J, Zhang Q, Wei N, Xu J, Sun H, Dong L, Liu D, Zhao S,Zhao X, Meng Q, Lan F, Huang X, Li Y, Fang L, Li C, Li D, Sun Y, Zhang Z, Yang Z, HuangY, Xi Y, Qi Q, He D, Huang H, Zhang X, Wang Z, Li W, Cao Y, Yu Y, Yu H, Li J, Ye J, ChenH, Zhou Y, Liu B, Wang J, Ye J, Ji H, Li S, Ni P, Zhang J, Zhang Y, Zheng H, Mao B, WangW, Ye C, Li S, Wang J, Wong GK, Yang H; Biology Analysis Group. 2004. A draft sequencefor the genome of the domesticated silkworm (Bombyx mori). Science 306:1937–1940.
Yamakawa M, Tanaka H. 1999. Immune proteins and their gene expression in the silkworm,Bombyx mori. Dev Comp Immunol 23:281–289.
Yoshida H, Kinoshita K, Ashida M. 1996. Purification of a peptidoglycan recognition proteinfrom hemolymph of the silkworm, Bombyx mori. J Biol Chem 271:13854–13860.
Yoshida H, Ochiai M, Ashida M. 1986. Beta-1,3-glucan receptor and peptidoglycan receptor arepresent as separate entities within insect prophenoloxidase activating system. BiochemBiophys Res Commun 141:1177–1184.
Zhao X, Ferdig MT, Li J, Christensen BM. 1995. Biochemical pathway of melanoticencapsulation of Brugia malayi in the mosquito, Armigeres subalbatus. Dev Comp Immunol19:205–215.
Zou Z, Wang Y, Jiang H. 2005. Manduca sexta prophenoloxidase activating proteinase-1 (PAP-1)gene: organization, expression, and regulation by immune and hormonal signals. InsectBiochem Mol Biol 35:627–636.
Gene Expression Profiling in B. mori � 33
Archives of Insect Biochemistry and Physiology





























