Embed Size (px)
Citation preview

GAS2L3, a target gene of the DREAM complex, isrequired for proper cytokinesis and genomic stability
Patrick Wolter1,*, Kathrin Schmitt1,*, Marc Fackler1,*, Heidi Kremling1, Leona Probst1, Stefanie Hauser1,Oliver J. Gruss2 and Stefan Gaubatz1,`
1Biozentrum, Universitat Wurzburg, Am Hubland, 97074 Wurzburg, Germany2Zentrum fur Molekulare Biologie der Universitat Heidelberg (ZMBH), DKFZ-ZMBH Alliance, Im Neuenheimer Feld 282, 69120 Heidelberg, Germany
*These authors contributed equally to this work`Author for correspondence ([email protected])
Accepted 2 January 2012Journal of Cell Science 125, 2393–2406� 2012. Published by The Company of Biologists Ltddoi: 10.1242/jcs.097253
SummaryThe mammalian DREAM complex is a key regulator of cell-cycle-regulated gene transcription and drives the expression of many gene
products required for mitosis and cytokinesis. In this study, we characterized GAS2L3, which belongs to the GAS2 family of proteinswith putative actin- and microtubule-binding domains as a target gene of DREAM. We found that GAS2L3 localizes to the spindlemidzone and the midbody during anaphase and cytokinesis, respectively. Biochemical studies show that GAS2L3 binds to and bundles
microtubules as well as F-actin in vitro. Strikingly, the RNAi-mediated knockdown of GAS2L3 results in chromosome segregationdefects in multinucleated cells and in cells with multi-lobed nuclei. Likewise, chronic downregulation of GAS2L3 causes chromosomeloss and aneuploidy. Time-lapse videomicroscopy experiments in GAS2L3-knockdown cells reveal abnormal oscillation of chromatin
and the spindle during cytokinesis. Taken together, our data reveal novel, important roles of GAS2L3 for faithful cell division. Our workthus contributes to the understanding of how DREAM regulates cytokinesis.
IntroductionCorrect progression through the cell cycle is essential for normal
development and differentiation. Its deregulation is associated
with the loss of genomic integrity and can contribute to
tumorigenesis. E2F transcription factors regulate the expression
of a large number of genes whose products play key roles in cell
cycle progression, synthesis of nucleotides, DNA replication and
apoptosis (Dimova and Dyson, 2005; van den Heuvel and Dyson,
2008). In the G1 phase of the cell cycle, E2F activity is regulated
by the binding to pRB, the product of the retinoblastoma tumor
suppressor gene, and by binding to two related ‘pocket proteins’
p107 and p130 (Burkhart and Sage, 2008; Cobrinik, 2005). Free,
uncomplexed E2F proteins function as transcriptional activators
with growth-promoting activities. By contrast, complexes
between E2F and pocket proteins act as active transcriptional
repressors and growth inhibitors (Trimarchi and Lees, 2002).
Importantly, it is believed that the deregulation of the pRB–E2F
pathway is involved in the pathogenesis of almost all human
tumors (Burkhart and Sage, 2008).
DREAM or LINC is a recently identified E2F pocket protein
complex in mammalian cells that undergoes a cell-cycle-dependent
switch of subunits (Litovchick et al., 2007; Schmit et al., 2007;
Pilkinton et al., 2007a). DREAM consists of the core subunits LIN9,
LIN37, LIN52, LIN54 and RbAp48. In quiescent cells, the core
complex is associated with p130 and E2F4 and contributes to the
repression of E2F-regulated genes. In late S phase, the interaction of
DREAM with p130–E2F4 is lost and DREAM then binds to the B-
MYB transcription factor. Genome-wide expression studies have
shown that DREAM–B-MYB is required for activation of a cluster
of genes required for entry into mitosis, spindle assembly and
cytokinesis (Osterloh et al., 2007; Knight et al., 2009; Reichert et al.,
2010; Schmit et al., 2009; Pilkinton et al., 2007b). Chromatin
immunoprecipitation experiments demonstrated that many of these
genes, such as Plk1 (polo-like kinase 1), Ccnb1 (cyclin B1) and
Kif20a, are direct target genes of DREAM (Osterloh et al., 2007;
Schmit et al., 2009).
One of the most prominent phenotypes after deletion of the
LIN9 subunit of DREAM in mouse embryonic fibroblasts is
cytokinesis failure, which results in binuclear cells (Reichert et al.,
2010). Similarly, in HeLa cells, RNAi-mediated silencing of
LIN54, another core subunit of DREAM, also causes cytokinesis
defects (Kittler et al., 2007). Cytokinesis is the final stage of cell
division after duplication and segregation of the genetic material
(Eggert et al., 2006). It is highly regulated to avoid unequal
chromosome segregation, which can result in aneuploidy and
tumor formation (Sagona and Stenmark, 2010). During
cytokinesis, a contractile actomyosin ring assembles at the cell
equator. Constriction of the cell membrane by the contractile ring
results in formation of the cleavage furrow. Subsequent membrane
fusion physically separates the cell into two daughter cells.
Formation of the contractile ring is controlled by the RhoA
GTPase (Narumiya and Yasuda, 2006; Piekny et al., 2005). The
centralspindlin complex, a component of the central spindle that
is formed by microtubule bundles between the segregating
chromosomes, recruits the Rho guanine nucleotide exchange
factor (GEF) Ect2 to activate RhoA at the overlying equatorial
cortex, which in turn activates the contractile ring and leads to
cleavage furrow ingression. Several mitotic kinases such as cyclin-
dependent kinases (CDKs), PLK1 and Aurora B also regulate
RhoA activation and cytokinesis. In addition, scaffolding proteins
such as anillin and septins play important roles in stabilizing
the cleavage furrow (Hickson and O’Farrell, 2008; Piekny and
Research Article 2393
Journ
alof
Cell
Scie
nce
Maddox, 2010). Although a large number of proteins involved in
cytokinesis have been identified in RNAi screens and through
proteomic studies, the molecular requirements for positioning and
stabilization of the contractile ring are not fully understood.
In this study, we characterized GAS2L3, a target gene of
DREAM and a member of the GAS2 protein family. We find that
GAS2L3, unlike other members of the GAS2 family is expressed
in mitosis and that it localizes the midbody during cytokinesis.
Biochemical studies indicate that GAS2L3 can bind to and
crosslink microtubule and actin filaments. Using RNAi we find
that loss of GAS2L3 leads to abnormal oscillation of chromatin
and the spindle during cytokinesis. Finally, we demonstrate
that RNAi-mediated depletion of GAS2L3 leads to genomic
instability. Thus we have identified GAS2L3 as an important
target gene of the DREAM complex that is required for proper
cytokinesis.
ResultsGAS2L3 is a DREAM target gene that is highly expressed
in G2 and mitosis
Transcriptional profiling of conditional LIN9-knockout MEFs
using Agilent DNA microarrays identified GAS2L3 as a target
gene of the mammalian DREAM complex (Reichert et al., 2010).
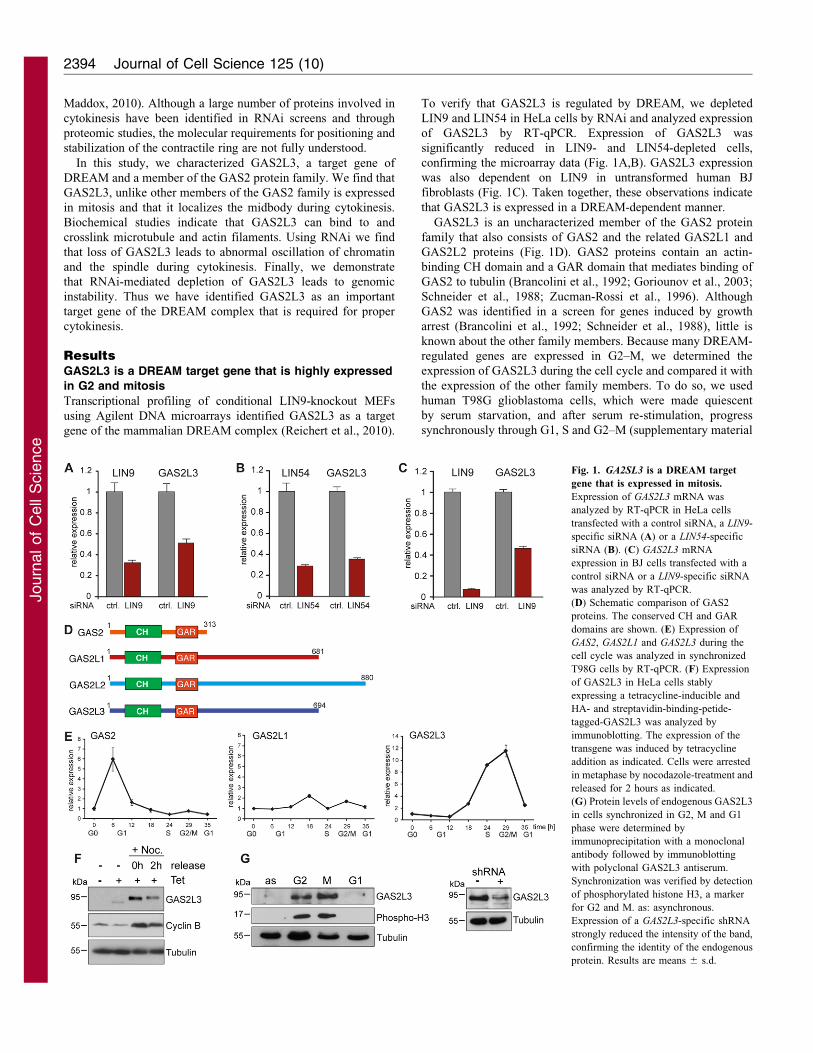
To verify that GAS2L3 is regulated by DREAM, we depleted
LIN9 and LIN54 in HeLa cells by RNAi and analyzed expression
of GAS2L3 by RT-qPCR. Expression of GAS2L3 was
significantly reduced in LIN9- and LIN54-depleted cells,
confirming the microarray data (Fig. 1A,B). GAS2L3 expression
was also dependent on LIN9 in untransformed human BJ
fibroblasts (Fig. 1C). Taken together, these observations indicate
that GAS2L3 is expressed in a DREAM-dependent manner.
GAS2L3 is an uncharacterized member of the GAS2 protein
family that also consists of GAS2 and the related GAS2L1 and
GAS2L2 proteins (Fig. 1D). GAS2 proteins contain an actin-
binding CH domain and a GAR domain that mediates binding of
GAS2 to tubulin (Brancolini et al., 1992; Goriounov et al., 2003;
Schneider et al., 1988; Zucman-Rossi et al., 1996). Although
GAS2 was identified in a screen for genes induced by growth
arrest (Brancolini et al., 1992; Schneider et al., 1988), little is
known about the other family members. Because many DREAM-
regulated genes are expressed in G2–M, we determined the
expression of GAS2L3 during the cell cycle and compared it with
the expression of the other family members. To do so, we used
human T98G glioblastoma cells, which were made quiescent
by serum starvation, and after serum re-stimulation, progress
synchronously through G1, S and G2–M (supplementary material
Fig. 1. GA2SL3 is a DREAM target
gene that is expressed in mitosis.
Expression of GAS2L3 mRNA was
analyzed by RT-qPCR in HeLa cells
transfected with a control siRNA, a LIN9-
specific siRNA (A) or a LIN54-specific
siRNA (B). (C) GAS2L3 mRNA
expression in BJ cells transfected with a
control siRNA or a LIN9-specific siRNA
was analyzed by RT-qPCR.
(D) Schematic comparison of GAS2
proteins. The conserved CH and GAR
domains are shown. (E) Expression of
GAS2, GAS2L1 and GAS2L3 during the
cell cycle was analyzed in synchronized
T98G cells by RT-qPCR. (F) Expression
of GAS2L3 in HeLa cells stably
expressing a tetracycline-inducible and
HA- and streptavidin-binding-petide-
tagged-GAS2L3 was analyzed by
immunoblotting. The expression of the
transgene was induced by tetracycline
addition as indicated. Cells were arrested
in metaphase by nocodazole-treatment and
released for 2 hours as indicated.
(G) Protein levels of endogenous GAS2L3
in cells synchronized in G2, M and G1
phase were determined by
immunoprecipitation with a monoclonal
antibody followed by immunoblotting
with polyclonal GAS2L3 antiserum.
Synchronization was verified by detection
of phosphorylated histone H3, a marker
for G2 and M. as: asynchronous.
Expression of a GAS2L3-specific shRNA
strongly reduced the intensity of the band,
confirming the identity of the endogenous
protein. Results are means 6 s.d.
Journal of Cell Science 125 (10)2394
Journ
alof
Cell
Scie
nce
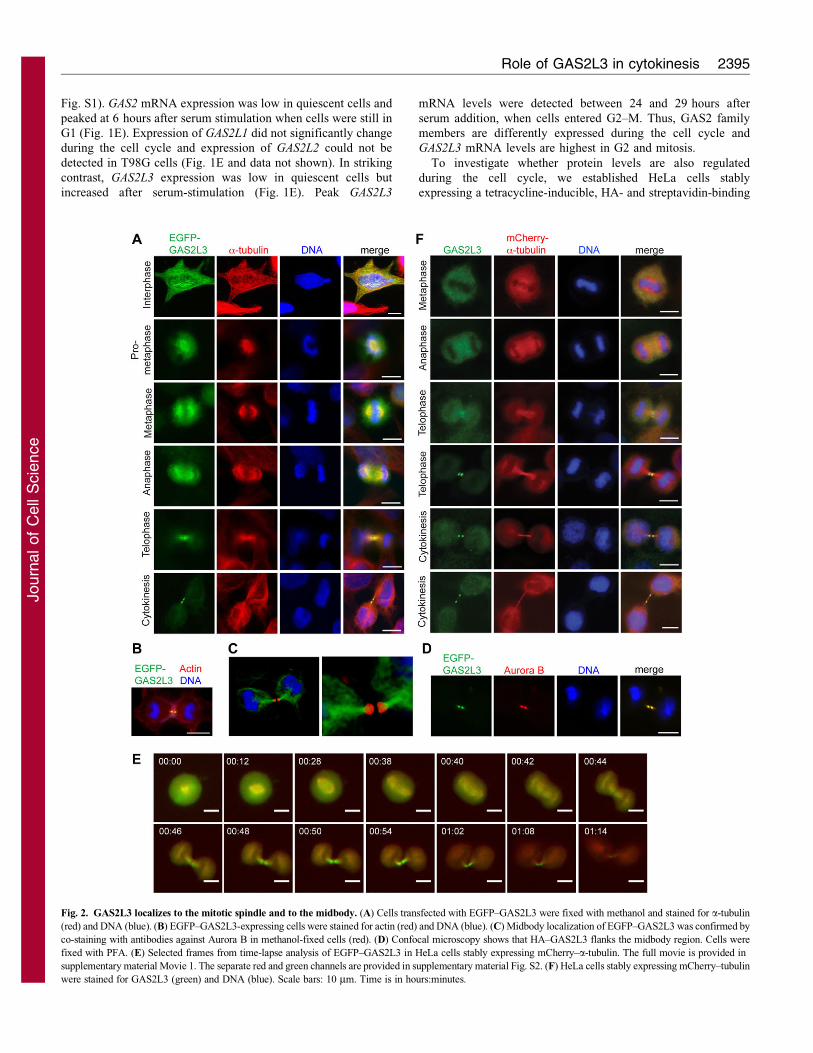
Fig. S1). GAS2 mRNA expression was low in quiescent cells and
peaked at 6 hours after serum stimulation when cells were still in
G1 (Fig. 1E). Expression of GAS2L1 did not significantly change
during the cell cycle and expression of GAS2L2 could not be
detected in T98G cells (Fig. 1E and data not shown). In striking
contrast, GAS2L3 expression was low in quiescent cells but
increased after serum-stimulation (Fig. 1E). Peak GAS2L3
mRNA levels were detected between 24 and 29 hours after
serum addition, when cells entered G2–M. Thus, GAS2 family
members are differently expressed during the cell cycle and
GAS2L3 mRNA levels are highest in G2 and mitosis.
To investigate whether protein levels are also regulated
during the cell cycle, we established HeLa cells stably
expressing a tetracycline-inducible, HA- and streptavidin-binding
Fig. 2. GAS2L3 localizes to the mitotic spindle and to the midbody. (A) Cells transfected with EGFP–GAS2L3 were fixed with methanol and stained for a-tubulin
(red) and DNA (blue). (B) EGFP–GAS2L3-expressing cells were stained for actin (red) and DNA (blue). (C) Midbody localization of EGFP–GAS2L3 was confirmed by
co-staining with antibodies against Aurora B in methanol-fixed cells (red). (D) Confocal microscopy shows that HA–GAS2L3 flanks the midbody region. Cells were
fixed with PFA. (E) Selected frames from time-lapse analysis of EGFP–GAS2L3 in HeLa cells stably expressing mCherry–a-tubulin. The full movie is provided in
supplementary material Movie 1. The separate red and green channels are provided in supplementary material Fig. S2. (F) HeLa cells stably expressing mCherry–tubulin
were stained for GAS2L3 (green) and DNA (blue). Scale bars: 10 mm. Time is in hours:minutes.
Role of GAS2L3 in cytokinesis 2395
Journ
alof
Cell
Scie
nce
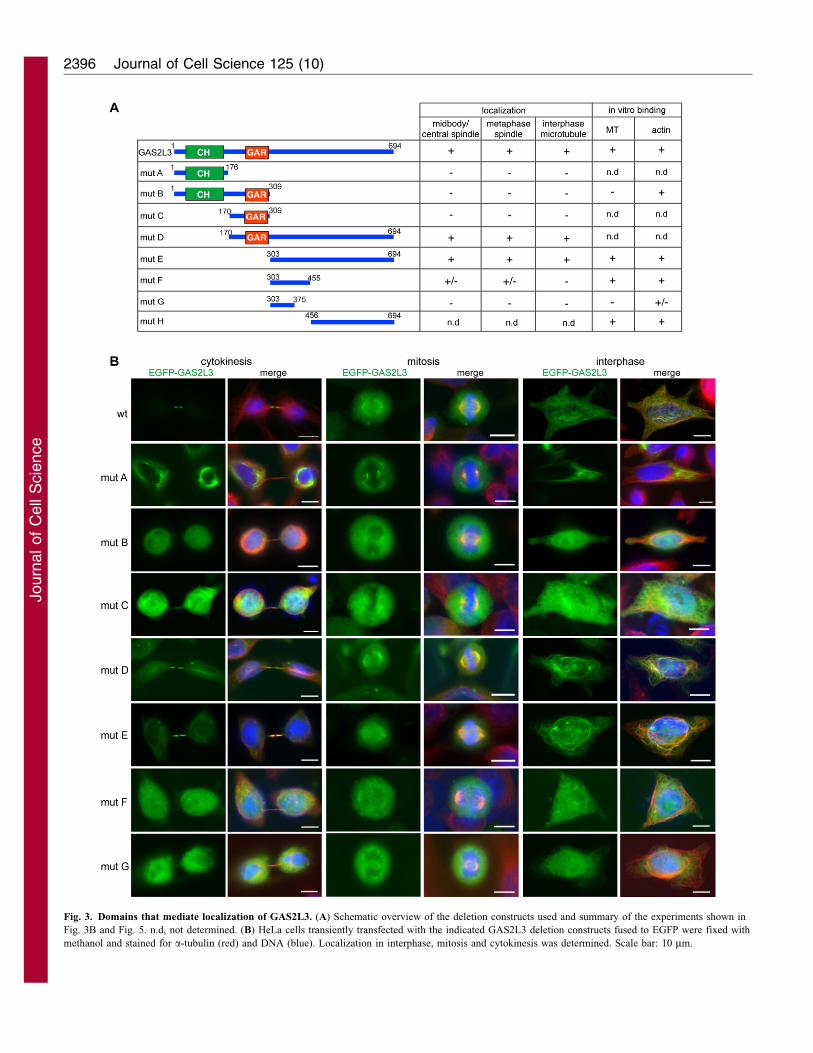
Fig. 3. Domains that mediate localization of GAS2L3. (A) Schematic overview of the deletion constructs used and summary of the experiments shown in
Fig. 3B and Fig. 5. n.d, not determined. (B) HeLa cells transiently transfected with the indicated GAS2L3 deletion constructs fused to EGFP were fixed with
methanol and stained for a-tubulin (red) and DNA (blue). Localization in interphase, mitosis and cytokinesis was determined. Scale bar: 10 mm.
Journal of Cell Science 125 (10)2396
Journ
alof
Cell
Scie
nce
peptide-tagged GAS2L3. Upon tetracycline induction, some
GAS2L3 protein could be detected in asynchronous cells, but
GAS2L3 protein levels were much higher in cells that were
arrested in mitosis by nocodazole (Fig. 1F). Upon exit from
mitosis, GAS2L3 protein levels decreased to a similar level as for
cyclin B1. We also noted a change in GAS2L3 mobility upon
nocodazole treatment, indicating that GAS2L3 is modified in
mitosis, possibly by phosphorylation. Endogenous GAS2L3
protein could only be detected in lysates of HeLa cells in G2
and M phase, suggesting that GAS2L3 is degraded after mitosis
(Fig. 1G).
Localization of GAS2L3 to the mitotic spindle and to
the midbody
The subcellular localization of transiently expressed EGFP–
GAS2L3 in HeLa cells was determined by fluorescence
microscopy. In interphase cells, EGFP–GAS2L3 was
cytoplasmic and localized to microtubules (Fig. 2A). In mitosis
EGFP–GAS2L3 was enriched at the mitotic spindle in prophase
and anaphase. In telophase EGFP–GAS2L3 was visible in the
spindle midzone. In cytokinesis EGFP–GAS2L3 was exclusively
detected at the midbody, a dense microtubule-rich region that
forms before abscission at the midpoint of the intercellular bridge.
GAS2L3 colocalized with a-tubulin, but not with actin at the
midbody (Fig. 2B). Midbody localization was confirmed by
colocalization of EGFP–GAS2L3 with Aurora B, a subunit of
the chromosomal passenger complex, which is known to localize
to the midbody (Fig. 2C). Confocal fluorescence microscopy of
HA-tagged mouse GAS2L3 revealed that GAS2L3 flanks the
midbody on both sides but is excluded from the central region of
the midbody (Fig. 2D). Live-cell imaging of EGFP–GAS2L3 in
HeLa cells stably expressing mCherry–a-tubulin confirmed the
localization of GAS2L3 observed in fixed cells, indicating that
localization of GAS2L3 is not an artifact of fixation (Fig. 2E;
supplementary material Movie 1 and Fig. S2).
The localization of endogenous GAS2L3 in relationship to
mCherry–a-tubulin and MKLP1, a subunit of centralspindlin
(Matuliene and Kuriyama, 2002; Mishima et al., 2002), was
determined by staining with a monoclonal antibody against
GAS2L3 (Fig. 2F; supplementary material Fig. S3). Endogenous
GAS2L3 localized to the spindle midzone and midbodies in
cytokinesis in HeLa and U2OS cells. In some cells, GAS2L3
also localized to the midbody ring that forms as a remnant of
the contractile ring around the midbody microtubules during
cytokinesis (supplementary material Fig. S4) (Gromley et al.,
2005; Pohl and Jentsch, 2008). Furthermore, in some interphase
cells, GAS2L3 was detected at midbody remnants in ring-like
structures (supplementary material Fig. S4). Co-staining with a-
tubulin showed that these ring-like structures were rich in
microtubules. Although spindle localization of endogenous
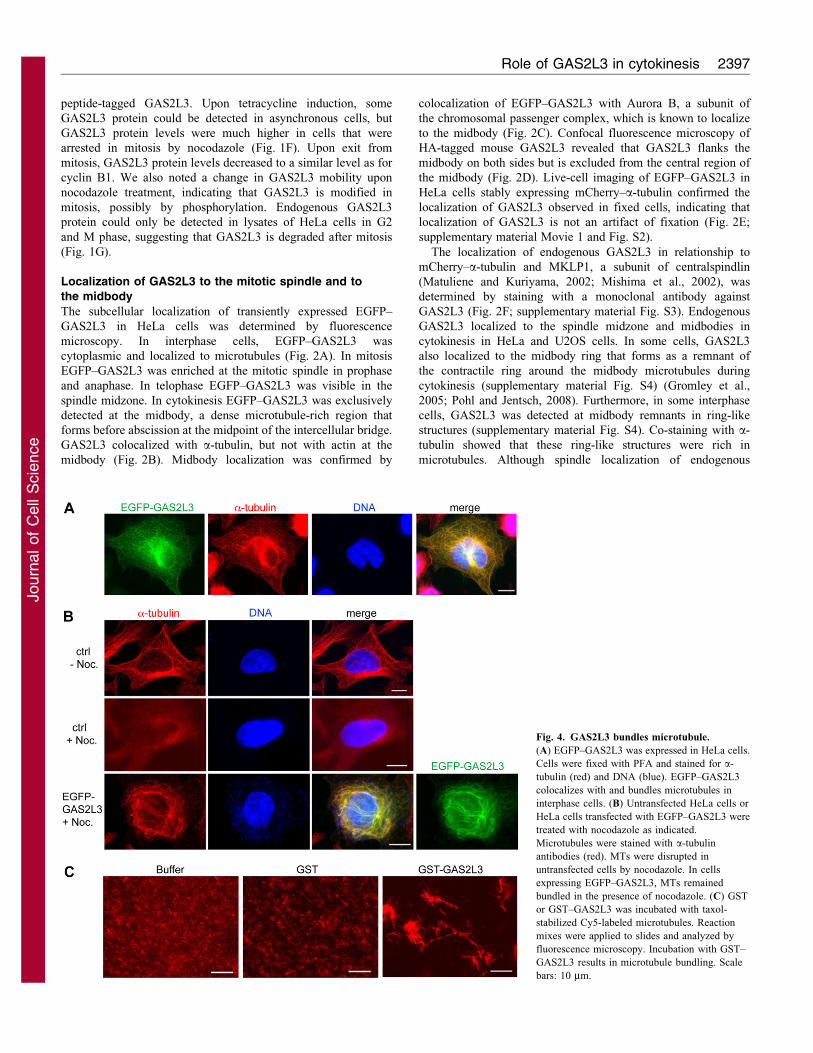
Fig. 4. GAS2L3 bundles microtubule.
(A) EGFP–GAS2L3 was expressed in HeLa cells.
Cells were fixed with PFA and stained for a-
tubulin (red) and DNA (blue). EGFP–GAS2L3
colocalizes with and bundles microtubules in
interphase cells. (B) Untransfected HeLa cells or
HeLa cells transfected with EGFP–GAS2L3 were
treated with nocodazole as indicated.
Microtubules were stained with a-tubulin
antibodies (red). MTs were disrupted in
untransfected cells by nocodazole. In cells
expressing EGFP–GAS2L3, MTs remained
bundled in the presence of nocodazole. (C) GST
or GST–GAS2L3 was incubated with taxol-
stabilized Cy5-labeled microtubules. Reaction
mixes were applied to slides and analyzed by
fluorescence microscopy. Incubation with GST–
GAS2L3 results in microtubule bundling. Scale
bars: 10 mm.
Role of GAS2L3 in cytokinesis 2397
Journ
alof
Cell
Scie
nce
GAS2L3 was not observed, this could be due to the difficulty in
detecting low amounts of the protein with the available
reagents.
Domains required for localization of GAS2L3
To investigate the role of the CH and GAR domains in
subcellular localization of the protein, we generated a set of
GAS2L3 deletion constructs (Fig. 3A). Deletion constructs were
transiently expressed as EGFP fusion proteins in HeLa cells and
their localization in interphase, mitosis and cytokinesis was
determined (Fig. 3B).
We found that mutants containing only the CH domain (mutant
A) or the CH and the GAR domain (mutant B) or the GAR
domain (mutant C) failed to colocalize with interphase
microtubule, mitotic spindles or spindle midzones. Instead, as
evidenced by Phalloidin co-staining, mutants with the CH
domain prominently induced and colocalized with actin stress
fibers (supplementary material Fig. S5), suggesting that the CH
domain can indeed function as an actin-interacting domain.
Mutants that contain the C-terminus but lack the CH domain
(mutant D) or the CH and the GAR domain (mutant E)
colocalized with microtubules in interphase and to metaphase
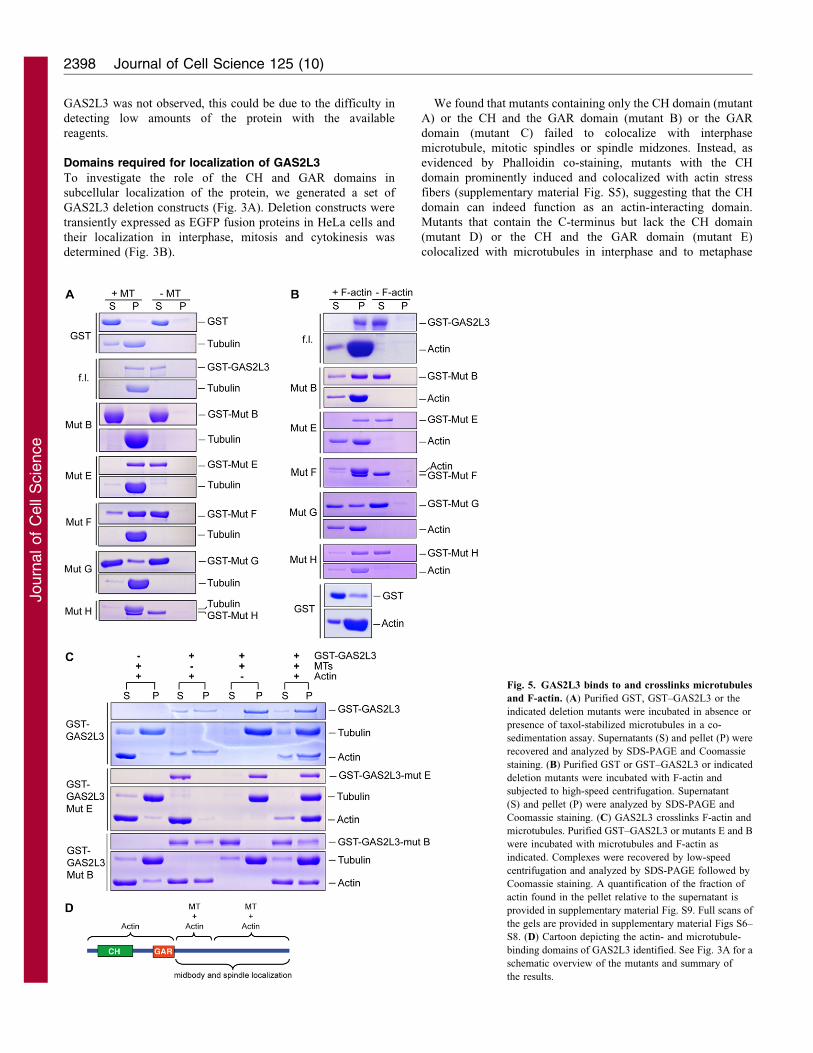
Fig. 5. GAS2L3 binds to and crosslinks microtubules
and F-actin. (A) Purified GST, GST–GAS2L3 or the
indicated deletion mutants were incubated in absence or
presence of taxol-stabilized microtubules in a co-
sedimentation assay. Supernatants (S) and pellet (P) were
recovered and analyzed by SDS-PAGE and Coomassie
staining. (B) Purified GST or GST–GAS2L3 or indicated
deletion mutants were incubated with F-actin and
subjected to high-speed centrifugation. Supernatant
(S) and pellet (P) were analyzed by SDS-PAGE and
Coomassie staining. (C) GAS2L3 crosslinks F-actin and
microtubules. Purified GST–GAS2L3 or mutants E and B
were incubated with microtubules and F-actin as
indicated. Complexes were recovered by low-speed
centrifugation and analyzed by SDS-PAGE followed by
Coomassie staining. A quantification of the fraction of
actin found in the pellet relative to the supernatant is
provided in supplementary material Fig. S9. Full scans of
the gels are provided in supplementary material Figs S6–
S8. (D) Cartoon depicting the actin- and microtubule-
binding domains of GAS2L3 identified. See Fig. 3A for a
schematic overview of the mutants and summary of
the results.
Journal of Cell Science 125 (10)2398
Journ
alof
Cell
Scie
nce
spindles, similar to full-length EGFP–GAS2L3 and consistent
with a recent study by Stroud and colleagues, who showed
that the C-terminus of GAS2L3 localizes to microtubules
in interphase (Stroud et al., 2011). These mutants also still
localized to the spindle midzone in cytokinesis; however, they
were not as sharply concentrated at the midbody as full-length
GAS2L3. Mutant F, which only contains part of the C-terminus,
weakly associated with mitotic spindles and with the central
spindle, but showed no colocalization with microtubules in
interphase. The smallest mutant (mutant G) showed no overlap
with microtubules.
Taken together, these results indicate that the C-terminus of
GAS2L3 is sufficient for microtubule colocalization and
localization to the spindle midzone, whereas the CH and GAR
domains are required for efficient midbody localization of
GAS2L3.
GAS2L3 stabilizes and bundles microtubule
EGFP–GAS2L3 appeared to bundle microtubules into rings
around the nucleus, indicative of hyperstable, buckling
microtubules (Fig. 4A). A similar rearrangement of microtubules
into perinuclear rings has been observed before, after
overexpression of microtubule-associated proteins such as PRC1
and Lis1 (Mollinari et al., 2002; Smith et al., 2000). To confirm
that GAS2L3 can stabilize microtubules in intact cells, we
expressed EGFP–GAS2L3 in HeLa cells and treated them with
low doses of nocodazole to depolymerize microtubules (Fig. 4B).
Strikingly, in cells that expressed high levels of EGFP–GAS2L3,
microtubules were resistant to depolymerization, indicating that
GAS2L3 can stabilize microtubules in vivo.
Next, to investigate whether GAS2L3 has microtubule-
bundling activity, taxol-stabilized microtubules were incubated
in vitro with GST or GST–GAS2L3 and investigated by
microscopy. In the presence of only GST, microtubules were
short and unbundled, as expected (Fig. 4C). By stark contrast,
GST–GAS2L3 induced a high degree of microtubule-bundling.
GAS2L3 is a microtubule- and actin-associated protein
We next determined the domains of GAS2L3 that mediate
direct binding to microtubules. To do so, we performed co-
sedimentation assays with in vitro polymerized microtubules and
purified recombinant GST–GAS2L3 wild-type or deletion
mutants. GST–GAS2L3, but not GST alone, co-sedimented
with microtubules, indicating that GAS2L3 can directly associate
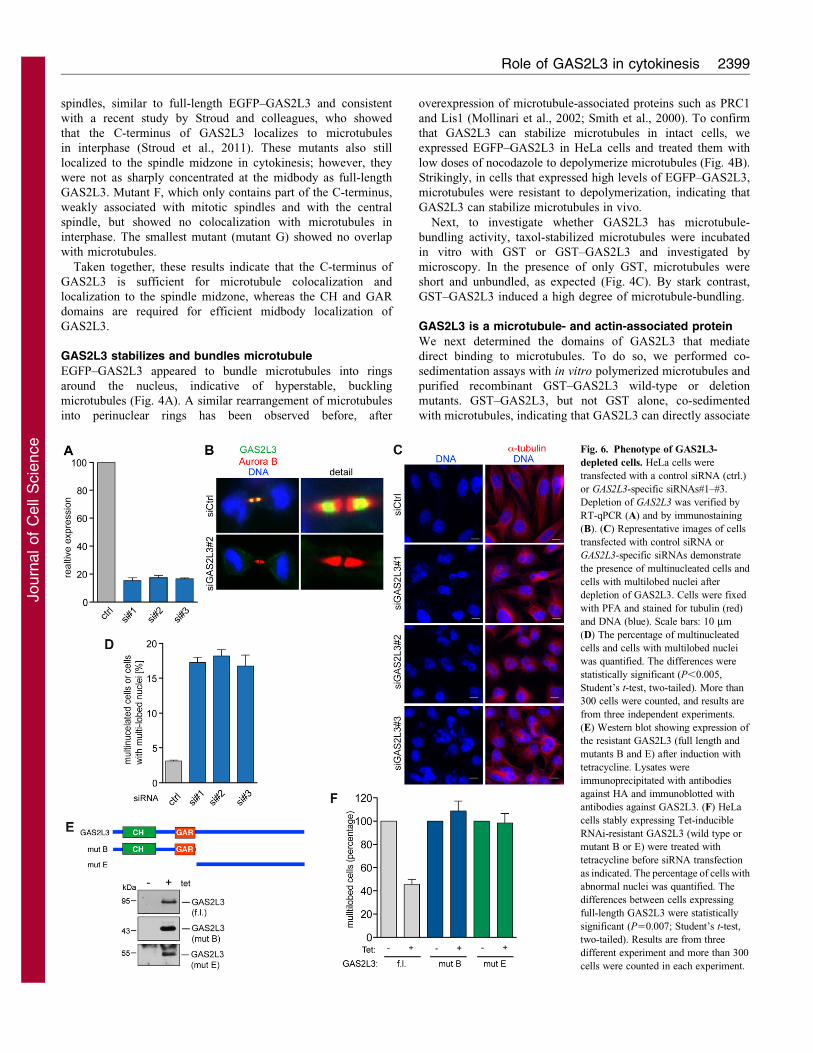
Fig. 6. Phenotype of GAS2L3-
depleted cells. HeLa cells were
transfected with a control siRNA (ctrl.)
or GAS2L3-specific siRNAs#1–#3.
Depletion of GAS2L3 was verified by
RT-qPCR (A) and by immunostaining
(B). (C) Representative images of cells
transfected with control siRNA or
GAS2L3-specific siRNAs demonstrate
the presence of multinucleated cells and
cells with multilobed nuclei after
depletion of GAS2L3. Cells were fixed
with PFA and stained for tubulin (red)
and DNA (blue). Scale bars: 10 mm
(D) The percentage of multinucleated
cells and cells with multilobed nuclei
was quantified. The differences were
statistically significant (P,0.005,
Student’s t-test, two-tailed). More than
300 cells were counted, and results are
from three independent experiments.
(E) Western blot showing expression of
the resistant GAS2L3 (full length and
mutants B and E) after induction with
tetracycline. Lysates were
immunoprecipitated with antibodies
against HA and immunoblotted with
antibodies against GAS2L3. (F) HeLa
cells stably expressing Tet-inducible
RNAi-resistant GAS2L3 (wild type or
mutant B or E) were treated with
tetracycline before siRNA transfection
as indicated. The percentage of cells with
abnormal nuclei was quantified. The
differences between cells expressing
full-length GAS2L3 were statistically
significant (P50.007; Student’s t-test,
two-tailed). Results are from three
different experiment and more than 300
cells were counted in each experiment.
Role of GAS2L3 in cytokinesis 2399
Journ
alof
Cell
Scie
nce
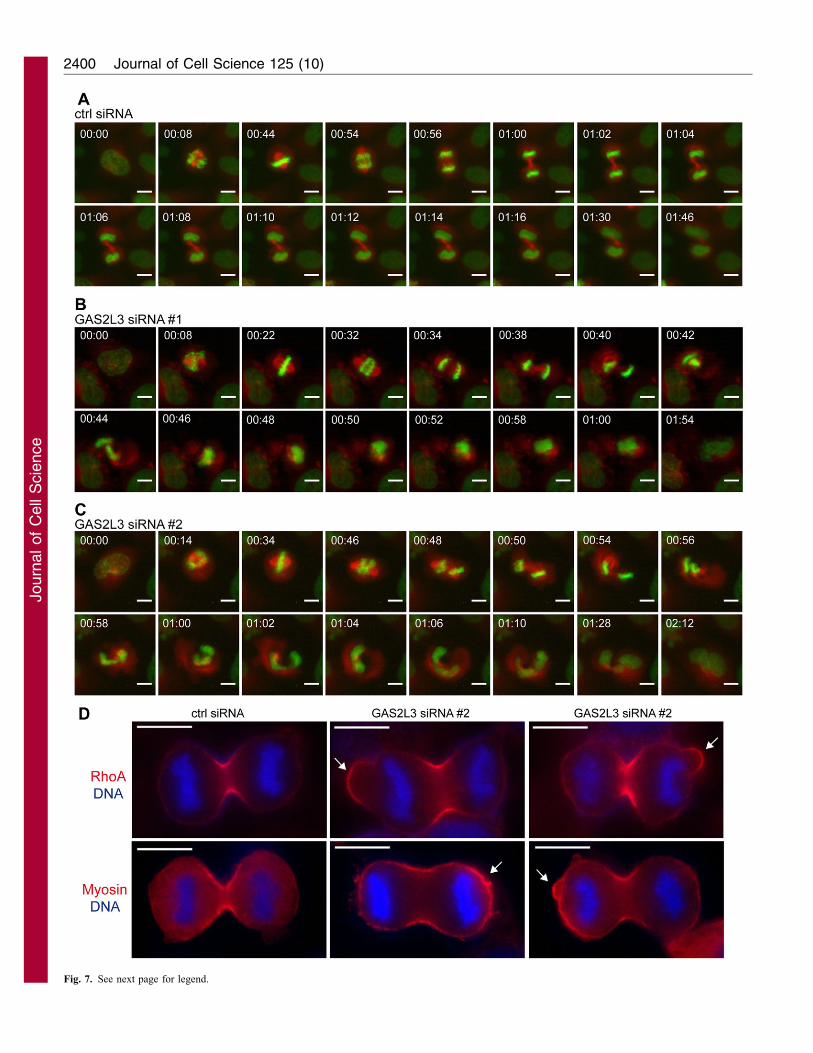
Fig. 7. See next page for legend.
Journal of Cell Science 125 (10)2400
Journ
alof
Cell
Scie
nce

with microtubules (Fig. 5A; supplementary material Fig. S6).
Whereas an N-terminal construct containing the CH and GAR
domain (mutant B) did not bind to microtubules, the C-terminus
(mutant E) was sufficient for this binding, consistent with the
localization data and with a previous study (Stroud et al., 2011).
The C-terminal domain of GAS2L3 contains several clusters
of positively charged amino acids that could mediate binding
to the negatively charged microtubules. Consistent with this
observation, two non-overlapping constructs, which both contain
basic clusters (mutant F and mutant H), independently associated
with microtubules. By contrast, a smaller construct (mutant G)
that contains fewer basic residues only very weakly associated
with microtubules. Therefore, whereas the GAR domain appears
not to function as microtubule-binding domain in this protein,
GAS2L3 appears to have several C-terminal microtubule-binding
domains.
Because GAS2L3 also has a putative actin-binding CH
domain, we performed F-actin co-sedimentation assays. GST
alone did not co-sediment with actin. In the absence of actin,
GST–GAS2L3 also remained in the supernatant (Fig. 5B,
supplementary material Fig. S7). However, when GST–
GAS2L3 was mixed with actin, it was found in the pellet
fraction, indicating that it can directly bind to F-actin. As
expected, the N-terminus of GAS2L3 containing the CH domain
(mutant B) was sufficient for binding to F-actin. Surprisingly, the
C-terminus lacking the CH domain (mutant E) efficiently
associated with actin as did the two non-overlapping C-
terminal parts tested (mutants F and H). Only the small C-
terminal mutant G associated with F-actin with reduced affinity.
We conclude that GAS2L3 has several actin-binding domains –
the CH domain and at least two different domains in the C-
terminus. The C-terminal F-actin-binding sites cannot be clearly
separated from the microtubule-binding domains.
As our data indicate that GAS2L3 has several microtubule-
and F-actin-binding domains, we next asked whether GAS2L3
can simultaneously bind to both microtubules and F-actin. This
possibility was addressed by low-speed centrifugation. When
microtubules and F-actin were centrifuged together, actin
remained in the supernatant after low-speed centrifugation
(Fig. 5C). When GAS2L3 was mixed with actin alone, a small
fraction of F-actin was found in the pellet, suggesting that
GAS2L3 has some actin-bundling activity. Importantly, when
wild-type GAS2L3, microtubules and F-actin were mixed, actin
bundles efficiently co-sedimented along with microtubules,
indicating that GAS2L3 can crosslink microtubules and F-actin
in vitro (Fig. 5C; supplementary material Fig. S8). Because
the N-terminus of GAS2L3 (mutant B) does not bind
to microtubules, it was unable to crosslink F-actin and
microtubules, although it appeared to have similar F-actin-
bundling activity as full-length GAS2L3. Interestingly, however,
the C-terminus of GAS2L3 (mutant E) was sufficient to crosslink
microtubules and F-actin, consistent with the observation that itcontains numerous F-actin- and microtubule-binding domains.
Taken together, our data indicate that GAS2L3 binds to andbundles F-actin and microtubules.
GAS2L3 is required for the completion of cytokinesis
The expression and localization of GAS2L3 suggests a role inmitosis or cytokinesis. To assess the in vivo role of GAS2L3, wedepleted GAS2L3 by RNA interference (RNAi) using three
different siRNAs. GAS2L3 mRNA levels were strongly reducedby all three siRNAs, as determined by RT-qPCR (Fig. 6A). Weconfirmed depletion of GAS2L3 to undetectable levels by
immunofluorescence on a single cell basis (Fig. 6B). All threesiRNAs caused a significant increase in multinucleated cells andcells with multilobed nuclei (Fig. 6C,D), suggesting defects inchromosome segregation and/or cytokinesis. The majority of
abnormal nuclei in GAS2L3-depleted cells were multilobed and asmall fraction (about 5%) of nuclei were binuclear. To confirmthat depletion of GAS2L3 caused the multilobed phenotype, we
performed rescue experiments with a siRNA-resistant GAS2L3construct that was created by introducing silent mutations into therecognition sequence for siRNA#2. GAS2L3 siRNA#2 reduced
expression of wild-type EGFP–GAS2L3 to undetectable levels,whereas the EGFP signal of the mutated construct was notreduced, confirming the resistance (supplementary material Fig.S10). HeLa cells stably expressing RNAi-resistant GAS2L3 were
cultured in the presence or absence of tetracycline and thentransfected with either control siRNA or GAS2L3-specificsiRNA#2. In the presence of tetracycline, all three proteins
were expressed (Fig. 6E). Expression of the RNAi-resistantGAS2L3 partially rescued the multilobed phenotype, indicatingthat the phenotype is indeed due to depletion of GAS2L3 and is
not an off-target effect (Fig. 6F). Neither the N-terminus (mutantB) nor the C-terminus (mutant E) of GAS2L3 was able to rescuethe multilobed phenotype. We conclude that both the N-terminus
and the C-terminal domains of GAS2L3 are crucial for properfunction of GAS2L3.
To investigate the role of GAS2L3 in mitosis and cytokinesis inmore detail, we performed time-lapse videomicroscopy in HeLa
cells engineered to stably express H2B–EGFP and a-tubulin–mRFP as markers for chromosomes and microtubules,respectively. Cells transfected with a control siRNA progressed
normally through mitosis and cytokinesis, as expected (Fig. 7A;supplementary material Movie 2). Cells depleted of endogenousGAS2L3 with either siRNA#1 or siRNA#2 formed a normal
mitotic spindle and progressed normally through metaphase andanaphase (Fig. 7B,C; supplementary material Movies 3,4). Intelophase, however, GAS2L3-depleted cells exhibited a spindle-rocking phenotype similar to the phenotype that has for example
been reported for cells depleted of anillin or containinghyperstabilized astral microtubules (Rankin and Wordeman,2010; Zhao and Fang, 2005). GAS2L3-depleted cells displayed
an oscillation of the entire spindle and the chromatin between theproto-daughter cells. Oscillations started just at the beginning ofcytokinesis, approximately 8 minutes after anaphase onset. In
oscillating cells, the chromatin and the spindle moved back andforth several times over a period of approximately 20 minutesbefore the separated sets of chromatin fell back into a single mass.
The oscillation phenotype was observed independently withsiRNA#1 in 45 of 214 cells (17.4%) and with siRNA#2 in 24 of143 cells (16.8%) but only in 1 out of 93 control-depleted cells
Fig. 7. GAS2L3 is required for cytokinesis. (A–C) Selected frames from
time-lapse movies of HeLa cells stably expressing H2B–EGFP and mRFP–a-
tubulin and transfected with a control (A) or GAS2L3-specific siRNA#1
(B) or siRNA#2 (C). Time relative to nuclear envelope breakdown is shown in
hours:minutes. Movies are provided in supplementary material Movies 2–4.
(D) Control and GAS2L3-knockdown cells were fixed with TCA and stained
for RhoA (red) and DNA (blue) or myosin IIA (red) and DNA (blue). Scale
bars: 10 mm. Arrows indicate abnormal cortical localization of RhoA or
myosin IIA.
Role of GAS2L3 in cytokinesis 2401
Journ
alof
Cell
Scie
nce
(1.1%). In addition, substantive membrane blebbing was observed
during anaphase in approximately 20% of GAS2L3-depleted cells
whereas less than 2% of control cells showed this phenotype (see
supplementary material Movies 3,4; Fig. 7D). In cells with large
blebs, RhoA localization was not restricted to the equator but was
detected at the blebs (Fig. 7D). Myosin IIA also accumulated at the
cortex in GAS2L3-depleted cells. Thus, GAS2L3 is required to
restrict RhoA and Myosin II to the cleavage furrow.
GAS2L3 protects cells from aneuploidy
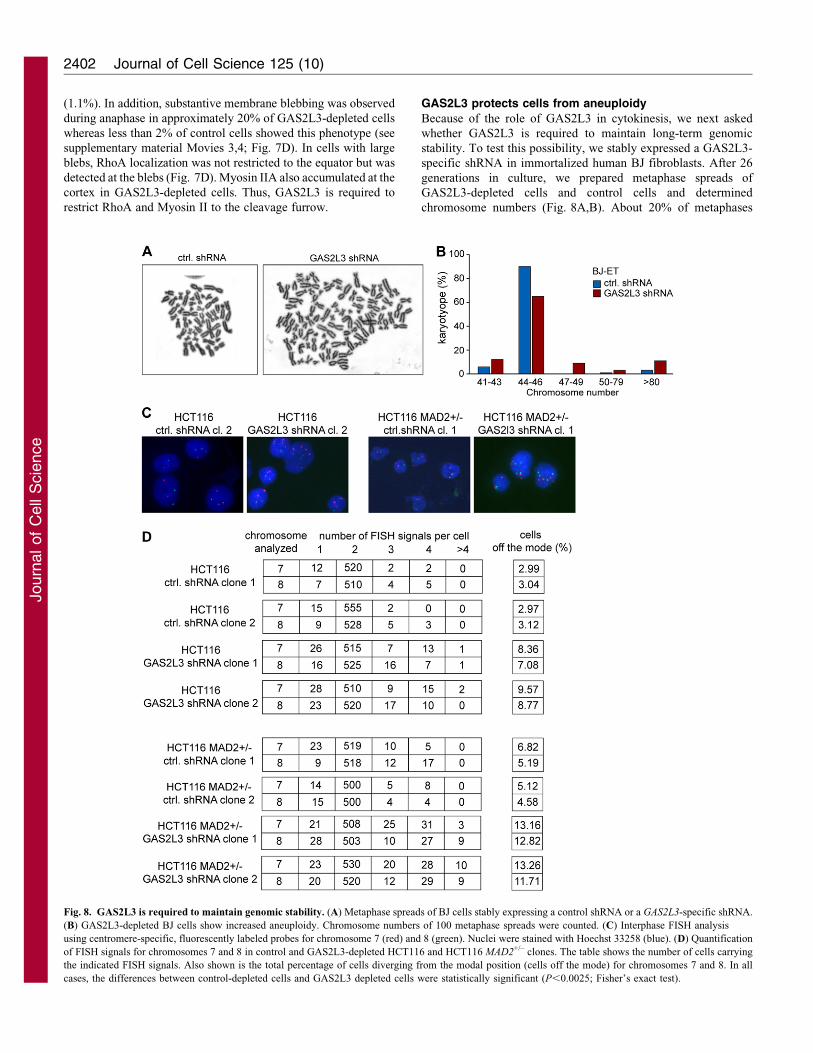
Because of the role of GAS2L3 in cytokinesis, we next asked
whether GAS2L3 is required to maintain long-term genomic
stability. To test this possibility, we stably expressed a GAS2L3-
specific shRNA in immortalized human BJ fibroblasts. After 26
generations in culture, we prepared metaphase spreads of
GAS2L3-depleted cells and control cells and determined
chromosome numbers (Fig. 8A,B). About 20% of metaphases
Fig. 8. GAS2L3 is required to maintain genomic stability. (A) Metaphase spreads of BJ cells stably expressing a control shRNA or a GAS2L3-specific shRNA.
(B) GAS2L3-depleted BJ cells show increased aneuploidy. Chromosome numbers of 100 metaphase spreads were counted. (C) Interphase FISH analysis
using centromere-specific, fluorescently labeled probes for chromosome 7 (red) and 8 (green). Nuclei were stained with Hoechst 33258 (blue). (D) Quantification
of FISH signals for chromosomes 7 and 8 in control and GAS2L3-depleted HCT116 and HCT116 MAD2+/– clones. The table shows the number of cells carrying
the indicated FISH signals. Also shown is the total percentage of cells diverging from the modal position (cells off the mode) for chromosomes 7 and 8. In all
cases, the differences between control-depleted cells and GAS2L3 depleted cells were statistically significant (P,0.0025; Fisher’s exact test).
Journal of Cell Science 125 (10)2402
Journ
alof
Cell
Scie
nce

from GAS2L3-depleted BJ cells showed polyploidy or
aneuploidy, whereas control cells had normal or nearly normalkaryotypes. We next investigated chromosomally stable HCT116cells, a near-diploid colon cancer cell line (Lengauer et al., 1997).
Single-cell clones of cells derived from cells stably expressing aGAS2L3-specific shRNA or a control shRNA were isolatedand passaged for five generations. Chromosome numberswere quantified by FISH using chromosome-specific satellite
enumeration probes in interphase nuclei. Control-transfectedcells were chromosomally stable, as judged by a small fraction ofabout 3% of cells with deviation from the modal value
(Fig. 8C,D). By contrast, 8–9% of cells treated with GAS2L3shRNA showed gains or losses of chromosome 7 and 8. Thesedata indicate that depletion of GAS2L3 is sufficient to induce low
levels of aneuploidy. We next asked whether a reduction inGAS2L3 enhances the effect of MAD2 heterozygosity, which isknown to result in genomic instability because of spindle
checkpoint defects (Michel et al., 2001). Stable clones ofHCT116 MAD2+/– cells transfected with a control shRNA or aGAS2L3-specific shRNA were analyzed by FISH. HCT116MAD2+/– cells showed higher levels of aneuploidy compared
with HCT116 wild-type cells, as expected. A total of 12–13% ofGAS2L3-depleted HCT116 MAD2+/– cells showed aneuploidyfor chromosome 7 and 8 (Fig. 8D). Thus aneuploidy is further
enhanced by combined inhibition of MAD2 and GAS2L3.
DiscussionIn this study we investigated GAS2L3, a target gene of the
DREAM complex. GAS2L3 belongs to the GAS2 family ofproteins, which are characterized by an actin-binding CH domainand a GAS2-related (GAR) domain. Previous studies have shown
roles for GAS2 proteins in interphase cells (Brancolini et al.,1992; Goriounov et al., 2003; Stroud et al., 2011; Zucman-Rossiet al., 1996). We now report that unlike the other members of theGAS2 family, GAS2L3 is expressed in M phase, localizes to
spindles, the spindle midzone and the midbody, and is requiredfor cytokinesis.
GAS2L3 is cell cycle regulated at several levels. First,
GAS2L3 mRNA levels are low during G1, increase in S phaseand peak in G2 and mitosis. Transcriptional activation ofGAS2L3 is dependent on the DREAM complex, a masterregulator of mitotic genes. Second, GAS2L3 is also regulated
at the protein level. GAS2L3 protein levels decrease when cellsexit from mitosis. The degradation of many unstable proteins atthe end of mitosis is mediated by the anaphase-promoting
complex (APC). Interestingly, GAS2L3 contains a consensus D-box, a known recognition sequence for the APC. Futuremutational analysis should reveal whether degradation of
GAS2L3 depends on this D-box motif.
We performed co-sedimentation assays to analyze theinteraction of GAS2L3 with F-actin and microtubules in vitro.Similar to a recent study, we found that the C-terminus but not the
GAR domain mediates the interaction with microtubules (Stroudet al., 2011). Further experiments revealed that there are at leasttwo non-overlapping microtubule-binding domains in the C-
terminus. In addition, GAS2L3 has several actin-binding domains– the N-terminal CH domain and at least two different domains inthe C-terminus that cannot clearly separated from the microtubule-
binding sites. The unstructured C-terminal domain of GAS2L3contains several clusters of positively charged amino acids thatcould mediate binding to the negatively charged microtubules and
actin. GAS2L3 not only binds to microtubule but also hasmicrotubule-bundling activity in vitro and, when overexpressed,
it can protect microtubule from nocodazole-induceddepolymerization. Bundling activity of GAS2L3 can bemediated by a single molecule because we found at least twonon-overlapping microtubule-binding sites in the C-terminus. Our
analysis also revealed that GAS2L3 can simultaneously bind to F-actin and microtubules, suggesting that it might function tocoordinate the actin and microtubule cytoskeletons.
The ability of GAS2L3 to localize to the spindle and spindlemidzone correlates well with its ability to bind to microtubules invitro (Fig. 3). Even a small C-terminal fragment with only one of
the two microtubule-binding domains displays some overlap withthe spindle and spindle midzone. However, only the full-lengthGAS2L3 shows the same midbody localization as endogenousGAS2L3, indicating that the N-terminal CH and GAR domains
in addition to the C-terminal actin- and microtubule-bindingdomains are required to localize GAS2L3 to the midbody. Thismight be due to the additional actin-binding activity of the CH
domain. Alternatively, it is possible that these domains mediateinteraction with other proteins that contribute to the localizationof GAS2L3 to the midbody.
So far, no loss-of-function studies of mammalian GAS2proteins have been reported. We found that depletion of GAS2L3by RNAi results in an increase in multinuclear cells and cellswith multi-lobed nuclei, indicating defects in cytokinesis, which
is consistent with its expression and localization during the cellcycle. Time-lapse videomicroscopy revealed that multinucleationwas not due to a failure to ingress the furrow. Instead, it was
caused by abnormal contractions and oscillations that started atthe beginning of cytokinesis. During these oscillations, thechromatin and the spindle were moved back and forth several
times between the future daughter cells. Furrows eventuallycollapsed and cells became multinucleated and multilobed. Wealso observed increased membrane blebbing after depletion of
GAS2L3 and found abnormal localization of myosin IIAand RhoA at the polar cortex. Interestingly, this phenotypeof GAS2L3-depleted cells is very similar to the oscillatingphenotype that has been reported after deletion of anillin, the
formin mDia2, the kinesin MCAK or the motor protein HSET,which is required for spindle midzone organization (Cai et al.,2010; Piekny and Glotzer, 2008; Straight et al., 2005; Watanabe
et al., 2008; Zhao and Fang, 2005). MCAK depletion leads to anincrease in the length of astral microtubules that invade andincrease the size of membrane blebs (Rankin and Wordeman,
2010). Oscillations are then triggered by Rho-dependent myosincontractions. Anillin or mDia2 depletion induces oscillationsbecause these proteins act as scaffolds that stabilize the positionof the contractile ring during cytokinesis (Straight et al., 2005;
Watanabe et al., 2008). Given the similar phenotypes, it is likelythat GAS2L3 is also involved in regulating components of thecontractile ring. Because GAS2L3 can simultaneously bind to
microtubules and actin filaments in vitro, GAS2L3 couldfunction as a scaffolding protein in the cleavage furrow tostabilize interactions between midbody microtubules and the
actomyosin ring. Consistent with such a function, RhoA andmyosin were not restricted to the cleavage furrow after GAS2L3depletion but also aberrantly localized at the sites of large
membrane blebs, where they might induce abnormal corticaloscillation. Although these observations support a role forGAS2L3 in stabilization of components of the contractile ring,
Role of GAS2L3 in cytokinesis 2403
Journ
alof
Cell
Scie
nce

we cannot exclude the possibility that other mechanisms, such as
changes in astral microtubules contribute to the observed
phenotype; however, there is no evidence that GAS2L3 can
localize to astral microtubules. To elucidate the precise function
of GAS2L3 in cytokinesis further studies such as the
identification of interacting proteins will be needed.
In this study, we also found that the long-term depletion of
GAS2L3 resulted in aneuploidy in human BJ fibroblasts and
HCT116 cells, indicating that GAS2L3 is required to maintain
genomic stability (Fig. 8). Given that genomic instability is
a hallmark of most cancers (Holland and Cleveland, 2009),
the deregulation of GAS2L3 could therefore contribute to
tumorigenesis. Interestingly, a search of the Oncomine database
revealed significant deregulation of GAS2L3 mRNA in certain
tumor types. For example, an upregulation of GAS2L3 in
glioblastoma has been reported in two recent studies (Sun et al.,
2006; Bredel et al., 2005). Conversely, GAS2L3 is significantly
downregulated in childhood T-cell acute lymphoblastic leukemia
(Andersson et al., 2007). In the future, it will be interesting to
investigate whether GAS2L3 plays a role in human cancer.
In conclusion, our data establish GAS2L3 as a target gene of
the DREAM complex that is required for proper cytokinesis and
maintenance of genomic stability. Downregulation of GAS2L3
probably contributes to the cytokinesis defects after inactivation
of DREAM.
Materials and MethodsTissue culture
All cells were cultured in DMEM (Invitrogen, Darmstadt, Germany) supplemented
with 10% FCS (Invitrogen, Darmstadt, Germany). HeLa cells stably expressingH2B–EGFP and a-tubulin–RFP were a gift from Patrick Meraldi (Toso et al.,
2009). HCT116 MAD2+/– cells were a gift from Robert Benezra (Michel et al.,2001). HeLa FlpIn-TRex cells were a gift from Stephen Taylor (Tighe et al., 2008).
HeLa cells stably expressing mCherry–a-tubulin were generated by stabletransfection with the plasmid pmCherry–a-tubulin–IRESpuro2 (Steigemann et al.,2009).
Plasmids, siRNA and transfections
Human GAS2L3 cDNA was obtained by RT-PCR from HeLa mRNA and inserted
into pCDNA3-EGFP and pGEX-4T-2 expression vectors. Mouse Gas2l3 cDNAwas obtained by RT-PCR from NIH3T3 cells and inserted into pCDNA3-HA. To
generate stable HeLa cells expressing double-tagged GAS2L3, the GAS2L3 cDNAwas inserted into pcDNA5/FRT/TO/nHASt-TAP encoding for a N-terminal HA
tag and a streptavidin-binding peptide (Wyler et al., 2011). The plasmid (0.1 mg)was co-transfected together with the flp-recombinase expression vector pOG44(0.9 mg) into HeLa Flp-In TRex cells (Tighe et al., 2008). Cells were selected for 2
weeks with 100 mg/ml hygromycin.
The following GAS2L3-specific siRNAs were used: siRNA#1, 59-
GGGAUACUCUUCAAGGAUU-39; siRNA#2, 59-CUAUGUCAGUCCGUUCU-AA-39; siRNA#3, 59-CAUUAAAUCCAGUAGGUAA-39.
siRNAs were purchased from MWG-Biotech (Ebersberg, Germany) and weretransfected with Lipofectamine 2000 (Invitrogen, Darmstadt, Germany) orMetafectene Pro (Biontex, Martinsried) according to the manufacturer’s
protocol. A retroviral GAS2L3 shRNA construct was generated by insertion ofan shRNA corresponding to the siRNA#2 into the retroviral vector pMSCV.
Retroviral infections with shRNA viruses were performed as described (Gagricaet al., 2004). A siRNA-resistant GAS2L3 was generated by introducing silent pointmutations that impair siRNA-mediated knockdown. Using site-directed
mutagenesis, nucleotides 1337–1339 of GAS2L3 encoding Ser371 were changedfrom TCT to AGC. The mutated cDNA was inserted into pcDNA5/FRT/TO/
nHASt-TAP and stable HeLa cells using the HeLa Flp-In TRex cells weregenerated as described above.
Antibodies
The following primary antibodies were used: GAS2L3 (Abnova, 1D4 and 1C8), a-
tubulin (Sigma, T6074), Actin (Santa Cruz, sc-47778), Aurora B (Abcam, ab2254),cyclin B (Santa Cruz, sc-245), HA (Covance, HA.11), MKLP1 (Santa Cruz, sc-
867), phospho-Histone H3 (Upstate, 06–570), RhoA (Santa Cruz, sc-418), myosinIIA (Sigma, M8064).
ImmunofluorescenceCells grown on coverslips were either fixed for 4 minutes at 220 C with methanolor for 10 minutes at room temperature with PFA (PBS, 3% paraformaldehyde, 2%sucrose). PFA-fixed cells were permeabilized for 5 minutes with 0.2% Triton X-100in PBS and washed with PBST (0.1% Triton X-100 in PBS). Slides were blocked for20– 60 minutes with 2–3% BSA in PBS, washed three times in PBS and incubatedwith primary antibodies. Coverslips were washed three times with PBS andincubated with secondary antibody (Invitrogen, Darmstadt, Germany) in PBST for30 minutes. Nuclei were stained with Hoechst 33258 (Sigma, Munich, Germany).
For the detection of RhoA and myosin IIA, cells were fixed for 15 minutes in10% TCA and processed as described above. For F-actin staining, cells were fixedwith 4% PFA in PBS, permeabilized and stained with Alexa Fluor 594 phalloidin(Molecular Probes).
Recombinant proteinsExpression of recombinant GST–GAS2L3 in BL21(DE3) pLysS or Rosetta (DE3)E. coli was induced by addition of 1 mM IPTG at 15 C overnight. The recombinantprotein was purified on glutathione-linked Sepharose beads according to standardprotocols. GST–GAS2L3 was dialyzed against BRB80 buffer (80mM PIPES,pH 6.8, 1mM MgCl2 and 1mM EGTA) overnight.
Microtubule and F-actin co-sedimentation assayTubulin was prepared from porcine brain following the protocol described(Mitchison and Kirschner, 1984). Tubulin was polymerized at 37 C for 30 minutesby addition of 0.5 volumes of glycerol and 1 mM GTP. The polymerized tubulinwas diluted to 10 mM with 20 mM taxol containing BRB80 buffer. For co-sedimentation assays, 20 ml of polymerized tubulin was mixed with 1 mM GST–GAS2L3 or GST protein and incubated for 15 minutes at room temperature.Mixtures were pelleted by centrifugation at 30,000 g for 30 minutes at 30 C over a30% glycerol cushion. The supernatant and pellet fractions were recovered andseparated by SDS-PAGE. Gels were stained with Coomassie Blue.
F-actin co-sedimentation assay was performed using the Non-Muscle ActinBinding Protein Biochem Kit (Cytoskeleton, Denver, CO). Actin was polymerizedat room temperature for 1 hour according to the manufacturer’s protocol. For co-sedimentation assays, 40 ml F-actin was mixed with 1 mM GST–GAS2L3 or GSTprotein and incubated for 30 minutes at room temperature. Mixtures were pelletedby centrifugation at 120,000 g for 60 minutes at 24 C. Supernatant and pelletfractions were recovered and separated by SDS-PAGE. Gels were stained withCoomassie Blue.
Microtubule-bundling assay5 mM tubulin and 0.4 mM Cy5-labeled tubulin were mixed and polymerized byaddition of 20 mM taxol and 1mM GTP in BRB80 at 37 C for 30 minutes. Afteraddition of 500nM GST–GAS2L3 or GST, the mixtures were incubated for anadditional 30 minutes at 37 C. 1.5 ml aliquots were pipetted onto slides, coveredwith 1.5 ml fixing solution (BRB80 buffer, 50% glycerol and 8% formaldehyde)and analyzed by fluorescence microscopy.
Microtubule and F-actin crosslinking assayThe microtubule–F-actin crosslinking assay was performed as describedpreviously (Miller et al., 2004). Briefly, 1 mM GST–GAS2L3 protein wasincubated with 5 mM polymerized tubulin in tubulin polymerization buffer andincubated for 15 minutes at room temperature. After addition of 5 mM F-actin, themixture was incubated for an additional 15 minutes at room temperature andcentrifuged at 5000 g for 10 minutes at 24 C. Supernatant (S) and pellet (P)fractions were recovered, separated by SDS-PAGE and stained with CoomassieBlue. The fraction of actin in the pellet was quantified using ImageJ.
Time-lapse microscopyHeLa cells stably expressing H2B–EGFP and a-tubulin–mRFP were transfectedwith control siRNA or GAS2L3-specific siRNA in 35 mm dish chambers (Ibidi,Munich, Germany). HeLa cells stably expressing mCherry–a-tubulin weretransiently transfected with EGFP–GAS2L3. Live-cell imaging was performedusing a Leica heating insert with attached incubator S-2. Images were capturedevery 2 minutes using Leica Application Suite.
RT-PCRTotal RNA was isolated with Trizol (Invitrogen, Darmstadt, Germany), reversetranscribed with 0.5 U M-MLV-RT Transcriptase (Thermo Scientific, Epsom, UK)and analyzed with quantitative real-time PCR with SYBR Green reagents fromThermo Scientific using the Mx3000 (Agilent technologies, Waldbronn, Germany)detection system. Expression differences were calculated relative to GAPDH asdescribed previously (Schmit et al., 2007).
ImmunoblottingCells were lysed in TNN [50 mM Tris-HCl (pH 7.5), 120 mM NaCl, 5 mMEDTA, 0.5% NP40, 10 mM Na4P2O7, 2 mM Na3VO4, 100 mM NaF, 10 mg/ml
Journal of Cell Science 125 (10)2404
Journ
alof
Cell
Scie
nce

phenylmethylsulfonyl fluoride, protease inhibitors (Sigma, Munich, Germany)].Proteins were separated by SDS-PAGE, transferred to PVDF membrane anddetected by immunoblotting.
Karyotype analysis and FISH
To prepare metaphase spreads, cells were treated with 10 ng/ml colcemid (Sigma,Munich, Germany) for 2 hours (HCT116 cells) or 5 hours (BJ cells) at 37 C. Cellswere collected by trypsinization, washed with DMEM and suspended in 0.8%sodium citrate at 37 C for 30–40 minutes. Next, cells were fixed in methanol andacetic acid (3:1) overnight at 220 C, dropped onto slides and stained with 5%Giemsa solution for 5 minutes. Metaphase spreads were observed under lightmicroscopy and chromosome numbers determined.
For FISH in interphase cells, poseidon chromosome 7 and 8 satelliteenumeration probes (Kreatech diagnostics, Amsterdam, Netherlands) were usedaccording to the manufacturer. DNA was counterstained with Hoechst 33258.Fluorescence signals of at least 500 nuclei were counted.
AcknowledgementsWe thank all members of the laboratory for their suggestions andcritical reading of the manuscript. We thank Susanne Spahr andAdelgunde Wolpert for their excellent technical assistance and MeikKunz for help with the time-lapse analysis. We thank PatrickMeraldi, Stephen Taylor, Robert Benezra, Holger Bastians, DanielGerlich and Ulrike Kutay for providing cell lines and plasmids. Wethank Olaf Stemmann for helpful discussions. We thank ClausSteinlein, Michael Schmid and Clemens Grimm for their help withkaryotype analysis and recombinant protein expression.
FundingThis work was supported by grants from the DeutscheForschungsgemeinschaft [grant numbers 575/6-1 and TR17-B1] toS.G.
Supplementary material available online at
http://jcs.biologists.org/lookup/suppl/doi:10.1242/jcs.097253/-/DC1
ReferencesAndersson, A., Ritz, C., Lindgren, D., Eden, P., Lassen, C., Heldrup, J., Olofsson, T.,
Rade, J., Fontes, M., Porwit-Macdonald, A. et al. (2007). Microarray-basedclassification of a consecutive series of 121 childhood acute leukemias: prediction ofleukemic and genetic subtype as well as of minimal residual disease status. Leukemia
21, 1198-1203.
Brancolini, C., Bottega, S. and Schneider, C. (1992). Gas2, a growth arrest-specificprotein, is a component of the microfilament network system. J. Cell Biol. 117, 1251-1261.
Bredel, M., Bredel, C., Juric, D., Harsh, G. R., Vogel, H., Recht, L. D. and Sikic,
B. I. (2005). Functional network analysis reveals extended gliomagenesis pathwaymaps and three novel MYC-interacting genes in human gliomas. Cancer Res. 65,8679-8689.
Burkhart, D. L. and Sage, J. (2008). Cellular mechanisms of tumour suppression by theretinoblastoma gene. Nat. Rev. Cancer 8, 671-682.
Cai, S., Weaver, L. N., Ems-McClung, S. C. and Walczak, C. E. (2010). Properorganization of microtubule minus ends is needed for midzone stability andcytokinesis. Curr. Biol. 20, 880-885.
Cobrinik, D. (2005). Pocket proteins and cell cycle control. Oncogene 24, 2796-2809.
Dimova, D. K. and Dyson, N. J. (2005). The E2F transcriptional network: oldacquaintances with new faces. Oncogene 24, 2810-2826.
Eggert, U. S., Mitchison, T. J. and Field, C. M. (2006). Animal cytokinesis: from partslist to mechanisms. Annu. Rev. Biochem. 75, 543-566.
Gagrica, S., Hauser, S., Kolfschoten, I., Osterloh, L., Agami, R. and Gaubatz, S.
(2004). Inhibition of oncogenic transformation by mammalian Lin-9, a pRB-associated protein. EMBO J. 23, 4627-4638.
Goriounov, D., Leung, C. L. and Liem, R. K. H. (2003). Protein products of humanGas2-related genes on chromosomes 17 and 22 (hGAR17 and hGAR22) associatewith both microfilaments and microtubules. J. Cell Sci. 116, 1045-1058.
Gromley, A., Yeaman, C., Rosa, J., Redick, S., Chen, C.-T., Mirabelle, S., Guha, M.,
Sillibourne, J. and Doxsey, S. J. (2005). Centriolin anchoring of exocyst andSNARE complexes at the midbody is required for secretory-vesicle-mediatedabscission. Cell 123, 75-87.
Hickson, G. R. X. and O’Farrell, P. H. (2008). Anillin: a pivotal organizer of thecytokinetic machinery. Biochem. Soc. Trans. 36, 439-441.
Holland, A. J. and Cleveland, D. W. (2009). Boveri revisited: chromosomal instability,aneuploidy and tumorigenesis. Nat. Rev. Mol. Cell Biol. 10, 478-487.
Kittler, R., Pelletier, L., Heninger, A.-K., Slabicki, M., Theis, M., Miroslaw, L.,
Poser, I., Lawo, S., Grabner, H., Kozak, K. et al. (2007). Genome-scale RNAiprofiling of cell division in human tissue culture cells. Nat. Cell Biol. 9, 1401-1412.
Knight, A. S., Notaridou, M. and Watson, R. J. (2009). A Lin-9 complex is recruited
by B-Myb to activate transcription of G2/M genes in undifferentiated embryonal
carcinoma cells. Oncogene 28, 1737-1747.
Lengauer, C., Kinzler, K. W. and Vogelstein, B. (1997). Genetic instability in
colorectal cancers. Nature 386, 623-627.
Litovchick, L., Sadasivam, S., Florens, L., Zhu, X., Swanson, S. K., Velmurugan, S.,
Chen, R., Washburn, M. P., Liu, X. S. and DeCaprio, J. A. (2007). Evolutionarily
conserved multisubunit RBL2/p130 and E2F4 protein complex represses human cell
cycle-dependent genes in quiescence. Mol. Cell 26, 539-551.
Matuliene, J. and Kuriyama, R. (2002). Kinesin-like protein CHO1 is required for the
formation of midbody matrix and the completion of cytokinesis in mammalian cells.
Mol. Biol. Cell 13, 1832-1845.
Michel, L. S., Liberal, V., Chatterjee, A., Kirchwegger, R., Pasche, B., Gerald, W.,
Dobles, M., Sorger, P. K., Murty, V. V. and Benezra, R. (2001). MAD2 haplo-
insufficiency causes premature anaphase and chromosome instability in mammalian
cells. Nature 409, 355-359.
Miller, A. L., Wang, Y., Mooseker, M. S. and Koleske, A. J. (2004). The Abl-related
gene (Arg) requires its F-actin-microtubule cross-linking activity to regulate
lamellipodial dynamics during fibroblast adhesion. J. Cell Biol. 165, 407-419.
Mishima, M., Kaitna, S. and Glotzer, M. (2002). Central spindle assembly and
cytokinesis require a kinesin-like protein/RhoGAP complex with microtubule
bundling activity. Dev. Cell 2, 41-54.
Mitchison, T. and Kirschner, M. (1984). Microtubule assembly nucleated by isolated
centrosomes. Nature 312, 232-237.
Mollinari, C., Kleman, J.-P., Jiang, W., Schoehn, G., Hunter, T. and Margolis, R. L.
(2002). PRC1 is a microtubule binding and bundling protein essential to maintain the
mitotic spindle midzone. J. Cell Biol. 157, 1175-1186.
Narumiya, S. and Yasuda, S. (2006). Rho GTPases in animal cell mitosis. Curr. Opin.
Cell Biol. 18, 199-205.
Osterloh, L., von Eyss, B., Schmit, F., Rein, L., Hubner, D., Samans, B., Hauser, S.
and Gaubatz, S. (2007). The human synMuv-like protein LIN-9 is required for
transcription of G2/M genes and for entry into mitosis. EMBO J. 26, 144-157.
Piekny, A. J. and Glotzer, M. (2008). Anillin is a scaffold protein that links RhoA,
actin, and myosin during cytokinesis. Curr. Biol. 18, 30-36.
Piekny, A. J. and Maddox, A. S. (2010). The myriad roles of Anillin during
cytokinesis. Semin. Cell Dev. Biol. 21, 881-891.
Piekny, A., Werner, M. and Glotzer, M. (2005). Cytokinesis: welcome to the Rho
zone. Trends Cell Biol. 15, 651-658.
Pilkinton, M., Sandoval, R. and Colamonici, O. R. (2007a). Mammalian Mip/LIN-9
interacts with either the p107, p130/E2F4 repressor complex or B-Myb in a cell cycle-
phase-dependent context distinct from the Drosophila dREAM complex. Oncogene
26, 7535-7543.
Pilkinton, M., Sandoval, R., Song, J., Ness, S. A. and Colamonici, O. R. (2007b).
Mip/LIN-9 regulates the expression of B-Myb and the induction of cyclin A, cyclin B,
and CDK1. J. Biol. Chem. 282, 168-175.
Pohl, C. and Jentsch, S. (2008). Final stages of cytokinesis and midbody ring formation
are controlled by BRUCE. Cell 132, 832-845.
Rankin, K. E. and Wordeman, L. (2010). Long astral microtubules uncouple mitotic
spindles from the cytokinetic furrow. J. Cell Biol. 190, 35-43.
Reichert, N., Wurster, S., Ulrich, T., Schmitt, K., Hauser, S., Probst, L., Gotz, R.,
Ceteci, F., Moll, R., Rapp, U. et al. (2010). Lin9, a subunit of the mammalian
DREAM complex, is essential for embryonic development, for survival of adult mice,
and for tumor suppression. Mol. Cell. Biol. 30, 2896-2908.
Sagona, A. P. and Stenmark, H. (2010). Cytokinesis and cancer. FEBS Lett. 584, 2652-
2661.
Schmit, F., Korenjak, M., Mannefeld, M., Schmitt, K., Franke, C., von Eyss, B.,
Gagrica, S., Hanel, F., Brehm, A. and Gaubatz, S. (2007). LINC, a human complex
that is related to pRB-containing complexes in invertebrates regulates the expression
of G2/M genes. Cell Cycle 6, 1903-1913.
Schmit, F., Cremer, S. and Gaubatz, S. (2009). LIN54 is an essential core subunit of
the DREAM/LINC complex that binds to the cdc2 promoter in a sequence-specific
manner. FEBS J. 276, 5703-5716.
Schneider, C., King, R. M. and Philipson, L. (1988). Genes specifically expressed at
growth arrest of mammalian cells. Cell 54, 787-793.
Smith, D. S., Niethammer, M., Ayala, R., Zhou, Y., Gambello, M. J., Wynshaw-
Boris, A. and Tsai, L. H. (2000). Regulation of cytoplasmic dynein behaviour and
microtubule organization by mammalian Lis1. Nat. Cell Biol. 2, 767-775.
Steigemann, P., Wurzenberger, C., Schmitz, M. H. A., Held, M., Guizetti, J., Maar, S.
and Gerlich, D. W. (2009). Aurora B-mediated abscission checkpoint protects against
tetraploidization. Cell 136, 473-484.
Straight, A. F., Field, C. M. and Mitchison, T. J. (2005). Anillin binds nonmuscle
myosin II and regulates the contractile ring. Mol. Biol. Cell 16, 193-201.
Stroud, M. J., Kammerer, R. A. and Ballestrem, C. (2011). Characterization of G2L3
(GAS2-like 3), a new microtubule- and actin-binding protein related to spectrapla-
kins. J. Biol. Chem. 286, 24987-24995.
Sun, L., Hui, A.-M., Su, Q., Vortmeyer, A., Kotliarov, Y., Pastorino, S., Passaniti, A.,
Menon, J., Walling, J., Bailey, R. et al. (2006). Neuronal and glioma-derived stem cell
factor induces angiogenesis within the brain. Cancer Cell 9, 287-300.
Tighe, A., Staples, O. and Taylor, S. (2008). Mps1 kinase activity restrains anaphase
during an unperturbed mitosis and targets Mad2 to kinetochores. J. Cell Biol. 181,
893-901.
Role of GAS2L3 in cytokinesis 2405
Journ
alof
Cell
Scie
nce

Toso, A., Winter, J. R., Garrod, A. J., Amaro, A. C., Meraldi, P. and McAinsh,A. D. (2009). Kinetochore-generated pushing forces separate centrosomes duringbipolar spindle assembly. J. Cell Biol. 184, 365-372.
Trimarchi, J. M. and Lees, J. A. (2002). Sibling rivalry in the E2F family. Nat. Rev.
Mol. Cell Biol. 3, 11-20.van den Heuvel, S. and Dyson, N. J. (2008). Conserved functions of the pRB and E2F
families. Nat. Rev. Mol. Cell Biol. 9, 713-724.Watanabe, S., Ando, Y., Yasuda, S., Hosoya, H., Watanabe, N., Ishizaki, T. and
Narumiya, S. (2008). mDia2 induces the actin scaffold for the contractile ring andstabilizes its position during cytokinesis in NIH 3T3 cells. Mol. Biol. Cell 19, 2328-2338.
Wyler, E., Zimmermann, M., Widmann, B., Gstaiger, M., Pfannstiel, J., Kutay, U. andZemp, I. (2011). Tandem affinity purification combined with inducible shRNA expressionas a tool to study the maturation of macromolecular assemblies. RNA 17, 189-200.
Zhao, W.-M. and Fang, G. (2005). Anillin is a substrate of anaphase-promotingcomplex/cyclosome (APC/C) that controls spatial contractility of myosin during latecytokinesis. J. Biol. Chem. 280, 33516-33524.
Zucman-Rossi, J., Legoix, P. and Thomas, G. (1996). Identification of new membersof the Gas2 and Ras families in the 22q12 chromosome region. Genomics 38, 247-254.
Journal of Cell Science 125 (10)2406
Journ
alof
Cell
Scie
nce





























