Embed Size (px)
DESCRIPTION
Protocols in Immunology
Citation preview

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX10.1146/annurev.immunol.23.021704.115756
Annu. Rev. Immunol. 2005. 23:337–66doi: 10.1146/annurev.immunol.23.021704.115756
Copyright c© 2005 by Annual Reviews. All rights reservedFirst published online as a Review in Advance on November 12, 2004
PENTRAXINS AT THE CROSSROADS BETWEEN
INNATE IMMUNITY, INFLAMMATION, MATRIX
DEPOSITION, AND FEMALE FERTILITY
Cecilia Garlanda,1∗ Barbara Bottazzi,1∗ Antonio Bastone,1
and Alberto Mantovani1,2
1Istituto di Ricerche Farmacologiche Mario Negri, 20157 Milan, Italy; 2Institute ofGeneral Pathology, Faculty of Medicine, University of Milan, Italy;email: [email protected]; [email protected]; [email protected];[email protected]
Key Words inflammatory response, immune response, extracellular matrix,fertility, neurodegeneration
■ Abstract C reactive protein, the first innate immunity receptor identified, andserum amyloid P component are classic short pentraxins produced in the liver. Longpentraxins, including the prototype PTX3, are expressed in a variety of tissues. Somelong pentraxins are expressed in the brain and some are involved in neuronal plasticityand degeneration. PTX3 is produced by a variety of cells and tissues, most notablydendritic cells and macrophages, in response to Toll-like receptor (TLR) engagementand inflammatory cytokines. PTX3 acts as a functional ancestor of antibodies, rec-ognizing microbes, activating complement, and facilitating pathogen recognition byphagocytes, hence playing a nonredundant role in resistance against selected pathogens.In addition, PTX3 is essential in female fertility because it acts as a nodal point forthe assembly of the cumulus oophorus hyaluronan-rich extracellular matrix. Thus, theprototypic long pentraxin PTX3 is a multifunctional soluble pattern recognition recep-tor at the crossroads between innate immunity, inflammation, matrix deposition, andfemale fertility.
The serum obtained from human beings and monkeys during the acutephase of diverse infections contains a protein which is precipitable by theC polysaccharide of pneumococcus (8).
∗Cecilia Garlanda and Barbara Bottazzi have equally contributed to the Pentraxin Projectand to this review.
0732-0582/05/0423-0337$14.00 337
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
338 GARLANDA ET AL.
INTRODUCTION
Innate immunity is a first line of resistance against pathogens, and it plays a keyrole in the activation and orientation of adaptive immunity and in the maintenanceof tissue integrity and repair. Recognition of pathogens and damaged tissues ismediated by pattern recognition receptors (PRRs) (1). Innate defense mechanismsconsist of a cellular and a humoral arm. Cellular PRRs belong to different functionaland structural groups, which include the Toll-like receptors (TLR), scavengerreceptors, lectin receptors, and G protein–coupled receptors for formyl peptides(2). The humoral arm of innate immunity is also diverse; it includes collectins(mannose-binding lectin, surfactant protein A and D, C1q), ficolins, and pentraxins.
Pentraxins are a superfamily of evolutionarily conserved proteins characterizedby a structural motif, the pentraxin domain (3–6). C reactive protein (CRP), whichtogether with serum amyloid P (SAP) component constitutes the short pentraxinarm of the superfamily, was the first PRR to be identified, as vividly illustrated bythe words of Abernethy & Avery, above (7, 8). CRP was originally described andnamed for its ability to bind in a Ca-dependent manner the C-polysaccharide ofStreptococcus pneumoniae.
PTX3 (9, 10) and subsequently identified long pentraxins (11–18) were iden-tified in the 1990s as cytokine-inducible genes or molecules expressed in specifictissues (e.g., neurons, spermatozoa). Short and long pentraxins are conserved inevolution from arachnids and insects to humans. Their conservation is testimonyto their role in complex organisms. Structural analysis and gene-modified micehave provided a new level of understanding of the role of pentraxins in immunityand homeostasis.
In this review, we summarize current understanding of the structure and functionof pentraxins. Our focus is primarily on the more recently discovered long pen-traxin family and its prototypic member PTX3, but we also discuss recent progresson the structure and functions of the classic short pentraxins CRP and SAP. Currentresearch suggests that pentraxins are an essential component of the humoral arm ofinnate immunity, activated following pathogen recognition by cellular PRR. More-over, pentraxins are multifunctional proteins at the crossroads between immunityand inflammation, extracellular matrix construction, and female fertility.
THE PENTRAXIN SUPERFAMILY
Members
Pentraxins are characterized by the presence in their carboxy-terminal of a 200amino acid pentraxin domain, with an 8 amino acid–long conserved pentraxin sig-nature (HxCxS/TWxS, where x is any amino acid) (Figure 1A). The first pentraxindescribed was CRP, identified in human serum during the 1930s as the prototypicacute phase response protein (7) produced by the liver in inflammatory or infec-tious conditions. Human SAP was subsequently identified as a relative of CRP for
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
PENTRAXINS IN INNATE IMMUNITY 339
the amino acid sequence homology (51%) and for the similar appearance in elec-tron microscopy (annular disc-like structure with pentameric symmetry) (3, 4, 19).CRP and SAP orthologs in different mammal species share substantial sequencehomology, with notable differences including serum basal levels and changes dur-ing the acute phase response, CRP and SAP being the main acute phase reactantsin human and mouse, respectively. In the arthropod Limulus polyphemus, differentforms of CRP and SAP were identified as abundant constituents of the hemolymph(20–22) involved in recognizing and destroying pathogens.
During the early 1990s, a new pentraxin domain–containing secreted proteinwas identified as an IL-1-inducible gene in endothelial cells (PTX3) or as a TNF-stimulated gene (TSG-14) in fibroblasts (9, 10). The main structural property ofthe long pentraxin PTX3 that differentiated it from CRP and SAP was the presenceof a 174 amino acid–long amino-terminal domain, not present in CRP and SAP,coupled to the pentraxin domain (Figure 1A).
The long pentraxins sharing the same general organization identified after PTX3include guinea pig apexin (14, 23), neuronal pentraxin (NP) 1 or NPTX1 (11, 13),NP2 (also called Narp or NPTX2) (12, 16), and neuronal pentraxin receptor (NPR),a transmembrane molecule (17, 18) (see below). The amino acid sequence iden-tity among members of this subfamily is relatively high in the carboxy-pentraxindomain and ranges from 28% between human PTX3 (hPTX3) and hNP1 to 68%between hNP1 and hNP2. By contrast, in the amino-terminal domain a low levelof similarity is found (about 10% for PTX3 versus NP1). However, identity in theamino-terminal domain among the neuronal pentraxins is higher and ranges be-tween 28% and 38%, suggesting the existence of subclasses of molecules amongthe long pentraxins. The homology between NP1 and PTX3 or between NP2 andPTX3 at the N-terminal level is restricted to the extreme N-terminus; this char-acteristic and the longer size of NP1 and NP2 suggest the presence of a thirddomain localized between the N-terminal and the pentraxin domains (24). Or-tholog molecules have been found so far for PTX3, NP1, NP2, and NPR not onlyin human, mouse, and rat, but also in lower vertebrates such as zebrafish and puffer-fish (Y. Martinez, unpublished results) (Figure 1B). The ortholog of apexin has notbeen clearly defined. We recently identified a new evolutionarily conserved mem-ber of the long pentraxin family, PTX4 (Y. Martinez, unpublished results). Longpentraxins have been identified in Xenopus laevis (XL-PXN1) (15) and in insects.In Drosophila melanogaster, Swiss cheese protein is a long (1425 amino acids)pentraxin that, when mutated, is responsible for age-dependent neurodegeneration(25).
Phylogeny
The pentraxin domain is highly conserved in mammals, in lower vertebrates (Xeno-pus laevis, Danio rerio, the zebrafish, Takifugu rubripes, the pufferfish), and inarthropods (the arachnids Limulus polyphemus and Tachypleus tridentatus, thehorseshoe crabs, and the insect Drosophila melanogaster). Phylogenetic tree anal-ysis allows the identification of five main groups of molecules (Figure 1B).
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
340 GARLANDA ET AL.
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
PENTRAXINS IN INNATE IMMUNITY 341
The first group of molecules includes the short pentraxins that diverged fromthe common ancestor of all pentraxins. The duplication event that gave rise toCRP and SAP may have occurred very early in evolution, as both can be found inchordates as well as in arthropods (20). Interestingly, XL-PXN1 Xenopus laevis,which is a long pentraxin, clusters with the short pentraxins, possibly because ofthe low level of homology between its N-terminal domain and that of the otherlong pentraxins. The second group includes the neuronal pentraxins that cluster asa subclass of long pentraxins found in mammals as well as in lower vertebrates.On the basis of branch length, NPR is the oldest group that diverged from a com-mon ancestor of neuronal pentraxins; subsequently NP2 and finally NP1 appeared.Apexin clusters with the neuronal pentraxins and in particular with NP2. The thirdgroup includes only PTX3, whose sequence has been identified in mammals as wellas in birds (Gallus gallus) and in the most ancient vertebrate Takifugu rubripes.PTX3 originated very early in the pentraxin evolution, directly from the commonancestor of the whole family. The fourth group includes PTX4 and its orthologs,found in mammals as well as in the zebrafish. Like PTX3, PTX4 originated di-rectly from the common ancestors of the pentraxins very early in the evolutionof pentraxins, at the divergence of vertebrates. The last group is represented byDrosophila melanogaster Swiss cheese protein, a long pentraxin involved in glia-neuron interaction (25). Swiss cheese protein originated from the common ancestorof pentraxins, and interestingly it localizes in the phylogenetic tree close to PTX3.The lack of relationship among the five groups of long pentraxins identified in thisanalysis suggests that they originated independently by fusion events between thegene encoding the ancestral pentraxin domain and other unrelated sequences.
SHORT PENTRAXINS
Short pentraxins are 25-kDa proteins characterized by a common structural or-ganization in five (six in Limulus polyphemus) or ten identical subunits arrangedin a pentameric radial symmetry. Human CRP and SAP are the prototypic shortpentraxins: they share approximately 51% amino acid sequence identity and arethought to have originated from a single gene duplication (26).
←Figure 1 The pentraxin superfamily. (A) Human short and long pentraxins. TM:transmembrane domain. (B) Phylogeny of pentraxins. The dendogram was organizedby the neighbor-joining (NJ) algorithm with molecular evolutionary genetics analysis(MEGA) version 2.1. Circles identify related molecules. Abbreviations are as follows:hosa, Homo sapiens (human); mumu, Mus musculus (house mouse); rano, Rattusnorvegicus (norway rat); gaga, Gallus gallus (domestic fowl); xela, Xenopus laevis(african clawed frog); dare, Danio rerio (zebrafish); taru, Takifugu rubripes (pufferfish);drome, Drosophila melanogaster (fruit fly); gp, Cavia porcellus (guinea pig); lipo,Limulus polyphemus (horshoe crab).
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
342 GARLANDA ET AL.
Structure
Human CRP is composed of five identical nonglycosilated protomers. The aminoacids sequence includes two cysteines in position 36 and 97 conserved in all themembers of the pentraxin family and involved in intrachain disulfide bonds. EachCRP protomer has a characteristic lectin fold composed of two-layered ß sheetswith flattened jellyroll topology; five protomers are noncovalently associated toform a pentamer with a total molecular weight of 115,135 Da (27).
The human CRP gene is located on chromosome 1q23 and is organized in twoexons, the first exon coding for the leader peptide and the first two amino acidsof the mature protein and the second exon coding for the remaining 204 aminoacids. The promoter region of human CRP comprises two acute phase responseelements, each containing a binding site for the liver-specific transcription factorHNF1 (28), and two C/EBP (CCAAT/enhancer binding protein β) binding sites,both necessary and sufficient for IL-6-induced transcription (29, 30). STAT3 hasalso been found to participate in transducing the effects of IL-6 on CRP expression(31). In addition, a nonconsensus κB site overlapping the proximal C/EBP bindingsite has been identified (32). Binding of Rel P50 to the κB site in response to IL-1enhances and stabilizes binding of C/EBP to the CRP promoter (33) and amplifiesCRP expression.
CRP levels in plasma of healthy adults are barely detectable (≤3 mg/l) butincrease as much as 1000-fold following an acute phase stimulus as a result ofaccelerated rates of transcription in the liver (4). Circulating CRP is producedonly by hepatocytes (4), mainly in response to the proinflammatory cytokineIL-6. IL-1 may also contribute as an additional signal acting synergistically withIL-6 in the induction of CRP mRNA (34). Other cells, such as lymphocytes andmonocyte/macrophages, are able to synthesize CRP, but apparently these cells donot contribute to CRP plasma levels.
SAP is a highly conserved plasma glycoprotein composed of 5 or 10 identicalsubunits noncovalently associated in pentameric rings interacting face to face (35).SAP is a normal component of basement membranes (36) and is the main acutephase protein in mice, whereas in human serum it is constitutively present at 30 to50 µg/ml. The human gene maps to chromosome 1 in close physical and geneticlinkage with the CRP gene and shares with CRP the same organization in twoexons, the first one coding for the signal peptide and the second for the matureprotein. The mature SAP protomer is 204 amino acids long and has a molecularmass of 25,462 Da; in the presence of physiological levels of calcium, humanSAP is a pentamer with a molecular mass of 127,310 Da, although at pH 8.0 inthe absence of calcium, SAP consists of pentameric and decameric forms (37–39).Unlike CRP, each SAP protomer is glycosilated with a single N-linked biantennaryoligosaccharide at Asn32 (38).
According to the three-dimensional structure derived by X-ray diffraction (35),human SAP has a tertiary fold, which resembles that of legume lectins as Con-canavalin A. SAP protomers have a flattened ß-jelly roll topology with a singlelong elix folded on the top of the ß-sheet. The five subunits are arranged in a
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
PENTRAXINS IN INNATE IMMUNITY 343
ring around a hole and are held together by hydrogen bonds and salt bridges. Thedecamer is stabilized by ionic interactions between the two pentamers. Each SAPsubunit can bind two calcium ions, and residues involved in calcium binding areconserved in all SAPs.
Ligands
The physiological functions attributed to CRP and SAP involve Ca2+-dependentligand binding (Table 1). The first ligand described for CRP was the C-polysac-charide of Streptococcus pneumoniae: This interaction is due to a direct binding
TABLE 1 Differential ligand recognition by PTX3 and short pentraxins
PTX3 CRP SAP
Membrane moieties:PC − + −PE − − +Galactomannana + NT NTLPS − − +OmpA + NT NT
Complement components:C1q + + +C4b-binding protein − − +
Cytokines—Growth factors:FGF2 + +/− NTIL-1; other growth factors, cytokines and chemokines − NT NT
Matrix proteins:TSG6 + NT NTLaminin − + +Type IV collagen − − +Fibronectin − + +Chondroitin sulfate − NT +Hyaluronic acid − NT NT
Microbes:Aspergillus fumigatus + + NTPseudomonas aeruginosa + NT NTSalmonella typhimurium + − +Paracoccidioides brasiliensis + NT NTZymosan + + +
Miscellaneous:Apoptotic cells + + +Histones + + +Heparin − − +MoβDGb − − +
abased on competition data.b4,6-O-(1-carboxyethylidene)-β-D-galactopyranoside.
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
344 GARLANDA ET AL.
of CRP to phosphorylcholine (PC), a major constituent of C-type capsule polysac-charides. Moreover, CRP binds various pathogens, including fungi, yeasts, andbacteria (40). CRP has additional calcium-dependent binding specificity for chro-matin, histones, and small nuclear ribonucleoprotein U1, as well as glycans andphospholipids (4). In the absence of calcium, CRP binds polycations such as poly-L-lysine, poly-L-arginine, and myelin basic protein.
SAP is a calcium-dependent lectin originally characterized for its binding toagarose and in particular for the agarose component 4,6-cyclin pyruvate acetal ofβ-D-galactose (41). Like CRP, SAP binds various bacteria, such as Streptococcuspyogens and Neisseria meningitidis (42, 43). Moreover, binding to influenza virus(44) and to lipopolysaccharide (LPS) has also been reported (42, 45). Other ligandsfor SAP include: carbohydrate determinants such as heparin, 6-phosphorylatedmannose, and 3-sulfated saccharides (46); matrix components such as laminin,type IV collagen, fibronectin, and proteoglycans; and C4b-binding protein andall forms of amyloid fibrils. In addition, SAP is the major DNA- and chromatin-binding protein in plasma (47, 48).
CRP and SAP, aggregated or attached to most of their ligands, can bind to C1q,the recognition subunit of the classical complement pathway (49), interacting toa specific region of its collagen-like domain and activating the classical comple-ment cascade (50, 51). Complement activation by short pentraxins may be oneof the mechanisms leading to the removal of cellular debris. Consistent with thishypothesis, both CRP and SAP bind in a calcium-dependent way apoptotic cellsenhancing phagocytosis by macrophages (52, 53).
The observation of enhanced phagocytosis of apoptotic cells opsonized withSAP and CRP in the absence of complement suggests the existence of specificreceptors for pentraxins on phagocytic cells. A specific and saturable binding toall three classes of Fcγ receptors has been demonstrated for both CRP and SAP (54,55), and the interaction with Fcγ seems able to mediate phagocytosis of apoptoticcells as well as zymosan (56, 57). However, some scientists have questioned theinterpretation of these data, suggesting instead that binding of CRP to Fcγ receptorsis not demonstrable when F(ab′)2 anti-CRP antibodies are used and that bindingmay be influenced by CRP contamination with traces of IgG (58, 59).
Functions
Although much is known about the biochemical characteristics of short pentraxinsand their use as markers in different pathological situations, the actual function ofthese molecules remains, to some extent, elusive. An important contribution to theunderstanding of the in vivo role of CRP and SAP comes from the generation ofgenetically modified mice. Here, a stumbling block in understanding the biologyof short pentraxins is represented by the different regulation of CRP and SAP inmouse and human.
Several reports concerning CRP transgenic (CRPtg) mice have been publishedin recent years, although no data are available on CRP knockout animals. Early
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
PENTRAXINS IN INNATE IMMUNITY 345
observations showed that administration of human CRP increases survival of miceinfected with Streptococcus pneumoniae and this effect is likely mediated bythe strong reaction of CRP toward the PC moiety present on the cell wall ofthese bacteria. According to these data, CRPtg mice infected with Streptococcuspneumoniae (60) are resistant to infections, showing longer survival time andlower mortality than non-tg littermates. Similarly, CRP administration protectsagainst Haemophilus influenzae, a pathogen expressing PC (61, 62). Finally, CRPtganimals are resistant to infections with the Gram-negative pathogen Salmonellaenterica (63), even in the absence of CRP binding.
The effects of CRP are not limited to protection against pathogens: CRPtg miceare also protected in a model of experimental allergic encephalomyelitis (64), andadministration of human CRP offers a transient protection against systemic lupuserythematosus (SLE) in SLE-prone mice (NZB/NZW) (65). The protective effectmay be mediated by CRP binding to nuclear antigens relevant for SLE, such aschromatin, histones, and small nuclear ribonucleoprotein U1, as well as by bindingand opsonization of apoptotic cells. Recent research shows that development ofdisease is delayed and survival is increased in NZB/NZW mice carrying the humanCRP transgene (66); this protection is associated with the ability of CRP to limitrenal damage by preventing deposition of immune complexes.
Conflicting reports concern a possible protective role of CRP toward bacterialLPS. Mice expressing the rabbit CRP gene are resistant to the lethal effect ofLPS (67); similarly, human CRP administration is protective in some experimen-tal models (42, 68), although protection of mice against LPS lethality by passiveadministration of CRP is not a general phenomenon (69). Such conflicting re-ports may reflect an inherent variability of models in which CRPs from differentspecies and purity were used, associated to differences in the animal strains andto heterogeneity of LPS preparations.
The availability of sap knockout (sap−/−) mice (70) helped to advance un-derstanding of the biological role of this protein. Surprisingly, SAP plays a dualrole in bacterial infections, exhibiting a host defense function against pathogensto which it does not bind. When SAP binds to bacteria, a strong antiopsoniceffect is observed, resulting in enhanced virulence of the infectious agent(42).
SAP is a universal constituent of the amyloid deposits that are characteristicof systemic amyloidosis, Alzheimer’s disease, and prion diseases. SAP binds toamyloid fibrils (71) and stabilizes the deposits, participating in the pathogenesisof the disease. The role of SAP in amyloidogenesis has been investigated in vivoin sap−/− animals: in these mice, the appearance of amyloid deposits is delayedand their quantity is reduced (70). Pharmacologic targeting of SAP may representan innovative therapeutic strategy (72). Furthermore, sap−/− mice spontaneouslydevelop antinuclear autoimmunity and severe glomerulonephritis (73), a phenotyperesembling human SLE. These results strongly support a role for SAP in theprotection against chromatin-induced autoimmunity.
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
346 GARLANDA ET AL.
THE PROTOTYPIC LONG PENTRAXIN PTX3
Gene and Protein Organization
The human PTX3 gene, localized on human chromosome 3 band q25, is organizedin three exons separated by two introns. The first two exons code for the leaderpeptide and the N-terminal domain of the protein, respectively, and the third exonencodes the pentraxin domain, matching exactly the other members of the pentraxinfamily (9).
The PTX3 protein is 381 amino acids long, including a signal peptide of 17amino acids. The mature secreted protein has a predicted molecular weight of40,165 Da and consists of a C-terminal 203 amino acids pentraxin-like domain cou-pled with an N-terminal portion of 178 amino acids unrelated to other known pro-teins. Scientists have observed a significant alignment between PTX3 C-terminaldomain and classical short pentraxins, with 57% of conserved amino acids and17% of identical amino acids. Analysis of human PTX3 sequence indicates thepresence of an N-linked glycosilation site in the C-terminal domain at Asn220 thataccounts for the higher molecular weight observed in SDS-PAGE under reducingconditions (45 kDa versus the predicted 40 kDa). The C-terminal domain containsa canonical pentraxin signature and two cysteines at amino acid positions 210 and271 of PTX3 conserved in all members of the pentraxin family.
Gel electrophoresis under native conditions demonstrates that PTX3 protomersare assembled to form multimers predominantly of 440 kDa apparent molecu-lar mass, corresponding to decamers; moreover, gel filtration reveals higher formseluting in a region corresponding to an apparent molecular mass of 900 kDa, whichsuggests that two decamers of PTX3 may associate to form an eicosamer (74). Incontrast to CRP and SAP pentamers, the decameric form of PTX3 (Figure 2)depends on interchain disulfide bonds, as demonstrated by SDS-PAGE in the ab-sence of reducing agents. A similar model was proposed for NP1 and Narp (75).
Human and murine PTX3, localized in the synthenic region of chromosome 3(q24–28), are highly conserved: Both proteins are 381 amino acids long and share82% of identical amino acids and 92% of conserved amino acids. According tomodelling, the PTX3 pentraxin domain well accommodates on the tertiary fold ofSAP, with almost all the ß-strands and the α-helical segments conserved (76).
Gene Regulation and Production
The human PTX3 proximal promoter contains Pu1, AP-1, NF-κB, Sp1, and NF-IL-6 sites. The NF-κB proximal site in the mouse and human promoter is essentialfor induction by IL-1 and TNF (77, 78). No induction of the human promoterwas observed with IL-6 (77). Therefore, analysis so far of the PTX3 proximalpromoter accounts for induction by TLR agonists, IL-1, and TNF, but not forrecently discovered costimuli (e.g., IL-10; see below).
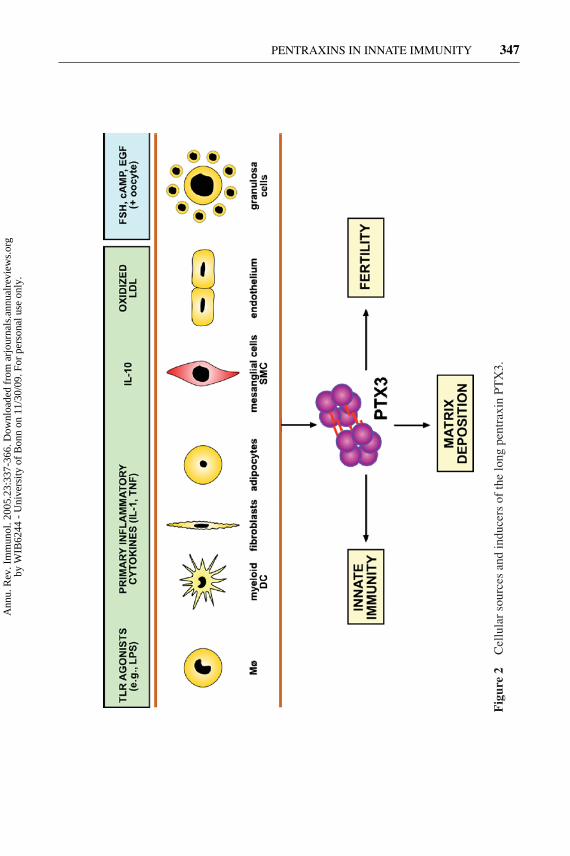
A variety of cell types can produce PTX3 in vitro upon exposure to primaryinflammatory signals (Figure 2). These include endothelial cells, smooth muscle
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.
14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
PENTRAXINS IN INNATE IMMUNITY 347
Fig
ure
2C
ellu
lar
sour
ces
and
indu
cers
ofth
elo
ngpe
ntra
xin
PTX
3.
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
348 GARLANDA ET AL.
cells (SMCs), adipocytes, fibroblasts, mononuclear phagocytes, and dendritic cells(DCs) (9, 10, 76, 79–84). We recently reassessed the relative capacity of differ-ent cell types to produce PTX3 and found that DCs produce the highest amounts(79). PTX3 production is induced by primary inflammatory signals, such as IL-1,TNF, and microbial moieties (e.g., LPS, lipoarabinomannans) (9, 85). Specifi-cally, agonists for different members of the TLR family stimulate PTX3 pro-duction. IL-6 is generally a poor inducer of PTX3 (80). However, PTX3 wasfound to be expressed in Castleman’s disease, propelled by IL-6 (86), and it isinduced in Kaposi’s sarcoma cells by viral IL-6 encoded by human herpesvirus-8(87).
IFN-γ and IL-10 have divergent effects on PTX3 production. IFN-γ inhibitsPTX3 expression and production in different cellular contexts (24, 82, 88). IL-4and IL-13 do not affect PTX3 production, whereas, surprisingly, IL-10 was foundby transcriptional profiling to induce PTX3 expression in DCs and monocytes andto costimulate PTX3 production with LPS (89; A. Doni, unpublished data). PTX3is therefore part of the genetic program expressed by M2 mononuclear phagocytesand IL-10-treated DCs. Given its role in matrix organization, PTX3 expression inthese cells is likely to be related to the orchestration of matrix deposition, tissuerepair, and remodeling (90).
DCs are major producers of PTX3 on a per-cell basis (79). Interestingly, produc-tion of PTX3 is restricted to myeloid DCs, whereas IFN-producing plasmacytoidDCs are unable to produce PTX3 in response to appropriate agonists, possiblybecause of autocrine inhibition by IFN. PTX3 production by DCs and neighbor-ing macrophages is likely to facilitate pathogen recognition and activation of anappropriate adaptive immune response (see below).
Vascular endothelial cells and SMCs produce copious amounts of PTX3 inresponse to inflammatory signals, including oxydized low density lipoproteins(ox-LDL) (9, 84) (Figure 2). PTX3-producing macrophages, endothelial cells, andSMCs have been identified in human atherosclerotic lesions (91). These resultshave provided a basis for testing the potential of PTX3 as a novel diagnostic toolin vascular disorders.
In vivo injection of LPS in mice induces PTX3 expression and high bloodlevels (76, 92). A variety of tissues express PTX3, with two aspects emerging.First, expression is most prominent in the heart and skeletal muscle. Second, incontrast to SAP, PTX3 is expressed at low levels in the liver. Unlike neuronal longpentraxins, PTX3 is not constitutively expressed in the central nervous system(CNS). Following exposure to inflammatory signals (LPS, IL-1, TNF), infectiousagents (Candida albicans, Cryptococcus neoformans), autoimmune reactions (ex-perimental allergic encephalomyelitis), or limbic seizures, PTX3 is expressed andproduced in the CNS (81, 93, 94).
Northern blot analysis revealed that ptx3 mRNA was expressed in the mouseovary during the preovulatory period, showing close temporal correlation to matrixdeposition by cumulus cells. In situ analysis revealed that ptx3 mRNA expressionwas confined to cumulus cells and to a few granulosa cells lining the follicle antrumof preovulatory follicles. Western blot and immunofluorescence analysis indicated
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
PENTRAXINS IN INNATE IMMUNITY 349
that PTX3 was associated with the extracellular matrix of wild-type cumuli. Inthe cumulus oophorus, expression of PTX3 is orchestrated by hormonal ovula-tory stimuli (follicle-stimulating hormone or human chorionic gonadotropin), byoocyte-derived soluble factors, and in particular a member of the transforminggrowth factor β (TGFβ) family, growth differentiation factor-9 (GDF-9) (95, 96)(see below).
In addition PTX3 has been identified in a variety of gene expression or proteomicprofiling efforts ranging from astrocyte secreted proteins to space flown fibroblasts(e.g., 97, 98).
Ligands
The first described and best characterized ligand of PTX3 is the complementcomponent C1q (74, 99) (Table 1). Unlike classical pentraxins, PTX3 interactswith C1q in a calcium-independent manner, without a previous aggregation of theprotein. Interaction of PTX3 with plastic-immobilized C1q induces activation ofthe classical complement pathway, as demonstrated by an increased deposition ofC3 and C4. In contrast, fluid-phase PTX3 binding to C1q inhibits complementactivation via competitive blocking of relevant interaction sites (99).
Like CRP and SAP, PTX3 binds to apoptotic cells, inhibiting their recognitionby DCs (100). Binding occurs late in the apoptotic process and, unlike classic shortpentraxins, PTX3 binding is calcium-independent. In addition, preincubation ofapoptotic cells with PTX3 enhances C1q binding and C3 deposition on the cellsurface, suggesting a role for PTX3 in the complement-mediated clearance ofapoptotic cells (99).
PTX3 also binds selected pathogens, including conidia of Aspergillus fum-igatus, Pseudomonas aeruginosa, Salmonella typhimurium, Paracoccidioidesbrasiliensis, zymosan, but not Escherichia coli, Burkholderia cepacia, Listeriamonocytogenes, or Candida albicans (101, 102).
The microbial moieties recognized by PTX3 have not been completely defined(Table 1). As expected on the basis of structural analysis (74, 76), PTX3 doesnot bind LPS; moreover, it does not bind classical short pentraxin ligands [PC,phosphoethanolamine (PE) and high pyruvate agarose]. Binding of PTX3 to A.fumigatus is competed by galactomannan (101). Mannan recognition is consistentwith binding to zymosan (102). We recently identified outer membrane proteinA (OmpA) as a bacterial moiety specifically bound by PTX3 (P. Jeannin & B.Bottazzi, unpublished observation). Matrix components (fibronectin, type IV col-lagen) are recognized by SAP but not by PTX3. In contrast, PTX3 binds the matrixcomponent TNFα-induced protein 6 (TNFAIP6 or TSG-6) (96), a multifunctionalprotein usually associated with inflammation (103, 104). By binding to TSG-6,PTX3 acts as a nodal point for the assembly of hyaluronic acid (HA)-rich extra-cellular matrix, which is essential for female fertility (96).
PTX3 binds fibroblast growth factor 2 (FGF2) but not other members of theFGF family or cytokines and chemokines (Table 1). The angiogenic activity ofFGF2 in vitro and in vivo is blocked by PTX3 (105).
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
350 GARLANDA ET AL.
Role in Innate Immunity
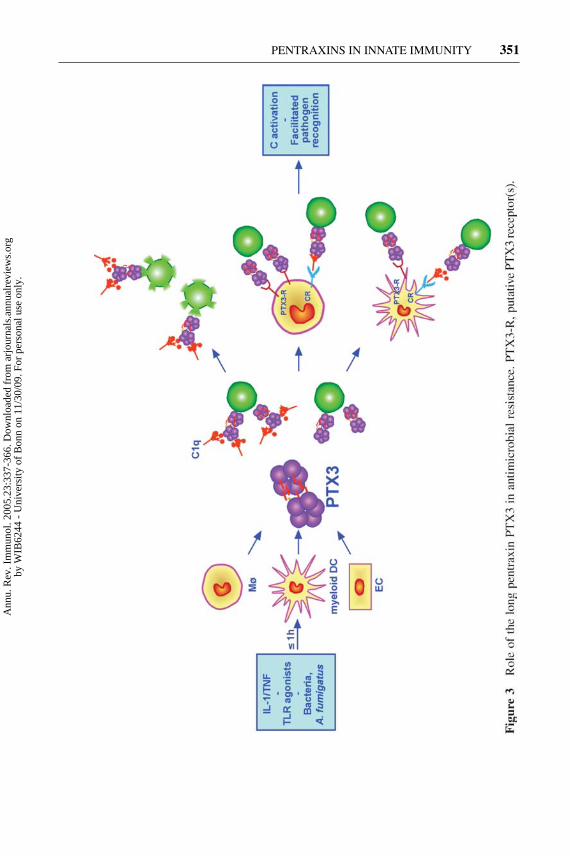
The investigation of ptx3−/− mice, generated by homologous recombination, hasbeen invaluable to understanding the in vivo function of PTX3 (96, 101). Ptx3-deficient mice are viable and display a normal life span in a conventional mousefacility. The only apparent abnormality is a severe deficiency in female fertility (seebelow).
As discussed above, PTX3 binds conidia of the opportunistic fungus A. fumiga-tus as well as selected Gram-positive and Gram-negative bacteria. Upon bindingto A. fumigatus conidia, recombinant PTX3 facilitated the internalization and thekilling of conidia, as well as the production of monocyte chemotactic protein-1 (MCP-1, also known as CCL-2) by mononuclear phagocytes. Conidia rapidlyinduced PTX3 production in vitro in murine and human mononuclear phago-cytes and DCs (Figure 3). PTX3 plasma levels were increased in mice infectedwith A. fumigatus and in neutropenic patients with hematologic malignancies andA. fumigatus systemic infection (101).
Ptx3−/− mice were extremely susceptible to invasive pulmonary aspergillosis(IPA) in terms of mortality, lung and brain colonization, and lung inflammatoryresponse. The specificity of the role played by PTX3 in the susceptibility to IPAis demonstrated by the complete protective effect of ptx3−/− mice treatment withrecombinant PTX3. The therapeutic potential of PTX3 in IPA is also suggestedby its protective effect in a murine IPA model of allogeneic, T cell–depleted bonemarrow transplantation (106).
In ptx3−/− mice, the defective recognition of A. fumigatus conidia by the host,and in particular by phagocytes and DCs, was associated with the lack of develop-ment of appropriate and protective T helper cell type 1 (Th1) antifungal responsesand with an unbalanced cytokine profile skewed toward a Th2 response. In thiscontext, the role played by PTX3 in regulating the development of detrimental Th2responses involved in allergic brochopulmonary aspergillosis needs to be furtherinvestigated.
In models of bacterial infections, such as P. aeruginosa lung infection (apathogen recognized by PTX3), ptx3−/− mice showed a partial increase in mor-tality and lung colonization. In contrast, susceptibility to L. monocytogenes and tointra-abdominal sepsis caused by caecal ligation and puncture were not affectedby PTX3 deficiency (101; C. Garlanda, unpublished results). These results suggestthat PTX3 deficiency does not cause a generalized impairment of host resistanceto microbial pathogens and that PTX3 is involved in recognition and resistanceagainst specific microorganisms.
These results demonstrate that PTX3 acts as a soluble PRR with an essentialrole in resistance against selected pathogens, in particular A. fumigatus (Figure 3).The regulated expression of this molecule in macrophages and DCs suggests thatPTX3, unlike the short pentraxins made in the liver, represents a mechanism ofamplification of innate resistance against pathogens mainly acting locally at thesite of infection and inflammation.
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.
14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
PENTRAXINS IN INNATE IMMUNITY 351
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
352 GARLANDA ET AL.
Role in Inflammation
PTX3 blood levels are low (about 25 ng/ml in mice, <2 ng/ml in humans) innormal conditions but increase rapidly (peak at 6–8 h) and dramatically (200–800 ng/ml) during endotoxic shock, sepsis, and other inflammatory conditions(107, 108).
The in vivo role of PTX3 in inflammatory conditions has been investigatedusing PTX3 transgenic mice overexpressing the murine gene under the control of itsown promoter (109) and in ptx3−/− mice generated by homologous recombination(101; C. Garlanda, unpublished results). PTX3 transgenic mice showed increasedresistance to LPS toxicity and to cecal ligation and puncture (109). Transgenicmice showed higher levels of IL-10, and macrophages were primed for increasednitric oxide production in response to IFN-γ and TNF (109).
In sharp contrast to the data on transgenic mice, ptx3−/− mice showed normalsusceptibility to LPS toxicity (in terms of mortality, cytokine levels, and PMNrecruitment in the lungs) and normal susceptibility to cecal ligation and puncture(101; C. Garlanda, unpublished). Moreover, PTX3 per se failed to affect cytokineproduction in vivo and in macrophages in vitro (C. Garlanda & B. Bottazzi, un-published observation). The reason for the divergence of results obtained in PTX3transgenic and deficient mice is not clear.
PTX3-overexpressing mice showed exacerbated inflammatory response fol-lowing intestinal ischemia reperfusion injury (110). In a model of skeletal muscleischemia and reperfusion injury, we observed PTX3 transcript induction, but wefailed to find differences between wild-type and PTX3-deficient mice in tissue dam-age, assessed as polymorphonuclear cell infiltration (myeloperoxidase activity) andCreatine Kinase activity in the injured muscle (R. Latini, unpublished results).
PTX3 is expressed in the CNS under a variety of inflammatory conditions (81,see above), including kainate-induced seizures (93). PTX3 is induced in glial cellsfollowing seizure-induced cytokine (IL-1) production, which amplifies damage.The susceptibility to kainate-induced seizures in ptx3−/− mice was similar to thatobserved in their wild-type littermates, but ptx3−/− mice had more widespreadand severe neuronal damage. These results suggest that PTX3 confers resistance toneurodegeneration, because its absence from the brain tissue is associated with anincreased number of degenerating neurons. In analogy with findings on apoptoticcells (100), PTX3 may bind to dying neurons and rescue them from otherwiseirreversible damage. Alternatively, it may interact with neuronal pentraxins andmodulate their function (see below).
PTX3 in Extracellular Matrix Architectureand Female Fertility
Ptx3−/− mice generated by homologous recombination displayed a severe defectin female fertility (95, 101). The infertility or severe subfertility of ptx3-deficientmice was tracked to an abnormal cumulus oophorus, characterized by an unstable
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
PENTRAXINS IN INNATE IMMUNITY 353
extracellular matrix in which cumulus cells were uniformly dispersed instead ofradiating out from a central oocyte (96).
As discussed above, ptx3 mRNA was expressed in the mouse ovary by cumu-lus cells during the preovulatory period, and the protein was associated with theextracellular matrix of the cumuli, where it plays a role in assembling the HA-enriched matrix of the cumulus. A crucial step of cumulus matrix assembly is thetransfer and covalent linkage of the heavy chains of serum inter α trypsin inhibitor(IαI) protein to HA or to the HA-binding glycoprotein TSG-6. Consistently, micedeficient in the light chain of IαI, bikunin, (111), or in TSG-6 (112) are infertilebecause of instability of the cumulus matrix and lack of oocyte fertilization, likeptx3−/− mice. Binding experiments performed to investigate a possible interactionbetween PTX3 and TSG-6, which is synthesized by cumulus cells in parallel withPTX3 (95), revealed that PTX3 binds to full-length human TSG-6 or to Link TSG-6 (i.e., its HA-binding domain) at a site distinct from its HA-binding surface (96).Competition studies suggest that each protomer in a PTX3 10/20-mer can bind anindividual TSG-6 molecule and, therefore, may form a multi-molecular complexthat acts as a “node” for cross-linking HA chains in the cumulus matrix, thereforeserving as an additional way of cross-linking the matrix.
Furthermore, cytofluorimetric and immunofluorescence analysis and sperm mi-gration/entrapment assays indicated that spermatozoa bound soluble and immobi-lized PTX3, suggesting that PTX3 embedded in the matrix of the cumulus couldalso direct guidance and entrapment of spermatozoa, thus facilitating the fertiliza-tion process (96).
PTX3 is expressed in the human periovulatory cumulus oophorus and is con-served between mouse and human. Therefore, it is likely that PTX3 may play thesame role in human female fertility and that PTX3 deficiency might be a cause ofunexplained infertility in women despite a normal ovulation.
Several lines of evidence point to analogies between the process of ovulationand inflammation (113, 114) and indicate that prostaglandins and cytokines playa central role in mediating the hormone regulation of cumulus matrix componentsynthesis. In addition to a hormonal ovulatory stimulus, oocyte soluble factors,and in particular GDF-9, a member of the TGFβ family (95, 115), are required foreliciting hyaluronan synthesis, cumulus expansion, and temporally and anatomi-cally restricted PTX3 expression during the periovulatory period (116, 117). Thesynthesis of PTX3 during both ovulation and inflammation adds a further elementlinking these fundamental processes (Figure 4).
PTX3 is assembled predominantly as a large multimer complex consisting oftwo decamers (74). PTX3/TSG-6 complexes might thus serve as anchoring sitesfor multiple HA molecules, thereby substantially strengthening and stabilizingthe HA network. In both physiological and pathological conditions, HA-proteininteraction is crucial for the formation and stability of extracellular matrix in severaltissues (118, 119). The finding that the long pentraxin PTX3 is a component of theextracellular matrix of the cumulus oophorus, essential for HA organization, raisesthe probability of a similar localization and function of this molecule in certain
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.
14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
354 GARLANDA ET AL.
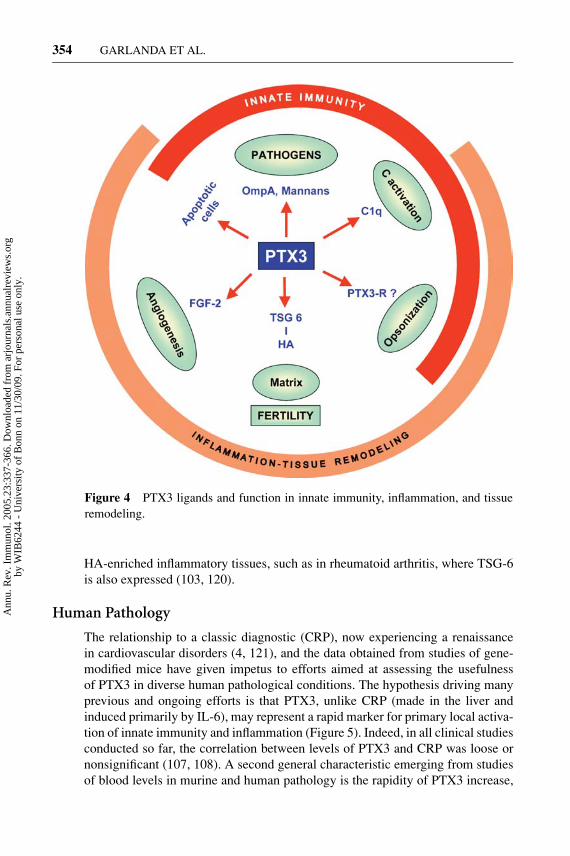
Figure 4 PTX3 ligands and function in innate immunity, inflammation, and tissueremodeling.
HA-enriched inflammatory tissues, such as in rheumatoid arthritis, where TSG-6is also expressed (103, 120).
Human Pathology
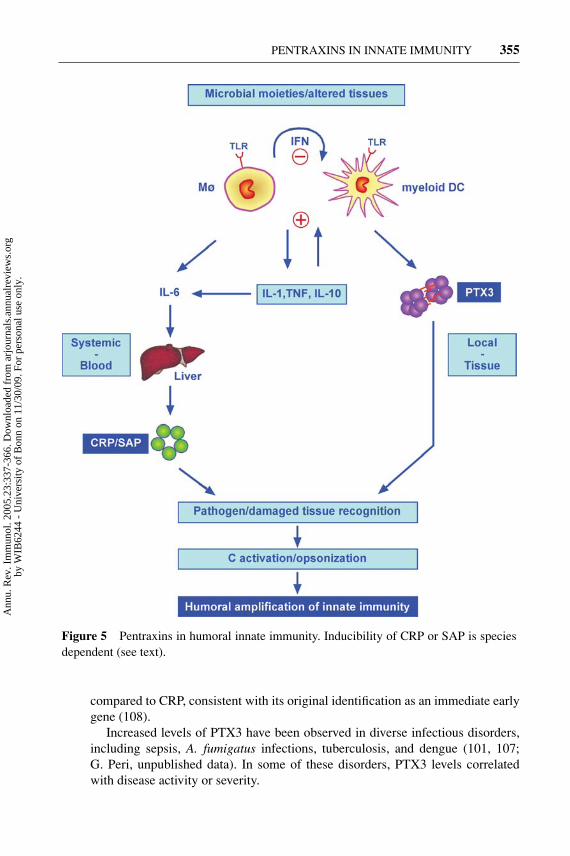
The relationship to a classic diagnostic (CRP), now experiencing a renaissancein cardiovascular disorders (4, 121), and the data obtained from studies of gene-modified mice have given impetus to efforts aimed at assessing the usefulnessof PTX3 in diverse human pathological conditions. The hypothesis driving manyprevious and ongoing efforts is that PTX3, unlike CRP (made in the liver andinduced primarily by IL-6), may represent a rapid marker for primary local activa-tion of innate immunity and inflammation (Figure 5). Indeed, in all clinical studiesconducted so far, the correlation between levels of PTX3 and CRP was loose ornonsignificant (107, 108). A second general characteristic emerging from studiesof blood levels in murine and human pathology is the rapidity of PTX3 increase,
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.
14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
PENTRAXINS IN INNATE IMMUNITY 355
Figure 5 Pentraxins in humoral innate immunity. Inducibility of CRP or SAP is speciesdependent (see text).
compared to CRP, consistent with its original identification as an immediate earlygene (108).
Increased levels of PTX3 have been observed in diverse infectious disorders,including sepsis, A. fumigatus infections, tuberculosis, and dengue (101, 107;G. Peri, unpublished data). In some of these disorders, PTX3 levels correlatedwith disease activity or severity.
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
356 GARLANDA ET AL.
Pentraxins, including SAP and PTX3, bind apoptotic cells and release nuclearcomponents, regulate their clearance, and gate the activation of autoimmunity(100). Increased levels of PTX3 have been observed in a restricted set of autoim-mune disorders (e.g., in the blood in small vessel vasculitis and in the synovial fluidin rheumatoid arthritis), but not in others (e.g., SLE, Crohn’s disease) (122, 123;G. Peri & A. Doni, unpublished results). In small vessel vasculitis, PTX3 levelscorrelate with clinical activity of the disease and represent a candidate marker formonitoring the disease (122).
Inflammation is a critical component of ischemic heart disorders, and CRP hasemerged as a valuable diagnostic tool (for a critical appraisal, see 4). The highlevels of expression in the heart during inflammatory reactions, the production byvascular cells in response to inflammatory signals (9) and oxLDL (84), and theoccurrence in atherosclerotic lesions (91) prompted studies on PTX3 levels in acutemyocardial infarction (108, 124). In a recent study with a cohort of 748 patients,PTX3, measured along with established markers including CRP, emerged as theonly independent predictor of mortality (124). It remains to be elucidated whetherthis impressive correlation with outcome actually reflects a role in the pathogenesisof damage, for instance by amplifying the complement and coagulation cascades(125, 126).
Results in gene-targeted mice have revealed a nonredundant function of PTX3in female fertility (95, 96). PTX3 is produced and present in the human cumu-lus oophorus, and it was recently suggested that cumulus PTX3 transcript levelscorrelate with oocyte quality (127).
PTX3 is a humoral effector molecule of innate immunity; it binds selectedpathogens, activates complement, facilitates cellular recognition and disposal, andacts as a matrix component. The molecular partner of PTX3 in the matrix iden-tified so far (Figures 4 and 5), TSG-6, is a candidate chondroprotective agent inrheumatoid arthritis (103). Current research suggests that PTX3 may well fulfill asimilar function.
PTX3 synergizes with conventional antifungal agents in the treatment ofA. fumigatus infections (106). This recent finding reinforces the hypothesis ofPTX3 as a candidate new agent for the treatment or profylaxis of A. fumigatusinfections in immunocompromised patients.
OTHER LONG PENTRAXINS
Researchers have identified a set of long pentraxins in neuronal tissues (Figure1). NP1 (or NPTXI) was originally identified as a protein binding taipoxin, thepresynaptic-acting snake venom neurotoxin (11). NP2 (or NPTXII) was cloned bylow stringency hybridization (12) or as Narp, an early gene induced by physiologicsynaptic activity (16). Unlike NP1, NP2/Narp is expressed in a wide range of non-nervous tissues, prominently in the testis (12). NP2 has homology with apexin,a guinea pig long pentraxin identified in the acrosome of spermatozoa (14, 23).
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
PENTRAXINS IN INNATE IMMUNITY 357
Recent analysis suggests that apexin is not the guinea pig ortholog of NP2 (Y.Martinez, unpublished data). NPR is an integral membrane pentraxin, coexpressedin the brain with NP1, that binds NP1 and taipoxin (17, 18).
Various functions have been attributed to neuronal pentraxins. On the basis ofits binding to taipoxin, NP1 was postulated to have a role in synaptic uptake. NP1 isinduced during low K+-induced apoptotic death of cerebellar granule cells (128)and during hypoxic-ischemic injury of the neonatal brain (129). NP1 antisensenucleotides protect neuronal cells against death induced by hypoxia or by the glu-tamate analog AMPA (α-amino-3-hydroxy-5-methyl-4 isoxazole-propionic acid)(129).
Narp, which binds agar (16), and NP1 form heterocomplexes in the brain via co-valent links involving cysteines in the N-terminal, nonpentraxin domain, as PTX3does. The pentraxin domains associate with AMPA-type gutamate receptors. NP1and Narp together have superadditive synaptogenic activity. These two neuronalpentraxins, and possibly NPR, would contribute to activity-dependent (Narp) and-independent (NP1) synaptogenesis and to synaptic plasticity (75). NP1-deficientmice have been generated but show no apparent phenotype (75). This negativefinding may reflect redundancy of neuronal pentraxins.
PTX3 is expressed by neuronal cells and astrocytes following exposure to in-fectious agents, inflammatory cytokines, stroke, experimental autoimmune en-cephalomyelitis, and limbic seizures (81, 93, 94). In limbic seizures, gene-targetedmice revealed a protective role for PTX3 in seizure-induced neurodegeneration(93). It remains to be established whether PTX3 can form heterocomplexes withneuronal pentraxins and regulate their function.
Scientists have identified pentraxin-like domains in diverse molecules expressedin peripheral tissues. For instance, vascular inducible G protein–coupled receptor(GPCR, VIGR) is induced by LPS or thrombin in endothelial cells and consists ofan N-terminal pentraxin-like domain on top of an adhesin-type GPCR (130). Simi-larly, the ectodomain of very large GPCR1 (VLGR-1) contains a pentraxin domain.Interestingly, VLGR-1 is expressed in the developing brain (131). The function ofthese and other pentraxin-like domain-containing molecules is at present unknown.
OVERVIEW AND PERSPECTIVE
Pentraxins are evolutionary conserved humoral PRRs. Strong evidence, includingevidence from studies of gene-modified mice, suggests that at least some of thesemolecules play a nonredundant role in innate resistance against microbes. Avail-able information suggests that the short pentraxins CRP and SAP and the longpentraxin PTX3 have different ligand specificity (Table 1). Thus, pentraxins pro-vide the innate immune system with a repertoire of diverse receptors with distinctspecificity.
In terms of regulation of production, members of the pentraxin family can bedistinguished as inducible (or inflammatory) and constitutive (or homeostatic).
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
358 GARLANDA ET AL.
Constitutive versus inducible production may vary in different species, as dis-cussed above for CRP and SAP in humans versus rodents. Copious amounts ofthe short pentraxins CRP and SAP are made in the liver as the end result of thepathogen recognition–cytokine cascade (Figure 5). In contrast, long pentraxinsare generally expressed at extrahepatic sites, and the prototypic molecule PTX3 isdirectly induced by TLR engagement and primary inflammatory cytokines. There-fore, liver-derived short pentraxins and tissue-expressed long pentraxins (PTX3)are likely to fulfill complementary functions, acting primarly at a systemic andlocal tissue level, respectively (Figure 5).
The regulation of production and effector functions of at least some mem-bers of the pentraxin superfamily (e.g., CRP or PTX3) are strongly reminescentof antibodies. Their production is induced by recognition via TLR of microbialnonself or altered self (oxLDL), directly (PTX3) or indirectly (short pentraxins)(Figure 5). Short pentraxins and PTX3 recognize microbial structures or alteredself (apoptotic cells), activate complement, and facilitate recognition and disposalby macrophages, thus acting as a humoral amplification loop of innate immunity.Therefore, inducible pentraxins act as bona fide functional proto-antibodies.
Innate resistance to pathogens is one of the many facets of pentraxins. PTX3is divergently regulated by IFN-γ and IL-10 and is essential for activation of apolarized type 1 response to A. fumigatus (89, 101). Pentraxins bind apoptoticcells, nucleosomes, and nuclear components, and SAP-deficient mice are proneto develop autoimmunity (73). Therefore, as for other innate immunity receptors,pentraxins can contribute to the orientation of adaptive immunity and to the vetoingof autoimmunity.
Members of the pentraxin family are involved in neuronal function (NP1, NP2,NPR), regulation of neurodegeneration (NP1, PTX3, SAP), construction of extra-cellular matrices (PTX3, SAP), and female fertility (PTX3). The latter two relatedfunctions, rather than immunity, most likely underlie the high conservation inmammals of PTX3. Therefore, pentraxins are soluble multifunctional PRRs at thecrossroads between innate and adaptative immunity, inflammation, matrix deposi-tion, and female fertility. It will be important to assess whether, in analogy to CRPand SAP, the newly discovered long pentraxins can contribute to clinical medicineas diagnostic or therapeutic agents.
ACKNOWLEDGMENTS
This work was supported by Associazione Italiana per la Ricerca sul Cancro(AIRC), Ministero Istruzione Universita e Ricerca (MIUR), CNR, European Com-mission. We wish to thank people who, over the years, were part of the Pen-traxin Project in our department (in particular Giuseppe Peri, Andrea Doni, andAnnunciata Vecchi) and external collaborators (in particular Luigina Romani,Roberto Latini, Antonia Salustri, and Giovanni Salvatori) for invaluable help,suggestions, and illuminating discussions. We thank Yeny Martinez for preparingFigure 1B.
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
PENTRAXINS IN INNATE IMMUNITY 359
The Annual Review of Immunology is online athttp://immunol.annualreviews.org
LITERATURE CITED
1. Medzhitov R. 2001. Toll-like receptorsand innate immunity. Nat. Rev. Immunol.1:135–45
2. Gordon S. 2002. Pattern recognition re-ceptors: doubling up for the innate im-mune response. Cell 111:927–30
3. Szalai AJ, Agrawal A, Greenhough TJ,Volanakis JE. 1999. C-reactive protein:structural biology and host defense func-tion. Clin. Chem. Lab. Med. 37:265–70
4. Pepys MB, Hirschfield GM. 2003. C-reactive protein: a critical update. J. Clin.Invest. 111:1805–12
5. Pepys MB, Baltz ML. 1983. Acute phaseproteins with special reference to C-reactive protein and related proteins (pen-taxins) and serum amyloid A protein. Adv.Immunol. 34:141–212
6. Mantovani A, Garlanda C, Bottazzi B.2003. Pentraxin 3, a non-redundant solu-ble pattern recognition receptor involvedin innate immunity. Vaccine 21(Suppl.2):S43–47
7. Tillet WS, Francis T Jr . 1930. Serologicalreactions in pneumonia with a non proteinsomatic fraction of pneumococcus. J. Exp.Med. 52:561–85
8. Abernethy TJ, Avery OT. 1941. The oc-curence during acute infections of a pro-tein not normally present in the blood. I.Distribution of the reactive protein in pa-tients’ sera and the effect of calcium onthe flocculation reaction with C. polysac-charide of pneumococcus. J. Exp. Med.73:173–82
9. Breviario F, d’Aniello EM, Golay J, PeriG, Bottazzi B, et al. 1992. Interleukin-1-inducible genes in endothelial cells.Cloning of a new gene related to C-reactive protein and serum amyloid Pcomponent. J. Biol. Chem. 267:22190–97
10. Lee GW, Lee TH, Vilcek J. 1993. TSG-
14, a tumor necrosis factor- and IL-1-inducible protein, is a novel member ofthe pentaxin family of acute phase pro-teins. J. Immunol. 150:1804–12
11. Schlimgen AK, Helms JA, Vogel H, PerinMS. 1995. Neuronal pentraxin, a secretedprotein with homology to acute phaseproteins of the immune system. Neuron14:519–26
12. Hsu YC, Perin MS. 1995. Human neu-ronal pentraxin II (NPTX2): conservation,genomic structure, and chromosomal lo-calization. Genomics 28:220–27
13. Omeis IA, Hsu YC, Perin MS. 1996.Mouse and human neuronal pentraxin 1(NPTX1): conservation, genomic struc-ture, and chromosomal localization. Ge-nomics 36:543–45
14. Noland TD, Friday BB, Maulit MT, Ger-ton GL. 1994. The sperm acrosomalmatrix contains a novel member of thepentaxin family of calcium-dependentbinding proteins. J. Biol. Chem. 269:32607–14
15. Seery LT, Schoenberg DR, Barbaux S,Sharp PM, Whitehead AS. 1993. Identifi-cation of a novel member of the pentraxinfamily in Xenopus laevis. Proc. R. Soc.London Ser. B 253:263–70
16. Tsui CC, Copeland NG, Gilbert DJ, Jenk-ins NA, Barnes C, Worley PF. 1996. Narp,a novel member of the pentraxin family,promotes neurite outgrowth and is dynam-ically regulated by neuronal activity. J.Neurosci. 16:2463–78
17. Dodds DC, Omeis IA, Cushman SJ,Helms JA, Perin MS. 1997. Neuronal pen-traxin receptor, a novel putative integralmembrane pentraxin that interacts withneuronal pentraxin 1 and 2 and taipoxin-associated calcium-binding protein 49. J.Biol. Chem. 272:21488–94
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
360 GARLANDA ET AL.
18. Kirkpatrick LL, Matzuk MM, Dodds DC,Perin MS. 2000. Biochemical interactionsof the neuronal pentraxins. Neuronal pen-traxin (NP) receptor binds to taipoxin andtaipoxin-associated calcium-binding pro-tein 49 via NP1 and NP2. J. Biol. Chem.275:17786–92
19. Gewurz H, Zhang XH, Lint TF. 1995.Structure and function of the pentraxins.Curr. Opin. Immunol. 7:54–64
20. Shrive AK, Metcalfe AM, Cartwright JR,Greenhough TJ. 1999. C-reactive proteinand SAP-like pentraxin are both present inLimulus polyphemus haemolymph: crys-tal structure of Limulus SAP. J. Mol. Biol.290:997–1008
21. Armstrong PB, Swarnakar S, Srimal S,Misquith S, Hahn EA, et al. 1996. A cy-tolytic function for a sialic acid-bindinglectin that is a member of the pen-traxin family of proteins. J. Biol. Chem.271:14717–21
22. Liu TY, Robey FA, Wang CM. 1982.Structural studies on C-reactive protein.Ann. NY Acad. Sci. 389:151–62
23. Reid MS, Blobel CP. 1994. Apexin, anacrosomal pentaxin. J. Biol. Chem. 269:32615–20
24. Goodman AR, Cardozo T, Abagyan R,Altmeyer A, Wisniewski HG, VilcekJ. 1996. Long pentraxins: an emerginggroup of proteins with diverse functions.Cytokine Growth Factor Rev. 7:191–202
25. Kretzschmar D, Hasan G, Sharma S,Heisenberg M, Benzer S. 1997. The swisscheese mutant causes glial hyperwrappingand brain degeneration in Drosophila. J.Neurosci. 17:7425–32
26. Rubio N, Sharp PM, Rits M, Zahedi K,Whitehead AS. 1993. Structure, expres-sion, and evolution of guinea pig serumamyloid P component and C-reactive pro-tein. J. Biochem. 113:277–84
27. Shrive AK, Cheetham GM, Holden D,Myles DA, Turnell WG, et al. 1996.Three dimensional structure of human C-reactive protein. Nat. Struct. Biol. 3:346–54
28. Toniatti C, Demartis A, Monaci P, NicosiaA, Ciliberto G. 1990. Synergistic trans-activation of the human C-reactive proteinpromoter by transcription factor HNF-1binding at two distinct sites. EMBO J.9:4467–75
29. Li SP, Goldman ND. 1996. Regulation ofhuman C-reactive protein gene expressionby two synergistic IL-6 responsive ele-ments. Biochemistry 35:9060–68
30. Ramji DP, Vitelli A, Tronche F, CorteseR, Ciliberto G. 1993. The two C/EBPisoforms, IL-6DBP/NF-IL6 and C/EBPδ/NF-IL6 β, are induced by IL-6 to pro-mote acute phase gene transcription viadifferent mechanisms. Nucleic Acids Res.21:289–94
31. Zhang D, Sun M, Samols D, KushnerI. 1996. STAT3 participates in transcrip-tional activation of the C-reactive pro-tein gene by interleukin-6. J. Biol. Chem.271:9503–9
32. Cha-Molstad H, Agrawal A, Zhang D,Samols D, Kushner I. 2000. The Relfamily member P50 mediates cytokine-induced C-reactive protein expression bya novel mechanism. J. Immunol. 165:4592–97
33. Agrawal A, Cha-Molstad H, Samols D,Kushner I. 2001. Transactivation of C-reactive protein by IL-6 requires synergis-tic interaction of CCAAT/enhancer bind-ing protein β (C/EBP β) and Rel p50. J.Immunol. 166:2378–84
34. Zhang D, Jiang SL, Rzewnicki D, SamolsD, Kushner I. 1995. The effect ofinterleukin-1 on C-reactive protein ex-pression in Hep3B cells is exerted at thetranscriptional level. Biochem. J. 310(Part1):143–48
35. Emsley J, White HE, O’Hara BP, OlivaG, Srinivasan N, et al. 1994. Structure ofpentameric human serum amyloid P com-ponent. Nature 367:338–45
36. Zahedi K. 1997. Characterization of thebinding of serum amyloid P to laminin. J.Biol. Chem. 272:2143–48
37. Aquilina JA, Robinson CV. 2003.
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
PENTRAXINS IN INNATE IMMUNITY 361
Investigating interactions of the pentrax-ins serum amyloid P component andC-reactive protein by mass spectrometry.Biochem. J. 375:323–28
38. Pepys MB, Rademacher TW, Ama-tayakul-Chantler S, Williams P, NobleGE, et al. 1994. Human serum amyloidP component is an invariant constituentof amyloid deposits and has a uniquelyhomogeneous glycostructure. Proc. Natl.Acad. Sci. USA 91:5602–6
39. Hutchinson WL, Hohenester E, PepysMB. 2000. Human serum amyloid P com-ponent is a single uncomplexed pentamerin whole serum. Mol. Med. 6:482–93
40. Szalai AJ. 2002. The antimicrobial activ-ity of C-reactive protein. Microbes Infect.4:201–5
41. Hind CR, Collins PM, Renn D, Cook RB,Caspi D, et al. 1984. Binding specificityof serum amyloid P component for thepyruvate acetal of galactose. J. Exp. Med.159:1058–69
42. Noursadeghi M, Bickerstaff MC, Gal-limore JR, Herbert J, Cohen J, Pepys MB.2000. Role of serum amyloid P compo-nent in bacterial infection: protection ofthe host or protection of the pathogen.Proc. Natl. Acad. Sci. USA 97:14584–89
43. Hind CR, Collins PM, Baltz ML, PepysMB. 1985. Human serum amyloid P com-ponent, a circulating lectin with speci-ficity for the cyclic 4,6-pyruvate acetal ofgalactose. Interactions with various bac-teria. Biochem. J. 225:107–11
44. Andersen O, Vilsgaard Ravn K, JuulSorensen I, Jonson G, Holm Nielsen E,Svehag SE. 1997. Serum amyloid P com-ponent binds to influenza A virus haemag-glutinin and inhibits the virus infection invitro. Scand. J. Immunol. 46:331–37
45. de Haas CJ, van der Tol ME, Van KesselKP, Verhoef J, Van Strijp JA. 1998.A synthetic lipopolysaccharide-bindingpeptide based on amino acids 27–39of serum amyloid P component inhibitslipopolysaccharide-induced responses inhuman blood. J. Immunol. 161:3607–15
46. Loveless RW, Floyd-O’Sullivan G,Raynes JG, Yuen CT, Feizi T. 1992.Human serum amyloid P is a multi-specific adhesive protein whose ligandsinclude 6-phosphorylated mannose andthe 3-sulphated saccharides galactose,N-acetylgalactosamine and glucuronicacid. EMBO J. 11:813–19
47. Pepys MB, Butler PJ. 1987. Serum amy-loid P component is the major calcium-dependent specific DNA binding proteinof the serum. Biochem. Biophys. Res.Commun. 148:308–13
48. Butler PJ, Tennent GA, Pepys MB. 1990.Pentraxin-chromatin interactions: serumamyloid P component specifically dis-places H1-type histones and solubilizesnative long chromatin. J. Exp. Med. 172:13–18
49. Nauta AJ, Daha MR, van Kooten C, RoosA. 2003. Recognition and clearance ofapoptotic cells: a role for complementand pentraxins. Trends Immunol. 24:148–54
50. Jiang H, Robey FA, Gewurz H. 1992.Localization of sites through which C-reactive protein binds and activates com-plement to residues 14–26 and 76–92 ofthe human C1q A chain. J. Exp. Med. 175:1373–79
51. Ying SC, Gewurz AT, Jiang H, GewurzH. 1993. Human serum amyloid P compo-nent oligomers bind and activate the clas-sical complement pathway via residues14–26 and 76–92 of the A chain collagen-like region of C1q. J. Immunol. 150:169–76
52. Gershov D, Kim S, Brot N, ElkonKB. 2000. C-Reactive protein binds toapoptotic cells, protects the cells fromassembly of the terminal complementcomponents, and sustains an antiinflam-matory innate immune response: implica-tions for systemic autoimmunity. J. Exp.Med. 192:1353–64
53. Familian A, Zwart B, Huisman HG,Rensink I, Roem D, et al. 2001.Chromatin-independent binding of serum
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
362 GARLANDA ET AL.
amyloid P component to apoptotic cells.J. Immunol. 167:647–54
54. Bharadwaj D, Stein MP, Volzer M, MoldC, Du Clos TW. 1999. The major recep-tor for C-reactive protein on leukocytes isFcγ receptor II. J. Exp. Med. 190:585–90
55. Bharadwaj D, Mold C, Markham E, DuClos TW. 2001. Serum amyloid P com-ponent binds to Fcγ receptors and op-sonizes particles for phagocytosis. J. Im-munol. 166:6735–41
56. Mold C, Baca R, Du Clos TW. 2002.Serum amyloid P component and C-reactive protein opsonize apoptotic cellsfor phagocytosis through Fcγ receptors.J. Autoimmun. 19:147–54
57. Mold C, Gresham HD, Du Clos TW.2001. Serum amyloid P component andC-reactive protein mediate phagocyto-sis through murine Fcγ Rs. J. Immunol.166:1200–5
58. Hundt M, Zielinska-Skowronek M,Schmidt RE. 2001. Lack of specificreceptors for C-reactive protein on whiteblood cells. Eur. J. Immunol. 31:3475–83
59. Saeland E, van Royen A, Hendriksen K,Vile-Weekhout H, Rijkers GT, et al. 2001.Human C-reactive protein does not bindto Fcγ RIIa on phagocytic cells. J. Clin.Invest. 107:641–43
60. Szalai AJ, Briles DE, Volanakis JE. 1995.Human C-reactive protein is protectiveagainst fatal Streptococcus pneumoniaeinfection in transgenic mice. J. Immunol.155:2557–63
61. Weiser JN, Pan N, McGowan KL, MusherD, Martin A, Richards J. 1998. Phos-phorylcholine on the lipopolysaccharideof Haemophilus influenzae contributes topersistence in the respiratory tract andsensitivity to serum killing mediated byC-reactive protein. J. Exp. Med. 187:631–40
62. Lysenko E, Richards JC, Cox AD, Stew-art A, Martin A, et al. 2000. The positionof phosphorylcholine on the lipopolysac-charide of Haemophilus influenzae af-
fects binding and sensitivity to C-reactiveprotein-mediated killing. Mol. Microbiol.35:234–45
63. Szalai AJ, VanCott JL, McGhee JR,Volanakis JE, Benjamin WH Jr. 2000.Human C-reactive protein is protectiveagainst fatal Salmonella enterica serovartyphimurium infection in transgenic mice.Infect. Immun. 68:5652–56
64. Szalai AJ, Nataf S, Hu XZ, BarnumSR. 2002. Experimental allergic en-cephalomyelitis is inhibited in transgenicmice expressing human C-reactive pro-tein. J. Immunol. 168:5792–97
65. Du Clos TW, Zlock LT, Hicks PS, MoldC. 1994. Decreased autoantibody levelsand enhanced survival of (NZB × NZW)F1 mice treated with C-reactive protein.Clin. Immunol. Immunopathol. 70:22–27
66. Szalai AJ, Weaver CT, McCrory MA, vanGinkel FW, Reiman RM, et al. 2003. De-layed lupus onset in (NZB × NZW)F1mice expressing a human C-reactive pro-tein transgene. Arthritis Rheum. 48:1602–11
67. Xia D, Samols D. 1997. Transgenic miceexpressing rabbit C-reactive protein areresistant to endotoxemia. Proc. Natl.Acad. Sci. USA 94:2575–80
68. Mold C, Rodriguez W, Rodic-Polic B, DuClos TW. 2002. C-reactive protein me-diates protection from lipopolysaccharidethrough interactions with Fcγ R. J. Im-munol. 169:7019–25
69. Hirschfield GM, Herbert J, Kahan MC,Pepys MB. 2003. Human C-reactiveprotein does not protect against acutelipopolysaccharide challenge in mice. J.Immunol. 171:6046–51
70. Botto M, Hawkins PN, Bickerstaff MC,Herbert J, Bygrave AE, et al. 1997. Amy-loid deposition is delayed in mice withtargeted deletion of the serum amyloid Pcomponent gene. Nat. Med. 3:855–59
71. Tennent GA, Lovat LB, Pepys MB.1995. Serum amyloid P component pre-vents proteolysis of the amyloid fibrils
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
PENTRAXINS IN INNATE IMMUNITY 363
of Alzheimer disease and systemic amy-loidosis. Proc. Natl. Acad. Sci. USA 92:4299–303
72. Pepys MB, Herbert J, Hutchinson WL,Tennent GA, Lachmann HJ, et al. 2002.Targeted pharmacological depletion ofserum amyloid P component for treatmentof human amyloidosis. Nature 417:254–59
73. Bickerstaff MC, Botto M, HutchinsonWL, Herbert J, Tennent GA, et al. 1999.Serum amyloid P component controlschromatin degradation and prevents antin-uclear autoimmunity. Nat. Med. 5:694–97
74. Bottazzi B, Vouret-Craviari V, Bastone A,De Gioia L, Matteucci C, et al. 1997. Mul-timer formation and ligand recognitionby the long pentraxin PTX3. Similaritiesand differences with the short pentraxinsC-reactive protein and serum amyloid Pcomponent. J. Biol. Chem. 272:32817–23
75. Xu DS, Hopf C, Reddy R, Cho RW, GuoLP, et al. 2003. Narp and NP1 form hetero-complexes that function in developmentaland activity-dependent synaptic plasticity.Neuron 39:513–28
76. Introna M, Alles VV, Castellano M, Pi-cardi G, De Gioia L, et al. 1996. Cloningof mouse ptx3, a new member of the pen-traxin gene family expressed at extrahep-atic sites. Blood 87:1862–72
77. Basile A, Sica A, d’Aniello E, BreviarioF, Garrido G, et al. 1997. Characteriza-tion of the promoter for the human longpentraxin PTX3. Role of NF-κB in tumornecrosis factor-α and interleukin-1β reg-ulation. J. Biol. Chem. 272:8172–78
78. Altmeyer A, Klampfer L, Goodman AR,Vilcek J. 1995. Promoter structure andtranscriptional activation of the murineTSG-14 gene encoding a tumor necrosisfactor/interleukin-1-inducible pentraxinprotein. J. Biol. Chem. 270:25584–90
79. Doni A, Peri G, Chieppa M, Allavena P,Pasqualini F, et al. 2003. Production of thesoluble pattern recognition receptor PTX3by myeloid, but not plasmacytoid, den-dritic cells. Eur. J. Immunol. 33:2886–93
80. Alles VV, Bottazzi B, Peri G, Golay J,Introna M, Mantovani A. 1994. Inducibleexpression of PTX3, a new member of thepentraxin family, in human mononuclearphagocytes. Blood 84:3483–93
81. Polentarutti N, Bottazzi B, Di Santo E,Blasi E, Agnello D, et al. 2000. Inducibleexpression of the long pentraxin PTX3 inthe central nervous system. J. Neuroim-munol. 106:87–94
82. Goodman AR, Levy DE, Reis LF, VilcekJ. 2000. Differential regulation of TSG-14 expression in murine fibroblasts andperitoneal macrophages. J. Leukoc. Biol.67:387–95
83. Abderrahim-Ferkoune A, Bezy O, Chiell-ini C, Maffei M, Grimaldi P, et al. 2003.Characterization of the long pentraxinPTX3 as a TNF α-induced secreted pro-tein of adipose cells. J. Lipid Res. 44:994–1000
84. Klouche M, Peri G, Knabbe C, Eck-stein HH, Schmid FX, et al. 2004.Modified atherogenic lipoproteins induceexpression of pentraxin-3 by human vas-cular smooth muscle cells. Atherosclero-sis 175:221–28
85. Vouret-Craviari V, Matteucci C, Peri G,Poli G, Introna M, Mantovani A. 1997.Expression of a long pentraxin, PTX3, bymonocytes exposed to the mycobacterialcell wall component lipoarabinomannan.Infect. Immun. 65:1345–50
86. Malaguarnera L, Pilastro MR, Vicari L,Di Marco R, Malaguarnera M, MessinaA. 2000. PTX3 gene expression in Castle-man’s disease. Eur. J. Haematol. 64:132–34
87. Klouche M, Brockmeyer N, Knabbe C,Rose-John S. 2002. Human herpesvirus8-derived viral IL-6 induces PTX3 ex-pression in Kaposi’s sarcoma cells. AIDS16:F9–18
88. Polentarutti N, Picardi G, Basile A, Cen-zuales S, Rivolta A, et al. 1998. Interferon-γ inhibits expression of the long pentraxinPTX3 in human monocytes. Eur. J. Im-munol. 28:496–501
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
364 GARLANDA ET AL.
89. Perrier P, Martinez FO, Locati M,Bianchi G, Nebuloni M, et al. 2004.Distinct transcriptional programs acti-vated by interleukin-10 with or withoutlipopolysaccharide in dendritic cells:induction of the B cell-activatingchemokine, CXC chemokine ligand 13.J. Immunol. 172:7031–42
90. Mantovani A, Sozzani S, Locati M,Allavena P, Sica A. 2002. Macro-phage polarization: tumor-associatedmacrophages as a paradigm for polarizedM2 mononuclear phagocytes. TrendsImmunol. 23:549–55
91. Rolph MS, Zimmer S, Bottazzi B, Gar-landa C, Mantovani A, Hansson GK.2002. Production of the long pen-traxin PTX3 in advanced atheroscleroticplaques. Arterioscler. Thromb. Vasc. Biol.22:E10–18
92. Lee GW, Goodman AR, Lee TH, Vilcek J.1994. Relationship of TSG-14 protein tothe pentraxin family of major acute phaseproteins. J. Immunol. 153:3700–7
93. Ravizza T, Moneta D, Bottazzi B, Peri G,Garlanda C, et al. 2001. Dynamic induc-tion of the long pentraxin PTX3 in theCNS after limbic seizures: evidence fora protective role in seizure-induced neu-rodegeneration. Neuroscience 105:43–53
94. Agnello D, Carvelli L, Muzio V, VillaP, Bottazzi B, et al. 2000. Increasedperipheral benzodiazepine binding sitesand pentraxin 3 expression in the spinalcord during EAE: relation to inflamma-tory cytokines and modulation by dexam-ethasone and rolipram. J. Neuroimmunol.109:105–11
95. Varani S, Elvin JA, Yan CN, DeMayo J,DeMayo FJ, et al. 2002. Knockout of pen-traxin 3, a downstream target of growthdifferentiation factor-9, causes femalesubfertility. Mol. Endocrinol. 16:1154–67
96. Salustri A, Garlanda C, Hirsch E, DeAcetis M, Maccagno A, et al. 2004. PTX3plays a key role in the organization ofthe cumulus oophorus extracellular ma-
trix and in in vivo fertilization. Develop-ment 131:1577–86
97. Lafon-Cazal M, Adjali O, Galeotti N,Poncet J, Jouin P, et al. 2003. Pro-teomic analysis of astrocytic secretion inthe mouse. Comparison with the cere-brospinal fluid proteome. J. Biol. Chem.278:24438–48
98. Semov A, Semova N, Lacelle C, MarcotteR, Petroulakis E, et al. 2002. Alterationsin TNF- and IL-related gene expressionin space-flown WI38 human fibroblasts.FASEB J. 16:899–901
99. Nauta AJ, Bottazzi B, Mantovani A, Sal-vatori G, Kishore U, et al. 2003. Biochem-ical and functional characterization of theinteraction between pentraxin 3 and C1q.Eur. J. Immunol. 33:465–73
100. Rovere P, Peri G, Fazzini F, Bottazzi B,Doni A, et al. 2000. The long pentraxinPTX3 binds to apoptotic cells and regu-lates their clearance by antigen-presentingdendritic cells. Blood 96:4300–6
101. Garlanda C, Hirsch E, Bozza S, Salustri A,De Acetis M, et al. 2002. Non-redundantrole of the long pentraxin PTX3 in anti-fungal innate immune response. Nature420:182–86
102. Diniz SN, Nomizo R, Cisalpino PS, Teix-eira MM, Brown GD, et al. 2004. PTX3function as an opsonin for the dectin-1-dependent internalization of zymosan bymacrophages. J. Leukoc. Biol. 75:649–56
103. Milner CM, Day AJ. 2003. TSG-6: a mul-tifunctional protein associated with in-flammation. J. Cell Sci. 116:1863–73
104. Wisniewski HG, Vilcek J. 2004.Cytokine-induced gene expression atthe crossroads of innate immunity,inflammation and fertility: TSG-6 andPTX3/TSG-14. Cytokine Growth FactorRev. 15:129–46
105. Rusnati M, Camozzi M, Moroni E, Bot-tazzi B, Peri G, et al. 2004. Selectiverecognition of fibroblast growth factor-2by the long pentraxin PTX3 inhibits an-giogenesis. Blood 104:92–99
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
PENTRAXINS IN INNATE IMMUNITY 365
106. Gaziano R, Bozza S, Bellocchio S, Perruc-cio K, Montagnoli C, et al. 2004. Com-bination therapy with pentraxin 3 andantifungals in experimental aspergillosis.Antimicrob. Agents Chemother. 48:4414–21
107. Muller B, Peri G, Doni A, Torri V, Land-mann R, et al. 2001. Circulating levels ofthe long pentraxin PTX3 correlate withseverity of infection in critically ill pa-tients. Crit. Care Med. 29:1404–7
108. Peri G, Introna M, Corradi D, Iacuitti G,Signorini S, et al. 2000. PTX3, a proto-typic long pentraxin, is an early indica-tor of acute myocardial infarction in man.Circulation 102:636–41
109. Dias AA, Goodman AR, Dos Santos JL,Gomes RN, Altmeyer A, et al. 2001. TSG-14 transgenic mice have improved sur-vival to endotoxemia and to CLP-inducedsepsis. J. Leukoc. Biol. 69:928–36
110. Souza DG, Soares AC, Pinho V, TorloniH, Reis LF, et al. 2002. Increased mor-tality and inflammation in tumor necro-sis factor-stimulated gene-14 transgenicmice after ischemia and reperfusion in-jury. Am. J. Pathol. 160:1755–65
111. Zhuo L, Yoneda M, Zhao M, YingsungW, Yoshida N, et al. 2001. Defect inSHAP-hyaluronan complex causes severefemale infertility. A study by inactivationof the bikunin gene in mice. J. Biol. Chem.276:7693–96
112. Fulop C, Szanto S, Mukhopadhyay D,Bardos T, Kamath RV, et al. 2003. Im-paired cumulus mucification and femalesterility in tumor necrosis factor-inducedprotein-6 deficient mice. Development130:2253–61
113. Richards JS, Russell DL, Ochsner S, Es-pey LL. 2002. Ovulation: new dimensionsand new regulators of the inflammatory-like response. Annu. Rev. Physiol. 64:69–92
114. Espey LL. 1994. Current status of thehypothesis that mammalian ovulation iscomparable to an inflammatory reaction.Biol. Reprod. 50:233–38
115. Elvin JA, Clark AT, Wang P, WolfmanNM, Matzuk MM. 1999. Paracrine ac-tions of growth differentiation factor-9 inthe mammalian ovary. Mol. Endocrinol.13:1035–48
116. Salustri A, Yanagishita M, Hascall VC.1990. Mouse oocytes regulate hyaluronicacid synthesis and mucification by FSH-stimulated cumulus cells. Dev. Biol. 138:26–32
117. Buccione R, Vanderhyden BC, Caron PJ,Eppig JJ. 1990. FSH-induced expansionof the mouse cumulus oophorus in vitrois dependent upon a specific factor(s) se-creted by the oocyte. Dev. Biol. 138:16–25
118. Day AJ, Prestwich GD. 2002. Hyaluro-nan-binding proteins: tying up the giant.J. Biol. Chem. 277:4585–88
119. Tammi MI, Day AJ, Turley EA. 2002.Hyaluronan and homeostasis: a balancingact. J. Biol. Chem. 277:4581–84
120. Wisniewski HG, Vilcek J. 2004. Cyto-kine-induced gene expression at the cross-roads of innate immunity, inflammationand fertility: TSG-6 and PTX3/TSG-14.Cytokine Growth Factor Rev. 15:129–46
121. Hirschfield GM, Pepys MB. 2003. C-reactive protein and cardiovascular dis-ease: new insights from an old molecule.Q. J. Med. 96:793–807
122. Fazzini F, Peri G, Doni A, Dell’AntonioG, Dal Cin E, et al. 2001. PTX3 in small-vessel vasculitides: an independent in-dicator of disease activity produced atsites of inflammation. Arthritis Rheum.44:2841–50
123. Luchetti MM, Piccinini G, Mantovani A,Peri G, Matteucci C, et al. 2000. Expres-sion and production of the long pentraxinPTX3 in rheumatoid arthritis (RA). Clin.Exp. Immunol. 119:196–202
124. Latini R, Maggioni AP, Peri G, GonziniL, Lucci D, et al. 2004. Prognostic sig-nificance of the long pentraxin PTX3 inacute myocardial infarction. Circulation110:2349–54
125. Napoleone E, Di Santo A, Peri G, Manto-vani A, de Gaetano G, et al. 2004. The long
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

14 Feb 2005 13:3 AR AR239-IY23-11.tex XMLPublishSM(2004/02/24) P1: JRX
366 GARLANDA ET AL.
pentraxin PTX3 up-regulates tissue fac-tor in activated monocytes: another linkbetween inflammation and clotting acti-vation. J. Leukoc. Biol. 76:203–9
126. Napoleone E, Di Santo A, Bastone A,Peri G, Mantovani A, et al. 2002. Longpentraxin PTX3 upregulates tissue fac-tor expression in human endothelial cells:a novel link between vascular inflamma-tion and clotting activation. Arterioscler.Thromb. Vasc. Biol. 22:782–87
127. Zhang XQJ, Moyse J, Jarari N, Confino E,Kazer R. 2003. Pentraxin-3 gene expres-sion in cumulus cells may be a molecu-lar marker for egg quality. Fertil. Steril.80(Suppl. 3):S81
128. DeGregorio-Rocasolano N, Gasull T,Trullas R. 2001. Overexpression of neu-ronal pentraxin 1 is involved in neuronal
death evoked by low K+ in cerebellargranule cells. J. Biol. Chem. 276:796–803
129. Hossain MA, Russell JC, O’Brien R, Lat-erra J. 2004. Neuronal pentraxin 1: a novelmediator of hypoxic-ischemic injury inneonatal brain. J. Neurosci. 24:4187–96
130. Stehlik C, Kroismayr R, Dorfleutner A,Binder BR, Lipp J. 2004. VIGR—a novelinducible adhesion family G-protein cou-pled receptor in endothelial cells. FEBSLett. 569:149–55
131. McMillan DR, Kayes-Wandover KM,Richardson JA, White PC. 2002. Verylarge G protein-coupled receptor-1, thelargest known cell surface protein, ishighly expressed in the developing centralnervous system. J. Biol. Chem. 277:785–92
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

P1: JRX
February 21, 2005 13:52 Annual Reviews AR239-FM
Annual Review of ImmunologyVolume 23, 2005
CONTENTS
FRONTISPIECE—Tadamitsu Kishimoto x
INTERLEUKIN-6: FROM BASIC SCIENCE TO MEDICINE–40 YEARS INIMMUNOLOGY, Tadamitsu Kishimoto 1
TNF/TNFR FAMILY MEMBERS IN COSTIMULATION OF T CELLRESPONSES, Tania H. Watts 23
DEVELOPMENT AND REGULATION OF CELL-MEDIATED IMMUNERESPONSES TO THE BLOOD STAGES OF MALARIA: IMPLICATIONSFOR VACCINE RESEARCH, Michael F. Good, Huji Xu, Michelle Wykes,and Christian R. Engwerda 69
THE T CELL RECEPTOR: CRITICAL ROLE OF THE MEMBRANEENVIRONMENT IN RECEPTOR ASSEMBLY AND FUNCTION,Matthew E. Call and Kai W. Wucherpfennig 101
CHEMOKINES, SPHINGOSINE-1-PHOSPHATE, AND CELL MIGRATION INSECONDARY LYMPHOID ORGANS, Jason G. Cyster 127
MARGINAL ZONE B CELLS, Shiv Pillai, Annaiah Cariappa,and Stewart T. Moran 161
HOW NEUTROPHILS KILL MICROBES, Anthony W. Segal 197
NK CELL RECOGNITION, Lewis L. Lanier 225
IPC: PROFESSIONAL TYPE 1 INTERFERON-PRODUCING CELLS ANDPLASMACYTOID DENDRITIC CELL PRECURSORS, Yong-Jun Liu 275
TYPE I INTERFERONS (α/β) IN IMMUNITY AND AUTOIMMUNITY,Argyrios N. Theofilopoulos, Roberto Baccala, Bruce Beutler,and Dwight H. Kono 307
PENTRAXINS AT THE CROSSROADS BETWEEN INNATE IMMUNITY,INFLAMMATION, MATRIX DEPOSITION, AND FEMALE FERTILITY,Cecilia Garlanda, Barbara Bottazzi, Antonio Bastone,and Alberto Mantovani 337
MAINTENANCE OF SERUM ANTIBODY LEVELS, Rudolf A. Manz,Anja E. Hauser, Falk Hiepe, and Andreas Radbruch 367
CATERPILLER: A NOVEL GENE FAMILY IMPORTANT IN IMMUNITY,CELL DEATH, AND DISEASES, Jenny P-Y. Tingand Beckley K. Davis 387
v
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

P1: JRX
February 21, 2005 13:52 Annual Reviews AR239-FM
vi CONTENTS
B CELL SIGNALING AND TUMORIGENESIS, Hassan Jumaa,Rudolf W. Hendriks, and Michael Reth 415
THE NOD MOUSE: A MODEL OF IMMUNE DYSREGULATION,Mark S. Anderson and Jeffrey A. Bluestone 447
ANTIGEN-SPECIFIC MEMORY B CELL DEVELOPMENT,Louise J. McHeyzer-Williams and Michael G. McHeyzer-Williams 487
THE B7 FAMILY REVISITED, Rebecca J. Greenwald, Gordon J. Freeman,and Arlene H. Sharpe 515
TEC FAMILY KINASES IN T LYMPHOCYTE DEVELOPMENT ANDFUNCTION, Leslie J. Berg, Lisa D. Finkelstein, Julie A. Lucas,and Pamela L. Schwartzberg 549
MOLECULAR GENETICS OF T CELL DEVELOPMENT, Ellen V. Rothenbergand Tom Taghon 601
UNDERSTANDING PRESENTATION OF VIRAL ANTIGENS TO CD8+T CELLS IN VIVO: THE KEY TO RATIONAL VACCINE DESIGN,Jonathan W. Yewdell and S.M. Mansour Haeryfar 651
IMMUNOLOGY OF MULTIPLE SCLEROSIS, Mireia Sospedraand Roland Martin 683
MAST CELLS AS “TUNABLE” EFFECTOR AND IMMUNOREGULATORYCELLS: RECENT ADVANCES, Stephen J. Galli, Janet Kalesnikoff,Michele A. Grimbaldeston, Adrian M. Piliponsky, Cara M.M. Williams,and Mindy Tsai 749
NETWORK COMMUNICATIONS: LYMPHOTOXINS, LIGHT, AND TNF,Carl F. Ware 787
ROLE OF C5A IN INFLAMMATORY RESPONSES, Ren-Feng Guoand Peter A. Ward 821
DNA DEGRADATION IN DEVELOPMENT AND PROGRAMMED CELLDEATH, Shigekazu Nagata 853
TOWARD AN UNDERSTANDING OF NKT CELL BIOLOGY: PROGRESSAND PARADOXES, Mitchell Kronenberg 877
MACROPHAGE RECEPTORS AND IMMUNE RECOGNITION,P.R. Taylor, L. Martinez-Pomares, M. Stacey, H-H. Lin, G.D. Brown,and S. Gordon 901
REGULATION OF LYMPHOID DEVELOPMENT, DIFFERENTIATION, ANDFUNCTION BY THE NOTCH PATHWAY, Ivan Maillard, Terry Fang,and Warren S. Pear 945
CELL BIOLOGY OF ANTIGEN PROCESSING IN VITRO AND IN VIVO,E. Sergio Trombetta and Ira Mellman 975
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.

P1: JRX
February 21, 2005 13:52 Annual Reviews AR239-FM
CONTENTS vii
INDEXESSubject Index 1029Cumulative Index of Contributing Authors, Volumes 13–23 1065Cumulative Index of Chapter Titles, Volumes 13–23 1072
ERRATAAn online log of corrections to Annual Review of Immunology chaptersmay be found at http://immunol.annualreviews.org/errata.shtml
Ann
u. R
ev. I
mm
unol
. 200
5.23
:337
-366
. Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
WIB
6244
- U
nive
rsity
of
Bon
n on
11/
30/0
9. F
or p
erso
nal u
se o
nly.
![Review Article Pentraxins: Structure, Function, and Role ...downloads.hindawi.com/journals/isrn/2013/379040.pdfISRN Inammation enzymaticattack[ ].SAPsharesmanystructuralandbiolog-icalcharacteristicswithCRP](https://img.pdfslide.us/doc/110x75/5cbc85a688c993f1698c6e67/review-article-pentraxins-structure-function-and-role-inammation-enzymaticattack.jpg)




























