Embed Size (px)
Citation preview

JOURNAL OF VIROLOGY, Oct. 2009, p. 10275–10279 Vol. 83, No. 190022-538X/09/$08.00�0 doi:10.1128/JVI.00949-09Copyright © 2009, American Society for Microbiology. All Rights Reserved.
Ganglioside GT1b Is a Putative Host Cell Receptor for the MerkelCell Polyomavirus�
Kimberly D. Erickson,1 Robert L. Garcea,1* and Billy Tsai2*Department of Molecular, Cellular, and Developmental Biology, University of Colorado at Boulder, 347 UCB, Boulder,
Colorado 80309,1 and Department of Cell and Developmental Biology, University of Michigan Medical School,109 Zina Pitcher Place, Room 3043, Ann Arbor, Michigan 481092
Received 13 May 2009/Accepted 9 July 2009
The Merkel cell polyomavirus (MCPyV) was identified recently in human Merkel cell carcinomas, anaggressive neuroendocrine skin cancer. Here, we identify a putative host cell receptor for MCPyV. We foundthat recombinant MCPyV VP1 pentameric capsomeres both hemagglutinated sheep red blood cells andinteracted with ganglioside GT1b in a sucrose gradient flotation assay. Structural differences between theanalyzed gangliosides suggest that MCPyV VP1 likely interacts with sialic acids on both branches of the GT1bcarbohydrate chain. Identification of a potential host cell receptor for MCPyV will aid in the elucidation of itsentry mechanism and pathophysiology.
Members of the polyomavirus (PyV) family, including sim-ian virus 40 (SV40), murine PyV (mPyV), and BK virus(BKV), bind cell surface gangliosides to initiate infection (2, 8,11, 15). PyV capsids are assembled from 72 pentamers (cap-someres) of the major coat protein VP1, with the internalproteins VP2 and VP3 buried within the capsids (7, 12). TheVP1 pentamer makes direct contact with the carbohydrateportion of the ganglioside (10, 12, 13) and dictates the speci-ficity of virus interaction with the cell. Gangliosides are glyco-lipids that contain a ceramide domain inserted into the plasmamembrane and a carbohydrate domain that directly binds thevirus. Specifically, SV40 binds to ganglioside GM1 (2, 10, 15),mPyV binds to gangliosides GD1a and GT1b (11, 15), andBKV binds to gangliosides GD1b and GT1b (8).
Recently, a new human PyV designated Merkel cell PyV(MCPyV) was identified in Merkel cell carcinomas, a rare butaggressive skin cancer of neuroendocrine origin (3). It is as yetunclear whether MCPyV is the causative agent of Merkel cellcarcinomas (17). A key to understanding the infectious andtransforming properties of MCPyV is the elucidation of itscellular entry pathway. In this study, we identify a putative hostcell receptor for MCPyV.
Because an intact infectious MCPyV has not yet been iso-lated, we generated recombinant MCPyV VP1 pentamers inorder to characterize cellular factors that bind to MCPyV. VP1capsomeres have been previously shown to be equivalent tovirus with respect to hemagglutination properties (4, 16), andthe atomic structure of VP1 bound to sialyllactose has dem-onstrated that the capsomere is sufficient for this interaction(12, 13). The MCPyV VP1 protein (strain w162) was expressed
and purified as described previously (1, 6). Briefly, a glutathi-one S-transferase-MCPyV VP1 fusion protein was expressed inEscherichia coli and purified using glutathione-Sepharose af-finity chromatography. The fusion protein was eluted usingglutathione and cleaved in solution with thrombin. The throm-bin-cleaved sample was then rechromatographed on a secondglutathione-Sepharose column to remove glutathione trans-ferase and any uncleaved protein. The unbound VP1 was thenchromatographed on a P-11 phosphocellulose column, andpeak fractions eluting between 400 and 450 mM NaCl werecollected. The purified protein was analyzed by sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE), fol-lowed by Coomassie blue staining (Fig. 1A, left) and immuno-blotting using an antibody (I58) that generally recognizes PyVVP1 proteins (Fig. 1A, right) (9). Transmission electron mi-croscopy (Philips CM10) analysis confirmed that the purifiedrecombinant MCPyV VP1 formed pentamers (capsomeres),which did not assemble further into virus-like particles (Fig.1B). In an initial screening of its cell binding properties, wetested whether the MCPyV VP1 pentamers hemagglutinatedred blood cells (RBCs). The MCPyV VP1 pentamers wereincubated with sheep RBCs and assayed as previously de-scribed (5). SV40 and mPyV recombinant VP1 pentamersserved as negative and positive controls, respectively. Wefound that MCPyV VP1 hemagglutinated the RBCs with thesame efficiency as mPyV VP1 (protein concentration/hemag-glutination unit) (Fig. 1C, compare rows B and C from wells 1to 11), suggesting that MCPyV VP1 engages a plasma mem-brane receptor on the RBCs. The recombinant murine VP1protein used for comparison was from the RA strain, a smallplaque virus (4). Thus, MCPyV VP1 has the hemagglutinationcharacteristics of a small plaque mPyV (12, 13).
To characterize the chemical nature of the putative receptorfor MCPyV, total membranes from RBCs were purified asdescribed previously (15). The plasma membranes (30 �g)were incubated with MCPyV VP1 (0.5 �g) and floated on adiscontinuous sucrose gradient (15). After fractionation, thesamples were analyzed by SDS-PAGE, followed by immuno-blotting with I58. VP1 was found in the bottom of the gradient
* Corresponding author. Mailing address for R. L. Garcea: Depart-ment of Molecular, Cellular, and Developmental Biology, Universityof Colorado at Boulder, 347 UCB, Boulder, CO 80309. Phone: (303)492-1669. Fax: (303) 492-1133. E-mail: [email protected] address for B. Tsai: Department of Cell and DevelopmentalBiology, University of Michigan Medical School, 109 Zina PitcherPlace, Room 3043, Ann Arbor, MI 48109. Phone: (734) 764-4167. Fax:(734) 764-5155. E-mail: [email protected].
� Published ahead of print on 15 July 2009.
10275
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 11
Feb
ruar
y 20
22 b
y 18
6.21
6.70
.159
.

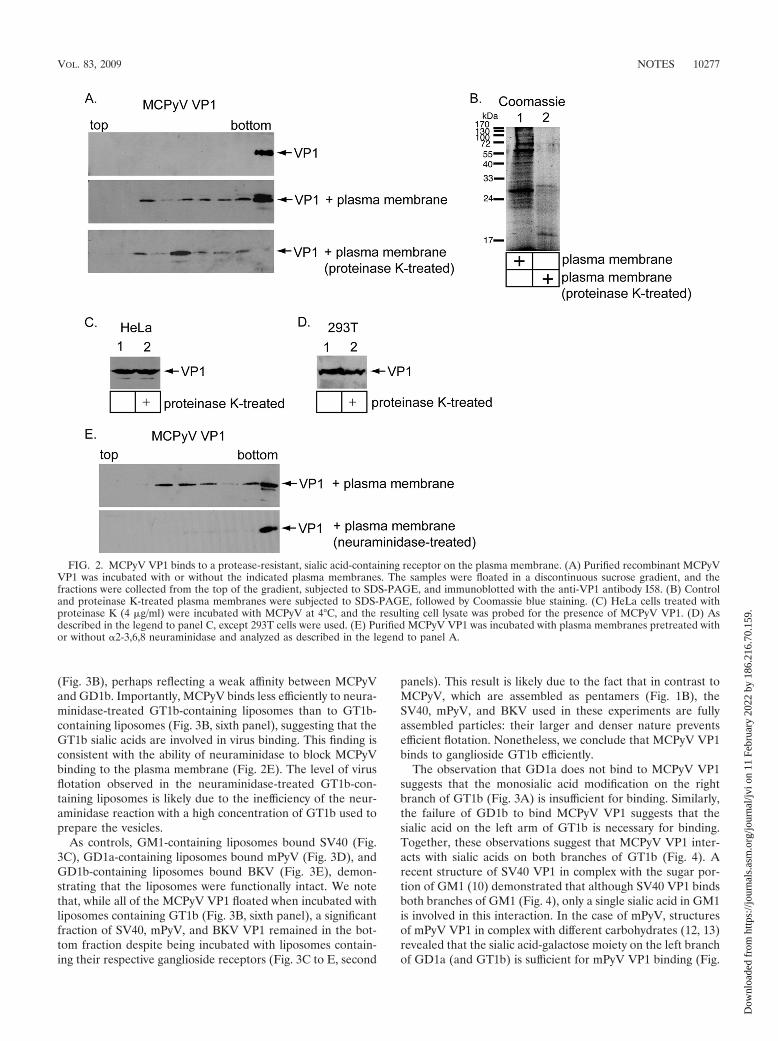
in the absence of the plasma membranes (Fig. 2A, first panel).In the presence of plasma membranes, a fraction of the VP1floated to the middle of the gradient (Fig. 2A, second panel),supporting the hemagglutination results that suggested thatMCPyV VP1 binds to a receptor on the plasma membrane.
To determine whether the receptor is a protein or a lipid,plasma membrane preparations (30 �g) were incubated withproteinase K (Sigma), followed by analysis with SDS-PAGEand Coomassie blue staining. Under these conditions, the ma-jority of the proteins in the plasma membranes were degradedby the protease (Fig. 2B, compare lanes 1 and 2). Despite thelack of proteins, the proteinase K-treated plasma membranesbound MCPyV VP1 as efficiently as control plasma mem-branes (Fig. 2A, compare the second and third panels), dem-onstrating that MCPyV VP1 interacts with a protease-resistantreceptor. The absence of VP1 in the bottom fraction in Fig. 2A(third panel) is consistent with the fact that the buoyant densityof the membranes is lowered by proteolysis. Of note, a similarresult was seen with binding of the mPyV to the plasma mem-brane (15). Binding of MCPyV to the cell surface of twohuman tissue culture cells (i.e., HeLa and 293T) was alsolargely unaffected by pretreatment of the cells with proteinaseK (Fig. 2C and D, compare lanes 1 and 2), further indicatingthat a nonproteinaceous molecule on the plasma membraneengages the virus.
We next determined whether the protease-resistant receptorcontains a sialic acid modification. Plasma membranes (10 �g)were incubated with a neuraminidase (�2-3,6,8 neuraminidase;Calbiochem) to remove the sialic acid groups. In contrast tothe control plasma membranes, the neuraminidase-treatedmembranes did not bind MCPyV VP1 (Fig. 2E, compare firstand second panels), indicating that the MCPyV receptor in-cludes a sialic acid modification.
Gangliosides are lipids that contain sialic acid modifications.We asked if MCPyV VP1 binds to gangliosides similar to otherPyV family members. The structures of the gangliosides usedin this analysis (gangliosides GM1, GD1a, GD1b, and GT1b)are depicted in Fig. 3A. To assess a possible ganglioside-VP1interaction, we employed a liposome flotation assay estab-lished previously (15). When liposomes (consisting of phos-phatidyl-choline [19 �l of 10 mg/ml], -ethanolamine [5 �l of 10mg/ml], -serine [1 �l of 10 mg/ml], and -inositol [3 �l of 10mg/ml]) were incubated with MCPyV VP1 and subjected to thesucrose flotation assay, the VP1 remained in the bottom frac-tion (Fig. 3B, first panel), indicating that VP1 does not interactwith these phospholipids. However, when liposomes contain-ing GT1b (1 �l of 1 mM), but not GM1 (1 �l of 1 mM) orGD1a (1 �l of 1 mM), were incubated with MCPyV VP1, thevesicles bound this VP1 (Fig. 3B). A low level of virus floatedpartially when incubated with liposomes containing GD1b
FIG. 1. Characterization of MCPyV VP1. Recombinant MCPyV VP1 forms pentamers and hemagglutinates sheep RBCs. (A) Coomassieblue-stained SDS-PAGE and an immunoblot of the purified recombinant MCPyV VP1 protein are shown. Molecular mass markers are indicated.(B) Electron micrograph of the purified MCPyV VP1. MCPyV VP1 (shown in panel A) was diluted to 100 �g/ml and absorbed onto Formvar/carbon-coated copper grids. Samples were washed with phosphate-buffered saline, stained with 1% uranyl acetate, and visualized by transmissionelectron microscopy at 80 kV. Bar � 20 nm. (C) Sheep RBCs (0.5%) were incubated with decreasing concentrations of purified recombinant SV40VP1 (row A), mPyV VP1 (row B), and MCPyV VP1 (row C). Wells 1 to 11 contain twofold serial dilutions of protein, starting at 2 �g/ml (well1). Well 12 contains buffer only and serves as a negative control. Well 7 (rows B and C) corresponds to 128 hemagglutination units per 2 �g/mlVP1 protein.
10276 NOTES J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 11
Feb
ruar
y 20
22 b
y 18
6.21
6.70
.159
.
(Fig. 3B), perhaps reflecting a weak affinity between MCPyVand GD1b. Importantly, MCPyV binds less efficiently to neura-minidase-treated GT1b-containing liposomes than to GT1b-containing liposomes (Fig. 3B, sixth panel), suggesting that theGT1b sialic acids are involved in virus binding. This finding isconsistent with the ability of neuraminidase to block MCPyVbinding to the plasma membrane (Fig. 2E). The level of virusflotation observed in the neuraminidase-treated GT1b-con-taining liposomes is likely due to the inefficiency of the neur-aminidase reaction with a high concentration of GT1b used toprepare the vesicles.
As controls, GM1-containing liposomes bound SV40 (Fig.3C), GD1a-containing liposomes bound mPyV (Fig. 3D), andGD1b-containing liposomes bound BKV (Fig. 3E), demon-strating that the liposomes were functionally intact. We notethat, while all of the MCPyV VP1 floated when incubated withliposomes containing GT1b (Fig. 3B, sixth panel), a significantfraction of SV40, mPyV, and BKV VP1 remained in the bot-tom fraction despite being incubated with liposomes contain-ing their respective ganglioside receptors (Fig. 3C to E, second
panels). This result is likely due to the fact that in contrast toMCPyV, which are assembled as pentamers (Fig. 1B), theSV40, mPyV, and BKV used in these experiments are fullyassembled particles: their larger and denser nature preventsefficient flotation. Nonetheless, we conclude that MCPyV VP1binds to ganglioside GT1b efficiently.
The observation that GD1a does not bind to MCPyV VP1suggests that the monosialic acid modification on the rightbranch of GT1b (Fig. 3A) is insufficient for binding. Similarly,the failure of GD1b to bind MCPyV VP1 suggests that thesialic acid on the left arm of GT1b is necessary for binding.Together, these observations suggest that MCPyV VP1 inter-acts with sialic acids on both branches of GT1b (Fig. 4). Arecent structure of SV40 VP1 in complex with the sugar por-tion of GM1 (10) demonstrated that although SV40 VP1 bindsboth branches of GM1 (Fig. 4), only a single sialic acid in GM1is involved in this interaction. In the case of mPyV, structuresof mPyV VP1 in complex with different carbohydrates (12, 13)revealed that the sialic acid-galactose moiety on the left branchof GD1a (and GT1b) is sufficient for mPyV VP1 binding (Fig.
FIG. 2. MCPyV VP1 binds to a protease-resistant, sialic acid-containing receptor on the plasma membrane. (A) Purified recombinant MCPyVVP1 was incubated with or without the indicated plasma membranes. The samples were floated in a discontinuous sucrose gradient, and thefractions were collected from the top of the gradient, subjected to SDS-PAGE, and immunoblotted with the anti-VP1 antibody I58. (B) Controland proteinase K-treated plasma membranes were subjected to SDS-PAGE, followed by Coomassie blue staining. (C) HeLa cells treated withproteinase K (4 �g/ml) were incubated with MCPyV at 4°C, and the resulting cell lysate was probed for the presence of MCPyV VP1. (D) Asdescribed in the legend to panel C, except 293T cells were used. (E) Purified MCPyV VP1 was incubated with plasma membranes pretreated withor without �2-3,6,8 neuraminidase and analyzed as described in the legend to panel A.
VOL. 83, 2009 NOTES 10277
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 11
Feb
ruar
y 20
22 b
y 18
6.21
6.70
.159
.

FIG. 3. MCPyV VP1 binds to ganglioside GT1b. (A) Structures of gangliosides GM1, GD1a, GD1b, and GT1b. The nature of the glycosidiclinkages is indicated. (B) Purified MCPyV VP1 protein was incubated with liposomes only or with liposomes containing the indicated gangliosides.The samples were analyzed as described in the legend to Fig. 2A. Where indicated, GT1b-containing liposomes were pretreated with �2-3,6,8neuraminidase and analyzed subsequently for virus binding. (C to E) The indicated viruses were incubated with liposomes and analyzed asdescribed in the legend to panel B.
10278 NOTES J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 11
Feb
ruar
y 20
22 b
y 18
6.21
6.70
.159
.

4). Although no structure of BKV in complex with the sugarportion of GD1b (or GT1b) is available, in vitro binding stud-ies (8) have suggested that the disialic acid modification on theright branch of GD1b (and GT1b) is responsible for bindingBKV VP1 (Fig. 4). Thus, it appears that the unique feature ofthe MCPyV VP1-GT1b interaction is that the sialic acids onboth branches of this ganglioside are likely involved in capsidbinding.
The identification of a potential cellular receptor forMCPyV will facilitate the study of its entry mechanism. Animportant issue for further study is to determine whetherMCPyV targets Merkel cells preferentially, and if so, whetherGT1b is found in higher levels in these cells to increase sus-ceptibility.
We acknowledge support from NIH grants AI064296 (to B.T.) andCA37667 (to R.L.G.).
REFERENCES
1. Bird, G., M. O’Donnell, J. Moroianu, and R. L. Garcea. 2008. Possible rolefor cellular karyopherins in regulating polyomavirus and papillomavirus cap-sid assembly. J. Virol. 82:9848–9857.
2. Campanero-Rhodes, M. A., A. Smith, W. Chai, S. Sonnino, L. Mauri, R. A.Childs, Y. Zhang, H. Ewers, A. Helenius, A. Imberty, and T. Feizi. 2007.N-glycolyl GM1 ganglioside as a receptor for simian virus 40. J. Virol.81:12846–12858.
3. Feng, H., M. Shuda, Y. Chang, and P. S. Moore. 2008. Clonal integration ofa polyomavirus in human Merkel cell carcinoma. Science 319:1096–1100.
4. Freund, R., R. L. Garcea, R. Sahli, and T. L. Benjamin. 1991. A single aminoacid substitution in polyomavirus VP1 correlates with plaque size and hem-agglutination behavior. J. Virol. 65:350–355.
5. Garcea, R. L., and T. L. Benjamin. 1983. Isolation and characterization ofpolyoma nucleoprotein complexes. Virology 130:65–75.
6. Kean, J. M., S. Rao, M. Wang, and R. L. Garcea. 2009. Seroepidemiology ofhuman polyomaviruses. PLoS Pathog. 5(3):e1000363.
7. Liddington, R. C., Y. Yan, J. Moulai, R. Sahli, T. L. Benjamin, and S. C.Harrison. 1991. Structure of simian virus 40 at 3.8-A resolution. Nature354:278–284.
8. Low, J. A., B. Magnuson, B. Tsai, and M. J. Imperiale. 2006. Identificationof gangliosides GD1b and GT1b as receptors for BK virus. J. Virol. 80:1361–1366.
9. Montross, L., S. Watkins, R. B. Moreland, H. Mamon, D. L. D. Caspar, andR. L. Garcea. 1991. Nuclear assembly of polyomavirus capsids in insect cellsexpressing the major capsid protein VP1. J. Virol. 65:4991–4998.
10. Neu, U., K. Woellner, G. Gauglitz, and T. Stehle. 2008. Structural basis ofGM1 ganglioside recognition by simian virus 40. Proc. Natl. Acad. Sci. USA105:5219–5224.
11. Smith, A. E., H. Lilie, and A. Helenius. 2003. Ganglioside-dependent cellattachment and endocytosis of murine polyomavirus-like particles. FEBSLett. 555:199–203.
12. Stehle, T., Y. Yan, T. L. Benjamin, and S. C. Harrison. 1994. Structure ofmurine polyomavirus complexed with an oligosaccharide receptor fragment.Nature 369:160–163.
13. Stehle, T., and S. C. Harrison. 1996. Crystal structures of murine polyoma-virus in complex with straight-chain and branched-chain sialyloligosaccha-ride receptor fragments. Structure 4:183–194.
14. Reference deleted.15. Tsai, B., J. M. Gilbert, T. Stehle, W. Lencer, T. L. Benjamin, and T. A.
Rapoport. 2003. Gangliosides are receptors for murine polyoma virus andSV40. EMBO J. 22:4346–4355.
16. Zullo, J. N., C. D. Stiles, and R. L. Garcea. 1987. Induction of c-myc and c-fosby polyomavirus: distinct roles for the capsid protein VP1 and the viral earlyproteins. Proc. Natl. Acad. Sci. USA. 84:1210–1214.
17. zur Hausen, H. 2008. Novel human polyomaviruses—re-emergence of a wellknown virus family as possible human carcinogens. Int. J. Cancer 123:247–250.
FIG. 4. A potential model of the different VP1-ganglioside interactions (see the text for discussion).
VOL. 83, 2009 NOTES 10279
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 11
Feb
ruar
y 20
22 b
y 18
6.21
6.70
.159
.





























