Embed Size (px)
Citation preview

NATURE MEDICINE • VOLUME 4 • NUMBER 10 • OCTOBER 1998 1115
NEWS & VIEWS
INVESTIGATIONS OF THE central nervous sys-tem (CNS) at the cellular and molecular
levels have generated some of the mostexciting breakthroughs in neuroscience.They have revealed the brain to be adynamic and constantly changing organthat is characterized by neural plasticity:the capacity of neural elements to altertheir electrical properties, grow newprocesses, and/or modify connectionsand circuits in order to adapt to an ever-changing world. Many of these studieshave focused on the synapse—thatmicro-molecular point of contactthrough which one neuron communi-cates with the next. Interest in synapticplasticity has grown, particularly withthe demonstration that the strength ofsynaptic connections can be altered bythe degree of activity at the synapse.This property is thought to provide thebasis for normal brain functions such aslearning and memory, as well asfor neurological disorders suchas epilepsy. Two papers, byBrooks-Kayal et al.1 on page1166 of this issue and Nusser et al.2 in Nature, suggest thathyperactivity at inhibitorysynapses—junctions that usethe neurotransmitter GABA (γ-aminobutyric acid)—leads toboth an increase in the numberof GABAA receptors in the post-synaptic membrane and analteration in their subunit com-position. These results shedlight on the involvement ofGABAergic transmission andassociated inhibitory synapticactivity in epileptic disorders.
In a widely studied example ofplasticity, called long-term po-tentiation (LTP), co-activation ofpre- and postsynaptic cells givesrise to a long-lasting increase inthe ability of the presynaptic cellto excite the postsynaptic neu-ron. Many laboratories are nowattempting to elucidate themechanisms underlying the LTPphenomenon3, and haveconcentrated particularly on ex-citatory synapses where gluta-mate is the neurotransmitter.Meanwhile, there has also been a
quiet revolution in our thinking about in-hibitory systems and how they contributeto many forms of synaptic plasticity.GABA is the major inhibitory neurotrans-mitter in the CNS, and much of the re-search into inhibitory activity hasinvolved the identification and descrip-tion of neurons that use GABA as theirmeans of communication. These cells—often called interneurons because theytend to communicate primarily withother cells in their local environment—are thought to modulate brain excitabil-ity and shape the output of the excitatorycells with which they interact.Investigation of the hippocampal forma-tion, a structure known for its involve-ment in learning and memory as well asits key role in temporal lobe epilepsy, re-
veals a rich complexity of GABAergic celltypes4, each with its own unique struc-ture, connectivity and (presumably) func-tion.
Perhaps not surprisingly, much of theinterest in GABAergic function hasarisen from research into the basicmechanisms underlying epileptic disor-ders5, for the epileptic state presentsapparently dramatic changes in the bal-ance between excitatory and inhibitoryactivity. The Brooks-Kayal and Nusserstudies are a continuation of efforts todecipher the basis of epilepsy. They bothuse a sophisticated combination of elec-trophysiological and molecular analysesthat provides new insights into the plas-ticity of GABA synapses. Much work hasshown that seizure activity results in achange in GABAA receptor subunit ex-pression6 and that the pharmacologicand kinetic properties of these receptors
depend on subunit composi-tion7. However, these twogroups provide for the first timea convincing molecular/mecha-nistic link between activity-de-pendent receptor changes andthe level of functional inhibi-tion in an intact CNS structure.
Their results bring interestingnew 'grist to the mill' withrespect to our understanding ofepileptogenesis (the develop-ment of a chronic seizure condi-tion) and the nature of theepileptic state. Although muchwork has centered on the rela-tive loss of inhibition in theepileptic brain, we have knownfor some time that inhibition isnot always decreased duringseizures. In some brain regions,and in some epileptic states, in-hibition seems to be increased.This has been observed not onlyin animal models but also inhuman (temporal lobe) epilep-sies. Both of the present studiesinvestigate the dentate regionof the hippocampal formationin which inhibition increasesafter treatments—electricalstimulation (kindling) or drugs(pilocarpine)—that lead to achronic seizure state. That there
GABA synapses enter the molecular big timeActivity-dependent changes in the number and composition of GABAA receptors provide the basis for inhibitory
synaptic plasticity and may be a key feature of the chronic epileptic state (pages 1166–1172).
PHILIP A. SCHWARTZKROIN
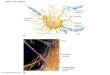
Example of GABAAreceptor subunit
compositionGABAA receptors
PresynapticPostsynaptic
Normalpre-seizurestate
Acutepost-seizure('latent')state
Chronicepilepticstate
α1
α2β1
β3 γ2
α4
α2δ
β3 γ2
α3
α2δ
β3 γ2
In the normal (pre-seizure) state (top), the GABAA receptorpopulation on hippocampal dentate granule cells is dominatedby α1, β1 and γ2 subunits. Shortly after an acute episode ofintense seizure activity, but before the animal exhibitsspontaneous chronic seizure behavior (that is, in the latentperiod, middle), the composition of the receptors shifts: α1 andβ1 are reduced relative to other α and β subunits, and α4 and β3
are increased along with δ and ε subunits (no significant changein γ2). In this period, GABA efficacy is not changed, suggestingthat the number of available receptors at the synapse has notbeen affected. In the chronically epileptic animal (bottom), thealtered subunit composition is maintained, and there is also anincreased number of GABAA receptors per synapse (both anincreased density of receptors and an increased area of thesynaptic junction)—a change that presumably gives rise toincreased GABA efficacy in these neurons1,2.
Bob
Crim
i

1116 NATURE MEDICINE • VOLUME 4 • NUMBER 10 • OCTOBER 1998
NEWS & VIEWS
is an increase in inhibition in the den-tate region at the same time that thebrain (as a whole) shows enhanced ex-citability may seem paradoxical. But thisobservation reminds us of the extremecomplexity of seizure states: clearly,seizures involve many cellular/molecu-lar variables and numerous interactionsbetween many areas of the brain.
What part do altered GABAA receptorsplay in epileptogenesis? Brooks-Kayaland co-workers attempt to address thisquestion by examining changes in themolecular make-up of GABAA receptorsat different times during the epilepto-genic process. They do this using a com-bination of patch-clamp and RNAamplification techniques, which allowsthem to study both electrical and molec-ular changes in a given neuron. The au-thors describe a change in the subunitcomposition of the GABAA receptors—adecrease in α1 and β1 subunits relative toother α and β subunits, respectively, anda concomitant increase in δ and ε sub-units (see figure)—weeks before the ani-mal demonstrates spontaneous seizureactivity. They speculate that thesechanges are involved in the evolution ofthe chronic seizure state. However, thestrength of response of single neurons toGABA (that is, GABA efficacy) increasesmuch later, after spontaneous seizureshave occurred. From the timing of thesechanges, one might view the later in-crease in GABA inhibitory power as acompensatory reaction to the high levelof excitation associated with theseizures, but not as a mechanism ofseizure development itself.
Although no comparable data areavailable from the Nusser study (whichexamined GABA receptor responsive-ness only after establishment of theepileptic state), it is tempting to corre-late the late increase in GABA efficacywith the increased number of GABA re-ceptors. Whether changes in GABAA re-ceptor subunit composition and/ornumber are early events in the epilep-togenic process, or whether they are,indeed, compensatory and triggered bythe repeated occurrence of sponta-neous seizures, remains a crucial ques-tion. Some recent efforts to developmore effective and specific antiepilep-tic drugs are directed towards targetingthe molecular subunit composition ofGABAA receptors. Thus, we need to
know whether to enhance or to coun-teract the influence of new receptorsfound in the epileptic brain.
The Brooks-Kayal and Nusser reports,in their focus on the postsynaptic GABAreceptor changes associated withseizures, also offer interesting commen-tary on the question of whether pre- orpostsynaptic changes are responsible forLTP-like synaptic plasticity—long a con-troversial issue in the excitatory (gluta-matergic) synapse literature. At leastunder the experimental conditions de-scribed in the new experiments, it seemsclear that postsynaptic changes in mole-cular structure contribute—and perhapsexplain entirely—seizure-related plastic-ity (although presynaptic contributionshave not been absolutely ruled out).
Whether such a conclusion can alsobe drawn for increased physiologic plas-ticity at GABAergic synapses in theabsence of the seizure state—indeed,whether LTP can be induced atGABAergic synapses—awaits further ex-perimentation. It is tempting to specu-late, however, that just as the nature ofthe postsynaptic glutamate receptors(the relative contribution of NMDA andnon-NMDA receptor subtypes) seems es-sential for a synapse’s potential for plas-ticity, so too might GABAergic synapticplasticity depend on the subunit compo-sition of the receptors at the synapse.The insertion of different GABAergic re-ceptor subunits into the postsynapticmembrane, and the increase in the num-ber of receptors at the postsynaptic site,are reminiscent of recent results, whichsuggest that glutamatergic receptornumber and subtype might be influ-enced by activity yielding changes insynapse strength8.
It is important to emphasize that thedemonstration of GABA receptor plastic-ity has implications for more than ourunderstanding and treatment ofepilepsy. Indeed, these two new reportssuggest that the properties of inhibitory(GABAergic) synapses can be altered byactivity—as has been shown for excita-tory (glutamatergic) synapses in studiesof LTP. Although the model systems de-scribed in both papers involve seizure (apathological extreme), the demonstra-tions of plasticity at GABAergic synapsessuggest that inhibitory points of contactmust be included in any analysis of achange in brain excitability.
How common this GABAergic plastic-ity might be, and under what physio-logical conditions it is induced, remainkey issues for investigation. The dra-matic GABA receptor plasticity inducedin association with seizures stronglysuggests that we should revisit previousattempts to demonstrate plasticity atthe level of inhibitory (GABAergic) in-terneurons9. Although these earlierstudies suggested that excitatory synap-tic contacts impinging on certaininterneurons undergo LTP-like potenti-ation—and thus indirectly promote in-hibition mediated by the GABAergiccells—the possibility that neuronal ac-tivity produces potentiation directly atGABAergic synapses has rarely been in-vestigated (but see ref. 10). GABAsynapses, and in particular the GABAA
receptors themselves, may providemore than gentle modulation of excita-tory neurotransmission—they, too,may be important points of synapticplasticity.
1. Brooks-Kayal, A.R., Shumate, M.D., Jin, H.,Rikhter, T.Y. & Coulter, D.A. Selective changes insingle cell GABAA receptor expression and functionin temporal lobe epilepsy. Nature Med. 4,1166–1172 (1998).
2. Nusser, Z., Hajos, N., Somogyi, P. & Mody, I.Increase in the number of synaptic GABAA recep-tors underlies change in synaptic efficacy at hip-pocampal inhibitory synapses. Nature 395,172–177 (1998).
3. Bliss, T.V.P. & Collingridge, G.L. A synaptic modelof memory: Long-term potentiation in the hip-pocampus. Nature 361, 31–39 (1993).
4. Freund, T.F. & Buzsaki, G. Interneurons of the hip-pocampus. Hippocampus 6, 347–470 (1996).
5. Fountain, N.B. & Lothman, E.W. Pathophysiologyof status epilepticus. J.Clin.Neurophysiol. 12,326–342 (1995).
6. Tsunashima, K., Schwarzer, C., Kirchmair, E.,Sieghart, W. & Sperk, G. GABAA receptor subunitsin the rat hippocampus. III. Altered messengerRNA expression in kainic-acid induced epilepsy.Neuroscience 80, 1001–1017 (1997).
7. Fisher, J.L. & Macdonald, R.L. Functional proper-ties of recombinant GABAA receptors composed ofsingle or multiple beta subunit subtypes.Neuropharmacology 36, 1601–1610 (1997).
8. Isaac, J.T., Nicoll, R.A. & Malenka, R.C. Evidencefor silent synapses: Implications for the expressionof LTP. Neuron 15, 427–434 (1995).
9. Ouardouz, M. & Lacaille, J.C. Mechanisms of se-lective long-term potentiation of excitatorysynapses in stratum oriens/alveus interneurons ofrat hippocampal slices. J. Neurophysiol. 73,810–819 (1995).
10. McLean, H.A., Caillard, O., Ben-Ari, Y. & Gaiarsa,J.L. Bidirectional plasticity expressed byGABAergic synapses in the neonatal rat hip-pocampus. J. Physiol. 496, 471–477 (1996).
Department of Neurological SurgeryUniversity of WashingtonBox 356470Seattle, Washington 98195-6470, USA