Embed Size (px)
Citation preview

F
F
Sl
Ca
b
a
ARRAHA
KSORSG
1
b1scscda(etaac
eFts
h0
ARTICLE IN PRESSG ModelISH-4562; No. of Pages 12
Fisheries Research xxx (2016) xxx–xxx
Contents lists available at ScienceDirect
Fisheries Research
journa l homepage: www.e lsev ier .com/ locate / f i shres
ull length article
patially intensive sampling by electrofishing for assessingongitudinal discontinuities in fish distribution in a headwater stream
éline Le Pichon a,∗, Évelyne Tales a, Jérôme Belliard a, Christian E. Torgersen b
Irstea, UR HBAN, 1 rue Pierre-Gilles de Gennes, CS10030, 92761 Antony Cedex, FranceU.S. Geological Survey, Forest and Rangeland Ecosystem Science Center, Cascadia Field Station, University of Washington, Seattle, WA, USA
r t i c l e i n f o
rticle history:eceived 17 December 2015eceived in revised form 26 August 2016ccepted 22 September 2016andled by Prof. George A. Rosevailable online xxx
a b s t r a c t
Spatially intensive sampling by electrofishing is proposed as a method for quantifying spatial variation infish assemblages at multiple scales along extensive stream sections in headwater catchments. We usedthis method to sample fish species at 10-m2 points spaced every 20 m throughout 5 km of a headwaterstream in France. The spatially intensive sampling design provided information at a spatial resolutionand extent that enabled exploration of spatial heterogeneity in fish assemblage structure and aquatichabitat at multiple scales with empirical variograms and wavelet analysis. These analyses were effective
eywords:tream fishbstaclesiverscapepatial discontinuity
for detecting scales of periodicity, trends, and discontinuities in the distribution of species in relation totributary junctions and obstacles to fish movement. This approach to sampling riverine fishes may beuseful in fisheries research and management for evaluating stream fish responses to natural and alteredhabitats and for identifying sites for potential restoration.
© 2016 Elsevier B.V. All rights reserved.
eostatistics. Introduction
Landscape perspectives in riverine ecology have increasinglyeen recognized in the last 30 years (Pringle et al., 1988; Schlosser991; Johnson and Host, 2010), leading to the concept of the “river-cape” (Malard et al., 2000; Ward et al., 2002; Wiens, 2002). Thisoncept considers the patchy nature of river systems and empha-izes the critical importance of considering the spatial and temporalontext of riverine organisms and their habitat. This contextualependency, expressed in the serial discontinuity concept (Wardnd Stanford, 1995) and the concept of the river discontinuumPoole, 2002), necessitates spatially explicit approaches. Fauscht al. (2002) highlighted the importance of (1) developing “a con-inuous view of a river. . .to understand how processes interactingmong scales set the context for stream fishes and their habitat”,nd (2) employing sampling approaches that enable the quantifi-ation of patterns in fish distribution at multiple scales.
The most commonly used method for sampling fish in rivers islectrofishing (Cowx and Lamarque, 1990; Dunham et al., 2009).
Please cite this article in press as: Le Pichon, C., et al., Spatially intensinuities in fish distribution in a headwater stream. Fish. Res. (2016), ht
or small- to medium-sized rivers, multiple-pass removal elec-rofishing performed on a relatively short river reach is thetandard method because it provides an accurate assessment of
∗ Corresponding author.E-mail address: [email protected] (C. Le Pichon).
ttp://dx.doi.org/10.1016/j.fishres.2016.09.026165-7836/© 2016 Elsevier B.V. All rights reserved.
fish abundance (Carle and Strub, 1978). Because of the preci-sion of local estimates that removal electrofishing provides, thissampling strategy is frequently recommended in large-scale mon-itoring programs, like those implemented within the EuropeanWater Framework Directive (CEN, 2003). However, this approachis typically conducted at relatively few discrete sites separated by>1 km of stream, leaving large portions of the river unsampled.Although regional fish patterns can be inferred using cumulativedata from discrete sampling locations (Smith and Jones, 2005)and predictive statistical modeling, there may be a knowledge gapbetween local scales (i.e., the scale of a sampled site: 100–500 m)and intermediate scales targeted by management plans (5–10 km),(Meador et al., 2003).
Spatially intensive approaches to sampling are needed to assessvariability in fish distribution in response to instream habitat struc-ture and spatial stream discontinuities of natural or anthropogenicorigin. Close spacing of samples longitudinally along the streamprovides the resolution necessary to describe spatio-temporal het-erogeneity in the physical stream environment (White et al., 2014)and to localize discontinuities in stream fish distribution causedby chemical, physical and thermal factors (Bateman et al., 2005;Baxter, 2002). Such approaches have been used to assess discon-tinuous fish distributions (Angermeier et al., 2002; Angermeier
ve sampling by electrofishing for assessing longitudinal disconti-tp://dx.doi.org/10.1016/j.fishres.2016.09.026
and Smogor, 1995) at tributary junctions (Kiffney et al., 2006)and below dams (Brenkman et al., 2012). Various methods to spa-tially intensive sampling have been employed to quantify spatial

ARTICLE IN PRESSG ModelFISH-4562; No. of Pages 12
2 C. Le Pichon et al. / Fisheries Research xxx (2016) xxx–xxx
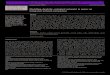
F hmen( il of ps oundi
hpvoaM2skeapassssa2u‘hcrcaissd
ig. 1. (a) Study area in the Seine River Basin, France. (b) Orgeval experimental catcc) Spatially intensive sampling by electrofishing (SIS) in the Rognon stream. A detapatial gap (400 m) in sampling (brackets) due to inaccessible private property surr
eterogeneity in rivers and stream fish distribution. For exam-le, low-level helicopter flights were used to survey salmon nestsisually (Isaak et al., 2007) and shore-based visual estimation meth-ds were tested to count age-0 Bonneville cutthroat trout (Whitend Rahel, 2008). Extensive snorkel surveys (Kiffney et al., 2006;ullner et al., 1998) and single-pass electrofishing (Bateman et al.,
005) have been used to provide the opportunity to evaluatepecies distribution at both small and large scales, from metres toilometres (Fausch et al., 2002; Gresswell et al., 2006; Lawrencet al., 2012; Torgersen et al., 2006). Snorkel surveys provide reli-ble estimates of fish abundance and community composition,articularly in remote locations and in rivers that are too largend deep for electrofishing (Chamberland et al., 2014). However,norkeling is generally not effective for detecting cryptic benthicpecies (Macnaughton et al., 2014; Plichard et al., 2016) and forampling streams that are shallow or turbid. The effectiveness ofingle-pass backpack electrofishing for sampling fishes has beenssessed in many studies (Bateman et al., 2005; Bertrand et al.,006; Meador et al., 2003; Reid et al., 2008). Brenkman et al. (2012)sed this method to highlight bull trout (Salvelinus confluentus)
hotspots’ (i.e., where the species was locally abundant) and fish-abitat relationships prior to dam removal. The expense of spatiallyontinuous single-pass electrofishing may preclude sampling longiver segments. To overcome this limitation, fractional samplingan be conducted randomly or systematically without altering theccuracy of fish abundance estimates (Mitro and Zale, 2000). For
Please cite this article in press as: Le Pichon, C., et al., Spatially intensinuities in fish distribution in a headwater stream. Fish. Res. (2016), ht
nstance, Torgersen and Close (2004) used a nested electrofishingampling design at fine (metres) and coarse (kilometres) spatialcales to quantify spatial patterns in larval Pacific lamprey abun-ance along a 55-km river section.
t (shaded in grey) and sampling reach on the Rognon stream (shaded in dark grey).ond A is provided, with locations of obstacles and connected inlet and outlet. Noteng pond B.
Point abundance sampling by electrofishing (PASE) is a frac-tional sampling design that is employed to collect fish at manysmall samples as opposed to relatively few large samples (Nelvaet al., 1979; Persat and Copp, 1990). The PASE approach is simi-lar to other fractional sampling methods, such as quadrat sampling(Williams et al., 2002), the abundance index (Prévost and Nihouarn,1999), and habitat unit sampling (Lamouroux et al., 1999). Rela-tive to multiple-pass removal by electrofishing, the PASE methodis more cost-effective and has less potential for causing injury andmortality to fish (Copp, 2010). Commonly used to sample 0+ fish(Copp and Garner, 1995; Scholten, 2003; Tales and Berrebi, 2007),its effectiveness has been shown for adult fish at the population orassemblage level (Laffaille et al., 2005; Pretty et al., 2003; Teixeira-de Mello et al., 2014; Tomanova et al., 2013).
The goal of this study was to propose and test a new samplingmethod in accordance with the riverscape approach (sensu Fauschet al., 2002) for assessing the distribution and discontinuities pat-terns in stream fish assemblages along a stream section (5 km) ina human-influenced headwater catchment. Specifically, we usedsystematic, spatially intensive sampling (SIS) based on PASE andspatial statistical analysis to quantify longitudinal patterns andspatial variability in stream fish abundance, species richness, andaquatic habitat at multiple scales.
2. Methods
ve sampling by electrofishing for assessing longitudinal disconti-tp://dx.doi.org/10.1016/j.fishres.2016.09.026
2.1. Study area
The study area was located in the headwaters of the Orgevalcatchment (104 km2) in the Seine River basin, 70 km east of Paris,

ING ModelF
es Res
Fl1sTactTaIrap
sPaauowoad
2
aspsw(cDhcdptwai
mgmdfi[awwaA
2
fiycft
ARTICLEISH-4562; No. of Pages 12
C. Le Pichon et al. / Fisheri
rance (Fig. 1a). The Orgeval is a rural experimental catchment andong-term environmental research observatory monitored since962 (Loumagne and Tallec, 2013). The Orgeval is a third-ordertream with an interannual average flow of 0.6 m3 s−1 (Fig. 1b).he climate in the study catchment is oceanic temperate, withn average annual rainfall of 700 mm; the average altitude of theatchment is 148 m and varies from 80 m to 186 m. Land use inhe catchment is agriculture (81%), forest (18%), and urban (1%).his rural catchment presents numerous straightened stretchesnd agricultural ditches in headwaters with imbedded substrates.n other segments, the habitats are dominated by glides, partlyelated to the presence of obstacles. Some of these obstacles are
legacy of historical uses of the catchment with run-of-the riveronds created for fish farming since the Middle Ages and flour mills.
In this catchment, fish assemblages and habitats were inten-ively sampled on a 5-km segment of the Rognon stream (Fig. 1c).otential obstacles to fish movement on this stream (i.e., natural orrtificial falls and chutes, mill weirs, spill-ways and culverts) werelso located, measured according to Ovidio et al. (2007), and contin-ously mapped using a Garmin GPSMAP 62 (±5 m) (see obstaclesn the Rognon stream depicted in Fig. 1c). Some of these obstaclesere associated with ponds connected to the stream by inlet and
utlet ditches; pond A is a fishing pond created in 1990 for recre-tional use and pond B is associated with an historic property thatates back to the seventeenth century (Fig. 1c).
.2. Spatially intensive sampling (SIS)
Based on point abundance sampling by electrofishing (PASE)pproaches (Nelva et al., 1979; Copp, 2010; Tomanova et al., 2013),patially intensive sampling (SIS) was implemented using a back-ack electroshocker (Martin Pecheur®, Dream Electronics). At eachample point, the anode was immersed for 10 s in a single pass,hich has been shown to be effective for electrofishing small fishes
Scholten, 2003). The sampled area for each sampled point was theircular electrical field around the anode estimated to be 10 m2.uring electrofishing, fish were captured with a small dip net andeld in a transportable container. Fish from each sample wereollected for identification and measurement and then releasedownstream so that they would not be caught in subsequent sam-les. SIS was conducted longitudinally in an upstream directionhroughout a 5-km segment of the Rognon stream (Fig. 1c). Samplesere collected systematically to facilitate spatial pattern analysis
nd were separated by 20 m to reduce the likelihood of fish movingnto subsequent samples.
For each sample, its lateral position in the channel (right, left andiddle) was randomly chosen using dice. Each sample location was
eoreferenced using a Garmin GPSMAP 62 (±5 m). Channel geo-orphic units (e.g., riffle, pool, glide) (Malavoi and Souchon, 2002),
epth, four classes of current velocity (zero, low, medium, high),ve dominant substrate types (silt [<0.6 mm], sand [<2 mm], gravel
2–64 mm], cobble [64–256 mm], boulders and block [>256 mm]),nd presence/absence of cover (large woody debris, under banksith roots, boulders) were recorded at each sample location. Theetted width in the sampled area ranged from 2 to 9 m and aver-
ged 3.5 ± 1.7. SIS was conducted at 264 sample points on 19–21ugust 2008 by a five-person team during stable flow conditions.
.3. Data analysis
To explore spatial patterns in the longitudinal distribution ofshes and habitats, graphical, multivariate and geostatistical anal-
Please cite this article in press as: Le Pichon, C., et al., Spatially intensinuities in fish distribution in a headwater stream. Fish. Res. (2016), ht
ses were applied and evaluated for analyzing high-resolutionommunity data. These analyses have been shown to be effectiveor identifying effects of tributaries on fish distribution in headwa-er streams (Torgersen et al., 2008).
PRESSearch xxx (2016) xxx–xxx 3
Samples and position of obstacles were located longitudinallyaccording to in-stream distance upstream from initiation of SIS.In-stream distances between samples were calculated with ArcGIS9.3 and Anaqualand 2.0, a freeware software package for calculat-ing directional hydrographic distances between points or patchesalong a river (Le Pichon et al., 2006). Fish and habitat data wereplotted by summarizing data in 100-m bins from downstreamto upstream. Species richness (distinguishing ecological guild ofrheophilic, and non-rheophilic species (Schiemer and Waidbacher,1992)) and abundance for the most common species were summedlongitudinally in 100-m bins. Habitat was described by the per-centage of each channel geomorphic units in 100-m bins. Thecumulative height of obstacles was determined by adding sequen-tial heights for each obstacle starting with the height of the obstaclefarthest downstream.
2.3.1. Multivariate analysisLongitudinal variations of fish assemblage and habitat on sam-
ples were described with multivariate analyses. Correspondenceanalysis (CA) was performed on the matrix of species abundanceby samples (10 species × 264 samples), after discarding rare species(relative occurrence below 1%) (Greenacre, 1984). The matrix offish abundances was log-transformed (log(x + 1)) before analy-sis. The samples scores on the first axis of the CA were thenplotted longitudinally and locally weighted scatterplot smooth-ing (Loess function, span = 0.5, 2nd degree polynomial curve) wasused to better visualize longitudinal pattern. This technique is anonparametric regression technique for identifying trends in het-erogeneous data (Trexler and Travis, 1993).
Likewise, an Hill and Smith analysis (HSA) (Hill and Smith,1976) was performed on the matrix describing habitat (depth,current velocity, bottom substrate, and shelters) by samples (4 vari-ables × 264 samples). HSA is close to a multiple correspondenceanalysis (MCA) and allows the use of both quantitative and qualita-tive variables. The samples scores on the first axis of the HSA wereplotted longitudinally and locally weighted scatterplot smoothing(Loess function, span = 0.5, 2nd degree polynomial curve) was usedto show the trends in the scores series and to facilitate the inter-pretation.
2.3.2. Geostatistical analysisGraphical analysis and multivariate analysis may not be suffi-
cient for detecting subtle trends. Empirical semivariograms werecomputed based on Perry et al. (2002) and Torgersen et al. (2004)to quantify patterns of spatial variability and scales of variation infish abundance and aquatic habitat. The semivariograms provideda graphical representation of the distances over which fish abun-dance and aquatic habitat were autocorrelated; this distance canbe determined by examining the inflection point in semivariance(Palmer, 2002). In geostatistics, this inflection point is referred to asthe “range” and is an indicator of the dominant spatial scale of vari-ation. A script in S-PLUS (Ganio et al., 2005) was used to computethe robust variogram estimator (Cressie, 1993) and to plot semi-variance versus separation distance (lag) between samples using(i) abundance data for the three most abundant fish species and(ii) the samples scores on axis 1 from CA and HSA. A maximumseparation distance of 4000 m was used, which enabled us to havebetween 92 and 658 pairs of points for each calculation of semivari-ance. This distance was longer than the recommended half of themaximum separation distance between points; however, calcula-tions of semivariance were based on a sufficient number of pairs(>50) for exploratory analysis (Rossi et al., 1992). The minimum
ve sampling by electrofishing for assessing longitudinal disconti-tp://dx.doi.org/10.1016/j.fishres.2016.09.026
separation distance and the lag class interval were 40 m, respec-tively, corresponding to twice the distance between samples. A verysmall value (10−5) was assigned to samples that had counts of zerofor a given species. Randomizations were used to generate 10th and

ARTICLE IN PRESSG ModelFISH-4562; No. of Pages 12
4 C. Le Pichon et al. / Fisheries Research xxx (2016) xxx–xxx
Table 1Species composition, abundance, and occurrence of fish collected with spatially intensive sampling by electrofishing (SIS) in the Rognon Stream, France. Fish were collectedat 264 point samples spaced every 20 m over 5 km of stream. Species are sorted in the table in ascending order of occurrence in the samples. Benthic species are in bold.
Species Abbreviation Type Abundance Relativeabundance RA(%)
Occurrence Relativeoccurrence RO(%)
Bullhead (Cottus gobio) BUL rheophilic 439 23.2 168 63.6Eurasian minnow (Phoxinus phoxinus) MIN rheophilic 1253 66.1 143 54.2Stone loach (Barbatula barbatula) LOA rheophilic 84 4.4 48 18.2Brown trout (Salmo trutta) BRT rheophilic 15 0.8 13 4.9European eel (Anguilla anguilla) EEL non-rheophilic 11 0.6 11 4.2Gudgeon (Gobio gobio) GUD rheophilic 14 0.7 9 3.4White bream (Blicca bjoerkna) WBR non-rheophilic 45 2.4 7 2.7Roach (Rutilus rutilus) ROA non-rheophilic 15 0.8 6 2.3European perch (Perca fluviatilis) PER non-rheophilic 6 0.3 6 2.3Chub (Squalius cephalus) CHU rheophilic 6 0.3 4 1.5Common bream (Abramis brama) CBR non-rheophilic 3 0.2 2 0.8Dace (Leuciscus leuciscus) DAC rheophilic 2 0.1 2 0.8
ic
ic
ic
9tewdsow
lWdwondwstsAwpap)opdcwti(
3
3
ttEstw
Northern pike (Esox lucius) PIK non-rheophilRudd (Scardinius erythrophthalmus) RUD non-rheophilTench (Tinca tinca) TEN non-rheophil
0th percentiles based on 5000 permutations for comparisons withhe observed empirical variogram (Ganio et al., 2005). Variogramstimates outside the percentiles indicated separation distances athich the spatial dependence was significantly different from ran-
om. To evaluate the influence of data spacing on the detection ofpatial structure in fish abundance, variograms based on every sec-nd sample (40-m spacing) and every fourth sample (80-m spacing)ere computed with a 100-m lag class interval.
Wavelet analysis was used to assess spatial heterogeneity andocal discontinuities in fish distribution (Cho and Chon, 2006).
avelet analysis provides insights into periodicity in fish abun-ance at nested spatial scales (Torgersen et al., 2004). This toolas used in an exploratory manner to examine spatial patterns
f three species, including benthic species, which typically haveot been sampled at a spatial resolution that is sufficient for con-ucting spatial analysis. The continuous wavelet transform (CWT)as applied to determine the dominant scale of variability in fish
pecies richness and abundance. The CWT can be used with datahat are systematically sampled along a one-dimensional transect,uch as data from spatially intensive longitudinal electrofishing.brupt transitions in one-dimensional data affect the continuousavelet coefficients at all scales and are statistically distinct from
atterns at small scales. Wavelet coefficients may be used to deriven estimate of the power spectrum. The Interactive Wavelet Plotrovided by Torrence and Compo (1998) (http://ion.exelisvis.com/
was used to compute the wavelet power spectrum with a second-rder Gaussian wavelet model, which is appropriate for periodiceaks. The Interactive Wavelet Plot includes a statistical test foretecting peaks in the wavelet power spectrum that are signifi-antly above those of the mean power spectrum (global wavelet)ith a 95% confidence level. The advantage of wavelet analysis is
hat it provides a method for visualizing multiple scales of variationn the data in relation to their longitudinal position in the streamTorgersen et al., 2004).
. Results
.1. Fish abundance, species richness and assemblage structure
A total of 1896 fish representing 15 species were captured inhe 240 samples that contained at least one fish in a 5-km sec-ion of the Rognon stream; 24 samples contained no fish (Table 1).
Please cite this article in press as: Le Pichon, C., et al., Spatially intensinuities in fish distribution in a headwater stream. Fish. Res. (2016), ht
urasian minnow (Phoxinus phoxinus) was the most abundantpecies based on relative abundance, followed by bullhead (Cot-us gobio) and stone loach (Barbatula barbatula), whereas bullhead
as the most commonly encountered species among samples,
1 0.1 1 0.41 0.1 1 0.41 0.1 1 0.4
followed by Eurasian minnow and stone loach (Table 1). Amonglow-occurrence species (relative occurrence <5%), brown trout(Salmo trutta), gudgeon (Gobio gobio) and chub (Squalius cephalus)were found, while non-rheophilic species included European eel(Anguilla Anguilla), white bream (Blicca bjoerkna), roach (Rutilusrutilus) and European perch (Perca fluviatilis). Rare species (rela-tive occurrence <1%) were mainly non-rheophilic species, Northernpike (Esox lucius), common bream (Abramis brama), rudd (Scardiniuserythrophthalmus) and tench (Tinca tinca); except common dace(Leuciscus leuciscus) (Table 1).
Longitudinal patterns in species richness of rheophilic andnon-rheophilic fishes were spatially heterogeneous and showedpronounced discontinuities ostensibly associated with connectedponds, in-stream obstacles, and, potentially, tributary junctions(Fig. 2). Species richness was low in the stream segment with themost dramatic changes in the cumulative height of obstacles, due toobstacles associated with connected ponds A and B (3200 m, 3700,3900 m, Fig. 2). Rheophilic species were the only species capturedin the lower 1100 m of the stream (Fig. 2). Peaks in non-rheophilicspecies richness at 2400 m and 3600 m were observed downstreamof outlets of connected ponds A and B, whereas rheophilic specieswere not observed in the area affected by falls (3900–4000 m)(Figure 2).
Fish assemblage analysis with multivariate methods revealeddistinct differences in assemblage structure. The first factorialaxis of the CA indicated a gradient of species occurrence, withlow-occurrence species (relative occurrence <4.9%) having posi-tive scores and prevalent species having negatives scores (Fig. 3a).This axis emphasized the opposition between a typical rheophilicassemblage of brown trout and bullhead, stone loach and Eurasianminnow, versus low-occurrence of non-rheophilic species that hadlikely colonized streams from connected ponds. Plotting samplescores from axis 1 on the longitudinal stream profile showed thespatial distribution of both assemblages (Fig. 3b).
The longitudinal patterns of the three most abundant speciesexhibited distinct peaks and troughs (Fig. 4). Bullhead occurrencewas high in the upstream segment of the survey (4000–4500 m),but this species was absent in the segment influenced by connectedponds (3600–4000 m) (Fig. 4a and d). Stone loach counts were high-est from 0 to 1400 m and were lower and more patchy upstreamof the confluence with the Bourgogne stream (1400 m) (Fig. 4b,e).Eurasian minnow were distributed throughout the entire segment,
ve sampling by electrofishing for assessing longitudinal disconti-tp://dx.doi.org/10.1016/j.fishres.2016.09.026
with several peaks of high abundance at 300, 3700, and 4600 m;this species was not observed at 3900–4000 m, where there wereconnected ponds (Fig. 4c,f). Counts of bullhead, stone loach, andEurasian minnow were highly variable at the scale of sample points,

ARTICLE IN PRESSG ModelFISH-4562; No. of Pages 12
C. Le Pichon et al. / Fisheries Research xxx (2016) xxx–xxx 5
Fig. 2. Longitudinal patterns of species richness from spatially intensive sampling by electrofishing (SIS) in the Rognon stream. Species richness is summarised in 100-mbins, with bars representing the total number of species and the composition of rheophilic, and non-rheophilic species. Locations of tributary junctions and connected pondsare indicated by vertical solid and dashed arrows, respectively. Locations of obstacles are indicated by solid black circles on the horizontal axis. The horizontal black lineunderneath an asterisk [*] indicates a spatial gap in sampling. Rheophilic species and non-rheophilic species are specified in Table 1.
F 0 specb in Tas ine.
slbpi
3
sat
ig. 3. Correspondence analysis (CA) of the log(x + 1) transformed abundances of 1lack triangles indicate sample coordinates, and species abbreviations are specifiedmoothing (Loess function, span = 0.5, degree = 2) of these data is shown as a black l
o binning these data at the 100-m scale facilitated the analysis ofongitudinal patterns. In particular, spatial patterns in the distri-ution of Eurasian minnow, a shoaling species, were particularlyronounced at the 100-m scale (Fig. 4f) compared to the scale of
ndividual sample points (Fig. 4c).
.2. Longitudinal patterns of aquatic habitat
Please cite this article in press as: Le Pichon, C., et al., Spatially intensinuities in fish distribution in a headwater stream. Fish. Res. (2016), ht
The longitudinal distribution of pool, riffle, and glide habitathowed a high degree of fine-scale spatial heterogeneity amongdjacent 100-m bins (Fig. 5). Longitudinal trends in aquatic habi-at in the 5-km section of stream were most apparent for pools,
ies in 264 samples. (a) Biplot of species and samples scores for the first two axes;ble 1. (b) Longitudinal variation in scores from Axis 1. Locally weighted scatterplot
which increased in an upstream direction (Spearman correlationtest p = 0.013) (Fig. 5). No significant trends were observed for rif-fles, although a large gap was noted (3300–4100 m), correspondingto high percentage of pools. The cumulative height of the 11 obsta-cles that were identified in the 5-km segment of stream (i.e., chutes,falls, logjams, and culverts) was 11 m (Fig. 5). Eighty percent of thecumulative height was the result of the six uppermost obstacles;the highest one (3.2 m) was located at 3750 m.
ve sampling by electrofishing for assessing longitudinal disconti-tp://dx.doi.org/10.1016/j.fishres.2016.09.026
The HSA on the habitat matrix has completed the analysis ofthe longitudinal distribution of channel geomorphic units. Cur-rent velocity and substrate had the main contribution on the firstaxis of HSA (Table 2). Samples with deep, slow and silted habitat

Please cite this article in press as: Le Pichon, C., et al., Spatially intensive sampling by electrofishing for assessing longitudinal disconti-nuities in fish distribution in a headwater stream. Fish. Res. (2016), http://dx.doi.org/10.1016/j.fishres.2016.09.026
ARTICLE IN PRESSG ModelFISH-4562; No. of Pages 12
6 C. Le Pichon et al. / Fisheries Research xxx (2016) xxx–xxx
Fig. 4. Longitudinal patterns of fish abundance at two spatial scales (left and right panels) in the Rognon stream for (a, d) bullhead, (b, e) stone loach, and (c, f) Eurasianminnow (black dots along the x-axis [a–c] indicate zero values, and a horizontal black line underneath an asterisk [*] indicates a spatial gap in sampling [d–f]). Panels on theleft (a–c) show fish counts at the spatial scale of a point sample (10 m2), and panels on the right (d–f) show fish counts at the 100-m scale.
Fig. 5. Longitudinal patterns of pool, riffle, and glide habitats in the Rognon stream. Percentages by length of pool, riffle, and glide habitat are summarised in 100-m bins.The horizontal black line underneath an asterisk (*) indicates a spatial gap in sampling. Cumulative height of in-stream obstacles is shown as a dark grey line.

ARTICLE IN PRESSG ModelFISH-4562; No. of Pages 12
C. Le Pichon et al. / Fisheries Research xxx (2016) xxx–xxx 7
Fig. 6. (a) Spatial variation in Axis 1 scores from multiple correspondence analysis (MCA) of depth, current velocity, bottom substrate, and presence of cover. Locally weightedscatterplot smoothing (Loess function, span = 0.5, degree = 2) is shown as a black line. Sesample scores from Axis 1. Minimum separation distance and the lag class interval were 4as black lines.
Table 2Variable contributions on axis 1 of the Multiple Correspondence Analysis.
Variable Modality Score on Axe 1
Depth – 0.858765350
Current velocity Nul 1.055020306Low −0.124326113Medium −1.016932288Strong −1.138305682
Substrate Silt 1.504472859Sand 0.215030578Gravel −0.005760549Pebble −0.626551069Boulder 0.915103195
whtacprpt((
3
Edh
Shelter (woody debris) Absent −0.364149443Present 0.353279311
ith cover had positive scores, whereas shallow, fast and pebbleabitat with no cover had negative scores (Fig. 6a). The longi-udinal distribution of scores and the loess curve show irregularnd contrasting segments; the downstream reach (1–1500 m) wasomposed of shallow and fast habitats, followed by an increasingroportion of deep and slow habitats (1500–3000 m); the upstreameach showed a pronounced peak in positive scores due to a highroportion of pools. The semivariogram of axis 1 scores fromhe HSA ordination of aquatic habitat variables showed fine-scale100–200 m) spatial structure nested within a broad-scale trend>3000 m) (Fig. 6b).
.3. Spatial discontinuities in fish distribution and aquatic habitat
Please cite this article in press as: Le Pichon, C., et al., Spatially intensinuities in fish distribution in a headwater stream. Fish. Res. (2016), ht
Semivariograms of fish counts for bullhead, stone loach, andurasian minnow varied with respect to their spatial structure andominant scales of variation (Fig. 5). The semivariogram for bull-ead revealed a complex, nested spatial structure at both small
e Table 2 for contributions of variables on Axis 1. (b) Empirical semivariogram of0 m. The 10th and 90th percentiles are based on 5000 permutations and are shown
and large scales (Fig. 5a). The low semivariance (�) at short lags(inflection point at 500 m) showed fine-scale patchiness, and theincreasing semivariance at long distances (2100 m) indicated abroad-scale spatial trend. Fig. 5a also showed potential period-icity (i.e., peaks and troughs in �) at fine and broad scales. Thesemivariogram for stone loach indicated significant spatial struc-ture at a large spatial scale (>2500 m) (Fig. 5b). In contrast, thesemivariogram for Eurasian minnow (Fig. 5c) showed no changein semivariance as a function of distance, indicating a random, orunstructured, spatial pattern.
When the lag class interval was increased to 100 m for bull-head, the shape of the semivariogram was smoother, but the overallpattern was the same (Figs. 7a and 8a ). When the calculationof the semivariogram used only half of the total number of sam-ples, the shape of the semivariogram was similar, but more of thepoints fell within the 10th and 90th percentiles (Fig. 8b). Whenonly 25% of the samples were used, the shape of the semivari-ogram showed a random, unstructured spatial pattern at distancesless than 3100 m (Fig. 8c). The width between the 10th and 90thpercentiles increased at distances >2500 m because the number ofpairs for � calculations at these distances was relatively low (<30).
Wavelet analyses of spatial patterns in the distribution ofbullhead, stone loach, and Eurasian minnow illustrated the loca-tions along the stream at which spatial structure was particularlypronounced (Fig. 9). For example, bullhead abundances exhib-ited spatial structure at nested scales: fine (<500 m), intermediate(500–3000 m), and large (>3000 m) (Fig. 9a). For stone loach, therewere primarily two scales of variability: less than 2000 m andgreater than 2000 m (Fig. 9b). In contrast, Eurasian minnow abun-dance showed a high degree of variability at small scales (<1000 m)and less pronounced patterns of spatial structure at broader
ve sampling by electrofishing for assessing longitudinal disconti-tp://dx.doi.org/10.1016/j.fishres.2016.09.026
scales. Statistically significant (p < 0.05) structure was observedin upstream reaches for bullhead, whereas for stone loach andEurasian minnow, statistically significant (p < 0.05) structure was

ARTICLE IN PRESSG ModelFISH-4562; No. of Pages 12
8 C. Le Pichon et al. / Fisheries Research xxx (2016) xxx–xxx
Fig. 7. Empirical semivariograms for (a) bullhead, (b) stone loach, and (c) Eurasianmtp
ac
4
4
dtmEicccrmGa
Fig. 8. Empirical semivariograms for bullhead with (a) all samples (250–1540 pairs),(b) every second sample (40-m spacing, 65–360 pairs), and (c) every fourth sam-
innow. Variograms depict the semivariance (solid circle) as a function of separa-ion distance between samples with a 40-m lag class interval. The 10th and 90thercentiles are based on 5000 permutations shown as black lines.
pparent in both upstream and downstream reaches (Fig. 9b and).
. Discussion
.1. Patterns in fish assemblage structure and habitat gradients
By using SIS, it was possible to examine spatial patterns andiscontinuities in the distribution of stream fishes and aquatic habi-at in the Rognon stream. Among the 15 species captured, the 3
ost common species were associated with brown trout: bullhead,urasian minnow and stone loach. Although brown trout was low
n abundance, these four associated species comprised the typi-al rheophilic assemblage in this headwater stream. They were notaptured on the section of the stream influenced by obstacles andonnected ponds in the upper section of our study reach. The other
Please cite this article in press as: Le Pichon, C., et al., Spatially intensinuities in fish distribution in a headwater stream. Fish. Res. (2016), ht
heophilic species such as gudgeon, chub and dace, found in theost downstream part of the Rognon, were likely coming from therand Morin River. The other less abundant species were eurytopicnd limnophilic species, living mainly in isolated ponds or slow-
ple (80-m spacing, 18–106 pairs, pairs >30 at separation distance <3150 m). The10th and 90th percentiles are based on 5000 permutations shown as black lines.Minimum separation distance and the lag class interval were 100 m.
flowing parts of large rivers (Schiemer and Waidbacher, 1992).In the Rognon stream, these non-rheophilic species were locateddownstream of two fishing ponds that were connected to the mainstream channel and may have colonized the stream through theoutlets of these ponds. The presence of non-rheophilic ecologicalguilds in small fast-flowing European headwaters has been shownto indicate an alteration of the fish assemblage (Pont et al., 2006).
The SIS approach made it possible to detect more species thancould have been captured with other sampling approaches, such astwo-pass electrofishing at relatively few sites. For example, in a pre-vious study, only 7 species were collected using two-pass removal
ve sampling by electrofishing for assessing longitudinal disconti-tp://dx.doi.org/10.1016/j.fishres.2016.09.026
methods in two sites (60–100 m in length) located in the 5 km-long studied segment of the Rognon stream (Le Pichon et al., 2012).None of the five rare species were detected in this earlier study. Thedetection of these rare species is important for identifying subtle

Please cite this article in press as: Le Pichon, C., et al., Spatially intensive sampling by electrofishing for assessing longitudinal disconti-nuities in fish distribution in a headwater stream. Fish. Res. (2016), http://dx.doi.org/10.1016/j.fishres.2016.09.026
ARTICLE IN PRESSG ModelFISH-4562; No. of Pages 12
C. Le Pichon et al. / Fisheries Research xxx (2016) xxx–xxx 9
Fig. 9. The wavelet power spectrum for species abundance using the Gaussian wavelet for (a) bullhead, (b) stone loach, and (c) Eurasian minnow. The colour gradient fromwhite to dark indicates that 75%, 50%, 25%, and 5% of the wavelet power spectrum is above each level, respectively. The wavelet power spectrum indicates increasing strengthof correspondence between the wavelet power function and the spatial structure of the data at a given scale and location. The black contour line shows the 95% confidencelevel, using the global wavelet as the background spectrum.

ING ModelF
1 es Res
gwwsmmatmn
taubmei(ahftedoo(oapewtu
4s
(2italstbd
opioumfcpaii2d
ARTICLEISH-4562; No. of Pages 12
0 C. Le Pichon et al. / Fisheri
radients in fish abundance and assemblage structure associatedith human impacts (Cao et al., 2001). The complex patterns thate observed in fish distribution may be poorly understood because
ampling is typically not conducted at such a fine resolution overultiple kilometres. Our analysis suggests that sampling in thisanner may be useful to detect anthropogenic influences such
s the effects of obstacles and associated ponds on fish distribu-ion. In addition, SIS could be a more relevant spatially intensive
ethod in rivers with rare or benthic species because snorkeling isot appropriate in this case as shown by Plichard et al. (2016).
The longitudinal distribution of the three common species ofhe typical assemblage, including two benthic species (bullheadnd stone loach), showed contrasting patterns associated with nat-ral and altered habitat gradients. These patterns were of interestecause the spatial distribution of benthic species over tens of kilo-etres is poorly understood due to sampling difficulties (Torgersen
t al., 2004). Bullhead and stone loach were generally associatedn samples in the segment downstream of the Bourgogne stream0–1500 m), where stone loach were abundant (data not shown),nd at the upstream limit of the study segment (>4000 m) where aotspot in bullhead was observed. These segments, dominated by
ast and shallow habitat with coarse substrate, were similar to habi-at preferences that have been documented for these species (Erost al., 2003). These two benthic species were absent in the segmentominated by high proportion of deep, silted and slow habitats. Thebserved distribution of bullhead indicated an increasing impact ofbstacle height on bullhead abundance. As shown by Utzinger et al.1998) bullhead is an indicator species for determining the impactf obstacles on longitudinal connectivity. European minnow weressociated with brown trout in the segment with both riffles andools (1500–3000 m), which are important habitat for trout (Ovidiot al., 2002) and European minnow (Eros et al., 2003). Brown troutere absent upstream of 3000 m, indicating its upstream coloniza-
ion limit, and European minnows were also uncommon in thesepstream reaches.
.2. Detecting patterns in stream fish distribution at multiplecales
By employing SIS at the spatial extent of a stream segment5 km) and at a fine resolution (i.e., spacing of point samples every0 m), we were able to detect a high degree of spatial heterogene-
ty in the distribution of fish species at multiple scales. The methodhat we employed provides additional information on the relativebundance of bullhead and stone loach, emphasizing both fine- andarge-scale trends. Other researchers have used spatially intensiveampling approaches to evaluate discontinuities in fish distribu-ion in relation to tributary confluences (Torgersen et al., 2008),ut the present study is the first to examine discontinuities in theistribution of benthic species with respect to in-stream obstacles.
The complementary use of longitudinal analysis, semivari-grams, and wavelet analysis has potential for detecting complexatterns in fish distribution related to natural and anthropogenic
nfluences. The shape of the semivariogram provides informationn fine-scale patchiness and broad-scale spatial trend that can besed to optimize sampling effort and to help identify factors thatay be influencing patterns of species abundance. Wavelet analysis
acilitated the identification of periodicity and abrupt longitudinalhanges in fish abundance with respect to the location of anthro-ogenic influences. Although wavelet analysis has not been widelypplied in rivers, this tool has been used for investigating variabil-
Please cite this article in press as: Le Pichon, C., et al., Spatially intensinuities in fish distribution in a headwater stream. Fish. Res. (2016), ht
ty at multiple scales simultaneously to detect fine-scale reductionsn water temperature variability caused by dams (Steel and Lange,007) and the influence of tributaries on coastal cutthroat troutistribution (Torgersen et al., 2004).
PRESSearch xxx (2016) xxx–xxx
Fine-scale periodicity in bullhead abundance at distances lessthan 500 m may be related to their longitudinal range of move-ment (Knaepkens et al., 2004; Ovidio et al., 2009) and the size ofstream segments that support subpopulations (Fisher and Kum-mer, 2000). The dispersion capacities of bullhead affect its spatialdistribution (Chaumot et al., 2006). Other studies have shown dif-ferences in bullhead abundance in short adjacent stream sections(below 200 m) in relation to flow conditions (Fischer and Kummer,2000). The large-scale trend (>2500 m) of increased abundance inupstream reaches may be related to higher quality habitat con-ditions. The semivariograms of bullhead distribution showed ahigh degree of spatial structure at fine (∼500 m), intermediate(1000–1500 m), and broad (3000 m) scales, which may indicate thatboth natural and anthropogenic factors may be limiting the distri-bution of this species. The spatial pattern of stone loach revealeda large-scale trend (>2000 m) of decreased abundance in upstreamreaches. The increasing number of obstacles in the intermediatesegment may be limiting the abundance of this species. In con-trast, Eurasian minnow have more general habitat preferences and,thus, had spatial patterns that were highly variable across multiplescales.
In this study, spatially intensive sampling (SIS) was evaluatedfor its capacity to quantify spatial patterns of stream fishes insmall rivers. These data provided information at a spatial resolutionand extent that enabled us to detect patterns over multiple scalesand to examine the potential impact of obstacles on fish assem-blage structure and species abundance. The SIS could contribute toput the “riverscape” perspective into practice, viewing the streamfish assemblage in its entirety over multiple kilometres. This spa-tially continuous approach to data collection was relatively labourintensive (e.g., with a five-persons team, 3 days of sampling wererequired for a 5-km survey), but it provided important and novelinformation on spatial patterns of abundance that has been lackingin studies employing more conventional sampling methods, partic-ularly on benthic fish species. This “riverscape” sampling approachenabled us to investigate fish distribution and habitat structure atintermediate spatial scales where headwater fish species completetheir life history and where restoration and management may bemost effective (Fausch et al., 2002). The development of this sam-pling approach for wadeable streams and the exploration of spatialpatterns described in this study are first steps in applying theseriverscape concepts in practice. The next steps will involve a moreextensive statistical analysis of associations between fish distribu-tion, aquatic habitat, and anthropogenic factors.
The large number of sample points (n = 264) and their spacing20 m apart in the present study were important for examining spa-tial patterns of fish distribution and determining the distances overwhich species abundance was spatially autocorrelated. However,a spatial resolution of 40 m may be sufficient for identifying thespatial scale of autocorrelation and evaluating broader-scale spa-tial trends. As recommended by Som et al. (2014), spatial samplingdesigns need to be optimized based on the objectives of a study. Forstream networks, Som et al. (2014) demonstrated the importanceof sampling directly upstream and downstream of confluences, aswell as having at least one sample at the stream outlet and onesample in the headwaters to better estimate fixed effects in spa-tial statistical models. These authors also suggested that fine-scaleclustering is needed to estimate spatial autocorrelation.
4.3. Management implications
The longitudinal extent and resolution of SIS provided detailed
ve sampling by electrofishing for assessing longitudinal disconti-tp://dx.doi.org/10.1016/j.fishres.2016.09.026
information on fish distribution in the Rognon stream for localmanagers. Localized environmental degradation in this stream wasdetected and has most likely influenced fine- and broad-scale pat-terns in fish assemblage structure and distribution. In addition, the

ING ModelF
es Res
piidpairusWgopmccl
mtctpiete
A
Miiwua
R
A
A
B
B
B
B
C
C
C
C
C
ARTICLEISH-4562; No. of Pages 12
C. Le Pichon et al. / Fisheri
resence of rare non-rheophilic species was an indicator of humanmpacts. In the Rognon stream, local managers may be able to usenformation on where rheophilic fish are locally abundant (e.g., theownstream- and uppermost sections) to establish conservationlans. SIS also was useful for quantifying the spatial extent of thelteration of the fish assemblage (around 500–1000 m for each fish-ng pond) in the Rognon stream. Furthermore, this method waselevant for detecting (i) locations of rare and previously undoc-mented species, (ii) colonization limits for endangered-speciesuch as brown trout, and (iii) hotspots for rheophilic assemblages.
hile multiple-pass electrofishing at distinct sites can be used toive a broad-scale view of fish assemblages in different streamsf the Orgeval catchment, SIS is complementary to elucidate localatterns in habitats and fish assemblage structure to provide infor-ation for local management within a single stream. The approach
ould thus contribute to bridging the gap between research andonservation by focusing on scales at which managers are mostikely to effect change.
Recent reviews have suggested that existing indicators andonitoring tools may be ineffective for evaluating restoration prac-
ices because (1) they have often been developed in a specificontext of organic pollution, and (2) they evaluate status ratherhan trends (Feld et al., 2011; Friberg et al., 2011). The SIS methodrovides comprehensive data to assess the entire fish assemblage
n its spatial context and offers an additional tool in fisheries sci-nce and management for evaluating human impacts and assessinghe effectiveness of riverine restoration efforts and monitoring theffects of stream restoration.
cknowledgements
We are grateful to Guillaume Gorges, Amandine Zahm, Danielira and Guilherm Jan for their invaluable assistance in carry-
ng out field work. Jeff Falke provided constructive comments thatmproved the presentation and clarity of the manuscript. This work
as supported by the CNRS scientific program “Piren-Seine”. Anyse of trade, product, or firm names is for descriptive purposes onlynd does not imply endorsement by the U.S. government.
eferences
ngermeier, P.L., Smogor, R.A., 1995. Estimating number of species and relativeabundances in stream-fish communities: effects of sampling effort anddiscontinuous spatial distributions. Can. J. Fish. Aquat. Sci. 52, 936–949.
ngermeier, P.L., Krueger, K.L., Dolloff, C.A., 2002. Discontinuity in stream-fishdistributions:implications for assessing and predicting species occurrences. In:Scott, J.M., Heglund, P.J., Morrison, M.L., Haufler, J.B., Raphael, M.G., Wall, W.A.,Samson, F.B. (Eds.), Predicting Species Occurrences Issues of Accuracy andScale. Island Press, Washington DC, USA, pp. 519–527.
ateman, D.S., Gresswell, R.E., Torgersen, C.E., 2005. Evaluating single-pass catch asa tool for identifying spatial pattern in fish distribution. J. Freshwater Ecol. 20,335–345.
axter, C.V., 2002. Fish Movement and Assemblage Dynamics in a PacificNorthwest Riverscape. Fish Sci. Corvallis. Oregon State University.
ertrand, K.N., Gido, K.B., Guy, C.S., 2006. An evaluation of single-pass versusmultiple-pass backpack electrofishing to estimate trends in species abundanceand richness in prairie streams. Trans. Kans. Acad. Sci. 109, 131–138.
renkman, S.J., Duda, J.J., Torgersen, C.E., Welty, E., Pess, G.R., Peters, R., Mchenry,M.L., 2012. A riverscape perspective of Pacific salmonids and aquatic habitatsprior to large-scale dam removal in the Elwha River, Washington, USA. Fish.Manage. Ecol. 19, 36–53.
EN, 2003. Water Quality—Sampling of Fish with Electricity, EN 14011:2003.Comité Européen de Normalisation.
ao, Y., Larsen, D., Thorne, R.S.-J., 2001. Rare species in multivariate analysis forbioassessment: some considerations. J. North Am. Benthol. Soc. 20, 144–153.
arle, F.L., Strub, M.R., 1978. A new method for estimating population size fromremoval data. Biometrics 34, 621–630.
hamberland, J.-M., Lanthier, G., Boisclair, D., 2014. Comparison between
Please cite this article in press as: Le Pichon, C., et al., Spatially intensinuities in fish distribution in a headwater stream. Fish. Res. (2016), ht
electrofishing and snorkeling surveys to describe fish assemblages inLaurentian streams. Environ. Monit. Assess. 186, 1837–1846.
haumot, A., Milioni, N., Abdoli, A., Pont, D., Charles, S., 2006. First step of amodeling approach to evaluate spatial heterogeneity in a fish (Cottus gobio)population dynamics. Ecol. Modell. 197, 263–273.
PRESSearch xxx (2016) xxx–xxx 11
Cho, E., Chon, T.-S., 2006. Application of wavelet analysis to ecological data. Ecol.Inf. 1, 229–233.
Copp, G.H., Garner, P., 1995. Evaluating the microhabitat use of fresh-water fishlarvae and juveniles with point abundance sampling by electrofishing. FoliaZool. 44, 145–158.
Copp, G.H., 2010. Patterns of diel activity and species richness in young and smallfishes of European streams: a review of 20 years of point abundance samplingby electrofishing. Fish Fish. 11, 439–460.
Cowx, I.G., Lamarque, P., 1990. Fishing with Electricity: Applications in FreshwaterFisheries Management. Cambridge.
Cressie, N.A.C., 1993. Statistics for Spatial Data. Wiley & Sons, New York.Dunham, J.B., Rosenberger, A.E., Thurow, R.F., Dolloff, C.A., Howell, P.J., 2009.
Coldwater fish in wadeable streams-chapter 8. Standard methods for samplingNorth American freshwater fishes American Fisheries Society, Bethesda,Maryland, pp. 119–138.
Eros, T., Botta-Dukat, Z., Grossman, G.D., 2003. Assemblage structure and habitatuse of fishes in a Central European submontane stream: a patch-basedapproach. Ecol. Freshwater Fish. 12, 141–150.
Fausch, K.D., Torgersen, C.E., Baxter, C.V., Li, H.W., 2002. Landscapes to riverscapes:bridging the gap between research and conservation of stream fishes.Bioscience 52, 483–498.
Feld, C.K., Birk, S., Bradley, D.C., Hering, D., Kail, J., Marzin, A., Melcher, A., Nemitz,D., Pedersen, M.L., Pletterbauer, F., Pont, D., Verdonschot, P.F.M., Friberg, N.,2011. From natural to degraded rivers and back again: a test of restorationecology theory and practice. In: Woodward, G. (Ed.), Adv. Ecol. Res. 44,119–209.
Fischer, S., Kummer, H., 2000. Effects of residual flow and habitat fragmentation ondistribution and movement of bullhead (Cottus gobio L.) in an alpine stream.Hydrobiologia 422, 305–317.
Friberg, N., Bonada, N., Bradley, D.C., Dunbar, M.J., Edwards, F.K., Grey, J., Hayes,R.B., Hildrew, A.G., Lamouroux, N., Trimmer, M., Woodward, G., 2011.Biomonitoring of human impacts in freshwater ecosystems: the good, the badand the ugly. In: Woodward, G. (Ed.), Adv. Ecol. Res. 44, 1–68.
Ganio, L.M., Torgersen, C.E., Gresswell, R.E., 2005. A geostatistical approach fordescribing spatial pattern in stream networks. Front. Ecol. Environ. 3, 138–144.
Greenacre, M.J., 1984. Theory and Applications of Correspondence Analysis.Academic Press, London.
Gresswell, R.E., Torgersen, C.E., Bateman, D.S., Guy, T.J., Hendricks, S.R., Wofford,J.E.B., 2006. A spatially explicit approach for evaluating relationships amongcoastal cutthroat trout, habitat, and disturbance in small Oregon streams. In:Hughes, R.M.L.W., Seelbach, P.W. (Eds.), Influences of Landscapes on StreamHabitats and Biological Assemblages. American Fisheries Society, Bethesda,Maryland.
Hill, M.O., Smith, A.J.E., 1976. Principal component analysis of taxonomic data withmulti-state discrete characters. Taxon 25, 249–255.
Isaak, D.J., Thurow, R.F., Rieman, B.E., Dunham, J.B., 2007. Chinook salmon use ofspawning patches: relative roles of habitat quality, size, and connectivity. Ecol.Appl. 17, 352–364.
Johnson, L.B., Host, G.E., 2010. Recent developments in landscape approaches forthe study of aquatic ecosystems. J. N. Am. Benthol. Soc. 29, 41–66.
Kiffney, P.M., Greene, C.M., Hall, J.E., Davies, J.R., 2006. Tributary streams createspatial discontinuities in habitat, biological productivity, and diversity inmainstem rivers. Can. J. Fish. Aquat. Sci. 63, 2518–2530.
Knaepkens, G., Bruyndoncx, L., Eens, M., 2004. Assessment of residency andmovement of the endangered bullhead (Cottus gobio) in two Flemish rivers.Ecol. Freshwater Fish. 13, 317–322.
Laffaille, P., Briand, C., Fatin, D., Lafage, D., Lasne, E., 2005. Point sampling theabundance of European eel (Anguilla anguilla) in freshwater areas. Arch.Hydrobiol. 162, 91–98.
Lamouroux, N., Capra, H., Pouilly, M., Souchon, Y., 1999. Fish habitat preferences inlarge streams of southern France. Freshwater Biol. 42, 673.
Lawrence, D.J., Olden, J.D., Torgersen, C.E., 2012. Spatiotemporal patterns andhabitat associations of smallmouth bass (Micropterus dolomieu) invadingsalmon-rearing habitat. Freshwater Biol. 57, 1929–1946.
Le Pichon, C., Gorges, G., Boot, P., Baudry, J., Goreaud, F., Faure, T., 2006. A spatiallyexplicit resource-based approach for managing stream fishes in riverscapes.Environ. Manage. 37, 322–335.
Le Pichon, C., Tales, E., Belliard, J., Gorges, G., Zahm, A., Clement, F., 2012. Ladistribution spatiale des peuplements de poissons dans les petits bassinsversants. Sci. Eaux Territoires, 24–33.
Loumagne, C., Tallec, G., 2013. L’observation long terme en environnement:Exemple du bassin versant de l’Orgeval. Versailles: Editions QUAE. 336p.
Macnaughton, C.J., Harvey-Lavoie, S., Senay, C., Lanthier, G., Bourque, G., Legendre,P., Boisclair, D., 2014. A comparison of electrofishing and visual surveyingmethods for estimating fish community structure in temperate rivers. RiverRes. Appl. 31, 1040–1051.
Malard, F., Tockner, K., Ward, J.V., 2000. Physico-chemical heterogeneity in aglacial riverscape. Landscape Ecol. 15, 679–695.
Malavoi, J.R., Souchon, Y., 2002. Description standardisée des principaux facièsd’écoulements observables en rivière: clé de détermination qualitative etmesures physiques. Standardized description of streams and rivers channel
ve sampling by electrofishing for assessing longitudinal disconti-tp://dx.doi.org/10.1016/j.fishres.2016.09.026
geomorphic units: qualitative description key and physical measurements.Bull. Fr. Peche Piscic. 365-366, 357–372.
Meador, M.R., McIntyre, J.P., Pollock, K.H., 2003. Assessing the efficacy ofsingle-pass backpack electrofishing to characterize fish community structure.Trans. Am. Fish. Soc. 132, 39–46.

ING ModelF
1 es Res
M
M
N
O
O
O
P
P
P
P
P
P
P
P
P
R
R
S
S
S
Interdiscip. Rev.: Water 1, 385–400.Wiens, J.A., 2002. Riverine landscapes: taking landscape ecology into the water.
ARTICLEISH-4562; No. of Pages 12
2 C. Le Pichon et al. / Fisheri
itro, M.G., Zale, A.V., 2000. Predicting fish abundance using single-pass removalsampling. Can. J. Fish. Aquat. Sci. 57, 951–961.
ullner, S.A., Hubert, W.A., Wesche, T.A., 1998. Snorkeling as an alternative todepletion electrofishing for estimating abundance and length-classfrequencies of trout in small streams. North Am. J. Fish. Manage. 18, 947–953.
elva, A., Persat, H., Chessel, D., 1979. New method for investigating fishpopulations in large streams by multiple point abundance sampling. C.R. Acad.Sci. 289, 1295.
vidio, M., Baras, E., Goffaux, D., Giroux, F., Philippart, J.C., 2002. Seasonalvariations of activity pattern of brown trout (Salmo trutta) in a small stream, asdetermined by radio-telemetry. Hydrobiologia 470, 195–202.
vidio, M., Detaille, A., Bontinck, C., Philippart, J.C., 2009. Movement behaviour ofthe small benthic Rhine sculpin Cottus rhenanus (Freyhof, Kottelat & Nolte,2005) as revealed by radio-telemetry and pit-tagging. Hydrobiologia 636,119–128.
vidio, M., Capra, H., Philippart, J.C., 2007. Field protocol for assessing smallobstacles to migration of brown trout Salmo trutta, and European graylingThymallus thymallus: a contribution to the management of free movement inrivers. Fish. Manage. Ecol. 14, 41–50.
almer, M.W., 2002. Scale detection using semivariograms and autocorrelograms.In: Gergel, S.E., Turner, M.G. (Eds.), Learning Landscape Ecology: A PracticalGuide to Concepts and Techniques. Springer, New York, pp. 129–144.
erry, J.N., Liebhold, A.M., Rosenberg, M.S., Dungan, J., Miriti, M., Jakomulska, A.,Citron-Pousty, S., 2002. Illustrations and guidelines for selecting statisticalmethods for quantifying spatial pattern in ecological data. Ecography 25,578–600.
ersat, H., Copp, G.H., 1990. Electric fishing and point abundance sampling for theichthyology of large rivers. In: Cowx, I.G. (Ed.), Developments in ElectricFishing Proceedings of an International Symposium on Fishing with Electricity.Fishing News Books Blackwell Scientific Publications, Oxford, pp. 197–210.
lichard, L., Capra, H., Mons, R., Pella, H., Lamouroux, N., 2016. Comparingelectrofishing and snorkelling for characterizing fish assemblages over timeand space. Can. J. Fish. Aquat. Sci. 0, 1–12, http://dx.doi.org/10.1139/cjfas-2015-0578.
ont, D., Hugueny, B., Beier, U., Goffaux, D., Melcher, A., Noble, R., Rogers, C., Roset,N., Schmutz, S., 2006. Assessing river biotic condition at a continental scale: aEuropean approach using functional metrics and fish assemblages. J. Appl. Ecol.43, 70–80.
oole, G.C., 2002. Fluvial landscape ecology: addressing uniqueness within theriver discontinuum. Freshwater Biol. 47, 641–660.
retty, J.L., Harrison, S.S.C., Shepherd, D.J., Smith, C., Hildrew, A.G., Hey, R.D., 2003.River rehabilitation and fish populations: assessing the benefit of instreamstructures. J. Appl. Ecol. 40, 251–265.
révost, E., Nihouarn, A., 1999. Relation entre indicateur d’abondance de type CPUEet estimation de densité par enlèvements successifs pour les juvéniles desaumon atlantique (Salmo salar L.) de l’année. Bull. Fr. Peche Piscic., 19–29.
ringle, C.M., Naiman, R.J., Bretschko, G., Karr, J.R., Oswood, M.W., Webster, J.R.,Welcomme, R.L., Winterbourn, M.J., 1988. Patch dynamics in lotic systems: thestream as a mosaic. J. North Am. Benthol. Soc. 7, 503–524.
eid, S.M., Jones, N.E., Yunker, G., 2008. Evaluation of single-Pass electrofishing andrapid habitat assessment for monitoring redside dace. North Am. J. Fish.Manage. 28, 50–56.
ossi, R.E., Mulla, D.J., Journel, A.G., Franz, E.H., 1992. Geostatistical tools formodeling and interpreting ecological spatial dependence. Ecol. Monogr. 62,277–314.
chiemer, F., Waidbacher, H., 1992. Strategies for conservation of a danubian fishfauna. In: Boon, P.J., Calow, P., Petts, G.E. (Eds.), River Conservation and
Please cite this article in press as: Le Pichon, C., et al., Spatially intensinuities in fish distribution in a headwater stream. Fish. Res. (2016), ht
Management. John Wiley &Sons, Chichester, pp. 363–382.chlosser, I.J., 1991. Stream fish ecology: a landscape perspective. Bioscience 41,
704–712.cholten, M., 2003. Efficiency of point abundance sampling by electro-fishing
modified for short fishes. J.Appl. Ichthyol. 19, 265–277.
PRESSearch xxx (2016) xxx–xxx
Smith, K.L., Jones, M.L., 2005. Watershed-level sampling effort requirements fordetermining riverine fish species composition. Can. J. Fish. Aquat. Sci. 62,1580–1588.
Som, N.A., Monestiez, P., Ver Hoef, J.M., Zimmerman, D.L., Peterson, E.E., 2014.Spatial sampling on streams: principles for inference on aquatic networks.Environmetrics 25, 306–323.
Steel, E.A., Lange, I.A., 2007. Using wavelet analysis to detect changes in watertemperature regimes at multiple scales: effects of multi-purpose dams in theWillamette River basin. River Res. Appl. 23, 351–359.
Tales, E., Berrebi, R., 2007. Controls of local young-of-the-year fish species richnessin flood plain water bodies: potential effects of habitat heterogeneity,productivity and colonisation-extinction events. Ecol. Freshwater Fish. 16,144–154.
Teixeira-de Mello, F., Kristensen, E., Meerhoff, M., González-Bergonzoni, I.,Baattrup-Pedersen, A., Iglesias, C., Kristensen, P., Mazzeo, N., Jeppesen, E., 2014.Monitoring fish communities in wadeable lowland streams: comparing theefficiency of electrofishing methods at contrasting fish assemblages. Environ.Monit. Assess. 186, 1665–1677.
Tomanova, S., Tedesco, P.A., Roset, N., Thomas, R.B.D., Belliard, J., 2013. Systematicpoint sampling of fish communities in medium- and large-sized rivers:sampling procedure and effort. Fish. Manage. Ecol. 20, 533–543.
Torgersen, C.E., Close, D.A., 2004. Influence of habitat heterogeneity on thedistribution of larval Pacific lamprey (Lampetra tridentata) at two spatial scales.Freshwater Biol. 49, 614–630.
Torgersen, C.E., Gresswell, R.E., Bateman, D.S., 2004. Pattern detection in streamnetworks: quantifying spatial variability in fish distribution. In: Nishida, T.,Kailola, P.J., Hollingworth, C.E. (Eds.), GIS-spatial Analyses in Fishery andAquatic Sciences (volume 2): Proceedings of the Second InternationalSymposium on GIS-Spatial Analyses in Fishery and Aquatic Sciences. 3–6September 2002. University of Sussex, Brighton, UK.
Torgersen, C.E., Baxter, C.V., Li, H.W., McIntosh, B.A., 2006. Landscape influences onlongitudinal patterns of river fishes: spatially continuous analysis offish-habitat relationships. In: Hughes, R.M.L.W., Seelbach, P.W. (Eds.),Influences of Landscapes on Stream Habitats and Biological Assemblages.American Fisheries Society, Bethesda, Maryland.
Torgersen, C.E., Gresswell, R.E., Bateman, D.S., Burnett, K.M., 2008. Spatialidentification of tributary impacts in river networks. In: Rice, S., Roy, A.,Rhoads, B. (Eds.), River Confluences, Tributaries and the Fluvial Network. JohnWiley & Sons, Chichester, England, pp. 159–181.
Torrence, C., Compo, G.P., 1998. A practical guide to wavelet analysis. Bull. Am.Meteorol. Soc. 79, 61–78.
Trexler, J., Travis, J., 1993. Nontraditional regression analyses. Ecology 74,1629–1637.
Utzinger, J., Roth, C., Peter, A., 1998. Effects of environmental parameters on thedistribution of bullhead Cottus gobio with particular consideration of theeffects of obstructions. J. Appl. Ecol. 35, 882–892.
Ward, J.V., Stanford, J.A., 1995. The serial discontinuity concept: extending themodel to floodplain rivers, regulated river. Res. Manage. 10, 159–168.
Ward, J.V., Malard, F., Tockner, K., 2002. Landscape ecology: a framework forintegrating pattern and process in river corridors. Landscape Ecol. 17, 35–45.
White, S.M., Rahel, F.J., 2008. Complementation of habitats for bonneville cutthroattrout in watersheds influenced by beavers, livestock, and drought. Trans. Am.Fish. Soc. 137, 881–894.
White, S.M., Giannico, G., Li, H., 2014. A ‘behaviorscape’ perspective on stream fishecology and conservation: linking fish behavior to riverscapes. Wiley
ve sampling by electrofishing for assessing longitudinal disconti-tp://dx.doi.org/10.1016/j.fishres.2016.09.026
Freshwater Biol. 47, 501–515.Williams, B.K., Nichols, J.D., Conroy, M.J., 2002. Analysis and management of
animal populations. Academic Press, New York.