Embed Size (px)
Citation preview

ARTICLE IN PRESS
0038-0717/$ - se
doi:10.1016/j.so
�CorrespondE-mail addr
1Present add
Tsukuba, Sanad2Present add
Sciences, Yokoh
Soil Biology & Biochemistry 38 (2006) 2743–2752
www.elsevier.com/locate/soilbio
Fungal colonization as affected by litter depth anddecomposition stage of needle litter
T. Osono�, D. Hirose1, R. Fujimaki2
Laboratory of Forest Ecology, Graduate School of Agriculture, Kyoto University, Kyoto 606-8502, Japan
Received 19 January 2006; received in revised form 10 April 2006; accepted 11 April 2006
Available online 15 May 2006
Abstract
The present study was designated to evaluate the relative effects of litter depth and decomposition stage of needles on fungal
colonization of needle litter in field experiments. The experiment was carried out in coniferous temperate forests in central Japan. Needle
litter of Chamaecyparis obtusa and Pinus pentaphylla var. himekomatsu at two decomposition stages (recently dead and partly
decomposed) were placed into the organic layer at two depths (on the surface of and beneath the litter layer). Fungal colonization of
needles after 1 year was examined in terms of hyphal abundance and frequency of fungal species. Total and live hyphal length on needles
were affected by the litter depth and (or) the decomposition stage of needles. Length of darkly pigmented hyphae on needles was 1.7–2.6
times greater beneath the litter layer than on the litter surface regardless of the decomposition stage of needles. Length of clamp-bearing
hyphae in Pinus pentaphylla was 5.0–5.2 times greater in partly decomposed needles than in recently dead needles regardless of the litter
depth. Frequencies of Pestalotiopsis spp. and Cladosporium cladosporioides were higher on recently dead needles than on partly
decomposed needles and (or) were higher on the litter surface than beneath the litter layer. Frequencies of Trichoderma, Penicillium, and
Umbelopsis species generally were higher on partly decomposed needles than on recently dead needles and were higher beneath the litter
layer than on the surface.
r 2006 Elsevier Ltd. All rights reserved.
Keywords: Chamaecyparis obtusa; Forest litter; Fungi; Hyphal length; Litterbag; Pinus pentaphylla; Succession
1. Introduction
Successional changes of fungal association are anobvious feature of leaf litter decomposition and have beendescribed on a wide variety of litter species (Hudson, 1968).According to the general schema summarized by Hudson(1968), early colonizers are characterized by ascomycetesand their anamorphs some of which are already present inliving tissues. These fungi soon disappear from decompos-ing leaves and are followed by variable species in otherascomycetes and basidiomycetes at late stages of decom-position. Factors proposed to affect the patterns of fungalsuccession include the variation of micro-environmental
e front matter r 2006 Elsevier Ltd. All rights reserved.
ilbio.2006.04.028
ing author. Tel./fax: +81 75 753 6080.
ess: [email protected] (T. Osono).
ress: Sugadaira Montane Research Centre, University of
a, Nagano 386-2201, Japan.
ress: Graduate School of Environment and Information
ama National University, Yokohama 240-8501, Kanagawa.
conditions within litter and soil horizons, such as moisture(Dix, 1984, 1985; Dix and Frankland, 1987), the changinglitter quality, such as chemical compositions (Osono andTakeda, 2001; Koide et al., 2005), and seasonal variation(Tokumasu, 1998a, b). During decomposition processesunder undisturbed conditions, however, these factorschange simultaneously. For example, the vertical positionof leaf litter within litter layers changes as decompositionprogresses and litter moves downward, which is alwaysaccompanied by the changes in chemical composition. Theincrease of basidiomycetes at later stages of decomposition,for example, thus can be attributable not only to theamelioration of moisture condition in lower litter layers(Dix, 1984; Dix and Frankland, 1987) but also to thedecreased availability of organic chemical components(Osono and Takeda, 2001). Clearly, it is difficult toevaluate the relative importance of individual factors infungal succession by means of observational and descrip-tive methods so far adopted. Manipulative experiments

ARTICLE IN PRESST. Osono et al. / Soil Biology & Biochemistry 38 (2006) 2743–27522744
that discriminate, for example, the effect of the litter depthand the decomposition stage of litter thus will provide auseful insight into the driving factor(s) responsible forfungal succession.
The purpose of the present study was to investigate therelative importance of litter depth and decomposition stageof needles in fungal colonization of needle litter by meansof field experiments. The experiment was carried out inconiferous temperate forests in central Japan. Needle litterof Japanese cypress (Chamaecyparis obtusa) and Japanesewhite pine (Pinus pentaphylla var. himekomatsu) at twodecomposition stages (recently dead and partly decom-posed) were placed into the organic layer at two positions(on the surface of and beneath the litter layer). Recentlydead needles and partly decomposed needles differed inlitter quality, such as chemical composition (Osono et al.,2003). Chamaecyparis obtusa is one of the most importanttrees subject to silvicultural practices in Japan, anddecomposition of Chamaecyparis obtusa needle litter andfungi associated with the decomposition have been studied(Takeda, 1995; Osono et al., 2002, 2003, 2006), whereasthere have been few studies on fungi associated with Pinus
pentaphylla (Hirose et al., 2004). Fungal colonization ofneedles of these tree species were examined after a 1-yearincubation period in terms of hyphal abundance andfrequency of occurrence of fungal species so as to discri-minate the effect of the litter depth and the decompositionstage of needles.
2. Materials and methods
2.1. Study site
The study was carried out in a natural forest ofChamaecyparis obtusa and a plantation of Pinus penta-
phylla var. himekomatsu (28 years old) at KamigamoExperimental Forest Station of Kyoto University (47 ha),about 12 km north of Kyoto City (351040N, 1351430E,140–200m above sea level), central Japan. Annual pre-cipitation of the study site was 1678mm and mean annualtemperature was approximately 15 1C. The study site wason sandstone and slate of the Paleozoic. The Chamaecy-
paris obtusa forest was situated at the top of a hill(Fujimaki et al., 2004). The overstorey of this forestconsisted mainly of Chamaecyparis obtusa. The organiclayer of Chamaecyparis obtusa forest developed well overthe mineral soil horizon and consisted of litter, fermenta-tion, and humus layers ranging from 2 to 5 cm in thickness.The Pinus pentaphylla forest was situated on a hill slope(average inclination 29.71) facing east-north-east (Hirose etal., 2004). The overstorey of this forest consisted only ofPinus pentaphylla. Seedlings of Pinus pentaphylla which wasnot indigenous to this area were grown in a nursery for 3years and then transplanted into a clear-cut site thatformerly had been a mixed forest of Pinus densiflora
and Quercus spp. The Pinus pentaphylla forest has organiclayer consisting of litter layer and thin fermentation layer
ranging from 0.5 to 4 cm in thickness. The two forests werelocated within 1 km of each other. A study plot of10� 10m was laid out within the Chamaecyparis obtusa
and was divided into 10 subplots of 5� 2m. A study plot of6� 16m was laid out within the Pinus pentaphylla forestand was divided into 96 subplots of 1� 1m.
2.2. Litterbag experiment
The litterbag method was used to study fungal coloniza-tion of needle litter (Osono et al., 2003). Recently deadneedles on the canopy and partly decomposed needles fromthe fermentation layer beneath the litter layer werecollected in the study plots in August 2001. The sampleswere taken to the laboratory and oven-dried at 40 1C for 1week. Then, 2 g of needles were enclosed in each litterbag(10� 10 cm), made of polypropylene shade cloth with amesh size of approximately 1mm for Chameacyparis obtusa
and 2mm for Pinus pentaphylla. A total of 80 bags, 40 withChamaecyparis obtusa needles and 40 with Pinus penta-
phylla needles, were prepared. Forty of the 80 bagscontained recently dead needles and the other 40 bagshad partly decomposed needles.Litterbags were placed in 10 subplots of Chamaecyparis
obtusa forest and 10 arbitrarily chosen subplots of Pinus
pentaphylla forest in August 2001. Litterbags with recentlydead and partly decomposed needles of Chamaecyparis
obtusa and Pinus pentaphylla were placed on the forestfloor of the respective forest stand at two depths: on thesurface of and beneath the litter layer. Twenty of the 40bags with recently dead needles and 20 of the 40 bags withpartly decomposed needles were placed on the surface oflitter layer. The other 20 bags with recently dead needlesand the other 20 bags with partly decomposing needleswere inserted beneath the litter layer and were covered bythe litter layer. The bags were attached to the soil by metalpins to prevent movement. Sampling of the bags took placeonce, at 12 months (August 2002) after the placement. All80 litterbags were retrieved and taken to the laboratory.Foreign plant remains attached to the outside of the bags
were carefully removed with forceps. Materials in thelitterbags were separated into needles and fine rootsmanually with forceps. One gram of fresh needles thenwas taken from each litterbag and used for hyphal lengthestimation. Two needles then were taken from eachlitterbag, weighed, and used for fungal isolation. Theseneedles were preserved at 4 1C in a refrigerator until theprocessing started. Hyphal length estimation and fungalisolation were carried out within 48 h after sampling. Theremaining needles were weighed and dried to a constantweight at 40 1C. Mass of needles remaining in the litterbagsthen were determined taking the mass of needles used forhyphal length and fungal isolation into account. Remain-ing mass of needles was expressed as a percentage of theoriginal mass. Water content of needles was calculatedaccording to the following equation: water content (%) ¼mass of water (g)/mass of water plus litter (g)� 100. Mean

ARTICLE IN PRESST. Osono et al. / Soil Biology & Biochemistry 38 (2006) 2743–2752 2745
values were calculated for mass loss and water content ofrecently dead or partly decomposed needles of Chamaecy-
paris obtusa and Pinus pentaphylla.
2.3. Hyphal length estimation
Hyphal length was measured using the agar film methodof Jones and Mollison (1948) but with several modifica-tions according to the method described in Osono et al.(2002). Samples of 1 g (fresh weight) were homogenized in ablender at 10,000 rev min�1 in 49ml of distilled water for3min. The suspension (20ml) was diluted with 20ml ofmolten agar solution (final concentration 1.5%, w/v) andmixed at low speed on a magnetic stirring plate. Two agarfilms were prepared from each suspension in a haemocyt-ometer (0.1mm depth), transferred to glass slides, anddried for 24 h. The films were dual-stained with fluorescentbrightener (FB) and acridine orange (AO), each for 1 h.Fluorescent brightener binds to chitin in fungal cell walls(West, 1988) and enables visualization of all hyalinehyphae that are live or ghost (empty). Acridine orangebinds to nucleic acids in live fungal cells (Rost, 1992) andallows visualization of live hyphae.
The stained films were mounted between slides andcoverslips with one drop of immersion oil (type DF,Cargille Laboratories, Inc., Cedar Grove, NJ, USA) andexamined with a Nikon Microphot-SA epifluorescentmicroscope equipped with a high-intensity mercury lightsource. A Nikon UV-1A filter cube was used forexamination of FB-stained hyphae, and a Nikon B-2Hfilter cube was used for AO-stained hyphae. Darklypigmented hyphae that were not stained with FB, wereobserved by bright field microscopy. Microscope fieldswere selected randomly and 25 fields were observed foreach slide at 1000�magnification. Hyphal lengths wereestimated using an eye-piece grid and a grid-intersectionmethod (Olson, 1950). Total hyphal length was calculatedas the sum of FB-stained hyphal length and the darklypigmented hyphal length. Acridine orange-stained hyphaewere regarded as live. Darkly pigmented hyphae wereregarded as those of dematiaceous fungi. Hyphae withclamp connections were classified into the Basidiomycota.It is recognized, however, that the fungal biomass of theBasidiomycota may have been underestimated because thefrequency of clamp formation varies among species. Thehyphal length was expressed as length per dry mass ofneedles using water content of needles for the conversion.
2.4. Fungal isolation
Fungi were isolated from needles by means of a modifiedwashing method (Harley and Waid, 1955; Tokumasu,1980). Needles were washed in a sterile test tube using avertical type shaker at 2000 rpm for 1.5min. The needleswere washed serially in 5 changes of 0.005% Aerosol-OT(di-2-ethylhexyl sodium sulfosuccinate) solution (w/v) andrinsed with sterilized distilled water 5 times. The needles
were transferred to sterile filter paper in Petri dishes (9 cmin diameter) and dried for 24 h to suppress vigorousbacterial growth after plating (Widden and Parkinson,1973). The needles then were placed on 9 cm Petri dishescontaining lignocellulose agar (LCA) (Miura and Kudo,1970), two needles per plate. LCA contains: glucose 0.1%,KH2PO4 0.1%, MgSO4 � 7H2O 0.02%, KCl 0.02%,NaNO3 0.2%, yeast extract 0.02%, and agar 1.3% (w/v).LCA was used because its low glucose content suppressesovergrowth of fast-growing species and because LCAeffectively induces sporulation and is useful for identifica-tion (Osono and Takeda, 1999). The needles wereincubated at 20 1C in darkness and observed at 3 daysand at 2, 4, and 8 weeks (Osono and Takeda, 1999). Anyfungal hyphae or spores appearing on the plates wereisolated onto fresh LCA plates and incubated andidentified.Mean values were calculated for species richness on each
needle. Frequency of individual species was calculated as apercentage based on the number of needles from which thespecies was found compared to the total number of needlesexamined (20). Fungal species with the frequency of morethan or equal to 25% on any needle types were arbitrarilyregarded as frequent species and the other fungal species asinfrequent species.
2.5. Statistical analysis
A two-way ANOVA was performed to evaluate differ-ences in the remaining mass of needles, water content ofneedles, hyphal length, and mean species richness using thelitter depth (the surface of the litter layer and beneath thelitter layer) and the decomposition stage of needles(recently dead and partly decomposed needles) as indepen-dent variables. Fisher’s exact probability test was per-formed to evaluate the difference in frequency of fungalspecies on needles between the litter depths or thedecomposition stages of needles because data were in theform of proportions. The statistical difference was eval-uated only for frequencies of the frequent species.
3. Results
3.1. Remaining mass and water content
Mass remaining after 1-year of decomposition rangedfrom 70% to 74% of the original mass of Chamaecyparis
obtusa needles and from 49% to 60% of Pinus pentaphylla
needles (Table 1). Remaining mass was not significantly(P40.10) different between the litter depths or thedecomposition stages of needles in both litter species.Water content of needles was 2.0–2.5 times higher in Pinus
pentaphylla than in Chamaecyparis obtusa. Water contentof needles was 1.1–1.3 times higher in needles beneath thelitter layer than in needles on the litter surface for bothlitter species, and the differences were significant (Po0.001for Chamaecyparis obtusa, Po0.10 for Pinus pentaphylla),
ARTICLE IN PRESS
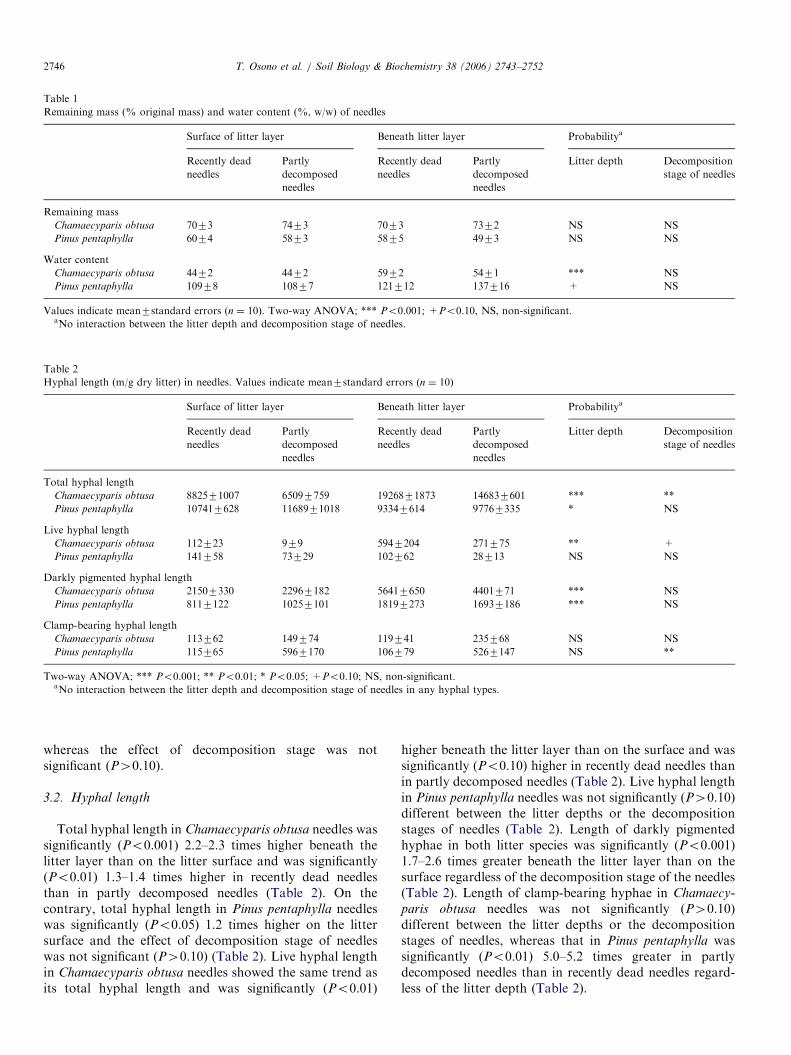
Table 1
Remaining mass (% original mass) and water content (%, w/w) of needles
Surface of litter layer Beneath litter layer Probabilitya
Recently dead
needles
Partly
decomposed
needles
Recently dead
needles
Partly
decomposed
needles
Litter depth Decomposition
stage of needles
Remaining mass
Chamaecyparis obtusa 7073 7473 7073 7372 NS NS
Pinus pentaphylla 6074 5873 5875 4973 NS NS
Water content
Chamaecyparis obtusa 4472 4472 5972 5471 *** NS
Pinus pentaphylla 10978 10877 121712 137716 + NS
Values indicate mean7standard errors (n ¼ 10). Two-way ANOVA; *** Po0.001; +Po0.10, NS, non-significant.aNo interaction between the litter depth and decomposition stage of needles.
Table 2
Hyphal length (m/g dry litter) in needles. Values indicate mean7standard errors (n ¼ 10)
Surface of litter layer Beneath litter layer Probabilitya
Recently dead
needles
Partly
decomposed
needles
Recently dead
needles
Partly
decomposed
needles
Litter depth Decomposition
stage of needles
Total hyphal length
Chamaecyparis obtusa 882571007 65097759 1926871873 146837601 *** **
Pinus pentaphylla 107417628 1168971018 93347614 97767335 * NS
Live hyphal length
Chamaecyparis obtusa 112723 979 5947204 271775 ** +
Pinus pentaphylla 141758 73729 102762 28713 NS NS
Darkly pigmented hyphal length
Chamaecyparis obtusa 21507330 22967182 56417650 4401771 *** NS
Pinus pentaphylla 8117122 10257101 18197273 16937186 *** NS
Clamp-bearing hyphal length
Chamaecyparis obtusa 113762 149774 119741 235768 NS NS
Pinus pentaphylla 115765 5967170 106779 5267147 NS **
Two-way ANOVA; *** Po0.001; ** Po0.01; * Po0.05; +Po0.10; NS, non-significant.aNo interaction between the litter depth and decomposition stage of needles in any hyphal types.
T. Osono et al. / Soil Biology & Biochemistry 38 (2006) 2743–27522746
whereas the effect of decomposition stage was notsignificant (P40.10).
3.2. Hyphal length
Total hyphal length in Chamaecyparis obtusa needles wassignificantly (Po0.001) 2.2–2.3 times higher beneath thelitter layer than on the litter surface and was significantly(Po0.01) 1.3–1.4 times higher in recently dead needlesthan in partly decomposed needles (Table 2). On thecontrary, total hyphal length in Pinus pentaphylla needleswas significantly (Po0.05) 1.2 times higher on the littersurface and the effect of decomposition stage of needleswas not significant (P40.10) (Table 2). Live hyphal lengthin Chamaecyparis obtusa needles showed the same trend asits total hyphal length and was significantly (Po0.01)
higher beneath the litter layer than on the surface and wassignificantly (Po0.10) higher in recently dead needles thanin partly decomposed needles (Table 2). Live hyphal lengthin Pinus pentaphylla needles was not significantly (P40.10)different between the litter depths or the decompositionstages of needles (Table 2). Length of darkly pigmentedhyphae in both litter species was significantly (Po0.001)1.7–2.6 times greater beneath the litter layer than on thesurface regardless of the decomposition stage of the needles(Table 2). Length of clamp-bearing hyphae in Chamaecy-
paris obtusa needles was not significantly (P40.10)different between the litter depths or the decompositionstages of needles, whereas that in Pinus pentaphylla wassignificantly (Po0.01) 5.0–5.2 times greater in partlydecomposed needles than in recently dead needles regard-less of the litter depth (Table 2).

ARTICLE IN PRESS
Table 3
Mean species richnessa on each needle
Surface of litter layer Beneath litter layer Probabilityb
Recently dead
needles
Partly
decomposed
needles
Recently dead
needles
Partly
decomposed
needles
Litter depth Decomposition
stage of needles
Chamaecyparis obtusa 3.070.3 5.170.4 2.370.2 4.670.3 NS ***
Pinus pentaphylla 3.670.4 4.970.4 4.470.4 6.470.4 ** ***
Values indicate mean7standard errors (n ¼ 20). Two-way ANOVA; *** Po0.001; ** Po0.01; NS, non-significant.aMean value for species richness on each needle.bNo interaction between the litter depth and decomposition stage of needles.
Table 4
Summary of the effect of litter depth and decomposition stage of needles
on the frequency of frequent fungi
1. No effect of litter depth or decomposition stage: Clonostachys rosea (P),
Lecanicillium psalliotae (P), Trichoderma koningii (P), Penicillium glabrum
(C), Pestalotiopsis foedans (C)
2. Effect of litter depth
Surface of litter layer Beneath litter layer
Cladosporium cladosporioides (C, P) Chaetomium globosum (Pa)
Penicillium thomii (C) Penicillium miczynskii (Pa)
Pestalotiopsis neglecta (Pa) Penicillium montanense (P)
Umbelopsis ramanniana (Ca) Trichoderma hamatum (P)
Trichoderma polysporum (P)
3. Effect of decomposition stage of needles
Recently dead needles Partly decomposed needles
Aspergillus japonicus (C) Chaetomium globosum (Pa)
Penicillium citreonigrum (P) Penicillium citrinum (P)
Pestalotiopsis neglecta (Pa) Penicillium miczynskii (Pa)
Penicillium thomii (P)
Trichoderma koningii (C)
Trichoderma viride (C, P)
Umbelopsis isabellina (C, P)
Umbelopsis ramanniana (Ca, P)
Fisher’s exact probability test was performed to evaluate the difference at
5% level (see appendices A and B). The species names were indicated when
their frequencies were not significantly different between litter depth or
decomposition stages (1), significantly higher on needles from the surface
of the litter layer or beneath the litter layer (2), or significantly higher on
recently dead needles or partly decomposed needles (3). (C) and (P)
indicate the effect was significant on Chamaecyparis obtusa and Pinus
pentaphylla needles, respectively.aFungal species the frequencies of which were affected by both litter
depth and decomposition stage.
T. Osono et al. / Soil Biology & Biochemistry 38 (2006) 2743–2752 2747
3.3. Species richness and frequency of fungi on needles
A total of 37 fungal species were isolated fromChamaecyparis obtusa needles and 9 of these were regardedas frequent species (Appendix A). A total of 39 fungalspecies were isolated from Pinus pentaphylla needles and 16of these were regarded as frequent species (Appendix B).Mean species richness on each needle of Chamaecyparis
obtusa was significantly (Po0.001) higher on partlydecomposed ones than on recently dead ones and theeffect of litter depth was not significant (P40.10) (Table 3).Mean species richness on each needle of Pinus pentaphylla
was significantly (Po0.01) higher on partly decomposedones than on recently decomposed ones and was signifi-cantly (Po0.001) higher beneath the litter layer than on thelitter surface (Table 3).
The effects of litter depth and decomposition stage ofneedles on the frequency of frequent fungal species aresummarized in Table 4. No significant effects of litter depthor decomposition stage were found for Penicillium glabrum
and Pestalotiopsis foedans on Chamaecyparis obtusa
needles and Clonostachys rosea, Lecanicillium psalliotae,
and Trichoderma koningii on Pinus pentaphylla needles.Frequencies of Cladosporium cladosporioides, Penicillium
thomii (on Chamaecyparis obtusa), Pestalotiopsis neglecta,and Umbelopsis ramanniana were significantly higher onneedles from the litter surface, whereas frequencies ofChaetomium globosum, Penicillium miczynskii, Penicillium
montanense, Trichoderma hamatum, and Trichoderma
polysporum were significantly higher on needles beneaththe litter layer. Frequencies of Aspergillus japonicus,
Penicillium citreonigrum, and Pestalotiopsis neglecta weresignificantly higher on recently dead needles, whereasfrequencies of Chaetamium globosum, Penicillium citrinum,
Penicillium miczynskii, Penicillium thomii (on Pinus penta-
phylla), Trichoderma koningii, Trichoderma viride, Umbe-
lopsis isabellina, and Umbelopsis ramanniana weresignificantly higher on partly decomposed needles.
4. Discussion
The experimental design used in the present study hasthe potential to answer the relative importance of micro-
environmental conditions and substrate quality on fungalcolonization of needles by discriminating between the effectof litter depth and decomposition stage of needles. Verticalpositions of organic layers usually differ in microenviron-mental conditions such as moisture content and tempera-ture. Taylor and Parkinson (1988), for example, found anincrease of moisture content with litter depth. Theexposure to the inoculum source of fungi in air or litterlayers is another factor that can change with litter depths

ARTICLE IN PRESST. Osono et al. / Soil Biology & Biochemistry 38 (2006) 2743–27522748
and potentially affect fungal colonization of litter. Sub-strate quality of needles changes as decompositionprogresses so that the needles at different stages ofdecomposition differ in substrate quality (Berg, 1986;Osono and Takeda, 2001, 2004, 2005; Osono et al., 2006).
4.1. Hyphal length
The higher total hyphal length on recently dead needlesof Chamaecyparis obtusa than on partly decomposedneedles can be primarily ascribed to the difference incontents of organic chemical constituents, such as solublecarbohydrates, extractives, and lignin (Osono et al., 2003).The higher total hyphal length on Chamaecyparis obtusa
beneath the litter layer can be ascribed to the morefavorable moisture condition for fungal growth than on thelitter surface (Table 1). On the contrary, however, totalhyphal length on Pinus pentaphylla needles was lowerbeneath the litter layer in spite of the higher water contentthan on the litter surface (Table 1). This may be possiblydue to the more water-saturated condition beneath thelitter layer that might impede fungal growth. These resultssuggest that not only the changes in substrate quality, aswas suggested by Hasegawa and Takeda (1996) and Osonoand Takeda (2001), but also the downward movement oflitter and concomitant changes in microenvironmentalconditions by themselves can influence the dynamics oftotal hyphal length on litter.
Similarly, the higher live hyphal length on Chamaecy-
paris obtusa needles beneath the litter layer is primarilyascribed to the more favorable moisture conditions(Table 1), and the higher live hyphal length on recentlydead needles of Chamaecyparis obtusa is primarily ascribedto the difference in substrate quality as discussed above fortotal hyphal length. These results are consistent with thoseof previous studies (Berg, 1991; Osono et al., 2003).
The litter depth by itself influenced the length of darklypigmented hyphae regardless of decomposition stages ofneedles. A possible explanation for this difference is themore intensive colonization of needles by dematiaceousfungi beneath the litter layer. This seems to contradict theresult that the frequent fungal species isolated were almostall hyaline (Tables 3 and 4), but it should be borne in mindthat isolation frequency does not necessarily provideinformation about hyphal abundance of individual fungalspecies. In fact, some authors reported the increase of somedematiaceous species with depth of organic layers inconiferous forests (Widden and Parkinson, 1973; Aokiet al., 1990; Tokumasu, 1996).
Clamp-bearing hyphal length (biomass of basidiomyce-tous fungi) was higher in partly decomposed needles ofPinus pentaphylla than in recently dead needles regardlessof the litter depth. Basidiomycetes colonizing Pinus
pentaphylla needle litter can include both litter-decompos-ing and mycorrhizal fungi (Hirose et al., 2004). The changein litter quality during decomposition could be responsiblefor the colonization by litter-decomposing basidiomycetes
(Osono and Takeda, 2001). Alternatively, the greaterlength of basidiomycetous hyphae in partly decomposedneedles suggests the colonization strategy of mycorrhizalbasidiomycetes in the Pinus pentaphylla forest. Hirose et al.(2004) reported that 60% of mycorrhizal biomass wasdistributed at the top soil layer (0–5 cm) in the study site.Mycorrhizal fungi may prefer partly decomposed needlesthat reached the ‘‘mineralization stage’’ of essentialnutrients, such as nitrogen, and had greater nutrientavailability than recently dead needles (Osono and Takeda,2004).
4.2. Species richness and frequency of fungi
Previous studies have indicated that species richness offungi increased at early stages of litter decomposition orincreased with depth of litter and fermentation layers(Visser and Parkinson, 1975; Swift, 1976; Christensen,1984; Osono, 2005). The results of the present study wereconsistent with this pattern and indicated that both litterdepth and decomposition stage of litter can influencespecies richness of fungi on decomposing litter.Species in Pestalotiopsis and Cladosporium are known as
early colonizers whose frequencies are initially high anddecrease during decomposition (Hudson, 1968; Osono andTakeda, 2001; Osono et al., 2003, 2004; Osono, 2005;Koide et al., 2005). The results of the present studysuggested the influence of litter quality for Pestalotiopsis
neglecta on the occurrence of Pinus pentaphylla needles,which is consistent with the previous findings (Osono andTakeda, 2001; Koide et al., 2005). The importance of litterdepth in the occurrence of Chamaecyparis cladosporioides
is noteworthy, as the result indicated that downwardmovement of needles rather than changing litter qualitycould cause the decrease of its frequency. One possibleexplanation for this is the decreasing exposure to airspora with forest floor depth as air spora is a major sourcefor litter colonization by this opportunistic fungalspecies (Osono, 2002, 2005). The higher frequency ofAspergillus japonicus on recently dead needles than onpartly decomposed needles regardless of the litter depthsuggests that litter quality, possibly the content of starch orreadily hydrolysable carbohydrates, is crucial for theoccurrence of this fungus on decomposing needles(Domsch et al., 1980).Species in Trichoderma, Penicillium, and Umbelopsis, on
the other hand, are known as late colonizers whosefrequencies show increasing trends during decomposition(Kendrick and Burges, 1962; Hudson, 1968; Tokumasu,1998a, b; Osono and Takeda, 2001; Osono et al., 2003;Osono, 2005). The higher frequencies of some Penicillium
and Trichoderma species and Chaetamium globosum onPinus pentaphylla needles beneath the litter layer than onthe litter surface suggest the possible importance of theirassociation with basidiomycetes (Hudson, 1968), as thehyphal length of basidiomycetes was greater on needlesbeneath the litter layer (Table 2). On the other hand, the
ARTICLE IN PRESST. Osono et al. / Soil Biology & Biochemistry 38 (2006) 2743–2752 2749
higher frequencies of other Penicillium, Trichoderma, andUmbelopsis species on partly decomposed needles than onrecently dead needles suggest the importance of changinglitter quality during decomposition (Osono and Takeda,2001). The frequency of Penicillium thomii on two litterspecies was affected by different factors: by the litter depthfor Chamaecyparis obtusa needles, whereas by the decom-position stage for Pinus pentaphylla needles. The reason forthis discrepancy is unclear, but other factors such asinteractions with other fungi or soil organisms on eachlitter species may influence the occurrence of fungi morethan the direct effect of microenvironmental conditions orlitter quality.
In conclusion, the present study demonstrated that bothlitter depth and decomposition stage of needles wereimportant factors affecting fungal colonization, but therelative importance of these factors differed depending onhyphal types and fungal species. In most previous studiesof fungal succession during litter decomposition, thereappeared a tendency to ascribe the patterns of fungalsuccession to one of these factors. The present study,however, showed that both litter depth and decomposition
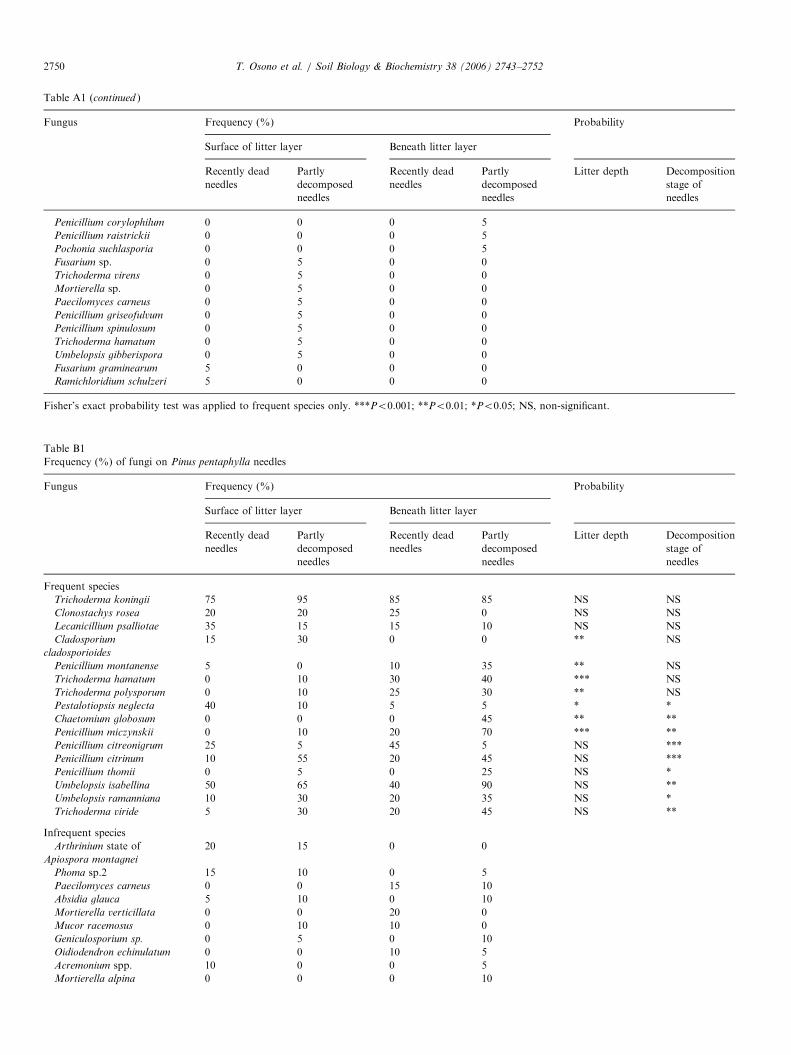
Table A1
Frequency (%) of fungi on Chamaecyparis obtusa needles
Fungus Frequency (%)
Surface of litter layer Be
Recently dead
needles
Partly
decomposed
needles
Re
ne
Frequent species
Penicillium glabrum 35 15 40
Pestalotiopsis foedans 30 25 15
Cladosporium
cladosporioides
20 30 0
Penicillium thomii 15 25 0
Umbelopsis ramanniana 25 65 0
Aspegillus japonicus 40 0 25
Trichoderma koningii 50 75 55
Trichoderma viride 35 80 70
Umbelopsis isabellina 15 45 0
Infrequent species
Lecanicillium psalliotae 5 20 0
Penicillium sclerotiorum 0 15 5
Penicillium miczynskii 0 10 15
Absidia glauca 0 0 0
Mucor hiemalis 0 0 0
Clonostachys rosea 0 5 0
Penicillium decumbens 0 5 5
Penicillium montanense 0 20 0
Penicillium janthinellum 0 0 0
Zygomycete sp. 0 0 0
Fusarium solani 5 5 0
Trichoderma polysporum 0 15 0
Mucor racemosus 0 5 0
Arthrinium state of
Apiospora montagnei
5 0 0
Penicillium citrinum 5 5 0
stage of litter can be driving factors responsible for fungalsuccession.
Acknowledgments
We thank Dr. S. Tokumasu for his helpful identificationof fungi; and Dr. H. Barclay for his critical reading of themanuscript. This study received partial financial supportfrom the Japanese Ministry of Education, Culture andSports (No. 14760099) to T.O.
Appendix A
The Fisher’s exact probability test was performed toevaluate frequency (%) of fungi on Chamaecyparis obtusa
(Table A1)
Appendix B
The Fisher’s exact probability test was performed toevaluate frequency (%) of fungi on Pinus pentaphylla
(Table B1)
Probability
neath litter layer
cently dead
edles
Partly
decomposed
needles
Litter depth Decomposition
stage of
needles
40 NS NS
10 NS NS
0 *** NS
10 * NS
35 ** ***
5 NS ***
90 NS **
75 NS *
40 NS ***
15
10
0
20
20
15
10
0
15
15
5
0
5
5
0
ARTICLE IN PRESS
Table A1 (continued )
Fungus Frequency (%) Probability
Surface of litter layer Beneath litter layer
Recently dead
needles
Partly
decomposed
needles
Recently dead
needles
Partly
decomposed
needles
Litter depth Decomposition
stage of
needles
Penicillium corylophilum 0 0 0 5
Penicillium raistrickii 0 0 0 5
Pochonia suchlasporia 0 0 0 5
Fusarium sp. 0 5 0 0
Trichoderma virens 0 5 0 0
Mortierella sp. 0 5 0 0
Paecilomyces carneus 0 5 0 0
Penicillium griseofulvum 0 5 0 0
Penicillium spinulosum 0 5 0 0
Trichoderma hamatum 0 5 0 0
Umbelopsis gibberispora 0 5 0 0
Fusarium graminearum 5 0 0 0
Ramichloridium schulzeri 5 0 0 0
Fisher’s exact probability test was applied to frequent species only. ***Po0.001; **Po0.01; *Po0.05; NS, non-significant.
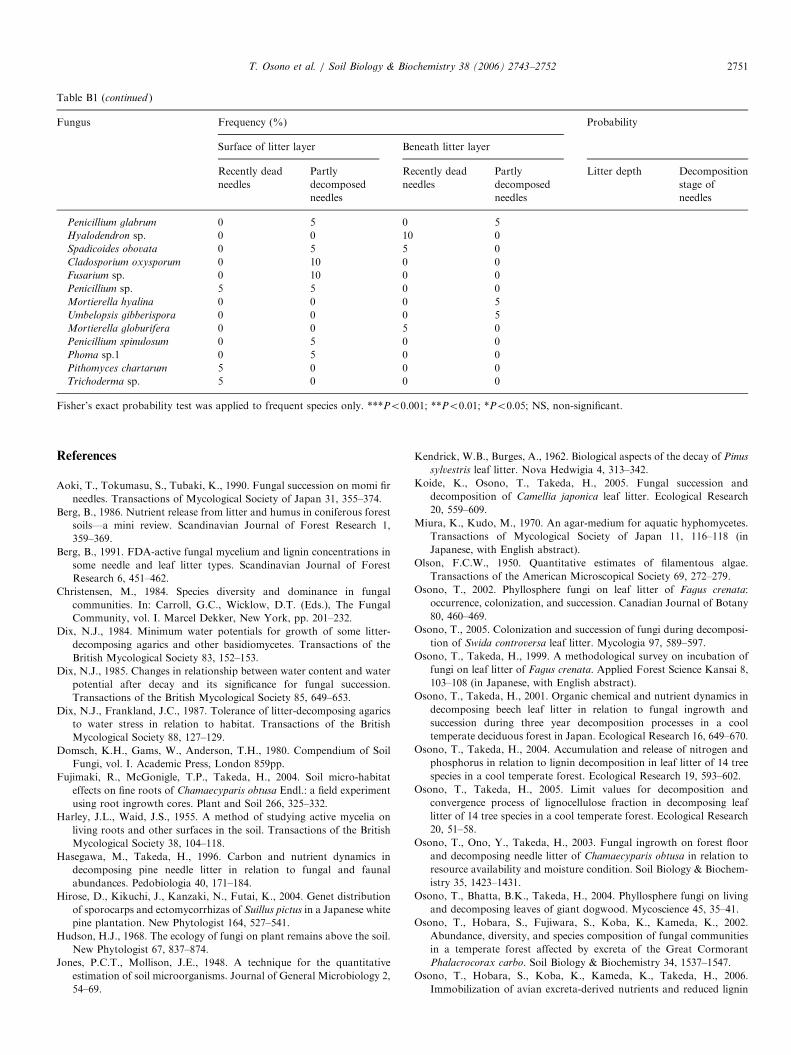
Table B1
Frequency (%) of fungi on Pinus pentaphylla needles
Fungus Frequency (%) Probability
Surface of litter layer Beneath litter layer
Recently dead
needles
Partly
decomposed
needles
Recently dead
needles
Partly
decomposed
needles
Litter depth Decomposition
stage of
needles
Frequent species
Trichoderma koningii 75 95 85 85 NS NS
Clonostachys rosea 20 20 25 0 NS NS
Lecanicillium psalliotae 35 15 15 10 NS NS
Cladosporium
cladosporioides
15 30 0 0 ** NS
Penicillium montanense 5 0 10 35 ** NS
Trichoderma hamatum 0 10 30 40 *** NS
Trichoderma polysporum 0 10 25 30 ** NS
Pestalotiopsis neglecta 40 10 5 5 * *
Chaetomium globosum 0 0 0 45 ** **
Penicillium miczynskii 0 10 20 70 *** **
Penicillium citreonigrum 25 5 45 5 NS ***
Penicillium citrinum 10 55 20 45 NS ***
Penicillium thomii 0 5 0 25 NS *
Umbelopsis isabellina 50 65 40 90 NS **
Umbelopsis ramanniana 10 30 20 35 NS *
Trichoderma viride 5 30 20 45 NS **
Infrequent species
Arthrinium state of
Apiospora montagnei
20 15 0 0
Phoma sp.2 15 10 0 5
Paecilomyces carneus 0 0 15 10
Absidia glauca 5 10 0 10
Mortierella verticillata 0 0 20 0
Mucor racemosus 0 10 10 0
Geniculosporium sp. 0 5 0 10
Oidiodendron echinulatum 0 0 10 5
Acremonium spp. 10 0 0 5
Mortierella alpina 0 0 0 10
T. Osono et al. / Soil Biology & Biochemistry 38 (2006) 2743–27522750
ARTICLE IN PRESS
Table B1 (continued )
Fungus Frequency (%) Probability
Surface of litter layer Beneath litter layer
Recently dead
needles
Partly
decomposed
needles
Recently dead
needles
Partly
decomposed
needles
Litter depth Decomposition
stage of
needles
Penicillium glabrum 0 5 0 5
Hyalodendron sp. 0 0 10 0
Spadicoides obovata 0 5 5 0
Cladosporium oxysporum 0 10 0 0
Fusarium sp. 0 10 0 0
Penicillium sp. 5 5 0 0
Mortierella hyalina 0 0 0 5
Umbelopsis gibberispora 0 0 0 5
Mortierella globurifera 0 0 5 0
Penicillium spinulosum 0 5 0 0
Phoma sp.1 0 5 0 0
Pithomyces chartarum 5 0 0 0
Trichoderma sp. 5 0 0 0
Fisher’s exact probability test was applied to frequent species only. ***Po0.001; **Po0.01; *Po0.05; NS, non-significant.
T. Osono et al. / Soil Biology & Biochemistry 38 (2006) 2743–2752 2751
References
Aoki, T., Tokumasu, S., Tubaki, K., 1990. Fungal succession on momi fir
needles. Transactions of Mycological Society of Japan 31, 355–374.
Berg, B., 1986. Nutrient release from litter and humus in coniferous forest
soils—a mini review. Scandinavian Journal of Forest Research 1,
359–369.
Berg, B., 1991. FDA-active fungal mycelium and lignin concentrations in
some needle and leaf litter types. Scandinavian Journal of Forest
Research 6, 451–462.
Christensen, M., 1984. Species diversity and dominance in fungal
communities. In: Carroll, G.C., Wicklow, D.T. (Eds.), The Fungal
Community, vol. I. Marcel Dekker, New York, pp. 201–232.
Dix, N.J., 1984. Minimum water potentials for growth of some litter-
decomposing agarics and other basidiomycetes. Transactions of the
British Mycological Society 83, 152–153.
Dix, N.J., 1985. Changes in relationship between water content and water
potential after decay and its significance for fungal succession.
Transactions of the British Mycological Society 85, 649–653.
Dix, N.J., Frankland, J.C., 1987. Tolerance of litter-decomposing agarics
to water stress in relation to habitat. Transactions of the British
Mycological Society 88, 127–129.
Domsch, K.H., Gams, W., Anderson, T.H., 1980. Compendium of Soil
Fungi, vol. I. Academic Press, London 859pp.
Fujimaki, R., McGonigle, T.P., Takeda, H., 2004. Soil micro-habitat
effects on fine roots of Chamaecyparis obtusa Endl.: a field experiment
using root ingrowth cores. Plant and Soil 266, 325–332.
Harley, J.L., Waid, J.S., 1955. A method of studying active mycelia on
living roots and other surfaces in the soil. Transactions of the British
Mycological Society 38, 104–118.
Hasegawa, M., Takeda, H., 1996. Carbon and nutrient dynamics in
decomposing pine needle litter in relation to fungal and faunal
abundances. Pedobiologia 40, 171–184.
Hirose, D., Kikuchi, J., Kanzaki, N., Futai, K., 2004. Genet distribution
of sporocarps and ectomycorrhizas of Suillus pictus in a Japanese white
pine plantation. New Phytologist 164, 527–541.
Hudson, H.J., 1968. The ecology of fungi on plant remains above the soil.
New Phytologist 67, 837–874.
Jones, P.C.T., Mollison, J.E., 1948. A technique for the quantitative
estimation of soil microorganisms. Journal of General Microbiology 2,
54–69.
Kendrick, W.B., Burges, A., 1962. Biological aspects of the decay of Pinus
sylvestris leaf litter. Nova Hedwigia 4, 313–342.
Koide, K., Osono, T., Takeda, H., 2005. Fungal succession and
decomposition of Camellia japonica leaf litter. Ecological Research
20, 559–609.
Miura, K., Kudo, M., 1970. An agar-medium for aquatic hyphomycetes.
Transactions of Mycological Society of Japan 11, 116–118 (in
Japanese, with English abstract).
Olson, F.C.W., 1950. Quantitative estimates of filamentous algae.
Transactions of the American Microscopical Society 69, 272–279.
Osono, T., 2002. Phyllosphere fungi on leaf litter of Fagus crenata:
occurrence, colonization, and succession. Canadian Journal of Botany
80, 460–469.
Osono, T., 2005. Colonization and succession of fungi during decomposi-
tion of Swida controversa leaf litter. Mycologia 97, 589–597.
Osono, T., Takeda, H., 1999. A methodological survey on incubation of
fungi on leaf litter of Fagus crenata. Applied Forest Science Kansai 8,
103–108 (in Japanese, with English abstract).
Osono, T., Takeda, H., 2001. Organic chemical and nutrient dynamics in
decomposing beech leaf litter in relation to fungal ingrowth and
succession during three year decomposition processes in a cool
temperate deciduous forest in Japan. Ecological Research 16, 649–670.
Osono, T., Takeda, H., 2004. Accumulation and release of nitrogen and
phosphorus in relation to lignin decomposition in leaf litter of 14 tree
species in a cool temperate forest. Ecological Research 19, 593–602.
Osono, T., Takeda, H., 2005. Limit values for decomposition and
convergence process of lignocellulose fraction in decomposing leaf
litter of 14 tree species in a cool temperate forest. Ecological Research
20, 51–58.
Osono, T., Ono, Y., Takeda, H., 2003. Fungal ingrowth on forest floor
and decomposing needle litter of Chamaecyparis obtusa in relation to
resource availability and moisture condition. Soil Biology & Biochem-
istry 35, 1423–1431.
Osono, T., Bhatta, B.K., Takeda, H., 2004. Phyllosphere fungi on living
and decomposing leaves of giant dogwood. Mycoscience 45, 35–41.
Osono, T., Hobara, S., Fujiwara, S., Koba, K., Kameda, K., 2002.
Abundance, diversity, and species composition of fungal communities
in a temperate forest affected by excreta of the Great Cormorant
Phalacrocorax carbo. Soil Biology & Biochemistry 34, 1537–1547.
Osono, T., Hobara, S., Koba, K., Kameda, K., Takeda, H., 2006.
Immobilization of avian excreta-derived nutrients and reduced lignin

ARTICLE IN PRESST. Osono et al. / Soil Biology & Biochemistry 38 (2006) 2743–27522752
decomposition in needle and twig litter in a temperate coniferous
forest. Soil Biology & Biochemistry 38 in press.
Rost, F.W.D., 1992. Fluorescence Microscopy, vol 2. Cambridge
University Press, Cambridge 473pp.
Swift, M.J., 1976. Species diversity and the structure of microbial
communities in terrestrial habitats. In: Anderson, J.M., Macfadyen,
A. (Eds.), The Role of Terrestrial and Aquatic Organisms in
Decomposition Processes. Blackwell, Oxford, pp. 185–222.
Takeda, H., 1995. A 5 year study of litter decomposition processes in a
Chamaecyparis obtusa Endl. forest. Ecological Research 10, 95–104.
Taylor, B.R., Parkinson, D., 1988. Does repeated wetting and drying
accelerate decay of leaf litter? Soil Biology & Biochemistry 20,
647–656.
Tokumasu, S., 1980. Observations on the fungal flora on pine needle litter.
In: Biseibutsu no Seitai, vol. 7. Gakkai Shuppan Center, Tokyo, pp.
129–144.
Tokumasu, S., 1996. Mycofloral succession on Pinus densiflora needles on
a moder site. Mycoscience 37, 313–321.
Tokumasu, S., 1998a. Fungal successions on pine needles fallen at
different seasons: the succession of surface colonizers. Mycoscience 39,
409–416.
Tokumasu, S., 1998b. Fungal successions on pine needles fallen at
different seasons: the succession of interior colonizers. Mycoscience 39,
417–423.
Visser, S., Parkinson, D., 1975. Fungal succession on aspen poplar leaf
litter. Canadian Journal of Botany 53, 1640–1651.
West, A.W., 1988. Specimen preparation, stain type, and extraction and
observation procedures as factors in the estimation of soil mycelial
lengths and volumes by light microscopy. Biology and Fertility of Soils
7, 88–94.
Widden, P., Parkinson, D., 1973. Fungi from Canadian coniferous forest
soils. Canadian Journal of Botany 51, 2275–2290.











![Buergers, Fungal Genomics & Biology · denture cleansers over mechanical brushing [4]. A great variety of ... C. albicans colonization on a soft denture relining material. Materials](https://img.pdfslide.us/doc/110x75/5b3a81367f8b9a4b0a8df977/buergers-fungal-genomics-biology-denture-cleansers-over-mechanical-brushing.jpg)

















