Embed Size (px)
Citation preview

92:331-342, 2002. ;J Appl Physiol Bandler and Ronald M. HarperLuke A. Henderson, Pearl L. Yu, Robert C. Frysinger, Jean-Philippe Galons, Richardfunctional magnetic resonance imagingNeural responses to intravenous serotonin revealed by
You might find this additional info useful...
49 articles, 13 of which you can access for free at: This article citeshttp://jap.physiology.org/content/92/1/331.full#ref-list-1
including high resolution figures, can be found at: Updated information and serviceshttp://jap.physiology.org/content/92/1/331.full
can be found at: Journal of Applied Physiology about Additional material and informationhttp://www.the-aps.org/publications/jappl
This information is current as of February 23, 2013.
http://www.the-aps.org/. © 2002 the American Physiological Society. ISSN: 8750-7587, ESSN: 1522-1601. Visit our website at year (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyrightphysiology, especially those papers emphasizing adaptive and integrative mechanisms. It is published 12 times a
publishes original papers that deal with diverse area of research in appliedJournal of Applied Physiology
at Penn S
tate Univ on F
ebruary 23, 2013http://jap.physiology.org/
Dow
nloaded from

Neural responses to intravenous serotonin revealed byfunctional magnetic resonance imaging
LUKE A. HENDERSON,1,2 PEARL L. YU,1 ROBERT C. FRYSINGER,1
JEAN-PHILIPPE GALONS,3 RICHARD BANDLER,2 AND RONALD M. HARPER1
1Department of Neurobiology, University of California at Los Angeles, California 90095-1763;2Department of Anatomy and Histology and Pain Management and Research Center,Royal North Shore Hospital, University of Sydney, Sydney, New South Wales 2006, Australia;and 3Department of Radiology, University of Arizona, Tucson, Arizona 85724Received 29 August 2001; accepted in final form 17 September 2001
Henderson, Luke A., Pearl L. Yu, Robert C. Fry-singer, Jean-Philippe Galons, Richard Bandler, andRonald M. Harper. Neural responses to intravenous sero-tonin revealed by functional magnetic resonance imaging. JAppl Physiol 92: 331–342, 2002.—We examined the sequenceof neural responses to the hypotension, bradycardia, andapnea evoked by intravenous administration of 5-hydroxy-tryptamine (serotonin). Functional magnetic resonance imag-ing signal changes were assessed in nine isoflurane-anesthe-tized cats during baseline and after a bolus intravenous lowdose (10 mg/kg) or high dose (20–30 mg/kg) of 5-hydroxytryp-tamine. In all cats, high-dose challenges elicited rapid-onset,transient signal declines in the intermediate portion of thesolitary tract nucleus, caudal midline and caudal and rostralventrolateral medulla, and fastigial nucleus of the cerebel-lum. Slightly delayed phasic declines appeared in the dentateand interpositus nuclei and dorsolateral pons. Late-develop-ing responses also emerged in the solitary tract nucleus,parapyramidal region, periaqueductal gray, spinal trigemi-nal nucleus, inferior olivary nucleus, cerebellar vermis, andfastigial nucleus. Amygdala and hypothalamic sites showeddelayed and prolonged signal increases. Intravenous seroto-nin infusion recruits cerebellar, amygdala, and hypothalamicsites in addition to classic brain stem cardiopulmonary areasand exhibits site-specific temporal patterns.
myocardial ischemia; Bezold-Jarisch reflex; bradycardia; ap-nea; hypotension
MECHANICAL OR CHEMICAL STIMULATION of cardiopulmo-nary afferents, such as the aortic plexus, myocardialreceptors, or bronchopulmonary C fibers, elicits a pro-found hypotension, bradycardia, and apnea known asthe Bezold-Jarisch depressor reflex. The reflex is ofconsiderable clinical interest, because loss of perfusionfrom the accompanying hypotension may be life threat-ening and rapid intervention is often necessary forsurvival (9, 29). The integrity of the reflex may play arole in compensatory cardiovascular responses in myo-cardial ischemia or aortic stenosis (38, 42). Althoughthe focus of attention for eliciting the reflex has beendirected principally on stimulation of afferents associ-
ated with myocardial damage, other conditions, suchas acute hypovolemia (19) or deep pain (24), may trig-ger similar physiological sequences.
Since the first description of the Bezold-Jarisch re-flex by Von Bezold and Hirt in the late 19th centuryand Jarisch in the early 20th century (23), a range ofsubstances have been identified that elicit this reflex,including plant alkaloids; nicotine; capsaicin; venomsfrom snakes, insects, and marine animals; histamine;and serotonin [5-hydroxytryptamine (5-HT)] (1). Studiesemploying electrophysiological recording, stimulation,and lesion techniques have begun to define the neuralstructures responsible for the reflex expression. Com-ponents of the circuitry responsible for the barorecep-tor reflex appear to mediate hypotensive and brady-cardic components of the Bezold-Jarisch reflex [e.g.,nucleus of the solitary tract (NTS), caudal ventrolat-eral medulla (CVLM), and rostral ventrolateral me-dulla (RVLM)] (11, 48). However, certain medullarystructures, such as the caudal midline medulla (CMM,including the nucleus raphe pallidus and obscurus),not normally associated with baroreflex control, appearto be recruited during stimulation by 5-HT (46). Theanatomic organization of medullary and more rostralsites involved in airway reflexes has been outlinedearlier (12). However, the sequence of activation ofthese structures responsible for expression of apneaduring airway reflexes and the extent of contributionsfrom these areas remain unknown. Although single-cell recording and stimulation and lesion studies canreveal specific functions of a localized brain region,determination of the relative onset of activity withinactivated neural sites during this challenge requiressimultaneous assessment of widespread neural areas,a difficult task for microelectrode recording or stimu-lation/lesion studies.
Functional magnetic resonance imaging (fMRI) pro-cedures provide a means to evaluate activity changesin widespread brain sites without use of systemic con-trast agents, ionizing radiation, implanted electrodes,
Address for reprint requests and other correspondence: R. M.Harper, Dept. of Neurobiology, Univ. of California, Los Angeles, CA90095-1763 (E-mail: [email protected]).
The costs of publication of this article were defrayed in part by thepayment of page charges. The article must therefore be herebymarked ‘‘advertisement’’ in accordance with 18 U.S.C. Section 1734solely to indicate this fact.
J Appl Physiol92: 331–342, 2002.
8750-7587/02 $5.00 Copyright © 2002 the American Physiological Societyhttp://www.jap.org 331
at Penn S
tate Univ on F
ebruary 23, 2013http://jap.physiology.org/
Dow
nloaded from

or other brain intervention. The origin and sequence ofneural activity responses to challenges can be assessedrapidly over the entire brain, assisting determinationof functional organization of responses to physiologicalchallenges. We used the blood oxygen level-dependent(BOLD) technique, which is based on the principle thatactivated brain areas undergo increased local bloodflow and volume, becoming relatively more oxygenatedthan surrounding areas. Because oxygenated blood ex-hibits different paramagnetic properties, activated re-gions show small magnetic signal changes (33). Unlikefunctional neuroanatomy procedures, such as c-Fos or2-deoxyglucose mapping techniques, fMRI proceduresallow repeatable assessments with minimal injury tothe subject.
We used BOLD imaging techniques in adult cats tovisualize brain regions that respond to activation of thecardiopulmonary afferents by intravenous administra-tion of 5-HT. Intravenous 5-HT administration hasbeen used to stimulate cardiopulmonary afferentswhile avoiding concerns of agent transmission throughthe blood-brain barrier (13, 44). Stimulation effectsfrom intravenous 5-HT administration are very shortlasting (50), thus minimizing accumulation of effectsfrom multiple trials. The primary objectives were to 1)determine the neural substrates involved in mediatingarterial pressure, heart rate, and respiratory responsesto stimulation of cardiopulmonary afferents and 2)evaluate the temporal sequence of neural activation indifferent areas after the afferent stimulation.
METHODS
Under isoflurane anesthesia, nine adult cats (3 male, 6female; 2–5.9 kg) were intubated, and the left or right carotidartery and jugular vein were cannulated for the measure-ment of arterial pressure and administration of pharmaco-logical agents. The tip of the jugular cannula was inserted tothe right atrium. Silver wire was placed subcutaneously inthe left and right thoracic walls for recording of electrocar-diographic (ECG) activity. Magnetic resonance imaging ofthe brain requires head immobility and a means to provideconsistent interanimal head positioning within the scannerto ensure comparable neural views across different subjectswith reference to a standard stereotactic atlas. Under isoflu-rane anesthesia, cats were placed in a Kopf stereotacticdevice, the dorsal surface of the skull was exposed via amidline incision, and a headpiece was attached to the skullby nylon screws and by dental cement in an opened frontalsinus. The headpiece consisted of a Plexiglas tube that laterslipped over a mating rod in a larger carrier to support theanimal. This carrier fit comfortably into a 60-mm-diameterDoty head coil (Doty Scientific, Columbia, SC) in the scanner(17).
Echo-planar fMRI procedures are especially sensitive tomagnetic field inhomogeneities. Such inhomogeneities arecreated by the bony tentorium between the cerebellum andcerebrum, cerebrospinal-brain tissue interfaces, air-filled si-nuses, and ferrous particles produced by the drill bits duringsurgery. This sensitivity can lead to excessive signal drop-outor to very large (.15%) signal changes, which were excludedfrom analysis in this series of studies. To reduce magneticfield inhomogeneities from ferrous particles, the frontal si-nuses were well flushed with saline, and, later in the series,
a diamond-tipped drill bit replaced the steel bit. A small bagof deuterium oxide (heavy water, 2H2O) was placed immedi-ately dorsal to the cerebellum on the back of the neck toreduce magnetic field inhomogeneities created by skull struc-tures near the caudal brain. These two procedures substan-tially reduced artifacts.
The ECG leads were led to a high common mode rejectionand low-noise operational amplifier inside the scanner areaand were coupled by fiber-optic cables to a receiver locatedoutside of the magnetic resonance imaging room (34). In theabsence of a magnetic resonance-compatible pneumotachom-eter, an indication of respiratory activity was provided byassessing thoracic wall breathing movements using a small,sealed, air-filled bag attached to nondistensible plastic tub-ing, placed between the thoracic wall of the cat and the wallof the carrier. The plastic tubing was then led external to thescanner to a pressure transducer (PX138 series, Omega En-gineering, Stamford, CT). Low-compliance arterial cannulaswere led outside the scanner shielding for connection to anarterial pressure monitor. Body temperature was assessed byusing an optically coupled rectal probe and was maintainedbetween 37.0 and 38.0°C by using a magnetic resonanceimaging-compatible heating blanket (Gaymar, OrchardPark, NY). Animals were maintained on 1% isoflurane dur-ing the entire procedure. A magnetic resonance imaging-compatible pulse oximeter was used to record O2 saturation.All physiological signals were recorded digitally by using theDaqEz analog-to-digital system (Quatech). The externaliza-tion of physiological signal transduction with fluid- or air-filled transfer media or optic coupling minimized noise intro-duced into the fMRI images and, conversely, reduced noiseintroduced into the physiological signals produced by thechanging scanner magnetic fields.
The Echo-planar technique (8) was used to visualize rapidbrain signal changes. A set of anatomic images was acquiredby a 4.7-T Bruker scanner using a transverse relaxation timerapid acquisition with relaxation enhancement (T2 RARE)protocol, followed by a time series of 45 gradient-echo imagesets, composed of 19 coronal sections, acquired during each5-HT or saline challenge (repetition time 5 1 s, 8 interleavedscans per image set, echo time 5 25 ms, flip angle 5 90°, fieldof view 5 8 cm, thickness 5 2 mm, no interslice gap). Voxelsizes were 0.625 3 0.625 3 2 mm (the latter, slice thickness).
The 45 image sets were acquired over an uninterrupted6-min period. The first 3 min consisted of baseline imageacquisition, after which a challenge of high-dose (20 or 30mg/kg) or low-dose (10 mg/kg) 5-HT was delivered intrave-nously as a single bolus; the challenge was followed by anadditional 3 min of image collection. All doses were dissolvedin 0.5 ml of saline and were followed immediately by a 5-mlsaline flush (5.5 ml total). An equivalent bolus volume ofnormal saline (5.5 ml) was administered over an equivalenttime period as the 5-HT bolus (4 s), as a control dose. At theend of the studies, the animals were euthanized with anoverdose of pentobarbital sodium. All procedures were ap-proved by the University of Arizona Institutional ReviewBoard.
The arterial pressure, ECG, and thoracic wall movementtraces were digitized at 1 kHz. Inspiratory and expiratoryonset and offset times and amplitude of thoracic wall move-ments were determined by a peak-searching algorithm, andthe extent of thoracic wall movements and respiratory rateswere determined from those calculations. Mean values for100-ms successive epochs of arterial pressure, heart rate,respiratory rate, and thoracic wall movement extent werecalculated over the entire baseline and challenge periods andare averaged over animals.
332 NEURAL CHANGES TO INTRAVENOUS 5-HT
J Appl Physiol • VOL 92 • JANUARY 2002 • www.jap.org
at Penn S
tate Univ on F
ebruary 23, 2013http://jap.physiology.org/
Dow
nloaded from

The first image set (8 s) was excluded from analysisbecause the technical characteristics of the scan procedurerequired a settling time for adequate signal assessment,leaving 44 image sets for analysis, 22 during baseline and22 during challenge. The MedX image analysis package(Sensor Systems, Sterling, VA) was used to evaluate func-tional image signal changes during each challenge, rela-tive to the corresponding baseline period. Animation ofsuccessive image sets was used to provide an initial visualassessment of movement during the scanning period;movement was also tracked by plotting values for thecenter of intensity of one slice per volume over time toassess whether significant motion emerged. No detectablemotion artifact, defined as sudden changes twice the base-line noise level, was apparent, largely because the animalswere anesthetized and because the head restraint mini-mized movement (17); therefore, no motion correction wasperformed. All image sets were ratio intensity normalizedso that mean intensity and standard deviation values wereequalized. Gaussian smoothing was performed with X 51.3, Y 5 1.3, Z 5 1 mm. Significant changes in voxelintensity were determined by using a t-test. Nonsignificantvoxels were removed from the volume, leaving only thosevoxels reaching a probability of P , 0.005. The percentchanges in signal intensity (threshold values: 60.5 to615% change from baseline) of these significant voxelswere then displayed onto a set of T2 RARE anatomicimages.
Regions of interest were selected, the boundaries of whichwere determined by anatomic criteria. In each animal, signalintensities of all voxels within a region of interest wereaveraged for each time point, and these signal intensitieswere then averaged across all animals. The mean signalintensity values (6SE) at each time point were expressed asa percent change from baseline. Statistical comparisons withbaseline were calculated for each time point using repeated-measures ANOVA (P , 0.005).
RESULTS
Physiology
Intravenous saline. The mean resting arterial pres-sure, heart rate, and respiratory rate values beforesaline administration were 109 6 9 mmHg, 147 6 10beats/min, and 23 6 5 breaths/min, respectively. Forall cats, intravenous saline resulted in a small pertur-bation in arterial pressure and heart rate but not inrespiratory rate (Fig. 1C).
Intravenous 5-HT. ARTERIAL PRESSURE. In seven ofeight animals, high-dose (n 5 11) and low-dose (n 5 7)5-HT evoked a triphasic arterial pressure response(Fig. 1, A and B) (1 of the 9 animals was excludedbecause of scanner-induced artifactual noise). Within5 s of high-dose 5-HT administration onset, arterialpressure fell 18 6 5 mmHg (baseline 5 120 6 6mmHg); the fall was transient (;10 s to maximumdecrease) and returned to 3 6 3 mmHg below baseline.A prolonged hypotension followed, reaching 25 6 5mmHg below baseline and returning to baseline ;500s after challenge onset. The remaining animal showeda similar initial decrease but no late hypotension. Pat-terns after low-dose administration were similar butwith reduced magnitude (Fig. 1B).
HEART RATE. In all animals, high-dose 5-HT adminis-tration resulted in a biphasic change in heart rate.Within 5 s of the high-dose challenge, heart rate beganto decline, reaching a nadir of 118 6 5 beats/min(baseline 5 144 6 6 beats/min). This transient brady-cardia was followed by a prolonged tachycardia, reach-ing a maximal increase of 29 6 6 beats/min abovebaseline. Heart rate then returned to baseline within
Fig. 1. Traces of arterial pressure, heart rate (in beats/min), respiratory rate (in breaths/min), and extent ofthoracic wall movement during baseline and after bolus intravenous delivery of 20–30 mg/kg 5-hydroxytryptamine(5-HT) (A), 10 mg/kg 5-HT (B), and saline (C). Vertical dashed lines indicate start of challenge. Values are means 6 SE.
333NEURAL CHANGES TO INTRAVENOUS 5-HT
J Appl Physiol • VOL 92 • JANUARY 2002 • www.jap.org
at Penn S
tate Univ on F
ebruary 23, 2013http://jap.physiology.org/
Dow
nloaded from

300 s of 5-HT delivery. Low-dose heart rate patternswere identical but were reduced in amplitude.
RESPIRATION. Breathing was markedly affected by5-HT in a dose-dependent fashion. Within 5 s of high-dose administration, an apnea ensued, with cessationof breathing efforts for 30 6 7 s (range 8–100 s). Afterthe apnea, respiratory rate returned to baseline (27 62 breaths/min) within 2 s and then increased to a level37 6 5 breaths/min above baseline for ;300 s. Themagnitude of thoracic wall movements also returned tobaseline, after an increase (34 6 11% above baseline)for ;150 s. Low-dose administration resulted in areduced apnea duration 12 6 3 s (range 5–30 s) and areduced compensatory later tachypnea (22 6 6% abovebaseline) for ;150 s.
Distribution of fMRI Signal Changes
Intravenous saline. Intravenous administration ofsaline resulted in no significant change in signal inten-sity in any area of the brain (Fig. 2).
Intravenous 5-HT. In contrast to intravenous saline,5-HT evoked significant changes in signal intensity ina number of brain structures. These changes were notconfined to medullary regions classically associatedwith mediation of changes in arterial pressure, heartrate, and breathing, but they also included rostralbrain structures and sites within the cerebellum. Thesignal changes, in most cases, were dose dependent.
Three general trends were observed to high-dose 5-HTchallenge. 1) Medullary and some cerebellar sites showed
a rapid-onset transient decline in signal intensity, fol-lowed by a fast recovery and sustained signal increase; insome cases, activity remained below baseline. 2) In thedentate and interpositus cerebellar nuclei and dorsolat-eral pons, the initial transient change was delayed. 3) Inrostral brain sites, as well as the inferior olive, parapy-ramidal region, and spinal trigeminal nucleus, activityshowed a delayed onset, sustained increase.
EARLY-ONSET TRANSIENT DECLINE. The changes in signalintensity are shown as pseudocolored overlays on ana-tomic views as well as mean 6 SE values across base-line and challenge for different areas in Figs. 3–5. Inmedullary sites of the intermediate NTS (NTS regionat the level of and rostral to the obex), RVLM, CVLM,and CMM (Fig. 3), an initial decline in signal intensityoccurred (intermediate NTS maximum fall 5 2.3 6 1%;RVLM maximum fall 5 0.8 6 0.8%); the declineemerged slightly later (first 16 s) within the CVLM(maximum fall 5 1.7 6 0.9%). A large transient de-crease in signal intensity also appeared within theCMM, falling 2.2 6 1% within the first 8 s and return-ing to baseline at 24 s. This transient decline was theonly response observed in the CMM; no significant latechange emerged. The fastigial nucleus also showed animmediate phasic decline (maximum decrease 5 1.4 60.4%); however, this decline was more prolonged thanother medullary sites.
After the initial phasic decline, and concomitantwith the end of the apnea and the onset of the pro-longed hypotensive phase (24 s after 5-HT), signal
Fig. 2. Signal-intensity changes in re-gions of the medulla (A), cerebellum(B), pons and midbrain (C), and hypo-thalamus and amygdala (D) after anequivalent volume of intravenous sa-line. Vertical dashed lines indicatestart of challenge. Values are means 6SE.
334 NEURAL CHANGES TO INTRAVENOUS 5-HT
J Appl Physiol • VOL 92 • JANUARY 2002 • www.jap.org
at Penn S
tate Univ on F
ebruary 23, 2013http://jap.physiology.org/
Dow
nloaded from
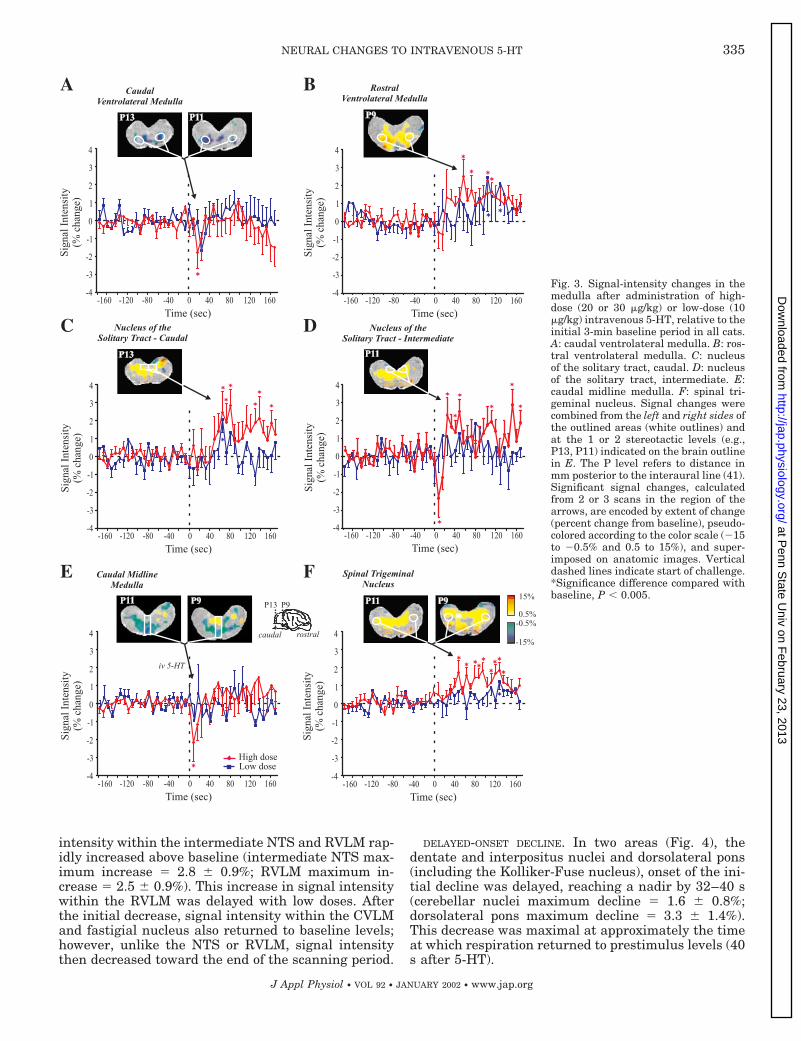
intensity within the intermediate NTS and RVLM rap-idly increased above baseline (intermediate NTS max-imum increase 5 2.8 6 0.9%; RVLM maximum in-crease 5 2.5 6 0.9%). This increase in signal intensitywithin the RVLM was delayed with low doses. Afterthe initial decrease, signal intensity within the CVLMand fastigial nucleus also returned to baseline levels;however, unlike the NTS or RVLM, signal intensitythen decreased toward the end of the scanning period.
DELAYED-ONSET DECLINE. In two areas (Fig. 4), thedentate and interpositus nuclei and dorsolateral pons(including the Kolliker-Fuse nucleus), onset of the ini-tial decline was delayed, reaching a nadir by 32–40 s(cerebellar nuclei maximum decline 5 1.6 6 0.8%;dorsolateral pons maximum decline 5 3.3 6 1.4%).This decrease was maximal at approximately the timeat which respiration returned to prestimulus levels (40s after 5-HT).
Fig. 3. Signal-intensity changes in themedulla after administration of high-dose (20 or 30 mg/kg) or low-dose (10mg/kg) intravenous 5-HT, relative to theinitial 3-min baseline period in all cats.A: caudal ventrolateral medulla. B: ros-tral ventrolateral medulla. C: nucleusof the solitary tract, caudal. D: nucleusof the solitary tract, intermediate. E:caudal midline medulla. F: spinal tri-geminal nucleus. Signal changes werecombined from the left and right sides ofthe outlined areas (white outlines) andat the 1 or 2 stereotactic levels (e.g.,P13, P11) indicated on the brain outlinein E. The P level refers to distance inmm posterior to the interaural line (41).Significant signal changes, calculatedfrom 2 or 3 scans in the region of thearrows, are encoded by extent of change(percent change from baseline), pseudo-colored according to the color scale (215to 20.5% and 0.5 to 15%), and super-imposed on anatomic images. Verticaldashed lines indicate start of challenge.*Significance difference compared withbaseline, P , 0.005.
335NEURAL CHANGES TO INTRAVENOUS 5-HT
J Appl Physiol • VOL 92 • JANUARY 2002 • www.jap.org
at Penn S
tate Univ on F
ebruary 23, 2013http://jap.physiology.org/
Dow
nloaded from

LATE-ONSET SUSTAINED RESPONSE. In contrast to the ini-tial decline of several brain stem sites, more rostralbrain areas of the amygdala, regions within the hypo-thalamus and the midbrain periaqueductal gray mat-ter (PAG), showed signal intensity elevations to 5-HTchallenge (Fig. 5). Although there was a short-livedinitial rise in the amygdala and PAG (;8 s after 5-HT),the major response was a slowly rising and prolongedincrease in signal intensity (hypothalamus maximum
increase 5 2.9 6 0.4%, amygdala maximum increase 51.5 6 0.4%, PAG maximum increase 5 1.4 6 0.2%).Within the parapyramidal region, signal intensity in-creased within 16 s of 5-HT administration to 2.6 6 2%above baseline. The only other sites to show such a latesustained increase were the caudal NTS (NTS regioncaudal to the obex), spinal trigeminal nucleus and theinferior olive (caudal NTS maximum increase 5 2.8 60.9%; spinal trigeminal nucleus maximum increase 5
Fig. 4. Signal-intensity changes at 2dose levels of 5-HT, combined with sig-nal changes overlaid on comparable an-atomic images in cerebellum [fastigial(A), interpositus and dentate nuclei(B), and cerebellar vermis (C)], and cer-ebellum-related regions [inferior oli-vary nucleus (D) and dorsolateral pons(E)], and the parapyramidal region (F).Stereotactic designations and pseudo-color calculations are as for Fig. 3. Ver-tical dashed lines indicate start of chal-lenge. *Significance difference comparedwith baseline, P , 0.005.
336 NEURAL CHANGES TO INTRAVENOUS 5-HT
J Appl Physiol • VOL 92 • JANUARY 2002 • www.jap.org
at Penn S
tate Univ on F
ebruary 23, 2013http://jap.physiology.org/
Dow
nloaded from
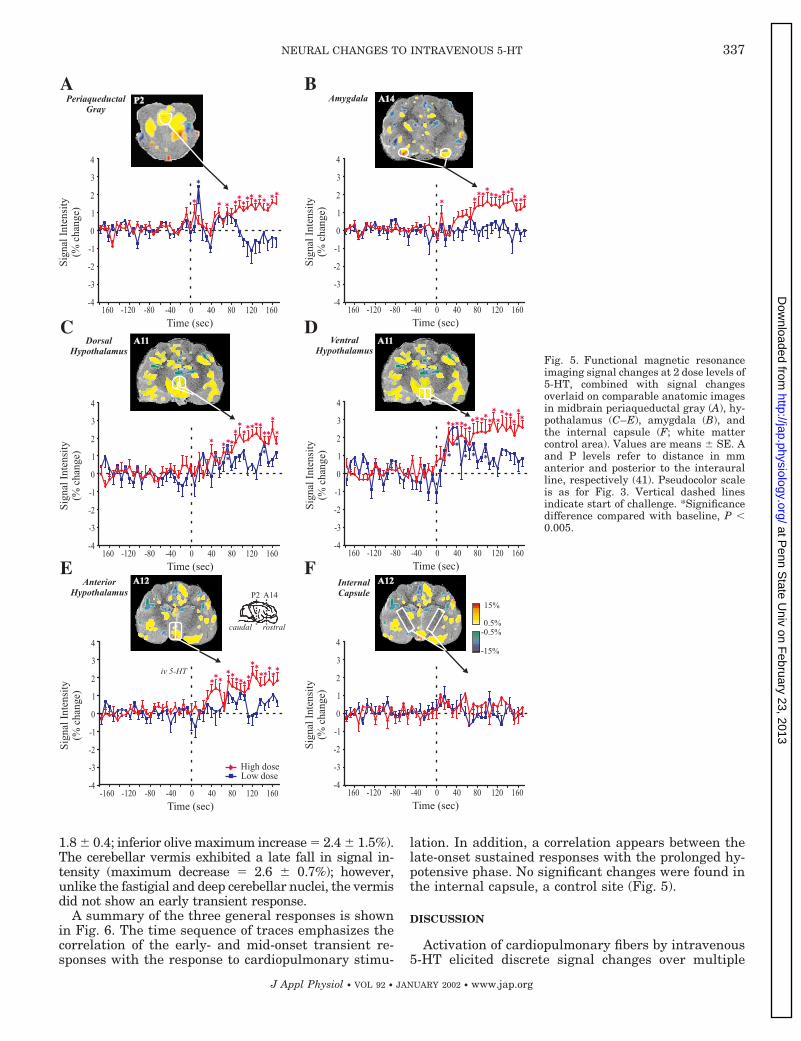
1.8 6 0.4; inferior olive maximum increase 5 2.4 6 1.5%).The cerebellar vermis exhibited a late fall in signal in-tensity (maximum decrease 5 2.6 6 0.7%); however,unlike the fastigial and deep cerebellar nuclei, the vermisdid not show an early transient response.
A summary of the three general responses is shownin Fig. 6. The time sequence of traces emphasizes thecorrelation of the early- and mid-onset transient re-sponses with the response to cardiopulmonary stimu-
lation. In addition, a correlation appears between thelate-onset sustained responses with the prolonged hy-potensive phase. No significant changes were found inthe internal capsule, a control site (Fig. 5).
DISCUSSION
Activation of cardiopulmonary fibers by intravenous5-HT elicited discrete signal changes over multiple
Fig. 5. Functional magnetic resonanceimaging signal changes at 2 dose levels of5-HT, combined with signal changesoverlaid on comparable anatomic imagesin midbrain periaqueductal gray (A), hy-pothalamus (C–E), amygdala (B), andthe internal capsule (F; white mattercontrol area). Values are means 6 SE. Aand P levels refer to distance in mmanterior and posterior to the interauralline, respectively (41). Pseudocolor scaleis as for Fig. 3. Vertical dashed linesindicate start of challenge. *Significancedifference compared with baseline, P ,0.005.
337NEURAL CHANGES TO INTRAVENOUS 5-HT
J Appl Physiol • VOL 92 • JANUARY 2002 • www.jap.org
at Penn S
tate Univ on F
ebruary 23, 2013http://jap.physiology.org/
Dow
nloaded from

brain regions. In addition to medullary regions pre-viously implicated by c-Fos or electrophysiologicalprocedures in expression of responses to cardiopul-monary stimulation, the sites included neural areasin the cerebellum and regions rostral to the medulla;the latter sites have been implicated in anatomicstudies of airway reflex and blood pressure control(12, 43). Whereas activation in medullary areas fol-lowed a sequence of an initial decline within the first
8 s and a later rise, rostral areas showed no suchphasic initial decline but only a late-emerging in-crease; these increases were long lasting (Fig. 6). Thetemporal patterning of responses in different sitessuggests specific roles for separate sites mediatingthe response. The marked apnea, hypotension, andbradycardia associated with 5-HT administration re-cruited an integrated pattern of neural responses inmultiple brain areas, some of which are not tradi-
Fig. 6. Traces of arterial pressure,heart rate (in beats/min), and respira-tory rate (in breaths/min) after high-dose 5-HT administration, togetherwith signal values from each sampledarea over the baseline and course ofthis challenge. Values are means 6 SE.Signal changes are grouped by timingof initial changes to the challenges:early, mid, or late onset. Early-onsetdeclines developed primarily frommedullary sites, as well as the cerebel-lar fastigial nucleus. Sites slightly de-layed in responses include the parapy-ramidal region, the interpositus anddentate cerebellar nuclei, and the dor-solateral pons. Late-onset response in-creases emerged principally from ros-tral brain stem and forebrain sites butalso from inferior olive, spinal trigemi-nal nucleus, and caudal nucleus of thesolitary tract. Note the late-onset de-creased response from the cerebellarvermis. Vertical dashed lines indicatestart of challenge, termination of ap-nea, and return of respiratory rate tobaseline.
338 NEURAL CHANGES TO INTRAVENOUS 5-HT
J Appl Physiol • VOL 92 • JANUARY 2002 • www.jap.org
at Penn S
tate Univ on F
ebruary 23, 2013http://jap.physiology.org/
Dow
nloaded from

tionally associated with cardiovascular and respira-tory control.
Physiology
As reported by others, intravenous 5-HT administra-tion evoked a short-latency hypotension and apnea,which, as Meller et al. (30) have shown, is elicited fromactivation of 5-HT3 receptors located on cardiopulmo-nary vagal afferents. In addition, 5-HT administrationevoked a late-onset prolonged hypotension. This laterhypotension has been presumed to result from activa-tion of 5-HT7 receptors, evoking a peripherally medi-ated vasodilation and a reflexive, baroreceptor-medi-ated increase in heart rate (39, 49). Thus 5-HTadministration evoked two distinct hypotensive reflex-es: an early-onset transient hypotension, bradycardia,and apnea and a late-developing, presumably periph-erally mediated hypotension and baroreflex-mediatedtachycardia.
Medulla
Consistent with previous c-Fos (11), electrophysiol-ogy (46), and lesion (26) evidence, 5-HT infusionevoked signal-intensity changes in several medullaryregions. Signal intensity immediately decreased withinthe intermediate NTS and CVLM. The cardiopulmo-nary afferent activity ascends through the vagus to theNTS (22, 31, 45) from which CVLM neurons receiveinput (47). It is of interest that the initial signal changeis a decline in both the NTS and CVLM; the decreasesmay reflect a combination of activation by cardiopul-monary afferent stimulation and deactivation by thebaroreceptor reflex as arterial pressure falls. Althoughboth the caudal and intermediate NTS increased signalintensity during the late hypotensive phase, only theintermediate NTS displayed an early fall. This mayreflect the disparate anatomic projections of vagal af-ferents (12, 40) and vagal C fibers, particularly thespecificity of termination of afferents within NTS re-gions at the level of, and slightly rostral to, the obex(25).
The direction of signal change in the CVLM and theinitial decrease within the RVLM support a role origi-nally hypothesized from optical imaging data obtainedfrom the ventral medullary surface (VMS) of cats andgoats. Optical studies reveal that stimulation of che-moreceptors by hypercapnia, simulated hypoxia withintravenous cyanide, or arterial pressure elevationelicits a decline in activity over regions of the VMS,with the extent of decline dependent on state (28, 36,37). Transection of the carotid sinus nerve eliminatesthe signal decline to cyanide administration, suggest-ing that the VMS receives inhibitory or dysfacilitatoryinfluences from carotid chemoreceptors. Stimulation ofC-fiber afferents by 5-HT also elicits a transient declinein activity in multiple medullary areas bordering theVMS, as well as the CMM, but not (at least at highdoses) in the NTS.
In contrast to the CVLM, CMM, and NTS, signalintensity in the parapyramidal region increased signif-
icantly, but it did so later in the cardiopulmonaryphase. Retrograde tracing (both viral transneuronaland nonviral) indicates that the parapyramidal regioncontains parasympathetic neurons innervating boththe phrenic motor nucleus and the peripheral airways(12, 16). Furthermore, the region responds to respira-tory stimuli such as hypercapnia, hypoxia, and anoxia(4, 6). The slightly delayed-onset and maintained na-ture of the increase in parapyramidal signal intensitysuggest a role in both the cessation of the apnea andcompensatory increase in respiratory drive during thelate-onset baroreflex phase. The region of the VMS,immediately lateral to the parapyramidal region, in-creases activity, as evaluated by optical procedures, tohypotension elicited by sodium nitroprusside adminis-tration or hypovolemia (14, 36).
The change in signal intensity in the CMM wasparticularly remarkable, because of its immediate on-set, profound extent of fall, and transient nature, withthe signal returning to baseline within 24 s. Unlikeother regions, such as NTS, RVLM, cerebellum, amyg-dala, or dorsal lateral pontine sites, only the initialphasic decline was apparent. The duration of signal-intensity decrease in CMM was concurrent with theinitial physiological responses, but the rapid return tobaseline did not parallel the long-lasting hypotension.The transient nature of the response suggests a signal-ing function, rather than a role in long-term mainte-nance of arterial pressure or respiration. The primaryrole of the CMM in regulation of breathing or arterialpressure control remains unclear. Although chemicalactivation of the CMM results in hypotension andbradycardia (15, 18), electrolytic lesions and chemicalinactivation of this region have no effect on restingarterial pressure or heart rate or on the arterial pres-sure or heart rate responses to activation of eithercardiopulmonary afferents or baroreceptor reflexes (19,26). However, Vayssettes-Courchay and colleagues (46)reported that single neurons within the CMM increaseor decrease firing rate after activation of cardiopulmo-nary afferents, suggesting that the structure plays arole in the apneic, rather than vasodepressor, compo-nents of the reflex. Chemical activation of the CMMalters respiratory patterning and enhances phrenicnerve discharge (3, 15), and, most strikingly, ibotenicacid lesions of the CMM results in respiratory failure(7), whereas comparable lesions in well-described re-spiratory regions (NTS, area postrema, lateral andventrolateral medulla, including the nucleus am-biguus) fail to elicit such a breathing cessation. Thesignal-intensity decrease in CMM may indicate anactivity decline that plays a role in the profound apneaafter 5-HT infusion.
Cerebellum and Related Structures
The signal changes found in the inferior olive (aprincipal afferent source to cerebellar Purkinje cells),the fastigial, dentate, and interpositius cerebellar nu-clei, and the vermis reinforce the significant role thatcerebellar-related structures play in mediating breath-
339NEURAL CHANGES TO INTRAVENOUS 5-HT
J Appl Physiol • VOL 92 • JANUARY 2002 • www.jap.org
at Penn S
tate Univ on F
ebruary 23, 2013http://jap.physiology.org/
Dow
nloaded from

ing and arterial pressure challenges, at least for thoseof an extreme nature (27, 51). Signal changes in theinferior olive, and also the fastigial, dentate, and inter-positus cerebellar nuclei reached their maximal valuesat approximately the termination of the initial arterialpressure fall and apnea from 5-HT stimulation (Fig. 6),suggesting participation with correction of the apnea(51) and/or the hypotension and bradycardia (27). To-gether with the inferior olive and fastigial nucleus, thevermis showed late-developing signal changes, possi-bly representing a role for error correcting the long-lasting hypotension accompanying the challenge (27).Cardiovascular and respiratory expression of fastigialaction could be mediated through the dorsolateralpons, which receives significant projections from thefastigial nucleus (43). Dorsal lateral pontine areasshowed both a delayed transient response and a laterresponse. The delayed response to the initial 5-HTcomponent may reflect action by this phase-switchingsite (5) to restore inspiration, whereas the late compo-nent may be acting on the late-phase hypotension.
Rostral Sites
In contrast to the initial signal declines in the brainstem, rostral brain areas showed only increased signalintensity after intravenous 5-HT. Apart from an initial,transient signal increase in the PAG and amygdala,the rise in signal intensity emerged late in the re-sponse for all rostral areas. This later activation likelyrepresents action to restore breathing after the apneafrom the 5-HT administration and, perhaps via thebaroreceptor reflex, to increase heart rate.
Studies using c-Fos procedures indicate that neuronsin the ventrolateral column of the PAG are selectivelyactivated by intravenous 5-HT (24). Furthermore,stimulation of the lateral and ventrolateral regions ofthe PAG has been found to modify baroreflex sensitiv-ity in anesthetized rats (21, 32). Thus the late sus-tained increase in PAG activity reported here mayrepresent actions to alter baroreflex sensitivity and, atthe same time, trigger late-phase cardiac responses.
Participation of both amygdala and hypothalamicsites as components of a neural network controlling theairway has been demonstrated anatomically (12). Hy-pothalamic sites all increased signal intensity to thechallenge, and these responses were late and sus-tained. Expression of c-Fos within the dorsal hypothal-amus (including paraventricular nucleus) and ventralhypothalamus (including the arcuate nucleus) in-creases after baroreceptor unloading (35). Further-more, during baroreceptor unloading, c-Fos is ex-pressed in norepinephrine-containing CVLM neuronsthat project directly to the paraventricular nucleus andare critical in mediating the vasopressin release duringbaroreceptor unloading (10). The late-onset increase indorsal hypothalamic signal intensity reported heremay reflect increased neural activity, possibly result-ing in increased vasopressin release.
The significant projections from the amygdala cen-tral nucleus to the NTS and parabrachial pons, as well
as areas surrounding the nucleus ambiguus (20), pro-vide considerable potential for this limbic structure tomodulate concurrent responses in brain stem sites to5-HT stimulation. The principal influences from theamygdala appear to arise later in the challenge (al-though a short-lasting initial activation was present).The late-developing response may influence compensa-tory responses for recovery from the late-developinghypotension.
Relationship to Anatomic Descriptions
The sites demonstrating signal changes to 5-HT ad-ministration closely overlap with neural regions iden-tified by transneuronal viral and nonviral retrogradelabeling as innervating airway structures and thephrenic motor nucleus (12, 16). Regions displayingearly-onset (CMM, CVLM, VMS, intermediate NTS) orlate-onset sustained (parapyramidal region, dorsolat-eral pons, amygdala, hypothalamus, PAG) changes insignal intensity contain neurons innervating both thephrenic motor nucleus and airway structures. Neuronssupplying the peripheral airways are also identifiedwithin the lateral paragigantocellular nucleus, an areain which we also observed signal changes with cardio-pulmonary activation and that aspect will be closelyexamined in future studies. Labeling within cerebellarsites was not examined in the anatomic studies.
Limitations
Although we used a relatively fast acquisition proce-dure, image collection was slower than signal changesobserved on ventral medullary surface areas with op-tical procedures (14). However, the focus of this studywas to survey brain areas involved in stimulation ofcardiopulmonary afferents and to provide a summaryview of the temporal patterning of those responses.More rapid image acquisition can be obtained withhigher field scanners or by using the current instru-mentation with fewer slices, i.e., by limiting scans tobrain areas shown by this initial study to be recruitedduring the challenge.
Cerebral perfusion is autoregulated so that littlechange in flow occurs between arterial pressures rang-ing from 50 to 150 mmHg (2). After high-dose admin-istration of 5-HT, arterial pressure fell from 120 to 95mmHg, well within the autoregulatory range. Mag-netic resonance studies directly addressing this con-cern show that, between 120 and 60 mmHg, there is nosignificant change in either blood volume or blood flowwithin the striatum, cortex, or the brain at the level ofbregma (52). Furthermore, we found that no regiondisplayed signal changes paralleling the time course ofarterial pressure changes and that opposite changes insignal intensity emerged in regions immediately bor-dering one another, e.g., the caudal ventrolateral me-dulla and the inferior olivary nucleus, an unlikelyevent if signal changes resulted from overall perfusionalterations. The changes we report here most likelyresulted from local blood flow changes in response tochanges in neural activity and were not due to overall
340 NEURAL CHANGES TO INTRAVENOUS 5-HT
J Appl Physiol • VOL 92 • JANUARY 2002 • www.jap.org
at Penn S
tate Univ on F
ebruary 23, 2013http://jap.physiology.org/
Dow
nloaded from

changes in cerebral perfusion following changes inarterial pressure.
The initial apnea induced by 5-HT could modifyarterial PO2 and end-tidal PCO2, with both modifica-tions having the potential to alter activity in medullaryand cerebellar sites. Because onset of the neural sig-nal changes was coincident with apnea onset, andchanges in arterial PO2 and end-tidal PCO2 require afinite time to develop, it appears unlikely that theinitial, transient changes in neural activity resultedfrom O2 and CO2 alterations. The late-developing char-acteristics, however, could well interact with blood-gaslevels. Partitioning of those late changes from contri-butions by gas levels, blood pressure changes, andother sources will be a necessary effort for future eval-uation after this initial overview description.
The mode of delivery of 5-HT, via an intravenousroute, has the potential to stimulate sensory afferentsother than C-fiber cardiopulmonary sensors located inthe atrium or thorax. Actions of 5-HT on the airway,such as bronchoconstriction and decreased lung com-pliance, can affect sensory activity other than directC-fiber stimulation. Some of the issues, particularly oftiming and localization of stimulation, could have beenmore readily accommodated with administration ofphenyldiguanide, a 5-HT3 agonist, rather than 5-HT,and direct administration of the agent onto primaryreceptors of interest, e.g., the right atrium.
We thank Drs. A. Gmitro and J. Alger for assistance with MRscanning and Dr. P. Macey for assistance with analysis.
This research was supported by National Heart, Lung, and BloodInstitute Grant HL-22418; the Flinn Foundation (Phoenix, AZ); andNational Institutes of Health/National Cancer Institute Grant CA-83148. Pearl L. Yu is a National Institute of Child Health andHuman Development Fellow of the Pediatric Scientist DevelopmentProgram (Grant K12-HD-00850).
REFERENCES
1. Aviado DM and Guevara Aviado G. The Bezold-Jarisch re-flex. A historical perspective of cardiopulmonary reflexes. AnnNY Acad Sci 940: 48–58, 2001.
2. Bates D and Sundt TM. The relevance of peripheral barore-ceptors and chemoreceptors to regulation of cerebral blood flowin the cat. Circ Res 38: 488–493, 1976.
3. Bernard DG. Cardiorespiratory responses to glutamate micro-injected into the medullary raphe. Respir Physiol 113: 11–21,1998.
4. Berquin P, Bodineau L, Gros F, and Larnicol N. Brainstemand hypothalamic areas involved in respiratory chemoreflexes: aFos study in adult rats. Brain Res 857: 30–40, 2000.
5. Bertrand F and Hugelin A. Respiratory synchronizing func-tion of nucleus parabrachialis medialis: pneumotaxic mecha-nisms. J Neurophysiol 34: 189–207, 1971.
6. Bodineau L, Cayetanot F, and Frugiere A. Possible role ofretrotrapezoid nucleus and parapyramidal area in the respira-tory response to anoxia: an in vitro study in neonatal rat. Neu-rosci Lett 295: 67–69, 2000.
7. Carruth MK, Fowler AA, Fairman RP, Mayer DJ, andLeichnetz GR. Respiratory failure without pulmonary edemafollowing injection of a glutamate agonist into the ventral med-ullary raphe of the rat. Brain Res Bull 28: 365–378, 1992.
8. Edelman RR, Weilopolski P, and Schmitt F. Echo planar MRimaging. Radiology 192: 600–612, 1994.
9. Fullerton DA, St Cyr JA, Clarke DR, Campbell DN, ToewsWH, and See WM. Bezold-Jarisch reflex in postoperative pedi-atric cardiac surgical patients. Ann Thorac Surg 52: 534–536,1991.
10. Gardiner SM and Bennett T. Endogenous vasopressin andbaroreflex mechanisms. Brain Res 396: 317–341, 1986.
11. Gieroba ZJ, MacKenzie L, Willoughby JO, and BlessingWW. Fos-determined distribution of neurons activated duringthe Bezold-Jarisch reflex in the medulla oblongata in consciousrabbits and rats. Brain Res 683: 43–50, 1995.
12. Hadziefendic S and Haxhiu MA. CNS innervation of vagalpreganglionic neurons controlling peripheral airways: a trans-neuronal labeling study using pseudorabies virus. J Auton NervSyst 76: 135–145, 1999.
13. Harper AM and MacKenzie ET. Cerebral circulatory andmetabolic effects of 5-hydroxytryptamine in anaesthetised ba-boons. J Physiol (Lond) 271: 721–733, 1977.
14. Harper RM, Richard CA, and Rector DM. Physiological andventral medullary surface activity during hypovolemia. Neuro-science 94: 579–586, 1999.
15. Haxhiu MA, Erokwu B, Bhardwaj V, and Dreshaj IA. Therole of the medullary raphe nuclei in regulation of cholinergicoutflow to the airways. J Auton Nerv Syst 69: 64–71, 1998.
16. Haxhiu MA, Erokwu BO, and Cherniack NS. The brainstemnetwork involved in coordination of inspiratory activity andcholinergic outflow to the airways. J Auton Nerv Syst 61: 155–161, 1996.
17. Henderson LA, Frysinger RC, Yu PL, Bandler R, andHarper RM. A device for feline positioning and stabilizationduring magnetic resonance imaging. Magnetic Resonance Imag-ing 19: 1031–1036, 2001.
18. Henderson LA, Keay KA, and Bandler R. The ventrolateralperiaqueductal gray projects to caudal brainstem depressor re-gions: a functional-anatomical and physiological study. Neuro-science 82: 201–221, 1998.
19. Henderson LA, Keay KA, and Bandler R. Caudal midlinemedulla mediates behaviourally-coupled but not baroreceptor-mediated vasodepression. Neuroscience 98: 779–792, 2000.
20. Hopkins DA and Holstege G. Amygdaloid projections to themesencephalon, pons and medulla oblongata in the cat. ExpBrain Res 32: 529–547, 1978.
21. Inui K, Murase S, and Nosaka S. Facilitation of the arterialbaroreflex by the ventrolateral part of the midbrain periaque-ductal grey matter in rats. J Physiol (Lond) 477: 89–101, 1994.
22. Jacobs L and Camroe JH Jr. Reflex apnea, bradycardia, andhypotension produced by serotonin and phenyldiguanide actingon the nodose ganglia of the cat. Circ Res 29: 145–155, 1971.
23. Jarisch A and Zotterman Y. Depressor reflex from the heart.Acta Physiol Scand 16: 31–351, 1948.
24. Keay KA, Clement C, and Bandler R. The neuroanatomy ofcardiac nociceptive pathways: differential representation of‘deep’ and ‘superficial’ pain. In: The Nervous System and theHeart. Totowa, NJ: Humana, 2000, p. 303–342.
25. Kubin L, Kimura H, and Davis RO. The medullary projec-tions of afferent bronchopulmonary C fibres in the cat as shownby antidromic mapping. J Physiol (Lond) 435: 207–228, 1991.
26. Lee TM, Kuo JS, and Chai CY. Central integrating mecha-nism of the Bezold-Jarisch and baroceptor reflexes. Am J Physiol222: 713–720, 1972.
27. Lutherer LO, Lutherer BC, Dormer KJ, Janssen HF, andBarnes CD. Bilateral lesions of the fastigial nucleus prevent therecovery of blood pressure following hypotension induced byhemorrhage or administration of endotoxin. Brain Res 269: 251–257, 1983.
28. Macey PM, Richard CA, Rector DM, Harper RK, andHarper RM. State influences on ventral medullary surface andphysiological responses to sodium cyanide challenges. J ApplPhysiol 89: 1919–1927, 2000.
29. Mark AL. The Bezold-Jarisch reflex revisited: clinical implica-tions of inhibitory reflexes originating in the heart. J Am CollCardiol 1: 90–102, 1983.
30. Meller ST, Lewis SJ, Brody MJ, and Gebhart GF. Vagalafferent-mediated inhibition of a nociceptive reflex by intrave-nous serotonin in the rat. II. Role of 5-HT receptor subtypes.Brain Res 585: 71–86, 1992.
31. Meller ST, Lewis SJ, Ness TJ, Brody MJ, and Gebhart GF.Vagal afferent-mediated inhibition of a nociceptive reflex by
341NEURAL CHANGES TO INTRAVENOUS 5-HT
J Appl Physiol • VOL 92 • JANUARY 2002 • www.jap.org
at Penn S
tate Univ on F
ebruary 23, 2013http://jap.physiology.org/
Dow
nloaded from

intravenous serotonin in the rat. I. Characterization. Brain Res524: 90–100, 1990.
32. Nosaka S, Murata K, Inui K, and Murase S. Arterial barore-flex inhibition by midbrain periaqueductal grey in anaesthetizedrats. Pflugers Arch 424: 266–275, 1993.
33. Ogawa S, Lee TM, Nayak AS, and Glynn P. Oxygenation-sensitive sensory contrast in magnetic resonance image of rodentbrain at high magnetic fields. Magn Reson Med 14: 68–78, 1990.
34. Parker JM, Alger JR, Woo MA, Spriggs D, and Harper RM.Acquisition of electrophysiologic signals during magnetic reso-nance imaging. Sleep 22: 1125–1126, 1999.
35. Potts PD, Polson JW, Hirooka Y, and Dampney RA. Effectsof sinoaortic denervation on Fos expression in the brain evokedby hypertension and hypotension in conscious rabbits. Neuro-science 77: 503–520, 1997.
36. Rector DM, Richard CA, Staba RJ, and Harper RM. Sleepstates alter ventral medullary surface responses to blood pres-sure challenges. Am J Physiol Regulatory Integrative CompPhysiol 278: R1090–R1098, 2000.
37. Richard CA, Rector D, Macey P, and Harper RM. Changesin ventral medullary light reflectance during hypercapnia inawake and sleeping cats. Neurosci Lett 286: 175–178, 2000.
38. Robertson RM and Robertson D. The Bezold-Jarisch reflex:possible role in limiting myocardial ischemia. Clin Cardiol 4:75–79, 1981.
39. Saxena PR and Villalon CM. Cardiovascular effects of seroto-nin agonists and antagonists. J Cardiovasc Pharmacol 15: 17–34, 1990.
40. Silva-Carvalho L, Paton JFR, Rocha I, Goldsmith GE, andSpyer KM. Convergence properties of solitary tract neuronsresponsive to cardiac receptor stimulation in the anesthetizedcat. J Neurophysiol 79: 2374–2382, 1998.
41. Snider RS and Niemer WT. A Stereotaxic Atlas of the CatBrain. Chicago, IL: Univ. of Chicago Press, 1961.
42. Somers VK and Abboud FM. Neurocardiogenic syncope. AdvIntern Med 41: 399–435, 1996.
43. Supple WF Jr and Kapp BS. Anatomical and physiologicalrelationships between the anterior cerebellar vermis and the
pontine parabrachial nucleus in the rabbit. Brain Res Bull 33:561–574, 1994.
44. Takahashi R, Sakai T, Furuyama Y, Kondo Y, Inoue CI,Onuma S, and Iinuma K. The vasocontractive action of nor-epinephrine and serotonin in deep arterioles of rat cerebral graymatter. Tohoku J Exp Med 190: 129–142, 2000.
45. Thurston-Stanfield CL, Ranieri JT, Vallabhapurapu R,and Barnes-Nable B. Role of vagal afferents and the rostralventral medulla in intravenous serotonin-induced changes innociception and arterial blood pressure. Physiol Behav 67: 753–767, 1999.
46. Vayssettes-Courchay C, Bouysset F, Laubie M, and Ver-beuren TJ. Central integration of the Bezold-Jarish reflex inthe cat. Brain Res 744: 272–278, 1997.
47. Verberne AJ, Beart PM, and Louis WJ. Excitatory aminoacid receptors in the caudal ventrolateral medulla mediate avagal cardiopulmonary reflex in the rat. Exp Brain Res 78:185–192, 1989.
48. Verberne AJ and Guyenet PG. Medullary pathway of theBezold-Jarisch reflex in the rat. Am J Physiol Regulatory Inte-grative Comp Physiol 263: R1195–R1202, 1992.
49. Villalon CM, Centurion D, Bravo G, De Vries P, SaxenaPR, and Ortiz MI. Further pharmacological analysis of theorphan 5-HT receptors mediating feline vasodepressor respon-ses: close resemblance to the 5-HT7 receptor. Naunyn Schmiede-bergs Arch Pharmacol 316: 665–671, 2000.
50. Whalen EJ, Johnson AK, and Lewis SJ. Functional evidencefor the rapid desensitization of 5-HT(3) receptors on vagal affer-ents mediating the Bezold-Jarisch reflex. Brain Res 873: 302–305, 2000.
51. Williams JL, Everse SJ, and Lutherer LO. Stimulating fas-tigial nucleus alters central mechanisms regulating phrenic ac-tivity. Respir Physiol 76: 215–227, 1989.
52. Zaharchuk G, Mandeville JB, Bogdanov AA Jr, WeisslederR, Rosen BR, and Marota JJ. Cerebrovascular dynamics ofautoregulation and hypoperfusion. An MRI study of CBF andchanges in total and microvascular cerebral blood volume duringhemorrhagic hypotension. Stroke 30: 2197–2204, 1999.
342 NEURAL CHANGES TO INTRAVENOUS 5-HT
J Appl Physiol • VOL 92 • JANUARY 2002 • www.jap.org
at Penn S
tate Univ on F
ebruary 23, 2013http://jap.physiology.org/
Dow
nloaded from



















![Analyzing neural responses to natural signals: Maximally ... · arXiv:physics/0212110v2 [physics.bio-ph] 19 Sep 2003 Analyzing neural responses to natural signals: Maximally informative](https://img.pdfslide.us/doc/110x75/5fa7b78d543d566cd753b2be/analyzing-neural-responses-to-natural-signals-maximally-arxivphysics0212110v2.jpg)









