Embed Size (px)
Citation preview

J. Pathol. 187: 530–534 (1999)
FUNCTIONAL LOSS OF E-CADHERIN ANDCADHERIN-11 ALLELES ON CHROMOSOME 16q22 IN
COLONIC CANCER
1, 2, 1, 2, - 3, ̈1,3 . 1*
1Institut für Pathologie, GSF-Forschungszentrum für Umwelt und Gesundheit, Neuherberg, Germany2Chirurgische Klinik der Technischen Universität München, München, Germany
3Institut für Allgemeine Pathologie und Pathologische Anatomie der Technischen Universität München, München, Germany
SUMMARY
Proteins of the cadherin family regulate cellular adhesion and motility and are believed to act as tumour suppressors. Previous studieshave identified frequent mutation and allelic inactivation of the E-cadherin (cadherin-1) locus in diffuse gastric cancer. At least two othercadherin genes, P-cadherin (cadherin-3) and OB-cadherin (cadherin-11), have been mapped close to the E-cadherin gene on chromosome16q22. As this region of the genome is frequently deleted in malignancy, multiple cadherin loci may be affected by losses of chromosome16q22. The expression of mRNA transcripts from polymorphic alleles of the E-cadherin and cadherin-11 genes was examined in 30 casesof colonic, gastric, and renal carcinoma. In gastric cancer, loss of expression of one allele was restricted to the E-cadherin locus, whilstin renal carcinoma neither locus was affected. In colonic cancers, loss of expression of one E-cadherin allele was detected in 5 of 22 cases,whilst loss of a cadherin-11 allele was seen in 5 of 23 cases. This functional loss of cadherin gene expression may be due to gene deletion,inactivation or recombination. As no evidence of cadherin gene mutation was observed in the remaining transcripts, we can conclude thatthese two genes are only indirectly involved in the pathogenesis of colorectal cancer. Copyright ? 1999 John Wiley & Sons, Ltd.
KEY WORDS—cadherins; functional allelic loss; colorectal cancer; chromosome 16q22
INTRODUCTION
The classical cadherins are a family of transmembraneglycoproteins mediating cell adhesion and intercellularsignalling. The extracellular domains engage in calcium-dependent homotypic binding, whilst the cytosolicdomains interact with cytoskeletal elements and theWnt/â-catenin (wingless/armadillo) signalling path-way.1,2 During embryogenesis, both the spatial–temporal distribution and the type of cadherinexpression are closely regulated, allowing the aggrega-tion, disaggregation, and migration of cell populationsthat is essential for morphogenesis.2,3 In adult tissues,expression of multiple cadherins maintains tissue integ-rity and allows complex interactions between differentcell populations.3
Inappropriate loss of cadherin expression can havedramatic consequences in both embryonic and differen-tiated tissues. Targeted deletion of the E-cadherin(cadherin-1)4 or N-cadherin (cadherin-2)5 genes resultsin early embryonic death, due to failure correctly toorganize embryonic tissues. Down-regulation and mu-tation of E-cadherin are associated with epithelialcarcinogenesis in diffuse stomach cancer,6 ovarian,7 andmammary carcinoma.8 Similar carcinogenic phenotypesare associated with mutation or abnormal expression ofother components of the Wnt/catenin signalling path-way, including Wnt 2 and Wnt 5A,9,10 â-catenin,11,12
and APC.13
Germany.
CCC 0022–3417/99/050530–05$17.50Copyright ? 1999 John Wiley & Sons, Ltd.
Using intragenic E-cadherin polymorphisms, we wereable to identify loss of expression of one allele in 42 percent of gastric carcinomas,14 consistent with a tumour/metastasis-suppressor role for the adhesion molecule.However, whilst mutation of E-cadherin was observedexclusively in diffuse-type gastric carcinomas, allelic losswas equally frequent in diffuse and intestinal-typetumours. This observation suggests that the affectedregion may extend beyond the E-cadherin locus toinclude other tumour-suppressor genes. Deletion ofchromosome 16q22 sequences is not restricted to gastriccancer and has been additionally reported in hepato-cellular,15 prostate,16 endometrial,17 and breast cancer.18
At least three cadherin genes (E-cadherin, P-cadherin,and cadherin-11) map to chromosome 16q22.19 (Notethat the anonymous human EST WI-2756, which mapsto the chromosme 16q22 YAC contig WC 16-5, is in factderived from the cadherin-11 sequence.) Consequently,this cluster of cadherin genes is a strong candidate to beaffected by allelic loss of chromosome 16q22. In thisinitial characterization, we present evidence establishingallelic inactivation affecting the E-cadherin andcadherin-11 loci in colonic cancer.
MATERIALS AND METHODS
*Correspondence to: Michael J. Atkinson, GSF-Institut fürPathologie, Ingolstaedter Landstrasse 1, D85764 Neuherberg,
Patient material
Thirty cases each of gastric, renal, and coloniccancer were obtained from the tumour/tissue bank ofthe Institutes of Pathology of the GSF-National
Research Centre and the Technical University ofReceived 4 March 1998Revised 3 July 1998
Accepted 27 November 1998

531FUNCTIONAL ALLELIC LOSS OF CADHERIN LOCI IN COLONIC CANCER
Munich. Non-tumour tissue samples were excisedmacroscopically from areas adjacent to tumour tissueand both tumour and non-tumour samples were snap-frozen in liquid nitrogen.
Histological analysis was performed to confirm theidentity of the samples before use.
extraction.
Molecular analysis of allelic status of cadherin geneexpression
Nucleic acids were extracted from frozen tissues usingguanidinium thiocyanate and phenol.20 Total RNA(1 ìg) was subjected to reverse-transcription; 1/25th ofthe reaction product was subsequently amplified bypolymerase chain reaction (RT-PCR), and the PCRproduct directly sequenced. Conditions and primers forthe analysis of polymorphic nucleotides within the cod-ing region of the E-cadherin gene were as previouslydescribed.14 For the identification of cadherin-11sequence polymorphisms, the PCR was performed togenerate four overlapping cDNA fragments, coveringthe entire coding region. Forty cycles of PCR wereperformed in a total volume of 50 ìl using 2·5 U ofTaq-polymerase (Eurobio, Raunheim, Germany), withdenaturing, annealing, and elongation times and tem-peratures of 1·5 min at 94)C, 1·5 min at 55)C, and1·5 min at 72)C. The following PCR primers were used:
Fragment 1 (nt 95 to nt 838)CDH11.1 5*-ACCGTACAGTTGGTGGAAGG-3*CDH11.2 5*-GCCTCCCTGTCCATGTTGGGTAG-3*
Fragment 2 (nt 781 to nt 1433)CDH11.3 5*-CGGTGGAAGCACAGACAGGT-3*CDH11.4 5*-GTCAGTGTGACGATCGATGG-3*
Fragment 3 (nt 1364 to nt 1893)CDH11.5 5*-GAGTGCATGCCAAAGACCCTG
ATG-3*CDH11.6 5*-GGATGCCGCCATCGCTGATCAC-3*
Fragment 4 (nt 1830 to nt 2568)CDH11.7 5*-GGGTCCAGTCGGCAGAAGCAG
GAC-3*CDH11.8 5*-GGCCAAATTTGTATCGTTATTG-3*
Reaction products were purified (Qiaquick affinitycolumns, Qiagen, Hilden, Germany), precipitated, anddirectly sequenced in both directions (Sequenase 2.0, U.S.Biochemicals, Braunschweig, Germany). Details of thesequencing primers used can be obtained from theauthors by e-mail. Polymorphisms within the open read-ing frame of the cadherin-11 gene were identified bycomparing the sequences obtained from non-tumourtissues of ten randomly selected study subjects. Thisallows the detection of polymorphic bases occurring at afrequency greater than 1 in 20 within the population. Theactual prevalences of the polymorphisms were subse-quently deduced from the sequences of all 90 non-tumourDNA samples included in the study. The allelic status ofthe polymorphic sites in tumour tissues was determinedby RT-PCR and sequencing of the appropriate region in
Copyright ? 1999 John Wiley & Sons, Ltd.
informative individuals. When functional allelic loss wasobserved, the entire cadherin-11 coding region wassequenced.
Cell culture
The human colonic carcinoma cell line SW480 (ATCCCCL 228), the E-cadherin and cadherin-11-negativehuman breast carcinoma MDA-MB-435S (ATCC HTB129), and the cadherin-11-expressing human rhabdo-myosarcoma cell line RD (ATCC CCL 136) werecultured in Dulbecco’s MEM containing 20 per centFCS. Total RNA was harvested from near-confluentcells using guanidinium thiocyanate lysis and phenol
20
Northern blot analysis
Ten micrograms of total cellular RNA per lane wasseparated on a 1 per cent agarose/formaldehyde gel,transferred onto nitrocellulose membrane (BoehringerMannheim, Mannheim, Germany), and hybridized at68)C with a digoxigenin-labelled cRNA probe (RNAlabelling kit, Boehringer Mannheim). The cadherin-11probe was generated by transcription of a linearizedpCRII vector (Invitrogen) containing a 1338 nt RT-PCRfragment generated using primers CDH11.1 andCDH11.4 (see above). Detection of hybridized probewas performed using the RNA–DIG detection kit(Boehringer Mannheim).
RESULTS
Cadherin-11 expression in the colonic carcinoma cell lineSW480
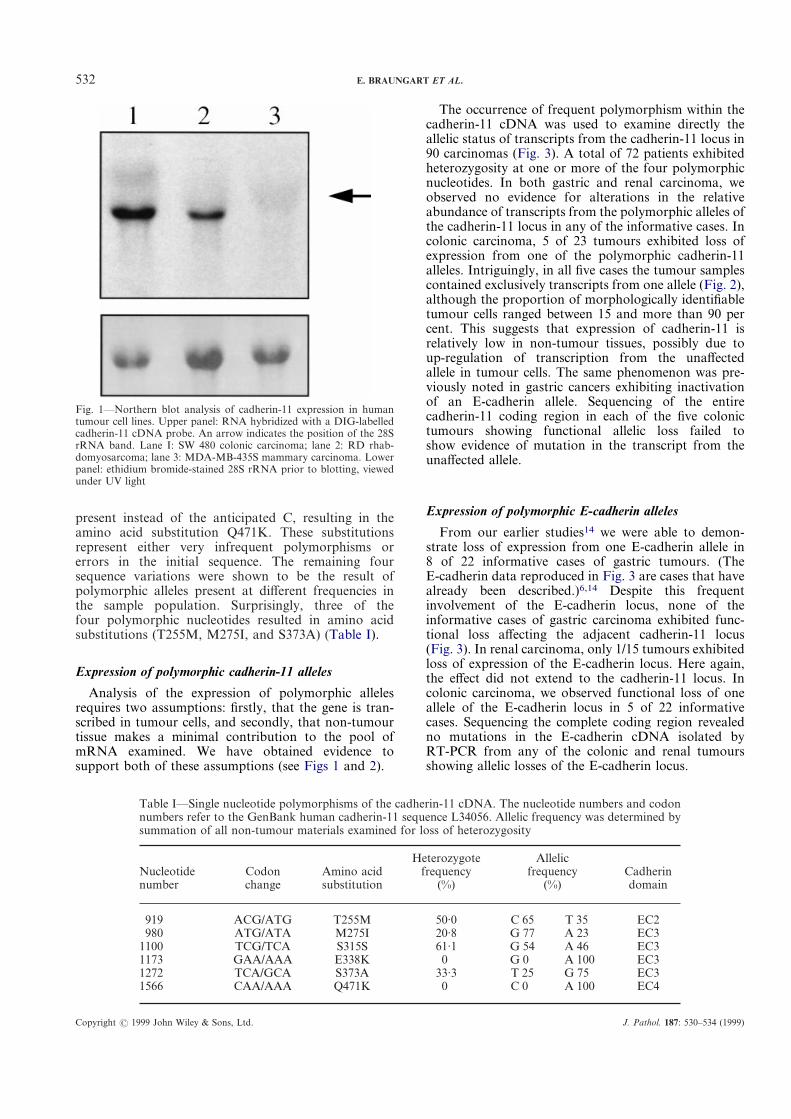
The use of the RT-PCR assay to examine directly theallelic status of cadherin genes demands that the gene betranscribed in the tumour cells under investigation.Cadherin-11 transcripts have previously been detected inboth renal and gastric tumour cell lines.21,22 To confirmthat cadherin-11 is indeed transcribed in colonic cancercells, we conducted a northern blot analysis using RNAextracted from the SW480 colonic carcinoma cellline. Figure 1 illustrates the positive expression ofcadherin-11 in both the human SW480 colonic cancerand RD rhabdomyosarcoma cell lines. In MDA-MB-435S human mammary cancer cells, no hybridizationsignal was observed.
Identification of intragenic cadherin-11 polymorphisms
Overlapping RT-PCR fragments, spanning the com-plete cadherin-11 open reading frame, were generatedand sequenced in both directions. A total of six nucleo-tide differences to the published human cadherin-11cDNA sequence (GenBank L34056) were identified.Two of these sequence variations were present in allsamples of non-tumour tissue analysed. At nucleotide1173, a G replacement of A resulted in the amino acidchange E340K, whilst at nucleotide 1566 an A was
J. Pathol. 187: 530–534 (1999)
532 E. BRAUNGART ET AL.
present instead of the anticipated C, resulting in theamino acid substitution Q471K. These substitutionsrepresent either very infrequent polymorphisms orerrors in the initial sequence. The remaining foursequence variations were shown to be the result ofpolymorphic alleles present at different frequencies inthe sample population. Surprisingly, three of thefour polymorphic nucleotides resulted in amino acidsubstitutions (T255M, M275I, and S373A) (Table I).
Expression of polymorphic cadherin-11 alleles
Analysis of the expression of polymorphic allelesrequires two assumptions: firstly, that the gene is tran-scribed in tumour cells, and secondly, that non-tumourtissue makes a minimal contribution to the pool ofmRNA examined. We have obtained evidence tosupport both of these assumptions (see Figs 1 and 2).
Copyright ? 1999 John Wiley & Sons, Ltd.
The occurrence of frequent polymorphism within thecadherin-11 cDNA was used to examine directly theallelic status of transcripts from the cadherin-11 locus in90 carcinomas (Fig. 3). A total of 72 patients exhibitedheterozygosity at one or more of the four polymorphicnucleotides. In both gastric and renal carcinoma, weobserved no evidence for alterations in the relativeabundance of transcripts from the polymorphic alleles ofthe cadherin-11 locus in any of the informative cases. Incolonic carcinoma, 5 of 23 tumours exhibited loss ofexpression from one of the polymorphic cadherin-11alleles. Intriguingly, in all five cases the tumour samplescontained exclusively transcripts from one allele (Fig. 2),although the proportion of morphologically identifiabletumour cells ranged between 15 and more than 90 percent. This suggests that expression of cadherin-11 isrelatively low in non-tumour tissues, possibly due toup-regulation of transcription from the unaffectedallele in tumour cells. The same phenomenon was pre-viously noted in gastric cancers exhibiting inactivationof an E-cadherin allele. Sequencing of the entirecadherin-11 coding region in each of the five colonictumours showing functional allelic loss failed toshow evidence of mutation in the transcript from theunaffected allele.
Fig. 1—Northern blot analysis of cadherin-11 expression in humantumour cell lines. Upper panel: RNA hybridized with a DIG-labelledcadherin-11 cDNA probe. An arrow indicates the position of the 28SrRNA band. Lane I: SW 480 colonic carcinoma; lane 2: RD rhab-domyosarcoma; lane 3: MDA-MB-435S mammary carcinoma. Lowerpanel: ethidium bromide-stained 28S rRNA prior to blotting, viewedunder UV light
Table I—Single nucleotide polymorphisms of the cadherin-11 cDNA. The nucleotide numbers and codonnumbers refer to the GenBank human cadherin-11 sequence L34056. Allelic frequency was determined bysummation of all non-tumour materials examined for loss of heterozygosity
Nucleotidenumber
Codonchange
Amino acidsubstitution
Heterozygotefrequency
(%)
Allelicfrequency
(%)Cadherindomain
919 ACG/ATG T255M 50·0 C 65 T 35 EC2980 ATG/ATA M275I 20·8 G 77 A 23 EC3
1100 TCG/TCA S315S 61·1 G 54 A 46 EC31173 GAA/AAA E338K 0 G 0 A 100 EC31272 TCA/GCA S373A 33·3 T 25 G 75 EC31566 CAA/AAA Q471K 0 C 0 A 100 EC4
Expression of polymorphic E-cadherin alleles
From our earlier studies14 we were able to demon-strate loss of expression from one E-cadherin allele in8 of 22 informative cases of gastric tumours. (TheE-cadherin data reproduced in Fig. 3 are cases that havealready been described.)6,14 Despite this frequentinvolvement of the E-cadherin locus, none of theinformative cases of gastric carcinoma exhibited func-tional loss affecting the adjacent cadherin-11 locus(Fig. 3). In renal carcinoma, only 1/15 tumours exhibitedloss of expression of the E-cadherin locus. Here again,the effect did not extend to the cadherin-11 locus. Incolonic carcinoma, we observed functional loss of oneallele of the E-cadherin locus in 5 of 22 informativecases. Sequencing the complete coding region revealedno mutations in the E-cadherin cDNA isolated byRT-PCR from any of the colonic and renal tumoursshowing allelic losses of the E-cadherin locus.
J. Pathol. 187: 530–534 (1999)

533FUNCTIONAL ALLELIC LOSS OF CADHERIN LOCI IN COLONIC CANCER
Fig. 2—Loss of cadherin-11 transcript heterozygosity in one case of human coloniccarcinoma. Left panel: loss of transcript heterozygosity at nucleotide 919. In non-tumourtissue, the polymorphic locus contains both T and C, whilst in tumour tissue, only the Cremains. Right panel: loss of polymorphic status at nucleotide 1100. In non-tumour tissue,the polymorphic locus contains G and A, whilst in the tumour tissue, only G is present. Notethe complete loss of expression of one allele in tumour tissue
Fig. 3—Loss of expression from polymorphic alleles of the cadherin-11 locus in colonic, gastric, and renal carcinomas. Afteranalysis of E-cadherin and cadherin-11 intragenic polymorphisms by RT-PCR, the allelic origin of cadherin expression inindividual patients was assessed. In this illustration, the allelic status of the tumour tissue is categorized as eithernon-informative ("), retention of expression from both alleles (open rectangles), or loss of expression from one allele (filledrectangles)
DISCUSSIONLoss of cadherin gene function in adult tissues has
been linked to both uncontrolled cell growth andincreased metastatic potential.6–8,23 The pattern ofE-cadherin mutation combined with loss of heterozy-gosity or allelic inactivation in diffuse gastric canceris consistent with a tumour-suppressor function ofthe E-cadherin gene situated on chromosome 16q22.14
However, in both gastric and mammary tumours, asignificant proportion of the malignancies showing
Copyright ? 1999 John Wiley & Sons, Ltd.
inactivation or loss of E-cadherin do not exhibitmutation of the remaining allele,8,14 suggesting thatadjacent loci on chromosome 16q22 may be involved inthe pathogenesis of the malignancy. Indeed, allelic lossesaffecting this region of chromosome 16 have been docu-mented in several different malignant tissues.16–19 Thepresence of at least three cadherin genes in the vicinity ofthe deleted region15 suggests that loss or inactivation ofthis gene region may involve additional members of thecadherin gene family.
J. Pathol. 187: 530–534 (1999)

534 E. BRAUNGART ET AL.
One such candidate is cadherin-11, a developmentallyregulated mesenchymal cadherin24,25 expressed in sev-eral adult tissues, including bone, heart, brain, andkidney.21 Expression of cadherin-11 has also been estab-lished in epithelial tumour cell lines derived from signet-ring gastric tumours and from renal carcinoma.21,22 Wereport here that cadherin-11 is also expressed in SW480colonic carcinoma cells. To allow a direct assay of theallelic status of the cadherin-11 locus, we identified foursequence polymorphisms in the coding region ofcadherin-11. Three of the polymorphic sites result inamino acid substitutions in the extracellular cadherinadhesion domains believed to be essential forhomophilic adhesion. These substitutions may influencethe adhesive function of the protein, leading to subtleheritable differences in cell adhesion. Indeed, a somaticpoint mutation in the comparable region of E-cadherinhas been reported in diffuse gastric carcinoma.6
In gastric carcinomas, the previously describedinvolvement of the E-cadherin locus6,14 did not extendto the nearby cadherin-11 locus. We identified eightinstances of functional loss of the E-cadherin locus in 22informative individuals, whilst the status of cadherin-11transcripts was unaltered in all 24 informative cases. Inrenal cell carcinoma, neither the E-cadherin nor thecadherin-11 loci exhibited a frequent involvement, withonly a single example affecting the E-cadherin gene.
In colonic carcinoma, 5 of 23 informative casesshowed loss of transcription from one of the polymor-phic cadherin-11 alleles, with no evidence of mutation ofthe remaining allele. A similar frequency of functionalallelic loss (5 of 22 informative cases) was noted at theE-cadherin locus, albeit in different cases. Again, thisinactivation of the E-cadherin locus occurred in theabsence of an accompanying gene mutation. A recentreport has described a somewhat lower rate (10 per cent)of E-cadherin loss of heterozygosity in colorectal can-cers. This may be attributable to differences in tumourmaterial or in the sensitivity of the methodologiesused.26 In all, a total of nine colonic cancers showed lossof heterozygosity of the transcripts from the E-cadherinor cadherin-11 genes. Only one tumour showed simul-taneous loss of both E-cadherin and cadherin-11 tran-scripts, suggesting that the chromosomal region affectedby this phenomenon is quite restricted. Indeed,gross cytogenetic losses in this region have not beenassociated with colonic cancer.27 Nevertheless, relativelysmall regions of inactivation or gene deletion, notreadily detectable by conventional cytogenetics orhybridization-based techniques, may affect all or part ofthe 16q22 locus containing E-cadherin, P-cadherin, andcadherin-11. As neither the E-cadherin nor cadherin-11genes exhibited mutations, we cannot exclude that theregion affected in colonic cancer extends to include otheradjacent loci, possibly including additional cadheringenes.
Whatever the mechanism, functional allelic loss couldserve to reduce the number of actively transcribedcadherin loci, thereby diminishing the levels of cadherinprotein. This would contribute to the excess of freeâ-catenin that is believed to be a critical stage incolorectal carcinogenesis.12 This may explain the
Copyright ? 1999 John Wiley & Sons, Ltd.
reduced level of E-cadherin expression that is seen tocorrelate with a worse prognosis in colonic cancer.28
REFERENCES1. Gumbiner BM. Signal transduction of beta-catenin. Curr Opin Cell Biol
1995; 7: 634–640.2. Takeichi M. Cadherin cell adhesion receptors as a morphogenetic regulator.
Science 1991; 251: 1451–1455.3. Gumbiner BM. Cell adhesion: the molecular basis of tissue architecture and
morphogenesis. Cell 1996; 84: 345–357.4. Larue L, Ohsugi M, Hirchenhain J, Kemler, R. E-cadherin null mutant
embryos fail to form a trophectoderm epithelium. Proc Natl Acad Sci U S A1994; 91: 8263–8267.
5. Radice GL, Rayburn H, Matsunami H, Knudsen KA, Takeichi M, HynesRO. Developmental defects in mouse embryos lacking N-cadherin. Dev Biol1997; 181: 64–78.
6. Becker KF, Atkinson MJ, Reich U, et al. E-cadherin gene mutationsprovide clues to diffuse type gastric carcinomas. Cancer Res 1994; 54:3845–3852.
7. Risinger J, Berchuck A, Kohler M, Boyd J. Mutations of the E-cadheringene in human gynecologic cancers. Nature Genet 1994; 7: 98–102.
8. Berx G, Cleton-Jansen AM, Nollet F, et al. E-cadherin is a tumour/invasionsuppressor gene mutated in human lobular breast cancers. EMBO J 1995;14: 6107–6115.
9. Vider BZ, Zimber A, Chastre E, et al. Evidence for the involvement of theWnt 2 gene in human colorectal cancer. Oncogene 1996; 12: 153–158.
10. Iozzo R, Eichstetter I, Danielson K. Aberrant expression of the growthfactor Wnt-5A in human malignancy. Cancer Res 1995; 55: 3495–3499.
11. Korinek V, Barker N, Morin PJ, et al. Constitutive transcriptional acti-vation by a beta-catenin–Tcf complex in APC"/" colon carcinoma.Science 1997; 275: 1784–1787.
12. Morin PJ, Sparks AB, Korinek V, et al. Activation of beta-catenin–Tcfsignalling in colon cancer by mutations in beta-catenin or APC. Science1997; 275: 1787–1790.
13. Miyoshi Y, Nagase H, Ando H, et al. Somatic mutations of the APC genein colorectal tumors: mutation cluster region in the APC gene. Hum MolGenet 1992; 1: 229–233.
14. Becker KF, Höfler H. Frequent somatic allelic inactivation of theE-cadherin gene in gastric carcinomas. J Natl Cancer Inst 1995; 87:1082–1084.
15. Yeh SH, Chen PJ, Lai MY, Chen DS. Allelic loss on chromosomes 4q and16q in hepatocellular carinoma: association with elevated alpha-fetoproteinproduction. Gastroenterology 1996; 110: 184–192.
16. Osman I, Scher H, Dalbagni G, Reuter V, Zhang ZF, Cordon-Cardo C.Chromosome 16 in primary prostate cancer: a microsatellite analysis. Int JCancer 1997; 71: 580–584.
17. Kihana T, Yano N, Murao S, et al. Allelic loss of chromosome 16q inendometrial cancer: correlation with poor prognosis of patients and lessdifferentiated histology. Jpn J Cancer Res 1996; 87: 1184–1190.
18. Iida A, Isobe R, Yoshimoto M, Kasumi F, Nakamura Y, Emi M.Localization of a breast cancer tumor suppressor gene to a 3 cM intervalwithin chromosome region 16q22. Br J Cancer 1997; 75: 264–267.
19. Bussemakers MJ, van Bokhoven A, Voller M, Smit FP, Schalken JA. Thegenes for the calcium-dependent cell adhesion molecules P- and E-cadherinare tandemly arranged in the human genome. Biochem Biophys Res Com-mun 1994; 203: 1291–1294.
20. Chomczynski P, Sacchi N. Single step method of RNA isolation by acidguanidinium isothiocyanate phenol chloroform extraction. Anal Biochem1987; 162: 156–159.
21. Shibata T, Ochiai A, Gotoh M, Machinami R, Hirohashi S. Simultaneousexpression of cadherin-11 in signet-ring cell carcinoma and stromal cells ofdiffuse-type gastric cancer. Cancer Lett 1996; 99: 147–153.
22. Shimazui T, Giroldi LA, Bringuier PP, Oosterwijk E, Schalken JA. Com-plex cadherin expression in renal cell carcinoma. Cancer Res 1996; 56:3234–3237.
23. Herminston ML, Gordon GJ. Inflammatory bowel disease and adenomas inmice expressing a dominant negative N-cadherin. Science 1995; 270: 1203–1207.
24. Hoffman I, Balling R. Cloning and expression analysis of a novel mesoder-mally expressed cadherin. Dev Biol 1995; 169: 337–346.
25. Kimura Y, Matsunami H, Inoue T, et al. Cadherin-11 expressed inassociation with mesenchymal morphogenesis in the head, somite, and limbbud of early mouse embryos. Dev Biol 1995; 169: 347–358.
26. Ilyas M, Tomlinson IP, Hanby A, Talbot IC, Bodmer WF. Allele loss,replication errors and loss of expression of E-cadherin in colorectal cancers.Gut 1997; 40: 654–659.
27. Bardi G, Johansson B, Pandis N, et al. Cytogenetic analysis of 52 colorectalcarcinomas—non-random aberration pattern and correlation withpathologic parameters. Int J Cancer 1993; 55: 422–428.
28. Dorudi S, Sheffield JP, Poulsom R, Northover JM, Hart IR. E-cadherinexpression in colorectal cancer. An immuno-cytochemical and in situhybridization study. Am J Pathol 1993; 142: 981–986.
J. Pathol. 187: 530–534 (1999)










![WallFlex Colonic Stent - Boston Scientific- US · WallFlex ™ Colonic Stent Visualization Expertise in combining stent materials has resulted ... (BTS). “The WallFlex™ [Colonic]](https://img.pdfslide.us/doc/110x75/5ae601bc7f8b9a8b2b8ca931/wallflex-colonic-stent-boston-scientific-us-colonic-stent-visualization-expertise.jpg)