Embed Size (px)
Citation preview

Functional
Ecology 1991.
5, 40-55
P. C. WAINWRIGHT:1:,C. W. OSENBERG* andG. G. MITTELBACHtDepartment of Ecology and EvolutionaryBiology, University of California, Irvine,California 92717, USA, *Marine ScienceInstitute and Department of Biological Sciences,University of California, Santa Barbara,California 93106, USA and tKellogg BiologicalStation and Department of Zoology, MichiganState University, Hickory Corners, Michigan49060, USA
Abstract. The effects of diet on morphology of thefeeding apparatus were explored in the pumpkin-seed sunfish, Lepomis gibbosus Linnaeus. Adultpumpkinseeds feed primarily on gastropod mol-luscs that they crush between their pharyngealjaws. Development of the muscles and bones of thepharyngeal jaws was contrasted in two Michigan(USA) lakes that differed both in snail density andin the numbers of snails in pumpkinseed diets. Inleast squares regressions of muscle masses, bone .masses and bone shape variables on body mass, allstructures examined showed isometry or positive .
allometry , with those structures most directlyinvolved in snail crushing tending to scale withpositive allometry .Of nine pharyngeal jawmuscles that were examined, analyses of cova-riance found six to be significantly larger in fishfrom snail-rich Three Lakes. The primary crushingmuscle, the levator posterior, was more than twiceas massive in Three Lakes fish than it was in fishfrom snail-poor Wintergreen lake. All three of thepharyngeal jaw bones used to exert crushing forceson snails were found to be significantly larger andof different shape in fish from Three Lakes. Themasses of one muscle and two bones that do notfunction in prey crushing showed no differencesbetween lakes. Teeth on the crushing surfaces ofthe jaw bones were worn down, and thus, shorter
*Present address: P. C. Wainwright, Department ofBiological Sciences, Florida International University,Miami, Florida 33199, USA.40
in Three Lakes fish. Ontogenetic variation ingrowth rates of two muscles, the levator posteriorand the retractor dorsalis, occurred in fish fromThree Lakes. These muscles exhibited strong pos-itive allometry between 40 and 80 mm standardlength (SL), the size range over which fish dietsshift from soft-bodied invertebrates to snails, butgrowth slowed to strong negative allometry in fishlarger than 80 mm. These differences betweenlakes in muscle and bone growth appear to be adirect consequence of the physical regime createdby repeatedly crushing snails during ontogeny.This plasticity indicates that transformations inthe feeding mechanism may originate as specificresponses to the demands of novel diets.
Key-words: Allometry, Lepomis gibbosus, ontogeny,phenotypic plasticity
Introduction
It is common for populations of a single speciesthat occur in different environments to divergephenotypically (e.g. Lindsey, 1981; Lavin &
; :McPhail, 1985; Grant, 1986; Hoogerhoud, 1986;!.0- Liveley, 1986; Magnan, 1988). The differences that...,emerge can be a result of shifts in the genetic~ake-up of the population (Carl & Healey, 1984;Grant, 1986; Trexler & Travis. 1990), via naturalselection. founder effects and other mechanisms,or differences may result from developmentalplasticity of the species responding to local con-ditions (Collins & Cheek, 1983; Liveley, 1986). Inthe latter case. population-level variation in func-tionally important characters provides an oppor-tunity to study the role of the environment indetermining phenotypic expression. Further. suchinstances permit a direct assessment of the degreeto which particular functional systems mayrespond independently from other anatomicalstructures, even those that lie in close physicalproximity .
In this study we examine the phenotypicresponse of the feeding mechanism in the pump-kinseed sunfish, Lepomis gibbosus Linnaeus, tointerlake variation in their prey resource. Adult

41
Trophic
polymorphism
in
pumpkinseed
sunfish
pumpkinseeds are typically trophic specialists ongastropod molluscs which they crush in theirpharyngeal jaws (Lauder. 1983a; Mittelbach.1984;Osenberg & Mittelbach. 1989). We take advantageof natural variation between lakes in the abun-dance of snails and document the influence of anenvironmentally imposed diet shift on the dimen-sions of the bones and muscles of the pharyngeal
jaw apparatus.Snail crushing in pumpkinseeds offers a parti-
cularly promising system to study the effects ofdiet on the feeding mechanism because of exten-sive previous research on both the functionalmorphology of snail crushing (Lauder. 1983a,b;Wainwright & Lauder. 1991) and the generaltrophic ecology of this species (Keast. 1978; Mit-telbach.1984.1988; Osenberg & Mittelbach.1989).Only rarely are studies of polymorphic speciesmade in the light of such detailed knowledge of thefunctional morphology and ecology of a complexfunctional system (i.e. Liem & Kaufman. 1984;Meyer. 1989, 1990). These pre-existing data notonly strengthen our interpretations of the func-tional and ecological consequences of morpho-logical differences. but. by understanding thefunctional morphology of snail crushing. we areable to infer the mechanistic processes that pro-duced the morphological variation.
levels and the density of competitors (Werner &Hall, 1979). In this study we take advantage ofpronounced differences in pumpkinseed dietsbetween two natural lakes in southern Michigan(USA) to examine the impact of consuming gastro-pods on the development of the pumpkinseeds'pharyngeal jaws. Wintergreen lake has a verydense population of pumpkinseeds and adepauperate snail fauna (C. W .Osenberg, G.G.Mittelbach & P .C. Wainwright, unpublished obser-vations). In this lake, gastropods make up less than10% of adult pumpkinseed diets by dry weight. Innearby Three Lakes II (hereafter simply calledThree Lakes), snail abundances are more typical ofthe region (about an order of magnitude higherthan in Wintergreen lake) and gastropods are thepredominant prey in the diets of large pumpkin-seeds (60-100% by dry weight for fish greater than75mm SL; Mittelbach, 1984; Osenberg, 1988). Wehypothesized that these interlake differences insnail abundance and pumpkinseed diets wouldaffect pharyngeal jaw anatomy. Below we comparethe musculature and bone structure of the pharyn-geal jaws of pumpkinseeds collected from the twolakes, documenting an increased robustness inthese structures with ontogeny in Three Lakesrelative to Wintergreen lake. No differences werefound between lakes in several structures notinvolved in snail crushing. Elsewhere we alsoshow that pumpkinseeds from Wintergreen lakeare less proficient at crushing snails and haveslower adult growth rates than do fish from ThreeLakes (C. W .Osenberg, G.G. Mittelbach & P .C..,Wainwrig~}"unpublished observations ).
~Materials a~d methods
Thirty-three pumpkinseeds, ranging in size from40 to 132mm SL, were collected with seines fromThree Lakes on 21 and 23 June 1988, and 20individuals (45-109mm SL) were collected fromnearby Wintergreen lake on 6 July 1988. Eviscer-ated fish were fixed in 10% buffered formalin forabout 4 weeks, transferred to 70% ethanol forstorage and weighed to the nearest 0.01 g.
Pharyngeal jaw anatomy was contrasted in thetwo lake populations through three types ofmeasurements: (1) the mass of pharyngeal jawmuscles; (2) the mass of pharyngeal jaw bones; and(3) aspects of the shape of pharyngeal jaw bones.The entire branchial apparatus was dissected fromeach preserved fish and the left side members ofnine paired muscles and one unpaired musclewere carefully removed and stored separately. Astrict protocol was followed during weighing to
The system
The pumpkinseed sunfish, L. gibbosus, and itssister species L. microlophus Gunther are the onlymolluscivorous members of the endemic NorthAmerican freshwater fish family Centrarchidae.Mollusc crushing in these species is associatedwith numerous morphological and neuromuscu-lar specializations (Lauder, 1983a,b, 1986; Wain-wright & Lauder, 1990), including hypertrophy ofthe pharyngeal jaw muscles, bones and teethbetween which snails are crushed and a novelpattern of muscle activity that is used during snailcrushing. As expected, gastropods commonlymake up a large fraction of adult pumpkinseeddiets in nature (often > 70% by volume or dryweight; Sadzikowski & Wallace, 1976; Keast,1978;Mittelbach, 1984; Osenberg & Mittelbach, 1989).Small pumpkinseeds, which are unable toeffectively crush snails (Mittelbach, 1984), feed oninsect larvae and other soft-bodied invertebrates(Sadzikowski & Wallace, 1976; Keast, 1978).
Although pumpkinseeds show considerablespecialization for feeding on snails, they remainflexible in their diet and habitat choice, and maychange diets in response to changes in resource

(b)42
P.C.
Wain
et al.
CB5(c
CB5
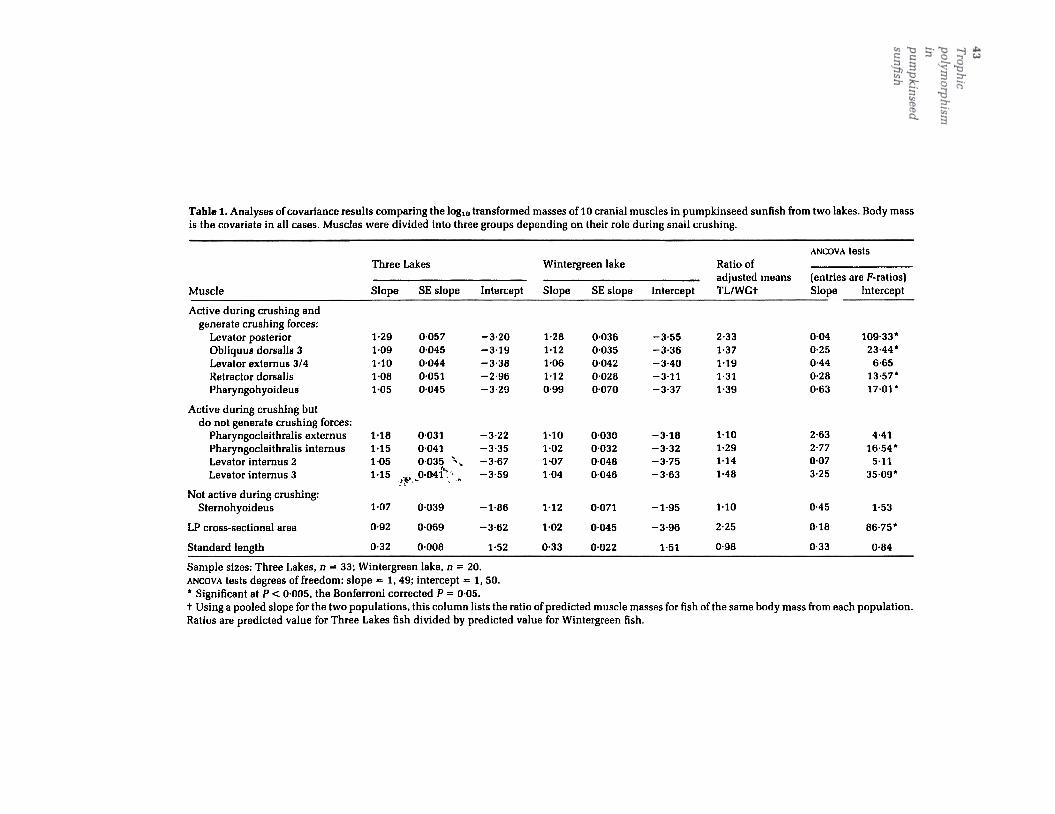
Fig. 1. Illustrations of the snail-crushing mechanism in the pumpkinseed sunfish. (a) Schematic lateral view of the headillustrating the position of the pharyngeal jaw apparatus and the muscles that function during its use. Muscles areshown as thick black lines indicating their attachments. [b) Posterior view diagram of the pharyngeal jaws illustratingthe mechanism of upper jaw depression that is crucial to snail crushing. During snail crushing the lower jaw is heldrelatively stationary and the upper jaw exerts the primary crushing force as it is pressed firmly against the snail shell.Upper jaw depression is caused by rotation of epibranchial4 about the insertion site of the obliquus posterior muscle.This rotation causes the epibranchial to press against the dorsal surface of the upper jaw. forcing it downward. Thiscrushing action is produced by several muscles. principally the levator posterior .levator externus 314. and the obliquusdorsalis. BC. buccal cavity; CB5. fifth ceratobranchial or lower pharyngeal jaw; EB4. fourth epibranchial; ES.oesophagus; LP. levator posterior; LV. fourth levator externus; NC. neurocranium; OD3. third obliquus doralis; OP.obliquus posterior; PB3. third pharyngobranchial or upper pharyngeal jaw; PCe. pharyngocleithralis externus; PCi.pharyngocleithralis internus; PEC. pectoral girdle; PH. pharyngohyoideus; RD. retractor dorsalis.
minimize measurement errors. Each muscle wasweighed twice to the nearest 0.01 mg, after firstbeing patted twice on a dry paper towel to removeexcess moisture. The average of these twomeasurements was used in subsequent analyseson the 10 muscle mass variables (Table 1). The 10muscles were (see Fig. 1): the levator posterior ,
(LP), a primary upper jaw depressor; the thirdobliquus dorsalis (OD3), also an upper jawdepressor; the third and fourth levator externus(LE3/4, these muscles are believed to functiontogether as upper jaw depressors [Lauder, 1983a;Wainwright, 1989a] and are difficult to separatephysically so they were treated together); thesecond levator internus (U2), an upper jaw eleva-tor and protractor; the third levator internus (U3),also an upper jaw elevator and protractor; theretractor dorsalis (RD), the only muscle thatretracts the upper jaws posteriorly; pharyn-gocleithralis internus (PCi), a lower jaw depressorand retractor; the pharyngocleithralis externus(PCe), also a lower jaw depressor; the pharyn-gohyoideus (PH), a lower jaw protractor; and thesternohyoideus (SH), the only muscle measuredthat is unpaired and that is not part of thepharyngeal jaw apparatus. The SH muscledepresses the hyoid bar during suction feeding,and therefore serves a general role during the
wright
capture of all prey types (Lauder, 1985). The SHwas selected as a control because it does notfunction during snail crushing. Thus, it enabled usto determine if any observed differences amonglakes in pharyngeal jaw muscle masses werespecific to the pharyngeal jaws or simply a cor-,r-elated response in all cranial muscles.
.;.. In addition to the masses of these 10 muscles, an-.'f;stimate of the physiological cross-sectional areaQf the levator posterior muscle was obtained sothat approximate differences among lakes in thescaling offorce-producing capacity of this primarycrushing muscle could be made. This is of interestbecause it is conceivable that muscle masses coulddiffer among lakes while, through organizationalchanges within the muscle, no differences result inthe force-producing capability of the muscle. Asforce production is presumably the feature ofprimary importance in limiting snail-crushingability in these fishes we explored this additionallevel of potential population divergence. Musclemass was divided by an estimate of muscledensity, 1.05g cm-3 (Lowndes, 1955), to obtainmuscle volume and this was then divided by theoverall length of the muscle, which was measuredto the nearest 0.1 mm, to obtain cross-sectionalarea. Such a procedure will only produce a reliableestimate of physiological cross-sectional area if the
"'"'coe>-
~0~.,;Q)
~~0~e.g.J::"'
==="'~Q
)Q
) .
"' O
D=
.=
~.J::p,"'e
2=
u
p,==
co
.-="' "'
Q)
OD
-=U
.-"'
..=
=
e~-Q
)C
O-
.-0=
..C
O ..
U
Q)
O.J::
-=O
O"'
OD
Q)
="'
.-"'~C
O =
e Q
)~
p,
Q) Q
)
e~.."'..9
§'"'
0=
..
~
:0
Q)
~
..O
D.J::
O
--0Q
) -
-=
.S
OD
~=
Q
).C
~
=
.>p,.-e~o
~U
Q
)
~
~'3
"'"'
Q)
Q)-
..UQ) "'
U
=
=~
CO
.
.-"'..Q
)C
O "'
>
CO
8 u
-=O
co"'
=Q
) .-"'
Q)
>--
-;a .~
=
a<
>
.0...uQ
I Q
):=
.J::co-
!-o.~
-;n-o
c..-.,-utC
...
';' .,
"' t.;.
-
-="' .,-
., ...
-tC<
"'>
.,
0 .-.,
u .=
c.z
=
0<
.,--C
/)
"'CJ
jCJ
CJ
~ Q)
~~=Q)
Q)
~Q)
=~
"'c:~Q)
s~+
-O
Q)t-'
ou;>.-~
;;..
co J
~~
!-
0)~0";ntzJC
/)0)u"';I
:2: Q)
~orn Q)
Po
o(jj Q)
~o";nt.J(/)'5.QJ
~~.5 Q.Q)
tJ..Q)
:5
;,; "'
"'Q
) =
=
~
=
=.s~
..--Q
) ..
w
..-QO
><-~
~=
"' =
' =
-
-Q)
-~=
Q).-
='.u
~
~.-
.-Q
O .."'
"(;; "'
QO
~ .~
.~ -.-~
ta
-=
0 "'
"' =
=
"'
--" "
"'5~
"'~Q
) .-2~
~"'"'
2 ~
~00.-ta=
ta~
~u~
~=
=
u ta
"'-=.."'
E.'.-
"' --=
=
"' -
2.-Q
) 0
2Q
) Q
O -ta
~-O
Q)O
>,
-Q)Q
)Q)Q
) -~
--
u"'"' tJta
~Q
) ~
...,
=ow
><
w...,
..tJtJ==
.t:"tJ
0 -
QO
..C.~
~..O
Q
O~
OO
=
'.- =
~
=u..=
..OQ
O
=~
QO
QO
-0
tJ ~
.-0 =
0
-=
.-Q)
=
=
0 0
w
>,
~
~
32!.-C'-~
>,
3QO
>,
ri--~
~
"' -
-cata.-ca tata
>0.;'
"tJw
..>->
-ta w
ota »
.-=
..Q
)a>Q
)~O
~a>
~
~=
~~
Q)Q
) U
.. ~
~
>=
...J ...J~
Q.
>0
Q.Q
J...J
taQ)
..~.-a>
.--tJ
~
uOO
U
"tJ 0(/)
Q.
:E
<
<
Z
...J (/)
C")~
"'I'-,-1C
")~co"'o
dI"'cD"'r:..
ON
,-1,-1
,-IC)C
)C~
~N
C
CC
.:..:..:..:..:.
~"'.,
"'"'.'..'.""'i'0000000000
oc.=~
c.N
"'c.N
';";";'N';'
I I
I I
I
=N
CC
NC
)N
... 0...
C)
~~
~~
O
CO
I/)N~
O"""~
N"'
0000066666
InCC
O
In(,) ~
...(')
.;,.;,.;,.;,.;,I
I I
I I
...r..CI
CI
N.:..:..:..:.
""""'~"'
°""""'tC66666
N"",Q
)N
"""",M
MM
MI
I I
I
ON
~~
...000~
~~
~
ON
~<
O"""~
~00006666
=N
Il)",,..."'t-.tC.:.0.:.0.:.0.:.0I
I I
I
QC
),#,=':'~
':"!'
"'1'-1'-1/)C
CI'-°"'
NN
6C:'
..~
aJ~
"' o
~rD
';";'"'
~
,,'.
"
~j;..;-
""""'~0000
6666",iv.~
,
~~
~~
0...
,:.,:.,:.,:.
r-. N
N
Q
=
"'.:.
6 6
a.a.~"'
~
oo
o o
6 6
6
~
N
N~
~
II)
.:. M
.:.
I I
N
N
C')
'"' Q
C
').:.
.:. 6
"' N
1-- .,.
NO
O
O
6 6
6
II) cc
...0'
0' II)
.:. ~
.:.
I I
0 ~
~
...N
ao~
N
6
Lt) ~
"'
~
..."'6
6 6
"0')
II) ~
II) C
Q.:.
tD
6C
Q
d.9-;"31:1.01:1.
~U~Q)
eo..t="'"'~e>,
~0.J:;Q
)e~"'Q
)-:S'-0~"'(J:....E"'Q
)"'"'~
.
e~Q)(J:
U
~"'
Q)
~
Q)
e ~..
~
Q)
Q)-
-~U
.-
:a~~
..I:I...E
'- Q
)0
~0-
.-~-; >
..~Q)
Q)
.~
-0
-U"'
.-..!?
~0
",Q)
"' N
0-1:1.
IIII
.~
-oe>'
~g.1I
~.J:;
.u,,-~Q
)..-°Q)
~Q
) U
~
~-~
"'.--C
:Q).->
c: .-U
~
.-
Q)
..Q)
-~
~~
t=,,;~
f.O
8§(J:Q
) .-"'
-II
.--Q)
c: c:.!S
~.-Q
) 0
~
~~
1:1. t=
I:I.~
...9.~
0 Q
)",",c:1:1.Q
)"'
..0 0
.E.
II e~
~L.
OQ
)-..~
~~
Q)..
.Q)-~
..E"'
Q)
.-
Q)..t=
"'..Q)
~,-g..E
..3~
0
.Q)
~Q
)"'°1:1.Q
)21V
O~
Q)
~..,,"'-
OO
-~U
E-o
Q)
-Q)
.-..~
~
-~",",-°Q
).~
-cn
; g,
~
"'~U
~~
(J: ..
~<
.-00~I:I.>
C:C
:"'"'
O
00 .-0
'" U
.-"'
.-
~Z
r/J:J-;r/J«<
+
~

(b)
PB3length
44
P.C.
Wain
et al.
2mm
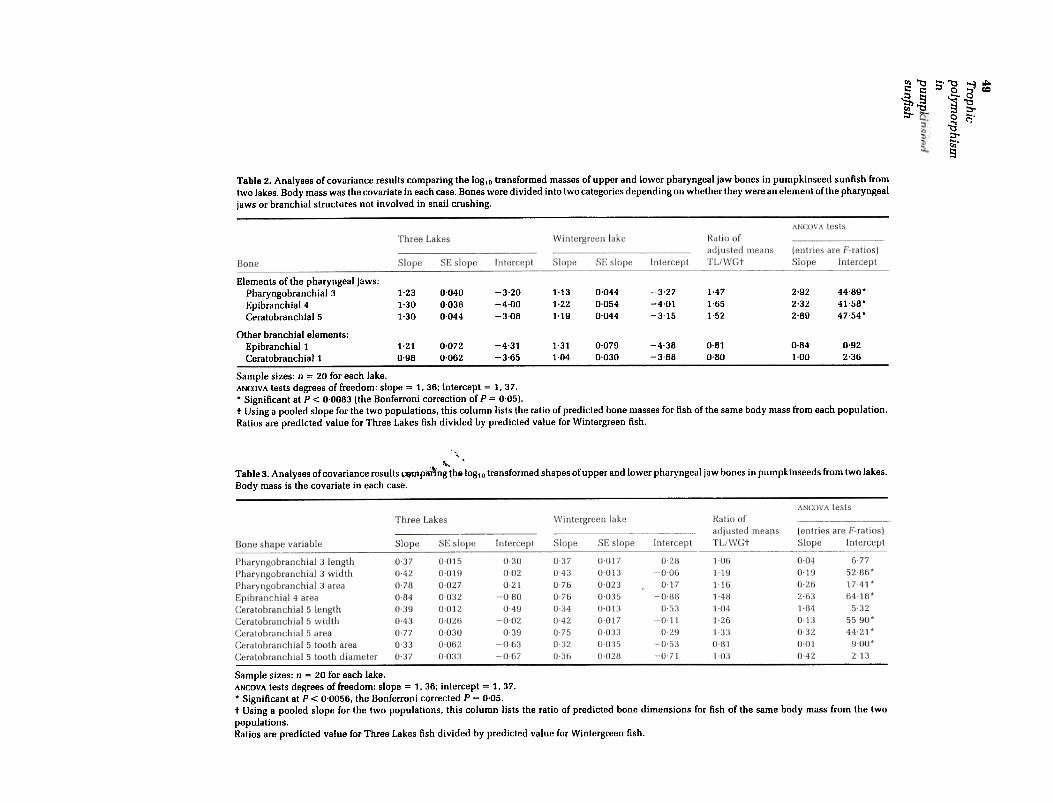
Fig. 2. Diagrams of the toothed pharyngeal jaw bones from a pumpkinseed sunfish illustrating the morphologicalmeasurements that were taken to summarize aspects ofbone shape. (a) Pharyngobranchial3. the upper pharyngeal jawbone and its associated tooth plate. shown in ventral view; (b) ceratobranchial5. the lower pharyngeal jaw bone and itsassociated tooth plate. shown in dorsal view; (c) ceratobranchial 5 shown in mesial view.
LP muscle has parallel fibres in the axis of action ofthe muscle, from origin to insertion. Smalldeviations from a parallel fibred arrangement willnot have a strong effect on estimates of physio-logical cross-sectional area (Powell et al., 1984).Observations under the dissecting microscopeconfirmed that the muscle fibres were orientedapproximately parallel to the muscle's axis ofmovement.
Following removal of the muscles, five boneswere dissected out of the branchial apparatus from20 fish from each lake. Each branchial apparatus ,was first cleared and double stained for bone andcartilage (Dingerkus & Uhler, 1977) to facilitate therecognition and separation of the bones from thepharyngeal jaws. The following bones wereremoved and weighed: the fifth ceratobranchial(CB5) or lower pharyngeal jaw bone; the thirdpharyngobranchial (PBJ) or upper pharyngeal jawbone; the fourth epibranchial (EB4); the first epi-branchial (EB1); and the first ceratobranchial(CB1). In each case the left side member of thepaired bones was taken. Of these bones only theCB5, PBJ and EB4 bones are thought to be directlyinvolved in the snail-crushing movements of thepharyngeal jaws (Lauder, 198Ja; Wainwright,1989a). The EB1 and CB1 bones were selected ascontrols to ensure that any differences that mightbe found between lakes in bone weights were notdue simply to effects seen in all skeletal elements.All bones were patted twice on a paper towel andweighed to the nearest 0.01 mg. The average of twomeasurements was used in subsequent analyses.
wright
To gain a more complete understanding of themorphological differences between the two lakepopulations, nine additional measurements weremade on the upper and lower pharyngeal jawbones (PB3 and CB5) and the bone that pushes theupper jaw ventrally during crushing (EB4). Eachbone was viewed under a dissecting microscopethat was equipped with a computer interfacedvideo camera. Desired views were digitized and acomputer program aided in making linear or arealmeasurements. Nine variables were measured that
, lf1ore finely determined the differences between
.rl~kes in bone shape (Fig. 2): from PB3 the anterior-~osterior length, maximum lateral width and areaof the toothed surface were measured; from EB4-the area projected by the surface articulating withPB3; from CB5 the anterior-posterior length, maxi-mum width, area projected in dorsal view, theaverage diameter of the four widest teeth and thelength of the teeth above the deepest portion of thekeel of CB5.
Statistical analyses
Because the samples of fish from the two lakesspanned a broad range of body sizes it wasnecessary to remove the effects of body size fromall variables before testing for differences betweenlakes. All data were loglo transformed to renderbivariate relationships approximately linear andto meet assumptions of the parametric statisticsused. For each group of variables (muscle mass,bone mass and bone shape) multivariate analyses

45
Trophic
polymorphismin
pumpkinseed
sunfish
pumpkinseed sunfish has been discussed in detailelsewhere (Lauder. 1983a; Wainwright. 1989a;Wainwright & Lauder, 1991) but a brief account ispresented here to provide a context in which tointerpret the morphological differences betweenthe two lakes. Snails are first captured by the oraljaws. using suction feeding. and are then passed tothe pharyngeal jaw apparatus for processing. Thekey movement in snail-crushing behaviour is thedepressive action of the upper pharyngeal jaw(PB3) against the relatively stationary lower phar-yngeal jaw (CB5; see Fig. 1). This actioncompresses the snail shell between the jaws.ultimately crushing the shell. Upper jaw depress-ion is accomplished through rotation of the fourthepibranchial about the insertion of the obliquusposterior muscle on its mid-ventral aspect (Wain-wright. 1989a). The rotating fourth epibranchialarticulates with the dorsal surface of the upperpharyngeal jaw and presses it ventrally. Severalmuscles, principally the levator posterior, fourthlevator externus and third obliquus dorsalis func-tion to depress the upper jaws. At the same timethat the upper jaw is depressed it is also retractedposteriorly by the retractor dorsalis muscle.During crushing the lower jaws are situated suchthat the posterior region is more dorsal than theanterior region. The snail is thus held against thissurface while the upper tooth plates press ven-trally and posteriorly on the snail.
Electromyographic studies of the patterns ofmuscle activity exhibited during snail crushing(Lauder. 1983a,b) have revealed a novel motorpattern in the 'pumpkinseed and its snail-crushingsister speci~s: the redear sunfish, L. microlophus.During crushing all pharyngeal jaw muscles are,active simult~eously in intense bursts. Thus, inaddition to the upper jaw depressors many antago-nistic muscles are active during crushing. pre-sumably to stabilize the jaws during the forcefulexertion of the crushing action.
of covariance (MANCOVA) were first used to makean overall comparison between the two lakes.Analysis of covariance (ANCOVA) was thenemployed to compare all morphological variablesbetween the two lake populations. Fish mass(without viscera) was selected as a proxy for bodysize and used as the covariate in all analyses.Because so many significance tests were con-ducted on highly correlated variables a Bonferronicorrection of the critical probability value of p <0.05 was used in each of the three data sets (Snede-cor & Cochran, 1980); muscle masses, bone massesand bone shape. For example, 10 muscle masseswere contrasted so the corrected critical prob-ability value was 0.05/10 = 0.005. This is a
conservative measure that reduces the chance offinding a significant lake effect when there is none.
Bivariate plots of the levator posterior andretractor dorsalis muscle mass against body masssuggested that these relationships were not linearon a log-log scale for the Three Lakes dataset (alsosee Results). This was explored further with acontinuous two-phase regression method (Nicker-son, Facey & Grossman, 1989; Yeager & Ultsch,1989) by progressing along the body mass a-xis,repeatedly dividing the Three Lakes dataset intotwo continuous groups and calculating regressionparameters for the two segments of data. Thecontinuous pair of segments that minimized thecombined residual sums of squares wasdetermined and used to estimate the scaling ofmuscle mass with body size within each sizerange. The point of intersection of the two scalingtrajectories was calculated to determine thecritical point, an estimate of the body size at whichthe rate of muscle growth changes.
An overall comparison of pharyngeal jaw mor-phology between the two lakes was made on the 24variables measured on 20 fish from each lake (10muscle masses, five bone masses and nine bone
shape variables). Principle components analysiswas used first to reduce the dimensionality of theoriginal dataset to four new variables thataccounted for most of the original variance. Adiscriminant analysis was then performed on thefour new variables to examine the distinctivenessof pharyngeal jaw morphology in fishes from thetwo lakes. All statistical calculations were carriedout using Systat version 4.0 (Wilkinson, 1986) on amicrocomputer .
Muscle mass
The relationship between standard length andbody mass was not different for fish from the twopopulations (Table 1). The MANCOVA on the 10muscle masses revealed a highly significant over-all difference between the two lakes ( MANCOV A testof lake by mass interaction, Wilks' A = 0.679, FlO.40= 1.898, p = 0.075; test for lake effect, Wilks' A =0.223, FlO.4l = 14.3, p < 0.001). In the univariate
ANCOVAs no differences were found between lakesin the slopes associated with any muscle (Table 1;but see below). With the Bonferroni corrected
Results
Mechanism of snail crushing
The functional morphology of snail crushing in
46
P. C.
Wain
et al.
-0. -1.0
C/IC/Ic
E -1.5.~OC/I5 -2.0
"0..c
-uc -2.5..
-
~
9 -3.0-0.0 0.5 1.0 1.5 2.0
log body mass (g )
CI -I
II)II)oE~II)~c..0)
-c
..
~0>
.$.
'POjo 00
000. .
",~ " ..
-1.5
-2.0
-2.5
-; -1.0 dJ
~ O~E -1.5 Q. 6"' ~.:. 0-E &66~ -2.0 0~ ~01
~ -2.5 ~0 ~ 8 -3.0 6 O~ 0'1.0 ~'O01 -3.0 01 -3.5 ' O ., ,o o-0-0 0.5 1.0 1.5 2.0 -0.0 0.5 1.0 1.5 2.0
log body mass ( g ) log body mass ( g )
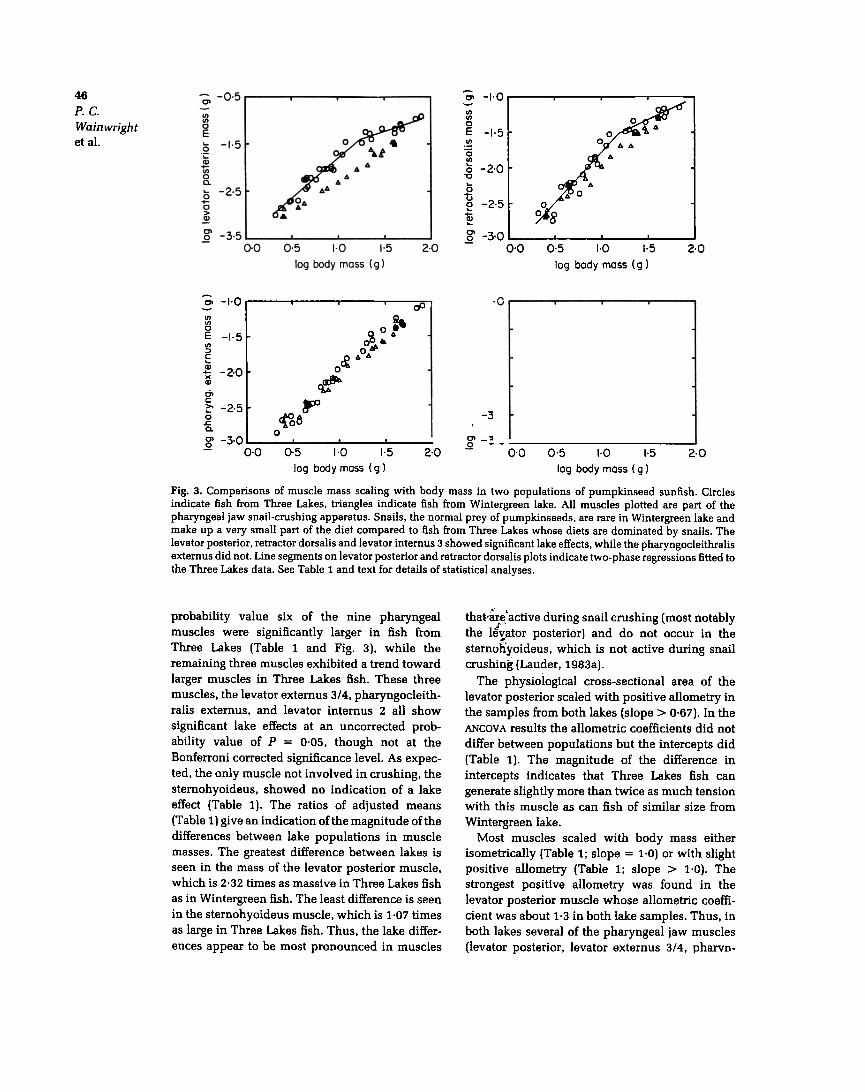
Fig. 3. Comparisons of muscle mass scaling with body mass in two populations of pumpkinseed sunfish. Circlesindicate fish from Three Lakes. triangles indicate fish from Wintergreen lake. All muscles plotted are part of thepharyngeal jaw snail-crushing apparatus. Snails. the normal prey of pumpkinseeds. are rare in Wintergreen lake andmake up a very small part of the diet compared to fish from Three Lakes whose diets are dominated by snails. Thelevator posterior. retractor dorsalis and levator intemus 3 showed significant lake effects. while the pharyngocleithralisextemus did not. Line segments on levator posterior and retractor dorsalis plots indicate two-phase regressions fitted tothe Three Lakes data. See Table 1 and text for details of statistical analyses.
probability value six of the nine pharyngealmuscles were significantly larger in fish fromThree Lakes (Table 1 and Fig. 3). while theremaining three muscles exhibited a trend towardlarger muscles in Three Lakes fish. These threemuscles. the levator externus 3/4. pharyngocleith-ralis externus. and levator internus 2 all showsignificant lake effects at an uncorrected prob-ability value of p = 0.05. though not at the
Bonferroni corrected significance level. As expec-ted. the only muscle not involved in crushing. thesternohyoideus. showed no indication of a lakeeffect (Table 1). The ratios of adjusted means(Table 1) give an indication of the magnitude of thedifferences between lake populations in musclemasses. The greatest difference between lakes isseen in the mass of the levator posterior muscle.which is 2.32 times as massive in Three Lakes fishas in Wintergreen fish. The least difference is seenin the sternohyoideus muscle. which is 1.07 timesas large in Three Lakes fish. Thus. the lake differ-ences appear to be most pronounced in muscles
that!~e: active during snail crushing (most notablythe le~ator posterior) and do not occur in thesternoI1yoideus, which is not active during snailcrushing (Lauder, 1983a).
The physiological cross-sectional area of thelevator posterior scaled with positive allometry inthe samples from both lakes (slope > 0.67). In theANCOV A results the allometric coefficients did notdiffer between populations but the intercepts did(Table 1). The magnitude of the difference inintercepts indicates that Three Lakes fish cangenerate slightly more than twice as much tensionwith this muscle as can fish of similar size fromWintergreen lake.
Most muscles scaled with body mass eitherisometrically (Table 1; slope = 1.0) or with slightpositive allometry (Table 1; slope > 1.0). Thestrongest positive allometry was found in thelevator posterior muscle whose allometric coeffi-cient was about 1.3 in both lake samples. Thus, inboth lakes several of the pharyngeal jaw muscles(levator posterior, levator externus 314, pharvn-
wright
i -800.
00 0
o~.t
~c9 A A
~A AA
47
Trophic
polymorphismin
pumpkinseed
sunfish
O6
~ ..". ...
0.0 0.5 1.0 1.5 2.0
log body moss (g )
01"'"'oE
0:c:(.)c:
e.0o
0'
~oE
-2 -2
+-0~u01O -
0.0 0.5 1.0 1.5 2.0 -0.0 0.5 1.0 1.5 2.0
log body mass (g) log body mass (g )
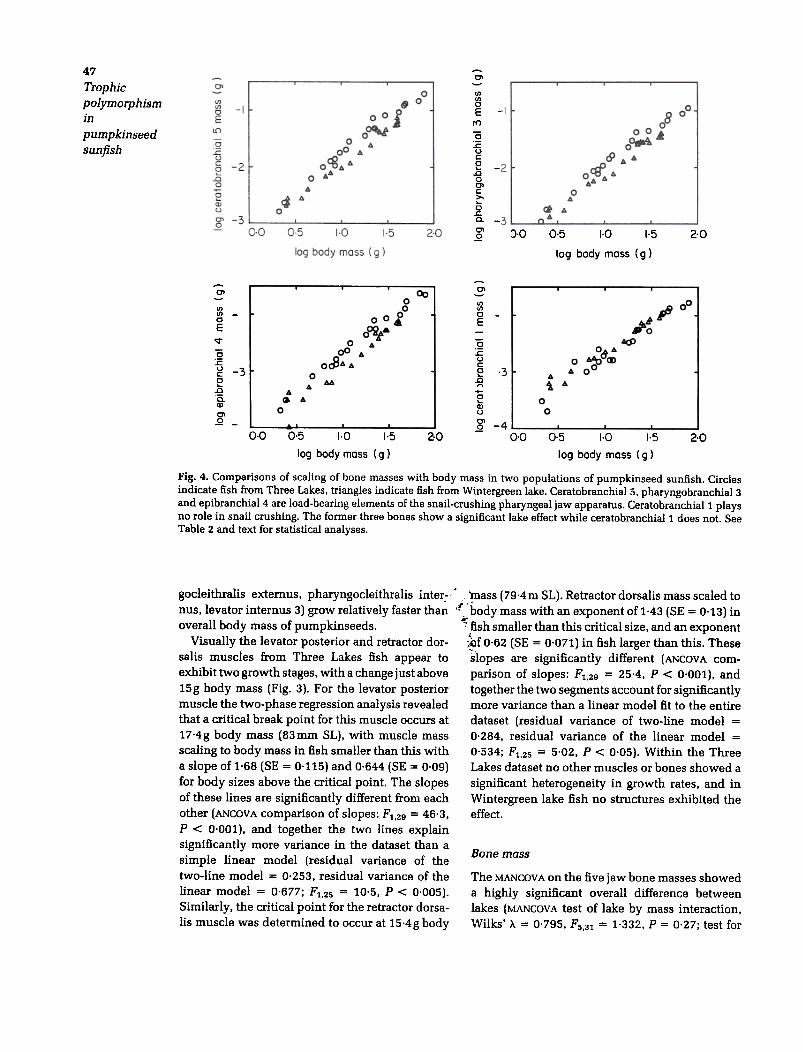
Fig. 4. Comparisons of scaling of bone masses with body mass in two populations of pumpkinseed sunfish. Circlesindicate fish from Three Lakes, triangles indicate fish from Wintergreen lake. Ceratobranchial 5, pharyngobranchial 3and epibranchial 4 are load-bearing elements of the snail-crushing pharyngeal jaw apparatus. Ceratobranchiall playsno role in snail crushing. The former three bones show a significant lake effect while ceratobranchiall does not. SeeTable 2 and text for statistical analyses.
'3"cEucc'-
.cCiQ)
0'..2 -4
gocleithralis externus. pharyngocleithralis inter- .
nus. levator internus 3) grow relatively faster than ,
overall body mass of pumpkinseeds.Visually the levator posterior and retractor dor-
salis muscles from Three Lakes fish appear toexhibit two growth stages. with a change just above15g body mass (Fig. 3). For the levator posteriormuscle the two-phase regression analysis revealedthat a critical break point for this muscle occurs at17.4g body mass (83mm SL). with muscle massscaling to body mass in fish smaller than this witha slope of 1.68 (SE = 0.115) and 0.644 (SE = 0.09)
for body sizes above the critical point. The slopesof these lines are significantly different from eachother (ANCOVA comparison of slopes: F1.Z9 = 46.3.p < 0.001). and together the two lines explainsignificantly more variance in the dataset than asimple linear model (residual variance of thetwo-line model = 0.253. residual variance of thelinear model = 0.677; F1.Z5 = 10.5, p < 0.005).
Similarly. the critical point for the retractor dorsa-lis muscle was determined to occur at 15.4g body
"0~(.)I:O....0 -c;001I:~.E0.01.9.
'mass (79.4 m SL). Retractor dorsalis mass scaled to','
r. body mass with an exponent of 1.43 (SE = 0.13) in...;0 fish smaller than this critical size, and an exponent,l;;(jf 0.62 (SE = 0.071) in fish larger than this. These
11opes are significantly different (ANCOV A com-parison of slopes: F1.29 = 25.4, p < 0.001), and
together the two segments account for significantlymore variance than a linear model fit to the entiredataset (residual variance of two-line model =0.284, residual variance of the linear model =0.534; F1.25 = 5.02, p < 0.05). Within the Three
Lakes dataset no other muscles or bones showed asignificant heterogeneity in growth rates, and inWintergreen lake fish no structures exhibited theeffect.
Bone mass
The MANCOV A on the five jaw bone masses showeda highly significant overall difference betweenlakes (MANCOVA test of lake by mass interaction.Wilks. >.. = 0.795, F5.31 = 1.332, p = 0.27; test for
2 o chial. the third pharyngobranchial and tooth dia-meter did not show a significant lake effect. Thegreatest difference was seen in the area of thearticulating surface of epibranchial 4. which is1.64 times as large in Three Lakes fish as inWintergreen fish. Scaling of the bone shape varia-bles produced a pattern where the bone width andarea variables scaled to body mass with positiveallometry (Table 3; for linear measurements slope> 0.33. for areal measurements slope > 0.67),while the bone length and tooth measurementsscaled isometrically.
0
48
P. C.
Wain
et al.0
66
6,.6'
"EQ)c00.
E
8 oa0.u.!:It
00..0
0
000 0A .
6
44
4 44 .-c. ' , , ,. , ,
-3 -2 -I O I 2 3Principal component 2
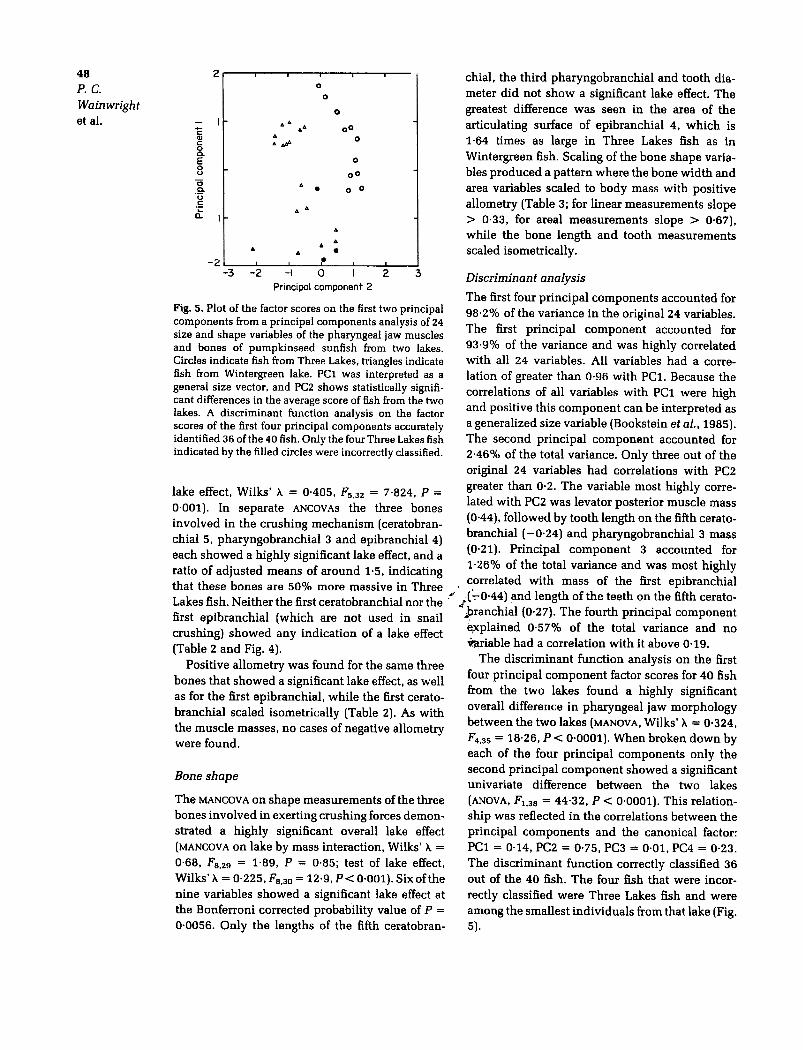
Fig. 5. Plot of the factor scores on the first two principalcomponents from a principal components analysis of 24size and shape variables of the pharyngeal jaw musclesand bones of pumpkinseed sunfish from two lakes.Circles indicate fish from Three Lakes. triangles indicatefish from Wintergreen lake. PCl was interpreted as ageneral size vector, and PC2 shows statistically signifi-cant differences in the average score of fish from the twolakes. A discriminant function analysis on the factorscores of the first four principal components accuratelyidentified 36 of the 40 fish. Only the four Three Lakes fishindicated by the filled circles were incorrectly classified.
lake effect, Wilks' A = 0.405, FS.32 = 7.824, p =
0.001). In separate ANCOVAS the three bones
involved in the crushing mechanism (ceratobran-
chial 5, pharyngobranchial 3 and epibranchial 4)
each showed a highly significant lake effect, and a
ratio of adjusted means of around 1.5, indicating
that these bones are 50% more massive in Three~Lakes fish. Neither the first ceratobranchial nor the -
first epibranchial (which are not used in snail
crushing) showed any indication of a lake effect
(Table 2 and Fig. 4).
Positive allometry was found for the same three
bones that showed a significant lake effect, as well
as for the first epibranchial, while the first cerato-
branchial scaled isometrically (Table 2). As withthe muscle masses, no cases of negative allometry
were found.
Bone shape
The MANCOVA on shape measurements of the threebones involved in exerting crushing forces demon-
strated a highly significant overall lake effect(MANCOVA on lake by mass interaction, Wilks' A =0.68, F8.29 = 1.89, p = 0.85; test of lake effect,
Wilks' A = 0.225, F8.30 = 12.9, p< 0.001). Six of the
nine variables showed a significant lake effect atthe Bonferroni corrected probability value of p =
0-0056. Only the lengths of the fifth ceratobran-
wright
Discriminant analysis
The first four principal components accounted for98.2% of the variance in the original 24 variables.The first principal component accounted for93.9% of the variance and was highly correlatedwith all 24 variables. All variables had a corre-lation of greater than 0.96 with PC1. Because thecorrelations of all variables with PC1 were highand positive this component can be interpreted asa generalized size variable (Bookstein et al.,1985).The second principal component accounted for2-46% of the total variance. Only three out of theoriginal 24 variables had correlations with PC2greater than 0-2. The variable most highly corre-lated with PC2 was levator posterior muscle mass(0-44), followed by tooth length on the fifth cerato-branchial ( -0.24) and pharyngobranchial 3 mass(0-21)- Principal component 3 accounted for1-26% of the total variance and was most highlycorrelated with mass of the first epibranchial
.("-:-0.44) and length of the teeth on the fifth cerato-./ pranchial (0.27). The fourth principal component
~plained 0.57% of the total variance and no$iable had a correlation with it above 0-19.
The discriminant function analysis on the firstfour principal component factor scores for 40 fishfrom the two lakes found a highly significantoverall difference in pharyngeal jaw morphologybetween the two lakes (MANOVA, Wilks' A = 0-324,F4.35 = 18.26, p < 0-0001). When broken down by
each of the four principal components only thesecond principal component showed a significantunivariate difference between the two lakes(ANOVA, F1.38 = 44-32, p < 0.0001). This relation-
ship was reflected in the correlations between theprincipal components and the canonical factor:PC1 = 0-14, PC2 = 0.75, PC3 = 0.01, PC4 = 0-23.
The discriminant function correctly classified 36out of the 40 fish. The four fish that were incor-rectly classified were Three Lakes fish and wereamong the smallest individuals from that lake (Fig.
5).
49Trophic
polymorphism
inpump---
sunfish
8-;o
.,.c
oc~
=
"' ~
(Q
'"=
~~
~
"' .,
""~.,-,,'-"'
0=
-
.~
c:~
.,
S"~
~-;;
~c:
=
'".~
.,
"' ..
., .,
c: ~
.8 i;'~
~'"
-
-.,'"~., -
oc.'=
~
~~
'" =
~
o~
oc..c:.,
.-
~""
0 =
-.,
""~c:~'"
"'..".,
.~
~..
~o
~
~'--0
'"u~
0
"' ~
"' -
'" o
8 -=
"" .~
.,""8
.,
..""0
.~>"'
.~=
""'"
.,-.,0
~
.~
~oc
OC
"'=0
., .~
-=~~
0 "'
-=~
~...
oc.'Uc:"'-
.~
'" .~
..u '"
'"~=
~u"'
8 '"
c:o
., .~
u=""
"' .~
.,
-" >
---~
'" 0
"' .~
>
., ..=
..~
.~"
0 -
u u
0c:
., =
'".~
~
"'
..-"'"
"' ..
>
'" ~
0 ~
-
u ~
u
'- "'
~0
"' =
"' '"
"'
.,8-"'
'">
- >
- .~
-""~'" o
u=
~
=
<
.~"'
.,,~N
~..
" '"
0
:=~
"'~
~
~
-.-
...C)=
...=
"""
~.:.,;.
0000':":":'
0"'.'..,."'.,.000
666
00=N
o0M
.;.MI
I I
MN
O)
':":":'
.,..,..,..,..".,.000
666
" "
NO
...M
...MI
I I
"'",N'1'~
L('
NN
O)
O)"'~
NN
N
;,;~.~-;; ..
., ..
00 -
Q",
Q
>,-
E.' ..'"'" .-"'
.,.c.c
---~
U...
'" .,
'"Q
~
'"-.c -;;-.c
-..'" U
.-'"
U-.0
.-Q
.c .-Q
'-o.C,"
U.C
,"°ooU
.. Q
U..
.o ,"Q
.o-~
'"
0 ..'"
0-~
..- ~
..-~
...0'" -.0'"
., '"
.-~
E.c~
.' .,~
.,
.,~~
u -5~
ui;J
0
NC
OC
)"'oN...~N
C)
.:.6
"""r-co0066
n",cQ
...0.I
I ,.,?9
C)o
,,"'0066 ~
~M
OC
.j.MI I
...0~
~66 ...0=
06.:.
=.9"iU"3Q
.0Q
..QU..Q
)
S0.t:......S>-
"00.cQ)
S....Q)
,s'0.Q..~.2~......Q
S~
Q)
CC
Q
)0
Q)
.c ..O
Q"0
..Q
) Q
)--U
C
:-g~Q..E
'0 Q
)0
='
:;:~..>~"0
..Q
J:,--
U."'
" 0
--"0
"
0...Q
).:::
.....II
C
Q.
"a.S>
--'-~
.cQ
.0.::"0Q
) 0
",
UC
U-o
~
0 -:;:
.->
C
U
.Q
.-.-Q
) -"0
..t: Q
to 0
C
..
MU
o~...: .~
:;:
..C
..Q
)II
e"3~
Q)..Q
.,""Q
.~
0O
CQ
.~.;-;n~
°.2~
eQ)~
'..-O
)...QU
"0 ~
.Q
.E
Q)
-
..Q)
M
..Q)
Q)
..CC
0
=
..'-0",,-
.E'O
,",~~
0.,00"0N
~V
-;nJ:,II
~
a. "0
.~
~Q
)-.J!."0"0
..0 Q
)
O;.'~
O~
Q)-~
Q.
N
., ca
Q)
.-Q)
U
..-~O
Q..
Q)
<
.-C
.,
~
>
::" .-0
-0
-., .-
" U
.-::J
-"
Z
cn ..
~<
. +
-a:
.;Q)
~0~E,.g~Q)
Q)
'"=~~E"~.5'"Q)
=o.c~.~""iUQ)
00=>-
"'".c~"Q
)
~
.Q~='"~~~"
'0'"Q)
~'".c'"
~Q
)
E.B'"='"l:o0'0.Q~
"' -:S
.{00=-
"..~e0- .
" Q
)'"
'"-'"-U~
.cQ
) U
" '"
Q)
Q)
U
=
=
.-'"
.C
2?'"
'">
.C
o '"
U
>
...OO
U
'" Q
)
Q).c
'"->-
-'"'" .-
=
'"
<
~
..; 8
Q)
>-
-~~
oE
-o~
0!..se
.g"'"'..e>-
"0
o
.Q
.,e.."'
..s0.I::"'
!C.E"'0:o.v;0:.,e
:a .
., .I::
0: "'
0 !C
.Q
0:
"0 .,
., ~
-00U
..
.-.,"0
-
., 0:
~
~
0 ...
o .E
=
.,
..~..-., ..
.I:: >
-"0
."' .,
"' .~
.~
.-"0~
0:
.,
II 6
e ~
-II
.E
>-
~
0 .Q
2j~U
"0
..."0"' .,
., .--
-., -~
0: .-U
->
.-~
'" :a
tC
"' o
O
..:-U
.-"'~
!C
II §"3
gj.,t:~
""
g-.s 8..
j.-0:
~
"' o
o ~
~E
.O!
~..:°~
.' E
-oU
"O-.I::
......,
--0.,.'tC
... "'"
t=oLn
.E
.,
0'- ~
...00.' -
0",6~
~
N.'
0
II~V
-;n -g
'=~
~"O
t5
--., .-
..~
..-"0"'
"' 0
.,
~
-=
0 .-gj
..~
~
~
"' -U
..0
.,
.,<
!C
-.-00 -..
~O>
o:0:~..
e00
U .-"'
~
0..ztl)~
~=
tI)<.
...0.. ~
~

50
P. C.
Waiu
et al.
Discussion
Pumpkinseed sunfish from the two study lakesshowed a distinct polymorphism in the anatomyof the pharyngeal jaw apparatus. In comparison toThree Lakes, fish from snail-poor Wintergreen lakeexhibited an overall reduction in the size of jawstructures; several pharyngeal jaw muscles weresmaller and the three skeletal elements of the jawsinvolved in snail-crushing actions were smaller,both in mass and in linear and areal aspects of bonedimensions. These reductions were coincidentwith the rarity of snails in Wintergreen lake andsuggest that infrequent snail crushing inWintergreen fish has resulted in a general atrophyof the crushing apparatus.
Important evidence in support of this interpre-tation is provided by the absence of polymorphismin the muscles and bones that are not involved inthe snail-crushing behaviour. Standard length,sternohyoideus muscle mass. epibranchiall massand ceratobranchial 1 mass all scaled similarlywith body mass in the two populations. This issignificant because anatomical differencesbetween lakes could be brought about by othereffects of the two environments. The differentdiets and reduced adult growth rates of fish inWintergreen lake (C. W .Osenberg. G.G. Mittelbach& P .C. Wainwright, unpublished observations)could conceivably result in general reductions ofanatomical structures. For example, becauseWintergreen fish rarely eat snails. limited calciumavailability could retard bone mineralization. Infact, of the numerous morphological features thatwere measured in addition to aspects of the ;
crushing apparatus none showed a significant lakeeffect. Only those components known to playacentral role in snail crushing varied between lakes.Previous studies of the functional morphology ofsnail crushing in pumpkinseeds and other perci-form fishes (Lauder, 1983a; Wainwright, 1989a)identified the key skeletal and muscular elementsof snail crushing and make it possible to infer theconsequences and probable causes of the particu-lar morphological differences that were found.Below we discuss the specific anatomical differ-ences between fish in the lakes. their causes andtheir consequences for pharyngeal jaw function.
Muscle morphology
The view that a reduction in the frequency of snailcrushing could lead to the observed morphologicaldifferences is supported by previous research onthe functional morphology of snail crushing in
wright
sunfishes (Lauder, 1983a) and pharyngeal jawmechanisms in other generalized perciform fishes(Wainwright, 1989a,b). The larger pharyngeal jawmuscles of Three Lakes fish can be tied to theirincreased use during snail crushing. Electro-myographic studies of snail crushing in pumpkin-seeds and the other known snail-eatingcentrarchid,L. microlophus, have shown that thisbehaviour involves a phylogenetically derivedpattern of muscle activity in which all pharyngealjaw muscles exhibit simultaneous, long, intensebursts of activity (Lauder. 1983a). The more gen-eralized pharyngeal transport behaviour that isused by pumpkinseeds and other sunfishes whenhandling other types of prey involves a differenttemporal pattern of muscle firing, with smaller.less intense bursts of activity. Our field observa-tions on the feeding rates of pumpkinseeds inThree Lakes suggest that adult fish consume morethan 200 snails in a 14-h summer day (C.W.Osenberg. unpublished observations). The intenseactivity of all pharyngeal jaw muscles during thesefrequent crushing events may exert a trainingeffect (Chapman & Troup. 1970; Ashton & Singh,1974) on these muscles through ontogeny thatresults in hypertrophy relative to the conditionseen in the Wintergreen fish that only rarely crushsnails.
To understand why some muscles that functionto abduct the pharyngeal jaws showed a lake effect(i.e. pharyngocleithralis internus and levatorinternus 3). in addition to muscles that exertcrushing forces. it is important to emphasize thatduring snail crushing all pharyngeal jaw muscles
~ are intensely active (Lauder. 1983a) and. hence, all~muscles are subject to any training effects of.crushing activity. The role of the abducting,,"muscles during snail crushing is presumably tostabilize structures during the forceful adductionof the jaws.
The maximum tension that a muscle candevelop is a combined function of its physio-logical cross-sectional area and the force-produc-ing capability of the muscle tissue per unit ofcross-sectional area (e.g. Calow & Alexander,1973). If muscle shape and the degree of fibrepinnation do not change during the growth of amuscle. then an increase in muscle mass willresult in an increase in the physiological cross-sectional area of the muscle. In general. whencomparing homologous muscles among individ-uals or closely related species. muscle mass hasbeen found to provide an accurate estimate ofrelative muscle strength (Powell et al., 1984;Wainwright. 1988). even if slight changes occur in

muscle fibre angle (Lauder & Reilly. 1991). Thisconclusion is further supported by our data forthe levator posterior. a primary crushing muscle.as both mass and physiological cross-sectionalarea were approximately twice as large in ThreeLakes fish relative to Wintergreen fish. Thus. thismuscle is about twice as strong in Three Lakes fish.and this differentiation was predicted based onmuscle mass. In general. then. the broad-scaledifferences between lakes in the masses of allpharyngeal muscles indicate that Three Lakes fishhave substantially stronger pharyngeal jaws thanfish from Wintergreen lake.
Plasticity or evolution?51
Trophic
polymorphism
in
pumpkinseed
sunfish
Bone morphology
Previous work on the functional morphology ofsnail crushing has indicated that the fifth cerato-branchial, third pharyngobranchial and fourthepibranchial are the primary force-bearingelements during snail crushing in pumpkinseedsand other perciform fishes (Lauder, 1983a; Wain-wright, 1989a). These elements showed thegreatest lake effect in the present study (Tables 2and 3). Frequent crushing actions against resistantsnail shells can be expected to cause these skeletalelements to experience intensified stress-strainregimes. Vertebrate bone is well known to respondto mechanical stresses through increased growthand remodelling (Goodship, Lanyon & McFie,1979; Bouvier & Hylander, 1981; Rubin & Lanyon,1981) and, thus, the more robust bones found inThree Lakes fish probably reflect such a response.
One result that does not fully conform to theexpectations of previous work concerns theresponse seen in the teeth of the lower jaw. Thoughthe large molariform teeth in Three Lakes fish wereshorter than teeth in Wintergreen fish there wereno differences in their thickness (Table 3). Thesimplest explanation for the difference in toothlength is that the shorter teeth in Three Lakes fishare a result of repeated snail-crushing actionswearing down the teeth. This result contrasts withwork on African and South American fishes in thefamily Cichlidae that has repeatedly found thatpharyngeal teeth exhibit strong plasticity in lengthand width, often in response to diet (Liem &Kaufrnan. 1984; Hoogerhoud, 1986; Meyer, 1990).Cichlid teeth can vary between papilliform andmolariform within a single species (e.g. Hooger-houd, 1986). It appears that the morphology ofpumpkinseed teeth is not as readily altered. exceptby wear .
Do the smaller anatomical features seen in pump-kinseeds that occur in the snail-poor environmentof Wintergreen lake simply represent the result ofontogenetic plasticity or has there been geneticdivergence between these two populations?Though examples of polymorphic animal speciesare abundant in the literature (e.g. Bernays, 1986;Liveley, 1986; Ehlinger & Wilson, 1988), mostcases do not distinguish between these twopotential causes of phenotypic separation (forexceptions see Turner et al., 1984; Lavin &McPhail, 1985; Grant, 1986; Trexler & Travis,1990). In the present case the answer to thisquestion is significant in that it determines theextent to which the observed changes can beviewed as a model for evolutionary change infunctional morphology of the feeding mechanism.
Distinctly polymorphic cichlid species that varyin pharyngeal jaw morphology are known fromAfrica (Greenwood, 1965; Hoogerhoud, 1986) andCentral America (Liem & Kaufman, 1984; Meyer,1989, 1990). Laboratory experiments show thattrophic morphology can be altered by rearing fishon different diets (Greenwood, 1965; Hoogerhoud,1986; Meyer,1987) and it appears that these effectscan sometimes be reversed during ontogeny by asecond experimental manipulation of diet (Meyer,1987). Though the issue of how morphologicaltypes are determined has not been entirelydetermined for any cichlid species, there is muchevidence that phenotypic plasticity plays an
.important role (Greenwood, 1965; Hoogerhoud,ji986; Meyer, 1987) and relatively less evidence..that morphological types are geneticallydetermined (e.g. Sage & Selander, 1975; Kornfieldttt al., 1982).
We are unable to definitely determine the rela-tive importance of phenotypic plasticity and gene-tic divergence in causing the trophicpolymorphism in pumpkinseeds. Evidenceregarding the contributions of the two factorscould be obtained in common garden experiments(e.g. Trexler & Travis, 1990) in which cohorts fromeach population could be reared in two groups,one going into a snail-rich environment and theother into a snail-poor environment. If fish fromthe two populations develop similar morphologi-cal patterns growing in each environment then thedifferences seen between lakes were due todevelopmental plasticity. Genetic divergencewould be implicated if fish from the two lakesdeveloped differently in the commonenvironments.
EE
.c--0
.i
I()
a:c:uc
e.co
+-0..Q)u0-
..2
o
oooeoA
o~.\A
52
PoCo
Wain
et alo
0!9
0000
04.4.4. 4.
4.
A
6 6O~
0.0 0.5 1.0 1.5
log body mass (g)
2.0
"'EE
o0>'-0
.0
00.5 tRIo
° ~~:a° A
O?A° A
A
.5
q-O 0.0Euco
:0 -0.50.Q)0-..2 -
r-. A
O
"'EE
o0)'-0
1'")"0:2 1.0uco.B8. 0.5c>-o-a. 0.0 I. ..
0.0 0.5 1.0 1.5 2.0 01 0.0 0.5 1.0 1.5 2.0.9.
log body mass (g ) log body mass (g )
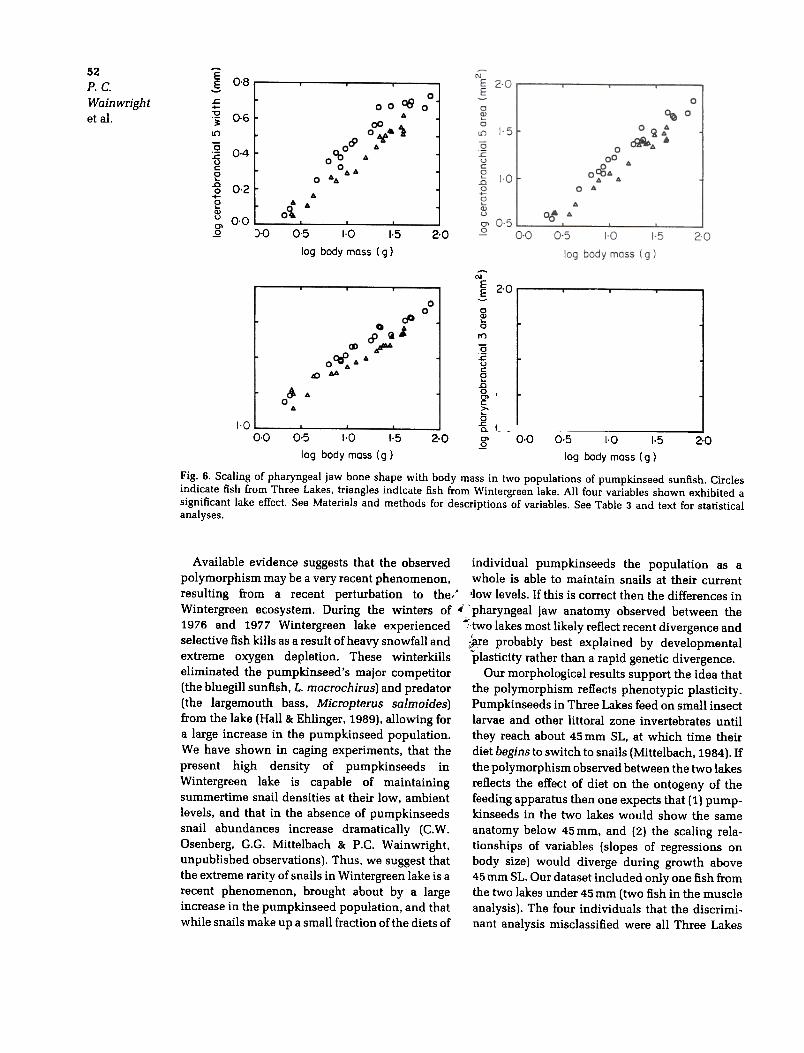
Fig. 6. Scaling of pharyngeal jaw bone shape with body mass in two populations of pumpkinseed sunfish. Circlesindicate fish from Three Lakes. triangles indicate fish from Wintergreen lake. All four variables shown exhibited asignificant lake effect. See Materials and methods for descriptions of variables. See Table 3 and text for statistical
analyses.
Available evidence suggests that the observed individual pumpkinseeds the population as apolymorphism may be a very recent phenomenon. whole is able to maintain snails at their currentresulting from a recent perturbation to the,'. 1ow levels. If this is correct then the differences inWintergreen ecosystem. During the winters of ~..pharyngeal jaw anatomy observed between the1976 and 1977 Wintergreen lake experienced ~two lakes most likely reflect recent divergence and
Iselective fish kills as a result of heavy snowfall and ;~e probably best explained by developmentalextreme oxygen depletion. These winterkills plasticity rather than a rapid genetic divergence.eliminated the pumpkinseed's major competitor Our morphological results support the idea that(the bluegill sunfish. L. macrochirus) and predator the polymorphism reflects phenotypic plasticity .(the largemouth bass. Micropterus salmoides) Pumpkinseeds in Three Lakes feed on small insectfrom the lake (Hall & Ehlinger. 1989). allowing for larvae and other littoral zone invertebrates untila large increase in the pumpkinseed population. they reach about 45 mm SL, at which time theirWe have shown in caging experiments. that the diet begins to switch to snails (Mittelbach, 1984). Ifpresent high density of pumpkinseeds in the polymorphism observed between the two lakesWintergreen lake is capable of maintaining reflects the effect of diet on the ontogeny of thesummertime snail densities at their low. ambient feeding apparatus then one expects that (1) pump-levels. and that in the absence of pumpkinseeds kinseeds in the two lakes would show the samesnail abundances increase dramatically (C.W. anatomy below 45mm. and (2) the scaling rela-Osenberg, G.G. Mittelbach & P.C. Wainwright, tionships of variables (slopes of regressions onunpublished observations). Thus, we suggest that body size) would diverge during growth abovethe extreme rarity of snails in Wintergreen lake is a 45 mm SL. Our dataset included only one fish fromrecent phenomenon. brought about by a large the two lakes under 45 mm (two fish in the muscleincrease in the pumpkinseed population. and that analysis). The four individuals that the discrimi-while snails make up a small fraction of the diets of nant analysis misclassified were all Three Lakes
0.8
0.6
0.4
0.2
0.0
wright

53
Trophic
polymorphism
in
pumpkinseed
sunfish
fish and were four of the five smallest fish exam-ined from that lake (Fig. 5; 40, 45, 64 and 65mm).This poor multivariate discrimination of pharyn-geal jaw structures in small fish supports the viewthat small fish in the two populations are similarand only diverge during the period of ontogenywhen snails are being eaten. However, we note thatwhen they were considered separately, none of themorphological variables showed heterogeneity ofslopes between lakes. Though the bivariate plotsvisually suggest greater similarity at small bodysizes (Figs. 3, 4 and 6) only the multivariateanalysis indicated that this was a significant trendthat obscures discrimination of fish that are lessthan about 65 mm. Below, we provide anadditional argument, based on the non-linearscaling of two muscles, that further corroboratesour interpretation that divergence is coincidentwith the diet shift to snails.
Differences between lakes in the ontogeneticscaling patterns of the levator posterior and retrac-tor dorsalis muscles suggest that the pattern ofgrowth in these muscles is related to the inclusionof snails in the diet. A significant change in the rateof growth of both muscles was found in fish fromThree Lakes but not in fish from Wintergreen lake(Fig. 3). The change in the rate of levator posteriorgrowth occurred at 17.4g body mass (83mm SL).The scaling exponent of this muscle in fish smallerthan this was 1.68, and 0.64 in fish larger than83 mm. Thus, levator posterior growth rate slowsby about two-thirds across this critical body size; astriking and significant change. As discussedabove, previous work has shown that pumpkin-seeds in Three Lakes begin including snails intheir diet at about 45 mm SL, about the minimumsize examined in this study, and increase theaverage amount of snails in the diet until this isnearly the only prey type eaten at about 80mm SL(Mittelbach, 1984; Osenberg, 1988). Thus, there isa strong coincidence in the body size at whichpumpkinseeds achieve a diet of nearly 100%snails and when the growth rates of the levatorposterior and retractor dorsalis slows. Further-more, it appears that by the time a pumpkinseedreaches 80mm it is capable of feeding on most, ifnot all of the available snails (Osenberg & Mittel-bach, 1989). The mean size of snails in the diets ofpumpkinseeds increases much between 45 and80mm SL, but exhibits little change after thistransition (Mittelbach, 1984; Osenberg & Mittel-bach, 1989). We suggest that the high growth ratesof the levator posterior and retractor dorsalismuscles below about 80mm SL reflect the periodof ontogeny when the diet is changing. As more
and larger snails are added to the diet the intensityof any training effect on the crushing musclesincreases but once pumpkinseeds achieve a dietdominated by snails and are able to crush mostavailable snails, the rate of growth in thesemuscles slows tremendously.
The pharyngeal jaw polymorphism reportedhere for pumpkinseeds seems to be a direct resultof differences in patterns of use of the pharyngealjaws during prey handling behaviour. While theneed to invoke a genetic basis for this polymor-phism is in doubt, this system provides a model forthe process of evolutionary change in a complexfunctional network. We observed no morphologi-cal differences that did not appear to be directlylinked to changes in the use of the pharyngealjaws. Thus, we found no evidence of other, indi-rect changes in the skeletal structures of thefeeding apparatus not used during snail crushing.The response of the feeding apparatus in theWintergreen population has been quite specific,with no evidence of complex, indirect effects ofthe diet shift (i.e. no lake effects were seen instructures not involved in snail crushing).However, we emphasize that our quantitativeobservations on morphology are mostly limited toseveral skeletal elements and muscles of thepharyngeal jaw apparatus. It is conceivable thatchanges have occurred in other parts of the head,as has been suggested for some cichlids (Barel,1983). If present, however, these changes must besmall as none were apparent in our observations ofdissected fish. In general, the extent of morpholo-gical variation seen among populations was not as
i extensive as has been found in cichlids (Hooger-
4'houd, 1986; Meyer, 1989, 1990), perhaps indicat-~g a fundamental difference in the capacity foranatomical plasticity between the phylogenetic
groups.Our results parallel the findings of comparative
studies of the functional basis of dietary diversifi-cation among species within various fish groups.Recent analyses have almost always found mor-phological transformations of the feeding appara-tus underlying trophic switches (Barel, 1983;Motta, 1988; Wainwright, 1988). Another level ofdesign of the feeding mechanism that is oftencontrasted among species differing in diet is pat-terns of muscle activity used during prey captureor prey handling (Liem, 1979; Lauder, 1983a;Wainwright, 1989b). Motor patterns haverepeatedly been found to be much more conserva-tive than morphology during the evolution oftrophic habits (e.g. Sanderson, 1988; Wainwright& Lauder,1991). A key goal in future work with the

pumpkinseed polymorphism will be to test the
generality of motor pattern conservatism at thislevel of comparison among populations.
Acknowledgments
We thank G.V. Lauder, S.M. Reilly, two anony-mous reviewers, and especially C.D.N. Barel for
insightful comments on an earlier draft of this
paper. Laboratory facilities were generously pro-vided by G. V. Lauder. Support for this research
was provided by NSF grants BSR 87-96261 to
G.G.M., BSR 89-05867 to G.G.M. and C. W .0., and
DCB 87 -10210 to G. V. Lauder. This is contribution
number 683 from the Kellogg Biological Station.
References
Ashton. T.E.J. & Singh. M. (1974) The effect of training onmaximal isometric back.lift strength and mean peakvoltage of the erector spinae. In Biomechanics IV (eds.R.C. Nelson & C.A. Morehouse), pp. 448-452. Uni-versity Park Press. Baltimore.
Barel. C.D.N. (1983) Towards a constructional morpho-logy of cichlid fishes (Teleostei, Perciformes). Nether-lands Journal of Zoology, 33, 357-424.
Bernays, E.A. (1986) Diet-induced head allometry amongfoliage-chewing insects and its importance for grami-nivores. Science. 231, 495-497.
Bookstein, F.. Chernoff, B.. Elder. R.. Humphries, J..Smith. G. & Strauss. R. (1985) Morphometrics inEvolutionary Biology. Special Publication number 15of the Academy of Natural Sciences of Philadelphia.
Bouvier, M. & Hylander. W.L. (1981) Effects of bonestrain on cortical bone structure in macaques. JournalofMorphology, 167, 1-12. .
Calow. L.J. & Alexander. R.M. (1973) A mechanical;analysis of a hind leg of a frog (Rana temporaria). .
Journal qfZoology, 171, 293-321.Carl, L.M. & Healey. M.C. (1984) Differences in enzyme
frequency and body morphology among three juvenilelife history types of the chinook salmon ( Oncorynch ustshawytscha) in the Nanaimo river, British Columbia.Canadian Journal of Fisheries and Aquatic Sciences.41,1070-1077.
Chapman. A.E. & Troup. J.D.G. (1970) Prolonged activityof lumbar erector spinae. An electromyographic anddynamometric study of the effect of training. Annals ofPhysiological Medicine. 10, 262-269.
Collins. J.P. & Cheek, J.E. (1983) Effect of food anddensity on development of typical and cannabilisticsalamander larvae in Ambystoma tigrinum nebulo-sum. American Zoologist. 23, 77-84.
Dingerkus. G. & Uhler, L.D. (1977) Enzyme clearing ofalcian blue stained whole vertebrates for demon-stration of cartilage. Stain Technology. 52, 229-232.
Ehlinger, T.J. & Wilson. D.S. (1988) A complex foragingpolymorphism in bluegill sunfish. Proceedings of theNational Academy of Sciences. 85, 1878-1882.
Goodship, A.E.. Lanyon, L.E. & McFie. H. (1979) Func-tional adaptation of bone to increased stress. Journal ofBone and Joint Surgery, 61A, 539-546.
54
P. C.
Wainwrightet al.
Grant. P.R. (1986) Ecology and Evolution of Darwin'sFinches. Princeton University Press. Princeton.
Greenwood. P.H. (1965) Environmental effects on thepharyngeal mill of a cichlid fish. Astatoreochromisalluadi. and their taxonomic implications. Proceed-ings of the Linnean Society of London, 176, 1-10.
Hall. D.J. & Ehlinger. T.J. (1989) Perturbation. plank-tivory and pelagic community structure: the conse-quence of winterkill in a small lake. Canadian Journalof Fisheries and Aquatic Sciences. 46, 2203-2209.
Hoogerhoud. R.J.C. (1986) The ecological and taxonomicaspects of morphological plasticity in molluscivoroushaplochromines. Annales. Musee Royal de l'AfriqueCentrole. Zoologie. 251, 131-134.
Keast. A. (1978) Trophic and spatial interrelationships inthe fish species of an Ontario temperate lake. Environ-mental Biology of Fishes. 3, 7-31.
Kornfield. I.L.. Smith. C.D.. Gagnon. P.S. & Taylor. J.N.(1982) The cichlid fishes of Cuatro Cienegas. Mexico:direct evidence of conspecificity among distinctmorphs. Evolution. 36, 658-664.
Lauder. G. V. (1983a) Functional and morphologicalbases of trophic specialization in sunfishes (Teleostei.Centrarchidae).lournal of Morphology, 178, 1-21.
Lauder. G.V. (1983b) Neuromuscular patterns and theorigin of trophic specialization in fishes. Science, 219,1235-1237.
Lauder, G. V. (1985) Aquatic feeding in lower vertebrates.In Functional Vertebrate Morphology (eds. M. Hilde-brand. D.M. Bramble. K.F. Liem & D.B. Wake). pp.210-229. The Belknap Press, Cambridge.
Lauder. G.V. (1986) Homology. analogy and the evolu-tion of behavior. In Evolution of Animal Behavior(eds. M.H. Nitecki & J.A. Kitchell). pp. 9-40. OxfordUniversity Press. New York.
Lauder. G.V. & Reilly. S.M. (1991) Metamorphosis of thefeeding mechanism in tiger salamanders (Ambystomatigrinum). Journal of Zoology. in press.
Lavin. P.A. & McPhail. J.D. (1985) The evolution of, freshwater diversity in the threespine stickleback
i' .(Gasterosteus aculeatus). Canadian Journal of Zoo-' logy, 63, 2632-2638.
~ Liem. K.F. (1979) Modulatory multiplicity in the feeding.' mechanism of the cichlids, as exemplified by the~: invertebrate pickers of Lake Tanganyika. Journal of
Zoology, 189. 93-125.Liem. K.F. & Kaufman, L.S. (1984) Intraspecific macro-
evolution: functional biology of the polymorphiccichlid species Cichlasoma minckleyi. In Evolution ofSpecies Flocks (eds. A.A. Echelle & I. Kornfield). pp.203-215. University of Maine Press. Orono.
Lindsey. C.C. (1981) Stocks are chameleons: plasticity ingill rakes of coregonid fishes. Canadian Journal ofFisheries and Aquatic Sciences, 38. 1497-1506.
Liveley. C.M. (1986) Predator-induced shell dimorphismin the acorn barnacle. Evolution, 40, 232-242.
Lowndes. A.G. (1955) Density of fishes. Some notes onthe swimming of fish to be correlated with density.sinking factor and the load carried. Annals Magazineof Natural History, 8,241-256.
Magnan. P. (1988) Interactions between brook char.Salvelinus fontinalis, and nonsalmonid species: eco-logical shift. morphological shift. and their impact onzooplankton communities. Canadian Journal ofFisheries and Aquatic Sciences, 45. 999-1009.
Meyer. A. (1987) Phenotypic plasticity and heterochrony

55
Trophic
polymorphism
in
pumpkinseed
sunfish
Sage. R.D. & Selander. R.K. (1975) Trophic radiationthrough polymorphism in cichlid fishes. Proceedingsof the National Academy of Sciences, 72,4669-4673.
Sanderson. S.L. (1988) Variation in neuromuscular act-ivity during prey capture by trophic specialists andgeneralists (Pisces: Labridae). Brain, Behavior, andEvolution, 32, 257-268.
Snedecor. C.W. &Cochran, W.C. (1980) Statistical Meth-ods. 7th edn. Iowa State University Press. Ames. Iowa.
Trexler. I.C. & Travis. I. (1990) Phenotypic plasticity inthe sail fin molly. Poecilia latipinna (Pisces:Poeciliidae). I. Field experiments. Evolution, 44. 143-156.
Turner. B.I.. Crudzien. T.A.. Adkisson. K.P. & White.M.M. (1984) Evolutionary genetics of trophic differen-tiation in Coodied fishes of the genus Ilyodon.Environmental Biology of Fishes, 9,159-172.
Wainwright. P.C. (1988) Morphology and ecology: func-tional basis of feeding constraints in Caribbean labridfishes. Ecology, 69, 635-645.
Wainwright. P.C. (1989a) Functional morphology of thepharyngeal jaw apparatus in perciform fishes: anexperimental analysis of the Haemulidae. Journal ofMorphology, 200, 231-245.
Wainwright. P.C. (1989b) Prey processing in haemulidfishes: patterns of variation in pharyngeal jaw muscleactivity. Journal of Experimental Biology. 141, 359-375.
Wainwright. P.C. & Lauder. C.V. (1991) The evolution offeeding biology in sunfishes. In Systematics, His-torical Ecology, and North American FreshwaterFishes (ed. R.L. Mayden). Stanford Press. Stanford. in
press.Werner. E.E. & Hall. D.I. (1979) Foraging efficiency and
habitat switching in competing sunfishes. Ecology, 60,256-264.
Wilkinson. L. (1986) Systat: The System for Statistics.Systat Inc.. Evanston.
Yeager. D.P. & Ultsch, C.R. (1989) Physiological regula-tion and conformation: a BASIC program for the deter-
.mination of critical points. Physiological Zoology, 62,888-907.
in Cichlasoma managuense (Pisces, Cichlidae) andtheir implications for speciation in cichlid fishes.Evolution. 41, 1357-1369.
Meyer, A. (1989) Cost of morphological specialization:feeding performance of the two morphs in the trophi-cally polymorphic cichlid fish, Cichlasomacitrinellum. Oecologia, 80, 431-436.
Meyer, A. (1990) Ecological and evolutionary aspects ofthe trophic polymorphism in Cichlasoma citrinellum(Pisces, Cichlidae). Biological Journal of the LinneanSociety. 39, 279-299.
Mittelbach, G.G. (1984) Predation and resource par-titioning in two sunfishes (Centrarchidae). Ecology,65, 499-513.
Mittelbach, G.G. (1988) Competition among refugingsunfishes and effects of fish density on littoral zonesinvertebrates. Ecology, 69, 614-623.
Motta, P.J. (1988) Functional morphology of feeding inbutterfly fishes (perciformes, Chaetodontidae): an eco-morphological approach. Environmental Biology ofFishes, 22, 39-67.
Nickerson, D.M., Facey, D.E. & Grossman, G.D. (1989)Estimating physiological thresholds with continuoustwo-phase regression. Physiological Zoology, 62, 866-887.
Osenberg, C. W. (1988) Body size and the interaction offish predation and food limitation in a freshwatersnail community. PhD dissertation, Michigan StateUniversity, East Lansing, Michigan, USA.
Osenberg, C. W. & Mittelbach, G.G. (1989) Effects ofbodysize on the predator-prey interaction between pump-kinseed sunfish and gastropods. Ecological Mono-graphs. 59, 405-432.
Powell, P.L., Roy, R.R., Kanim, P., Bello, M.A. & Edger-ton, V. (1984) Predictability of skeletal muscle tensionfrom architectural determinations in guinea pighindlimbs. Journal of Applied Physiology, 57, 1715-1721.
Rubin, C.T. & Lanyon, L.E. (1981) Bone remodelling inresponse to applied dynamic loads. Journal of Boneand Joint Surgery. 5, 237-238. ,
Sadzikowski, M.R. & Wallace, D.C. (1976) A comparison .iof the food habits of size classes of three sunfishes(Lepomis macrochirus, L. gibbosus and L. cyanellus).American Midland Naturalist. 95, 220-225.
.;Received 22 January 1990; revised 9 April 1990;'DcceDted 18 A Dril1990





























