Embed Size (px)
Citation preview

Frustration and Adenylate Cyclase
Michael E. Maguire, Thomas W. Sturgill, and Alfred G. Gilman
M UCH OF THE work from this laboratory during the past few years has involved attempts to explore mechanisms that control cyclic AMP
synthesis in intact cells cultured in vitro. Our hope is that a combination of pharmacologic, biochemical, and genetic manipulations of somatic cells with specific properties and hybrid cells derived therefrom will facilitate our under- standing of this complex system. Progress is slow, and it appears too early to decide if observations of a truly fundamental nature will be secured. We have reviewed certain of our efforts recently.’
Since our basic goal is obviously to understand the nature of the components that are essential to this system, we attempt to observe closely the progress of others who are following perhaps more reasonable paths-dissection of the sys- tem by examination of kinetic properties or purification and reconstitution of components. The invitation to write a critical review for this issue of Metabo- lism has provided us with an opportunity that we are apparently too foolish to pass by; we have therefore decided to discuss selected aspects of the subject of adenylate cyclase. The past few years have witnessed an extraordinary amount of information gathering on this subject, and we review it primarily with the goal of the clarification of our own thinking. It should be noted that, with the cerebral equipment available to us, this goal is left almost entirely unachieved.
If this prolix introduction sounds pessimistic, it is intentional. While the past two decades have demonstrated the presence of adenylate cyclase in a myriad of cell types, remarkably few fundamental observations have been made on this enzyme system since its elementary properties were described by Sutherland and Rall and their associates between 1957 and 1962. After over a decade of work, Robison, Butcher, and Sutherland’ observed:
“One of the reasons for our lack of understanding of the relation between receptors and adenyl cyclase derives from our lack of understanding of the nature of adenyl cyclase itself. Adenyl cyclase from mammalian sources has so far resisted all attempts to purify it extensively, and indeed it is not even clear that it represents a single enzyme . . . it may be best to refer to adenyl cyclase not as an enzyme but as an enzyme system, and subsequent references to ‘adenyl cyclase’ in this essay should be considered with this meaning in mind.” Several years later, this statement remains entirely appropriate. Virtually no
model has, we feel, been excluded. The system may possess the relative simplic- ity of a monomeric enzyme; more likely it is far more complex, and we must
From the Department of Pharmacology, University of Virginia School of Medicine, Charlottes-
ville, Va.
Receivedfor publication October II, 1974.
Supported in part by USPHS grants NS-10193 and GM 02139.
Reprint requests should be addressed to Dr. M. E. Maguire. Department of Pharmacology, Univer-
sity of Virginia School of Medicine, Charlottesville, Va. 22903.
o 1975 by Grune & Stratton, Inc.
Metabolism, Vol. 24, No. 3 (March), 1975 287

288 MAGUIRE, STURGILL, AND GILMAN
consider dimeric hormone receptor-catalytic unit models, multiple-subunit
models, and multienzyme systems. The components of the latter may include both soluble and membrane-bound elements. This question, conceptually simple, begs for an answer: of what order of complexity is the adenylate
cyclase system? We will attempt to keep this question in view as we discuss two aspects of the adenylate cyclase system-the action of guanine nucleotides and
that of fluoride.
GUANINE NUCLEOTIDES AND THE ADENYLATE CYCLASE SYSTEM
It is logical that adenylate cyclase should be regulated from within the cell as well as from without, although it is not that easy to cite examples of this logic. The ability of the system to become refractory to hormonal stimulation might represent such a phenomenon, as may the fact that certain hormones may cause relatively stable increases in adenylate cyclase activity. Since hor-
monal activation of this second messenger system represents the initial step in
a series of reactions that ultimately alters the performance of the cell, there may well be feedback mechanisms whereby the “slaves” can overrule the apparent
“master.” Numerous stimulatory and inhibitory effects of guanine nucleotides
have been described on adenylate cyclase activity and on the ability of hor- mones to stimulate this catalytic activity. Another fundamental question must also not drop from view. Do guanine nucleotides play a functional role in vivo
to regulate the activity of adenylate cyclase? Some consideration of guanine nucleotide concentrations is necessary. Cel-
lular concentrations of GTP are on the order of 0.1 mM, whereas in vitro ef-
fects of this nucleotide on the cyclase can be demonstrated, in many cases, at concentrations of O.Ol- 1 PM. This naively implies that the system would always be saturated in vivo, and saturation of the system seems inconsistent with
regulation. While saturation is consistent with a permissive effect, one may
question the probability of the preservation of such a role through evolution. If this inconsistency is not artifactual (resulting, for example, from an inabil-
ity to reproduce in vivo conditions in standard assays), the rationalizations must involve compartmentalization of guanine nucleotides or alterations of the
apparent affinity of these compounds by unknown factors operating on the adenylate cyclase system.
Another difficulty in postulating a regulatory role for guanine nucleotides is encountered when one considers the concentrations of ATP that prevail in vivo. Under these conditions, effects of guanine nucleotides are usually less than
dramatic, hypothetically because ATP can interact at the guanine nucleotide
binding site (see below). If answers to these questions are found, control of the system might then be
in the hands of reactions that regulate guanine nucleotide concentrations, per- haps particularly those involved in de novo synthesis. Alternatively, fluctua- tions in the relative concentrations of mono-, di-, and triphosphates could as- sume importance, but these questions are currently beyond our ability to pose them.
Regardless of possible significance in vivo, the effects of guanine nucleotides observed in vitro do demonstrate the existence of a modulatory site. Although

FRUSTRATION AND ADENYLATE CYCLASE 289
many investigators have assumed that this site is on the catalytic unit of the
system, it is currently completely impossible to specify this. An example of data that may reasonably be interpreted to suggest that guanine nucleotides
may act on a separate unit of the adenylate cyclase system includes an apparent dissociation of both guanine nucleotide and catecholamine activation from
cyclase catalytic activity, using the polyene antibiotic filipin.3 At least one possible explanation of the data presented is that filipin disrupts the interaction
between the catalytic unit and a unit binding guanine nucleotides. The latter
could be a regulatory subunit in an oligomeric allosteric adenylate cyclase sys-
tem, a separate hormone-binding subunit, or even another enzyme in a multi- enzyme complex. Alternatively, if the regulatory and catalytic sites are present
on the same polypeptide chain, an environmental change such as that produced by a polyene or a detergent could conceivably mask the modulatory effect. The location of the site will clearly require separation and reconstitution of the in-
dividual components. A minimal description of the adenylate cyclase system requires three inter-
acting sites, one each for hormone, guanine nucleotide, and substrate. Al- though no final proof has been obtained for any hormone capable of stimulat-
ing the cyclase system, it is usually assumed that the substrate and hormone
sites reside on different polypeptide chains, rather than on an integral protein
spanning the membrane. However, the evidence for this is as weak as that men- tioned above for guanine nucleotides, and, furthermore, some “hormones”
(e.g., prostaglandins) clearly have the physical properties necessary for pene- tration to more internal sites of action.
It has seemed possible that guanine nucleotides could act as permissive
agents4 by binding to a regulatory site and allowing interaction between those sites specific for hormone and substrate. This may call to mind roles for gua-
nine nucleotides in other phenomena involving translocation-on the ribosome and in hypothetical actions of microtubules. As a permissive agent, it is not necessary that the nucleotides affect either hormone or substrate binding; they
simply must be present for hormone receptor-cyclase interaction to be produc-
tive. However, the observations that guanine nucleotides themselves activate the cyclase system and that hormones may stimulate activity in the absence of
a guanine or other nucleotide bound to this hypothetical site seem incompat- ible with this hypothesis.
Since adenylate cyclase is a regulatory enzyme, it is reasonable to expect that the system should be at least of the order of complexity of other regulatory enzymes; the example of aspartate transcarbamylase has been used to advan- tage in the past. Most of these systems are allosteric* in nature, and this has
been suggested for adenylate cyclase.4 An enzyme system of this level of com-
plexity (i.e., multiple ligand-mediated states) is capable of adequately describ-
*Allosteric effects refer simply to indirect interactions between distinct ligand-binding sites. A
possible molecular mechanism is by ligand-linked conformational transitions, e.g., the Monod-
Changeux-Wyman or Koshland models. An allosteric enzyme here refers to an enzyme whose
activity is subject to thermodynamic control by virtue of existing in multiple (2 2) ligand-
linked thermodynamic states.

290 MAGUIRE, STURGILL, AND GILMAN
ing the observed interactions with hormones, guanine nucleotides, and sub- strate. It is appropriate to consider, therefore, the extent to which available data is compatible with a model of an allosteric enzyme.
In an allosteric system, the binding of one ligand influences the binding of another. GTP has been shown to affect the binding of glucagon, although it has not been proven that this binding was to the hormonal receptor.* If guanine
nucleotides are to be considered as heterotropic allosteric ligands, it must be
possible to show that glucagon also affects GTP binding. A minimum thermo- dynamic description of the adenylate cyclase system requires that at least eight states of the system be considered:
ES - E - EH - EHS
t $ $ $ EGS - EG - EGH - EGHS
where E, G, H, and S represent enzyme, regulatory nucleotide, hormone, and
substrate, respectively.?
Since these states are assumed to be in equilibrium with each other, there must be a reciprocal effect, if the binding of one ligand affects the binding of
another. To our knowledge, such reciprocal interactions have not been demon- strated. While it may not be feasible to study specifically GTP binding to the appropriate site, it should be possible to determine the apparent K, for GTP activation at several different glucagon concentrations, as a first approximation of the data required.
There are anomalies in the literature that require explanation. In a study of the effect of GTP on glucagon binding, the nucleotide was shown to increase the rate of dissociation of the hormone and to decrease the amount bound at
equilibrium; there is thus a decrease in the binding constant for glucagon.’
However, kinetic data demonstrated that GTP decreased the K, for glucagon activation of the cyclase,6 implying an increased binding affinity. Although the
K, for glucagon activation is not equivalent to its dissociation constant, it is surprising that the effects should be opposite in direction.
Well-studied allosteric regulatory enzymes show homotropic effects for at least one ligand. While not a necessary phenomenon, this has not yet been demonstrated for any ligand thought to interact with the adenylate cyclase sys-
tern. For the most part, appropriate studies have not been designed to test this
possibility, and, when acquired, such data have not always been fully analyzed. An example of a phenomenon that has been interpreted as a type of homotropic
effect should be discussed. When adenylate cyclase activity is determined at various substrate concentra-
tions in the presence and absence of activators, it is possible to plot a ratio of activity in the presence of a maximally effective concentration of the stimula- tory agent to the basal activity versus substrate concentration. In hepatic plasma membranes, the ratio of fluoride-stimulated activity to basal activity
tThe equilibria shown were arbitrarily chosen and do not imply restrictive reaction pathways
for interaction between the species. All other possible equilibria between the species shown can
be expressed in terms of the constants for the equilibria chosen.

FRUSTRATION AND ADENYLATE CYCLASE 291
appears to be independent of substrate. The ratio in the presence of glucagon, however, gives a sigmoidal curve with increasing substrate concentration.’ The authors interpreted these results in light of their demonstration that gua- nine nucleotides modulate glucagon-stimulated adenylate cyclase activity, Thus, it was proposed that at sufficiently high concentrations, ATP could bind to a GTP site, and that ATP could imitate the effect of GTP to permit activa- tion of the enzyme by glucagon.’ Such an effect of ATP would constitute a type of homotropic effect.
While we feel that this interpretation is reasonable, we would like to show that these data are consistent with other simple kinetic considerations that do not assume an interaction of ATP at a guanine nucleotide binding site.
The cyclase activity is assumed to be governed by classic Michaelis-Menten kinetics, with the velocity, v, of the unstimulated cyclase given by
PI v = v.- PI + K
where V is the maximal velocity at saturating substrate concentration, [S], and K is the apparent Michaelis constant. Likewise, the velocity, v’, of the stimu- lated (hormone or fluoride) cyclase is
v’ = V’. [Sl [Sl + K’
The data are then plotted as the ratio v’/v versus [S]:
V’ V’ [S] + K - _ -. V V [Sl + K’
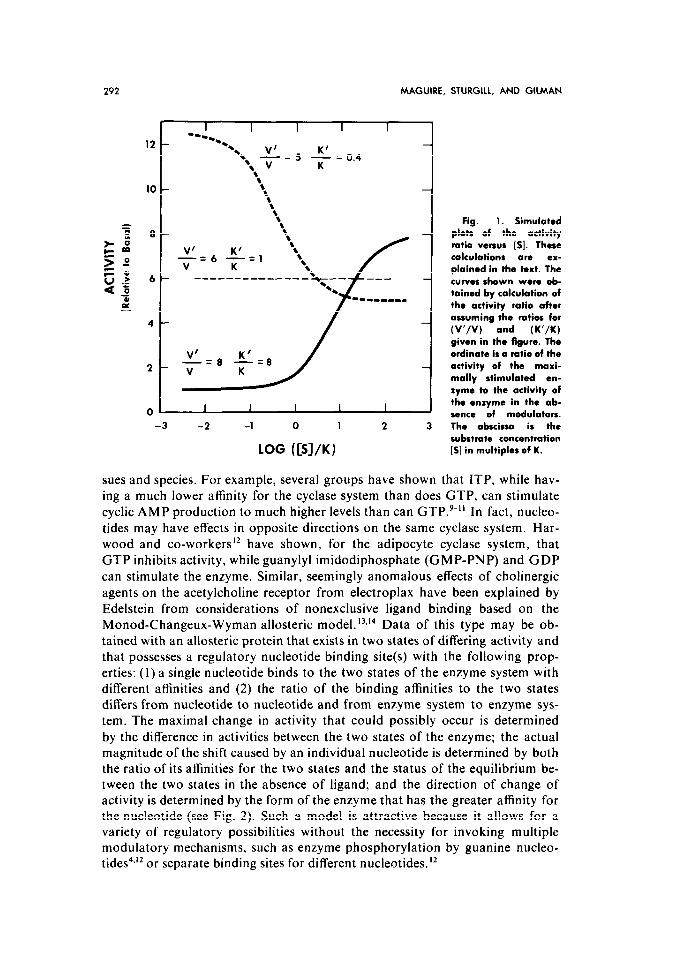
It is apparent that a plot of the activity ratio versus [S] is dependent on the ratios V’/V and K’/K, as shown in Fig. 1. If the stimulatory agent increases maximal velocity (V’ > V) and also increases substrate affinity (K’ < K), a de- crease in the activity ratio with increasing [S] is observed.
If K = K’, the activity ratio at any substrate concentration is logically given by VI/V, and the plot is a line parallel to the abscissa.
Finally, if the maximal velocity in the presence of the perturbing agent is increased (V’ > V) while the apparent substrate affinity is decreased (K’ > K), a plot of the activity ratio versus [S] generates the sigmoidal curve given by the solid line in Fig. 1. The simulated data obtained from these simple kinetic con- siderations are of exactly the same form as the published data cited,’ and there is no apparent need to invoke a regulatory site in order to explain the obser- vations.
We must emphasize that an analysis such as this does not (and cannot) show
that the GTP site is incapable of binding ATP. Indeed, Bockaert and associates’ have presented data that they correctly conclude cannot be adequately ex- plained by the type of analysis shown above.
If it is assumed that the cyclase system is an allosteric one, it becomes pos- sible to explain the varied and confusing effects of different guanine nucleotides (and other nucleotides) on hormonally stimulated cyclases from multiple tis-
292 MAGUIRE, STURGILL, AND GILMAN
% 8
10 - 8 8 8 8 8 8 8
8- 8 8
-_=(j -_=I
-3 -2 -1 0 1 2 3
LOG (CSl/K)
Fig. 1. Simulated plots of the activity ratio versus [S]. These calculations are ex- plained in the text. The curves shown were ob- tained by calculation of the activity ratio after assuming the ratios for (V’/V) and (K’/K)
given in the figure. The ordinate is a ratio of the activity of the maxi- mally stimulated en- zyme to the activity of the enzyme in the ab- sonce of modulators. The abscissa is the substrate concentration [S] in multiples of K.
sues and species. For example, several groups have shown that ITP, while hav-
ing a much lower affinity for the cyclase system than does GTP, can stimulate cyclic AMP production to much higher levels than can GTP.‘-” In fact, nucleo- tides may have effects in opposite directions on the same cyclase system. Har-
wood and co-workers I2 have shown, for the adipocyte cyclase system, that GTP inhibits activity, while guanylyl imidodiphosphate (GMP-PNP) and GDP
can stimulate the enzyme. Similar, seemingly anomalous effects of cholinergic
agents on the acetylcholine receptor from electroplax have been explained by
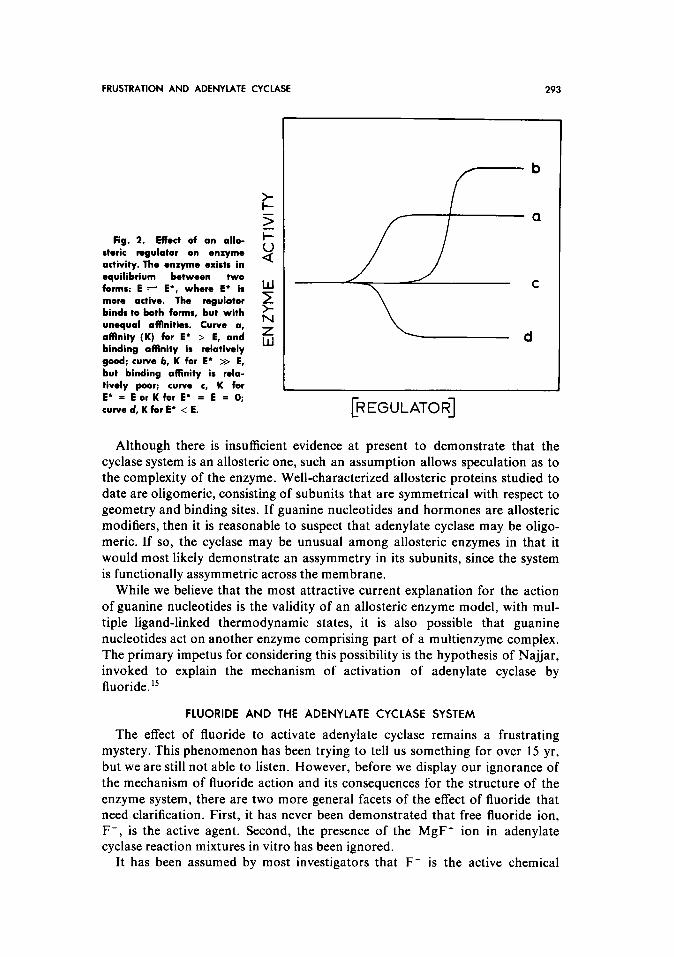
Edelstein from considerations of nonexclusive ligand binding based on the Monod-Changeux-Wyman allosteric model.‘3*‘4 Data of this type may be ob- tained with an allosteric protein that exists in two states of differing activity and that possesses a regulatory nucleotide binding site(s) with the following prop- erties: (1) a single nucleotide binds to the two states of the enzyme system with different affinities and (2) the ratio of the binding affinities to the two states differs from nucleotide to nucleotide and from enzyme system to enzyme sys- tem. The maximal change in activity that could possibly occur is determined
by the difference in activities between the two states of the enzyme; the actual magnitude of the shift caused by an individual nucleotide is determined by both the ratio of its affinities for the two states and the status of the equilibrium be- tween the two states in the absence of ligand; and the direction of change of activity is determined by the form of the enzyme that has the greater affinity for the nucleotide (see Fig. 2). Such a model is attractive because it allows for a variety of regulatory possibilities without the necessity for invoking multiple modulatory mechanisms, such as enzyme phosphorylation by guanine nucleo- tides4.12 or separate binding sites for different nucleotides.”
FRUSTRATION AND ADENYLATE CYCLASE 293
tr: 3
Fig. 2. Effect of an allo- i=
steric regulator on *nxymo activity. The enxyme exists in $:
equilibrium between two forms: E + E*, where E* is more active. The regulator ? binds to both forms, but with unequal affinities. Curve a,
L
affinity (K) for E’ > E, and Z binding affinity is relatively
w
good; curve b, K for E* >> E, but binding affinity is rela- tively poor; curve c, K for E* = E or K for E* = E = 0; curve d, K for E’ < E.
b
a
,
[REGULATOR]
Although there is insufficient evidence at present to demonstrate that the cyclase system is an allosteric one, such an assumption allows speculation as to the complexity of the enzyme. Well-characterized allosteric proteins studied to date are oligomeric, consisting of subunits that are symmetrical with respect to geometry and binding sites. If guanine nucleotides and hormones are allosteric modifiers, then it is reasonable to suspect that adenylate cyclase may be oligo- merit. If so, the cyclase may be unusual among allosteric enzymes in that it would most likely demonstrate an assymmetry in its subunits, since the system is functionally assymmetric across the membrane.
While we believe that the most attractive current explanation for the action of guanine nucleotides is the validity of an allosteric enzyme model, with mul- tiple ligand-linked thermodynamic states, it is also possible that guanine nucleotides act on another enzyme comprising part of a multienzyme complex. The primary impetus for considering this possibility is the hypothesis of Najar, invoked to explain the mechanism of activation of adenylate cyclase by fluoride. ”
FLUORIDE AND THE ADENYLATE CYCLASE SYSTEM
The effect of fluoride to activate adenylate cyclase remains a frustrating mystery. This phenomenon has been trying to tell us something for over 15 yr, but we are still not able to listen. However, before we display our ignorance of the mechanism of fluoride action and its consequences for the structure of the enzyme system, there are two more general facets of the effect of fluoride that need clarification. First, it has never been demonstrated that free fluoride ion, F-, is the active agent. Second, the presence of the MgF+ ion in adenylate cyclase reaction mixtures in vitro has been ignored.
It has been assumed by most investigators that F- is the active chemical

294 MAGUIRE, STURGILL, AND GILMAN
species in the stimulation of the cyclase system. Drummond and Duncan’6 demonstrated that fluoroorganic compounds did not activate; only NaF or NH,F.HF, both of which release F-, could stimulate activity. These data
demonstrate the F- is necessary, but do not eliminate fluoride complexes (Mg-F, Mg-F-P04) from consideration. However, we believe that a strong
argument can be made, based on a reinterpretation of existing data, that such
complexes are not necessary and that F- is indeed the active agent. First, when particulate preparations are incubated with fluoride alone in the
absence of Mg ‘+ ATP or phosphate and are subsequently washed to remove , the ion, adenylate cyclase is activated; ” this obviously suggests that F- is the
active species. However, it is possible that fluoride could be tightly bound and that the active complex forms later, upon addition of enzyme assay reagents.
Such tight binding of fluoride is known to occur (see below). Second, free phos- phate is apparently not necessary, since little or no phosphate is presumably
present in washed membrane fractions, and since fluoride can activate the cyclase when adenylyl imidodiphosphate is the substrate, without generation of
phosphate from pyrophosphate. (Phosphate, in millimolar concentrations,
actually inhibits fluoride activation.) Finally and most importantly, fluoride can activate the cyclase system when Mn2+ or Co’+ are substituted for Mg’+.
Relative to Mg2+, these ions do not form appreciable quantities of complexes with F- (e.g., CoF+) at the concentrations of cation that are usually used
(Kvg z 60, Kfl” z 6 2 KF”, see reference 18). The implication is that F-
is the active agent in the presence of Co’+ and Mn2+ and thus, by analogy, in
the presence of Mg ‘+ Further experimentation would be desirable to establish . this point.
There is, however, another potential complication. Since the stability con- stant for formation of MgF+ is significant, both free Mg2+ and free F- can be
reduced considerably under some conditions for the assay of adenylate cyclase.
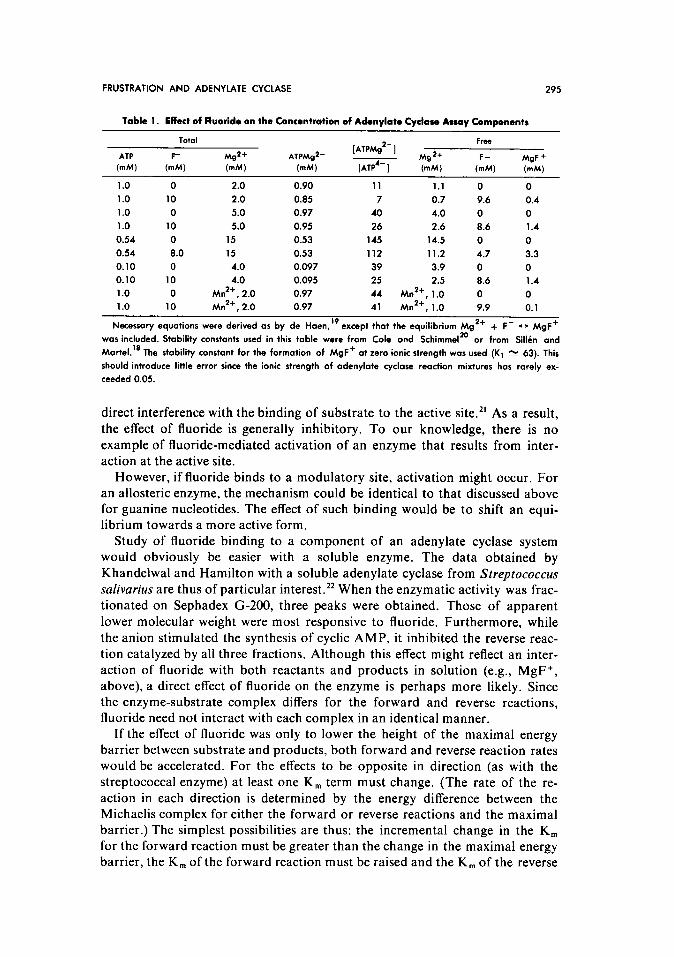
Selected examples are presented in Table 1. The data were obtained by an ex- tension of de Haen’s recent analysis” to include the reaction Mg2+ + F- -
MgF+. Attempted kinetic analysis of the effects of Mg2+ or F- on adenylate cyclase thus must be strictly controlled, since free Mg2+ or F- are certainly not identical with total Mg2+ or F- added to the system. A further effect of
fluoride may be noted in Table 1. Although the [ATPMg’-] is not appreciably affected, the ratio of [ATPMg’-]/[ATP4-] is changed significantly by the addi- tion of F-. If, as de Haen has suggested, free ATP4- is an inhibitor of adenylate
cyclase (by forming an abortive complex at the catalytic site), kinetic analysis
in the presence of fluoride is more complicated. It is also of interest to note than Mn2+ does not affect the ratio [ATPMg’-]/[ATP4-1, since it does not form
large amounts of MnF+.
The Possible Interaction of Fluoride With an Allosteric Enzyme
If fluoride interacts with an allosteric enzyme, we must consider possible sites of binding: to the active site, to a modulatory site, or to a unique location, perhaps most accessible to fluoride because of its size and charge.
Of the well-characterized enzymes with which fluoride interacts (e.g., enolase, glutamine synthetase, fructose- 1,6-diphosphatase), it apparently does so by
FRUSTRATION AND ADENYLATE CYCLASE 295
Table 1. Effact of Fluoride on the Concentration of Adenylate Cyclase Assay Components
ATP
MU
[ATPMg’-] Frm
h4g2+ MgF +
(mM) fmM)
1.0 0
1 .o 10
1 .o 0
1 .o 10
0.54 0
0.54 8.0 0.10 0
0.10 10
1 .o 0
1 .o 10
2.0 0.90
2.0 0.85
5.0 0.97
5.0 0.95
15 0.53
15 0.53
4.0 0.097
4.0 0.095
Mm2+, 2.0 0.97
ktn2+, 2.0 0.97
11 1.1 0 0
7 0.7 9.6 0.4
40 4.0 0 0
26 2.6 8.6 1.4
145 14.5 0 0
112 11.2 4.7 3.3
39 3.9 0 0
25 2.5 8.6 1.4
44 fMn2+, 1.0 0 0
41 Mn2+, 1.0 9.9 0.1
Necessary equations were derived as by de Haen,” except that the equilibrium Mg ‘+ -I- F- - MgF+
was included. Stability constants used in this table were from Cole and Schimmel 20
or from Sillin and
Maftel.‘* The stability constant for the formation of MgF+ at zero ionic strength was used (K’ % 63). This
should introduce little error since the ionic strength of adenylate cyclose reaction mixtures has rarely ex-
ceeded 0.05.
direct interference with the binding of substrate to the active site.” As a result,
the effect of fluoride is generally inhibitory. To our knowledge, there is no example of fluoride-mediated activation of an enzyme that results from inter-
action at the active site. However, if fluoride binds to a modulatory site, activation might occur. For
an allosteric enzyme, the mechanism could be identical to that discussed above for guanine nucleotides. The effect of such binding would be to shift an equi- librium towards a more active form.
Study of fluoride binding to a component of an adenylate cyclase system would obviously be easier with a soluble enzyme. The data obtained by Khandelwal and Hamilton with a soluble adenylate cyclase from Streptococcus safivarius are thus of particular interest. 22 When the enzymatic activity was frac-
tionated on Sephadex G-200, three peaks were obtained. Those of apparent
lower molecular weight were most responsive to fluoride. Furthermore, while the anion stimulated the synthesis of cyclic AMP, it inhibited the reverse reac-
tion catalyzed by all three fractions. Although this effect might reflect an inter- action of fluoride with both reactants and products in solution (e.g., MgF+, above), a direct effect of fluoride on the enzyme is perhaps more likely. Since the enzyme-substrate complex differs for the forward and reverse reactions, fluoride need not interact with each complex in an identical manner.
If the effect of fluoride was only to lower the height of the maximal energy barrier between substrate and products, both forward and reverse reaction rates would be accelerated. For the effects to be opposite in direction (as with the
streptococcal enzyme) at least one K, term must change. (The rate of the re- action in each direction is determined by the energy difference between the Michaelis complex for either the forward or reverse reactions and the maximal barrier.) The simplest possibilities are thus: the incremental change in the K, for the forward reaction must be greater than the change in the maximal energy barrier, the K, of the forward reaction must be raised and the K, of the reverse

296 MAGUIRE, STURGILL, AND GILMAN
reaction lowered, or the K, of the reverse reaction must be lowered more than is the maximal energy barrier. While the observed opposite effects of fluoride
on the forward and reverse rates are thus consistent with a direct effect on the
enzyme, necessary data for a decision are missing, i.e., K, and V for the reverse
reaction in the presence and absence of fluoride. Examinations of the kinetics of fluoride stimulation of adenylate cyclases
from eukaryotic species have revealed only an increase in V with no effect on
K, for the forward reaction. However, with no knowledge of the characteristics of the reverse reaction catalyzed by cyclases from eukaryotes, it is currently impossible to compare the mechanisms of fluoride activation of streptococcal and various eukaryotic cyclases, and we cannot decide if the bacterial enzyme
is a valid model. Since binding of F- could be sufficient to activate the cyclase system, we
need answers to two questions: can it bind, and, if so, is the available data on
fluoride activation (e.g., irreversibility) compatible with such binding? Fluoride
can clearly bind to several enzymes.” The more important problem is to recon-
cile the apparently irreversible nature of fluoride activation” with presumably
reversible binding of the ion. In at least one instance, it has been demonstrated
that treatment of a purified carboxyesterase with fluoride was not reversible over short periods of time. 23 When the esterase was diluted such that the F-
should have become noninhibitory, reactivation of the enzyme required hours of incubation. Such slow reversal of a fluoride effect could apply to the adenyl- ate cyclase system, since fluoride-treated enzyme has been assayed only at rela- tively brief intervals after removal of the bulk of the ion. Furthermore, the
assays for fluoride that have been attempted after washing of the treated enzyme have not been sufficiently sensitive to detect low levels of bound
halide. ” It has also been suggested that fluoride can inhibit enzymes by allosteric
mechanisms.24v25 Such an action could be involved in the adenylate cyclase
system if it is exerted on a separate enzyme that is inhibitory to the cyclase.
The Possible Interaction of Fluoride With a Multienzyme System
By a multienzyme system, we mean to imply an adenylate cyclase system comprised of enzymatic activities that directly interact with and modify the
catalytic unit. These modifying enzymes might coexist with the catalytic unit in the membrane as a multienzyme complex. Alternatively, some modulatory enzymes could be soluble. It is distinctly possible that fluoride could affect the
cyclase via inhibition or activation of a membrane-bound component or by functional replacement of a lost soluble component.
Modification of adenylate cyclase activity by another enzyme would pre- sumably involve a covalent alteration, and, indeed, the most likely reactions would probably involve phosphorylation or dephosphorylation of some com- ponent of the cyclase system.
In this field, one is sensitized to the possibilities that F- could activate that system by stimulation of a chain of positive reactions or by inhibition of a sequence of double or quadruple negative influences. Evidence that fluoride can

FRUSTRATION AND ADENYLATE CYCLASE 297
inhibit phosphatases and kinases abounds and need not be further discussed here.” Evidence as to the existence of a phosphorylated adenylate cyclase must await further experimentation.
Najjar has recently proposed that fluoride stimulates adenylate cyclase either by nucleophilic displacement of phosphate from a less active, phosphorylated enzyme form or by stimulation of a phosphatase which then removes the phosphate from the same form of the cyclase. ” It should be noted that the first proposal is entirely consistent with the known organic chemistry of both fluo- ride and phosphate. Indeed, Jencksz6 has stated:
“Fluoride . . . is an extremely effective nucleophile toward phosphate; it is one of the few compounds which competes effectively with water for reaction with monosubstituted phosphates, and it is more reactive than ethoxide, phenoxide, and thiophenoxide toward diisopropyl phosphorochloridate. This high reactivity parallels the high thermodynamic stability of fluorophos- phates. . . .” Even with this in mind, we feel that the hypothesis of nucleophilic displace-
ment of phosphate by fluoride is subject to reservation. Najjar and co-workers reported that fluoride could release 32P. from the active serine phosphate of purified phosphoglucomutase,27 and hale suggested a similar mechanism for the fluoride activation of adenylate cyclase. However, the effect of fluoride on phosphoglucomutase requires a divalent cation, and there is no evidence of such a need for activation of adenylate cyclase. Furthermore, Mg*+ is a poor divalent cation to meet this requirement in the reaction with phosphoglucomu- tase. The presumed ability of fluoride to dephosphorylate both phosphoglu- comutase and adenylate cyclase also predicts that the dephosphorylation of other enzymes, such as glycogen synthase, phosphorylase, and phosphorylase kinase, should occur with corresponding changes in enzymatic activity.
An additional insight into the nature of adenylate cyclase and its activation by fluoride is perhaps found in the work of Robison and co-workers.28 Fluoride stimulation of adenylate cyclase from brain was shown to be a function of age in developing rats. Fluoride was without effect until between days 5 and 9 postpartum. After day 9, activity in the absence of fluoride dropped, while activity in the presence of fluoride continued to rise. The decrease in activity in the absence of huoride suggests the appearance of an inhibitory influence on the cyclase, while the apparent independence of fluoride stimulation and adenylate cyclase activity may imply their localization on macromolecular components that are under independent genetic regulation.
The pessimistic tone used to introduce this limited review compels us to offer these concluding remarks. Too large a portion of published work, our own included, is devoted to demonstrations of effects of multiple agents on adenyl- ate cyclase systems derived from the entire spectrum of living organisms. The temptation is obvious. It is clear, however, that the elucidation of the com- plexity of adenylate cyclase necessitates the opposite course: the extensive analysis of a limited number of model systems. Such an approach has been taken by only a few groups, notably, in our opinion, by Rodbell and colleagues and Bockaert et a1.8*29-3’ Most of our detailed knowledge of adenylate cyclase derives from such efforts.

298 MAGUIRE, STURGILL, AND GILMAN
ACKNOWLEDGMENT
The authors wish to thank Dr. Rodney Biltonen for a critical reading of this manuscript.
REFERENCES
1. Maguire ME, Sturgill TW, Anderson HJ,
Minna JD, Gilman AG: Hormonal control of
cyclic AMP metabolism in parental and hybrid
somatic cells, in Drummond GI, Greengard P,
Robison GA (eds): Advances in Cyclic Nucleo-
tide Research, vol 5. New York, Raven Press
(in press)
2. Robison GA, Butcher RW, Sutherland
EW: On the relation of hormone receptors to
adenyl cyclase, in Danielli JF, Moran JF,
Triggle DJ (eds): Fundamental Concepts in
Drug-Receptor Interaction. New York, Aca-
demic Press, 1970, pp 59-91
3. Puchwein G, Pfeuffer T, Helmreich EJM:
Uncoupling of catecholamine activation of
pigeon erythrocyte membrane adenylate cyclase
by filipin. J Biol Chem 249:3232-3240, 1974
4. Rodbell M: The problem of identifying
the glucagon receptor. Fed Proc 32: 1854- 1858,
I973
5. Rodbell M, Krans HMJ. Pohl SL, Birn- baumer L: The glucagon-sensitive adenyl
cyclase system in plasma membranes of rat
liver. IV. Effects of guanyl nucleotides on bind-
ing of ‘2sI-glucagon. J Biol Chem 246:1872-
1876, 1971
6. Rodbell M, Lin MC, Salomon Y: Evi-
dence for interdependent action of glucagon
and nucleotides on the hepatic adenylate
cyclase system. J Biol Chem 249:59-64, 1974
7. Birnbaumer L, Pohl SL, Rodbell M,
Sundby F: The glucagon-sensitive adenyl
cyclase system in plasma membranes of rat
liver. IV. Hormonal stimulation: reversibility
and dependence on concentration of free hor-
mone. J Biol Chem 247:2038-2043, 1972
8. Bockaert J, Roy C. Jard 5: Oxytocin-
sensitive adenylate cyclase in frog bladder
epithelial cells. Role of calcium, nucleotides,
and other factors in hormonal stimulation. J
Biol Chem 247:7073-7081, 1972
9. Leray F. Chambaut A-M, Perrenoud M-L, Hanoune J: Adenylate-cyclase activity of
rat-liver plasam membranes. Hormonal stimu-
lations and effect of adrenalectomy. Eur J Bio-
them 38:185-192, 1973
IO. Bilezikian JP, Aurbach CD: The effects of nucleotides on the expression of fl-adrenergic
adenylate cyclase activity in membranes from
turkey erythrocytes. J Biol Chem 249: I57- 161,
1974.
I I Wolff J, Cook GH: Activation of thyroid
membrane adenylate cyclase by purine nucleo-
tides. J Biol Chem 248:350-355. 1973
12. Harwood JP, Low H, Rodbell M: Stimu-
latory and inhibitory effects of guanyl nucleo-
tides on fat cell adenylate cyclase. J Biol Chem
248:6239-6245, 1973
13. Edelstein SJ: An allosteric mechanism
for the acetylcholine receptor. Biochem Biophys
Res Commun 48:1160-l 165, 1972
14. Monod J, Wyman J, Changeux J-P: On
the nature of allosteric transitions: A plausible
model. J Mol Biol 12:88-l 18, I965
15. Najjar VA, Constantopoulos A: The
activation of adenylate cyclase. I. A postulated
mechanism for fluoride and hormone activation
of adenylate cyclase. Mol Cell Biochem 2:87-
93, 1973
16. Drummond Cl, Duncan L: Adenyl
cyclase in cardiac tissue. J Biol Chem 245:976--
983, 1970
17. Perkins JP: Adenyl cyclase, in Green-
gard P, Robison GA (eds): Advances in Cyclic
Nucleotide Research, vol 3. New York, Raven
Press, 1973, pp l-64
18. Sill&n LG. Martell AE: Stability con-
stants of metal-ion complexes. London, The
Chemical Society, 1964, and Suppl I. 1971
19. De Haen C: Adenylate cyclase. A new
kinetic analysis of the effects of hormones and
fluoride ion. J Biol Chem 249:2756-2762. 1974
20. Cole FX. Schimmel PR: lsoleucyl trans-
fer ribonucleic acid synthetase. The role of mag-
nesium in amino acid activation. Biochemistry
9:314333148, 1970
21. Wiseman A: Effect of inorganic fluoride
on enzymes, in Smith FA (ed): Handbuch der
experimentellen Pharmakologie Xx/2. Phar-
macology of Fluorides. Berlin, Springer-Verlag,
1970, pp 48897
22. Khandelwal RL. Hamilton IR: Purifica-
tion and properties of adenyl cyclase from
Streptococcus salivarius. J Biol Chem 246~3297~
3304,1971
23. Haugen DA, Suttie JW: Fluoride inhibi-
tion of rat liver microsomal esterases. J Biol
Chem 249~2723~2731, I974
24. Farias RN, Goldemberg AL. Trucco RE:
The effect of fat deprivation on the allosteric
inhibition by fluoride of the (Mg2+)-ATPase
and (Na+ + K+)-ATPase from rat erythro-
cytes. Arch Biochem Biophys 139:39-44. 1970
25. Morero RD. Bloj B, Farms RN. Trucco

FRUSTRATION AND ADENYLATE CYCLASE 299
RE: The allosteric transitions from membrane- bound enzymes: Behavior of erythrocyte acetyl- cholinesterase from fat-deficient rats. Biochim Biophys Acta 282: 157- 165, 1972
26. Jencks WP: Catalysis in Chemistry and Enzymology. New York, McGraw-Hill. 1969, p 103
27. Layne PP, Najjar VA: Dephosphoryla- tion of phosphoglucomutase by fluoride, cysteine, and hydroxylamine. Fed Proc 32:667, 1973
28. Robison GA, Schmidt MJ, Sutherland EW: On the development and properties of the brain adenyl cyclase system, in Greengard P, Costa E (eds): Role of Cyclic AMP in Cell
Function. Advances in Biochemical Psycho- pharmacology, vol 3. New York, Raven Press, 1970, pp I l-30
29. Bockaert J, Jard S, Morel F, Montegut M: Uptake of oxytocin-‘H by frog skin and bladder epithelial cells. Am J Physiol 219: 1514-1521, 1970
30. Bockaert J, Imbert M, Jard S, Morel F: ‘H-oxytocin binding sites in the isolated frog skin epithelium: Relation to the physiological response. Mol Pharmacol8:230-240, I972
31. Bockaert J, Roy C, Rajerison R, Jard S: Specific binding of [ ‘Hllysine-vasopressin to pig kidney plasma membranes. J Biol Chem 2485922-5931, 1973












![Development of Glucagon Sensitivity Neonatal Rat Liver · activity of ['3P]ATP. Protein was determined fluorometri-cally (18) and adenylate cyclase activity was expressed as picomoles](https://img.pdfslide.us/doc/110x75/5cc4ce6288c993474e8c3ac5/development-of-glucagon-sensitivity-neonatal-rat-liver-activity-of-3patp.jpg)






![Index [link.springer.com]978-3-642-38487...Index A ACA. See Adenylyl cyclase acaA. See Adenylate cyclase A ACC. See 1-aminocyclo-propane-1-carboxylic acid Acinetobacter baumannii,](https://img.pdfslide.us/doc/110x75/5b47af067f8b9af5078c45af/index-link-978-3-642-38487index-a-aca-see-adenylyl-cyclase-acaa-see-adenylate.jpg)









