Embed Size (px)
Citation preview

Journal of the History of Biology 30: 1–29, 1997. 1c 1997 Kluwer Academic Publishers. Printed in the Netherlands.
From the Regulatory Vision of Cancer to the OncogeneParadigm, 1975–1985
MICHEL MORANGEDepartment of BiologyEcole Normale Superieure46 rue d’Ulm75230 Paris Cedex 05, France
The discovery and characterization of cellular oncogenes – the limited groupof genes involved in tumor formation – was one of the most brilliant achieve-ments made possible by the new recombinant DNA technology (see Table 1for a chronology of events).1 The new model of oncogenesis was advanta-geously substituted for the previous unsuccessful theories, which had empha-sized the role of viruses.2
I would like to argue that the most dramatic change that occurred duringthese crucial years (1975–1985) was not the renunciation of the viral theoryof oncogenesis – in fact, in the new model, cancer might also result from aviral infection – but the disappearance of the previous mundane regulatoryvision of cancer that was dominant when the oncogene paradigm came intobeing.3 My aim will be to characterize this regulatory vision. Then, I will
1 Michel Morange, “The Discovery of Cellular Oncogenes,” Hist. Phil. Life Sci., 15 (1993),45–59; Giancarlo Vecchio, “Oncogenes of DNA and RNA Tumor Viruses and the Origin ofCellular Oncogenes,” ibid., pp. 59–74.
2 Joan H. Fujimura, “The Molecular Biological Bandwagon in Cancer Research: WhereSocial Worlds Meet,” Soc. Prob., 35 (1988), 261–283; Jean-Paul Gaudilliere, “Oncogenes asMetaphors for Human Cancers: Articulating Laboratory Practices and Medical Demands,” inMedicine and Change: Historical and Sociological Studies of Medical Innovation, ed. IlanaLowy (Paris: INSERM J. Libbey Eurotext, 1992), pp. 213–247. For a general history ofcancer in the twentieth century, see David Cantor, “Cancer,” in Companion Encyclopedia ofthe History of Medicine, vol. I, ed. W. F. Bynum and Roy Porter (London and New York:Routledge, 1993), pp. 537–561; and J. Michael Bishop, “Cancer: The Rise of the GeneticParadigm,” Genes Devel., 9 (1995), 1309–1315.
3 I use the expression “oncogene paradigm” to refer to the limited set of theories that, around1985, were considered by scientists to provide an “explanation” of cancer. This paradigm didnot replace another paradigm, but was a substitute for a long-lasting, mundane regulatoryvision of cancer. In a previous article (Morange, “Discovery of Cellular Oncogenes” [above, n.1]) I characterized the main components of the oncogene paradigm and demonstrated that itsacceptance took place much later than the discovery of cellular oncogenes. Readers unfamiliarwith this domain may refer to Harold Varmus and Robert A. Weinberg, Genes and the Biologyof Cancer (New York: Scientific American Library, 1992). A more complete description of
VICTORY: PIPS No.: 125343 LAWKAPhist3011.tex; 30/05/1997; 9:35; v.5; p.1

2 MICHEL MORANGE
Table 1. The birth of the oncogene paradigm.
1911 Discovery by P. Rous of the first cancer virus1961 Models of gene regulation (F. Jacob and J. Monod)1969 The Provirus Model (R. J. Huebner and G. J. Todaro)1976 Discovery of the first cellular oncogene, homologons to a retroural
oncogene (D. Stehelin, H. Varmus, M. Bishop)1979–1980 Intercellular passage of the transformed phenotype by a single gene
transfer (G. Cooper, R. Weinberg)1982 Transformation may result from a point mutation in a cellular oncogene1983–1984 Oncogenes code for growth factors or growth-factor receptors1986 Discovery of the first antioncogene (tumor suppressor gene)
describe the major transition corresponding to the development of the newoncogene paradigm, and will look for the mechanisms responsible for it.
Joan Fujimura has offered some explanations for the birth of the oncogeneparadigm, directly connecting the “interests” of molecular biologists andthe rise of genetic engineering technology.4 I will discuss the value of thesemodels and will insist on the necessity of alternative tools to follow moreclosely the progressive shaping of the oncogene theory. Rather than the strate-gies of molecular biologists, I will emphasize the major role played by therhythm and form of discoveries in the elaboration of the new paradigm.
Cancer as a Dysregulation (1960–1980)
The regulatory molecular models elaborated in bacteria in 1960 had a pro-found influence on biology as a whole. They opened the way to the studyof differentiation and development by molecular biologists.5 But they alsodeeply affected the theories of carcinogenesis constructed between 1960 and1980.
The vision of cancer as a dysregulation was widely accepted between 1960and 1980. In his answer to an amateur scientist who had proposed a model ofoncogenesis, Jacques Monod (who, along with Francois Jacob, fathered thegenetic models of regulation in bacteria) admitted in 1968 that cancer was
the dominant oncogenes may be found in Geoffrey Cooper, Oncogenes (Boston: Jones andBartlett, 1990).
4 Fujimura, “Molecular Biological Bandwagon” (above, n. 2); Joan Fujimura, “Constructing‘Do-able’ Problems in Cancer Research: Articulating Alignment,” Soc. Stud. Sci., 17 (1987),257–293.
5 Michel Morange, “Construction of the Developmental Gene Concept: The Crucial Years,1960–1980,” Biol. Zentral bl., 115 (1996), 132–138; idem, Histoire de la biologie moleculaire(Paris: La Decouverte, 1994), chap. 14.
hist3011.tex; 30/05/1997; 9:35; v.5; p.2

FROM THE REGULATORY VISION OF CANCER TO THE ONCOGENE PARADIGM 3
almost certainly “a disease of genetic expression.”6 In 1966, Harry Rubinin Phage and the Origins of Molecular Biology wrote that cancer was “adisruption of the regulatory processes of cells.”7 The understanding of cancertherefore relied on a “natural” extension to higher organisms of the regulatorymodels elaborated in bacteria. This probably explains why, between 1962 and1975, Jacques Monod attended many symposia on cancer.8
The provirus model of Robert Huebner and George Todaro, which was themost popular model of oncogenesis at the end of the sixties, was a transposi-tion of the results obtained at the Pasteur Institute in Paris by Andre Lwoff,Elie Wollman, and Francois Jacob on the prophage, to higher organisms andto the specific problem of oncogenesis.9 It also incorporated studies concern-ing oncogenic viruses, in particular those of Ludwik Gross on the murineleukemia viruses.10 In this model, oncogenesis resulted from the “derepres-sion” of a silent provirus by different viral or chemical agents.
An even more direct transposition of the bacterial regulatory models led in1973 to the elaboration by David Comings of a “general theory of carcino-genesis”: cells possess multiple structural genes (Tr) coding for transformingfactors that can release the cell from its normal constraints to growth; theaction of these transforming genes is suppressed by a pair of regulatory genesthat code for a repressor.11 This regulatory vision of cancer was stronglysupported by results obtained by the hybridization of transformed, cancerouscells with normal cells. Despite some initial conflicting results and interpre-tations, it was widely accepted from these experiments that the transformedstate was negatively controlled – repressed – by the action of normal genes.12
6 “Le cancer est a peu pres certainement une maladie de l’expression genetique”: JacquesMonod to G. Xerri, November 4, 1968, Institut Pasteur Archive Center, Paris.
7 H. Rubin, “Quantitative Tumor Virology,” in Phage and the Origins of Molecular Biology,ed. John Cairns, Gunther S. Stent, and James D. Watson (Cold Spring Harbor, N.Y.: Cold SpringHarbor Laboratory Press, 1966), p. 299.
8 Jacques Monod, “Conclusive Remarks,” in Symposium of Basic Problems in NeoplasticDisease, ed. Alfred Gellhorn and E. Hirschberg (New York: Columbia University Press, 1962),pp. 218–237. Health problems prevented Monod from attending the Dedication Symposium forthe new cancer research facilities at the Wistar Institute: Correspondence of Jacques Monod,Institut Pasteur Archive Center.
9 Robert J. Huebner and George J. Todaro, “Oncogenes of RNA Tumor Viruses as Deter-minants of Cancer,” Proc. Nat. Acad. Sci. USA, 64 (1969), 1087–1094; George J. Todaroand Robert J. Huebner, “The Viral Oncogene Hypothesis: New Evidence,” ibid., 69 (1972),1009–1015; Charles Galperin, “Virus, provirus et cancer,” Rev. Hist. Sci., 47 (1994), 7–56.
10 Ludwik Gross, Oncogenic Viruses (Oxford and New York: Pergamon Press, 1961).11 David E. Comings, “A General Theory of Carcinogenesis,” Proc. Nat. Acad. Sci. USA, 70
(1973), 3324–3328.12 Henry Harris, O. J. Miller, George Klein, P. Worst, and T. Tachibana, “Suppression of
Malignancy by Cell Fusion,” Nature, 223 (1969), 363–368; Boris Ephrussi, Richard L. David-son, and Mary C. Weiss, “Malignancy of Somatic Cell Hybrids,” Nature, 224 (1969), 1314–
hist3011.tex; 30/05/1997; 9:35; v.5; p.3

4 MICHEL MORANGE
This regulatory vision was still dominant when the new oncogene paradigmtook shape. The first experiments leading to the acceptance of the latter weretherefore interpreted within the framework of the theory that cancer was dueto the dysregulation of some specific genes, the oncogenes.
The groups of Robert Weinberg and Geoffrey Cooper were both workingon the mechanisms leading to cell transformation (cells extracted from tumorsand put into culture in vitro have properties that distinguish them from normalcells: they are called “transformed cells”).13 In 1979 and 1980, both groupsshowed that DNA extracted from chemically transformed tumor cells, whentransferred to “normal” cells (a process called transfection), was able totransform the recipient cells.14 However, the results obtained by the twolaboratories were not identical. For Weinberg, DNA extracted from normalcells was unable to act in a similar way. Cooper obtained results that weredifferent: in his case, the critical parameter was the size of the DNA usedfor the transfection experiments; large fragments of DNA from normal ortransformed cells were unable to transform, but when DNA was disruptedinto small fragments, whatever its origin, it acquired the capacity to transformat a low efficiency. Cooper suggested that the activity of the transferredoncogenes was controlled by the DNA sequences surrounding the oncogene.This explanation was in complete agreement with the regulatory models ofcancer derived from the work of Jacob and Monod: in transformed cells, thechemicals may have mutated and inactivated the genes coding for the trans-acting repressor of the transforming gene; the transfected oncogene wouldtherefore never be expressed in a recipient cell in which this repressor wasstill functional, unless the oncogene is separated from the DNA sequences onwhich the repressor binds.
Almost simultaneously, many other studies confirmed that some of thecellular oncogenes (genes that had been shown for the first time by Dominique
1315; Eric J. Stanbridge, “Suppression of Malignancy in Human Cells,” Nature, 260 (1976),17–20; Dorothy A. Miller and Orlando J. Miller, “Chromosomes and Cancer in the Mouse:Studies in Tumors, Established Cell Lines, and Cell Hybrids,” Adv. Cancer Res., 39 (1983),153–182; J. A. Witkowski, “Somatic Cell Hybrids: A Fusion of Biochemistry, Cell Biologyand Genetics,” Trends Biochem. Sci., 11 (1986), 149–152; Henry Harris, The Cells of the Body:A History of Somatic Cell Genetics (Cold Spring Harbor, N.Y.: Cold Spring Harbor LaboratoryPress, 1995), pp. 222–224.
13 It is also possible to transform normal cells in vitro by different treatments. Even if thesein vitro – transformed cells share many characteristics with the transformed cells coming fromtumors, it is often problematic to ascertain that they are totally identical to cancer cells (seebelow).
14 Chiaho Shih, Ben-Zion Shilo, Mitchell P. Goldfarb, Ann Dannenberg, and Robert A.Weinberg, “Passage of Phenotypes of Chemically Transformed Cells via Transfection of DNAand Chromatin,” Proc. Nat. Acad. Sci. USA, 76 (1979), 5714–5718; Geoffrey M. Cooper,Sharon Okenquist, and Lauren Silverman, “Transforming Activity of DNA of ChemicallyTransformed and Normal Cells,” Nature, 284 (1980), 418–421.
hist3011.tex; 30/05/1997; 9:35; v.5; p.4

FROM THE REGULATORY VISION OF CANCER TO THE ONCOGENE PARADIGM 5
Stehelin, Harold Varmus, Michael Bishop, and Peter Vogt to be homologousto the transforming genes carried by retroviruses)15 were able to transformrecipient cells when placed under control elements allowing the synthesis oftheir products (their expression, as molecular biologists say) at a very highlevel.16 In a similar way, Robert Weinberg admitted that, in 1982, he wasastonished when his collaborators found that the level of expression of thegene they had isolated by transfection assay, ras, was not altered in bladdercarcinoma cells) and that the mutation leading to its activation was not locatedin the promoter, but rather in the coding portion of the gene, leading to thereplacement of one amino acid by another in the protein.17
I shall return more explicitly to these experiments below. Here I want onlyto emphasize the wide acceptance of this regulatory vision of cancer, and tounderline that it was not a precise one. Many different choices still remainedopen. This vision was shared by biologists such as Charles Heidelberger whosupported an epigenetic model of cancer,18 as well as by those convincedof the mutational origin of cancer. The transfer of the regulatory models tohigher organisms led Alfred Knudson and David Comings to the convictionthat mutations leading to cancer were recessive.19 On the other hand, thetransfection experiments of Weinberg demonstrated the dominant nature ofthe cancer lesions. In fact, if we adopt the models of Jacob and Monod toexplain the origin of cancers, it is possible to simultaneously accommodatethe existence of dominant and recessive mutations able to cause cell transfor-mation and cancer. Interestingly, however, these explicit explanations wererarely mentioned. The regulatory vision of cancer was often a very vague one.
15 Dominique Stehelin, Harold E. Varmus, J. Michael Bishop, and Peter K. Vogt, “DNARelated to the Transforming Gene(s) of Avian Sarcoma Viruses Is Present in Normal AvianDNA,” Nature, 260 (1976), 170–173.
16 Marianne Oskarsson, William L. McClements, Donald G. Blair, J. V. Maizel, and GeorgeVande Woude, “Properties of a Normal Mouse Cell DNA Sequence (sarc) Homologous tothe src Sequence of Moloney Sarcoma Virus,” Science, 207 (1980), 1222–1224; D. G. Blair,et al., “Activation of the Transforming Potential of a Normal Cell Sequence: A MolecularModel for Oncogenesis,” Science, 212 (1981), 941–943; Deborah De Feo et al., “Analysisof Two Divergent Rat Genomic Clones Homologous to the Transforming Gene of HarveyMurine Sarcoma Virus,” Proc. Nat. Acad. Sci USA, 78 (1981), 3328–3332; Esther H. Chang,Mark E. Furth, Edward M. Scolnick, and Douglas R. Lowy, “Tumorigenic Transformation ofMammalian Cells Induced by a Normal Human Gene Homologous to the Oncogene of HarveyMurine Sarcoma Virus,” Nature, 297 (1982), 479–483.
17 Natalie Angier, Natural Obsessions: The Search for the Oncogene (Boston: HoughtonMifflin, 1988).
18 Henry C. Pitot and Charles Heidelberger, “Metabolic Regulatory Circuits and Carcinogen-esis,” Cancer Res., 23 (1963), 1694–1700.
19 Alfred G. Knudson, Jr., “Mutation and Cancer: Statistical Study of Retinoblastoma,” Proc.Nat. Acad. Sci USA, 68 (1971), 820–823; Alfred G. Knudson, “Mutation and Human Cancer,”Adv. Cancer Res., 17 (1973), 317–352; Comings, “General Theory of Carcinogenesis” (above,n. 11).
hist3011.tex; 30/05/1997; 9:35; v.5; p.5

6 MICHEL MORANGE
References to precise models of regulation were rare, and the experimentaldifficulties that prevented regulatory models from being extended to higherorganisms do not appear sufficient to explain this apparent lack of interest.20
My hypothesis is that the regulatory vision of cancer was an implicitly accept-ed component of the thinking of cancer researchers. Although it was not anefficient concept that could lead to an entirely new vision of cancer, it was along-term stable concept as described by Peter Keating, Alberto Cambrosio,and Michael Mackenzie.21
Finally, I would like to insist upon the close link between the regulatoryvision of cancer and the regulatory vision of embryogenesis and development.Development was also considered to be a regulatory process involving the dif-ferential repression of genes in different tissues. Moreover, numerous studiesshowed that cancer cells shared properties with early embryonic cells: cancercells reexpress fetal and embryonic characteristics.22 Dysregulation of cancercells was therefore considered as a temporally and locally incorrect transitionof the cells from their differentiated state towards their embryonic state ofgene expression.23 Thus, cancer was conceived as a disease of development.
20 The evidence for the existence in eukaryotes of regulatory circuits analogous to thoseexisting in prokaryotes was always indirect, often resulting from the analysis of the propertiesof hybrid cells. For instance, see Richard L. Davidson, Boris Ephrussi, and Kohtaro Yamamoto,“Regulation of Pigment Synthesis in Mammalian Cells as Studied by Somatic Hybridization,”Proc. Nat. Acad. Sci. USA, 56 (1966), 1437–1440; Richard Davidson and Kohtaro Yamamoto,“Regulation of Melanin Synthesis in Mammalian Cells as Studied by Somatic Hybridization, II.The Level of Regulation of 3,4-DihydroxyPhenylalanine Oxidase,” ibid., 60 (1968), 894–901;and Thomas B. Shows and Frank H. Ruddle, “Function of the Lactate Dehydrogenase B Genein Mouse Erythrocytes: Evidence for Control by a Regulatory Gene,” ibid., 61 (1968), 574–581. A complete description of the early attempts to extend the operon model to eukaryotesand its associated difficulties may be found in Jean-Paul Gaudilliere, “Biologie moleculaireet biologistes dans les annees soixante: La naissance d’une discipline, le cas francais,” Ph.D.thesis, University of Paris, 1991. Whatever the real faith in the bacterial regulatory models,dysregulation was always considered as synonymous to activation, in agreement with the firstmodels of Jacques Monod and Francois Jacob, but also with some much older conceptions ofgene regulation (Edgar Stedman and Ellen Stedman, “Cell Specificity of Histones,” Nature,166 [1950], 780–781).
21 Peter Keating, Alberto Cambrosio, and Michael Mackenzie, “The Tools of the Discipline:Standards, Models, and Measures in the Affinity/Avidity Controversy in Immunology,” in TheRight Tools for the Job: At Work in Twentieth-Century Life Sciences, ed. Adele E. Clarke andJoan H. Fujimura (Princeton: Princeton University Press, 1992), 312–354.
22 Some examples, from a very abundant literature: Elliott H. Stonehill and Aaron Bendich,“Retrogenetic Expression: The Reappearance of Embryonal Antigens in Cancer Cells,” Nature,228 (1970), 370–371; Fanny Schapira, “Resurgence of Fetal Isozymes in Cancer: Study ofAldolase, Pyruvate Kinase, Lactic Dehydrogenase and �-Hexosaminidase,” in Isozymes, Curr.Top. Biol. Med. Res., 5 (1981), 27–75; Edward J. Benz et al., “Embryonic Fetal ErythroidCharacteristics of a Human Leukemic Cell Line,” Proc. Nat. Acad. Sci. USA, 77 (1980),3509–3513.
23 Van R. Potter, “Oncogeny Is Blocked Ontogeny,” reported in Nature, 225 (1970), 588–589; Clement L. Markert, ‘Neoplasia: A Disease of Cell Differentiation,” Cancer Res., 28
hist3011.tex; 30/05/1997; 9:35; v.5; p.6

FROM THE REGULATORY VISION OF CANCER TO THE ONCOGENE PARADIGM 7
Evidence for a New Paradigm: The Nature of the Change
In order to show how the previously described continuity of the models ofcancer contrasts with the “oncogene revolution” of the 1980s, I will rapidlydescribe the main components of the oncogene paradigm. In this paradigm,cancer is due to the increased expression or structural modification of alimited number of genes called the oncogenes. These oncogenes can be acti-vated by different mechanisms: direct mutation, transcriptional activation bythe close insertion of a retrovirus, translocation that places them under thecontrol of very active promoters, or insertion into a retrovirus, which leads tovery high levels of expression. Oncogenes may also be activated by ampli-fication of the corresponding DNA sequences. Cancer is due to a multiplealteration of oncogenes, and therefore to the cooperative action of differentoncogenes. Oncogenes code for proteins involved in the control of cell divi-sion: growth factors, which activate cell division, the receptor proteins forthese growth factors, the intermediary proteins relaying the extracellular sig-nals of these growth factors toward the cell nucleus; and nuclear transcriptionfactors responsible for the state of gene expression characteristic of rapidlyreplicating cells. (I have not mentioned in this description the antioncogenesor tumor suppressor genes; their discovery came later, and only slightly alteredthe main components of this oncogene paradigm.)24
What best distinguishes this new oncogene paradigm from the previousstudies of cancer is the limited number of hypotheses forming the new model,in contrast to the abundance of contradictory, but not conflicting, theories anddata on cancer at the end of the 1970s. I will give a very brief summary ofthem.
The Virus Cancer Program was initiated in 1964 in the National CancerInstitute, and was developed extensively after the announcement by PresidentRichard Nixon that the same tactics that had been successful in the Apolloprogram would be applied against cancer.25 This program was centered onthe search for cancer viruses. It ended in 1978 with the general feeling thatmost human cancers were not due to the action of viruses, even if somecancers had been unambiguously shown to result from a viral infection.26
(1968), 1908–1914; Thomas Graf, Norbert Ade, and Hartmut Beug, “Temperature-SensitiveMutant of Avian Erythroblastosis Virus Suggests a Block of Differentiation as Mechanism ofLeukaemogenesis,” Nature, 275 (1978), 496–501.
24 On the antioncogenes or tumor suppressor genes, see George Klein, Tumor SuppressorGenes (New York: Marcel Dekker, 1990).
25 Jean-Paul Gaudilliere, “Le cancer entre infection et heredite: Genes, virus et souris auNational Cancer Institute (1937–1977),” Rev. Hist. Sci., 47 (1994), 57–89.
26 Some DNA viruses, such as the Epstein-Barr virus belonging to the family of herpesviruses, had been clearly demonstrated to be responsible for some very specific forms of
hist3011.tex; 30/05/1997; 9:35; v.5; p.7

8 MICHEL MORANGE
Since cancer rates were highly variable from one country to another, manybiologists thought that oncogenesis probably resulted from the action ofthe environment; there were probably not one but several mechanisms ofoncogenesis, leading to different forms of cancer.27 Cancer was conceived asan epigenetic phenomenon, or as due to actual mutations.28 These mutationswere suggested to be either dominant or recessive; they could be either pointmutations or large alterations of genes, such as translocation of differentchromosome fragments. It was also not clear whether a higher incidenceof cancer resulted from an increased rate of mutation, or from a decreasedrate of repair of DNA damage.29 The relevance of molecular studies of thetransformed state to the understanding of the formation of human tumors wasfar from being widely accepted. It was also advocated that the real cause ofcancer was a deficiency in the immune response of the host against the tumorcells.30 Therefore it was not really evident in 1980 whether the studies oncancer ought to be targeted toward environmental studies, the preparationof magic immune bullets against cancerous cells, or molecular studies oftransformed cells.
These different conceptions of cancer were linked with different researchstrategies, different disciplines, and different tools. This link was very looseand precluded a clear distinction between schools. Moreover, as we saw pre-viously, these different conceptions of cancer were not in conflict, and someexpressions, such as dysregulation, made it possible to reconcile opposingviews.
The consequence is that all the structural or functional components of theoncogene paradigm had already been proposed as explanations for the for-mation of tumors. The oncogene hypothesis had been cited in its presentlyaccepted form – in which the mutation of cellular genes leads to the for-mation of cancer – by Howard Temin in 1974 as one of many models ofoncogenesis.31 The major role of gene translocation had been proposed by
human cancer; see George Klein, “Herpesviruses and Oncogenesis,” Proc. Nat. Acad. Sci.USA, 69 (1972), 1056–1064.
27 John Cairns, Cancer, Science, and Society (San Francisco: Freeman, 1978).28 The careful experiments of Bruce Ames and colleagues showing that carcinogens were
mutagens when metabolized by the organism increased the number of biologists convincedthat cancer resulted from mutations: Joyce McCann, Edmund Choi, Edith Yamasaki, and BruceN. Ames, “Detection of Carcinogens as Mutagens in the Salmonella Microsome Test: Assayof 300 Chemicals,” Proc. Nat. Acad. Sci. USA, 72 (1975), 5135–5139.
29 John Cairns, “Mutation Selection and the Natural History of Cancer,” Nature, 255 (1975),197–200; R. B. Setlow, “Repair Deficient Human Disorders and Cancer,” Nature, 271 (1978),713–717.
30 Robert A. Good, “Relation between Immunity and Malignancy,” Proc. Nat. Acad. Sci.USA, 69 (1972), 1026–1032.
31 Howard M. Temin, “On the Origin of the Genes for Neoplasia: G. H. A. Clowes MemorialLecture,” Cancer Res., 34 (1974), 2835–2841.
hist3011.tex; 30/05/1997; 9:35; v.5; p.8

FROM THE REGULATORY VISION OF CANCER TO THE ONCOGENE PARADIGM 9
cell biologists who had highlighted the numerous chromosomal rearrange-ments accompanying cell transformation and tumor formation.32 The ideathat neoplasia could be due to the inappropriate production of ligands withstructural similarity to growth factors by cells that possess receptors for thoseligands – and are therefore sensitive to their action – was a long-lived idea,but it was explicitly proposed as a mechanism of oncogenesis by GeorgeTodaro and others as early as 1978 and 1979.33 Immediately upon its discov-ery in 1976, the first oncogene was suggested to play a role in the regulation ofgrowth and development.34 The idea that oncogenesis was a multistep processwas widely believed to explain the rarity of cancer, its exponential increasewith age, and the separate but additive action of promoting and initiatingagents in the case of chemically induced carcinogenesis.35
Therefore the acceptance of the oncogene paradigm can be explainedneither by the appearance of new theories or models, nor (as we shall seelater) by the efficiency of new tools, but, on the contrary, by the selectionof a limited number of hypotheses and the acceptance of this limited num-ber of hypotheses as central to the explanation of cancer. Cancer had beenoverexplained by the models existing at the end of 1970s. The reorganizationof theories and practices associated with the diffusion of the oncogene para-digm therefore appears as an original case study. The advent of the oncogeneparadigm represented the reorganization of a cognitive field as much as theconstruction of a new object.
To provide a complete description of this reorganization, we must notforget to mention the theories, models, or tools that were not incorporatedinto the oncogene paradigm, or the lines of research that were promising butwere abandoned, or at least did not play a major role in the formation of thenew paradigm. For instance, Elisabeth Gateff’s studies on Drosophila hadled to the characterization of genes whose deletion or inactivation induced
32 John Cairns, “The Origin of Human Cancers,” Nature, 289 (1981), 353–357. (This articleappeared after some components of the oncogene paradigm had already been formulated;however, its conclusion relied mainly on earlier data.)
33 Robert W. Holley, “Control of Growth of Mammalian Cells in Cell Culture,” Nature, 258(1975), 487–490; George J. Todaro and J. E. De Larco, “Growth Factors Produced by SarcomaVirus-Transformed Cells,” Cancer Res., 38 (1978), 4147–4154; J. E. De Larco and George J.Todaro, “Growth Factors from Murine Sarcoma Virus-Transformed Cells,” Proc. Nat. Acad.Sci. USA, 75 (1978), 4001–4005; Michael B. Sporn and George J. Todaro, “Autocrine Secretionand Malignant Transformation of Cells,” New Eng. J. Med., 303 (1980), 878–880; MichaelS. McGrath and Irving L. Weissman, “AKR Leukemogenesis: Identification and BiologicalSignificance of Thymic Lymphoma Receptors for AKR Retoviruses,” Cell, 17 (1979), 65–75.
34 Stehelin et al., “DNA Related to the Transforming Gene(s)” (above, n. 15).35 Cairns, Cancer, Science, and Society (above, n. 27).
hist3011.tex; 30/05/1997; 9:35; v.5; p.9

10 MICHEL MORANGE
the formation of tumors.36 The richness of the genetic background in thisorganism might have suggested that the way to understanding carcinogenesiswas paved by these initial studies in Drosophila; but such was not the case,and these studies were partially reevaluated only after the discovery of theantioncogenes. In a similar way, the evidence of a specific tumor antigen invirally transformed cells as well as in uninfected cancer cells appeared tobe a favored path toward a molecular understanding of cancer. Moreover,this antigen was associated with the protein coded by the transforming geneof the SV40 DNA oncovirus. Yet these observations were included in theoncogene paradigm only after a long delay and following the discovery ofantioncogenes.37
The abundance of theories and the complexity of data led at the end ofthe seventies to a pessimistic vision of cancer research. However, in 1982Jonothan Logan and John Cairns wrote that “this year may one day be seenas the year when a strong sense of order came to cancer research and the longdrive to understand the cancer cell really got underway.”38 What happened?What allowed the transition from the vague idea that gene dysfunction wasinvolved in oncogenesis to the conviction that the molecular basis of cancerhas been largely unveiled?
Sociological Explanation for the Development of the OncogeneParadigm
Joan Fujimura explains the development of the oncogene paradigm by thespreading of a new “theory-method package” that enhanced the articulationbetween the interests of biologists and the demands of medicine.39 This pack-age, built by molecular biologists, included standardized methods (recombi-nant DNA techniques) and theoretical “boundary objects,” the most importantof which was the oncogene concept.
The construction of this package was the result of a strategy by whichbiologists invested their expertise in areas promising them the most fruitfulreturn for their investment. Fujimura rightly outlines that the new program was
36 Elisabeth Gateff, “Malignant Neoplasms of Genetic Origin in Drosophila melanogaster,”Science, 200 (1978), 1448–1459.
37 There was an abundant literature on this tumor antigen. For example, see Daniel I. H.Linzer and Arnold J. Levine, “Characterization of 54K Dalton Cellular SV40 Tumor AntigenPresent in SV40-Transformed Cells and Uninfected Embryonal Carcinoma Cells,” Cell, 17(1979), 43–52; D. P. Lane and L. V. Crawford, “T Antigen Is Bound to a Host Protein inSV40-Transformed Cells,” Nature, 278 (1979), 261–263.
38 Jonothan Logan and John Cairns, “The Secrets of Cancer,” Nature, 300 (1982), 104.39 Fujimura, “Molecular Biological Bandwagon” (above, n. 2).
hist3011.tex; 30/05/1997; 9:35; v.5; p.10

FROM THE REGULATORY VISION OF CANCER TO THE ONCOGENE PARADIGM 11
an advantageous replacement for the Virus Cancer Program, which at the endof the 1970s was clearly a failure. Very few human cancers are due to infectionby a virus. Moreover, the Virus Cancer Program was criticized because it hadneglected the fundamental aspects of oncogenesis, the study of normal andtransformed cells.40 In this perspective, working on the oncogenes was, formany biologists, a good strategy for exploiting the results obtained by theirstudies of oncogenic viruses, and for recycling their expertise with new scopeand possibilities of medical applications in diagnostics and therapeutics.
However, specific criticisms can be addressed to the previous scheme,lowering its explanatory value. First, it is not true that the new paradigm wasan association of new techniques (recombinant DNA technology) and newconcepts such as cellular oncogenes. In fact, recombinant DNA technologywas “old” and had already been extensively used to characterize the oncogenicDNA or RNA viruses.41 The role of the new molecular technology in the areaof cancer research was justified very simply by John Cairns in 1979: “Biologyand cancer research have developed together. Invariably, at each stage, thecharacteristics of the cancer cell have been ascribed to some defect in whateverbranch of biology happens at the time to be fashionable and exciting; today,it is molecular genetics.”42 The concept of oncogenes also was not new;the suggestion that normal genes might become oncogenic, tumorigenic, bymutation had already been put forward years earlier.43
The explanation of the oncogene paradigm by the construction of a pack-age including a technology (recombinant DNA) and a concept (oncogene) istherefore mundane, unless the components of this package are more preciselydescribed. Such a precise description is necessary because other associationsbetween the oncogene concept and the new tools of DNA recombinant tech-nology were simultaneously proposed. For instance, George Todaro in 1980suggested using retroviruses to identify the genes involved in oncogenesis.44
By infecting transformed cells with retroviruses, it would have been possible
40 The criticisms concerning the scientific content of the program and its organization wereraised as early as 1971: “Special virus Cancer Program: Travails of a Biological Moonshot,”Science, 174 (1971), 1306–1310.
41 However, it is true that the development of the oncogene paradigm was parallel to thestandardization of the recombinant DNA technology, which allowed the rapid diffusion of thenew vision.
42 Cairns, Cancer, Science, and Society (above, n. 27), p. 63.43 Temin, “On the Origin of the Genes for Neoplasia” (above, n. 31).44 Ulf R. Rapp and George J. Todaro, “Generation of Oncogenic Mouse Type C Viruses:
In Vitro Selection of Carcinoma-Inducing Variants,” Proc. Nat. Acad. Sci. USA, 77 (1980),624–628. This is one among many other previous packages associating molecular technologiesand concepts from the cancer field. As early as 1972, Maurice Green wrote: “Until recently, theprospects for understanding the nature of cancer and for developing a rational basis to combatthis disease were dim. But now we can be more optimistic, for we are rapidly acquiringthe knowledge, techniques, experimental systems, and scientific insight to investigate and
hist3011.tex; 30/05/1997; 9:35; v.5; p.11

12 MICHEL MORANGE
to retrieve new genes involved in transformation and oncogenesis. The pro-posed program of research was a package including retroviruses and recom-binant DNA technology, together with the concept of cellular oncogene. Thisprogram of research did not lead to important developments. This exampleclearly shows that the explanation in terms of a technology/concept packageis presently underdetermined.
Two concepts used by Joan Fujimura deserve further scrutiny. The firstis the consideration of oncogenes as standardized and transportable bound-ary objects (immutable mobiles, according to Bruno Latour).45 Boundaryobjects are used to translate the interests of and facilitate the interactionsbetween members of different social worlds. In these different social worlds,the boundary objects have different structures, but the common structure issufficient to render them recognizable. It is true that the oncogene objectallowed different specialists, with their own separate viewpoints, to tacklethe same problem.46 However, it is hard to prove that the construction of theboundary object is causally explained by a deliberate strategy.
One important result of the social studies of science has been the attentionpaid to the tools used by scientists. To paraphrase the title of the book editedby Adele Clarke and Joan Fujimura, scientific successes result from the choiceof the right tool for the job.47 In the discovery of oncogenes, the transfectionassay is often mentioned as having played a major role. In the hands of RobertWeinberg, it allowed the first characterization of the mutations leading tothe formation of an active oncogene. It also allowed the demonstration thatmultiple steps, due to different classes of oncogenes, are necessary for celltransformation.48 The transfection assay has a higher demonstrative valuethan other technologies. Whereas other discoveries, such as the evidence for
to understand the molecular basis of neoplasia and to effectively use the new knowledgefor the prevention and cure of cancer” (Maurice Green, “Molecular Basis for the Attack onCancer,” Proc. Nat. Acad. Sci. USA, 69 [1972], 1036–1041); the technique was DNA-RNAhybridization, and the conceptual model the provirus model of Huebner and Todaro.
45 Susan Leigh Star and James R. Griesemer, “Institutional Ecology, ‘Translations’ andBoundary Objects: Amateurs and Professionals in Berkeley’s Museum of Vertebrate Zoology,1907–39,” Soc. Stud. Sci., 19 (1989), 387–420; Bruno Latour, “Visualization and Cognition:Thinking with Eyes and Hands,” in Knowledge and Society: Studies in the Sociology ofCulture Past and Present, ed. Henrika Kuklick and Elizabeth Long (Greenwich, Conn.: JAIPress, 1986), pp. 1–40.
46 Morange, “Discovery of Cellular Oncogenes” (above, n. 1).47 Clarke and Fujimura, Right Tools for the Job (above, n. 21).48 Hartmut Land, Luis F. Parada, and Robert A. Weinberg, “Tumorigenic Conversion of
Primary Embryo Fibrolasts Requires at Least Two Cooperating Oncogenes,” Nature, 304(1983), 596–602; H. Earl Ruley, “Adenovirus Early Region 1A Enables Viral and CellularTransforming Genes to Transform Primary Cells in Culture,” ibid., pp. 602–606; Hartmut Land,Luis F. Parada, and Robert A. Weinberg, “Cellular Oncogenes and Multistep Carcinogenesis,”Science, 222 (1983), 771–778.
hist3011.tex; 30/05/1997; 9:35; v.5; p.12

FROM THE REGULATORY VISION OF CANCER TO THE ONCOGENE PARADIGM 13
amplification or rearrangement of oncogenes in tumors, only suggested thatmodifications of oncogenes were associated with tumorigenicity, transfectiondemonstrated the causal relationship between oncogene alteration and celltransformation. Moreover, the transfection experiments were identical to theexperiments performed by Oswald Avery in the forties showing that DNAwas responsible for bacterial transformation.49 Robert Weinberg exploitedthis historical parallel in his review article published in 1983 in ScientificAmerican.50
However, the reference to transfection as the right tool for the job of char-acterizing oncogenes must be restricted. First, the transfection assays werenot new. They had already been used to characterize the provirus state ofoncogenic viruses.51 Moreover, the transfection tool never reached “trans-parency”: the moment when instrumentation is viewed as a reliable transmit-ter of nature’s messages;52 on the contrary, the real significance of in vitrotransformation and its relevance to tumorigenecity always remained the sub-ject of lively discussions. In fact, as we shall see later, the first transfectionexperiments by Weinberg and Cooper did not attract the interest of a largegroup of biologists. Only when the transfected oncogenes were shown to beidentical to the retrovirally encoded oncogenes – and, above all, when themodification leading to oncogenicity was demonstrated to be the replacementof only one amino acid in the protein – did these experiments arouse theinterest of biologists.
In conclusion, the description or explanations proposed thus far for thediscovery and characterization of oncogenes have underlined the role ofsome practical or theoretical tools and objects. However, these studies haveneglected to describe the complexity of the history of oncogene discovery,masking the precise timing of events in the creation of a mundane theo-ry/practice package. Moreover, as we shall see, these studies have completelyneglected the main component of the oncogene paradigm, its functional side.
49 Oswald T. Avery, Colin M. MacLeod, and Maclyn McCarty, “Studies on the ChemicalNature of the Substance Inducing Transformation of Pneumococcal Types,” J. Exp. Med., 79(1944), 137–158.
50 Robert A. Weinberg, “A Molecular Basis of Cancer,” Sci. Amer., 249 (November 1983),102–116.
51 See, for instance, one of the first successful assays in the domain of oncogenicity: M. Hilland Jana Hillova, “Virus Recovery in Chicken Cells Tested with Rous Sarcoma Cell DNA,”Nature New Biol., 237 (1972), 35–39. The efficiency of the technique was improved by F.L. Graham and A. J. Van der Eb, “A New Technique for the Assay of Infectivity of HumanAdenovirus DNA,” Virology, 52 (1973), 456–467.
52 Simon Schaffer, “Glass Works: Newton’s Prisms and the Uses of Experiment,” in TheUses of Experiment: Studies in the Natural Sciences, ed. David Gooding, Trevor Pinch, andSimon Schaffer (Cambridge: Cambridge University Press, 1989), pp. 67–104.
hist3011.tex; 30/05/1997; 9:35; v.5; p.13

14 MICHEL MORANGE
Alternative Tools and the Result of Their Use
In this section I shall propose other tools, some partially new and othersalready largely exploited by historians of science, to try to explain the accep-tance of the oncogene paradigm by the community of biologists. I shalldescribe, for each of these tools, the kind of information they provide, andshall emphasize their interests for the history of the oncogene concept.
Examination of the Scientific Literature
I want first to underline the importance of an accurate examination of thescientific literature. Many historians of science have attracted attention tothe major role of practice and informal exchanges in science. The interestof published literature has been questioned, as it is an already elaboratedform of science.53 Journal articles would represent a final stage of scienceconstruction, where the foundations have been deliberately rubbed out. If thiscriticism depreciates articles and reviews as the unique source of informationfor the historian, it does not justify their neglect in many science studies.Moreover, the criticisms addressed to the published literature are less pertinentwhen they concern a period of intense discoveries during which a new theoryis built: the hurry with which the papers are published makes them moretransparent.
In fact, a careful examination of published documents offers clues to recon-stitute the way in which a new theory was constructed. For instance, somearticles appear self-contradictory or deliberately equivocal: a subtle shiftoccurs between the reported results and the conclusions or summary of thearticle. I would like to give two examples of these internal contradictions.In their short review in Nature, Jonothan Logan and John Cairns describedthe two joint articles of Mariano Barbacid’s and Robert Weinberg’s groupsthat showed that, in transfection assays, the oncogenicity of the transferredoncogene, ras, was due to a point mutation in the coding sequence of the geneand, therefore, to a modification of the protein itself. In this previously citedreview, the authors placed the recent discoveries in the general frameworkof cancer studies, outlining the importance of the studies on tumorigenicretroviruses:
The tumorigenic retroviruses can cause cancer in at least two differentways. The virus can bring in an oncogene, which, because it is under thecontrol of the viral promoter of transcription, is inevitably transcribed at
53 Publication occurs at the end of a long process of simplification; see Susan Leigh Star,“Simplification in Scientific Work: An Example from Neuroscience Research,” Soc. Stud. Sci.,13 (1983), 205–228.
hist3011.tex; 30/05/1997; 9:35; v.5; p.14

FROM THE REGULATORY VISION OF CANCER TO THE ONCOGENE PARADIGM 15
too high a level. Alternatively, the virus need not contain an oncogene,but merely provide a strong promoter which, by chance, can then becomeintegrated next to one of the cell’s oncogenes; this is a much less efficientform of tumorigenesis and usually succeeds only after a long time interval.To summarize, the product of each cellular proto-oncogene seems to becapable of making a cell into a cancer provided that it is produced toexcess or has undergone a change in amino acid sequence.54
Even the poorly informed reader will realize that there is a discrepancybetween the conclusion (which in fact is imposed by the two linked articles),and the previously described work on retroviruses, which suggested thatoncogenes are activated by dysregulation and overexpression. This internalincoherence is easily explainable by the previous “faith” of the authors in thedysregulation model of cancer.
A similar ambiguity can been found in a review written by Michael Bishopin 1983. In the perspectives and summary, Bishop states that “their function[of oncogenes] in normal cells is not known, although the prevailing specu-lation holds that they are active in differentiation.”55 In the section entitled“The Role of Cellular Oncogenes in Normal Cells,” the reader is advised thatthe “pursuit of this question (are cellular oncogenes involved in differentia-tion?) : : : has been difficult and ineffectual. : : : No coherent picture emergesfrom these findings. : : : The discovery of cellular oncogenes in invertebrateshas further obscured the relationship between these genes and organismaldevelopment.56 Yet at the end of this section, in a very elegant about-face,Bishop concludes that “the paradox may be more apparent than real, how-ever, and its resolution may reveal unexpected principles of developmentalbiology,”57 This review was beautifully written and made an exhaustive sur-vey of the growing results on oncogenes. Its ambiguity cannot be attributedto some form of awkwardness, but reflects a conflict between the prevailingtheory that oncogenes were involved in cell differentiation and the actualexperimental results on the expression of oncogenes in different tissues andorganisms.
The contradiction may also lie, not within a single article or review, butbetween the content of an article and the way this article is referred to in theliterature. As an illustration, I have chosen a small group of articles showingthat cellular oncogenes – specifically, c-mos and c-ras – can transform cells
54 Logan and Cairns, “Secrets of Cancer” (above, n. 38), p. 104.55 J. Michael Bishop, “Cellular Oncogenes and Retroviruses,” Ann. Rev. Biochem., 52 (1983),
303.56 Ibid., pp. 332–333.57 Ibid., p. 334
hist3011.tex; 30/05/1997; 9:35; v.5; p.15

16 MICHEL MORANGE
when they are overexpressed.58 In most articles that cite these experiments,it is assumed that they clearly demonstrate that overexpression of a cellularoncogene leads to transformation.59 Therefore the transition of an oncogenefrom its normal state to its oncogenic state is due only to its overexpression.But close examination of the reported experiments shows that they have beenoverinterpreted: the level of transformation obtained by overexpression of anormal cellular oncogene remained lower than that obtained with the viralor mutated oncogene; moreover, the integrity of the cellular oncogenes wasnot properly checked in these experiments. Once more, it would be absurd todeduce that scientists are unable to correctly read an article. Most scientistswere conscious of the limits of these experimental proofs, but due to theirbelief in the dysregulation model of cancer, they underrated these limitations,considering them as minor and transient.
Analysis of Published Errors
Yet probably the most informative use of the published literature is the col-lecting of published errors, most of which are systematically eliminated fromsubsequent literature and frequently omitted in interviews.60 Natalie Angierhas pinpointed many errors occurring between 1977 and 1984 concerningthe structure, characterization, and function of oncogenes. It is not knownwhether all of these irreproducible results were errors, or whether some ofthem were due to a deliberate intent of fraud. The accusations of fraud havebeen consistently rejected by the main authors. Whatever the truth may be, Ithink that the distinction is of minor importance for the historian of science.The interest lies elsewhere. The multiplicity of errors and/or frauds revealsa state of agitation in this area of research, and the conviction of scientiststhat there is something to be “discovered.” This sense of an imminent break-through relaxes ordinary barriers. Moreover, errors and frauds both representinefficient strategies of scientists when the negotiations with the nonhumanactors have been deficient or even absent.
58 See above, n. 16.59 For instance, in George Klein, “The Role of Gene Dosage and Genetic Transpositions in
Carcinogenesis,” Nature, 294 (1981), 313–318; or in Geoffrey M. Cooper, “Cellular Trans-forming Genes,” Science, 218 (1982), 801–806. In a similar way, the fact that cellular onco-genes were roughly similar to viral oncogenes has been extrapolated to an identity of structure.All these extrapolations were pinpointed by Peter Duesberg: Peter H. Duesberg, “RetroviralTransforming Genes in Normal Cells?” Nature, 304 (1983), 219–226; idem, “Cancer Genes:Rare Recombinants Instead of Activated Oncogenes (A Review),” Proc. Nat. Acad. Sci. USA,84 (1987), 2117–2124.
60 Except in the book of Natalie Angier, Natural Obsessions (above, n. 17), where I havefound most of the following information. See also, for the “Spector affair,” William Broad andNicholas Wade, Betrayers of the Truth (New York: Simon and Schuster, 1982).
hist3011.tex; 30/05/1997; 9:35; v.5; p.16

FROM THE REGULATORY VISION OF CANCER TO THE ONCOGENE PARADIGM 17
The best way to look at the forces that were operative in the formation ofthe oncogene model under its present shape is to compare it to the erroneousmodels that were proposed during the same years as “unified visions of onco-genesis.” The most significant results leading to the triumph of the oncogeneparadigm are those present during the successful story and absent during theunsuccessful repetitions and anticipations.
The following list of errors is far from being exhaustive:
(1) The transfer of oncogenicity by isolated chromosomes in hamster cells.61
The first author, Demetrios Spandidos, was subsequently accused of fraudby the head of the laboratory, Louis Siminovitch. These results are there-fore not considered an anticipation of the transfection experiments per-formed by Cooper and Weinberg two years later.
(2) The characterization by Mark Spector of a cascade of a specific class ofenzymes, the tyrosine protein kinases, which was responsible for trans-formation, and in particular for the biochemical modifications occurringin cancer cells.62 Efraim Racker, the head of the group, along with someof Spector’s coworkers, made a public retraction.63
(3) The isolation and characterization by Shelly Bernstein in Robert Wein-berg’s laboratory of a human metastatic tumor DNA clone that was a newkind of oncogene responsible, not for transformation, but for the ability ofthe tumor cells to disseminate in the organism.64 The isolated clone was,in fact, ras, an oncogene previously studied in the laboratory. Despite the
61 Demetrios A. Spandidos and Louis Siminovitch, “Transfer of Anchorage Independenceby Isolated Metaphase Chromosomes in Hamster Cells,” Cell, 12 (1977), 675–682; idem,“Transfer of the Marker for Morphologically Transformed Phenotype by Isolated MetaphaseChromosomes in Hamster Cells,” Nature, 271 (1978), 259–261; idem, “The Relationshipbetween Transformation and Somatic Mutation in Human and Chinese Hamster Cells,” Cell,13 (1978), 651–662.
62 Mark Spector, Steven O’Neal, and Efraim Racker, “Reconstitution of the Na+K+ Pumpof Ehrlich Ascites Tumor and Enhancement of Efficiency by Quercetin,” J. Biol. Chem., 255(1980), 5504–5507; idem, “Phosphorylation of the �-Subunit of Na+K+-ATPase in EhrlichAscites Tumor by a Membrane-bound Kinase,” ibid., pp. 8370–8373; idem, “Regulation ofPhosphorylation of the �-Subunit of the Ehrlich Ascites Tumor Na+K+-ATPase by a ProteinKinase Cascade,” ibid., 256 (1981), 4219–4227; Mark Spector, Robert B. Pepinsky, VolkerM. Vogt, and Efraim Racker, “A Mouse Homolog to the Avian Sarcoma Virus src ProteinIs a Member of a Protein Kinase Cascade,” Cell, 25 (1981), 9–21; Efraim Racker and MarkSpector, “Warburg Effect Revisited: Merger of Biochemistry and Molecular Biology,” Science,213 (1981), 303–307.
63 Volker M. Vogt, Robert B. Pepinsky, and Efraim Racker, “Src Protein and the KinaseCascade,” Cell, 25 (1981), 827; Efraim Racker, “Warburg Effect Revisited,” Science, 213(1981), 1313.
64 Shelly C. Bernstein and Robert A. Weinberg, “Expression of the Metastatic Phenotype inCells Transfected with Human Metastatic Tumor DNA,” Proc. Nat. Acad. Sci. USA, 82 (1985),1726–1730.
hist3011.tex; 30/05/1997; 9:35; v.5; p.17

18 MICHEL MORANGE
vivid description of Natalie Angier, the origin of the confusion and anexplanation for the first promising results remain elusive.65
(4) The aforementioned discovery by Geoffrey Cooper that cellular onco-genes from normal cells transform with high efficiency when isolatedfrom their surrounding DNA sequences.66 There are no explanations forwhy these results differed from those of Robert Weinberg and the othergroups that used the same transfection assay.
(5) The evidence that the ras oncogene product forms a molecular com-plex with a receptor for a protein involved in iron transportation insidethe cell, transferrin.67 This apparent association was an artifact of theimmunoprecipitation technique used for the assay.68
(6) The molecular cloning and sequencing of a transforming gene from achicken lymphoma. These experiments could not be reproduced.69
Some of these “errors” were limited to one publication, and correspondedto overpreliminary results that were insufficiently checked. Such is the caseof the discovery of the oncogene involved in metastasis. Others, in contrast,relied on a continuous line of research and appeared in a series of publications.These errors were supported by a coherent set of experiments. Such is thecase of the results of Demetrios Spandidos, who published a series of differentworks using the chromosome transfer technique.70 The initial observation ofMark Spector concerned the modification of energy metabolism characteristicof transformed cells, and its explanation by the phosphorylation of a mem-brane enzyme. Later, he characterized the cascade of tyrosine protein kinasesresponsible for this modification, which he demonstrated to be the productsof different oncogenes. The results of Geoffrey Cooper on transfection werealso the end of a long line of research that showed that the oncogenic viruseswere subjected to a negative control by the flanking DNA sequences of the
65 Angier, Natural Obsessions (above, n. 17).66 Cooper, Okenquist, and Silverman, “Transforming Activity of DNA” (above, n. 14).67 Toren Finkel and Geoffrey M. Cooper, “Detection of a Molecular Complex between ras
Proteins and Transferrin Receptor,” Cell, 36 (1984), 1115–1121.68 Joe Harford, “An Artefact Explains the Apparent Association of the Transferrin Receptor
with a ras Gene Product,” Nature, 311 (1984), 673–675.69 Geoffrey M. Cooper and Paul E. Neiman, “Transforming Genes of Neoplasms Induced
by Avian Lymphoid Leukosis Viruses,” Nature, 287 (1980), 656–659; idem, “Two DistinctCandidate Transforming Genes of Lymphoid Leukosis Virus-Induced Neoplasms,” Nature,292 (1981), 857–858; Gerard Goubin, Debra S. Goldman, Judith Luce, Paul E. Neiman, andGeoffrey M. Cooper, “Molecular Cloning and Nucleotide Sequence of a Transforming GeneDetected by Transfection of Chicken B-Cell Lymphoma DNA,” Nature, 302 (1983), 114–119.
70 Demetrios A. Spandidos and Louis Siminovitch, “Linkage of Markers Controlling Con-secutive Biochemical Steps in CHO Cells as Demonstrated by Chromosome Transfer,” Cell,12 (1977), 235–242.
hist3011.tex; 30/05/1997; 9:35; v.5; p.18

FROM THE REGULATORY VISION OF CANCER TO THE ONCOGENE PARADIGM 19
host.71 Even the last two errors represent a coherent line of research, since theoncogene isolated from lymphoma coded for a protein with high homologywith transferrin.72 Moreover, many parallel experiments had shown that thetransferrin receptor was more active in transformed cells.73
What does each of these errors or frauds teach us about the history of theoncogene paradigm?
First the experiments of Demetrios Spandidos show that the idea of trans-fecting the transformed phenotype was not new (see earlier). In fact, it was anextension of the transfection of integrated retroviruses in the form of provirus-es. However, the transfection technique always required a large amount oftinkering. It never became mundane, never reached the status of a black box.74
These difficulties – the controversial results of Demetrios Spandidos, and thecomplex link between transformation and cancer – explain why this techniqueand its results were considered with suspicion. This preliminary conclusioncan be confirmed, as we shall see later, by an analysis of the Science CitationIndex.
Second the results obtained by Mark Spector show another version of whatmight have been the oncogene paradigm. Spector’s model was well received,and his cascade of protein kinases formed the cover of the collection ofabstracts of the RNA Tumor Viruses meeting in Cold Spring Harbor in May1981. This model was a biochemical model of oncogenesis. Regulation by aprotein kinase cascade was a sort of paradigm in metabolism. Moreover, asEfraim Racker insisted in his article sent to Science, this model confirmedthe observations of the eminent biochemist Otto Warburg and constituted themerger of biochemistry and molecular biology.75 In the oncogene paradigm,in agreement with the new vision of molecular biologists, the functions ofthe oncogenes are also important, but their action is interpreted at the level of
71 Geoffrey M. Cooper and Howard M. Temin, “Lack of Infectivity of the EndogenousAvian Leukosis Virus Related Genes in the DNA of Uninfected Chicken Cells,” J. Virol., 17(1976), 422–430; Geoffrey M. Cooper and Lauren Silverman, “Linkage of the EndogenousAvian Leukosis Virus Genome of Virus-Producing Chicken Cells to Inhibitory Cellular DNASequences,” Cell, 15 (1978), 573–577; Neal G. Copeland, Andrew D. Zelenetz, and GeoffreyM. Cooper, “Transformation by Subgenomic Fragments of Rous Sarcoma Virus DNA,” Cell,19 (1980), 863–870.
72 Goubin et al., “Molecular Cloning” (above, n. 69).73 References can be found in Finkel and Cooper, “Detection of a Molecular Complex”
(above, n. 67).74 For a critical discussion of the “black box” concept, see Kathleen Jordan and Michael
Lynch, “The Sociology of a Genetic Engineering Technique: Ritual and Rationality in thePerformance of the ‘Plasmid Prep,’ ” in Clarke and Fujimura, Right Tools for the Job (above,n. 21), pp. 77–114.
75 Racker and Spector, “Warburg Effect Revisited” (above, n. 62).
hist3011.tex; 30/05/1997; 9:35; v.5; p.19

20 MICHEL MORANGE
intracellular communication and regulation, not at the level of biochemicalreactions.
Third, the aforementioned transfection experiments of Geoffrey Cooperthat, in contrast to the results of Robert Weinberg, showed that DNA fromnormal cells could transform recipient cells, outline once again the dominantconception of cancer as dysregulation. In fact, this dominant regulatory visionstarted to lose ground only when Weinberg and other groups showed that themutation affects the protein coded by the oncogene, not its level of expression.
The three other errors are of a different nature. They occurred later, whenthe oncogene paradigm had already been accepted. They reflect the regu-lar tendency of scientists to generalize their results:76 oncogenes must beextractable from all transformed cells. The link between oncogene productsand growth-factor receptors (see below) ought to be extended to the impor-tant ras product. Metastasis – which, clinically speaking, is one of the mostdramatic aspects of cancer – also had to be explained in terms of onco-genes. These examples show the subtle “negotiations” necessary to define theboundaries of the new theory and its domain of application.
From the previous examples, some of the contextual elements surroundingthe birth of the oncogene paradigm have already been drawn: the feeling that anew theory was right around the corner, the previous dominant regulatory andbiochemical vision of cancer, the already mundane concept of the oncogene.The real question now is to determine how these different elements wereassembled, but also altered, in the final paradigm.
Analysis of the “Science Citation Index”
Assistance in following the active part of this process can be provided byanalysis of the Science Citation Index. This tool gives interesting informationon the extent and rate of diffusion of the new results.77 I would like to mentiononly the results of three studies that shed some new light on the weight of thedifferent contributions to the oncogene paradigm.
In the first study I looked at the diffusion of the results obtained by theWeinberg group (Figure 1). I have chosen three different articles: the first isthe successful transmission by transfection of the transformed character ofa chemically induced cancer (noted Shih et al. in the figure); the second isthe identification of the transforming gene of a human bladder carcinoma as
76 Lindley Darden, “Essay Review: Generalizations in Biology,” Stud. Hist. Phil. Sci., 27(1996), 409–419.
77 My use of the SCI is similar to that of Bruno Latour and Steve Woolgar in Laboratory life:The Social Construction of Scientific Facts, 2nd ed. (Princeton: Princeton University Press,1986); their book contains an extensive bibliography concerning the use of SCI.
hist3011.tex; 30/05/1997; 9:35; v.5; p.20
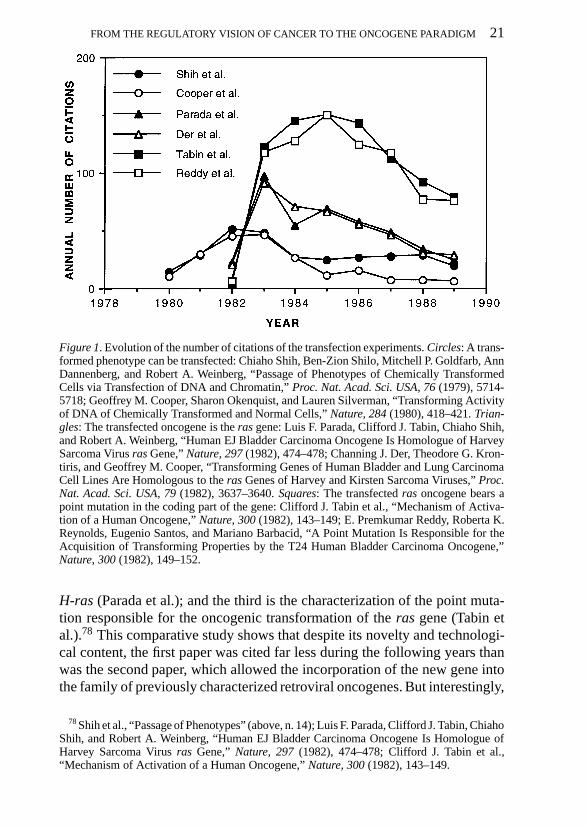
FROM THE REGULATORY VISION OF CANCER TO THE ONCOGENE PARADIGM 21
Figure 1. Evolution of the number of citations of the transfection experiments. Circles: A trans-formed phenotype can be transfected: Chiaho Shih, Ben-Zion Shilo, Mitchell P. Goldfarb, AnnDannenberg, and Robert A. Weinberg, “Passage of Phenotypes of Chemically TransformedCells via Transfection of DNA and Chromatin,” Proc. Nat. Acad. Sci. USA, 76 (1979), 5714-5718; Geoffrey M. Cooper, Sharon Okenquist, and Lauren Silverman, “Transforming Activityof DNA of Chemically Transformed and Normal Cells,” Nature, 284 (1980), 418–421. Trian-gles: The transfected oncogene is the ras gene: Luis F. Parada, Clifford J. Tabin, Chiaho Shih,and Robert A. Weinberg, “Human EJ Bladder Carcinoma Oncogene Is Homologue of HarveySarcoma Virus ras Gene,” Nature, 297 (1982), 474–478; Channing J. Der, Theodore G. Kron-tiris, and Geoffrey M. Cooper, “Transforming Genes of Human Bladder and Lung CarcinomaCell Lines Are Homologous to the ras Genes of Harvey and Kirsten Sarcoma Viruses,” Proc.Nat. Acad. Sci. USA, 79 (1982), 3637–3640. Squares: The transfected ras oncogene bears apoint mutation in the coding part of the gene: Clifford J. Tabin et al., “Mechanism of Activa-tion of a Human Oncogene,” Nature, 300 (1982), 143–149; E. Premkumar Reddy, Roberta K.Reynolds, Eugenio Santos, and Mariano Barbacid, “A Point Mutation Is Responsible for theAcquisition of Transforming Properties by the T24 Human Bladder Carcinoma Oncogene,”Nature, 300 (1982), 149–152.
H-ras (Parada et al.); and the third is the characterization of the point muta-tion responsible for the oncogenic transformation of the ras gene (Tabin etal.).78 This comparative study shows that despite its novelty and technologi-cal content, the first paper was cited far less during the following years thanwas the second paper, which allowed the incorporation of the new gene intothe family of previously characterized retroviral oncogenes. But interestingly,
78 Shih et al., “Passage of Phenotypes” (above, n. 14); Luis F. Parada, Clifford J. Tabin, ChiahoShih, and Robert A. Weinberg, “Human EJ Bladder Carcinoma Oncogene Is Homologue ofHarvey Sarcoma Virus ras Gene,” Nature, 297 (1982), 474–478; Clifford J. Tabin et al.,“Mechanism of Activation of a Human Oncogene,” Nature, 300 (1982), 143–149.
hist3011.tex; 30/05/1997; 9:35; v.5; p.21
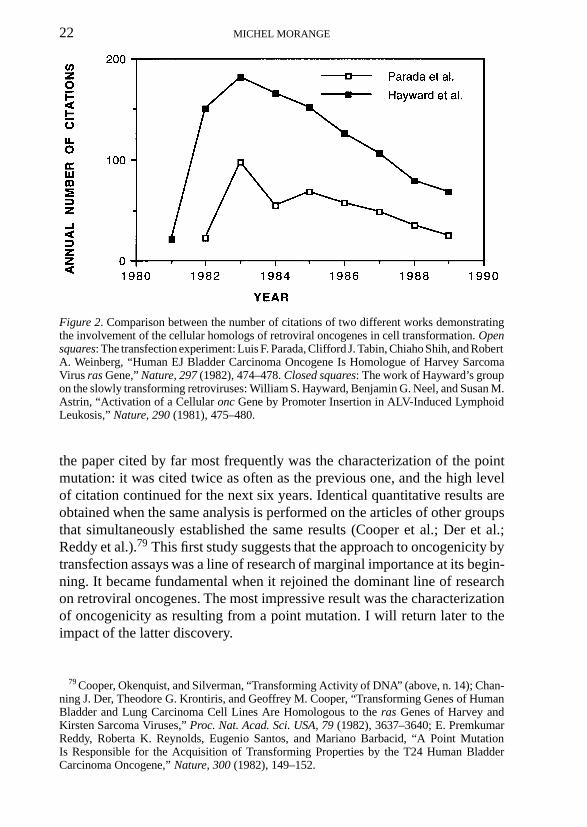
22 MICHEL MORANGE
Figure 2. Comparison between the number of citations of two different works demonstratingthe involvement of the cellular homologs of retroviral oncogenes in cell transformation. Opensquares: The transfection experiment: Luis F. Parada, Clifford J. Tabin, Chiaho Shih, and RobertA. Weinberg, “Human EJ Bladder Carcinoma Oncogene Is Homologue of Harvey SarcomaVirus ras Gene,” Nature, 297 (1982), 474–478. Closed squares: The work of Hayward’s groupon the slowly transforming retroviruses: William S. Hayward, Benjamin G. Neel, and Susan M.Astrin, “Activation of a Cellular onc Gene by Promoter Insertion in ALV-Induced LymphoidLeukosis,” Nature, 290 (1981), 475–480.
the paper cited by far most frequently was the characterization of the pointmutation: it was cited twice as often as the previous one, and the high levelof citation continued for the next six years. Identical quantitative results areobtained when the same analysis is performed on the articles of other groupsthat simultaneously established the same results (Cooper et al.; Der et al.;Reddy et al.).79 This first study suggests that the approach to oncogenicity bytransfection assays was a line of research of marginal importance at its begin-ning. It became fundamental when it rejoined the dominant line of researchon retroviral oncogenes. The most impressive result was the characterizationof oncogenicity as resulting from a point mutation. I will return later to theimpact of the latter discovery.
79 Cooper, Okenquist, and Silverman, “Transforming Activity of DNA” (above, n. 14); Chan-ning J. Der, Theodore G. Krontiris, and Geoffrey M. Cooper, “Transforming Genes of HumanBladder and Lung Carcinoma Cell Lines Are Homologous to the ras Genes of Harvey andKirsten Sarcoma Viruses,” Proc. Nat. Acad. Sci. USA, 79 (1982), 3637–3640; E. PremkumarReddy, Roberta K. Reynolds, Eugenio Santos, and Mariano Barbacid, “A Point MutationIs Responsible for the Acquisition of Transforming Properties by the T24 Human BladderCarcinoma Oncogene,” Nature, 300 (1982), 149–152.
hist3011.tex; 30/05/1997; 9:35; v.5; p.22

FROM THE REGULATORY VISION OF CANCER TO THE ONCOGENE PARADIGM 23
The previous conclusions may now be confirmed by turning to the relativeimpact of two different discoveries: the already-studied characterization ofthe bladder carcinoma oncogene as being H-ras (Parada et al.), and the obser-vation of William Hayward and colleagues in 1981 that the avian leukosisvirus, which is a slowly transforming virus harboring no oncogene, induceslymphoma by integrating in the proximity of the c-myc proto-oncogene andenhancing its expression (Figure 2).80 This last paper was cited twice as oftenas the paper of Robert Weinberg, reaching the very high value of 180 citationsin 1983. Interestingly, this article was not even mentioned by Weinberg in thereview he wrote for Cell in 1982.81 The result obtained by William Haywardwas in support of dysregulatory models of oncogenesis, but it also demon-strated that the retroviral oncogenes were implicated in forms of tumors otherthan those directly induced by the transforming retroviruses. Retrospectively,however, it was a more limited extension of the oncogene paradigm than theone proposed by Weinberg.
Finally, the third analysis of citations demonstrates that the functional sideof the oncogene paradigm was perhaps the most important to its acceptance.I compared the impact of five discoveries (Figure 3): the discovery in 1976 ofcellular oncogenes by Dominique Stehelin, Harold Varmus, Michael Bishop,and Peter Vogt (Stehelin et al.); the first characterization of cellular onco-genes by transfection by the groups of Weinberg and Cooper (Shih et al.;Cooper et al.); and three discoveries shedding some light on the functionsof oncogenes: the discovery in 1980 that the src protein is a kinase specificfor tyrosine (Hunter et al.), the evidence obtained by computerized sequencecomparisons that the sis oncogene is similar in structure to the already-knownPlatelet Derived Growth Factor (Waterfield et al.; Doolittle et al.), and thedemonstration, also by sequence analysis, that one of the oncogenes presentin the transforming avian erythroblastosis virus is a truncated form of thereceptor for Epidermal Growth Factor (Downward et al.).82 The results of my
80 William S. Hayward, Benjamin G. Neel, and Susan M. Astrin, “Activation of a Cellularonc Gene by Promoter Insertion in ALV-Induced Lymphoid Leukosis,” Nature, 290 (1981),475–480.
81 Robert A. Weinberg, “Fewer and Fewer Oncogenes,” Cell, 30 (1982), 3–4.82 Stehelin et al., “DNA Related to the Transforming Gene(s)” (above, n. 15); Shih et al.,
“Passage of Phenotypes” (above, n. 14); Cooper, Okenquist, and Silverman, “TransformingActivity of DNA” (above, n. 14); Tony Hunter and B. M. Sefton, “Transforming Gene Productof Rous Sarcoma Virus Phosphorylates Tyrosine,” Proc. Nat. Acad. Sci. USA, 77 (1980), 1311–1315; Michael D. Waterfield et al., “Platelet-Derived Growth Factor Is Structurally Relatedto the Putative Transforming Protein p28sis of Simian Sarcoma Virus,” Nature, 304 (1983),35–39; Russell F. Doolittle et al., “Simian Sarcoma Virus onc Gene v-sis Is Derived from theGene (or Genes) Encoding a Platelet-Derived Growth Factor,” Science, 221 (1983), 275–277;J. Downward et al., “Close Similarity of Epidermal Growth Factor Receptor and v-erb BOncogene Protein Sequences,” Nature, 307 (1984), 521–527.
hist3011.tex; 30/05/1997; 9:35; v.5; p.23
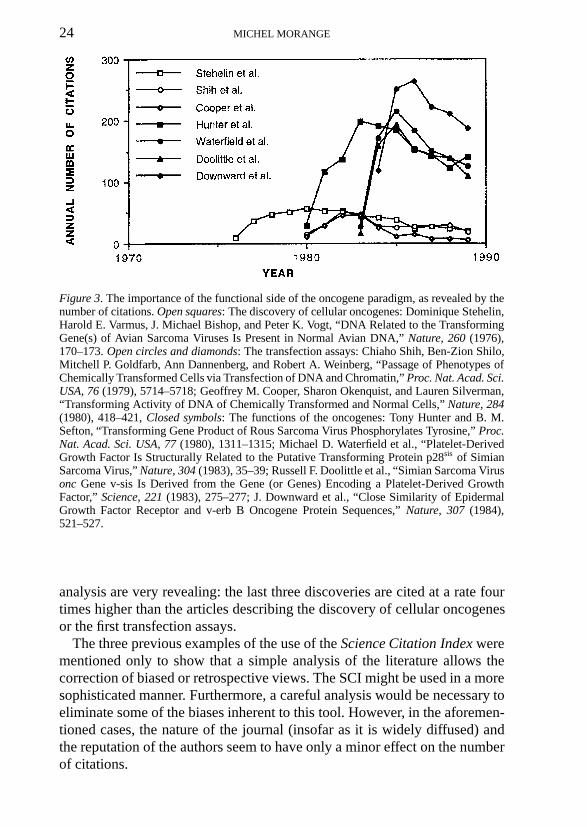
24 MICHEL MORANGE
Figure 3. The importance of the functional side of the oncogene paradigm, as revealed by thenumber of citations. Open squares: The discovery of cellular oncogenes: Dominique Stehelin,Harold E. Varmus, J. Michael Bishop, and Peter K. Vogt, “DNA Related to the TransformingGene(s) of Avian Sarcoma Viruses Is Present in Normal Avian DNA,” Nature, 260 (1976),170–173. Open circles and diamonds: The transfection assays: Chiaho Shih, Ben-Zion Shilo,Mitchell P. Goldfarb, Ann Dannenberg, and Robert A. Weinberg, “Passage of Phenotypes ofChemically Transformed Cells via Transfection of DNA and Chromatin,” Proc. Nat. Acad. Sci.USA, 76 (1979), 5714–5718; Geoffrey M. Cooper, Sharon Okenquist, and Lauren Silverman,“Transforming Activity of DNA of Chemically Transformed and Normal Cells,” Nature, 284(1980), 418–421, Closed symbols: The functions of the oncogenes: Tony Hunter and B. M.Sefton, “Transforming Gene Product of Rous Sarcoma Virus Phosphorylates Tyrosine,” Proc.Nat. Acad. Sci. USA, 77 (1980), 1311–1315; Michael D. Waterfield et al., “Platelet-DerivedGrowth Factor Is Structurally Related to the Putative Transforming Protein p28sis of SimianSarcoma Virus,” Nature, 304 (1983), 35–39; Russell F. Doolittle et al., “Simian Sarcoma Virusonc Gene v-sis Is Derived from the Gene (or Genes) Encoding a Platelet-Derived GrowthFactor,” Science, 221 (1983), 275–277; J. Downward et al., “Close Similarity of EpidermalGrowth Factor Receptor and v-erb B Oncogene Protein Sequences,” Nature, 307 (1984),521–527.
analysis are very revealing: the last three discoveries are cited at a rate fourtimes higher than the articles describing the discovery of cellular oncogenesor the first transfection assays.
The three previous examples of the use of the Science Citation Index werementioned only to show that a simple analysis of the literature allows thecorrection of biased or retrospective views. The SCI might be used in a moresophisticated manner. Furthermore, a careful analysis would be necessary toeliminate some of the biases inherent to this tool. However, in the aforemen-tioned cases, the nature of the journal (insofar as it is widely diffused) andthe reputation of the authors seem to have only a minor effect on the numberof citations.
hist3011.tex; 30/05/1997; 9:35; v.5; p.24

FROM THE REGULATORY VISION OF CANCER TO THE ONCOGENE PARADIGM 25
Examination of Sources Other Than Science Publications
The last tools for the historian are the information sources distinct from thepublished literature: correspondence, interviews, notes, and so forth. NatalieAngier’s book is an invaluable source on the errors or frauds that accompaniedthe discovery and characterization of cellular oncogenes.83 Moreover, byfollowing the reactions of Robert Weinberg on a day-to-day basis, Angiervery vividly illustrates that experiments do not always find what they aredesigned to look for. The comparison of the transforming oncogene isolatedby transfection with the previously described retroviral oncogenes was forWeinberg only a control, a somewhat superfluous experiment left to a student:the idea that the number of oncogenes was limited was not “natural.” In asimilar way, Weinberg insisted on looking for the mutation giving rise to thetransforming oncogene in the promoter of the gene, demonstrating that hehimself was convinced that cancer was a dysregulation. Yet documents suchas Angier’s book are not sufficient: they give a biased vision of the oncogenestory. Angier, for instance, by focusing on the laboratory of Robert Weinberg,overevaluates the work on transfection.
Lessons and Conclusions from This Study
The first conclusions concern the oncogene paradigm itself. This paradigmwas established between 1980 and 1984. It therefore did not immediatelyfollow the discovery of the cellular oncogenes, nor even the first transfectionexperiments.84 The latter became fundamental when they revealed the inter-vention in chemically induced or natural cancers of the same oncogenes asthose carried by retroviruses, and defined the molecular defects at the root oftransformation.
83 Angier, Natural Obsessions (above, n. 17).84 This point has already been discussed in Morange, “Discovery of Cellular Oncogenes”
(above, n. 1). Whereas the cellular oncogenes were discovered as early as 1976, the convictionthat cancer was due, whatever the triggering agent, to the modification of a limited number ofgenes involved in intracellular signaling, was only acquired later. One demonstration of thisis the fact that reviews on oncogenes appeared only after 1981. Three articles were publishedbetween 1982 and 1984 in the widely circulated review Scientific American: J. Michael Bishop,“Oncogenes,” Sci. Amer., 246 (March 1982), 69–78; Weinberg, “Molecular Basis of Cancer”(above, n. 50); and Tony Hunter, “The Proteins of Oncogenes,” Sci. Amer., 251 (August1984), 60–69. These three articles reflect the two main lines of research (on retroviruses andtransfection) that led to the characterization of oncogenes, as well as the functional side of theoncogene paradigm. Many other reviews were published in more specialized journals duringthe same years. Besides those already cited: J. Michael Bishop, “Enemies Within: The Genesisof Retrovirus Oncogenes,” Cell, 23 (1981), 5–6; Robert A. Weinberg, “Oncogenes of HumanTumor Cells,” Trends Biochem. Sci., 7 (1982), 135–136; Michael Bishop, “Cancer GenesCome of Age,” Cell, 32 (1983), 1018–1020.
hist3011.tex; 30/05/1997; 9:35; v.5; p.25

26 MICHEL MORANGE
In fact, two components of the oncogene paradigm were essential forthe adoption of the new vision. The first was the “discovery” that the numberof oncogenes was limited, and that the same oncogenes were altered indifferent forms of cancer. This is clearly revealed in the titles and contents ofthe reviews that appeared in 1982 or 1983: “Fewer and Fewer Oncogenes,”announced Weinberg in 1982.85 The fact that the number of oncogenes waslimited gave hope that a complete list of oncogenes was within the reach ofbiologists. This hope of simplification even led to the early erroneous viewthat this small number of oncogenes belonged to only one or two structuralfamilies.86
The second fundamental component of the oncogene paradigm was func-tional: the products of the oncogenes participated in the pathways, present inevery cell with a nucleus, that convey extracellular messages from the cellularmembrane to the nucleus. This functional part of the oncogene paradigm wasnot totally new, but it received strong experimental support and was rein-forced by the simultaneous characterization of these intracellular signalingpathways.87
On the other hand, the involvement of oncogenes in differentiation some-how vanished. The presence of these oncogenes in simple organisms such asyeast showed that the link between oncogenes and differentiation was not a
85 Weinberg, “Fewer and Fewer Oncogenes” (above, n. 81). This sense of limitation camefrom the discovery that the oncogenes discovered by different experimental approaches wereidentical: cellular oncogenes characterized by transfection were identical to retroviral onco-genes (Parada et al., “Human EJ Bladder Carcinoma Oncogene” [above, n. 78]; Der, Krontiris,and Cooper, “Transforming Genes” [above, n. 79]). The insertion of the Avian Leukosis Virusled to the activation of another cellular oncogene, myc (Hayward, Neel, and Astrin, “Activationof a Cellular onc Gene” [above, n. 80]). The multiplication of transfection assays showed thatthe same oncogenes were activated in various tumors from different animal species.
In 1982, it was also demonstrated that the same cellular oncogenes are amplified andoverexpressed in cells derived from some human tumors: Steven Collins and Mark Groudine,“Amplification of Endogenous myc-Related DNA Sequences in a Human Myeloid LeukemiaCell Line,” Nature, 298 (1982), 679–681.
Finally, the location of the same oncogenes at the breakpoints of the chromosomal translo-cations observed in human leukemias, and the demonstration that these translocations led tothe activation of their expression, were anticipated by G. Klein as early as 1981, before beingconfirmed step by step: Klein, “Role of Gene Dosage” (above, n. 59); Philip Leder et al.,“Translocation among Antibody Genes in Human Cancer,” Science, 222 (1983), 765–771.
86 See, for instance, Bishop, “Cellular Oncogenes and Retroviruses” (above, n. 55).87 These pathways extend from the growth-factor receptors to the nuclei. Essential were
the discovery of a new intracellular second messenger, IP3, and the characterization of proteinkinase C: Michael J. Berridge and Robin F. Irvine, “Inositol Triphosphate, a Novel Second Mes-senger in Cellular Signal Transduction,” Nature, 312 (1984), 315–321; Yasutomi Nishizuka,“The Role of Protein Kinase C in Cell Surface Signal Transduction and Tumour Promotion,”Nature, 308 (1984), 693–698.
hist3011.tex; 30/05/1997; 9:35; v.5; p.26

FROM THE REGULATORY VISION OF CANCER TO THE ONCOGENE PARADIGM 27
tight one.88 The role of oncogenes was not limited to one enzymatic reaction(such as in the kinase cascade model of Mark Spector), but linked with thedifferent intracellular signaling pathways. The discovery that a point mutationin the ras gene was sufficient to induce transformation was fundamental torecenter the general mundane vision of cancer as dysregulation to the morespecific conception of cancer as an alteration either in the level or in theactivity of the components forming the intracellular signaling network.
In fact, many conceptual and empirical difficulties of the new model weretransiently forgotten, “swept under the rug.” For instance, only a limited num-ber of cancerous cells gave rise to transforming genes in transfection assays.Also, and probably linked with the previous limit, only dominant oncogeneshad been characterized, whereas cell hybridization studies and many observa-tions on the transmission of human cancers suggested the existence of reces-sive oncogenes or antioncogenes.89 Moreover, the correspondence betweenthe nature of the activated oncogenes and the nature of the tumor was notsimple: the dominant oncogenes were neither universal nor limited to onekind of tumor. The complex interactions of cancer cells with the organism,and in particular with the immune system, and their progressive evolutiontoward higher levels of malignancy, had not been completely forgotten, butrather were discarded from the front line of research.
What does this very rapidly summarized story of oncogene research tell usnow about the tools and models used by the historians of science?
Two important aspects of the oncogene paradigm have been omitted fromprevious attempts to describe the explosive development of studies on onco-genes. The first is the existence of numerous studies on the functions of onco-genes. This omission is probably not an accident, but a deliberate strategy tomask the rationality at work in scientific research. Scientists are rationalists –but this does not prevent them from being opportunists, from exploiting whatis at hand. Their rationalism is open. It allows for wide flexibility: as we saw,concurrent and incompatible theories can be simultaneously cited withoutany scruples.90 But scientists are very satisfied when they have the impres-sion (perhaps the illusion?) that some natural mystery has been unraveled
88 Dieter Gallwitz, Cornelia Donath, and Christian Sander, “A Yeast Gene Encoding a ProteinHomologous to the Human c-has/bas Proto-Oncogene Product,” Nature, 306 (1983), 704–707;D. DeFeo-Jones, Edward M. Scolnick, R. Koller, and Ravi Dhar, “Ras-Related Gene SequencesIdentified and Isolated from Saccharomyces cerevisiae,” ibid., pp. 707–709.
89 See Alfred G. Knudson, Jr., “ Hereditary Cancer, Oncogenes and Antioncogenes,” CancerRes., 45 (1985), 1437–1443.
90 As beautifully described by Steve Woolgar, scientists are neither rationality-dopes, norinterest-dopes: Steve Woolgar, “Interests and Explanation in the Social Study of Science,”Soc. Stud. Sci., 11 (1981), 375.
hist3011.tex; 30/05/1997; 9:35; v.5; p.27

28 MICHEL MORANGE
and explained: the discovered functions of oncogenes explained the specificproperties of cancer cells.
The second important, previously omitted aspect of the establishment ofthe oncogene paradigm is the rhythm and form of discoveries and the natureof the results. One must recall that in a period of one year, the myc gene, apartfrom its previously demonstrated translocation by retroviruses, was found tobe the target of the slowly transforming viruses, to be overexpressed in manytumors, to be amplified in some of them, and to be precisely located at thepoint of chromosomal translocation occurring in one well-known form ofleukemia!91 Even the skeptic could hardly deny that there was coherence inthese very rapidly accumulating results. The nature of discovery also playeda role in the acceptance of the oncogene paradigm. The fact that the mutationactivating the ras oncogene in different tumors modified the same aminoacid and, moreover, produced the same replacement of a glycine by a valine,might not be due to chance.92 This conviction was strong, because the resultwas totally different from the predictions, which located the alteration in thegene promoter. The subsequent experiments that showed that other mutationsmight occur, affecting different positions in the protein, were insufficient toerase the initial revelation. As one of the biologists involved in oncogeneresearch remarked, “you couldn’t ask for a nicer, cleaner system than that totry to understand how the protein works.”93
Moreover, the way in which the functions of oncogenes were discoveredalso strengthened the feeling that the mechanisms of cancer had been unveiled.It was the computer-assisted comparison of sequences that suddenly providedthe simplest response for the long-awaited functions of oncogenes.94 Suchfortuitous discoveries reinforce the spontaneous realistic philosophy of sci-entists and give them greater confidence in the value of the results.
I do not want to argue that the establishment of the oncogene paradigmresulted only from the attainment by the biologists of a conviction. I simplywant to suggest that the rhythm and form of discoveries were importantparameters that cannot be neglected. In other words, the history of science isexactly that– history – and an understanding of scientific discoveries cannotbe reached without a careful account of the events.
91 Hayward, Neel, and Astrin, “Activation of a Cellular onc Gene” (above, n. 80); Collinsand Groudine, “Amplification of Endogenous myc-Related DNA Sequences” (above, n. 85);Leder et al., “Translocations among Antibody Genes in Human Cancer” (above, n. 85).
92 Tabin et al., “Mechanism of Activation of a Human Oncogene” (above, n. 78); Reddy etal., “Point Mutation” (above, n. 79).
93 Fujimura, “Constructing ‘Do-able’ Problems in Cancer Research” (above, n. 4), p. 268.94 Doolittle et al., “Simian Sarcoma Virus” (above, n. 82); Waterfield et al., “Platelet-Derived
Growth Factor” (above, n. 82); Downward et al., “Close Similarity of Epidermal GrowthFactor Receptor” (above, n. 82).
hist3011.tex; 30/05/1997; 9:35; v.5; p.28

FROM THE REGULATORY VISION OF CANCER TO THE ONCOGENE PARADIGM 29
Acknowledgments
I am indebted to Rosemary Sousa Yeh and David Cantor for critical readingof the manuscript; to Charles Galperin, Daniel Kevles, and Lindley Dardenfor giving me the occasion to discuss the ideas presented in this paper; andto Denise Ogilvie and Madeleine Brunerie for their help in consulting J.Monod’s correspondence.
hist3011.tex; 30/05/1997; 9:35; v.5; p.29

![Chromosomal Rearrangements and Oncogene Amplification ... · [CANCER RESEARCH 61, 1214–1219, February 1, 2001] Chromosomal Rearrangements and Oncogene Amplification Precede Aneuploidization](https://img.pdfslide.us/doc/110x75/6044c5937a1f9344c165f56e/chromosomal-rearrangements-and-oncogene-amplification-cancer-research-61-1214a1219.jpg)












![Presentazione standard di PowerPointmedia.aiom.it/userfiles/files/doc/AIOM-Servizi/slide/...paronychia, dermatitis, stomatitis,..) 63% (vs 41%) Oncogene Addicted [ARCHER 1050] Oncogene](https://img.pdfslide.us/doc/110x75/5e8e913ff7852e421e584c5b/presentazione-standard-di-paronychia-dermatitis-stomatitis-63-vs-41.jpg)














