Embed Size (px)
Citation preview

From Microbiology and Tumor Biology Center Karolinska Institutet, Stockholm, Sweden
Health hazards associated with dissemination of bacterial strains in waste water recycling
By
Mokhlasur Rahman
Stockholm 2005

All previously published papers were reproduced with permission from the publisher. Published and printed by Karolinska University Press Box 200, SE-171 77 Stockholm, Sweden © Mokhlasur Rahman, 2005 ISBN 91-7140-468-6

This work is dedicated
TO MY FAMILY


ABSTRACT
Treated waste effluents with low levels of chemical and microbiological contents are
used for domestic, industrial, agricultural and aquacultural purposes worldwide, and it is
estimated that one tenth or more of the world’s population consume food produced through
irrigation with wastewater. Treated hospital waste effluents may contain pathogenic and drug
resistant bacteria, which constitutes the most dangerous single risk factor for dissemination of
pathogenic and drug resistant organisms to the environment. This thesis focuses on the
possibility of persistence and transmission of pathogenic and drug resistant bacteria like
Aeromonas and Enterococcus from wastewater to environment and to humans, especially in
relation to a sewage treatment process based on waste water recycling.
In one study (Paper I), we have shown that a prevalent ampicillin and ciprofloxacin
resistant clonal lineage of Enterococcus faecium in Swedish hospital sewage water may be
transmitted from hospital patients to hospital sewage water. Recycling of hospital sewage water
may in the second step disseminate the disease-causing organisms to the environment and in the
third step to the human food chain. In other studies (Papers III, V), we have investigated the
persistence of Aeromonas in a duckweed aquaculture based hospital sewage water treatment
plant, where sewage grown duckweed is used as fish food. In this treatment system, the numbers
of Aeromonas organisms, diversities and resistance patterns to different antibiotics were found to
reasonably decrease through the purification process, with the exception of the fish culture pond
where a high prevalence of resistance was observed. We found that a phenotypically and
genotypically stable clonal lineage of Aeromonas could survive in the treatment system for
years. Both virulent and avirulent clonal lineages of Aeromonas could be transmitted from
hospitalised children suffering from diarrhoea to the fish that will be consumed by human
through recycling of hospital sewage water. It was also shown that transport vehicles such as
duckweed and amoebas (if present) might play a role in this transmission. We were able to
detect a gentamicin resistant (multiresistant) clonal lineage that was transmitted from hospital
sewage water to fish. Furthermore, we found that a transmitted clonal lineage of Aeromonas had
a high biofilm formation ability compared to other Aeromonas clones. As in other bacteria, we
could show that transition between biofilm formation and motility of one strain of this clone is
regulated by the cyclic di-guanosine-monophosphate concentration. We also characterized
Aeromonas from fish with Epizootic Ulcerative Syndrome disease and showed that one
pathogenic clonal group was associated with this fish disease in Bangladesh (Paper II).
To facilitate the monitoring of resistance in different sites of the waste treatment plant,
we developed a user-friendly semi automated MIC determination method, the ScanMIC method,

to screen a large number of isolates in a short time (Papers IV, V). We compared this method
with a reference method and in both cases the ScanMIC method was found to meet the
performance criteria guidelines for automated susceptibility testing.
In conclusion, the results indicate that sewage water recycling in aquaculture constitutes
a potential health hazard through dissemination of pathogenic and drug resistant bacteria to the
environment and/or to human.


TABLE OF CONTENTS
LIST OF PUBLICATIONS............................................................................................ 3
ABBREVIATIONS ........................................................................................................ 4
1 INTRODUCTION .................................................................................................... 5 1.1. Waste water and treatment systems 5
1.1.1 Waste water................................................................................................. 5 1.1.2 Treatment of waste water ............................................................................ 5 1.1.2.1 Activated sludge .......................................................................................... 6 1.1.2.2 Waste stabilization ponds ............................................................................. 6 1.1.2.3 Duckweed based waste treatment system....................................................... 7
1.2. Aquaculture 7 1.2.1 Aquaculture as a source of protein.............................................................. 7 1.2.2 Duckweed as a feed for aquaculture ........................................................... 8
1.3. Transmission and health 8 1.3.1 Transmission of pathogen from waste water to human .............................. 8 1.3.2 Water and/or food borne pathogen.............................................................. 9 1.3.3 Hazard identification for food safety .......................................................... 9 1.3.4 Bacterial typing in epidemiological surveillance...................................... 10 1.3.5 The host-pathogen relationship on waste related infections ..................... 11
1.4. The Bacterial Genus Aeromonas 12 1.4.1 Opportunistic pathogen ............................................................................. 13 1.4.2 Historical aspects of Aeromonas taxonomy.............................................. 13 1.4.3 New Species of Aeromonas ...................................................................... 13 1.4.4 Genospecies .............................................................................................. 14 1.4.5 Identification to the genus level ................................................................ 14 1.4.6 Identification to the species level.............................................................. 14 1.4.7 Ecology of Aeromonas .............................................................................. 16 1.4.7 Pathogenicity of Aeromonas ..................................................................... 17 1.4.8 Putative virulence factors of Aeromonas .................................................. 18 1.4.9. Biofilm formation of Aeromonas ............................................................. 21
1.5. Enterococci 22 1.5.1 Genus description of Enterococcus........................................................... 22 1.5.2 Habitat of Enterococcus ............................................................................ 22 1.5.3 Clinical diseases and nosocomial infections by Enterococcus ................. 22
1.6. Antibiotic resistance 23 1.6.1 Antibiotic susceptibility testing and MIC breakpoint ............................... 23
2 AIMS OF THE STUDY ......................................................................................... 25
3 MATERIALS AND METHODS............................................................................ 27
4 RESULTS AND DISCUSSION.................................................................... 32 4.1 Transmission of Enterococcus faecium from hospital patients to environment 32 4.2 A clonal lineage of Aeromonas is associated with fish diseas 34 4.3 Persistence and transmission of Aeromonas in the sewage treatment plant 36 4.4. General comments on duckweed aquaculture based sewage treatment plant 45 4.5 Usefulness of “ScanMIC” method for antibiotic related hazard identification 48
1

4.6 Cyclic-di-GMP signalling in Aeromonas (preliminary data) 50 5 ABSTRACTS OF INCLUDED PAPERS..............................................................54
6 ACKNOWLEDGEMENTS ....................................................................................57
7 REFERENCES.......................................................................................................58
2

LIST OF PUBLICATIONS
This thesis is based on the following original papers, which will be referred to in the text by
their Roman numerals:
I. Aina Iversen, Inger Kühn, Mokhlasur Rahman, Anders Franklin, Lars G. Burman,
Barbro Olsson-Liljequist, Erik Torell and Roland Möllby. Evidence for
transmission between humans and the environment of a nosocomical strain of
Enterococcus faecium. Environmental Microbiology 2004; 6: 55-59.
II. Mokhlasur Rahman, Patricia Colque-Navarro, Inger Kühn, Geert Huys, Jean Swings
and Roland Möllby. Identification and characterization of pathogenic
Aeromonas veronii biovar sobria associated with Epizootic Ulcerative Syndrome
in fish in Bangladesh. Applied and Environmental Microbiology 2002; 68: 650-655.
III. Mokhlasur Rahman, Geert Huys, Motiur Rahman, John Albert, Inger Kühn and
Roland Möllby. Persistence and transmission of Aeromonas in a duckweed
aquaculture-based hospital sewage water recycling plant. Submitted to the
Applied and Environmental Microbiology.
IV. Mokhlasur Rahman, Inger Kühn, Motiur Rahman, Barbro Olsson-Liljequist and
Roland Möllby. Evaluation of a scanner assisted colorimetric MIC method for
susceptibility testing of Gram negative fermentative bacteria. Applied and
Environmental Microbiology 2004; 70: 2398-2403.
V. Mokhlasur Rahman, Inger Kühn, Motiur Rahman, Geert Huys, and Roland Möllby.
Influence of a duckweed-aquaculture based hospital sewage water recycling
plant on the antibiotic resistance of Aeromonas spp. as determined by the
ScanMIC method. (Manuscript)
3

ABBREVIATIONS
AFLP Amplified Fragment Length Polymorphism
ARE Ampicillin Resistance Enterococci
ASL N-Acyl-L-Homoserine Lactones
c-di-GMP Cyclic di Guanosine Monophosphate
EPS Extracellular Polymeric Substance
EUS Epizootic Ulcerative Syndrome
FAME Fatty Acid Methyl Ester
GDP Gross Domestic Products
GFP Green Fluorescent Protein
HACCP Hazard Analysis Critical Control Point
HSL N-butanoyl-L-Homoserine Lactone
MIC Minimum Inhibitory Concentration
PFGE Pulsed Field Gel Electrophoresis
UPGMA Unweight Pair Group Method
WSP Waste stabilization pond
4

INTRODUCTION
Due to growing populations, increased human consumption and urbanization,
freshwater reserves are decreasing in many developing countries (33, 102). To address these
problems, treated sewage water with a low level of chemical and microbiological
contaminations has been used for domestic, industrial, and agricultural purposes (33, 100).
Worldwide it is estimated that 40% of all food is produced with irrigated water and at least
20 million hectars of land in 50 countries are irrigated with raw or partially treated waste
water (33, 59). One tenth or more of the world’s population consume foods produced by
irrigation with wastewater (172). There are several kinds of waste purification systems
available, but most of them are too costly to provide solution for sewage water purification
in growing developing countries (61). In most developing countries only 10% of all
wastewater is subject to purification (79). Duckweed based waste treatment systems are a
relatively new concept of waste purification since sewage grown duckweed could be used as
feed in fish farm. This thesis focuses on the possibility of persistence and transmission of
pathogenic and drug resistant bacteria like Aeromonas and Enterococci from the wastewater
to environment and to humans, especially in relation to a sewage treatment process based on
waste water recycling.
1.1. Waste water and treatment systems 1.1.1 Waste water
Water from households, industries, and run off water comprise the municipal waste
water. Most of the waste water contains a number of organic substances that are more or less
biodegradable (64). In the waste water, microorganisms consume oxygen to use the organic
substance as energy sources and combust them to carbon dioxide and water. As a result, the
reduction of oxygen concentration creates anaerobic conditions in the waste water that may
be toxic to higher forms of life in the natural recipient, which is one of the major problem
with the effluent of untreated waste water (49). Furthermore, excessive nutrient enrichment,
mainly nitrogen and phosphorous contributes to the eutrophication of the natural water. The
consequences of the eutrophication are large algae blooms, sometimes poisonous, and
oxygen depletion in the bottom sediments, which leads to a diminishing bottom fauna (64). It
is thus of great importance to decrease the nutrient loads in order to restore the natural
conditions of water.
1.1.2 Treatment of waste water Treatment of waste water implies reduction of the level of floating and suspended
solids, organic materials such as nitrogen and phosphorus and elimination of the load of
pathogenic microorganisms (81). In a conventional municipal waste treatment plant, the
5

sewage is first passed through a series of screens or strainers to remove floating materials
and solids. After this removal, suspended solids are settled in the primary sedimentation. The
wastewater then enters the biological step in the treatment plants. There are two main types
of biological treatment systems: biofilters and activated sludge. Both of them are based on
removal of organic substances with biological removal of nitrogen. The biological unit is
sometimes followed by a second sedimentation and in some cases also chemical treatment
before letting out the effluent into the receiving water.
1.1.2.1 Activated sludge Activated sludge treatment of wastewater has been around for about 100 years in one
form or another, and is hither the most successful mean of achieving pollution control. The
activated sludge separator is composed of two basic compartments: the aerator reactor and
the sludge separator. Aerator reactor is used to mix the microorganisms and sludge that lives
in the plant. The sludge contains microorganisms, inorganic and organic substances. A mass
of activated sludge is kept moving in the water by stirring. The overall goal of the activated-
sludge process is to remove substances that have a demand for oxygen from the system.
Microorganisms play an important role in the waste treatment process and are responsible for
removal of organic substances, such as nitrogen. Bacteria constitute the majority of
microorganisms present in activated sludge. The organic material is oxidised by
heterotrophic bacteria to CO2 and water. Nearly every city in the world uses a form of this
treatment, because it is a relatively simple technology. But these systems are energy
consuming and they require trained personal for operation and maintenance, therefore this
system is too costly for developing countries.
1.1.2.2 Waste stabilization ponds Waste stabilization ponds (WSP) are generally the waste water treatment process of
first choice in developing countries. They are suitable for both large and small populations
(from a few hundreds to hundreds of thousands). WSP have several important advantages for
developing countries: low capital costs, simple operation and maintenance, and high
performance. They can easily be designed to produce high-quality effluents suitable to
irrigate crops, support fish farms, for aquatic vegetable culture or simply to control river
pollution. The principal of the system are it is an entirely natural method of wastewater
treatment (they obtain all their energy directly from the sun) but the disadvantage is that it
requires much more land than conventional processes such as activated sludge. WSP
treatment systems have been used in different countries, such as Turkey, Cyprus, Israel,
Jordan, Egypt, Tunisia, Algeria, Morocco, Portugal, Spain, France, Greece, as well as at
several sites in India. However, land increases in value over time and in an overpopulated
developing country like Bangladesh municipalities cannot afford the land needed for waste
6

stabilization ponds. Moreover, in Bangladesh, like in many other developing countries,
planning for domestic waste water reuse is one area that has received special attention.
1.1.2.3 Duckweed based waste treatment system Current mainstream technologies for sewage water treatment are too costly to
provide a satisfactory solution for the growing sewage water problem of developing
countries (61). Cost effective, environmental friendly, macrophyte-based sewage water
treatment systems like the duckweed based waste treatment system is a relatively new
concept and is becoming more common practice in different south East Asian countries (
such as Thailand, Malaysia, Taiwan and Bangladesh) as well as in China and USA (7, 80).
Duckweeds are small aquatic plants belonging to the family Lemnaceae, which may be used
as food for livestock and fish. The use of free-floating duckweed in sewage water treatment
systems has been advocated for, since it is fast growing and easy to harvest, has a low fiber
and a high protein content, and shows a high efficiency in removal of nitrogen and
phosphorous from sewage water (111). The duckweed based waste treatment system uses
sewage water as a nutrient source for duckweed production. In this way waste water is
transferred into high quality protein instead of being disposed in the environment thus
solving the waste purification problem. The growing interest from the potential cost recovery
through the use of duckweed as animal feed mostly for intensively cultured duck, chicken or
aquaculture farms. Thus, the duckweed serves two purposes: it is purifying the wastewater as
well as being used as animal feed.
1.2. Aquaculture 1.2.1 Aquaculture as a source of protein
Aquaculture is the fastest growing sector among the world’s food production sectors.
According to the latest FAO statistics, the contribution of aquaculture to global supplies of
fish, crustaceans and molluscs continues to grow, increasing from 3.9% of total production
by weight in 1970 to 29.9% in 2002. Total world aquaculture production was reported to be
51.4 million tons by volume and 60.0 US$ billion by values in 2002 and countries in Asia
accounted for 91.2% of the production and 82% of these values. The major aquaculture
productions of fish, crustaceans and molluscs continue to come from the freshwater
environment (57.7% by volume and 48.4% by values) (177).
Fish is an important part of the diet for a large proportion of the people living in the
developing world. Many types of fish and aquatic products are available at affordable prices
in developing countries. At present, food fish represents the primary source of animal protein
(contribution more than 25% of the total animal protein supply) for a one billion people
within 58 countries worldwide. Fish are important source of many nutrients, including
protein of very high quality, Vitamin A, vitamin D, vitamin E, iodine and selenium.
7

However, affordability is a key factor explaining why aquaculture is making an essential
contribution in the developing world (177). In the overall agriculture based economy of
Bangladesh, aquaculture supplied 80 % of all animal protein produced, and it contributed to
4.7 % of Gross Domestic Products (GDP). The contribution of fisheries in Bangladesh is
very promising and important for creating job opportunities for unemployed people, earning
foreign exchange, alleviating poverty and improving nutritional status of the people.
1.2.2 Duckweed as a feed for aquaculture In developing countries like Bangladesh, in spite of limited resources, aquaculture
provides considerable amounts of high quality protein. Environmental pollution, abused use
of pesticides (about 7000 metric tons per year in agricultural fields, of which 25% are being
washed into open water bodies) and fish diseases are now depleting the fisherie’s resources
in the open water bodies in Bangladesh. In the future, farmed fish will be a more important
source of protein than they are today. Day by day integrated fish farming is getting more
popular in Bangladesh and also throughout Southeast Asia.
The integrated fish farming system are relatively confined units with a high density
fish farming. The fish used to receive supplementary feed and, in most cases, fresh extract of
animals which supports the growth of plankton, the natural food for fish, thus reduced the
cost of feed and fertilizers for fish ponds. But using animal waste in fish culture is not
socially accepted. Thus there is an increased demand of low cost or natural production of
fish food in many developing countries like Bangladesh. In this context, the use of duckweed
as fish feed has attracted much attention, since it can grow on waste water, which does not
cost any investment. The duckweed aquaculture based fish farming system maximizes
production through recycling of waste water, optimum utilization of resources and a
maximum return with minimum cost involved in fish culture.
1.3. Transmission and health 1.3.1 Transmission of pathogen from waste water to human
Protection of public health is the fundamental purpose of waste treatment.
Environmental protection is the second major purpose. Waste water is known to be possible
reservoirs of pathogenic and/or drug resistant bacteria and other pathogenic organisms.
Pathogenic organisms can be transmitted from wastewater to human through direct and
indirect transmission. Direct transmission of pathogens could happen when drinking water or
swimming water is contaminated with waste water. Indirect transmission could happen when
waste water is used for agricultural and aquaculture purposes.
Management and treatment of waste water have largely been insignificant in
preventing pollution from affecting surface water quality on both local and global scales (6).
More recent studies in developing nations have demonstrated that breaking pathogen
8

transmission cycles through proper sanitation and sewage management improve health
benefits more than only simple provision of clean drinking water (50) The treatment and
disposal of waste water is becoming more and more important since recent studies have
shown that health problem often are related to inadequate sewage treatment. Due to the lack
of proper treatment of sewage water in developing countries, inadequately treated sewage
water has been used/recycled directly or indirectly in agriculture and aquaculture.
1.3.2 Water and/or food borne pathogen In developing countries with inadequate water supplies, transmission of pathogen
from waste water to human is mainly responsible for food and water borne diseases (faecal
oral transmission). For the past decade, the increase in food born and water borne infections
has become an important public health concern worldwide (133). Besides the classical food
and water borne pathogens such as Clostridium, Salmonella, Shigella, Vibrio a number of so
called new or emerging pathogens have recently been identified as food poisoning organisms
such as E.coli 0157:H7, Camplylobacter, Listeria and Aeromonas (133). The number of
identified food and water borne pathogens since the 1950s has nearly trebled. Several newly
recognized food and water borne pathogens have contributed greatly to the occurrence of
infectious diseases during the last two decades.
Public health authorities have estimated that half the population of the developing
world is suffering from the major microbial diseases associated with water supply and
sanitation (194, 195). Infection by pathogenic bacteria, protozoa, and viruses are the most
prevalent global health risks associated with waste water. However, common organisms in
drinking and waste water that have been identified as posing major threats to human health
include (i) bacteria: enteropathogenic Escherichia coli (notably E. coli 0157:H7), Vibrio
cholerae, Shigella, Campylobacter jejuni, Salmonella, Yersinia enterocolitica, (ii)
protozoans: Giardia lamblia, Cryptosporidium parvum, Entamoeba histolytica, Toxoplasma
gondii, Balantidium coli, and (iii) viruses: Norwalk and Norwalk-like, Rotavirus, Hepatitis A
and E; (26, 53, 68, 99, 130, 137)
1.3.3 Hazard identification for food safety Control and prevention of food and water borne diseases can be difficult. To
increase food safety, it is important to identify pathogens or situations that may lead to food
borne illness and to determine the magnitude of the impact these have on human health
(118). Such information is needed for the effective use of resources, since hazard analysis
critical control point (HACCP) programs can identify the hazards and might play an
important role in reducing food borne pathogens. The term “hazard” refers to a biological
agent, that is, the microorganism and/or its toxins, that has the potential to cause adverse
health effects (118). A systematic examination is necessary to effectively manage of food
9

safety. Risk assessment is a process that provides an estimation of the probability and impact
of adverse health effects attributable to potentially contaminated foods (124). Risk
assessment is a science-based investigation consisting of four steps: hazard identification,
exposure assessment, hazard characterization and risk characterization. Thus the hazard
identification is the primary and fundamental step in risk assessment (37).
Hazard Identification What agents are present in the food and capable of causing adverse health effects
Exposure Assessment Consumption, Pathogen in food serving
(occurrence, density), Predictive microbiology (growth/decline), ingested number of
pathogens
Hazard Characterization What is the nature of adverse effect ?
Dose response assessment (non-threshold model, threshold model)
Risk Characterization and Risk Estimation Probability and severity of illness attributable to the
food/pathogen source; e.g. no of illness per year and per 100 K population
1.3.4 Bacterial typing in epidemiological surveillance
Identification and characterization of isolates below the species level is important in
epidemiological studies, e.g. for tracing the source of infections and for identification of
epidemic clones. Bacterial typing has been used for different purposes in epidemiology, but
most reports concern the use of bacterial molecular typing method for investigation of an
outbreaks and route of transmission (18, 44, 67). In the investigation of outbreak, a limited
number of isolates from a limited number of samples/persons are typed to identify which
isolates are clonally related (i.e., directly descended from a common ancestor). In this case
each isolate is compared to each other. This application implies that the same
epidemiological markers are used consistently: markers must be standardized and
reproducible over time. Such typing is called library typing. Any library typing results has a
universal meaning for the epidemiologist.
Molecular epidemiology is based on one postulate. When several isolates are
deemed identical using a molecular typing method with a high discriminatory power, it
10

assumed that they are derived from a common ancestor. Thus in a molecular epidemiological
investigation, identification of cases will be with identical strains for which the transmission
can be confirmed by epidemiological links. When a typing method with a high
discriminatory power is tested on a set of epidemiological unrelated isolates, one must
considered the existence of highly prevalent clones in the population of the microorganisms
under study. In this situation, epidemiological unrelated cases might be harbouring a strain of
identical genotype. The predominant clone might have been present for a long period of
time, have had time to spread without significant mutations or might have spread very
rapidly compared to other strains. Several examples of such predominant clones have been
described. For example, epidemiological unrelated S. aureus isolates with similar PFGE
patterns have been found in different geographical regions suggesting that some genotypes
are ubiquitous (65, 165). Another example is the species Legionella pneumophila where
isolates showing similar PFGE patterns have been recovered from many patients and from
different parts of Paris water systems (119). Moreover, this predominate genotype, called the
“Paris strain” has been found between 1998 and 2002 in patients from all over France (15).
Thus molecular typing enables us the tracking of the dissemination of specific clones with or
without epidemiological links (19)
1.3.5 The host-pathogen relationship in waste related infections Waste related microbial water and food borne disease may occur when a susceptible
individual consume water and/or food contaminated by a viable microbial pathogen(s) and/or
microbial toxins . However, not every exposure to a pathogen in water and/or food will result
in infection or illness and not all individuals in a given population are equally susceptible to
all pathogens. Therefore the risk of food and/or water borne disease is a combination of the
likelihood that exposure will result in infection or intoxication and subsequently illness and
the severity of the illness.
11

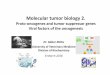
Figure 1 The pathogen-host relationship and possible transmission routes for sewage related
infections. Adapted from Martin Strauss (126)
1.4. The Bacterial Genus Aeromonas
The genus Aeromonas were at first recognized as pathogens of cold blooded animals
(fish, frogs, snakes, lizards) that suffered from Aeromonas associated septicaemia (16, 112,
168,169). In fact, nonmotile psychrophilic Aeromonas (originally named Bacterium
salmonicida but today known as Aeromonas salmonicida) was first identified as the primary
cause of fish furunculosis, a disease leading to severe economical losses in fish farm (174,
175, 197). The first human infection with an Aeromonas strain was reported by Hill and
colleagues in 1954 (78). From that date to until today, clinical microbiologists recovered
various Aeromonas species from a wide range of infection sites. Hundreds of reports,
varying from single case studies to long term monitoring surveys, have revealed a
relationship between high Aeromonas densities in drinking water and diarrhea in humans
(14, 21, 31). Gastrointestinal infections of Aeromonas are generally considered waterborne;
for this reason, Aeromonas has been placed on the United States Environmental Protection
Agency candidate list of emerging pathogens in drinking water. Furthermore, well
documented field studies have changed the traditional belief that Aeromonas associated
gastroenteritis mainly occurs in African, South American and South East Asian countries (8,
9, 104, 114, 129, 143)
12

1.4.1 Opportunistic pathogen Several degrees of pathogenicity have been observed among the thousands of
bacterial species described so far. Some of the bacteria have a very high probability of
developing diseases in any susceptible host, for example, Bordetella pertussis that causes
whooping cough, is regarded as a primary pathogen. In contrast, other bacteria, such as
Pseudomonas aeruginosa, enter the human body virtually on a daily basis through
consumption of drinking water and vegetables, yet they only rarely cause diseases in healthy
individuals. However P. aeruginosa strains have been known to produce severe infections
(e.g. meningitis) with high fatality (45, 162) especially in hospitalized patients under
immunosuppressive treatment. Bacteria that show this kind of behaviours are called
opportunistic pathogens. Aeromonas spp. are considered as opportunistic pathogen.
1.4.2 Historical aspects of Aeromonas taxonomy The Aeromonas group of bacteria was first described over 100 years ago by Sanarelli
in 1891(163). But the genus name Aeromonas was first proposed by Kluyver and van Niel in
1936 (109). This group of bacteria was allocated too many different bacterial genera in
different time table such as Bacillus (1891, 1901, 1917), Bacterium (1891), Aerobacter
(1900), Achromobacter (1923), Pseudomonas (1930, 1936, 1954), Proteus (1936) and Vibrio
(1955) (52). However, in the seventh edition of Bergeys Manual Determinative
Bacteriology, Snieszko (1957) citated the genus Aeromonas in the family
Pseudomonadaceae (four species, namely the motile species A. hydrophila, A. punctata, A.
liquefaciens and the nonmotile species A. salmonicida)(176). In the eighth edition of
Bergeys Manual of Determinative Bacteriology (1974), the genus Aeromonas was
considered a member of the Vibrionaceae, with a new taxonomy [Aeromonas hydrophila,
(with the subspecies hydrophila, anaerogenes and proteolitica) and A. punctata and the non-
motile species A. salmonicida (with the subspecies salmonicida, achromogenes and
masoucida], which also included the genera Vibrio, Photobacterium, Plesiomonas (192).
Subsequently, phylogenetic investigations indicate that the genus Aeromonas is not closely
related to Vibrio but rather forms a monophyletic unit in the γ-3 subgroup of the class
proteobacteria (128, 161). These conclusions necessitated the removal of Aeromonas from
the family Vibrionaceae and transfer to a new family, the Aeromonadaceae (38).
1.4.3 New Species of Aeromonas The genus Aeromonas has undergone a number of taxonomic and nomenclature
revisions over the past 15 years. Since 1987, a number of new Aeromonas species have been
identified that include A. veronii, A. schubertii, A janda, A torta, A. allosaccharophila, A.
encheleia, A. bestiarum, and recently A. popoffii (94). Most recently, Aeromonas hydrophila
subsp. Dhakensis, one clonal group of Aeromonas, suspected to cause diarrhoea in patients
13

in Bangladesh (85). But the major problem associated with several of the more recently
described Aeromonas species is the extremely limited number of strains. For example, for A.
allosaccharophila, only three strains are known to exist; A. encheleia is based on 4 strains
isolated from European eels; A. popoffii is based on 8 strains isolated from drinking water
(85). In such cases, it is unclear whether the isolates analyzed were in fact separate strains or
whether they simply represented multiple isolates of the same strains.
1.4.4 Genospecies Aeromonas bacteria could be classified into phenotypically defined phenospecies
and into genospecies delineated on the basis of DNA-DNA hybridisation studies (107). Since
the advent of polyphasic molecular approaches in the study of bacterial systematics, the
genus Aeromonas has undergone a number of significant nomenclature changes. Several
approaches have been proposed in order to differentiate between Aeromonas species,
including ribotyping and multilocus enzyme electrophoresis, restriction fragment length
polymorphism (RFLP) (144), amplified fragment length polymorphism (AFLP) (84), the
complete 16S rRNA gene sequences (24), Pulse field gel electrophoresis (PFGE) (23, 181).
Nevertheless, the use of genomic studies based on classical DNA-DNA hybridisation
method is necessary to determine the taxonomic position of Aeromonas species. Aeromonas
genospecies can be defined on the basis of the re-associational kinetics between individual
strains and isotopically labelled DNA extracted from reference strains (93).
1.4.5 Identification to the genus level Identification of Aeromonas to the genus level is relatively simple and
straightforward. Aeromonas species can grow on any standard media such as nutrient agar,
brain heart infusion agar and tryptic soy agar within overnight incubation at 25-30oC for
mesophilic Aeromonas and 22-25o C for psychrophilic Aeromonas (153). Aeromonas genus
can also be easily distinguished from other gram negative, facultative anaerobic genera (i.e.
Vibrio, Plesiomonas, and Enterobacteriaceae) through oxidation and vibriostatic reagent test.
1.4.6 Identification to the species level One of the major difficulties in the identification of Aeromonas strains to species
level concerns the current number of recognized taxa (n=15) and the lack of clear-cut
phenotypic characters useful in distinguishing each species from the others (1). Genospecies
identification of Aeromonas to the species level is unpractical in most clinical laboratories
because of the time required, reagents involved, high instrument cost (2). To facilitate,
routine identification in hospital and clinical laboratories, at least 14 different commercial
systems are available in the USA but the resolving power of most of those system are often
too low to deal with high phenotypic relatedness among Aeromonas taxa (135). For example,
API 20E system can correctly identify only 52% of the Aeromonas strains (98). However,
14

Janda et al proposed that Aeromonas should simple be reported as “Aeromonas species” or
“Aeromonas hydrophila group” or “Aeromonas hydrophila complex”(94). Another
alternative is to continue to biochemically identify strains of the three major species namely
“A. hydrophila, A. caviae, A. sobria”. However, a feasible alternative is to identify
Aeromonas to the species level by a set of reliable biochemical tests. In the early 1990s,
Carnahan et al (31) and Abott et al (1) recommended the use of 20 to 25 selected phenotypic
features for correct allocation of mesophilic Aeromonas strains. However, subsequent
systematic investigations have resulted in nine biochemical key tests as a primary battery to
identify Aeromonas to HG levels (1, 95). The use of these nine phenotypic characters,
however still do not allow differentiation at HG level within A. hydrophila complex (HGs 1
to 3) and A. caviae complex (HGs 4 to 6).
Table 1. Species and DNA hybridization groups currently situated in the genus Aeromonas
HG a, b
(Geno-species)
Species name a, b
(Phenospecies)
Pathogenic status C Main source of isolation b
References b
1 A. hydrophila Major pathogen Humans, animals, water
(152)
2 A. bestiarum Environmental Animal, water, humans
(10)
3 A. hydrophila Environmental Water (52) A. salmonicida Major pathogen Fish (152)
4 A. caviae Major pathogen Humans, animals, water
(152)
5A A. caviae Environment Water (75) 5B A. media Environment Water (75) 6 A. eucreniophila Environment Water (166) 7 A. sobria Environment Fish (152) 8 A. veronii biogroup
sobria Major pathogen Humans, animals,
water (77)
9 A. jandai Minor pathogen Humans (29) 10 A. vronii biogroup
veronii Minor pathogen Humans (77)
11 A. veronii like Environment Human, water (77) 12 A. schubertii Environment Humans, water (76) 13 A .trota Environment Human, water (32) 14 A. allosaccharophila Environment Human, fish (127) 15 A. encheleia Environment Fish (51)
a Genospecies and Phenospecies represent taxa that are delineated on the basis of DNA hybridization data and phenotypic characterization (30) HG, DNA hybridization groups.
b Data adapted from the thesis by Geert Huys (83). C associated with human diseases based upon clinical frequency rather than disease
presentation; data presented from Janda and Abbott (94).
15

1.4.7 Ecology of Aeromonas Aeromonas spp. are ubiquitous organisms which can be readily isolated from a
variety of sources such as water, soil, food, animal and human. Recent progress in the
development of selective media for Aeromonas has increased our knowledge of quantitative
Aeromonas from different sources (148). Despite the new Aeromonas species, most of the
Aeromonas bacteria appear to be environmental species. Only six of the Aeromonas species
have been unquestionably established as human and animal pathogens. Within this five
species, three species accounts for more than 85% of all clinical isolates and they include
HG groups 1, 4 and 8 (94).
Environmental Water: Aeromonas spp are widely distributed in the aquatic
environment. Numerous ecological investigations have showed that Aeromonas occur in
virtually all kinds of aquatic environments throughout the world but in varying
concentrations (13, 98). They can be found in both polluted and unpolluted water, sewage
water, drinking water, and chlorinated and non chlorinated water distribution systems and
even in bottled mineral water. These species were first considered to be absent from the
marine water, but a number of recent reports demonstrated that they are widespread in
marine environments too (136).
Plant and phytoplankton: Aeromonas spp not only have been found in aquatic
water but also in aquatic plants, soil sediments and phytoplankton. Islam et al reported that
Aeromonas sobria could be isolated in plant and phytoplankton’s during hot weather with
the highest isolation rates (47.5%) (90).
Mineral water: Aeromonas was isolated from mineral waters in Spain, which was
published in two papers (134). Gonzales detected this organism in one of three investigated
mineral water sources (60), while Quevedo et al (154) referred that approximately 10% of all
bacteria recovered from mineral water are belonged to Aeromonas spp. In another report,
these organisms were detected in 4.9% mineral water samples and three of these (1.8 %)
with A. hydrophila counts >103 CFU/ml (41). In Taiwan, Aeromonas were isolated from 31
of 88 domestic and 11 of 48 imported mineral water samples (188).
Foods: Aeromonas have been isolated from a wide range of foods including raw
and processed meat, poultry, seafood, raw milk, vegetables and pre-prepared salads (113,
132). The isolation of Aeromonas from vegetables have been shown to represent a potential
risk to consumers health, due to their possible pathogenicity (production of cytotoxin at
refrigerator temperature) and ability to grow at refrigerator’s temperature (132). Organic
vegetables have been considered to represent an increased risk to public health, where
natural fertilizers such as animal manure and sewage water are used and where no chemical
treatment is employed to reduce the microbiological load of the raw products (132).
16

Seasonal abundance: The abundance of Aeromonas in the aquatic ecosystem has
been shown to be seasonally distributed. Islam et al (90) showed a high incident of
Aeromonas sobria during the warmer months (May to July) and the beginning of the winter
in November in a Bangladeshi pond. Different studies also show that the number of
Aeromonas increased with the temperature, with a maximum occurring during the summer
and the lowest incidence during the winter. Monteil reported that Aeromonas were probably
one of the leading causes of bacterial enteritis during summer months, affecting mainly
young children and travellers (138). The correlation between summer seasons, increased
water contamination levels and increased number of gastroenteritis cases may be associated
(54, 138).
1.4.7 Pathogenicity of Aeromonas Cold blooded animals: The pathogenicity of Aeromonas was first established
with cold-blooded animals, A. hydrophila can cause red leg disease in frogs. But recent
studies have shown that turtles, snakes, crocodiles and lizards can also be infected with
Aeromonas with a high rate of mortality (189). Recently, Turutoglu et al described a case
report of skin lesions and septicaemia associated with A. hydrophila in a Nile crocodile
(Crocodylus niloticus) (189). Psychrophilic species Aeromonas salmonicida is known as the
etiological agent of furunculosis in salmon worldwide. Furunculosis has caused enormous
economical losses in commercial salmon and trout fish farms during the last decades. Recent
studies indicate that motile mesophilic Aeromonas play a major role as opportunistic
pathogen in freshwater fishes worldwide. McGarey reported that 50-80% of daily catches of
fish from the St. Jons River system exhibited UDS (Ulcerative Diseases Syndrome) lesions
(131). This disease is frequently occurring not only in different intensive culture fish farms
but also in open water bodies (i.e lakes, rivers) in Indo-Pacific regions (155). Different
studies have shown that Aeromonas is frequently associated with diseases in carp, eels,
milkfish, channel catfish and tilapia (156) .
Warm blooded animals: Aeromonas species have been isolated from a wide
range of mammalians including cats, dogs, horses, rabbits, monkeys, guinea pigs, chicken (4,
57, 146). Recently Andre reported the pure culture of Aeromonas hydrophila from liver and
kidney of a dog suffering from leptospirosis with septicaemia due to this bacteria (11).
Aeromonas infections in human: Aeromonas associated major human infection
could be divided into two major categories. One of them is gastroenteritis that occurs mainly
in young children or aging people but also in travellers (72). Besides of gastroenteritis extra-
intestinal may occur infection in adult individuals (94).
17

Gastroenteritis: The first Aeromonas associated Gastroenteritis was reported by
Ronser et al 1964 (159). From 1964 until today, numerous studies have demonstrated a
strong correlation between high number of Aeromonas in the gastrointestinal tract and
human diarrhoeal diseases. Several case control studies have shown the epidemiological
links between Aeromonas and acute diarrhoea (9, 27, 149). However, none of these
investigations have been able to identify Aeromonas as the true causative agent of
gastroenteritis (94). Thus the single greatest topic “the association of Aeromonas with
gastrointestinal disease” is still unresolved. The fact is that no well documented
epidemiologically linked outbreaks of diarrhoeal disease attributed to Aeromonas have been
reported as yet (94). Even volunteer studies that involve the ingestion of Aeromonas of
various strains showed that only certain strains were associated with colonization in most
individuals when the doses of challenge exceeded 109 CFU (139) .
Extra intestinal infections
Wound infection: Wounds are the second most common infections by Aeromonas
after the gastrointestinal infections (94). Wound infections caused by Aeromonas often
associated with aquatic accidents or occupational accident where the source of Aeromonas
was shown to be water and soil (94).
Septicaemia: Septicaemia is generally considered to be the most life threatening
infection caused by Aeromonas. Aeromonas sepsis usually occurs in immunocompromised
patients but also in patients suffering from malignancy, hepatobiliary diseases and diabetes
(96). Mortality rates for Aeromonas associated septicaemia are usually ranged between 25-
50% (46, 94).
Other infections: Aeromonas species have been shown to be associated with
several other types of infection including meningitis, peritonitis, catheter related infection,
myonecrosis (94, 105, 140, 170).
1.4.8 Putative virulence factors of Aeromonas
Virulence often refers to a measure of the capacity of a microorganism to infect or
damage a host and is often quantified by the size of the dose required to infect (ID50) or to
kill (LD50) 50% of hosts (121). Molecular studies have shown that virulence is a composite
trait and virulence will vary often be selected independently, and selective pressures may
vary between different macro- and microenvironments within a single host or even within a
single tissue (121). Due to the lack of adequate animal models, the significance of the precise
putative virulence in Aeromonas remains to be established. It is well known that some of the
motile Aeromonas possess a number of putative virulence factors. The Table 2 shows the
putative virulence factors described in the literature. Some of these virulence factors were
18

19
found to be distributed in strains isolated from the environment and from the cases of
infections. Thus an association of prevalence of putative virulence factors and infection
could not be established. However, certain species are more frequently isolated from
infections than from environments.

20
Table 2 Putative virulence factors of Aeromonas Virulence factor Gene Prot. Size Effect Ref.
Toxins
Aerolysin aerA 48 kD channel form. (5) HlyA hlyA hemolysis (74) AexT aexT ADP ribosyltransferase (25)
Cytotonic enterotoxin alt cAMP and PGE2 levels* (36) ast 32 kD
(35) Cytolytic enterotoxin act 52 kD haemo- /cytolytic Shiga-like toxin 1 (plasmid encoded) (73)
Enzymes
Acetylcholinesterase 45 kD (151)
Lipase Phospholipase C
Serine protease aspA 70 kD (193) Metalloproteinase asaP1 20 kD caseinolys (190) GCAT (glycerophospholip:cholesterol acetyltransferase) lipase Metallo-gelatinase P2 Aminopeptidases Amonabactin T and P fstA and fsp (150)
Surface proteins/outer membrane proteins/fimbriae S-layer
Mucin binding component 22-95 kD Type IV pili
*: elevated cAMP and prostaglandin (a receptor-mediated activator of adenylate cyclase) levels in CHO cells.
Involved in cell differentiation and apoptosis.

1.4.9. Biofilm formation of Aeromonas In general, biofilms are defined as assemblages of microorganisms associated by
their extracellular products. A typical biofilms are communities of microorganisms attached
to an abiotic or biotic surface (43). Bacteria found in environmental, clinical and industrial
settings often exist as surface associated biofilms (40) and can be beneficial or deleterious.
Biofilms give the bacteria a significant survival advantage, since bacteria in biofilms can be
resistant to antibiotics, chlorination, disinfections (39). Bacteria residing within a biofilms
receive certain degree of shelter since cells in biofilm are known to enmeshed within a
matrix of extracellular polymeric substance (EPS) by the organisms themselves and, EPS
vary not only upon the organism, but also within the same organisms when other
environmental conditions are applied (43, 147).
Biofilms are extremely important in the waste water treatment process since they
remove many kind of key waste water contaminants such as nitrogen and phosphorous
compounds (142). On the other hand biofilms control is now recognised as an important part
of the operation of drinking water plants and distribution systems in developed and
developing countries Aeromonas was identified in 7.7% of biofilm samples in a water
distribution system (34), a finding that may be related to future outbreak of gastroenteritis.
In natural settings biofilms form in aqueous environments, and aquatic environments are also
known as reservoirs of different pathogenic and non-pathogenic bacteria such as V. cholera,
V. parahaemolyticus, Aeromonas spp. In the aquatic environment, these bacterial may form
biofilms on the aquatic flora and fauna, persist for a long time and be are transmitted within
the environment by using the aquatic flora and fauna as a vehicle. Genetic exchange between
community members in environmental biofilms is another key aspects environmental
biofilms harbouring pathogens
Bacteria sense the environmental signal and settle on the surface. Sensing of cell
density (quorum sensing) plays a major role in bacterial biofilm formation (28). Quorum
sensing could be define as accumulation of a low molecular mass signal enables individual
cells to sense when the minimal population unit (’quorum’) has been achieved (for a
concerted population response to be initiated) (180). Quorum sensing in gram-negative
bacteria is mainly based on N-acyl-homoserine lactones (AHLs) and, its co-regulatory
receptors which control the expression of genes (97). AHLs are the mostly studied quorum
sensing signalling molecules in Gram negative bacteria (178). Chemically, AHLs consist of
a homoserine lactone ring, an amide bond and acyl side chain of variable length (to date 4 to
18 carbons in length have been identified). Aeromonas spp are known to produce C4-HSL
(N-butanoylhomoserine lactone) (108, 123, 179) .
21

1.5. Enterococci 1.5.1 Genus description of Enterococcus
Enterococcus are gram positive cocci, arranged single, in pairs or as short chains.
They are facultative anaerobes with a growth range from 10o to 45oC. All Enterococci
hydrolyze esculin in the presence of bile salts, grow in broth containing 6.5% NaCl, produce
leucine aminopeptidase, and most enterococci but not all, and hydrolyze pyrrolidonyl-b-
naphtylamide (PYR). At present there are 23 distinct Enterococcus species based on
molecular characterization techniques such as DNA-DNA reassociation, 16s RNA gene
sequences, and whole cell protein analysis.
1.5.2 Habitat of Enterococcus Enterococci are natural habitants of the human intestinal microflora, and can be
found in faeces of most adults. However, they represent only a minority, i.e. less than 1% of
the microorganisms found in faeces of healthy individuals (167). E. faecalies and E. faecium
are the most commonly detected with E. faecalies as the dominating species but E.
casseliflavus, E. durans and E. avium have also been isolated from small proportions of
human individuals (12, 48).
Enterococci are less frequently isolated from other human sites, but have been found
in vagina, oral and skin specimens. In addition they can be found in soil, on plants, in waters
and in various food products (47). Enterococci have a hardy nature and can endure harsh
environmental conditions, such as 65oC for 10 minutes and a pH from 4.8 to 9.6 (103). They
can survive in nutrient deficient environments and have been recovered from dry surfaces
after weeks and from water after several months (103, 191).
1.5.3 Clinical diseases and nosocomial infections by Enterococcus The most commonly isolated enterococci of clinical significance are E. faecalis and
E. faecium, both of which may be the causative agent of different clinical diseases such as
urinary tract infections, wound infections and serious blood stream infections. E. faecalis is
most frequently encountered in infections acquired in the community, in long term care
facilities and in hospitals (20, 58, 110).
During the last few decades, enterococci have emerged as important nosocomial
pathogens with great ability to acquire antibiotic resistance. Enterococci are in the USA the
third and in Europe the fifth most common clinical isolate causing serious nosocomial
infections such as bacteraemia. They also spread their resistance genes to other bacteria
outside the genus. Transmission of resistant enterococci strains between patients treated in
different settings has also been noted. The hardy nature of Enterococcus makes them able to
survive in the environment for a long time. Increased movements of colonized patients
between different units and hospitals is partly responsible for their spread.
22

1.6. Antibiotic resistance
Antibiotic resistance can be divided into intrinsic resistance and acquired resistance.
Intrinsic or inherent resistance can be regarded as a specie’s characteristics and occurs in
most stains of a particular species. The genes of this type of resistance typically reside on the
chromosome. Acquired resistance is the result of mutations in the existing DNA or
acquisition of new DNA from other organisms. Basically antibiotic resistance in bacteria
disseminate in two ways: clonal spread of resistant clonal lineage and horizontal transfer of
resistance genes. The most important vehicles for resistance gene transfer are mobilisable or
self transmissible plasmids in conjugation with transposons, IS-elements and integrons.
Previous studies have shown that class 1 integrons associated resistance gene cassettes are
highly prevalent on conjugative R-plasmids within Aeromonas population in different fish
culture environment from different countries. The occurrence of integrons associated
resistance gene cassettes located on antibiotic resistance plasmids was also isolated from
waste water treatment plant. It has been shown that plasmid mediated horizontal gene
transfer play an important role in waste water purification facilities.
1.6.1 Antibiotic susceptibility testing and MIC breakpoint
In order to find out if an antibiotic exhibits activity against a particular isolate of an
organism, antibiotics susceptibility testing is performed. The susceptibility testing is done by
determining the minimal inhibitory concentration (MIC) of the drug for the isolates of
interest. The MIC results are then interpreted in a breakpoint system and isolates are divided
into susceptible (S), intermediate (I) and resistance (R). Ideally, a breakpoint system should
satisfy both the need to guide therapy and the need to detect biological resistance. From the
clinical point of view this means that if an organism is classified as susceptible to a particular
antibiotic, it can be assumed that the drug will inhibit the organism and therapy will be
successful. However, clinical breakpoints do not take into account whether or not the
organisms harbour resistance mechanisms. To meet the needs of epidemiological monitoring
of emerging resistance, the MIC breakpoint should be able to divide the susceptible wild
type strains from those one that have acquired resistance mechanisms.
Many different methods are available for bacterial susceptibility testing, including
disk diffusion, agar dilution, broth microdilution and antibiotic gradient disks. Broth
microdilution is a convenient and widely used for susceptibility testing of several antibiotics
on a large number of bacterial isolates in a short time.
23

24

AIMS OF THE STUDY
1 To study the prevalence of FMSE1 like PhP clonal group of Enterococci in hospital
sewage water.
2 To characterise the Aeromonas strains isolated from Epizootic Ulcerative Syndrome
in fish in Bangladesh.
3 To study the persistence and transmission of Aeromonas in a duckweed aquaculture
based hospital sewage water recycling plant.
4 To develop a user-friendly semiautomatic susceptibility testing method to screen
large numbers of isolates in a short time.
5 To study the influence of a duckweed aquaculture based hospital sewage water
recycling plant on the antibiotic resistance of Aeromonas.
6 To study the cyclic- di - GMP signalling in Aeromonas.
25

26

MATERIALS AND METHODS
Sample and Isolates
Study I. PhP fingerprinting of 9676 Enterococci from different kinds of sources
(such as untreated urban sewage water, treated urban sewage water, hospital sewage water,
surface water, pig, pig faeces, pig manual, broiler chicken, cattle, hospitalised patients,
outpatients, healthy children) from our databases were used in this study (117). The number
of isolates of Enterococci from the different studies and sample types are described in Table
1 in Paper I.
Study II. Fourteen Aeromonas spp isolated from different diseased fish from several
fish farms, with deep hemorrhagic ulcers in the mid-body and tail regions, were obtained
from a collection of Bangladesh Agricultural University fish diseases laboratory. For
comparison, 26 environmental Aeromonas isolates were randomly chosen from the same
disease ponds, and twelve randomly selected human diarrheal isolates of Aeromonas spp
from an earlier investigation (114) on Bangladeshi children suffering from diarrhoea were
included in the study. The bacteriological methods used in this study are described in Paper
II.
Study III, IV, V. The sampling sites were a model system of duckweed aquaculture-
based hospital sewage (Kumudini hospital and its residential area complex) water treatment
plant located in Tangail, Bangladesh. A total of 1,318 presumptive Aeromonas from a total
of 691 samples from sewage water, duckweed, fish, patients suffering from diarrhea, control
ponds and healthy humans were isolated over a period of three years (Papers III, V).
Furthermore, a total of 1,005 coliform bacterial were also isolated from the same samples
(paper IV). The procedures for samples collection, samples processing and bacteriological
analysis are described in the corresponding papers.
Biochemical Phenotyping with the PhenePlate™ system (study I, II, III, IV, V)
Bacterial isolates were typed through a biochemical phenotyping method,
PhenePlate™ system (PhP; PhPlate Microplate Techniques AB, Stockholm, Sweden)
according to the manufacturer’s instructions. The biochemical reactions of the isolates were
compared pair wise and a similarity matrix consisting of the correlation coefficient between
all possible pairs was constructed. The similarity matrix was clustered according to the
unweight pair group method using average linkages (UPGMA)(173). Isolates with a level of
similarity greater than 0.97 were assigned to the same PhP phenotypes. All data processing,
including optical reading and calculation of correlation coefficient, as well as clustering and
27

printing of dendrograms, was performed with PhP software (PhPlate Microplate Techniques
AB, Stockholm, Sweden).
FAME analysis (Study II, III, IV)
Gas-liquid chromatographic analysis of cellular fatty acid methyl esters (FAMEs)
were prepared and extracted by using the standardized protocol of the Microbial
Identification Systems (MIDI; Microbial ID Inc., Newark, Del.). The extracts were analysed
with a Hewlett-Packard model HP5890 gas chromatograph as described previously (145).
Identification of the FAME fingerprinting, generation of database and unweighted arithmetic
average cluster analysis were performed using the MIDI software package. Aeromonas
isolates were identified by comparing their FAME profiles with the laboratory-based
identification library AER48C (86).
AFLP analysis (Study II, III)
Whole-genome fingerprinting were carried out using amplified fragment length
polymorphism (AFLP) analysis according to the method of Huys and Swings (84). The
AFLP profiles of unknown isolates were compared with the laboratory-based identification
library AEROLIB comprising AFLP profiles generated from a collection of well-
characterized type and reference strains encompassing all currently recognized Aeromonas
taxa (87).
PFGE analysis (Study I)
DNA fingerprinting by pulsed field gel electrophoresis (PFGE) was performed
according to the method described by Torell et al (186) and the resulting DNA band pattern
was interpreted according to criteria proposed by Tenover et al (182).
Cytotoxic and hemolysin activity (study II, III)
Cytotoxic activity of the isolates was tested on the fish cell line EPC (epithelioma
papulosum of carp, Cyprinus carpio ) or Hep- 2 cell lines as described previously (56).
Briefly, confluent monolayers of the cell were grown in 24 well tissue culture plates (Coster,
Corning, NY, USA) in minimal essential medium supplemented with 10 % fetal bovine
serum, 1% (w/v) glutamine and 1% (w/v) penicillin-streptomycin. The cells were incubated
for 6 h at 18°C with 100 µL of sterile culture supernatant (containing 150 mgl-1 protein as
determined by Bio-Rad protein assay) serially two-fold diluted in supplemented MEM. The
cytotoxic activity was measured as rounding up, detachment and loss of viability of the cells
as seen in the light microscope within 6 h. The titer was determined as the highest dilution of
28

supernatant affecting at least 50 % of the cells. Isolates showing a cytotoxic effect at a
dilution of 1/8 (final concentration) or more were regarded as positive.
Hemolysin activity (study II, III)
Hemolytic activity of the isolates was measured on 1% (v/v) human and rabbit
erythrocytes as described earlier (101). Isolates were considered positive for haemolysin
production when the culture supernatant at a final concentration of 1/8 lysed at least 50 % of
the erythrocytes as determined by eye. Known positive (BD2-9) and negative (BD12)
isolates from an earlier investigation were included as controls into both cytotoxin and
haemolysin assay (115).
Hemagglutination (Study II)
Hemagglutination tests were performed on glass slides by mixing a loopful bacteria
with a 3 % (v/v) suspension of erythrocytes from rabbit, human or fish (Labeo rohita) in
PBS. Visible agglutination within five min was considered as positive. Agglutination
inhibition test was performed using a dilution of 1 % (w/v) D-mannose, D-galactose and L-
fucose sugar in PBS (66).
AHCYTOEN genes detection (study II, III)
The AHCYTONE gene, a cytolytic enterotoxin gene (AHCYTOEN) has been
reported as a multivirulence gene involved in lethality in mice, hemolysins, cytotoxicity and
enterotoxicity, were detected by PCR method using the primer AHCF1 ( 5 / – GAG AAG
GTG ACC ACC AAG AAC A–3 / ) and AHCR1 ( 5 / –AAC TGA CAT CGG CCT TGA
ACT C–3 / ) (107). The specificity of the primer combination was corroborated with positive
and negative controls during PCR experiments.
Antimicrobial susceptibility testing
The tested agents and range of concentrations for tested antimicrobial agents were:
ampicillin (2-128 mg /L), tetracycline (1-64 mg/L), chloramphenicol (2- 128 mg/L),
nalidixic acid (2-128 mg/L), cephalothin (1-64 mg/L), erythromycin (2-128 mg/L),
streptomycin (2-128 mg/L), and gentamicin (1-64 mg/L). Preparation of microdilution plates
were performed according to the NCCLS recommendations (141). Furthermore, in the
“ScanMIC” method, a growth indicator (TTC) was also added to the microplates and the
microplates were dried overnight by dry air at 25o C.
Bacteria were cultured over night on blood agar plates. Three colonies were
transferred to 5 ml Mueller Hilton broth (Becton Dickinson, USA) without blood and
29

incubated at 37o C for four hours to reach the exponential phase of growth. From these
cultures, bacterial turbidity was adjusted to 0.5 on the McFarland turbidity standard as
measured by absorbance (0.08-0.1 at 625 nm) in spectrophotometer (Hitachi U-1100),
corresponding to approximately 10 8 CFU/ml. The above adjusted bacterial suspensions were
first diluted 1: 200 in Mueller-Hilton broth, and 100 µl were added to each well (5 X 104
CFU/ well) in the pre prepared microplate (broth microdilution or “ScanMIC” microplate)
followed by incubation for 16 h at 37 oC. For “ScanMIC”, the microplates were scanned
with a reflective flat bed scanner (UMAX-Astra 6450, USA) connected to a Windows based
PC. The images were analysed by an in-house developed software and MICs were
determined by software. For broth microdilution, MIC was defined as the lowest
concentration of an antimicrobial agent at which no growth was detected (100% inhibition)
as turbidity seen visually. The detailed methods are described in paper IV. The “ScanMIC”
method was compared to the reference broth microdilution method, in two different ways. i)
The distribution of differences in MICs results ii) Interpretative categorical results, obtained
according to the NCCLS recommended interpretive standards (SIR=Susceptible,
Intermediate and Resistant) for Enterobacteriaceae (141), were compared for the “ScanMIC”
method versus the reference method. The details of these calculations were described in
paper IV.
Interaction of Acanthamoeba castellanii and Aeromonas
Acanthamoeba castellanii (ATCC 30234) and Aeromonas hydrophila (HG1) strain
(AEW 44) isolated from child suffering with diarrhea and AEW 20 isolated from the
environment were used in this study. A. castellanii was grown without shaking at 30°C in
ATCC medium no. 712 (ATCC). For invasion experiment, invasion positive strains Dublin
strain SH9325 and invasion negative strain TG1 were used as reference strains (183). For
visualization of the bacteria by fluorescence microscopy we transformed the Green
fluorescent protein (GFP), vector pGFPuv (Clonetech, BD Bioscience) to Aeromonas
hydrophila AEW 20 by electropoeration. The Co culture experiments and invasion
experiment were carried out as described previously (3, 183).
Biofilm and c-di-GMP related assays.
One Aeromonas veronii strain (AEW43), isolated from a child suffering from
diarrhoea, was chosen to study the c-di-GMP signalling pathway in this study. The strain
representative a clonal group of Aeromonas (MT7), which showed persistence ability in
different niches, such as environment, human, fish, and was positive for all the tested
virulence factors.
30

The detailed of the construction of the plasmids (GGDEF domain containing protein
AdrA cloned into p LAFR3 and named pWJB9; EAL domain containing protein YhJH
cloned into p LAFR3 and named pRGS3 and pLAFR3 itself as vector control) has been
described by Simm et al (171). The E. coli DH5 α strains carried those plasmids were
transcongugated to the selected Aeromonas strains (AEW43) through the plate mating
methods resulting the AdrA, YhJH and vector expressing Aeromonas.
Isolation and detection of c-di-GMP from Aeromonas were performed as described
by Simm et al (171). For phenotypic assessment, the AdrA, YhJH and vector expressing
Aeromonas was grown on Congo Red (CR) agar plates for 20 h at 37°C as described by
Römling et al (157). Swimming motility was observed using 0.3% LB agar plates inoculated
with a constant cell number from overnight cultures. Biofilm formation was observed in
glass tubes. Overnight cultures of Aeromonas were inoculated in LB medium for 16 h with
shaking 150 r.p.m. After staining the glass tubes with crystal violet, the biofilms were
dissolved in dimethylsulfoxide and the absorbance at 540 nm was measured. In some cases,
a score was assigned on the absorbance values.
Furthermore, to observe the adhesion of constructed strains to duckweed surface,
duckweeds were washed several times with gentamicin. The duckweeds were then incubated
with a bacterial suspension of 107 CFU/ml for 1 hour at room temperature and washed five
times with PBS in order to eliminate non-attached bacteria. For the quantification, the
duckweed and attached bacteria were then lysed in 0.1% Triton in sterile water for 10 min.
The ability of the bacteria to adhere to the duckweed was determined by spreading the Triton
induced cell lysine on nutrient agar plate and counting the number of colonies after
incubation overnight at 37oC.
The quorum sensing molecules, HSL were extracted from AdrA, YhjH and vector
expressing strain as method described by Lynch el al (123). To estimate the bioluminescence
of extracted HSL, overnight culture of Chromobacterium violaceum CV026 or E. coli
JM109 (pSB536) were diluted in LB containing antibiotics to a final optical density at
600nm within the range of 0.005 - 0.006. A 100 µl of the above bacterial culture were added
to a microtiter plate which containing HSL extraction, and incubated for overnight at 30oC
with shaking at 150 r.p.m. Bioluminescence was measured in the luminometer (VICTOR
1420 Multilab counter, Wallac) and relative light units as a function of C4-HSL
concentration was expressed.
31

RESULTS AND DISCUSSION
4.1 Transmission of Enterococcus faecium from hospital patients to environment (Paper I)
During the 1990s a constant increase in the incidence of clinical Ampicillin
Resistant Enterococci (ARE) would be observed in many Swedish hospitals (69-71). Torell
et al showed that 22% of the hospitalized patients and 6% of the outpatients were faecal
carries of ARE (185). Biochemical fingerprinting of these isolates revealed that, 73% of 180
ARE isolates from hospitalised patients and 54% of 39 ARE isolates from outpatients
belonged to the same PhP types (arbitrary named as FMSE1) and almost all isolates of this
type showed high level resistance to ampicillin and ciprofloxacin (186). Thus there was
evidence for a faecal carriage of clonally related ARE isolates among Swedish hospitalised
patients and outpatients nationwide.
The prevalence of Enterococcus faecium FMSE1 (an ampicillin and ciprofloxacin
resistant) strain of was searched for in the PhP typing databases in materials collected
(databases comprising 9,676 isolates of Enterococci) from different sewage waters,
slaughtered animals, pig farms, humans, etc. The FMSE1 type was identified in 1.4% of all
isolates. The FEMS1 type was most commonly found in samples collected from hospital
sewage (50%), followed by surface water (35%), treated urban sewage water (28%),
untreated urban sewage water (17%) and rarely in healthy children (0.8%) and animals (2%)
(Table 1, Paper I). The high prevalence of FMSE1 in different sewage and surface water
samples might be related to a spread of faecal bacteria from hospital patients to sewage and
then to surface water.
The available FMSE1 PhP type isolates were further typed through the PFGE typing
and it was verified that the isolates from hospital sewage and the FMSE1 reference isolates
were genetically closely related (Figure 1, Paper I), whereas the FMSE1 isolate from surface
water that was similar according to PhP typing, showed a different banding pattern, though it
could still be regarded as related. The antibiotic resistance pattern of this isolate also showed
a slightly different profile, while all hospital sewage isolates showed identical profiles
(Figure 1, Paper I). Thus, there is a possible transmission of ampicillin and ciprofloxacin
resistant strains from hospital patients to hospital sewage water, and also possibly to the
surface water. The hospital sewage seems likely the source of resistant bacteria. Thus it
could be possible that ampicillin and ciprofloxacin resistant Enterococcus faecium might be
transmitted to the environment from hospital patients to hospital sewage water and further
through urban sewage waters to treated sewage water and surface water.
32

Drug resistant Enterococcus strains may be carried for months or even years in the
faecal flora of humans (22, 122) and have the potential to spread through colonization of
patients (106, 187). Drug resistant isolates such as FMSE1 like isolates (in Sweden), might
have acquired resistance to ciprofloxacin in hospitals, giving an increased incidence in
hospitalised patients through clonal spread (184, 186). Further on, this same clonal group
transmitted to the environment through hospital sewage water. Current infectious disease
control measures and antibiotic usage routine controls might not prevent this silent spread of
FMSE1 like isolates in the community. But in near future, early detection of new FMSE
isolates with acquired antibiotic resistance might contribute to the control of the clonal
spread in the society.
Table 3 Prevalence of FMSE1-like isolates from PhP Enterococcus database.
% (Positive/number)
FMSE l like
isolates in*
Hospital
sewage**
Surface water Untreated urban
sewage
Treated urban
sewage
Samples 50 (7/14) 35 (13/35) 17 (6/35) 28 (9/32)
Isolates 6.4 (24/374) 3.8 (24 /625) 1.5 (10/677) 1.2 (10/869)
* Samples and isolates analysed for FMSE l like type;
** Samples or isolates were positive within the investigated samples or isolates
In paper I, the samples have been taken from a broad spectrum of sources, and
unfortunately a limited number of isolates were available for further typing. Therefore, we
could not precisely confirm the transmission route(s) of the bacteria from the hospitalised
patient to the environment. Further sampling within a narrow spectrum may permit the
detection of the precise transmission route of the bacteria from hospital to the human
environment. Thus a multidisciplinary project has been chosen, where hospital sewage water
was recycled for production of food, to be consumed by human (Paper III).
33

4.2 A clonal lineage of Aeromonas is associated with fish disease (Epizootic Ulcerative Syndrome) (Paper II)
Epizootic Ulcerative Syndrome, a fish disease, frequently occurs in many fish farms
and open water bodies (lakes, rivers) in Bangladesh as well as in many other countries of the
Indo-Pacific region. The disease generally develops with ulcers on the fish bodies, especially
on the head, middle of the body and dorsal regions (Figure 2). The fish usually die within
one week of infection. The diseases have caused substantial economical losses to fish
farmers and open water fisheries in Bangladesh.
In this study (Paper II), a collection of Aeromonas strains isolated from fish ulcer
scraps were characterized. Biochemical phenotyping with the PhP system revealed that the
isolates belonged to one PhP type. It is shown in the dendrogram that EUS associated
isolates belonged to a PhP clonal group of Aeromonas when 52 Aeromonas isolates from
fish, from environment and from human diarrhoea patients were clustered (figure 1, Paper
II). This EUS associated PhP type was searched for in PhP-AE database, containing a
collection of 1,600 Aeromonas isolates, which comprised Aeromonas isolated from human
diarrhoea and the environment in Bangladesh. But the EUS associated PhP type was not
identical to any type in the database, which indicates that this PhP type might be a unique
clone.
Figure 2 Fish suffering from Epizootic Ulcerative Syndrome (EUS) with dermal ulcer in the middle of the body.
Figure 3 Infected fish are ready to sell in the market from a fish farm; inside infected fish
34

When we compared the virulence properties of the EUS isolates to those of human
diarrhoea and environmental isolates, we found that all the EUS isolates produced all the
virulence factors tested (haemolysin, cytotoxin, the presence of cytolytic enterotoxin genes,
agglutination to different erythrocytes). But some of the human diarrheal and the
environmental isolates also showed the same characteristics. Interestingly, only the EUS
isolates had the ability to agglutinate fish erythrocytes (Table 1, Paper II). In a previous
study by our group we have shown that one phenotypic clonal group of Aeromonas (BD 2,
later assigned a new subspecies Aeromonas hydrophila sub. dhakensis) was associated with
children suffering from diarrhoea and this group was distinguishable from environmental
phenotypes. All of the isolates from that clonal group, except one were positive when tested
for putative virulence factors. Similar to that study, we assumed that the specific EUS
associated clonal group in this study was the causative agent in the fish disease in
Bangladesh.
To test the pathogenicity of the isolates (unpublished data), one representative
isolate of the human diarrhoeal type and one of the EUS associated isolates were subjected to
pathogenicity testing in rainbow trout. The mortality rate was found 100% when challenged
with 104 cells/fish for the diarrhoeal isolate. The infected fishes displayed several ulcerations
with muscle necrosis spreading from the site of injection and the fish kidneys were liquefied.
However, only 67% mortality was found upon challenge with 104 cells/fish for the EUS
associated isolate. The disease signs included haemorrhaging in the muscle around the
injection sites and also the fish kidneys were liquefied. Our results showed that EUS
associated isolates are pathogenic to fish, and human diarrhoeal Aeromonas may also cause
infection in fish. It seems that human diarrhoea associated isolates were more pathogenic to
fish than the fish isolates. Thus if human pathogenic Aeromonas are transmitted to fish
environment, they may increase the risk of infection in fish. But for practical reasons, we
could not detect whether fish associated isolates were pathogenic to human.
The origin of the EUS associated clone in the fish culture environment is not known.
But because of the remarkable survival potential of Aeromonas in the environment, it would
be guess that the EUS associated clone originates from human diarrhoea and was transmitted
to the fish culture pond. In a visit to a fish farm (anonymous) in Bangladesh, we observed
that infected fish was caught for sale in the market indicates that there must be a circulation
of strains between human and fish environment (Figure 3). It is not documented whether
Aeromonas infected fish would be considered as a public health hazard or not. However, fish
culture environment should be aware from contamination of human sewage, since human
pathogenic Aeromonas could increase the rate of fish infection. Our result showed the
evidence for circulation of strains between human and fish in Paper III.
35

36
4.3 Persistence and transmission of Aeromonas in the sewage treatment plant (Papers III, V)
To provide the evidence for possible transmission and/or circulation of bacteria from
hospitalised patients to environment and/or to the human food chain through the sewage
water, we have sampled from a duckweed aquaculture based hospital sewage water treatment
plant and Aeromonas species were chosen as the indicator bacteria (Figure 4). In this studied
treatment plant, the hospital sewage water was first allowed to settle in a settlement pond
and, was then transferred to a lagoon where the duckweed was grown. By growing the
duckweed in the sewage water, organic substances such as, nitrogen and phosphorus loads
were decreased; thus purifying the sewage water. After the duckweed treatment of the
sewage water, treated sewage water effluent was discharged into the nearby river and the
sewage grown duckweed was used as fish food in intensive culture fishponds within the
treatment plant.
4.3.1 Persistence of Aeromonas strains in different samples
For the bacteria, to be able transmit from wastewater to human, the first criteria is survival
and persistence in the treatment system. Among our collection of 1,318 Aeromonas isolates, only 57%
(arbitrarily defined and named to 23 major types) were recovered at several occasions and at several
sites by our testing methods and the remaining isolates occurred sporadically. These findings suggest
that only members of these major types were able to survive and persist during the purification
process (Table 2, Paper III), i.e. were present or possible to detect in different samples. For example,
PhP type MT1 (described as major type in Paper III) was detected in 6 out of 19 Aeromonas positive
hospitalised children suffering from diarrhoea while type MT13 was detected in 4 out of 10
Aeromonas positive human workers. Few major types (MT:s 2, 4 and 11) were found to be distributed
in different sites of the treatment plant, in hospitalised children suffering with diarrhoea and in the
control ponds. Furthermore, some interesting types (MT:s 3, 7 and 8) were detected in the treatment
plants and in the hospitalised children suffering with diarrhoea but never in the environment, i.e. the
control ponds. It is not well understood why certain types where only detected in specific sites but
possibly some strains were adapted to a specific niche e.g. the human gut, the fish pond, while other
strains survive in several niches. For example, some strains were only found in the human samples but
not in the environment, and certain strains survived only in sewage water while others in all niches.
However, our data indicate that phenotypically and genetically stable certain Aeromonas strains
(MT10 type) can survive and persist in the sewage water treatment plant for years (Figure 4, Paper
III). The remarkable survival and persistence of Aeromonas in different environments, such as water
microcosm for 180 days and water distribution systems for years, is documented (116, 134).

37
Hospital
Waste settlement pond
Duckweed lagoon
Aeromonas
Figure 4 Different sampling sites of the duckweed aquaculture based hospital sewage water treatment plant.
Fish pond and fish feed

When the PhP profile of our collected 1318 isolates were compared to a PhP references
database for Aeromonas it was found that A. hydrophila was the dominating species (45 %)
in the human samples while its relative frequency decreased to 11 % in fish (Figure 2, Paper
III). On the other hand, A. veronii was found at the lowest percentage (22 %) in human
samples but this percentage increased throughout the purification process up to 51 % in the
fish. We found both A. hydrophila and A. veronii were common in the control ponds.
4.3.2 Evidence for transmission of Aeromonas from hospitalised patients to human food through sewage water recycling
One of the main concerns of public health in recycling of waste water is that
pathogenic and drug resistant bacteria may persist in the treatment plant and from the plant
possibly disseminate to the environment or spread to the community through the food chain.
To be able to track the transmission of single Aeromonas strains a large number of isolates
were characterized. A series of different identification and typing methods were used to
understand the clonal relationships of the isolates, since identification of Aeromonas species
is not straightforward. This study was based on PhP-AE typing, and it was assumed that
isolates belonging to the same PhP-AE type probably belonged to the same clonal group of
strains. The consequent AFLP analysis of the PhP typed strains not only confirmed the
previous PhP grouping but also the subdivision of the PhP types into multiple genotypes
(Figure 4, Paper III).
4.3.2.1 Transmission of a virulent clonal lineage of Aeromonas bacteria
The PhP clonal groups of bacteria that persisted in different parts of the sewage
treatment plant were subjected to AFLP DNA fingerprinting to confirm the genetic
relatedness. High AFLP fingerprinting homology was found within the isolates belonging to
PhP major types 7 and 11 (Figure 5). One of these clonal groups (MT7) of strains that was
found in children suffering from diarrhoea, but also found in sewage water, duckweed, and
fish. The isolates of this clonal group were positive for the putative virulence factors tested.
On the other hand, the other clonal group (MT11) was found to be distributed in all parts of
the treatment plant samples. But the isolates of this latter type appeared to be avirulent. This
finding shows that both virulent and avirulent clonally related Aeromonas strains were able
to transmit from the hospital sewage water to the fish for human consumption through the
treatment plant.
38

39
4.3.2.2 Transmission of multiresistant clonal lineages of Aeromonas bacteria
Hospital sewage water receives bacteria that were previously exposed to different
antibiotics used for treatment human infections and is known to be a reservoir for many
multiresistant bacteria. Although only 6% of the tested isolates were gentamicin resistant,
these isolates were also resistant to all the eight antibiotics tested. PhP data analysis revealed
that they belonged to two PhP phenotypic groups. Further AFLP analysis of ten isolates
indicated a molecular homogeneity of the isolates thought they could be divided into two
AFLP types (Figure 6). The isolates originated from sewage water, duckweed growing
lagoon, duckweed, fish pond and fish. But these two AFLP types could not be detected
either in the human samples nor in control ponds. Our data revealed that two specific
multiresistant clonal lineages of Aeromonas bacteria persisted in the waste treatment plant
and that the same clonal groups were transmitted from hospital sewage water to cultured fish
for human consumption. Thus through recycle of sewage water virulent and multiresistance
strains of Aeromonas were disseminated to the environment through the sewage water
effluent.
"Imagination is more important than knowledge” Albert Einstein

Isolate no PhP type Origin* Isolation Year Species**
Virulence
107 MT7 Sewage 2001 A. veronii HG8/10 ++ 108
MT7 Lagoon 2001 A. veronii HG8/10 ++
104 MT7 Human 2001 A. veronii HG8/10 ++
103 MT7 Fish 2001 A. veronii HG8/10 ++
56 MT11 F. Pond 2001 A. veronii HG8/10 --
3 MT11 Sewage 2001 A. veronii HG8/10 --
54 MT11 P. Duckweed 2001 A. veronii HG8/10 --
55 MT11 L. Duckweed 2001 A. veronii HG8/10 --
53 MT11 Fish 2001 A. veronii HG8/10 --
40
Figure 5 UPGMA/Pearson correlation clustering of digitalized and normalized AFLP patterns of a selection of gentamicin resistant Aeromonas isolates from different
sites of the sewage water treatment plant.
Origin Amp Tet Ery Chl Nali Cep Stp Gen
Fish 128 32 128 64 128 64 128 64
Duckweed
128 32 128 64 128 64 128 64
Sewage 128 32 128 64 128 64 128 64
Lagoon 128 32 128 64 128 64 128 64
Fish pond 128 32 128 64 128 64 128 64
Lagoon 128 32 128 64 128 64 128 8
Duckweed 128 32 128 64 128 64 128 8
Fish pond 128 32 128 64 128 64 128 8
Fish 128 32 128 64 128 64 128 8
Sewage 128 32 128 64 128 64 128 8
100
959085807570
....AEW-119
....AEW-120
....AEW-116
....AEW-118
....AEW-117
....AEW-112
....AEW-115
....AEW-114
....AEW-113
....AEW-111
Figure 5 AFLP patterns of a selection of Aeromonas isolates from different sites of the sewage water treatment plant with isolation year, species and virulence.

41
4.3.3 Performance of the duckweed based sewage treatment plant
Protection of public health is the fundamental purpose of the waste water treatment
and aim is the reduction and elimination of the load of pathogenic microorganisms.
From the sites sampled at the sewage water treatment plant, the highest diversity
value of Aeromonas was found in the sewage water and the diversity of the populations
appeared to decrease further down the sewage water system from the duckweed lagoon to
the fish. In the treatment plant, the highest number of Aeromonas was found in the duckweed
lagoon and the lowest number in the fish pond. The high number of Aeromonas observed in
the duckweed lagoon was due to the fact that the samples from duckweed contained the
highest numbers of Aeromonas. However, the bacterial load was decreased from the sewage
water to the fish culture pond in the duckweed aquaculture based sewage treatment plant
(Table1 Paper III).
0,00
0,20
0,40
0,60
0,80
1,00
1,20
Sewage Lagoon Duckweed Fishpond Fish Patient Control
Tet Ery Chl Nal Stp Gen
Figure. 7 Relative frequency of resistant bacteria found in different sampling sites. The percentage of
resistant bacteria found in the initial stage of the treatment plant i.e. in sewage water was referred to 1
for all tested antibiotics.
The use of sewage water grown duckweed as fish feed might have an impact on the
occurrence of antimicrobial resistant bacteria in the fish pond. The percentage of resistant
bacteria was found to decrease for all antibiotics throughout the purification process in the
sewage water treatment process, i.e. from the sewage water to the duckweed to be used as
fish feed (Table 2 Paper V). The relative frequencies of resistance are shown to illustrate the
changes within the treatment system (Figure 7). In the sewage water treatment system we
studied here, the duckweed lagoon water is discharged to the river. There is a considerable
decrease of resistance in the lagoon water as compared to the sewage water.

42
In conclusion, duckweed based waste treatment system had a positive effect on both
the number and the resistance pattern of Aeromonas bacteria. Despite the fact that we found
high numbers Aeromonas attached to the duckweed.
4.3.4. Roles as vehicles in transmission
Duckweed as a vehicle for bacterial transmission: In this study, we followed up
the two unexpected results we found. Firstly, in the duckweed lagoon as well as in the fish
pond the duckweed itself appeared to contain higher numbers (>105 cfu/mL) of Aeromonas
(Table1, Paper III). Possibly, the high numbers of Aeromonas found in the duckweed might
be due to attachment and biofilm formation on the duckweed. In the sewage treatment
system studied here, the duckweed grown on sewage water is used as fish feed, which means
that the fish might become colonized with Aeromonas and provide a reservoir for further
transmission to humans. In fact, we were able to isolate specific Aeromonas clones in both
duckweed and fish (MT11 in Figure 4 Paper III), which supports the hypothesis that these
organisms can use duckweed as transmission vehicles. This could make up a disadvantage of
duckweed-based sewage treatment plants if such strains are pathogenic. In previous studies it
has been reported that aquatic animals and plants may function as reservoirs of pathogenic
bacteria, for example Vibrio cholerae may survive longer on duckweed than in water on
which duckweed was floating or in control water (88, 91, 92). Parveen and colleagues (148)
also observed high Aeromonas numbers on aquatic plants and phytoplankton in Bangladesh.
Furthermore, Vibrio cholerae were transmitted as far as from the Philippines to California by
using seaweed as vehicles (198).
Plasmids as a vehicle for resistance transfer: Secondly, although resistance
decreased generally from sewage water to the duckweed, there was an increase in resistance
frequency in the fish pond (Table 2 and Figure 2, Paper V). In our studied sewage water
treatment system the intensive fish culture ponds makes up a relatively confined unit with
little exchange of water. One explanation could be that the antimicrobial drugs, or their
residues together with multiresistant bacteria present in the sewage, enter the fish pond
through the use of duckweed for fish production. Another explanation could be the
horizontal transfer of resistance genes through plasmids, as it is known that vehicles for
resistance gene transfer are mobilisable or self-transmissible plasmids in conjugation with
transposons, IS-elements and integrons. In this study, 24 isolates were investigated for
transfer of resistance and plasmid, 21 of which were shown to contain plasmids and 18 of
these were positive in the conjugative transfer analysis. A single plasmid of 12 Kb, designed
pAEW1, was found in 81 % of the plasmid positive strains including all of the tested isolates
from the fish pond water. The presence of pAEW1 (arbitrary named in this study) was found

43
in isolates of different PhP and AFLP types. A multiresistant clonal group of Aeromonas
caviae also carried that plasmid. Altogether, conjugative transfer analyses in our laboratory
experiments indicate that the horizontal transfer of an R-plasmid might have taken place in
the sewage treatment system. Our results support the hypothesis that plasmid-mediated
spread of resistance in the duckweed aquaculture based sewage water facilities may play a
role in the antibiotic resistance trails. Thus the increased levels of resistant bacteria in the
fish pond may be the results of the introduction of resistant bacteria, a selective pressure
favouring growth of resistant bacteria and/or spread of resistance plasmids within the
Aeromonas population.
Amoeba as a vehicle for Aeromonas transmission (preliminary results):
Free-living amoebas have attracted much interest during the past decades as reservoirs for
different amoeba resistant bacteria (17). The role free-living amoeba plays in the
transmission of both pathogenic and non-pathogenic bacteria have been the subjects of
discussions. To our present knowledge amoeba may increase the transmission rate of
bacteria by acting as an endosymbiont or as a vehicle by carrying a huge number of
microorganisms (62, 63, 125, 18, 19, 123). Like Aeromonas, Acanthamoeba have previously
been isolated from different sewage water systems. To be able to understand the possible
transfer routes of Aeromonas via free-living Acanthamoeba or the utilization of
Acanthamoeba as a natural host, we performed co-culture experiments. We found a mutual
interference of growth between the amoebae and the Aeromonas strains, dependent on the
ratio of bacteria to amoebae cells. But we found that at the high concentrations (107 to 109
CFU ml-1) Aeromonas was able to repress amoeba growth, and within one week the amoeba
cells decreased with 95%. Interestingly, Aeromonas was never able to lyse all amoeba cells,
and even after two weeks of time 100 cell/ml were still left. To investigate the direct
interaction between Aeromonas and A. castellanii, we tagged A. hydrophila (AEW 20) with
GFP and observed the adherence of the bacteria to the amoeba by fluorescent and phase-
contrast microscopy. We observed that the bacteria were always attached in clumps on the
amoeba surfaces, i.e. thus forming a biofilm on the amoebal cell wall (Figure 8 A,
Aeromonas biofilm on Amoeba). These observations show that in a natural setting,
Aeromonas spp may adhere to the amoeba surface and may use amoeba as a vehicle for
transmission. To study the invasion ability of bacteria into Acanthamoeba castellanii, we
carried out gentamicin protection assay, using A. hydrophila (AEW44), A. veronii biovar
sobria (AEW98) and A. caviae (AEW1) at a multiplicity of infection rate of 1:100. We were
not able to recover any intracellular bacteria from the amoeba two hours post infection. But
we recovered a significant number of bacteria 16 hours post infection from the amoebae.
However, we obtained the highest recovery rates of intracellular bacteria after three days of

44
incubation. Fluorescent microscopic observations of the GFP-tagged bacteria indicated
internalization of Aeromonas by amoeba (Figure 5B). Our results suggest that free-living
amoeba may increase the transmission rate of Aeromonas as a vehicle or in endosymbiosis,
wherein they may multiply and later lyse the amoeba in order to disperse. In summary, we
show here that the co-existence of Aeromonas and amoeba in the sewage water increase the
risk of transmission as Aeromonas may use the amoeba as a natural host.
Figure 8 A. Phase-contrast microscopic images of Aeromonas AEW20 adherence to the Acanthamoeba within 15 minutes of co-culture. B-C. Fluorescent microscope images show internalization of Aeromonas AEW 20 by Acanthamoeba castellanii and the increased number of intracellular bacteria with time.
A B C
4.3.5 Biofilm formation of Aeromonas (unpublished data)
Prevalence of biofilm formation of Aeromonas: A total of 92 Aeromonas isolates
were tested for biofilm formation in glass tubes. 25 of them (27%) showed strong biofilm
formation, 41 (44%) showed moderate biofilm formation and the remaining 26 (29%)
isolates did not show any biofilm formation in our setting (Figure 9). Species distribution on
biofilm formation is shown in figure 9. Interestingly, it was found that 100% of the
Aeromonas hydrophilia tested were able to form biofilm either strongly or moderately.
0
10
20
30
40
50
60
"++++ to +++++ "++ to +++ "-- to +
Arbitrary biofilm formation score
Pece
ntag
es
A.hydrophila A.veronii A.caviae
Figure 9 Distribution of species on Aeromonas biofilm formation.

Strong biofilm formation in persistent clonal groups: When the biofilm formation
scores (arbitrary) were analysed in relation to the PhP clonal types, it was found that all
members of PhP MT types 7 and 11 showed strong biofilm formation (Figure 10).
Interestingly, members of these clonal groups were also been shown to be persistent in
different sites of the hospital sewage water treatment plant and to belong to the clonal groups
that are transmitted from the hospitalised patients to the fish to be consumed by human,
through the sewage water treatment system (Paper III). Taken together, our data suggest that
a strong biofilm formation capacity of PhP MT types 7 and 11 clonal groups might have
contributed to the successive persistence and transmission.
0
1
2
3
4
5
6
Arb
itrar
y bi
ofilm
form
atio
n
Fig
Aeromonas.
4.4. Genetreatmen
Wa
organisms f
of microorg
microorgan
according t
removing s
different en
the waste
organisms.
MT7 MT 8 MT 11 MT 3 MT 2
45
ure 10 Biofilm formation capacity of the members of different PhP clonal group of
ral comments on duckweed aquaculture based sewage t plant
ste water treatment plants seldom have been planned to remove pathogenic
rom waste water. Conventional waste water treatment plants reduce the numbers
anisms but the waste water effluents may still contain high numbers of faecal
isms. The removal efficiency of pathogenic and indicator organisms may vary
o the treatment process type, retention time, temperature and the efficiency of
uspended solids. Survival and persistence abilities of different organisms in
vironments may vary and organisms have been demonstrated even multiply in
water treatment system. Thus removal efficiencies may also vary between
Efficient removal of pathogens from waste water is a critical task, since waste

46
water discharge may increase the pathogen contamination in surface water and result in the
increase of food and water borne infections.
The mainstream sewage water treatment, such as activated sludge and waste
stabilization pond systems are too costly for developing countries (61). Duckweed
aquaculture based sewage water treatment systems are a relatively new concept for waste
treatment. The successful implementation of duckweed aquaculture based waste treatment
systems with the benefit of using duckweed for cost recovery far out weighted the problem
with attendant risks of recycling of pathogens. Using the indicator bacteria Aeromonas, we
have shown that virulent and avirulent, and drug resistant bacterium bacteria could be
transmitted from the hospital sewage water to the fish to be consumed by humans through
the recycling of sewage water in a duckweed aquaculture based hospital sewage water
treatment plant in Bangladesh.
Unfortunately, there is no data available on the prevalence of gastroenteritis in
relation to consumption of fish associated with sewage water treatment but there is well-
documented data on shellfish as vectors of infectious agents in USA. Since 1800s, more than
400 infectious disease outbreaks and 14,000 cases have been reported in USA. These
outbreaks are attributed to bacterial and viral agents that are associated either with human
wastes (sewage effluents that have received variable levels of treatment) or to bacterial
pathogens indigenous to coastal marine environments (e.g., Vibrio spp). Before the 1950s,
the most common illness associated with the consumption of shellfish was typhoid fever. In
the mid-1920s there were several outbreaks of typhoid fever (more than 1,500 cases and 150
deaths) in several US cities. After the outbreak, the U.S public health service established
regulations for the sanitary control of shellfish. National shellfish sanitation program appears
to have been effective since then no shellfish associated typhoid cases have been reported in
USA in almost 40 years. The incidence of typhoid fever began to decline for at least two
reasons, i) the technology of the sewage treatment was improved, particularly the removal of
pathogen associated particles, ii) the water quality standard was developed for the shellfish
growing area. These epidemiological results suggest that aquaculture (shellfish) related
infectious diseases could be controlled through proper management of waste water and
aquaculture water.
In our studied duckweed aquaculture based sewage treatment plant, there is a clear
vertical transmission of pathogenic and drug resistant Aeromonas from the sewage water to
the fish through the duckweed. Duckweed itself was also shown to contained high amount of
Aeromonas. These results necessitate a few additional steps to reduce the load of pathogens

47
from duckweed before using it as fish feed. The additional steps could include drying of the
weed and/or washing with the fresh water. Another additional step could be that the water in
the fish culture pond was changed more often. Furthermore, to decreases the gastroenteritis
in relation to consumption of fish associated with sewage water treatment i) Fish should not
be marketed from waste water grown area unless they are shown to be free from any
suspected or dangerous disease producing organisms; ii) Epidemiological studies should be
conducted for all outbreaks (epidemics) that implicate fish as sources, so that the sources of
the fish could be accurately traced.
This investigation serves as a first step of risk assessment, i.e. potential hazard
identification in the duckweed aquaculture based sewage water treatment system. It is
therefore recommended that a quality assurance system based on the HACCP principles
should be introduced to ensure the microbiological safety of the waste recycled food. Thus
our results support that sewage water recycling in aquaculture constitutes a potential health
hazard and further studies are needed for a proper characterization of the risks involved.
Although a detailed assessment of the epidemiological relationship between the
prevalence of enteric disease and the consumption of fish produced in the sewage treatment
system fell outside the scope of the present investigation, this study suggests that it would be
important to perform a community-based study on the prevalence of gastroenteritis in
relation to consumption of fish associated with sewage water treatment.

48
4.5 Usefulness of “ScanMIC” method for antibiotic related hazard identification (Paper IV, V)
Increased bacterial resistance to antimicrobial agents has initiated the monitoring of
the resistance in isolates from humans, foods, animals and environments through national
surveillance and research programs. For antibiotic related hazard identification surveillances
in human food chains, large numbers of isolated are to be investigated. Therefore, a rapid
and effective method is needed to screen large numbers of isolates for antibiotic resistance
patterns in a short time.
In this study, a “ScanMIC” method, i.e. a colorimetric MIC (minimum inhibitory
concentration) method has been developed for susceptibility testing of gram-negative
fermentative bacteria. The method is a slight modification of the NCCLS (National
Committee for Clinical Laboratory Standard, USA) recommended broth microdilution
method, using a redox indicator (TTC) to facilitate the estimation of the bacterial growth
inhibition in a microplate, and a flatbed scanner to capture the microplate image. An in-
house software was developed to transform the microplate image into numerical values
based on the amount of bacterial growth and to generate the MIC values automatically. The
general concept of the ScanMIC method is shown below.
To evaluate the ScanMIC method, we compared the “ScanMIC” method to the
NCCLS recommended broth microdilution method using 197 coliform strains and 288
Aeromonas strains in duplicate against seven and eight antimicrobial agents respectively.
The absolute interpretative categorical agreement was obtained in 92.4 % and 96.7 % of the
assays for coliform and Aeromonas respectively. The percent agreement for MICs
differences (within ± 1 log2 dilution) was obtained in 96% and 97 % for “ScanMIC” versus
broth microdilution for coliform and Aeromonas respectively (Paper IV, Table 1). In both
cases the ScanMIC method was found to meet the performance criteria guideline for
automated susceptibility testing, which has been suggested in the NCCLS recommendations.
The ScanMIC method has been used in this practical study, i.e. epidemiological
surveys of resistance patterns from the hospital sewage water treatment plant, and it was
found to be easy to use for large number of isolates. The main advantage of this method was
the use of a growth indicator, a cheap normal flat bed scanner to capture the microplate
image and the software that directly generated the MIC values from the microplate image.
Besides, the microplate images could be saved and final results sheet with all MIC values
were automatically created. Thus the method could be a useful method for epidemiological
surveys of resistance patterns of large numbers of bacteria in the hospital as well as in the
environment.

49
Schematic presentation of the ScanMIC method
Antibiotic microdilution in microplate with added bacteria
Overnight growth
Scanned the microplate
Software analysis of the scanned microplate; pellet values transferred to numerical values Final result sheet provided by the software
1 2 3 4 5 6 7 8 9 10 11 12A 89 90 84 80 87 84 84 84 81 89 90 94B 86 88 82 67 81 81 83 81 0 82 85 85C 83 90 78 0 80 78 71 81 0 83 90 89D 82 87 82 0 82 78 0 78 0 81 89 94E 80 85 79 0 82 82 0 76 0 83 85 93F 80 0 80 0 82 82 0 80 0 84 93 90G 79 0 81 0 79 75 0 75 0 86 84 90H 76 0 71 0 80 0 0 70 0 88 82 85
Amp Tet Chl Nal Cep Stp Gen Amp Tet Chl Nal CepMIC >128 16 >128 4 >64 128 4 >128 <1 >128 >128 >64
Isolate no Amp Tet Chl Nal Cep Stp Gen Amp Tet Chl Nal Cep 1 256 64 8 8 8 64 2 128 4 16 16 8 2 256 16 4 4 4 16 64 2 2 4 32 4 3 16 2 64 4 2 8 8 32 1 4 16 64 4 4 2 32 4 4 2 4 64 1 8 8 16 5 256 64 4 8 8 64 2 4 64 16 32 1 6 256 16 16 4 4 16 2 4 2 8 32 32 7 16 2 4 4 2 8 2 16 1 32 8 64 8 4 2 8 4 4 2 4 64 1 8 32 2

50
4.6 Cyclic-di-GMP signalling in Aeromonas (preliminary data)
Cyclic nucleotides, such as cyclic adenosine monophosphate (cAMP) and cyclic
guanosine monophosphate (cGMP) are important low molecular weight signalling molecules
in both prokaryotes and eukaryotes (82, 120, 164). In eukaryotes, cGMP is known to be
involved in different mechanisms such as regulation of metabolic processes, cell
differentiation, immunity, and behavioural processes (82, 105, 171). Prokaryotes in general
do not seem to use cGMP for signalling, but recent discoveries suggest that bacteria make
extensive use of a cyclic guanosine compound, cyclic-guanosine–monophosphate (c-di-
GMP) to regulate cell surface associated trials and community behaviours like biofilm
formation in a number of bacterial species such as Yersinia pestis, Pseudomonas aeruginosa,
Salmonella enterica , Vibrio cholera, Gluconacetobacter xylinus (42). It has also previously
been shown that GGDEF and EAL domains are involved in the turnover of cyclic-di-GMP in
pathogenic Salmonella aeruginosa, Pseudomonas aeruginosa and E. coli strains (171). But
nothing is known about the c-di-GMP signalling in Aeromonas spp.
GGDEF and EAL domains protein regulate the cyclic di-GMP levels in
Aeromonas: To elucidate the role of GGDEF and EAL domains in Aeromonas, we
introduced AdrA (GGDEF domain from S. typhimurium) and YhjH (EAL domain from S.
Typhimurium) domains on plasmid carriers to our selected strain. The c-di-GMP
concentration rose to a level of 0.297 nmol mg-1 cells when AdrA was introduced compared
to 0.002 nmol mg-1 cells by vector control strain, and when YhjH was introduced the levels
of c-di-GMP was 0,005 nmol mg-1 cells. Thus our results showed that c-di-GMP level of
Aeromonas are significantly enhanced when the GGDEF domain containing protein AdrA is
expressed. When the EAL domain containing protein YhjH is expressed, c-di-GMP levels
were only slightly enhanced. This phenomenon was also seen in S .typhimurium when YhjH
was expressed. (171).
Phenotypic regulation: GGDEF and/ or EAL domain proteins are known to
regulate the phenotypic trial in different bacterial species such as wrinkled colonies
behaviour for Pseudomonas, radar colonies behaviour for Salmonella, rugose colonies
behaviour for Vibrio (42). In this study, the phenotypic trail was altered when AdrA was
introduced to Aeromonas strains. Figure 11A shows the pink colony of Aeromonas veronii
when AdrA was expressed. Furthermore, when the bacterial colony was stirred with a tooth
pick, it was found that AdrA expressing colonies were highly sticky but neither the YhjH nor
the vector control expressing strain showed such behaviour. A dramatically behaviour
change was observed when swimming motility was tested with the GGDEF and EAL

51
expressing stains. Figure 11B shows that the swimming motility was increased when YhjH
was expressed and the motility was decreased when AdrA was expressed.
AAdrA
B
Dow
nregulated by AdrA
Upr
egul
ated
by
YhJ
H
Vector Y
hjH
Vector Control
Fig 11. A. Phenotyping regulation of Aeromonas when AdrA was expressed; B. Swimming behaviours of Aeromonas was repressed when AdrA was expressed and enhanced when YhjH was expressed.
Adherence on abiotic surface: Biofilm formation on glass test tubes was tested
with the AdrA and YhjH expressing strains. Biofilm formation was found to be increased
when AdrA was expressed and decreased when Yhjh was expressed (Figure 12), as shown
by previous studies on the bacterial species Salmonella typhimurium and Pseudomonas
aeruginosa (171).
CBA
Figure 12. Pattern of adherence of Aeromonas to glass surface; A, Vector control; B, AdrA expressing
strain; YhjH expressing strain.
Adhesion to plant surfaces: c-di-GMP signalling has been suggested to play a
conserved ecological role in bacterial adhesion to plant and animal surfaces with the GGDEF
type regulators (55, 158, 160, 196). In this study, the influence of c-di-GMP on Aeromonas
attachment to duckweed was studied. Figure 13 shows the adherence ability to the duckweed

52
surface. It can easily be seen that there is a considerably higher adherence ability of AdrA
expressing Aeromonas than of the YhjH or vector control strains. Thus our data indicate that
GGDEF type regulatory protein AdrA may have an effect on Aeromonas attachment to the
duckweed surface.
0
10000
20000
30000
40000
AdrA YhjH Vector
CFU
/duc
kwee
d (a
pp. 0
.01g
)
Figure 13 Adherence ability of AdrA, YhjH and Vector control expressing Aeromonas strains to the duckweed surface
Influences on quorum sensing molecules: Quorum sensing is a mechanism for
controlling gene expression in response to an expanding bacterial population. For
identification and analysis of quorum sensing in Aeromonas bacteria i..e, C4 -HSL
bioluminescent sensors have been developed (Figure 14), and are based on the LuxR
regulatory mechanism, which reflects the need to accumulate sufficient pheromones to
trigger the induction of gene expression of LuxR homologues (178, 179)
Biosensor strain
Pheromone producer (AdrA) Vector control
YhJH
Figure 14 Pheromone production of AdrA, YhJH and vector expressing strains against biosensor strain Chromobacterium violaceum CV026 on agar plate
To study the influence of c-di-GMP on quorum sensing molecules, HSL were
extracted from AdrA, YhjH and vector expressing Aeromonas strains. To estimate the
extracted HSL, bioluminescence were measured as relative light units using the detector E.
coli (pSB536) in a microtiter plate counter luminometer. It can easily be seen that AdrA

expressing strains produce considerably higher luminescence that the YhjH and vector
control strains, i.e. they contain higher level of C4-HSL (Table 4). Our results suggest that,
the level of c-di-GMP is related to the level of quorum sensing molecules C4-HSL. Thus c-
di-GMP may influence the quorum sensing of Aeromonas spp.
Table 4. Relative light units were measured as a function of C4-HSL from extracted HSL of AdrA, YhJH and vector expressing strains.
Strains Relative light units
expressed by the sensor
AdrA 31874
YhjH 19486
Vector 18667
Altogether, the AdrA and YhJH expressing strains of Aeromonas were constructed
that contain different levels of c-di-GMP. They showed different morphotypes, motility,
biofilm activities, adhesion abilities to plant surfaces and amounts of quorum sensing
molecules C4-HSL. Thus, c-di-GMP may play an important role in the physiology of
Aeromonas.
53

ABSTRACTS OF INCLUDED PAPERS I. Evidence for transmission between humans and the environment of a
nosocomial strain of Enterococcus faecium. The prevalence of an ampicillin and
ciprofloxacin resistant strain of Enterococcus faecium, named FMSE1, with a characteristic
biochemical fingerprinting pattern was investigated from database comprising 9,676 isolates
of enterococci from 13 different kinds of sources. The FMSE1 type was identified in 1.4% of
all isolates. The FEMS1 type was most commonly found in samples collected from hospital
sewage (50%), followed by surface water (35%), treated urban sewage water (28%),
untreated urban sewage water (17%) and rarely in healthy children (0.8%) and animal origin
(2%). PFGE typing of the isolates showed that they were genetically closely related. This
suggests that a possible transmission route for nosocomial E. faecium from patients in
hospital is through hospital and urban sewage waters and further via treatment plants to
surface water to healthy humans.
II. Identification and Characterization of Pathogenic Aeromonas veronii
Biovar Sobria Associated with Epizootic Ulcerative Syndrome in Fish in
Bangladesh. Epizootic ulcerative syndrome is a fish disease characterized by the presence
of severe, open dermal ulcer on the head, on the middle of the body and on the dorsal regions
of the fish. We identified a pathogenic unique clonal group of Aeromonas veronii biovar
sobria associated with Epizootic Ulcerative Syndrome (EUS) in fish in Bangladesh by the
aid of the Pheneplate biochemical fingerprinting system, fatty acid methyl ester analysis
(FAME) and amplified fragment length polymorphism analysis (AFLP). In this study, the
EUS associated isolates produced haemolysin and cytotoxin, carried the cytolytic enterotoxin
gene, exerted adhesion and invasion ability to fish cell lines and a unique ability to
agglutinate fish erythrocytes. Our results suggest that this clonal group may be considered a
possible causative agent of EUS in fish in Bangladesh.
III. Persistence and Transmission of Aeromonas in a Duckweed
Aquaculture-Based Hospital Sewage Water Recycling Plant. To study the
persistence and transmission of Aeromonas spp. in a duckweed aquaculture-based hospital
sewage water treatment plant in Bangladesh, 1,364 presumptive Aeromonas isolates from a
total of 691 samples were characterised. Aeromonas spp. were found at all sampled sites of
the sewage treatment plant, in 40% of the control environmental samples, in 8.5% of the
faeces from hospitalised children suffering from diarrhoea as well as in 3.5% of the faeces
from healthy control humans. The mean CFU/ml and diversities of Aeromonas species were
found to decrease throughout the purification process indicating that a part of the Aeromonas
population could be eradicated in the sewage treatment system. High numbers of Aeromonas
54

were found associated to the duckweed, which may point to the fact, that aquatic weed may
serve as a reservoir of Aeromonas in the environment. Certain PhP phenotypes from both
duckweed and fish or phenotypes isolated during two subsequent years or from hospitalised
humans suffering from diarrhoea, from different stages of the sewage treatment and from
human consumable fish, were found to be indistinguishable by the AFLP fingerprinting
method, and some of these clonal groups showed virulence properties. Thus, our data
indicate that a few genetically stable Aeromonas clones may have the potential to persist in
the sewage treatment plant for a prolonged period and can possibly be transmitted from the
sewage water to the fish for human consumption via the harvesting of duckweed in the
treatment system.
IV. Evaluation of a scanner assisted colorimetric MIC method for
susceptibility testing of Gram-negative fermentative bacteria. To facilitate the
screening of antibiotic resistance patterns of large numbers of isolates in a short time, we
developed a semi automated colorimetric MIC method, the “ScanMIC” method. The method
used concentration gradients of antibiotics in 96-well microplates, tetrazolium salt as growth
indicator and a flat bed scanner to read the results and an in-house developed software for
image analysis and to calculate and store the MIC values. We compared the “ScanMIC”
method to the NCCLS recommended broth micro dilution method using 197 coliform strains
against seven antibacterial agents and a high agreement (96%) between our “ScanMIC”
method and the NCCLS reference method for MIC determination was obtained. We
conclude that the “ScanMIC” method might become an acceptable and useful method for
epidemiological surveys of resistance patterns of large numbers of coliform bacteria.
V. Influence of a duckweed-aquaculture based hospital sewage water
recycling plant on the antibiotic resistance of Aeromonas spp. as determined by
the ScanMIC method. To understand the influence of duckweed-aquaculture based
hospital sewage water treatment on the resistance patterns of the Aeromonas spp, we
determined the resistance for 288 Aeromonas strains against eight antibacterial agents by the
ScanMIC method as well as with the NCCLS reference method. The ScanMIC method was
again found to be an acceptable (97% agreement) and useful method for susceptibility
determinations in epidemiological surveys on environmental bacteria. The frequencies of
resistant strains decreased during the purification process in the duckweed ponds and even
lower frequencies of resistant bacteria were found attaching to the duckweed used in fish
feed. However, frequencies of resistance were found to increase again in the fish culture
pond. A multiresistant biochemical phenotype was found to persist in all sites of the
treatment plant, and this type was verified by molecular fingerprinting (AFLP) to consist of
one single clone of Aeromonas caviae. Such a clone might spread resistance to the
55

environment, both through the outlet of purified wastewater and through the human
consumption of cultivated fish.
56

ACKNOWLEDGEMENTS
This work has been performed with financial support from Swedish International
Development Agencies (SIDA) Project No 1999-255.
I would like to thank each and everyone who helped me during these years to finish my PhD
works - colleagues, family, and friends. Especially I would like to thank:
My supervisor Roland Möllby, for giving me the freedom and opportunities to carry out my
ideas while still keeping me on track, for teaching me how to write a manuscript, for listening to all
my stupid research ideas, for helping me to write all the project proposals and for giving me a new
name Mokhi (which I did not accept in the beginning !). I have enjoyed this journey with you. I
would also like to thank Gunneli (Roland’s wife) as I have stolen some Roland’s time from her.
My co-supervisor Inger Kuhn, although I still have a problem to writing her name. I must
thank to her for helping me improving the project proposals and manuscripts, and thanks for carefully
listening to all the PhP associated problems.
My unofficial supervisor Ute Römling, for listening to my all ideas, for helping me to
improve my knowledge in molecular biology, for improving my project proposals in the middle of the
night and during weekends and for letting me use her laboratory materials.
The members of Roland Möllby’s group; Patricia Colque (lab mum) for being nice
sometimes !!!, and for encouraging me during my hard times (Mokhi fight fight !!!), Jenny Gabrielson
for helping me with all the computer problem (magic fingers), Aina Iversen for giving me advice
about different things (family, research) and for revising my thesis, Beatrix for being a nice colleague
and helping me revising my thesis, Sara for being a nice colleague, Lena Guldevall for being a lab
partner and all the nice music in the Lab, Maj Ringman for being a nice friend, Magnus for helping
me with all the computer problems. Also thanks to Ruth, Felix, Margarita, Mats (Pling !), Anna Karin,
Hadi Abd, Lena Gezelius, Barbro Olsson-Liljequist and Malin Grape for being to nice with me. Geert
Huys and members of his group for carefully typing the bacteria. The members of Ute’s group: Kader,
Roger, Ulrich, Dinesh, Xiaoda, Devrim, Agaristi for being nice with me and for sharing the lab and
writing space.
Professor Monica Thelestam, Professor Staffan Arvidsson, Professor Gunnar Sandstrom,
Docent Agneta Richter-Dahlfors, Docent Elisabeth Norin, Docent Öjar Melefors and their group
members, MTC administrative people: Eva Pijnenburg, Anna Lögdberg, Leena Norenius, MTC
prefect Mats Wahlgren for helping me with different things in different times.
Especial thanks to Motiur and his lab members, the members of ICDDR,B for laboratory
support and Prism Bangladesh Ltd Staffs and Kumudini Hospital staffs for sampling support.
My family - my parents, my brothers and sisters, my wife Shapla and all my friends for
encouraging me during these years.
57

REFERENCES 1. Abbott, S., W. K. Cheung, S. K. Bystrom, T. Malekzadeh, and J. M. Janda. 1992.
Identification of Aeromonas strains to the genospecies level in the clinical laboratory. J. Clin Microbiol. 30:1262-1266.
2. Abbott, S. L., W. K. Cheung, and J. M. Janda. 2003. The genus Aeromonas: biochemical characteristics, atypical reactions, and phenotypic identification schemes. J Clin Microbiol 41:2348-57.
3. Abd, H., T. Johansson, I. Golovliov, G. Sandstrom, and M. Forsman. 2003. Survival and growth of Francisella tularensis in Acanthamoeba castellanii. Appl Environ Microbiol 69:600-6.
4. Abdullah, A. I., C. A. Hart, and C. Winstanley. 2003. Molecular characterization and distribution of virulence-associated genes amongst Aeromonas isolates from Libya. J Appl Microbiol 95:1001-7.
5. Abrami, L., M. Fivaz, P. E. Glauser, N. Sugimoto, C. Zurzolo, and F. G. van der Goot. 2003. Sensitivity of polarized epithelial cells to the pore-forming toxin aerolysin. Infect Immun 71:739-46.
6. Abu-Zeid, M. A. 1998. Water and sustainable development: the vision for world water, life and the environment. Water Policy 1:9-19.
7. Alaerts G. J., M. Rahman. P. Kelderman. 1996. Performance analysis of a full scale duckweed covered lagoon. Water Res 30:843-852.
8. Albert, M. J., M. Ansaruzzaman, K. A. Talukder, A. K. Chopra, I. Kuhn, M. Rahman, A. S. Faruque, M. S. Islam, R. B. Sack, and R. Mollby. 2000. Prevalence of enterotoxin genes in Aeromonas spp. isolated from children with diarrhea, healthy controls, and the environment. J Clin Microbiol 38:3785-90.
9. Albert, M. J., A. S. G. Faruque, S. M. Faruque, R. B. Sack, and D. Mahalanabis. 1999. Case-Control Study of Enteropathogens Associated with Childhood Diarrhea in Dhaka, Bangladesh. J. Clin. Microbiol. 37:3458-3464.
10. Ali, A., A. M. Carnahan, M. Altwegg, J. LuthyHottenstein, and S. W. Joseph. 1996. Aeromonas bestiarum sp nov (formerly genomospecies DNA group 2 A-hydrophila), a new species isolated from non-human sources. Med Microbiol Letters 5:156-165.
11. Andre-Fontaine, G., P. Monfort, M. Buggin-Daubie, C. Filloneau, and J. P. Ganiere. 1995. Fatal disease mimicking leptospirosis in a dog, caused by Aeromonas hydrophila. Comp Immunol Microbiol Infect Dis 18:69-72.
12. Anita, H. 2005. Thesis. Royal Institute of Technology (KTH), Stockholm. 13. Araujo, R. M., R. M. Arribas, and R. Pares. 1991. Distribution of Aeromonas species in
waters with different levels of pollution. J Appl Bacteriol 71:182-6. 14. Aulicino, F. A., and F. Pastoni. 2004. Microorganisms surviving in drinking water systems
and related problems. Ann Ig 16:265-72. 15. Aurell, H., J. Etienne, F. Forey, M. Reyrolle, P. Girardo, P. Farge, B. Decludt, C. Campese,
F. Vandenesch, and S. Jarraud. 2003. Legionella pneumophila serogroup 1 strain paris: Endemic distribution throughout France. J Clin Microbiol 41:3320-3322.
16. Austin, B., and D. Allen-Austin. 1985. Bacterial pathogens of fish. J Appl Bacteriol 58:483-506.
17. Barker, J., and M. R. Brown. 1994. Trojan horses of the microbial world: protozoa and the survival of bacterial pathogens in the environment. Microbiology 140 :1253-9.
18. Blackwood, R. A., C. K. Rode, C. L. Pierson, and C. A. Bloch. 1997. Pulsed-field gel electrophoresis genomic fingerprinting of hospital Escherichia coli bacteraemia isolates. J Med Microbiol 46:506-10.
19. Blanc, D. S. 2004. The use of molecular typing for epidemiological surveillance and investigation of endemic nosocomial infections. Infect Genet Evol 4:193-7.
20. Blondeau, J. M. 2004. Current issues in the management of urinary tract infections: extended-release ciprofloxacin as a novel treatment option. Drugs 64:611-28.
21. Bomo, A. M., M. V. Storey, and N. J. Ashbolt. 2004. Detection, integration and persistence of aeromonads in water distribution pipe biofilms. J Water Health 2:83-96.
22. Bonten, M. J., M. K. Hayden, C. Nathan, T. W. Rice, and R. A. Weinstein. 1998. Stability of vancomycin-resistant enterococcal genotypes isolated from long-term-colonized patients. J Infect Dis 177:378-82.
58

23. Borchardt, M. A., M. E. Stemper, and J. H. Standridge. 2003. Aeromonas isolates from human diarrheic stool and groundwater compared by pulsed-field gel electrophoresis. Emerg Infect Dis 9:224-8.
24. Borrell, N., S. G. Acinas, M. J. Figueras, and A. J. Martinez-Murcia. 1997. Identification of Aeromonas clinical isolates by restriction fragment length polymorphism of PCR-amplified 16S rRNA genes. J Clin Microbiol 35:1671-4.
25. Braun, M., K. Stuber, Y. Schlatter, T. Wahli, P. Kuhnert, and J. Frey. 2002. Characterization of an ADP-ribosyltransferase toxin (AexT) from Aeromonas salmonicida subsp. salmonicida. J Bacteriol 184:1851-8.
26. Bryan, F. L., and M. P. Doyle. 1995. Health risks and consequences of Salmonella and Campylobacter jejuni in raw poultry. J Food Protection 58:326-344.
27. Burke, V., M. Gracey, J. Robinson, D. Peck, J. Beaman, and C. Bundell. 1983. The microbiology of childhood gastroenteritis: Aeromonas species and other infective agents. J Infect Dis 148:68-74.
28. Camara, M., P. Williams, and A. Hardman. 2002. Controlling infection by tuning in and turning down the volume of bacterial small-talk. Lancet Infect Dis 2:667-76.
29. Carnahan, A., G. R. Fanning, and S. W. Joseph. 1991. Aeromonas jandaei (formerly genospecies DNA group 9 A. sobria), a new sucrose-negative species isolated from clinical specimens. J Clin Microbiol 29:560-4.
30. Carnahan, A. M., and M. Altwegg. 1996. Taxonomy, p. 1-38. In M. A. Eds. B Austin, P.J. Gosling and S. Joseph (ed.), In The genus Aeromonas. Wileys and Sons; Chichester.
31. Carnahan, A. M., S. Behram, and S. W. Joseph. 1991. Aerokey II: a flexible key for identifying clinical Aeromonas species. J. Clin Microbiol. 29:2843-9.
32. Carnahan, A. M., T. Chakraborty, G. R. Fanning, D. Verma, A. Ali, J. M. Janda, and S. W. Joseph. 1991. Aeromonas trota sp. nov., an ampicillin-susceptible species isolated from clinical specimens. J Clin Microbiol 29:1206-10.
33. Carr, R. M., U. J. Blumenthal, and D. D. Mara. 2004. Guidelines for the safe use of wastewater in agriculture: revisiting WHO guidelines. Water Sci Technol 50:31-8.
34. Chauret, C., C. Volk, R. Creason, J. Jarosh, J. Robinson, and C. Warnes. 2001. Detection of Aeromonas hydrophila in a drinking-water distribution system: a field and pilot study. Can J Microbiol 47:782-6.
35. Chopra, A. K., C. W. Houston, J. W. Peterson, and G. F. Jin. 1993. Cloning, expression, and sequence analysis of a cytolytic enterotoxin gene from Aeromonas hydrophila. Can J Microbiol 39:513-23.
36. Chopra, A. K., X. Xu, D. Ribardo, M. Gonzalez, K. Kuhl, J. W. Peterson, and C. W. Houston. 2000. The cytotoxic enterotoxin of Aeromonas hydrophila induces proinflammatory cytokine production and activates arachidonic acid metabolism in macrophages. Infect Immun 68:2808-18.
37. Coleman, M. E. 2003. Interactions of predictive microbiology and risk assessment. Risk Anal 23:175-8.
38. Conwell, R.R, M.T McDonell and J. De Ley.1986. Proposal of recognize the family Aeromonadaceae farm. nov. Int. J. Syst. Bacteriol. 36:473-477..
39. Costerton, J. W., Z. Lewandowski, D. E. Caldwell, D. R. Korber, and H. M. Lappin-Scott. 1995. Microbial biofilms. Annu Rev Microbiol 49:711-45.
40. Costerton, J. W., P. S. Stewart, and E. P. Greenberg. 1999. Bacterial biofilms: a common cause of persistent infections. Science 284:1318-22.
41. D.W. Warburton, B. H., C. Crawford, R. Foster, C. Fox, L. Gour and P. Krol. 1998. A further review of the microbiological quality of bottled water sold in Canada: 1992–1997 survey results. Int J Food Microbiol 39:3–17.
42. D'Argenio, D. A., and S. I. Miller. 2004. Cyclic di-GMP as a bacterial second messenger. Microbiology 150:2497-502.
43. Davey, M. E., and A. O'Toole G. 2000. Microbial biofilms: from ecology to molecular genetics. Microbiol Mol Biol Rev 64:847-67.
44. Doring, G., M. Ulrich, W. Muller, J. Bitzer, L. Schmidt-Koenig, L. Munst, H. Grupp, C. Wolz, M. Stern, and a. Botzenhart et. 1991. Generation of Pseudomonas aeruginosa aerosols during handwashing from contaminated sink drains, transmission to hands of hospital personnel, and its prevention by use of a new heating device. Int. J Hygiene Environ Med 191:494-505.
45. Dronda, F., A. Gonzalez-Lopez, J. I. Fernandez-Martin, M. Alonso-Sanz, F. Chaves, and L. Lopez-Cubero. 1996. Primary cutaneous cryptococcosis and bacterial pneumonia caused by
59

Pseudomonas aeruginosa in AIDS: clinical case and review of the literature. Enferm Infecc Microbiol Clin 14:101-5.
46. Duthie, R., T. W. Ling, A. F. Cheng, and G. L. French. 1995. Aeromonas septicaemia in Hong Kong species distribution and associated disease. J Infect 30:241-4.
47. Eaton, T. J., and M. J. Gasson. 2002. A variant enterococcal surface protein Esp(fm) in Enterococcus faecium; distribution among food, commensal, medical, and environmental isolates. FEMS Microbiol Lett 216:269-75.
48. Edlund, C., L. Barkholt, B. Olsson-Liljequist, and C. E. Nord. 1997. Effect of vancomycin on intestinal flora of patients who previously received antimicrobial therapy. Clin Infect Dis 25:729-32.
49. Enfors Sven-Olof, L. H. 2000. Bioprecess Technology and Applications. Royal Institute of Technology, Stockholm.
50. Esrey, S. A. 1996. Water, waste, and well-being: A multicountry study. American J Epidemiol 143:608-623.
51. Esteve, C., M. C. Gutierrez, and A. Ventosa. 1995. Aeromonas encheleia Sp-Nov, Isolated from European Eels. Int J Sys Bacteriol 45:462-466.
52. Farmer III J.J., F.W. Hickman-Brenner, G.R. Fanning, M.J. Arduino, and D.J. Brenner.1986. Analysis of Aeromonas and Plesiomonal by DNA-DNA hybridization and phenotype. Abstr. P1; !st Int Workshop on Aeromonas and Plesiomonas, Manchester.
53. Ford and Mac Kenzie, 2000. How safe is our drinking water ?.Postgraduate Medicine Journal.108:http://www.postgradmed.com/issues/2000/09_00/sep15.htm.
54. Fricker and Tompsett, 1989. Aeromonas spp. in foods: a significant cause of food poisoning. J. Food Microbiol. 9 .17–23.
55. Gal, M., G. M. Preston, R. C. Massey, A. J. Spiers, and P. B. Rainey. 2003. Genes encoding a cellulosic polymer contribute toward the ecological success of Pseudomonas fluorescens SBW25 on plant surfaces. Mol Ecol 12:3109-21.
56. Gentry, M. K., and J. M.Dalrymple. 1980. Quantative microtiter cytotoxicity assay for Shigella toxin. J. Clin. Microbiol. 12:361-366.
57. Ghenghesh, K. S., S. S. Abeid, M. M. Jaber, and S. A. Ben-Taher. 1999. Isolation and haemolytic activity of Aeromonas species from domestic dogs and cats. Comp Immunol Microbiol Infect Dis 22:175-9.
58. Giamarellou, H., and A. Antoniadou. 1996. Epidemiology, diagnosis, and therapy of fungal infections in surgery. Infect Control Hosp Epidemiol 17:558-64.
59. Gleick, P. H. 2000. The world's water 2000-2001, 315p. In Resources (ed.) Island Press, Washington, DC, USA.
60. Gonzalez, C., C. Gutierrez, and T. Grande. 1987. Bacterial flora in bottled uncarbonated mineral drinking water. Can. J. Microbiol. 33:1120-1125.
61. Grau, P. 1994. What's next. Water qual Intl 4:29-32. 62. Greub, G., B. La Scola, and D. Raoult. 2004. Amoebae-resisting bacteria isolated from
human nasal swabs by amoebal coculture. Emerg Infect Dis 10:470-7. 63. Greub, G., and D. Raoult. 2004. Microorganisms resistant to free-living amoebae. Clin
Microbiol Rev 17:413-33. 64. Grunditz, C. 1999. Thesis. Bioassays for the determination of nitrification inhibition. KTH,
Stockholm. 65. Grundmann, H., A. Hahn, B. Ehrenstein, K. Geiger, H. Just, and F. D. Daschner. 1999.
Detection of cross-transmission of multiresistant Gram-negative bacilli and Staphylococcus aureus in adult intensive care units by routine typing of clinical isolates. Clin Microbiol Infect 5:355-363.
66. Gunilla, K., and R. Mölby. 1979. Adhesion of Escherichia coli to human periurethral cells correlated to mannose-resistant agglutination of human erythrocytes. FEMS Microbiol. lett. 5:295-299.
67. Guyot, A., S. P. Barrett, E. J. Threlfall, M. D. Hampton, and T. Cheasty. 1999. Molecular epidemiology of multi-resistant Escherichia coli. J Hosp Infect 43:39-48.
68. Haas, C.N., 2001. Drinking Water Outbreaks.http//water.sesep.drexel.edu/outbreaks/ 69. Hanberger, H., M. Hoffmann, S. Lindgren, and L. E. Nilsson. 1997. High incidence of
antibiotic resistance among bacteria in 4 intensive care units at a university hospital in Sweden. Scand J Infect Dis 29:607-14.
70. Hanberger, H., and L. E. Nilsson. 1997. High frequency of antibiotic resistance among Gram-negative isolates in intensive care units at 10 Swedish hospitals. Clin Microbiol Infect 3:208-215.
60

71. Hanberger, H., and L. E. Nilsson. 1997. A multicenter study shows alarming results. High antibiotic resistance within Swedish intensive care Lakartidningen 94:2381-2, 2385-6.
72. Hanninen, M. L., S. Salmi, L. Mattila, R. Taipalinen, and A. Siitonen. 1995. Association of Aeromonas spp. with travellers' diarrhoea in Finland. J Med Microbiol 42:26-31.
73. Haque, Q. M., A. Sugiyama, Y. Iwade, Y. Midorikawa, and T. Yamauchi. 1996. Diarrheal and environmental isolates of Aeromonas spp. produce a toxin similar to Shiga-like toxin 1. Curr Microbiol 32:239-45.
74. Heuzenroeder, M. W., C. Y. Wong, and R. L. Flower. 1999. Distribution of two hemolytic toxin genes in clinical and environmental isolates of Aeromonas spp: correlation with virulence in a suckling mouse model. FEMS Microbiol Lett 174:131-6.
75. Hickman-Brenner, F. W., G. R. Fanning, M. J. Arduino, D. J. Brenner, and J. J. Farmer, 3rd. 1988. Aeromonas group 501, a new mannitol negative species found in human clinical specimens, Abstr. P-20, 2nd Int. Workshop on Aeromonas and Plesiomonas, Miami Beach, Florida.
76. Hickman-Brenner, F. W., G. R. Fanning, M. J. Arduino, D. J. Brenner, and J. J. Farmer, 3rd. 1988. Aeromonas schubertii, a new mannitol-negative species found in human clinical specimens. J Clin Microbiol 26:1561-4.
77. Hickman-Brenner, F. W., K. L. MacDonald, A. G. Steigerwalt, G. R. Fanning, D. J. Brenner, and J. J. Farmer, 3rd. 1987. Aeromonas veronii, a new ornithine decarboxylase-positive species that may cause diarrhea. J Clin Microbiol 25:900-6.
78. Hill, K. R., F. H. Caselitz, and L. M. Moody. 1954. A case of acute, metastatic, myositis caused by a new organism of the family: Pseudomonadaceae; a preliminary report. West Indian Med J 3:9-11.
79. Homsi, J. 2000. The present state of sewage treatment. International report. Water Supply 18:325-327.
80. Homsi, Z. J., and R. S.C. 1988. The use of duckweed for wastewater treatment. J. WPCF 60:1253-1258.
81. Horan, N. J. 1990. Biological waste water treatment systems, Theory and operation. John Wiley & Sons, Chichester; England.
82. Houslay, M. D., and G. Milligan. 1997. Tailoring cAMP-signalling responses through isoform multiplicity. Trends Biochem Sci 22:217-24.
83. Huys, G. 1996. Thesis. Classification and Identification of mesophilic Aeromonas. Gent Universiteit.
84. Huys, G., R. Coopman, P. Janssen, and K. Kersters. 1996. High resolution genotypic analysis of the genus Aeromonas by AFLP fingerprinting. Int. J. Syst. Bacteriol 46:572-580.
85. Huys, G., P. Kampfer, M. J. Albert, I. Kuhn, R. Denys, and J. Swings. 2002. Aeromonas hydrophila subsp. dhakensis subsp. nov., isolated from children with diarrhoea in Bangladesh, and extended description of Aeromonas hydrophila subsp. hydrophila (Chester 1901) Stanier 1943 (approved lists 1980). Int J Syst Evol Microbiol 52:705-12.
86. Huys, G., M. Avncanneyt, R. Coopman, P. Jansson, E. Falsen, M. Altwegg, and K. Kersters. 1994. Cellulor fatty acid, Composition as a chemotaxonomic marker for the identification of phenospecies and the hybridization groups in the genus Aeromonas. Int. J. Syst. Bacteriol. 44:651-658.
87. Huys, G., and J. Swings. 1999. Evaluation of a fluorescent amplified fragment length polymorphism (FAFLP) methodology for the genotypic discrimination of Aeromonas taxa. FEMS Microbiol. Lett. 177:83-92.
88. Islam, M. S. 1990. Increased toxin production by Vibrio cholerae O1 during survival with a green alga, Rhizoclonium fontanum, in an artificial aquatic environment. Microbiol Immunol 34:557-63.
89. Islam MS, Drasar BS; Bradley DJ.1990. Survival of toxigenic Vibrio cholerae O1 with a common duckweed, Lemna minor, in artificial aquatic ecosystems. Trans R Soc Trop Med Hyg. 1990 May-Jun;84(3):422-4.
90. Islam, M. S., M. J. Alam, and S. Tzipori. 1992. Abundance of Aeromonas in various components of pond ecosystems in Dhaka; Bangladesh. Int J. Env Studies 39:297-304.
91. Islam, M. S., B. S. Drasar, and D. J. Bradley. 1989. Attachment of toxigenic Vibrio cholerae 01 to various freshwater plants and survival with a filamentous green alga, Rhizoclonium fontanum. J Trop Med Hyg 92:396-401.
92. Islam, M. S., B. S. Drasar, and R. B. Sack. 1994. The aquatic flora and fauna as reservoirs of Vibrio cholerae: a review. J Diarrhoeal Dis Res 12:87-96.
61

93. Janda, J. M. 1991. Recent advances in the study of the taxonomy, pathogenicity, and infectious syndromes associated with the genus Aeromonas. Clin. Microbiol. Rev. 4:397-410.
94. Janda, J. M., and S. L. Abbott. 1998. Evolving Concepts Regarding the Genus Aeromonas: An Expanding Panorama of Species, Disease Presentations, and Unanswered Questions. Clin. Infect. Dis. 27:332-344.
95. Janda, J. M., S. L. Abbott, S. Khashe, G. H. Kellogg, and T. Shimada. 1996. Further studies on biochemical characteristics and serologic properties of the genus Aeromonas. J Clin Microbiol 34:1930-3.
96. Janda, J. M. A. S. 1996. p. 151-173. In A. M. Austin B, Gosling PJ, Joseph S (ed.), Human Pathogen, vol. John Willey & Sons. Chichester, England.
97. Joint, I., K. Tait, M. E. Callow, J. A. Callow, D. Milton, P. Williams, and M. Camara. 2002. Cell-to-cell communication across the prokaryote-eukaryote boundary. Science 298:1207.
98. Joseph, S. W., and A. Carnahan. 1994. The isolation, identification, and systematics of themotile Aeromonas species. Ann. Rev. Fish. Dis 4:315-343.
99. Juranek, D. D. 1995. Cryptosporidiosis: Sources of infection and guidelines for prevention. Clinical Infectious Diseases 21:S57-S61.
100. Kahana, Y. 2000. Technologicla solution in support of sustainable use of water in metropolitan areas.
101. Kanclerski, K., and R. Mollby. 1987. A simple and exact two-point interpolation method for determination of haemolytic activity in microtiter plates. Acta. Pathol. Microbiol. Immunol Scand 95:175-9.
102. Karl, T. R., and K. E. Trenberth. 2003. Modern global climate change. Science 302:1719-23. 103. Kearns, A. M., R. Freeman, and N. F. Lightfoot. 1995. Nosocomial enterococci: resistance to
heat and sodium hypochlorite. J Hosp Infect 30:193-9. 104. Kehinde, A. O., R. A. Bakare, A. A. Oni, and A. O. Okesola. 2001. Childhood gastroenteritis
due to Aeromonas hydrophila in Ibadan, Nigeria. Afr J Med Med Sci 30:345-6. 105. Kelly, K. A., J. M. Koehler, and L. R. Ashdown. 1993. Spectrum of extraintestinal disease
due to Aeromonas species in tropical Queensland, Australia. Clin Infect Dis 16:574-9. 106. Kim, W. J., R. A. Weinstein, and M. K. Hayden. 1999. The changing molecular
epidemiology and establishment of endemicity of vancomycin resistance in enterococci at one hospital over a 6-year period. J Infect Dis 179:163-71.
107. Kingombe, C. I. B., G. Huys, M. Tonolla, M. J. Albert, J. Swings, R. Peduzzi, and T. Jemmi. 1999. PCR detection, characterization, and distribution of virulence genes in Aeromonas spp. Appl. Environ. Microbiol. 65:5293-302.
108. Kirke, D. F., S. Swift, M. J. Lynch, and P. Williams. 2004. The Aeromonas hydrophila LuxR homologue AhyR regulates the N-acyl homoserine lactone synthase, AhyI positively and negatively in a growth phase-dependent manner. FEMS Microbiol Lett 241:109-17.
109. Kluyver, A. J., and C.B. van Niel .1936. Prospects for a natural system of classification of bacteria. Zentrallbl.Bakteriol. Parasitenk. infektionskr. Hyg. Abt. II 94:369-403.
110. Koch, S., M. Hufnagel, and J. Huebner. 2004. Treatment and prevention of enterococcal infections--alternative and experimental approaches. Expert Opin Biol Ther 4:1519-31.
111. Korner, S., J. E. Vermaat, and S. Veenstra. 2003. The capacity of duckweed to treat wastewater: ecological considerations for a sound design. J Environ Qual 32:1583-90.
112. Krovacek, K., F. Ahmed, W. Ahne, and I. Mansson. 1987. Adhesion of Aeromonas hydrophila and Vibrioo anguillarum to fish cells and to mucus-coated glass slides. FEMS Microbiol. Lett. 42:85-89.
113. Krovacek, K., A. Faris, S. B. Baloda, M. Peterz, T. Lindberg, and I. Mnsson. 1992. Prevalence and characterization of Aeromonas spp. isolated from foods in Uppsala, Sweden. Food Microbiology 9:29-36.
114. Kuhn, I., M. J. Albert, M. Ansaruzzaman, N. A. Bhuiyan, S. A. Alabi, M. S. Islam, P. K. Neogi, G. Huys, P. Janssen, K. Kersters, and R. Mollby. 1997. Characterization of Aeromonas spp. isolated from humans with diarrhea, from healthy controls, and from surface water in Bangladesh. J Clin Microbiol 35:369-73.
115. Kuhn, I., M. J. Albert, M. Ansaruzzaman, N. A. Bhuiyan, S. A. Alabi, M. S. Islam, P. K. Neogi, G. Huys, P. Janssen, K. Kersters, and R. Mollby. 1997. Characterization of Aeromonas spp. isolated from humans with diarrhea, from healthy controls, and from surface water in Bangladesh. J. Clin. Microbiol. 35:369-73.
116. Kuhn, I., G. Allestam, G. Huys, P. Janssen, K. Kersters, K. Krovacek, and T. X. Stenstrom. 1997. Diversity, persistence and virulence of Aeromonas strains isolated from drinking water distribution systems in Sweden. App. Env. Microbiol 63:2708-2715.
62

117. Kuhn, I., A. Iversen, and R. Mollby. 2003. The PhenePlate system for studies of the diversity of enterococcal populations from the food chain and the environment. Int J Food Microbiol 88:189-96.
118. Lammerding, A. M., and A. Fazil. 2000. Hazard identification and exposure assessment for microbial food safety risk assessment. Int J Food Microbiol 58:147-57.
119. Lawrence, C., M. Reyrolle, S. Dubrou, F. Forey, B. Decludt, C. Goulvestre, P. Matsiota-Bernard, J. Etienne, and C. Nauciel. 1999. Single clonal origin of a high proportion of Legionella pneumophila serogroup 1 isolates from patients and the environment in the area of Paris, France, over a 10-year period. J Clin Microbiol 37:2652-2655.
120. Lengeler, K. B., R. C. Davidson, C. D'Souza, T. Harashima, W. C. Shen, P. Wang, X. Pan, M. Waugh, and J. Heitman. 2000. Signal transduction cascades regulating fungal development and virulence. Microbiol Mol Biol Rev 64:746-85.
121. Lipsitch, M., and E. R. Moxon. 1997. Virulence and transmissibility of pathogens: what is the relationship? Trends Microbiol 5:31-7.
122. Livornese, L. L., Jr., S. Dias, C. Samel, B. Romanowski, S. Taylor, P. May, P. Pitsakis, G. Woods, D. Kaye, M. E. Levison, and et al. 1992. Hospital-acquired infection with vancomycin-resistant Enterococcus faecium transmitted by electronic thermometers. Ann Intern Med 117:112-6.
123. Lynch, M. J., S. Swift, D. F. Kirke, C. W. Keevil, C. E. Dodd, and P. Williams. 2002. The regulation of biofilm development by quorum sensing in Aeromonas hydrophila. Environ Microbiol 4:18-28.
124. Marks, H. M., M. E. Coleman, C. T. Lin, and T. Roberts. 1998. Topics in microbial risk assessment: dynamic flow tree process. Risk Anal 18:309-28.
125. Marolda, C. L., B. Hauroder, M. A. John, R. Michel, and M. A. Valvano. 1999. Intracellular survival and saprophytic growth of isolates from the Burkholderia cepacia complex in free-living amoebae. Microbiology 145 ( Pt 7):1509-17.
126. Martin, S. 1995. Health (Pathogen) considerations regarding the use of human waste in aquaculture. Swiss federal Institute for Environmental Science & Technology.
127. Martinez-Murcia, A. J., S. Benlloch, and M. D. Collins. 1992. Phylogenetic interrelationships of members of the genera Aeromonas and Plesiomonas as determined by 16S ribosomal DNA sequencing: lack of congruence with results of DNA-DNA hybridizations. Int J Syst Bacteriol 42:412-21.
128. Martinez-Murcia, A. J., N. M. Harland, and M. D. Collins. 1993. Phylogenetic analysis of some leuconostocs and related organisms as determined from large-subunit rRNA gene sequences: assessment of congruence of small- and large-subunit rRNA derived trees. J Appl Bacteriol 74:532-41.
129. Martins, L. M., R. F. Marquez, and T. Yano. 2002. Incidence of toxic Aeromonas isolated from food and human infection. FEMS Immunol Med Microbiol 32:237-42.
130. Matsunaga and Okochi. 1998. T. Matsunaga and M. Okochi, Bacterial pathogens in drinking water. In: R.A. Meyers, Editor, Encyclopedia of Environmental Analysis and Remediation vol. 1, Wiley, New York (1998), pp. 607–621.
131. McGarey, D. J., L. Milanesi, D. P. Foley, B. J. Reyes, L. C. Frye, and D. V. Lim. 1991. The role of motile Aeromonads in the fish disease, ulcerative disease syndrome (UDS). Experientia,Rev. May:441-4.
132. McMahon, M. A. S., and I. G. Wilson. 2001. The occurrence of enteric pathogens and Aeromonas species in organic vegetables. International Journal of Food Microbiology 70:155-162.
133. Meng, J., and M. P. Doyle. 1997. Emerging issues in microbiological food safety. Annual Review of Nutrition 17:255-275.
134. Messi, P., E. Guerrieri, and M. Bondi. 2002. Survival of an Aeromonas hydrophila in an artificial mineral water microcosm. Water Research 36:3410-3415.
135. Millership. S.E.1996. Identification. In The genus Aeromonas, Eds. B Audtin, M. Altwegg, P.J. Gosling and S. joseph, pp. 85-107. Wiley and Sons, Chichester.
136. Monfort, P., and B. Baleux. 1990. Dynamics of Aeromonas hydrophila, Aeromonas sobria, and Aeromonas caviae in a sewage treatment pond. Appl. Environ. Microbiol. 56:1999-2006.
137. Monge, R., and M. Chinchilla. 1996. Presence of Cryptosporidium oocysts in fresh vegetables. Journal of Food Protection 59:202-203.
138. Monteil and C. H Monteil. 1997. Aeromonas infections. Presse Med. 26:1790–1797.
63

139. Morgan, D. R., P. C. Johnson, H. L. DuPont, T. K. Satterwhite, and L. V. Wood. 1985. Lack of correlation between known virulence properties of Aeromonas hydrophila and enteropathogenicity for humans. Infect Immun 50:62-5.
140. Munoz, P., V. Fernandez-Baca, T. Pelaez, R. Sanchez, M. Rodriguez-Creixems, and E. Bouza. 1994. Aeromonas peritonitis. Clin Infect Dis 18:32-7.
141. National Committee for Clinical Laboratory Standards. Approved standard M7-A5, v., no 2. National Committee for Clinical Laboratory Standards, Wayne, Pa. 2000. Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically.
142. Nicolella, C., M. C. van Loosdrecht, and J. J. Heijnen. 2000. Wastewater treatment with particulate biofilm reactors. J Biotechnol 80:1-33.
143. Obi, C. L., and P. O. Bessong. 2002. Diarrhoeagenic bacterial pathogens in HIV-positive patients with diarrhoea in rural communities of Limpopo Province, South Africa. J Health Popul Nutr 20:230-4.
144. Ormen, O., P. E. Granum, J. Lassen, and M. J. Figueras. 2005. Lack of agreement between biochemical and genetic identification of Aeromonas spp. Apmis 113:203-7.
145. Osterhout, G. J., V. H. Shull, and J. D. Dick. 1991. Identification of clinical isolates of gram-negative nonfermentative bacteria by an automated cellular fatty acid identification system. J Clin Microbiol 29:1822-30.
146. P.J, G. 1996. Aeromonas speciees in diseases of animal, p. 175-95. In M. A. B. Austin, P.j gosling And S. Joseph (ed.), The genus Aeromonas. Wiley and Sons, Chichester.
147. Parsek, M. R., and C. Fuqua. 2004. Biofilms 2003: emerging themes and challenges in studies of surface-associated microbial life. J Bacteriol 186:4427-40.
148. Parveen, S., M. S. Islam, and A. Huq. 1995. Abundance of Aeromonas spp. in river and lake waters in and around Dhaka, Bangladesh. J Diarrhoeal Dis Res 13:183-6.
149. Pazzaglia, G., R. B. Sack, E. Salazar, A. Yi, E. Chea, R. Leon-Barua, C. E. Guerrero, and J. Palomino. 1991. High frequency of coinfecting enteropathogens in Aeromonas-associated diarrhea of hospitalized Peruvian infants. J Clin Microbiol 29:1151-6.
150. Pemberton, J. M., S. P. Kidd, and R. Schmidt. 1997. Secreted enzymes of Aeromonas. FEMS Microbiol Lett 152:1-10.
151. Perez, M. J., L. A. Rodriguez, A. Fernandez-Briera, and T. P. Nieto. 2002. A 45-kDa acetylcholinesterase protoxin of Aeromonas hydrophila: purification and immunogenicity in fish. FEMS Microbiol Lett 211:23-7.
152. Popoff, M.Y. 1984.Genus III, Aeromonas Klyuver and van Neil 1936. In Bergey's manual of Systematic Bacteriology 1st ed., Eds N.R. Kreig and J.G. Holt, pp.545-548. Williams & Wilkins, Baltimore.
153. Popoff, M., and M. Veron. 1976. A taxonomic study of the Aeromonas hydrophila-Aeromonas punctata group. J Gen Microbiol 94:11-22.
154. Quevedo-Sarmiento, J., A. Ramos-Cormenzana, and J. Gonzalez-Lopez. 1986. Isolation and characterization of aerobic heterotrophic bacteria from natural spring waters in the Lanjaron area (Spain). J Appl Bacteriol 61:365-372.
155. Rahman, M., P. Colque-Navarro, I. Kuhn, G. Huys, J. Swings, and R. Mollby. 2002. Identification and characterization of pathogenic Aeromonas veronii biovar sobria associated with epizootic ulcerative syndrome in fish in Bangladesh. Appl Environ Microbiol 68:650-5.
156. Rahman, M. H., and K. Kawai. 2000. Outer membrane proteins of Aeromonas hydrophila induce protective immunity in goldfish. Fish Shellfish Immunol 10:379-82.
157. Romling, U. 2001. Genetic and phenotypic analysis of multicellular behavior in Salmonella typhimurium. Methods Enzymol 336:48-59.
158. Romling, U. 2002. Molecular biology of cellulose production in bacteria. Res Microbiol 153:205-12.
159. Rosner, R. 1964. Aeromonas hydrophila as the Etiologic Agent in a Case of Severe Gastroenteritis. Am J Clin Pathol 42:402-4.
160. Ross, P., R. Mayer, and M. Benziman. 1991. Cellulose biosynthesis and function in bacteria. Microbiol Rev 55:35-58.
161. Ruimy, R., V. Breittmayer, P. Elbaze, B. Lafay, O. Boussemart, M. Gauthier, and R. Christen. 1994. Phylogenetic analysis and assessment of the genera Vibrio, Photobacterium, Aeromonas, and Plesiomonas deduced from small-subunit rRNA sequences. Int J Syst Bacteriol 44:416-26.
162. Sadikot, R. T., T. S. Blackwell, J. W. Christman, and A. S. Prince. 2005. Pathogen-host interactions in Pseudomonas aeruginosa pneumonia. Am J Respir Crit Care Med 171:1209-23.
64

163. Sanarelli, G. 1891.Uber einen neuen Mikroorganisums des Wassers, welcher fur thiere mit veränderlicher und konstanter Temperature pathogen ist. Zentralbl. Bakteriol. Parasitenk. 9:222-228.
164. Saran, S., M. E. Meima, E. Alvarez-Curto, K. E. Weening, D. E. Rozen, and P. Schaap. 2002. cAMP signaling in Dictyostelium. Complexity of cAMP synthesis, degradation and detection. J Muscle Res Cell Motil 23:793-802.
165. Schlichting, C., C. Branger, J. M. Fournier, W. Witte, A. Boutonnier, C. Wolz, P. Goullet, and G. Doring. 1993. Typing of Staphylococcus aureus by pulsed-field gel electrophoresis, zymotyping, capsular typing, and phage typing: resolution of clonal relationships. Journal Of Clinical Microbiology 31:227-232.
166. Schubert, R. H. W., and M. Hegazi. 1988. Aeromonas Eucrenophila Species Nova Aeromonas-Caviae a Later and Illegitimate Synonym of Aeromonas-Punctata. Zentralblatt Fur Bakteriologie Mikrobiologie Und Hygiene Series a-Medical Microbiology Infectious Diseases Virology Parasitology 268:34-39.
167. Sghir, A., G. Gramet, A. Suau, V. Rochet, P. Pochart, and J. Dore. 2000. Quantification of bacterial groups within human fecal flora by oligonucleotide probe hybridization. Appl Environ Microbiol 66:2263-6.
168. Shotts, E. B., Jr. 1987. Bacterial diseases of fish associated with human health. Vet Clin North Am Small Anim Pract 17:241-7.
169. Shotts, E. B., Jr., J. L. Gaines, Jr., L. Martin, and A. K. Prestwood. 1972. Aeromonas-induced deaths among fish and reptiles in an eutrophic inland lake. J Am Vet Med Assoc 161:603-7.
170. Siddiqui, M. N., I. Ahmed, B. J. Farooqi, and M. Ahmed. 1992. Myonecrosis due to Aeromonas hydrophila following insertion of an intravenous cannula: case report and review. Clin Infect Dis 14:619-20.
171. Simm, R., M. Morr, A. Kader, M. Nimtz, and U. Romling. 2004. GGDEF and EAL domains inversely regulate cyclic di-GMP levels and transition from sessility to motility. Mol Microbiol 53:1123-34.
172. Smit, J., and N. J. 1992. Urban agriculture for sustainable cities : using wastes and idle land and water bodies as resources. Environment and Urbanization 4:141-152.
173. Sneath, P. H. A., and R. R. Sokal. 1973. Numerical taxonomy. W. H. Freeman & Co., New York.
174. Snieszko, S. F., S. B. Friddle, and P. J. Griffin. 1951. Successful treatment of ulcer disease in brook trout (Salvelinus fontinalis) with terramycin. Science 113:717-8.
175. Snieszko, S. F., E. M. Wood, and W. T. Yasutake. 1957. Infectious pancreatic necrosis in trout. AMA Arch Pathol 63:229-33.
176. Snieszo. 1957. Huys. Genus IV, Aeromonas Klyuver and van Neil 1936. In Bergey's Manual of Determinative Bacteriology 7th ed., R.S. Breed, E.G.D. murray, and N.R. Smith, p.189. Williams & Wilkins, Baltimore.
177. Subasinghe, R. P. 2005. Epidemiological approach to aquatic animal health management: opportunities and challenges for developing countries to increase aquatic production through aquaculture. Prev Vet Med 67:117-24.
178. Swift, S., N. J. Bainton, and M. K. Winson. 1994. Gram-negative bacterial communication by N-acyl homoserine lactones: a universal language? Trends Microbiol 2:193-8.
179. Swift, S., A. V. Karlyshev, L. Fish, E. L. Durant, M. K. Winson, S. R. Chhabra, P. Williams, S. Macintyre, and G. S. Stewart. 1997. Quorum sensing in Aeromonas hydrophila and Aeromonas salmonicida: identification of the LuxRI homologs AhyRI and AsaRI and their cognate N-acylhomoserine lactone signal molecules. J Bacteriol 179:5271-81.
180. Swift, S., J. P. Throup, P. Williams, G. P. Salmond, and G. S. Stewart. 1996. Quorum sensing: a population-density component in the determination of bacterial phenotype. Trends Biochem Sci 21:214-9.
181. Talon, D., M. J. Dupont, J. Lesne, M. Thouverez, and Y. Michel-Briand. 1996. Pulsed-field gel electrophoresis as an epidemiological tool for clonal identification of Aeromonas hydrophila. J Appl Bacteriol 80:277-82.
182. Tenover, F. C., R. D. Arbeit, R. V. Goering, P. A. Mickelsen, B. E. Murray, D. H. Persing, and B. Swaminathan. 1995. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol 33:2233-9.
183. Tezcan-Merdol, D., M. Ljungstrom, J. Winiecka-Krusnell, E. Linder, L. Engstrand, and M. Rhen. 2004. Uptake and replication of Salmonella enterica in Acanthamoeba rhysodes. Appl Environ Microbiol 70:3706-14.
65

184. Torell, E., O. Cars, and A. Hambraeus. 2001. Ampicillin-resistant enterococci in a Swedish university hospital: nosocomial spread and risk factors for infection. Scand J Infect Dis 33:182-7.
185. Torell, E., O. Cars, B. Olsson-Liljequist, B. M. Hoffman, J. Lindback, and L. G. Burman. 1999. Near absence of vancomycin-resistant enterococci but high carriage rates of quinolone-resistant ampicillin-resistant enterococci among hospitalized patients and nonhospitalized individuals in Sweden. J Clin Microbiol 37:3509-13.
186. Torell, E., I. Kuhn, B. Olsson-Liljequist, S. Haeggman, B. M. Hoffman, C. Lindahl, and L. G. Burman. 2003. Clonality among ampicillin-resistant Enterococcus faecium isolates in Sweden and relationship with ciprofloxacin resistance. Clin Microbiol Infect 9:1011-9.
187. Trick, W. E., M. J. Kuehnert, S. B. Quirk, M. J. Arduino, S. M. Aguero, L. A. Carson, B. C. Hill, S. N. Banerjee, and W. R. Jarvis. 1999. Regional dissemination of vancomycin-resistant enterococci resulting from interfacility transfer of colonized patients. J Infect Dis 180:391-6.
188. Tsai, G.-J., and S.-C. Yu. 1997. Microbiological evaluation of bottled uncarbonated mineral water in Taiwan. Int J Food Microbiol.37:137-143.
189. Turutoglu, H., S. Ercelik, and M. Corlu. 2005. Aeromonas hydrophila-associated skin lesions and septicaemia in a Nile crocodile (Crocodylus niloticus). J S Afr Vet Assoc 76:40-2.
190. Wegener, H. C., F. M. Aarestrup, L. B. Jensen, A. M. Hammerum, and F. Bager. 1999. Use of antimicrobial growth promoters in food animals and Enterococcus faecium resistance to therapeutic antimicrobial drugs in Europe. Emerg Infect Dis 5:329-35.
191. Wendt, C., B. Wiesenthal, E. Dietz, and H. Ruden. 1998. Survival of vancomycin-resistant and vancomycin-susceptible enterococci on dry surfaces. J Clin Microbiol 36:3734-6.
192. Veron, M. 1965. The taxonomic position of Vibrio and certain comparable bacteria. C R Acad Sci Hebd Seances Acad Sci D 261:5243-6.
193. Whitby, P. W., M. Landon, and G. Coleman. 1992. The cloning and nucleotide sequence of the serine protease gene (aspA) of Aeromonas salmonicida ssp. salmonicida. FEMS Microbiol Lett 78:65-71.
194. WHO. 1996. Water and sanitation fact sheet. N112. http://www.who.int/inf-fs/en/fact112.html.
195. WHO. 1997. Surveillance and Control of Community Supplies, Geneva. Geneva vol. 3. 196. Williams, W. S., R. E. Carron. 1989. Alternative environmental roles for cellulose produced
by Acetobacter xylinum. Appl Environ Microbiol 55. 197. Wood, E. M., S. F. Snieszko, and W. T. Yasutake. 1955. Infectious pancreatic necrosis in
brook trout. AMA Arch Pathol 60:26-8. 198. Vugia, D. J., A. M. Shefer, J. Douglas, K. D. Greene, R. G. Bryant, and S. B. Werner. 1997.
Cholera from raw seaweed transported from the Philippines to California. J Clin Microbiol 35:284-5.
66