Embed Size (px)
Citation preview

FOUNDATIONS FOR A LONG TERM DRACAENA BREEDING PROGRAM:
FLOWER INDUCTION, IRRADIATION, AND POLYPLOIDIZATION
A THESIS SUBMI I I ED TO THE GRADUATE DIVISION OF THE UNIVERSITY OF HAW AI'I IN PARTIAL FULFILLMENT OF THE
REQUIREMENTS FOR THE DEGREE OF
MASTER OF SCIENCE
IN
TROPICAL PLANT AND SOIL SCIENCES
MAY 2007
By
Emily Shih-wen Teng
Thesis Committee:
Kenneth W. Leonhardt, Chairperson Richard A. Criley
John L. Griffis

We certify that we have read this thesis and that, in our opinion, it is satisfactory
in scope and quality as a thesis for the degree of Master of Science in Tropical
Plant and Soil Sciences.
THESIS COMMITTEE
~rI/It to ~QAjJ--Chairperson
~/47
ii

ACKNOWLEDGEMENTS
I would like to express my appreciation to my committee chairperson, Dr. Ken
Leonhardt, for his guidance during my time at UH. His knowledge, advice, and support
have been very helpful over the course of my research project.
I would also like to give thanks to Dr. Richard Criley and Dr. John Griffis for
their help and for serving on my thesis committee.
Associates on Maui and the Big Island have provided much assistance on this
project which is much appreciated. Joanne Lichty and Patty Nakao collaborated on many
of the experiments including several of the flower induction trials and Andrew Kawabata
has provided support in many ways.
The following people and organizations have donated plant materials and/or use
of their facilities for my research experiments and their generosity is gratefully
acknowledged. Ray Fukunaga, Ray Baker and Lyon Arboretum, Bill Durston and
Leilani Nursery, Jake Henny, California and Hawaii Foliage Growers. A & K Nursery,
Sandy Baehr from Alpha Genesis Laboratories, Andrew Gruttadauro from Adauro
Builders, Stuart Stein, Dean Shimonishi, and Michael Swiderski at the USDA Irradiation
Facility in Waimanalo, and Hawaiian Sunshine Nursery.
Thanks to the staff at Magoon Research Facility, especially Craig Okazaki and
Ronald Matsuda, for all their assistance. The assistance from Roger Corrales and his
staff at the Waimanalo Experimental Station is also much appreciated.
Many other faculty and staff members have been helpful in many aspects of my
research and education. Dr. Kheng-tuan Cheah's advice and knowledge in tissue culture
has been extremely helpful to my project. Thanks to Dr. Yoneo Sagawa for his assistance
iii

and use of his laboratory. Dr. Mark Wright provided excellent knowledge and advice in
statistical analysis. Dr. Kent Kobayashi was generous in loaning temperature recording
devices. Dr. Karen Selph's assistance and time during flow cytometric analysis is also
greatly appreciated. In addition, Susan Takahashi, Shirley Ishihara, Lynn Horiuchi, and
Elsie Sun in the TPSS Department office have provided much appreciated assistance over
the years.
Funding for my graduate assistantship and research supplies was provided in part
by T -STAR Project No. HA W00839-1 017S.
I would also like to thank my labmates Xuebo Shi, Thomas Littleton and
especially Susana Vanzie-Canton for their help, advice, and friendship. Thanks to Craig
Koyanagi, Kekaha Spencer, Natalie Pullano, Alison Render, and Jackie Chan-Halbrendt,
the student employees in our lab and Magoon for their assistance. And thanks to my
fellow graduate student Peter Toves for his help and friendship.
Finally, I would like to express my thanks to my husband Mark for supporting our
move to Hawaii in order for me to attend graduate school. He has also provided me
assistance on my project and emotional support and encouragement during my graduate
education. Thanks to my family - my parents and brother - for their support and
encouragement.
iv

ABSTRACT
Dracaenas are important foliage plants not only in the United States, but also
worldwide, and new varieties are needed to maintain consumer demand. Flower
induction, mutation induction with gamma rays, and polyploidization using oryza1in were
attempted on various dracaenas to lay down the foundations for a long-term breeding
program to create new varieties for the foliage industry.
Using traditional breeding methods for breeding requires the availability of
flowers on plants intended for breeding. Two methods were employed to attempt flower
induction - Gibberellic acid (GA3) applications and cold temperature treatments.
GA3 solutions ranging in concentration from 0 to 6000 ppm were applied to nine
different varieties of Dracaena to attempt out of season flower induction. Dracaena
fragrans 'Massangeana' flowered at the rate of 8% in the 0, 500, and 1000 ppm
treatments and 16% in the 2000 ppm treatment. Dracaena xmasseffiana flowered at rates
of 0%, 41.67%,58.33% and 62.5% in the 0,500,1000, and 2000 ppm treatments,
respectively. None of the other treated plants flowered during the experiments.
Three cold temperature experiments using temperatures ranging from 8 to 20°C
and treatment durations from 3 to 40 days were performed on seven different Dracaena
varieties. D. xmasseffiana plants chilled for 0, 3, 6, or 9 days at 12°C flowered at the
rate of 0%, 16.9%,39.2%, and 86.7% of stems, respectively. None of the other plants
treated had significant flowering percentages.
Mutation induction using irradiation is a useful method for creating new varieties
of ornamental plants that are vegetatively propagated. Unrooted cuttings of four
Dracaena varieties were irradiated using Cesium-137 gamma rays at dosages ranging v

from 0 to 500 Grays (Gy) in the first round and 0 to 50 Gy in the second round in order to
estimate the LDso dosages for root and shoot formation. A general trend of decreased
root and shot formation with increased radiation dosage was observed. For root
formation, the LDso dosages were estimated as 14.6,13.8,5.7, and 17.7 Gy and for shoot
formation, the dosages were estimated as 19.4,16.6,22.1, and 10.9 Gy for D. deremensis
'Santa Rosa', D. fragrans 'Massangeana', D.fragrans 'Victoriae', and D. xmasseffiana,
respectively. Visually detectable mutations for each treatment were observed at rates of
0% to 93.3% with chlorophyll mutations being the most common.
Polyploid forms of plants often have horticulturally desirable characteristics such
as more compact growth habit, thicker and more robust leaves, and a deeper green color.
Two methods of polyploidization on Dracaena were attempted. Both methods employed
six treatments consisting of three concentrations of oryzalin ranging (0%, 0.05% and
0.01 %) and two durations of treatment (24 and 48 h). The first method treated
developing axillary buds of D. deremensis 'Santa Rosa' in vivo by placing oryzalin
soaked cotton on the meristem and covering the meristem in plastic. The second method
treated call us tissue of D. deremensis 'Lisa' in vitro by soaking the calli in oryzalin
solution. The developed shoots from the axillary buds and the regenerated shoots from
the callus tissue were tested for conversion to polyploidy using flow cytometry with leaf
tissue nuclei. In vivo treatments resulted in only one mixoploid. In vitro treatments
resulted in one mixoploid and one tetraploid plant. The tetraploid has shorter internodes
and shorter leaves than its diploid counterpart and is being further evaluated for
suitability as a new variety or for use in hybridizing efforts.
vi

TABLE OF CONTENTS
ACKN"OWLEDGEMENTS ............................................................................................ m ABSTRACT ...................................................................................................................... V
LIST OF T ABLES .......................................................................................................... IX
LIST OF FIGURES ...•....•................................................................................................ X
CHAPTER 1. INTRODUCTION ................................................................................. 11
CHAPTER 2. LITERATURE REVIEW ..................................................................... 12
2.1. DRACAENA TAXONOMY, MORPHOLOGY, AND PROPAGATION METHODS ......•.••....• 12 2.2. FLOWER INDUCTION USING GIBBERELLIC ACID ••.••••.••••.•••••.••••.•.•...•.•.....••....•..•....• 14
2.3. FLOWER INDUCTION USING COLD TEMPERATURES •...•....•.•....•........•.•...•..•....•.••....• 15 2.4. IRRADIATION .......................................................................................................... 17 2.5. POLYPLOID INDUCTION .............•..•.••.••.•••••••••••••••.•••...•..................................•......•• 22 2.6. FLOW CYTOMETRY ••.•...•............•..............••.•..•••••••••••••••••••••••••••••••••••••.••.••...•••.....•• 26
CHAPTER 3. ORJECfIVES ........................................................................................ 29
CHAPTER 4. FLOWER INDUCTION ....................................................................... 31
INTRODUCTION ..••..•••..•.•.........•.••..••••....••.••...•.•...•.•....•.•..•.•....................•................••••••• 31
MATERIALS AND METHODS ......................•.....•...•••..•.•••..••••.•••••••••••••••.••••.••••....••.•....••.•. 32
RESULTS ................•.••.••••.•••••..•.••..••.•....•..........•.........•.•..•.•..•.•...•.•..•.•................•.•....••.•• 35 DISCUSSION AND CONCLUSIONS ....•........•........•...•.•...•.•.............•.......................•.••..•••••• 38 TABLES AND FIGURES ..•.•..•......•............•.•.......•...•••••..••••.••••••••••••••••••••.•••••..•••..•.••.•....•.•. 45
CHAPTER 5. IRRADIATION ..................................................................................... 49
INTRODUCTION ..•....•....•....•......•.....•.....................•.....••.•..•.........•.........................•..••.•.•• 49 MATERIALS AND METHODS .•...........................................•..•.•..•••....•.•.•....•••.......•......••... 50 RESULTS ..........•.....•..•.••••.••••••••••••.••••.•••••••........••..................•••••••••••••••••••••••••.••••••••••••... 52
DISCUSSION AND CONCLUSIONS .........................................•....•.•....•........•......•.........•.... 56 TABLES AND FIGURES •••..•••....•.•.....................•.....•......•..•.•..•.•..•.•...•....•....•..•...•..•....•.••... 62
CHAPTER 6. POLypLOIDIZATION ........................................................................ 73
INTRODUCTION •••••••••.••••..•••...••..........................•.....•..•..•.•..•.•........•••..•••...•••....••••...•...... 73 MATERIALS AND METHODS .•.••.....••.....••......•.....•.....•.....•......•..••••••••••••••••••••••••••••••••••••.. 74
RESULTS ..........••........••.•..•...•..••....••.....•..•....•...........•.....••...••••..••••••.....•••••••••...•......••....• 78 DISCUSSION AND CONCLUSIONS .................................................................................... 79 TABLES AND FIGURES .•....•...••••.••••••••••••••••••••••••••••••••.••••••••••.....................•......•.......•..... 83
APPENDIX A. FLOWER INDUCTION RAW DATA .............................................. 89
APPENDIX B. FLOWER INDUCTION SIGMASTAT OUTPUTS ........................ 91
APPENDIX C. FLOWER INDUCTION LINEAR REGRESSION ......................... 94
APPENDIX D. IRRADIATION RAW DAT A ............................................................ 97
vii

APPENDIX E. IRRADIATION LINEAR REGRESSION ...................................... 101
APPENDIX F. MUTATION RATE CONTINGENCY TABLES ........................... 109
LITERA TU"RE CITED ................................................................................................ 113
viii

LIST OF TABLES
Table Page
I Setup of GA3 flower induction experiments .................................... , 45
2 Setup of cold temperature flower induction experiments -Experiment #2.... .. ......... ................... .... ..... ....................... ... 46
3 Setup of cold temperature flower induction experiments -Experiment #3....................... .......... ......................... ........ ...... 46
4 Linear regression equations, R2, and P-values for the flower induction in Dracaena using GA3 and cold temperature treatments...... ................... 46
5 Results of gamma ray irradiation treatments - Experiment #1.......... ........ 62
6 Results of gamma ray irradiation treatments - Experiment #2................ 63
7 Linear regression equations, R2, and P-values for rooting percentages in 64 second round gamma ray irradiation treatments ............................... ..
8 Linear regression equations, R2, and P-values for sprouting percentages in second round gamma ray irradiation treatments... ... ............ ............... 64
9 LDso and LDIOO values of rooting and sprouting of the four Dracaena . . d·th 137C vaneties treate WI s gamma rays ........................................ .. 64
10 Mutations observed in the four Dracaena varieties treated with 137CS
gammarays ......................................................................... .. 65
11 Results of in vivo treated D. deremensis 'Santa Rosa' buds with oryzalin... 83
12 Results of in vitro treated D. deremensis 'Lisa' callus with oryzalin.......... 83
ix

LIST OF FIGURES
Figure Page
1 Flowering Response of D. xmasseffiana to GA3 treatments............... .... 47
2 Flowering Response of D.fragrans 'Massangeana' to GA3 treatments...... 47
3 Flowering Response of D. xmasseffiana to cold temperature treatments... 48
4 Rooting over time of irradiated Dracaena varieties......... .................... 66
5 Sprouting over time of irradiated Dracaena varieties... ...... .............. .... 67
6 Rooting percentages of irradiated Dracaena varieties.......................... 68
7 Sprouting percentages of irradiated Dracaena varieties..... ....... ......... ... 69
8 Number of mutations in irradiated Dracaena varieties............ ............. 70
9 D.fragrans 'Massangeana' - Normal shoots and chlorophyll mutants...... 71
10 Hormesis in D. xmasseffiana............................................... ....... 72
11 In vivo oryza1in treatment procedures of D. deremensis 'Santa Rosa' buds 84
12 In vitro oryzalin treatment procedures of D. deremensis 'Lisa' callus....... 84
13 DNA fluorescence of D. deremensis 'Santa Rosa' mixoploid obtained from in vivo oryzalin treatment........................... ........................ 85
14 DNA fluorescence of D. deremensis 'Lisa' mixoploid obtained from in vitro oryza1in treatment....................................... ...... ................ 86
15 DNA fluorescence of D. deremensis 'Lisa' tetraploid obtained from in vitro oryza1in treatment................................................ ............. 87
16 Diploid and tetraploid D. deremensis 'Lisa' in vitro plantlets................. 88
x

CHAPTERl. ~ODUCTION
Dracaenas originate from tropical areas of Africa and Asia and are popular as
houseplants and interior landscaping plants (Henny and Chen, 2003). They are known to
have been cultivated in Europe since at least the mid-1700's (Cialone, 1984). Major
production areas in the United States include Puerto Rico, Florida and Hawaii, where
they are one of the most important commercially produced genera. In Hawaii in 2005, 26
fanns sold seven million dollars worth of potted dracaena, the second largest of all
floriculture and nursery products (Hawaii Department of Agriculture, 2006). Much of
their popularity is due to the fact that they can tolerate low light levels and have few
insect and disease problems (Cialone, 1984).
Almost all of the dracaenas commercially grown as foliage plants are cultivars of
only six species. These are Dracaenafragrans, D. deremensis, D. marginata, D.
sanderiana. D. reflexa, and D. surculosa (Chen et al., 2002). Despite the economic
importance of dracaenas to the foliage industry, there is currently no known organized
breeding program to create new varieties. All releases of new cultivars to the industry
since 1975 have been due to selections of sports from cuttings by growers (Chen et al.,
2002). Yet not all of the introductions are commonly grown, and many are quite similar
in morphology and coloration. Recent interviews with foliage growers in Hawaii show
that there is a demand for new Dracaena varieties (K. Leonhardt, personal
communication). The lack of new introductions, ever changing consumer preferences,
and demand by the growers implies a good market potential for new varieties of
Dracaena.
11

CHAPTER 2. LITERATURE REVIEW
2.1. Dracaena Taxonomy, Morphology, and Propagation Methods
The genus Dracaena is a monocotyledonous group of plants that consists of
approximately 35 species of tree-like or shrub-like plants. The diploid chromosome
number of the commonly grown species is 2n = 38, 40 or 42 (Mathew and Vijayavaili,
1989). There has been debate over whether it belongs in the Liliaceae or the Agavaceae
family, but most recently, Takhtajan placed it in a separate family, Dracaenaceae (Henny
and Chen, 2003).
Kingdom: Plant
Division: Spermatophyta
Class: Angiosperm
Subclass: Monocotyledon
Order: Liliaies
Family: Dracaenaceae
Genus: Dracaena
Depending on the species, dracaenas can range from 1 m to more than 3 m tall.
Leaves of Dracaena range from lanceolate to sword shaped, and coloration ranges from
solid to variegated to speckled The growth habit of many species is a single treelike
stem with leaf rosettes at the tip of the cane.
Dracaena inflorescences occur in terminal panicles with flowers usually in
clusters. The flowers are radially symmetrical, bisexual, have six tepais, a superior
ovary, and tubular bases with free lobes (Staples and Herbst, 2005). Because
12

inflorescences are terminal, the stem is unable to continue growth and further
development is by branching from buds in the axils of leaves close to the base of the
inflorescence (Bos, 1984). According to Bos (1984), the inflorescences are branched but
the flowers are grouped in clusters or glomerules because of the reduced nature of the
branches. The exception is D. surcuiosa, in which the inflorescences are not branched
and bear a single terminal glomerule.
Propagation of dracaenas in commercial practice is mainly by cuttings. Canes are
often purchased from stock farms in Central America, South America, or the Carribean
and shipped to Florida for propagation. In Hawaii, many growers have stock plants in
their own fields for propagation. Canes are cut into various sizes ranging from one to six
feet and grown in configurations of staggering heights. For example, a twelve inch pot
may have four, three, and two foot canes, or a ten inch pot may have three. two and one
foot canes. Lateral buds at the top of each cane generally develop, producing the most
commonly seen look in nurseries around the country. Rooting hormones such as Indole-
3-Butyric acid (IBA) can be used to speed up rooting (Cialone, 1984). Canes are usually
planted directly into the final container and media. In Hawaii, a mix consisting mainly of
volcanic cinder supplemented with peat moss or coir is used while elsewhere a mix of
bark, sand and peat moss is used. Air-layering and seeds are possible propagation
methods, but have limited practice in the industry. Until recent years, tissue culture was
only used for research, but some growers are now using it to produce dracaenas for
wholesale and retail sale.
13

2.2. Flower Induction using Gibberellic Acid
One approach that has been successful in initiating floral development in other
foliage genera is the use of Gibberellic Acid (GA3) treatments (Henny, 1980, 1981, 1983,
1988, 1991; Henny et al., 1999). GA3 is a naturaIly occurring plant growth regulator that
can cause a variety of effects, including the stimulation of flowering. Gibbere11ins also
have roles in regulating nucleic acid and protein synthesis, and may also suppress root
initiation by interfering with transcription (Hartmann et al., 2001). Corr and Widmer
(1987) suggested that gibberellins may act on metabolically useful substrates available to
the apex at the time of flower initiation. The increase in the availability of these
substrates promotes flower initiation in shoots that might normally have insufficient
substrate.
Foliar sprays of GA3 have induced flowering in many ornamental aroids. In a
study on Aglaonema, an average of5.6 inflorescences were produced in GA3treated
plants, but only 0.3 in the control (Henny, 1983). InAnthurium, an average of 1.8
inflorescences were formed in GA3 treated plants verses 0.2 in untreated plants (Henny
and Hamilton, 1992). The trend is similar in Cryptocoryne, Dieffinbachia, Homalomena,
and Syngonium. In each of these studies, the number of inflorescences formed in GA3
treated plants was significantly higher than in non-treated plants, and the control plants
did not produce any inflorescences (Henny, 1980, 1988; Henny et al., 1999; Kane et al.,
1995). The induction of flowering using GA3 as a foliar spray was also studied in
Spathiphyllum using concentrations of250 to 1000 ppm. After 20 weeks, 10% of
controls, 90% of 250 ppm treated plants, and 100% of 500 and 1000 ppm treated plants
had flowered (Henny, 1981).
14

Another method of GA3 application, the tuber or rhizome soak, has also been
quite effective in inducing flowering in members of the Araceae family. Harbaugh and
Wilfret (1979) found that soaking Caladium tubers in 250 to 1000 ppm GA3 for 4 to 64 h
increased the average number of inflorescences from 0.2 in the control to 2.6 in treated
plants. The rhizome soak was more effective than the foliar spray for Zantedeschia in a
study by Corr and Widmer (1987). Spraying the foliage with 50 to 1000 ppm GA3
solutions did not cause an increase in the number of flowers, but soaking the rhizomes in
the same concentrations for 10 min increased the percentage of shoots with flowers from
47.9% in the control to an average of 69.2% in the treated plants.
Fisher (1980) conducted a study on the effects of GA3 on flowering in Cordyline
and Dracaena species. Daily applications of three drops of solution were applied on
three consecutive days to clones of Cordy line terminalis (currently c.jruticosa), D.
thalioides, D. deremensis, D. marginata, and D. reflexa. None of the dracaenas
responded to the treatments, but the Cordyline species did respond. Untreated plants did
not flower while treated plants flowered at rates ranging from 3% to 97% at different
concentrations of GA3. This study is the only known published attempt to induce
flowering using GA3 on any Dracaena species.
2.3. Flower Induction using Cold Temperatures
Cold temperature treatments have also successfully been used to induce plants to
flower. The transition from vegetative to reproductive growth is largely under genetic
control but is also influenced by environmental factors (Zeevart, 1976). Many plants
respond to environmental conditions such as temperature once they have reached the
capacity to flower (Runkle et al., 1999). Extensive studies on the cold requirements of
15

many flower crops have been carried out. Peony species produced more flowers when
plants were treated to colder temperatures and longer chilling durations. Potted plants
were placed into temperatures of I, 4, or 7°C for 3,6,9, or 12 weeks and then grown at
18°C to measure flowering production. Control plants that were not subjected to cold
temperature treatments did not produce flowers, while increased flowering was reported
for cooler temperatures and longer chilling durations (Fulton et aI., 2001).
A study on Cape Daisy (Osteospermum jucundum) also found that plants required
a cold period prior to flowering. Plants not subjected to low temperatures and plants
subjected to 1 week at a low temperature did not flower, while plants SUbjected to a 2
week period at a temperature of 12°C flowered after only 43 days (pearson et aI., 1995).
Rhododendrons are stated to require cold temperature storage below 10°C to
break flower bud dormancy (French and Alsbury, 1988). A study on the effects of two
cold storage treatments for 6 and 8 weeks was undertaken on four cultivars of
Rhododendron. The four cultivars responded differently to the treatments, but the
general finding was that plants stored at the colder temperatures of 5 °C for the longer
period of 8 weeks had a decreased time to flowering when compared to plants stored at
6.8 °C (French and Alsbury, 1988).
In a simple experiment to study the effects of chilling on Astilbe, plants were
chilled for 0,3,6,9, or 12 weeks at 5°C. The finding was that the plants chilled for 12
weeks produced the most flowers, while plants that were not chilled and plants chilled for
3 weeks did not produce any flowers (Beattie and Holcomb, 1983). Another plant that
showed similar results in studies was Chinese holly (Rex cornuta Lindl. Paxt cv.
16

Bufordii). Plants were stored at 3 to 4 °C for 0, 2, 4, 6, or 8 weeks. Those that received 6
or 8 weeks of chilling produced the greatest number of flowers and flowered in the least
number of days (Wright and Milbocker, 1979).
These studies show that many plants require cold periods prior to flowering and
that longer durations and colder temperatures have the greatest effect. The published
studies on this subject are mostly on temperate species, and no relevant papers on cold
temperature treatment effects on flowering in tropical foliage plants could be found. The
only information on inducing Dracaena flowering that could be found was a publication
in which the objective of the experiment was to regenerate inflorescences of D. jragrans
'Massangeana' from in vitro ca\lus. The article points out that flowering of Dracaena
doesn't occur naturally in the area, and plants were induced to flower by placing them in
temperatures of 12 or 15 °C for 20 or 30 days with 8 h oflight daily (Lu, 2002). Specific
results were not given, but it is mentioned that frequency of inflorescence formation
could reach 100%, and that temperatures higher than 15 °C and durations shorter than 20
days decreased the inflorescence frequency significantly.
2.4. Irradiation
Inducing mutations is a well established method to create new varieties.
According to one summary, over 1700 mutant varieties from chemical and radiation
mutation have been released (Larkin, 1998). However, it is believed that the actual
number of new varieties created from mutation is much higher than reported because
some breeders are not willing to disclose their methods, and also because many are just
not reported (Schum and Preil, 1998). Recently, the interest in mutation breeding has
decreased due to the shift to new molecular techniques (Schum, 2003). Molecular 17

techniques are highly sophisticated and require large investments in equipment and
training. The use of these expensive molecular methods on ornamental species often
cannot be justified because although ornamental crops as a whole make up a large portion
of cultivated crops, individually they are small in value compared to the widely grown
agronomic crops. Mutation breeding is therefore still a very useful method in ornamental
crops. Furthermore, desired traits are easily monitored and propagated in ornamental
species, especially those that are vegetatively propagated (Nybom, 1961).
One of the most widely used methods of mutation induction is irradiation. The
most common type is called ionizing radiation because it produces ionization when it
interacts with matter and has the ability to ionize or rupture chemical bonds (Sparrow,
1961). There are two main types of ionizing radiation. Particulate radiations are alpha,
beta, protons, and deuterons. Electromagnetic radiations include gamma rays, which will
be the focus of this study, and X-rays.
Gamma rays are produced by radioactive substances which give off the high
energy photons called gamma rays. There are many different sources of gamma rays, the
most common of which are Cobalt-60 rOCo) and Cesium-137 e37Cs). The main
difference between these two sources is their half-life. 6OCo has a halflife of 5.3 years,
while 137Cs has a half-life of33 years. In addition, 137CS is easier to shield than 6OCo
because it is less energetic. Gamma sources are advantageous because high intensities of
radiation can be reached in a restricted area near the source. In this way, higher dosages
can be applied in a shorter time than with x-ray machines, which require large amounts of
electricity, have lower maximum intensities and penetration than other sources of
18

radiation, cannot accommodate larger objects, and also cannot be operated for long
periods of time economically (Sparrow, 1961).
Mutation induction in ornarnentaI plants has been used for many purposes such as
improving or creating new flower characters including color, size, morphology and
fragrance, leaf characters such as form, size, and pigmentation, and also growth habit
(Schum and Preil, 1998). Other traits researchers attempt to improve using mutation are
flowering time and tolerance of biotic and abiotic stresses. For example, irradiation
produced improved seed viability and fertility in rice (Miyahara, 1997) and also produced
low temperature tolerant cultivars of Chrysanthemum (Broertjes et aI., 1983). Of all
these characteristics, flower color mutations are probably the most sought after. An
evaluation of the irradiation literature shows that 55% of the pUblications concern
changes in flower color (Schum, 2003). This large number of studies is probably due to
the fact that flower color is such a prominent and important feature of many ornarnentaIs.
In fact, the first official mutant released in 1936 was a tulip in which the flower color was
altered by irradiation (Schum and Preil, 1998).
Some of the most common crops in irradiation studies are chrysanthemum,
carnation, and poinsettia One study conducted in India on the chrysanthemum used
dosages of 15 to 25 Oy (Banerji and Datta, 1990). With increased dosage, survival, plant
height, number of branches and leaves per plant, leaf length and width all decreased.
Flower color mutations were recorded in plants exposed to 15 Oy and 20 Oy at rates of
27.26% and 20%, respectively. These mutations were lighter color, striped color, and a
white color, all of which were maintained and planned for release as new cultivars.
19

Similar results were obtained in a study on a different cultivar of chrysanthemum.
Rooted cuttings of Dendranthema morifolium 'Navneet' (fonnerly Chrysanthemum
morifolium 'Navneet') were exposed to 15,20 and 25 Gy of gamma rays. One new
cultivar was released with a yellow flower color as a result (Banerji et aI., 1996).
Another successful example of radiation induced mutants in Chrysanthemum was
a long tenn program conducted in the Netherlands. Dendranthema morifolium 'Horim'
cuttings were irradiated at 17.5 Gy and rooted. After cutting back the plants two to three
times, cuttings were taken, rooted and planted, then flowers were induced. About 50,000
plants were produced, from which 244 mutants were selected. Not only were new flower
colors obtained, but improvements in quality of stems and larger inflorescences with
better fonn were also obtained (Broertjes et aI., 1980).
Begonias are another ornamental often exposed to irradiation in order to create
new flower colors. Leaf cuttings of Begonia x Hiemalis clones were treated at 0, 15,20,
or 25 Gy. Increased dosages resulted in smaller numbers of adventitious buds per
cutting, increased number of days from planting to removal of first shoot, and increased
percentages of shoots with one or more mutations (Doorenbos and Karper, 1975).
Mutations included changes in flower color, growth habit, leaf color and size. and flower
size and shape. One mutant was introduced as a new commercial variety with a more
vivid hue than the original.
An irradiation study on Hibiscus was also focused on flower mutations. Hibiscus
rosa-sinensis' Alipur Beauty' stem cuttings were exposed to gamma rays from a 6Oeo
source at dosages of 0, 10, 20, or 40 Gy (Banerji and Datta, 1986). With increased
dosage, time to sprouting increased, number of sprouts decreased, and leaf abnonnalities
20

--------
increased. The most significant result of these experiments was a mutation from the
double to the single flower form in two of the plants.
Leaf mutations are often observed in studies involving floral crops, but genera1ly
are not studied long term since it is not the important characteristic of the plant. In other
types of ornamentals, however, changes in leaf characteristics are highly significant
results. For example, two cultivars of double-bracted Bougainvillea cuttings, 'Los Banos
Beauty' and 'Roseville's Delight,' were irradiated with gamma rays from a 6OCO source at
dosages from 0 to 10 Gy (Banerji and Datta, 1987; Banerji et al., 1987). Effects on
sprouting, plant height, leaf number and size were similar to irradiation studies in other
crops with decreases in these characteristics correlated with increases in dosages. Also,
the leaves of plants of both cultivars treated with 7.5 Gy became variegated. These were
both propagated and released as new cultivars.
In most of the above mentioned studies, plant parts were exposed to a relatively
wide range of irradiation dosages. A desirable mutation in at least one plant was obtained
at one or more of these dosages, propagated, and released as a new cultivar. In a
structured irradiation program, however, the first step is to evaluate the "sensitivity" of
the plant material to irradiation. Generally, the best dosage for mutation induction is that
at which 50% of the sample dies, or the LD50. At this dosage a larger number of
mutations can be expected with a sufficient number of samples surviving. For example,
in sweet potato (Ipomoea batatas), stem cuttings were exposed to 60Co gamma radiation
from 0 to 350 Gy. At each dosage, 60 cuttings were treated along with a non-irradiated
control. In this case the LD50 dosage was found to be 200 Gy, the dosage at which 50%
of the stem cuttings survived (Cuevas-Ruiz, 1973). Similarly, in Muscat grape (Vitis
21

vinifera), the LDso was found to be between 20 and 25 Gy (Ponnuswami, 1991).
Experiments in Malaysia have found the LDso for Cordyline cuttings to be 30 Gy, and 15
Gy for Dracaena (Basiran and Ariffin, 2002). However, they specify that this dosage is
highly dependent on the type of plant material used. Green cuttings of these same species
may not be as resistant to irradiation as woodier cuttings. The types of cuttings used in
the experiments are not specified. Of course, the LDso dosages will be different for every
type of plant and can only be determined with experimentation. Once it is determined,
large numbers of plants can be irradiated to attempt to obtain desirable mutants.
Numerous studies have been conducted using irradiation mutation and show the
general trend that higher dosages of radiation result in lower plant survival, growth,
number of buds, flowers, and leaves, and increased numbers of mutations of all types.
Over 600 cultivars in 41 species have been officially released from irradiation mutation
alone (Schum, 2003).
2.5. Polyploid Induction
A polyploid plant is simply defined as a plant combining three or more basic
genomes of the taxonomic group to which it belongs (Dewey, 1980). Polyploid plants
are useful in breeding for many reasons. One of the most important is the characteristics
they tend to have. When compared to their diploid counterparts, polyploid plants are
generally more robust and sturdy. In addition, they often have thicker leaves and stems, a
deeper green color, an increased width-to-length ratio ofieaves, larger flowers, a longer
or later flowering period, and a more compact growth habit (Gao et al., 1996). Another
very useful breeding feature of polyploidy is that it often restores fertility to sterile
hybrids. On the flip side, a tetraploid plant can be crossed with its diploid counterpart to
22

create sterile triploids, which can be useful if seed production in the species is
undesirable.
Polyploidy can be induced or can occur spontaneously in nature. The discovery
in 1937 that colchicine could artificially double the chromosome number in plants was
very exciting and numerous studies were conducted on many different species of crops.
By 1979, the chromosome number of over 150 plant species had been doubled using
colchicine (Dewey, 1980). A few successes in crops were triploid sugar beets, tetraploid
clovers, and the rye x wheat hybrid triticale. There were also many successes in
improving floral crops such as snapdragons, marigolds, zinnias, impatiens, lilies,
delphiniums, and daylilies (Hancock, 1997).
One method of inducing polyploidy is applying chemicals such as colchicine in
vivo. Seeds can be soaked in aqueous solutions, or these solutions can be applied to the
growing points of seedlings or newly emerging buds. Sangowawa (1994) soaked
Solanum melongena seeds in 0.4% colchicine for 2, 4, 6, or 8 days. Tetraploid cells were
found in the seedlings that developed, but the treatments greater than 2 days had negative
effects on seedling emergence. In another study, Crotolariajuncea and Crotolaria retusa
seeds were soaked in 0.025% to 0.30% colchicine solutions for 6 or 12 h. Mixoploids
(plant structures that have originated from meristematic tissue comprised of cells with
varying ploidy levels) and tetraploids were obtained, and tetraploids had smaller plant
heights, shorter and wider leaves, and larger stomata. The percentage of tetraploids
obtained ranged from 0 to 10%, with the best treatment being 0.025% for 12 h (Gupta
and Gupta, 1975). However, a later study of the same species found the best treatment of
Crotolaria seeds for induction of tetraploids was 0.5% for 24 h (Dnyansagar and
23

Nadkami,1983). Seeds of many other species have also been treated by soaking in
solutions of varying concentrations of colchicine. In all these studies, however, the
percentages of conversion to polyploidy were quite low. Treating seeds has not been
shown to be a very efficient method of treatment to induce polyploidy.
Treating the growing point of seedlings or emerging buds is another method
which has been used in numerous studies for inducing polyploidy. Colchicine is applied
to the growing point directly in aqueous solution or in lanolin paste, or the aqueous
solution is applied with a cotton ball and covered to prevent evaporation. Several studies
have shown the improved effectiveness of these methods over treating seeds. More than
16% of Abelmoschus seedlings were converted to tetraploidy while less than 1 % of
treated seeds were converted (Jambhale, 1983). Similar results were found when seeds
and seedlings of Areca palm (Areca catechu) were treated with colchicine, with many
more in the seedling treatment being converted to tetraploids than from treated seeds
(Nair and Ratnambal, 1974). In Cyamopsis psoraloides, none of the seeds treated with
colchicine were converted while up to 33% of seedlings treated were converted to
polyploidy (Biswas and Bhattacharyya, 1971). More positive results were found in
muskmelon (Cucumis melo) with the cotton swab method of application. In this study,
44.5% of treated plants doubled their chromosome number. Also, axillary buds of coffee
were successfully treated to create polyploids in a study conducted on the Ivory Coast
(Berthou, 1975).
When treating emerging buds, another factor which affects the success of
treatment is how close the solution is applied to the apical meristem. The effectiveness of
treatment is dependent on the solution reaching the meristem. In newly emerging buds,
24

the apical meristem is often undemeath many layers of newly forming leaves. A study
conducted on Kiwi (Actinidia spp) treated buds with colchicine, but only obtained
chimeras and no solid tetraploids (Han et al., 1998). In contrast, a study on a camellia
hybrid (Camellia lutchuensis x Camellia rusticana 'Yoshida') successfully obtained a
fertile polyploid from the previously sterile diploid (Ackerman and Dermen, 1996). The
key difference between the procedures of the two studies was that the camellia bud tips
were cut off as close as possible to the apical meristem before treatment while nothing
was done before treatment of the kiwi buds.
Colchicine is a compound that disrupts mitosis by binding to tubulin dimers.
These dimers are then unable to form microtubules and no spindle fibers are formed.
While quite effective in doubling chromosomes, it has some disadvantages. It is very
toxic to humans and also causes some unwanted mutations in plants other than changing
chromosome number. Recently, chemicals other than colchicine have been tested for
induction ofpolyploids such as oryzalin, a dinitroaniline herbicide (3,5 -dinitro-N2, N4-
dipropylsulfanilamide). The mechanism is similar to colchicine in that it also disrupts
mitosis. Oryzalin binds to tubulins, preventing their polymerization into microtubules.
Subsequently, spindle fibers are unable to form, the cell doesn't divide, and the
chromosome number is doubled in that cell (Strachan and Hess, 1983). Advantages of
oryza1in over colchicine are that it is less toxic and is also needed in lower concentrations
for effectiveness. Micromolar concentrations of oryza1in are as effective as millimolar
concentrations of colchicine. Both of these advantages are mainly due to the fact that
oryza1in binds more specifically to plant tubulins than colchicine, which binds more
25

specifically to animal tubulins (Strachan and Hess, 1983). Therefore it is more effective
in plants while being less toxic to humans.
Most recently, in vitro methods of polyploid induction have been studied to a
great extent. The most common method of treatment is addition of the chemical to the
growth medium. Colchicine has been successfully used for induction of polyploidy using
in vitro cultures in a number of plants including butterfly bush (Budd/eia globosa), lilac
(Syringa vulgaris), mountain ash (Sorbus aucuparia), and lily (Lilium spp) (Rose et aI.,
2001; Tuyl et aI., 1992). However, because of the concerns involving colchicine, other
chemicals have been tested in vitro and oryzalin has also emerged as one of the most
effective altematives to colchicine. It has been tested directly against colchicine in vitro
to compare their relative effectiveness and results have shown that oryzalin at much
lower concentrations is generally as effective as colchicine. In oil seed rape (Brassica
napus), comparable rates of chromosome doubling were obtained when using colchicine
at rates up to 3 mM and oryzalin at rates of only up to 30 !lM (Hansen and Andersen,
1996). Similar results were found in Alocasia at concentrations of 0.01 % to 0.1% for
colchicine and 0.005% to 0.05% for oryzalin (Thao et aI., 2003). In this case oryzalin
actually induced more tetraploids than colchicine (15.4% vs. 10.5%) and the data also
showed that oryzalin was less phytotoxic. Studies on Gerbera, Miscanthus sinensis, and
Rhododendron reinforce the data showing the effectiveness of oryzalin at lower
concentrations (petersen et aI., 2003; Tosca et aI., 1995; Vainola, 2000).
2.6. Flow cytometry
Flow cytometry is a technique which can count, examine and sort microscopic
particles suspended in a stream of fluid and allows for the determination of the amount of 26

nuclear DNA content. The technique was originally developed for use in human and
animal cells and is a relatively new method for plants. Single cells or nuclei are isolated
in suspension and stained with DNA fluorochromes. The cell suspension is
hydrodynamically focused through an opening typically sized between 50 to 100 ",m,
allowing the cells to travel downstream, one after another. The flow cytometer is
equipped with a light source, usually a laser or a mercury arc lamp, which is directed into
the fluid stream. Generally, the cell suspension has been stained with a fluorescent dye
which results in fluorescence emission from the cells in the flow stream. A detector in
the flow cytometer then measures the fluorescence emitted (Dolozel, 1991). The amount
of fluorescence emitted corresponds to the DNA content of the sample.
Flow cytometry in plants is used for many purposes, the most common being the
measurement of somatic DNA content, ploidy analysis, and cell-cycle analysis. Somatic
DNA content is estimated by the simultaneous measurement of the fluorescence of
stained nuclei of the species and of a reference standard with known DNA content. A
reference standard is needed for comparison because flow cytometry only provides the
relative amount of DNA content. Ploidy analysis is similar except that the reference
standard used is of the same species with known ploidy level. Cell-cycle analysis
involves determining the fractions of cells in the G I, S and G2 phases of the cell cycle by
using computer programs to analyze the measurements taken by the flow cytometer
(Dolozel, 1991).
A crucial step in flow cytometric analysis is the preparation of the sample.
Galbraith has been a key researcher and the method he described in 1983 was the first
report of the use of flow cytometry for analysis of plant genome sizes (Sklar, 2005). The
27

basic procedure he described involves placing the plant tissue in a "chopping buffer" in a
petri dish and chopping with a single-edged razor blade for less than 2 min. Numerous
chopping buffers have now been described and some are available ready-made from
manufacturers. They generally contain similar components to Galbraith's buffer, which
consisted of magnesium chloride, sodium citrate, 4-morpholinepropane sulfonate, and
Triton X-lOO. After chopping, the mixture is passed through a nylon filter and then
stained with fluorescent dye. Many different types of dyes are used, including propidium
iodide, Hoechst 33342, and DAPI. The entire procedure is performed on ice and the
sample is kept on ice until analysis in the flow cytometer.
The ability to make reliable and rapid determinations of DNA content is the major
advantage of flow cytometry. In plants, leaf tissue is a good source of material for flow
cytometry. This is another advantage because leaf tissues are easy to obtain. Ploidy
analysis via flow cytometry is much less time-consuming and simpler than traditional
chromosome counts via root cell squashing. In addition, active root tips are not always
easily obtained for chromosome counts. The accuracy of flow cytometry has been
proven and the number of samples which can be tested with low statistical error is much
greater than with traditional chromosome counts (Barker et aI., 1998).
28

CHAPTER3. OBJECTfVES
The overall long term objective of this research is to develop new varieties of
Dracaena for the foliage industry. Some of the desirable new characteristics are new
colors and new and unique variegation or morphology. Breeding new varieties of any
plant is a long term endeavor and the focus of this project is to build the foundations for a
long term breeding program.
In order to use traditional breeding (the cross pollination of plants with desirable
characteristics) as the approach to developing new varieties, flowering is essential.
Information pertaining to flowering stimuli in Dracaena is scarce due to the facts that
dracaenas are mainly vegetatively propagated and that flowering in dracaenas is
commercially undesirable. The desired plant architecture of most dracaenas is a single
shoot with no branching. Flowering is terminal, and often results in branching when the
new shoot starts to grow. This results in the destruction of the desired plant symmetry.
In Hawaii and elsewhere, it has been observed that dracaenas flower from late fall
through late winter, though not consistently. This sporadic flowering is a great barrier to
breeding efforts. One of the objectives of this project is to develop an efficient method to
stimulate flowering of Dracaena "on-demand." A large range of methods are used to
induce flowering for various plants, but none have been successfully used on Dracaena.
If a successful method of inducing flowering in dracaena could be found, traditional
breeding methods could be used to cross pollinate different Dracaena species with each
other or even with related genera that normally do not flower simultaneously.
Nontraditional breeding methods are also an important part of any breeding
program. The basis of many nontraditional breeding methods is induced mutation. The
29

objectives of other experiments conducted for this project revolve around nontraditional
breeding methods and the main focus will be to induce mutations in Dracaena. Inducing
mutations is very broad in scope and involves many methods. Mutations can be induced
with an extensive range of mutagens including irradiation and a host of chemicals.
Mutation breeding is a relatively new method compared to conventional cross
pollination, but has now become a well established method to create new varieties. It is
especially useful in vegetatively propagated plants because small phenotypical changes
can be detected and propagated easily (Nybom, 1961).
This project will use irradiation to attempt mutation induction in Dracaena.
Different types of irradiation such as gamma rays and x-rays are commonly used. The
best dosage to use for inducing mutations in an irradiation program is the dosage at which
50% of the treated plants survive, called the LDso. In order to set up a long term
irradiation breeding program, this dosage must be determined. The objective of this part
of the project will be to determine the LDso for Dracaena using gamma rays. Knowing
the LDso will enable future studies to irradiate large numbers of Dracaena to obtain
useful mutants.
Another mutagen used in this project will be the chemical mutagen oryzalin. The
compound oryzalin has been successful in inducing polyploidy in many plants.
Commercially grown dracaenas are diploids. However, it is well known that polyploid
forms of plants generally have desirable characteristics (Gao et al., 1996). The objective
of using oryzalin as a mutagen is to create a tetraploid form of dracaena which hopefully
will have desirable new characteristics or will be useful in creating variation in
hybridizing efforts.
30

CHAPTER 4. FLOWER INDUCTION
Introduction
Traditional breeding requires the aVailability of flowers on the desired parent
plants. An efficient method to stimulate flowering of dracaena "on-demand" would be
very helpful in hybridizing efforts to create new Dracaena varieties.
GA3 treatments have successfully induced flowering in a number of tropical
foliage crops (Henny, 1980, 1981, 1983, 1988, 1991; Henny et al., 1999). However, one
study that used GA3 on Dracaena met with little success (Fisher, 1980). Although the
results of that study were not positive, they were only preliminary. The treatments were
done at only one concentration with one replication. The many reported successes by
Henny on tropical foliage crops warrant further study on the use of GA3 to induce
flowering in Dracaena.
The only published information about the flowering of Dracaena aside from
botanical descriptions is that of Bos in 1984. According to his description, abundant
nectar is produced below the pedicels but the production bodies cannot be seen. The
buds more than double in size during the 24 h prior to anthesis, which occurs in the
evening. A sweet heavy odor is produced in the later hours, and he personally observed a
certain species of Hawk moth hovering outside the glass roof above where dracaenas
with inflorescences were kept. Dracaena flowers that open during the night fade by
dawn.
According to observational accounts, flowering in Dracaena occurs most
regularly during late fall and winter in Hawaii when temperatures are lower. However, I
31

have personally observed that not all species flower during the same time periods and
some species will flower at multiple times of the year. In addition, while some individual
plants are flowering in a large group, not all will do so at the same time. These
observations imply that flowering is not exclusively due to a requirement for photoperiod
since there is no consistency in timing of flowering. Since flowering mainly occurs
during cooler temperature periods, there is good reason to believe that flowering is
associated with exposure to cold temperatures. In addition, cold temperatures have been
used to induce flowering in many plants(Beattie and Holcomb, 1983; Wright and
Milbocker, 1979). These facts point to the possible success of inducing flowering in
Dracaena using cold temperatures.
Materials and Methods
All flower induction treatments (GA3 and cold temperature) were made during
spring and summer months when dracaenas do not flower to ensure that floral initiation
had not already occurred.
GA3 treatments
Potted plants or field-grown plants of nine different Dracaena varieties were
treated (D. deremensis 'Bausei', Janet Craig, 'Janet Craig Compacta', 'Santa Rosa', and
'Wameckei', D. fragrans, D. fragrans 'Massangeana' and 'Massangeana Compacta', and
D. xmassefflana) (Table 1). A total of 13 trials were carried out on the nine varieties with
concentrations of GA3 ranging from 0 to 6000 ppm. GA3 solutions were prepared by
dissolving Gibberellin A3 (Sigma-Aldrich, St. Louis, MO) in distilled water and diluting
to the desired concentrations. Treatments were made by applying the solution to the
apical cup of each plant. A total of five (1 mL each) applications were made to each plant 32

at three day intervals. The experiments were replicated between 2 to five times
depending on the variety being treated. The number of plants (or stems if there were
multiple stems per plant) which formed inflorescences was recorded biweekly for 16
weeks after treatment and the percentage of flowering stems was calculated. Several of
the trials were performed by Patricia Nakao, a collaborator on this project at the Maui
Agricultural Research Center in Kula, Hawaii and by Joanne Lichty, another collaborator
at the Beaumont Agricultural Research Center in Hilo, Hawaii. All other trials were
performed on Oahu at the University of Hawaii Waimanalo Experiment Station, Pope
Laboratories at the University of Hawaii in Manoa, and the Magoon Research Station at
the University of Hawaii in Manoa.
Cold temperature treatments
Experiment #1
Plants in 2S cm pots were placed in a cooler unit in Waimanalo, Hawaii and
provided with an 8 h photoperiod from nine 60 Watt incandescent bulbs. Four different
dracaena cultivars were used in the experiment with five plants per treatment Plants
were treated for 0, 3, 6, or 9 days and the experiment was replicated three times. D.
deremensis 'Santa Rosa' was treated at 8,12,16, and 20°C, D.fragrans 'Massangeana'
at 12 and 16°C, D. deremensis 'Janet Craig' at 8 and 12°C, and D.xmasseffiana at 12
°C. D. deremensis 'Santa Rosa' and D. fragrans 'Massangeana' plants were
approximately I.S m tall and had been rooted for S months. D. deremensis 'Janet Craig'
plants were approximately 2.5 m tall and had been rooted for over 2 years.
D. Xmasseffiana plants were approximately 1.5 m tall and had been rooted for 1 year.
Since only one cooler unit was available, treatments were made successively at each 33

temperature. The number of plants (or stems if there were multiple stems per plant)
which formed inflorescences was recorded biweekly for 16 weeks after treatment and the
percentage of flowering stems was calculated.
Experiment #2
Plants in 25 cm pots were placed in a cooler unit at the University of Hawaii Pope
Laboratories for seven consecutive nights. Plants were put into the unit at 5:00 PM and
taken out the next morning at 8 :00 AM. Four chambers were used for four temperatures
(6, 9, 12, and 15°C) and four different varieties were treated (D. deremensis 'Janet
Craig' and 'Santa Rosa', D. fragrans 'Massangeana', and D. Xmassefftana). There were
8 stems of D. deremensis 'Janet Craig', 9 stems of D. deremensis 'Santa Rosa', 6 stems
of D. fragrans 'Massangeana', and 30 stems of D. xmassefftana per treatment (Table 2).
In addition, there was a control treatment in which plants were placed in darkness
for 14 h for seven nights but at a temperature of 23 :I: 1°C. After treatment, the 64 plants
were placed in a greenhouse at the University of Hawaii Magoon Research Station for
observation. The number of plants (or stems if there were multiple stems per plant)
which formed inflorescences was recorded biweekly for 16 weeks after treatment and the
percentage of flowering stems was calculated.
Experiment #3
Plants were placed in a cooler unit at the University of Hawaii Pope Laboratory
for two durations at two different temperatures for a total of five treatments with a
control.
1. Control 2. 12°C for 20 days 3. 12°C for 40 days
34

4. 15°C for 20 days 5. 15°C for 40 days
Potted plants offive different varieties (D. fragrans 'Massangeana Compacta' and
'Massangeana', D. Xmasseffiana, D. surculosa, and D. sp. 'Fantasy' (Accession number
L-8210373» were used in this experiment with at least six plants per treatment (Table 3).
After treatment, plants were placed in a greenhouse at the University of Hawaii
Magoon Research Station for observation. Plants were placed in 50% shade and irrigated
once a day for three minutes with overhead sprinklers. The number of plants (or sterns if
there were multiple stems per plant) which formed inflorescences was recorded biweekly
for 16 weeks after treatment and the percentage of flowering stems was calculated.
Results
GAJ treatments
The first inflorescences of D. xmasseffiana started emerging 2 weeks after the last
GA3 application. There was flowering in all treatments except the control 4 weeks after
the last application. Higher concentrations produced increased percentages of flowering
stems (Figure 1). Flowering percentages ranged from 0% in the control to 66.7% in the
2000 ppm treatment. Single factor analysis of variance (ANDV A) was performed on the
results and the mean flowering percentages for the treatments were found to be
significantly different (P = 0.023). Subsequently, Tukey's test was performed on the
means of the four treatments. The 1000 ppm and 2000 ppm treatments were significantly
different from the control treatment (Figure 1). In addition, linear regression was
calculated and determined to be highly significant (p < 0.01) (Table 4).
35

The percentages of stems of D. /ragrans 'Massangeana' flowering in the control,
500 ppm. and 1000 ppm treatments were all 20%. The 2000 ppm treatment was higher
with 40% of the stems flowering (Figure 2). Single factor ANOV A showed no
significant difference between the mean flowering percentages (P = 0.551). In addition,
linear regression was calculated and was not significant (p = 0.19) (Table 4).
No flowering was initiated in the control and treated plants of the other trials
performed on D. deremensis 'Bausei', 'Janet Craig, 'Janet Craig Compacta', 'Santa
Rosa', and 'Warneckei', D./ragrans. and 'Massangeana Compacta'.
Cold temperature treatments.
Experiment #1
Inflorescences of D. xmassefJiana plants treated at 12°C for 3,6, and 9 days
emerged within 3 weeks after the plants were removed from the cooler. Flowers were
visible approximately 30 days after removal from the cooler. None of the control plants
flowered. The percentage of stems that flowered increased with increased treatment
duration with the average flowering percentages for the treatments being 16.9%, 39.2%,
and 86.7% for the 3, 6, and 9 day treatments, respectively (Figure 3). Single factor
ANOV A determined that the differences in the mean values among the treatments were
highly significant (p < 0.001). Tukey's test was performed on the means of the four
treatments (Figure 3). In addition, linear regression was calculated and determined to be
highly significant (P < 0.01) (Table 4).
Neither the controls nor treated plants of D. deremensis 'Santa Rosa' and D.
/ragrans 'Massangeana' initiated any inflorescences.
36

One plant of D. deremensis 'Janet Craig' treated for 6 days at 12°C produced an
inflorescence within 4 weeks after being removed from the cooler. None of the control
plants and none of the other treated plants initiated any inflorescences.
Experiment #2
One stem of one plant of D. xmassefflana treated at 15°C flowered 4 weeks after
removal from the cold chambers. Neither the controls nor the other treated plants formed
any flowers. None of the treated plants showed any signs of cold damage.
Experiment #3
Neither the controls nor treated plants of D. fragrans 'Massangeana',
'Massangeana Compacta', and D. sp. 'Fantasy' L-8210373 formed any flowers.
One stem of one D. surculosa plant treated at 15°C for 20 days formed an
inflorescence 4 weeks after removal from the cold temperature chamber. In addition, two
inflorescences formed on one D. surculosa control plant.
D. xmassefflana plants treated at 15°C for 20 days were the only ones that formed
inflorescences. A total of 11 inflorescences were produced on 39 stems. Out of the 11
formed, three were formed 4 weeks after removal, and eight were formed 10 weeks after
removal from the cold storage.
All the D. jragrans 'Massangeana' and D. fragrans 'Massangeana Compacta'
plants treated at 12°C for 40 days suffered cold damage and eventually died. In the other
three species treated (D. xmassefflana, D. surculosa, and D. sp. 'Fantasy' L-8210373),
damage was apparent on plants treated at 12°C. The damage was most apparent on the
leaves, many of which formed large necrotic spots and eventually dropped off. This
37

particular cold temperature chamber was previously used in experiments with chlorine
and it is possible that the chlorine remaining in the chamber may have damaged the
plants also. However, these plants all recovered and resumed normal growth.
Discussion and Conclusions
The GA3 treatments were ineffective in promoting flowering in the majority of
Dracaena species used in these experiments, contrary to previous successes in other
tropical foliage plants (Corr and Widmer, 1987; Henny, 1980, 1981, 1983). The GA3
treatment methods reported by Fisher (1980) to be very successful with Cordyline species
were employed because of the close taxonomic relationship between Cordyline and
Dracaena. In addition, the morphology of Dracaena is especially conducive for GA3
applications. GA3 solutions were applied to the apical cup of the plants because the apical
cup holds solutions well, and because it is the location where an inflorescence would
develop. The method allows maximum contact and absorption by the apical meristem
and it was thought that this method would be the most effective. Past successes in eight
different genera of the Araceae family used the foliar spray method of application and a
larger volume of GA3 solution (between 10 and 25 mL) for each treatment on each plant
(Henny, 1981, 1983, 1988). In our experiments, 1 mL per application was used, which is
a larger amount of solution than Fisher (1980), but not nearly as much Henny (1980;
1981; 1983; 1988). Also, the concentrations used in previous studies on Araceae plants
ranged from 100 to 1000 ppm. Higher concentrations of GA3 solution were used in
several of our trials on Dracaena, but these were not effective either. In addition, many
of the GA3 treatments were carried out on plants that were unprotected from such
conditions as wind and rain. It was not feasible in these cases to cover the plants because
38

of their large sizes. In addition to completely protecting treated plants from the elements,
it is possible that using larger amounts of solution, using the foliar spray method of
application or using different types of GA such as G~ or GA7 may be a more effective
method in inducing flowering in Dracaena. For example, it has been shown in azaleas
that GA3 is more effective in inducing anthesis while GA7 has a role in inducing
budbreak (Nell et ai., 1983).
The small amount of flowering which occurred in D . .fragrans 'Massangeana'
may not have been due to the GA3 treatments. The untreated control plants flowered at
the same rate as the 500 ppm and 1000 ppm treated plants and no significant difference
was found between treatments (Figure 2). Therefore it is possible that the plants had
already initiated flowers prior to the treatments and that the flowering observed during
the experiment would have occurred regardless of treatment.
D. xmasseffiana, the only known interspecific hybrid of Dracaena, was the most
responsive to the GA3 treatments. Higher concentrations of solution corresponded with a
higher percentage of flowering (Figure 1). However, other studies have shown that
hybrids are less responsive to applied GA3 than their parents. In a study on pearl millet
(Pennisetum glaucum) hybrids and their parents, the hybrids showed a lower response to
applied GA3 than their parents (Sadasivam and Saxena, 1998). Similar results have been
reported in maize, with the inbred response to GA3 much greater than the hybrid (Rood et
ai., 1983). D . .fragrans 'Massangeana', one of the parents of the hybrid D. xmasseffiana,
was SUbjected to GA3 treatments in this study and showed a low flowering response.
Slightly more flowering occurred in the GA3 treated plants than in control plants. The
other parent, D. surculosa, should be tested for flowering response to GA3 treatments to
39

determine if it is responsive. A high flowering response of D. surculosa to GA3 and the
fact that Bos (1984) has reported that D. surculosa is the most readily flowering species
in cultivation might point to the flowering responsiveness of D. xmasseffiana being due
to having D. surculosa as one ofits parents.
The mechanisms of signal transduction triggered by GA3 are still poorly
understood. The level of sensitivity to exogenous GA3 applications can be an indicaror of
the endogenous levels of gibbere1lins present in that plant. Ashikari and Matsuoka
(2002) have done extensive studies on the genetics of rice (Oryza sativa). According to
their reports, many GA-related mutants from numerous plant species have been isolated
and can be split into two general categories, GA-sensitive and GA-insensitive. A rice
mutant that responds to exogenously applied gibberellins does so because it cannot
produce or produces insufficient gibberel1ins. One study used a GA-insensitive mutant in
which the levels of endogenous gibberellins (various types, including GAl and GA2o)
were found to be 100 times higher than the levels found in wild-type plants.(Ashikari and
Matsuoka, 2002). In fact, exogenous GA3 applications are often used to determine
relative levels of endogenous gibberellins between different plants. In the study on pearl
millet, hybrids and their parents were tested for level of response to exogenously applied
GA3 and these results were used to estimate endogenous gibberellin levels. The hybrids
showed a lower response to applied GA3 and had higher levels of bound GA-like
substances than their parents (Sadasivam and Saxena, 1998). A study by Alamu and
McDavid (1978) on the promotion of flowering in edible aroids using GA3 found that it
was more effective in promoting flowering in dasheen (Colocasia esculenta) than in
tannia (Xanthosoma sagittifolium). They hypothesized that the differential effect of GA3
40

treatments was due to a difference in endogenous gibberellin levels. It is possible that
the cultivated species used in this study that did not respond to GA3 treatments had high
levels of endogenous gibbere11ins, while the hybrid does not. This would help to explain
the high response of the hybrid and the low response of the species. The endogenous
levels of gibberellins in Dracaena were not determined.
It is evident that plant response to gibberel1ins is complex. Jones (1973) once
stated that "ail aspects of the growth and development of higher plants from seed
germination to fruit set can be affected by GAs." To add to this complexity is the fact
that floral initiation can be inhibited by application of GA3. Research reports show that
flowering of Hedera helix, Malus domestica, and Euphorbia pulche"ima have been
inhibited after exogenous applications of GA3 (Evans et ai., 1992). A possible
explanation for the inhibition is that nutrients are diverted from the shoot apex. This
explanation was confirmed by King (2003) when measured sucrose levels at the shoot
apex decreased after GA application. Extensive physiological studies would have to be
performed to determine if these scenarios could be occurring when GA is applied to
Dracaena, but they serve as possibilities to be considered for any future work in flower
induction.
Cold temperature treatments were ineffective in promoting flowering in three of
the four dracaena used in Experiment #1. Previous studies have been published on
successful induction of flowering after storage in cold temperatures on plants such as
rhododendron, peony and Cape Daisy, but these plants were already known to require a
cold period for initiation of floral development (French and Alsbury, 1988; Kamenetsky,
2003; Pearson et ai., 1995). Cold temperature treatments were attempted because of
41

observations that dracaenas tend to flower only during the coolest seasons of the year.
Plants are not able to flower or will not respond to a low temperature stimulus until they
have reached a critical biomass (Vince-Prue, 1975), and the D. deremensis 'Santa Rosa'
and D. fragrans 'Massangeana' plants used in this experiment may not have accumulated
enough biomass to respond to cold temperature treatments. The roots were well
established, but the plants did not have a large amount of new growth because they had
been propagated only 3 months before treatment. In contrast, rooted cuttings of D.
xmasseffiana that responded to the cold treatments in this experiment had been growing
in pots for over a year before this treatment.
Lower temperatures were used in Experiment #2 in an attempt to induce
flowering in the varieties which didn't flower in Experiment # 1. The treatments were
also set up to simulate natural conditions where night temperatures are cooler than day
temperatures. However, the desired results were not achieved as only one stem of one
treated plant formed an inflorescence. The procedure used in this study was based on the
observation that flowering in dracaena occurs 7 to 8 weeks after several hours of 13 to 16
°C and/or longer periods of 16 to 18 °C (J. Lichty, personal communication). Although
temperatures below this observed critical range were used in the experiment, it is possible
that one week of exposure was too short.
Experiment #3 was based on the work done by Lu (2003), in which D. fragrans
'Massangeana' plants were induced to flower using temperatures of 12 and 15 °C for 20
and 30 days with 8 h of light and 16 h of darkness. The treatments used in Experiment #3
replicated those described in Lu's experiment but used 20 and 40 days as the durations in
cold temperature. Lu didn't report the exact results of their experiment except to say that
42

frequency of inflorescence formation could reach 100%. However, flowering frequencies
in Experiment #3 were not close to 100%. It is unclear why this experiment didn't garner
similar results when the treatments used were the same.
The only positive results obtained in the cold temperature treatments were with D.
xmassefJiana. The control plants did not flower even 13 weeks after the treatment ended.
In chilled plants, the number of stems with flowers was positively correlated with the
amount of time the plants were exposed to the cold temperature. The difference in the
results between the hybrid and the species treated may be due to the hybrid requiring
different conditions for flowering from the species. This observation has been made in
other families of flowering plants. Menninger observed that Cymbidiumjloribundum
(formerly C. pumilum) species have a narrower temperature range tolerance than C.
jloribundum hybrids. In addition, C. jloribundum hybrids flower over a longer period of
time and a wider temperature range than the species (Menninger, 1965). It is pgssible
that the dracaena hybrid will flower in a wider range of temperatures than Dracaena
species. Less restrictive environmental requirements for flowering would be an
advantageous trait for breeding purposes.
Neither GA3 treatments nor cold temperature treatments successfully induced
Dracaena flowering in the majority of the varieties tested. Other factors may be involved
in initiation of flowering in Dracaena such as photoperiod or the effects of dry and wet
seasons. The flowering requirements for a specific species are complex, and often depend
on more than one factor. Dracaena may need a very specific combination of photoperiod
and temperature to flower. Other types of treatments could be tested such as ethephon or
perhaps a combination of GA and cold temperature treatments.
43

However, both types of treatments were effective in promoting flowering in the
single known interspecific hybrid, D. xmassefJiana. It is possible that these positive
results were due to the fact that the plant is an interspecific hybrid. More work should be
done to determine a reliable method for inducing dracaenas to flower, the results of
which would aid in Dracaena breeding efforts.
44
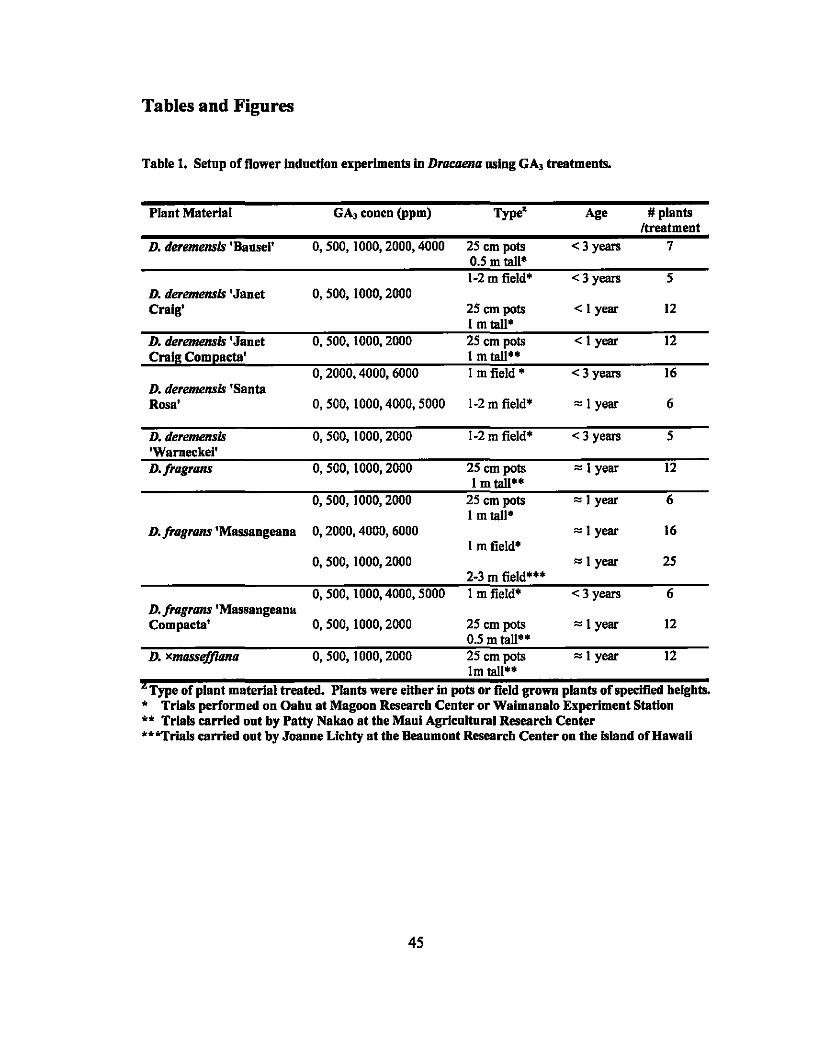
Tables and Figures
Table 1. Setup of flower indnction experiments in Dracaena using GA, treatments.
Plant Material GA, concn (ppm) Type" Age # plants Itreatment
D. deremensis 'Bansel' 0,500,1000,2000,4000 25 em pots <3 years 7 0.5 m tall' 1-2 m field" <3 years 5
D. deremensis 'Janet 0, 500, 1000,2000 Craig' 25 em pots < I year 12
I m tall· D. tieremensis 'Janet 0, 500, 1000, 2000 25 em pots < I year 12 Craig Coml!acta' 1 m tall'·
0,2000,4000,6000 I m field' <3 years 16 D. deremensis 'Santa Rosa· 0,500,1000,4000,5000 1-2 m field' '" I year 6
D. deremensis 0, 500, 1000, 2000 1-2 m field' < 3 years 5 'Warneckel' D·fragrans 0, 500, 1000, 2000 25 em pots '" I year 12
I m tall·· 0, 500, 1000, 2000 25 em pots '" 1 year 6
I m tall" D. fragrans 'Massangeana 0,2000,4000,6000 '" I year 16
I m field" 0, 500, 1000, 2000 '" I year 25
2-3 m field'·· 0,500,1000,4000,5000 I m field" <3 years 6
D·fragrans'Massangeana Compacta' 0, 500, 1000, 2000 25 em pots '" 1 year 12
0.5 m tall" D. xmassefj1ana 0, 500, 1000, 2000 25 em pots '" I year 12
1m tall·· t Type of plant material treated. Plants were eitber in pots or field grown plants of specified belgbts.
" Trials performed on Oabn at Magoon Researcb Center or Waimanalo Experiment Station "" Trials carried ont by Patty Nakao at the Manl Agrlcnltural Researcb Center ""*Trials carried ont by Joanne Licbty at the Beaumont Researcb Center on tbe Island of Hawaii
45
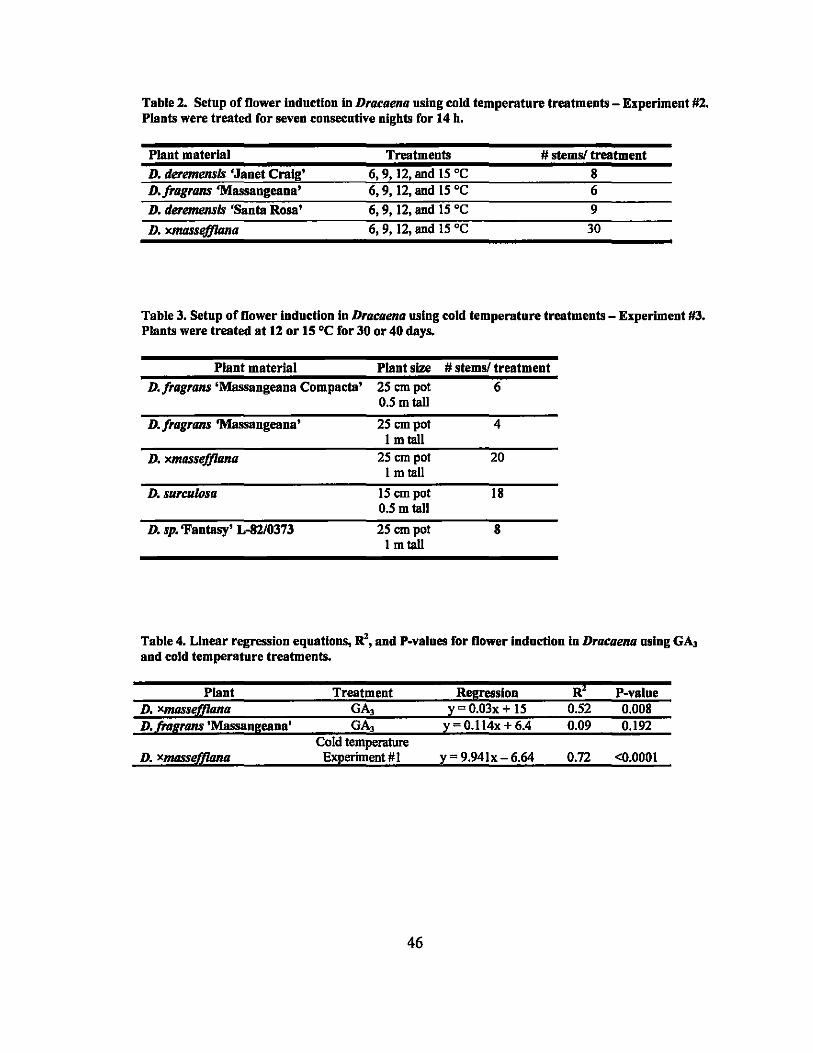
Table 2. Setup of flower Induction In Dracaena using cold temperature treatments - Experiment #2. Plants were treated for seven consecutive nights for 14 h.
Plant material Treatments # stems! treatment
D. deremensls 'Janet Craig' 6, 9, 12, and 15 DC 8 D·fragrans'Massangeana' 6,9, 12, and 15 DC 6
D. deremensls 'Santa Rosa' 6,9, 12, and 15 DC 9
D. XJt/I1SSe/flllna 6,9, 12, and IS DC 30
Table 3. Setup of flower Induction In Dracaena using cold temperature treatments - Experiment #3. Plants were treated at 12 or 15 DC for 30 or 40 days.
Plant material Plant size # stems! treatment
D. 'ragrans 'Massangeana CompaetB' 25 em pot 6 0.5 m tall
D. 'ragrans 'Massangeana' 25 em pot 4 I m tall
D. xmassefflana 25 em pot 20 I m tall
D. surculosa 15 em pot 18 0.5 m tall
D. sp. 'Fantasy' L-82/0373 25 em pot 8 I m tall
Table 4. Linear regression equations, R', and P-values for flower indnction In Dracaena using GA, and cold temperature treatments.
Plant Treatment Regression R' P-value D. xl1IIJSstif(lana GA, y 0.03x + IS 0.52 0.008 D. (ragrans'Massangeana' GA, y-0.114x+6.4 0.09 0.192
Cold temperature D. Xl1IIJSSfd(iona Experiment # I y = 9.94lx - 6.64 0.72 <0.0001
46
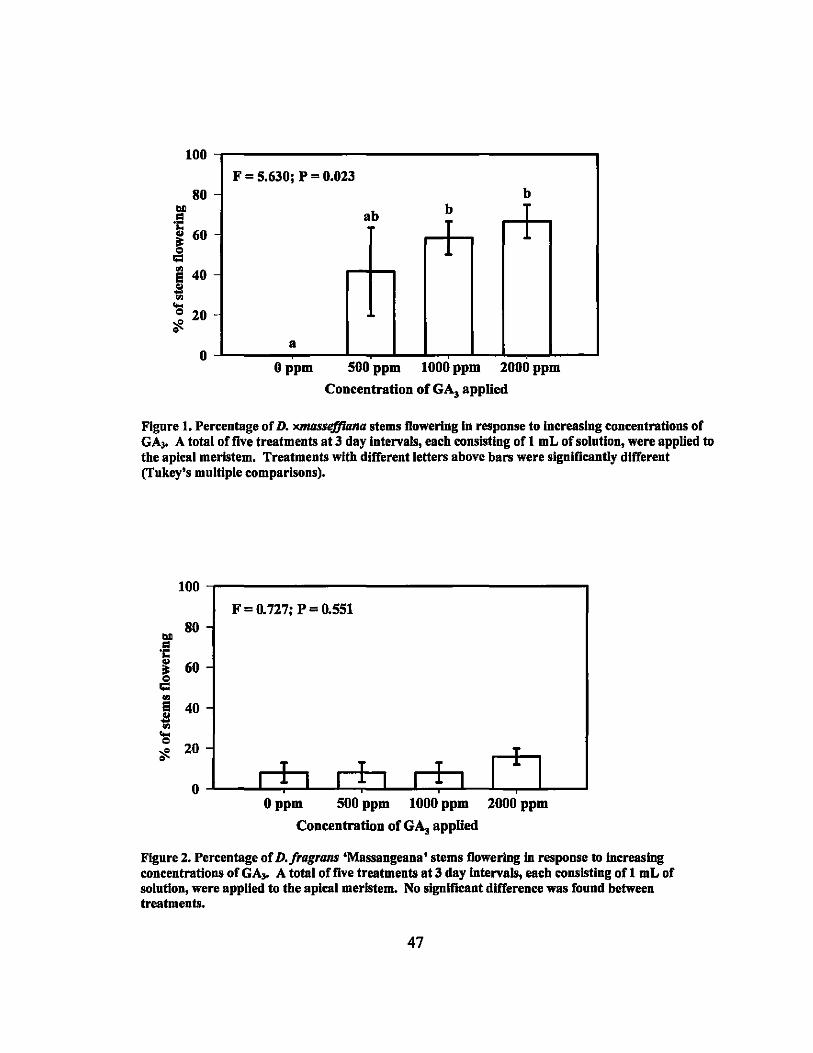
100
F = 5.630; P = 0.023 80 - b
ab b T T 1 1
a o o ppm 500 ppm 1000 ppm 2000 ppm
Concentration of GA, applied
Figure I. Percentage of D. xmasse.ffiana stems flowering In response to increasing concentrations of GA" A total of live treatments at 3 day Intervals, each consisting of 1 mL of solution, were applied to the apical meristem. Treatments with different letters above bars were significantly different (Tukey's multiple comparisons).
100
F = 0. 727; P = 0.551 80
bll .. "t:
~ 60
-= ..,
! 40
... = 20 ~
0
T
T T T I ~ I I J. I I J. I I J. I o ppm 500 ppm 1000 ppm 2000 ppm
Concentration nf GA, applied
Figure 2. Percentage of D. fragrans 'Massangeana' stems flowering In response to increasing concentrations ofGA .. A total offive treatments at3 day Intervals, each consisting ofl mL of solution, were appUed to the apical meristem. No slguificant difference was found between treatments.
47
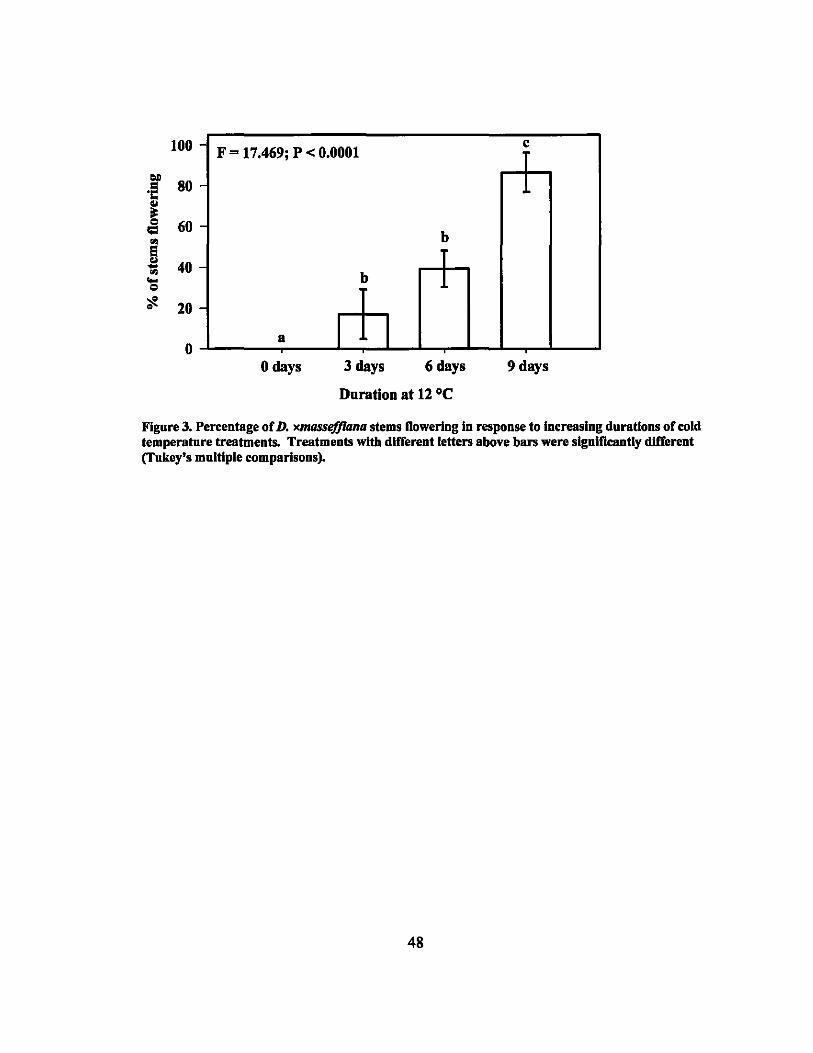
100 - F = 17.469; P < 0.0001 c
IOJ)
80 .~ -.. ~ 60 = ."
-b
El .. 40 ...
." ... -b co
"I- 20 -
0 a I I
o days 3 days 6 days 9 days
Duration at 12°C
Figure 3. Percentage of D. xmassefJlana stems flowering In response to increasing durations of cold temperature treatments. Treatments with different letters above bars were significantly different (Tukey's multiple comparisons).
48

CHAPTERS. IRRADIATION
Introduction
Nontraditional breeding methods such as using irradiation as a mutagen can be an
important part of an organized breeding program. The use of x-rays and gamma rays has
proven effective in producing new varieties of a number of ornamental crops, including
chrysanthemum, hibiscus, and bougainvillea (Banerji and Datta, 1986; Banerji et al.,
1987; Broertjes et al., 1980). Although the mutations induced are "accidental," designed
experiments allow for selection of useful phenotypes. The first step in creating an
organized mutation program is determining the dosage at which 50% of the plants treated
die (LDso). This dosage is the optimum dosage to use when trying to create mutations
because a large number of mutations can be obtained while still retaining a large number
of surviving plants.
Electromagnetic radiation (x-rays and gamma rays) are widely used in irradiation
mutation programs because they are more readily available than other types of radiation.
However, many x-ray machines can only accommodate eight to ten five-inch long
Dracaena cuttings at one time, and the time it takes to reach equivalent dosage levels can
be 75 times longer for an x-ray machine than for gamma rays. Therefore gamma ray
irradiation is a more practical treatment for Dracaena than x-rays.
Very few irradiation studies have been conducted on Dracaena. One study stated
that the lethal dosage was 15 Gy using gamma rays from 6OCO, but did not specify the
species or the type of plant material that was used (Basiran and Ariffin, 2002). Another
study reported new mutants of D. sanderiana and D. surculosa (formerly D. godsefJiana)
49

were obtained at dosages of up to 30 Gy (Lapade et aI., 2002). It is difficult to interpret
these results because cuttings of different species and varieties of Dracaena can vary
greatly in'size and woodiness. Therefore, the objective of this experiment is to conduct a
systematic study of irradiation on Dracaena species to determine the LDso, which will
provide the foundation for a long term irradiation program to create new varieties.
Materials and Methods
Experiment #1
Cuttings were obtained from field grown stock plants, cleaned, and cut to size.
The cane sections used did not have any leaves. They were graded for uniformity and
randomly assigned to treatments. The top end of each cutting was covered in paraffin
wax to prevent desiccation. In addition, a control group of untreated cuttings were cut at
the same time, stored in the same .conditions, then rooted at the same time as treated
plants. Cuttings of D. deremensis 'Santa Rosa' and D. fragrans 'Massangeana' were
approximately 12 to 13 cm long and 2 to 2.5 cm in diameter. Cuttings of D. xmassefJlana
were approximately 12 to 13 cm long and 1 to 1.5 em in diameter.
The unrooted cuttings were treated at the USDA Irradiation facility in
Waimanalo, Hawaii on 21 Dec. 2005 with \31Cs. After treatment, the cuttings were
immersed in water for 30 seconds and planted in metal trays in Pro-Mix BX® medium
(Premiere Horticulture, Quakertown, PA) for rooting and observation. The three
replications of each treatment were planted in one tray. The trays were placed in a
glasshouse covered with 50% shadecloth and irrigated three times a day for 2 min with
overhead sprinkIers.
50

The experimental design was a completely randomized design with seven
treatments and three replications for each variety treated. The seven treatments were the
irradiation dosages of 0, 50, 100, 150,200,250 and 500 Gy. The treated varieties were
D. deremensis 'Santa Rosa', D.fragrans 'Massangeana', and D. xmasseffiana. Each
replication consisted of 10 cuttings.
The shoot and root formation percentages were recorded, and observations were
taken on any abnorma1ities in leaf color, shape, contortion, size, etc.
Experiment #2
Cuttings were obtained from field grown stock plants, cleaned, and cut to size.
They were graded for uniformity and randomly assigned to treatments. The top end of
each cutting was covered in tree sealant to prevent desiccation. In addition, a control
group of untreated cuttings was cut at the same time, stored in the same conditions, then
rooted at the same time as treated plants. Cuttings of D. deremensis 'Santa Rosa', D.
fragrans 'Massangeana', and D.fragrans 'Victoriae' were approximately 12 to 13 cm
long and 2 to 2.5 cm in diameter. Cuttings of D. xmasseffiana were approximately 12 to
13 cm long and 1 to 1.5 cm in diameter.
The unrooted cuttings were treated at the USDA Irradiation facility in
Waimanalo, Hawaii on 24 May 2006 with 137 Cs. After treatment, the cuttings were
treated with Dip 'N Grow™ Liquid Rooting Hormone (Dip 'N Grow, Inc., Clackamas,
OR) consisting of 1000 ppm rnA and 500 ppm naphthalene acetic acid (NAA) and
planted in metal trays in Pro-Mix HP® medium (premiere Horticulture, Quakertown,
PA) for rooting and observation. The three replications of each treatment were planted in
51

one tray. The trays were placed in a glasshouse in an area covered with 50% shadecloth
and irrigated three times a day for 2 min with overhead sprinklers.
The experimental design was a completely randomized design with seven
treatments and three replications for each variety treated. The seven treatments were the
irradiation dosages of 0, 5, 10, 20, 30, 40, and 50 Gy. The treated varieties were D.
deremensis 'Santa Rosa', D. fragrans 'Massangeana', D. fragrans 'Victoriae', and D.
xmasseffiana. Each replication consisted of 10 cuttings.
The shoot and root formation percentages were recorded, and observations were
taken on any abnormalities in leaf color, shape, contortion, size, etc.
Results
Experiment #1
None of the treated cuttings in the first round of irradiation treatments formed
roots or shoots. The untreated cuttings had nearly 100% root and shoot formation (Table
5). For both D. deremensis 'Santa Rosa' and D. fragrans 'Massangeana', the shoots of
two cuttings in the control set were all green with none of the variegation usually
observed in these cultivars.
Experiment #2
The second round of irradiation showed a general trend of decreased root and
shoot formation with increased dosage with each variety showing a slightly different
response (Table 6).
The number of cuttings in each treatment with at least one root formed was
recorded approximately every 2 weeks. This number was used to calculate percentage of
52

cuttings with roots and plotted against days since treatment to show the different rooting
responses of the different varieties (Figure 4).
In D. deremensis 'Santa Rosa', the percentage of cuttings with roots in the control
reached 100% after only 12 weeks. The percentage of cuttings with roots formed
decreased as the dosage of irradiation increased. At 30 Gy and higher, no roots formed
on any of the cuttings.
D. fragrans 'Victoriae' showed a similar trend in which the percentage of cuttings
with roots formed decreased as the irradiation dosage increased. However, the controls
reached only 70% of cuttings with roots and no roots formed in any treatments of20 Oy
and higher.
D. fragrans 'Massangeana' showed a slightly different trend of root formation.
During the first 8 weeks of data collection, the lOGy treatment had greatest number of
cuttings form roots, followed by the 5 Oy treatment, followed by the controls. After
these 8 weeks, though, no additional cuttings in the 5 Gy and 10 Oy treatments formed
any roots, while the control group had additional cuttings forming roots. At the end of
data collection, the controls still had the greatest number of cuttings with roots, and the
percentage decreased with increased radiation dosage.
The root formation trend of D. xmassejJiana was also different from the other
varieties treated. At the beginning of the data collection period, the percentage of
cuttings with roots was highest in the 5 Gy and 10 Oy treatments, followed by the
controls and 20 Oy treatments. About 8 weeks into the experiment, the percentages of
cuttings with roots in the control,S Oy, 10 Oy and 20 Oy treatments were almost equal.
By the end of the experiment, the treatment with the highest percentage of cuttings with
53

roots formed was the 10 Gy treatment, followed by 5 Gy, 20 Gy, the control, 30 Gy, and
40 Gy treatments. No cuttings formed roots in the 50 Gy treatment.
The number of cuttings with at least one shoot visible was recorded
approximately every 2 weeks. This number was used to calculate the percentage of
cuttings with shoots and plotted against the number of days since treatment to show the
different sprouting responses of different varieties (Figure 5). The trends of shoot
formation were similar but not identical to the trends in root formation.
In D. deremensis 'Santa Rosa', the rate of shoot formation decreased with
increased radiation dosage (Figure 5). In the controls, 100% of cuttings formed shoots 16
weeks into the experiment, which was about 4 weeks after 100% of the cuttings formed
roots. In the 5 Gy treatment, 100% of cuttings had formed shoots 18 weeks into the
experiment, but not all had formed roots at the end of data collection. In general, root
formation occurred more quickly than shoot formation for this variety.
The percentage of cuttings with shoots formed in D. fragrans 'Victoriae' for the
control and 5 Gy treatments were similar during the first 8 weeks of the experiment. After
this initial period, the percentage of cuttings with shoots formed in the control treatment
was higher than that in the 5 Gy treatment. By the end of data collection the percentages
in the two treatments were almost equal, with 100% of the controls and 96.7% of the 5
Gy treatments with shoots (Figure 5). The higher dosages followed the general trend of
decreased shoot formation with increased dosage, with none of the cuttings forming
shoots in the 50 Gy treatment. In general, shoot formation occurred more quickly than
root formation for D. fragrans 'Victoriae' (Figures 4 and 5).
54

In D. fragrans 'Massangeana' , the percentage of cuttings with shoots decreased
with increased dosage. Shoot fonnation in the treatments was slightly slower than root
formation for this variety (Figures 4 and 5).
The percentage of cuttings fonning shoots followed a similar trend to root
fonnation for D. xmassejJiana in that the 5 Gy and lOGy treatments fonned shoots
before the control and 20 Gy treatments (Figures 4 and 5). By the end of the experiment,
the lOGy treatment had the highest percentage of cuttings with shoots, followed by the 5
Gy, the control, 30 Gy, and 20 Gy treatments. The percentage of cuttings with shoots
was the lowest in the 40 Gy treatment and no cuttings fonned shoots in the 50 Gy
treatment. Shoot formation was slower than root fonnation during the first 8 weeks of
the experiment in D. xmassejJiana, but were approximately equal by the end of data
collection. At dosages of 20 Gy and above, the percentages of cuttings with shoots at the
end of the experiment were not nearly as large as the percentage of cuttings with roots.
For each variety, the percentage of cuttings with roots was plotted against the
dosage of irradiation (Figure 6). Linear regressions were calculated and tested for
significance (Table 7). For all varieties, regression was significant with P < 0.01. The
same procedures were applied to the shoot fonnation percentages (Figure 7). All
regression lines for shoot formation were significant with P < 0.01 (Table 8). The linear
regressions were used to estimate the LDso and LD)oo values for both root and shoot
formation (Table 9). The LDso values for root fonnation were 14.6, 13.8,5.7 and 17.6 Gy
for D. deremensis 'Santa Rosa', D. fragrans 'Massangeana', D. fragrans 'Victoriae', and
D. xmassejJiana, respectively. The LDso values for shoot formation were 19.7, 16.6,
55

22.1, and 10.9 Gy, D. deremensis 'Santa Rosa', D. fragrans 'Massangeana', D. fragrans
'Victoriae', and D. xmasseffiana, respectively.
The total number of shoots for each treatment and the number of shoots showing
mutations were recorded (Table 10). D. fragrans 'Massangeana' had the most mutated
shoots, with a total of 41, followed closely by D.fragrans 'Victoriae' with 40. D.
deremensis 'Santa Rosa' had 23 mutated shoots while D. xmasseffiana had only 5
mutated shoots. To determine if the mutation rate was significantly associated with the
irradiation dosage, 2 x 2 contingency tables were used to calculate chi-square values. For
D. deremensis 'Santa Rosa', the numbers of mutations in each treatment were all
significantly different from each other. For D.fragrans 'Massangeana' and D. fragrans
'Victoriae', the numbers of mutations in the 5 Gy and lOGy treatments were not
significantly different from each other, but were different from the control and 20 Gy
treatments (Figure 8). There were two classes of mutations observed - chlorophyll and
morphological. The mutations exhibited by the developed shoots were classified by type
of mutation (Table 10). Chlorophyll mutations were further separated into three
categories - albino (no chlorophyll), viridis (no variegation - all green) and other.
Chlorophyll mutations were the most numerous, mostly of the viridis type.
Discussion and Conclusions
The dosages (50 to 500 Gy) used in the first round of irradiation were too high
and none of the treated cuttings formed roots or shoots after the treatments. Close to
100% of the control treatments rooted and formed shoots for all varieties treated, showing
that the failure to root and shoot was due to the irradiation treatments. Numerous
publications about the irradiation of ornamentals have clearly shown that survival and
56

regeneration capacity decrease with increasing irradiation dosage (F AO and IAEA,
1977). The results of the second round of irradiation using dosages ranging from 5 to 50
Gy are generally in agreement as there was a decrease in the percentage of cuttings
forming roots and shoots with increased dosage of gamma irradiation. There was also an
increase in the percentage of mutated shoots with increased radiation (up to the point
when shoot formation was inhibited), which is also consistent with previous reports.
The LDso estimations for the four varieties irradiated ranged from 5.7 to 17.6 Gy
for rooting, with the order of sensitivity to radiation from highest to lowest being D.
/ragrans 'Victoriae' > D. /ragrans 'Massangeana' > D. deremensis 'Santa Rosa' >
D. xmasseffiana (Table 9). The LDso estimations for shoot formation ranged from 10.9 to
22.1 Gy, and the order of sensitivity was D. xmasseffiana > D. /ragrans 'Massangeana'
> D. tieremensis 'Santa Rosa' > D. /ragrans 'Victoriae' (Table 9). Root formation and
shoot formation sensitivity to radiation differ, even for the same variety. The LDso for D.
/ragrans 'Victoriae' shoot formation was almost 15 Gy higher than that for root
formation. Conversely, the LDso of D. xmasseffiana rooting was almost 7 Gy higher
than that for shoot formation. The radiosensitivities of different plants have been shown
to correlate with what has been termed average interphase chromosome volume, the
average volume occupied by each chromosome in the nucleus of a cell during interphase
(Sparrow et aI., 1963). This number is calculated by dividing the nuclear volume by the
number of chromosomes, but is not a very useful indicator of radiosensitivity in our
studies because the varieties studied have similar genetic constitutions and the difference
between chromosome numbers is at most one. Flow cytometry performed on the four
varieties also indicated similar DNA contents. The different sensitivities are likely due to
57

other factors such as the age of the plant, length of mitotic cycle, percentage of cells
dividing, concentration of growth hormones, and nutritional state (Sparrow et aI., 1968).
It is difficult to speculate what caused the chlorophyll and morphological
mutations and the changes in the ability to form roots and shoots in the irradiated
cuttings. The exact mechanisms and causes of the effects of radiation are still in question
despite the fact that the end results of radiation on plants have been well studied and
documented since irradiation of plants first gained interest almost a century ago. The
number of crops in which researchers know which genes control which characteristics is
relatively small, and ornamentals are studied to an even lesser extent than major
agricultural crops such as maize and rice. In addition, with molecular techniques now at
the forefront of genetic research, irradiation research has taken a backseat. It is known
that the major effect of ionizing radiation such as gamma rays is caused by its ability to
rupture chemical bonds, and the produced effect is proportional to the energy absorbed in
the tissue irradiated. Gamma rays can penetrate many centimeters into tissue which is
more than x-rays and beta particles can penetrate (Sparrow, 1961). When radiation
penetrates tissue, it loses energy by interacting with atoms and molecules and the energy
is dissipated along the track of the particle. The results are random changes in the
nuclear DNA or cytoplasmic organelles, producing gene, chromosomal or genomic
mutations. In comparison to chemical mutagens, radiation mutation produces more
chromosomal aberrations and rearrangements (Donini and Sonnino, 1998). Many
theories have been offered to explain specific irradiation effects. Nybom (1961) attributed
most induced somatic changes to loss or duplication of genetic material. Some
researchers claim that abnormalities are due to physiological disturbances rather than
58

mutation (Fereol et al., 1996). Disturbances in auxin synthesis or mineral metabolism
and amino acid accumulation have also'been offered as explanations for specific
mutations. Casarett (1968) specifically attributes root growth inhibition to the disruption
of auxin synthesis and cell division.
The percent of mutated shoots ranged from 0% in some treatments to as high as
93.3% in the 10 Oy treated D. fragrans 'Victoriae'. The greatest number of mutations
genera11y occurred at dosages in the range of the estimated LDso values for each variety.
The majority of mutations observed in these experiments were chlorophyll mutations
(Table 10). Specific mutations observed included viridis (green) and albino (colorless -
white or very pale yellow) types. Viridis mutations were the overwhelming majority with
albino following far behind. Albino types of mutations are not useful as the shoot
genera11y dies after several weeks because of the lack of chlorophyll. Viridis mutations
are also not desirable because they resemble wild types and are less attractive than
variegated cultivars. These types of chlorophyll mutations could be due to a disruption in
the chlorophyll synthesis or mineral metabolism pathways (Rao, 1999). The most
interesting chlorophyll mutations obtained were those that had variegation different from
that normally observed in the species (Figure 9). For example, several mutated leaves of
D. fragrans 'Massangeana' were half yellow and half green. These shoots may be
chlorophyll chimeras and may not be stable. They wiU continue to be monitored, and if
they are stable mutants, their potential as new cultivars can be evaluated. The large
number of mutations obtained is encouraging for future research involving induced
mutations using gamma ray irradiation in Dracaena. It confirms that irradiation can
indeed give rise to different phenotypes. Also, since the mutations mostly occurred in the
59

LDso range, future experiments with larger numbers of cuttings treated at that dosage will
probably induce large numbers of mutations.
One exception to the trend of decreasing root and shoot formation with increased
dosage occurred in D. xmasseffiana. Our results showed that the highest root and shoot
formation occurred in the lOGy treatment followed by the 5 Gy treatment (Figures 6, 7,
and 10). This phenomenon has been well documented and is called hormesis, a term
coined in 1942 to describe the stimulation of fungi by a naturally occurring antibiotic
which at higher concentrations suppresses fungal growth (Sagan, 1987). Hormesis is
defined as a concept that describes the nature of dose-response relationships in biological
systems as displaying a stimulatory response at low doses and an inhibitory response at
higher doses (Calabrese and Baldwin, 2000). Numerous studies have documented cases
in which growth was stimulated in plants when exposed to low doses of radiation (Skok
et al., 1965). Reported effects include taller plants, greater numbers of flowers, fruits,
and stems, longer roots, faster germination, increased leaf thickness, and earlier
flowering. For example, Gupta et al. (1982) found that at 15 Gy, Costus speciosus
showed an increase in rhizome yield and equal or higher levels of a useful chemical,
diosgenin, than the controls. Miller and Miller (1987) reviewed over 200 articles
claiming hormetic effects in plants. Skok et al (1965) conducted a study specifically to
examine these stimulatory effects in plants, but concluded that although sma1l but
significant growth increases were produced in certain instances, these results were not
always reproducible. Despite the numerous reports ofhormesis, the phenomenon has
largely been ignored or discredited in the scientific community because many influential
scientists were strong opponents, citing weak studies with inadequate sample sizes and
60

poor replication of findings. Thus, studies tapered off and there is no true understanding
of the mechanism of the response (Calabrese and Baldwin, 2000; Miller and Miller,
1987). One of the explanations offered for the cause of this stimulatory effect is that the
radiation disturbs the activity of certain enzymes involved in the synthesis of auxin
(Casarett, 1968). Ehrenberg (1955) suggested that low doses of radiation destroys an
inhibitory condition in the plant, allowing the subsequent increase in growth. However,
these are all theories which have not been expanded on, and the phenomenon remains
largely unexplained.
In our experiment, only D. xmasseffiana exhibited a hormetic response in the
form of higher percentages of cuttings forming roots and shoots at low dosages of
radiation. It would be interesting for future research to determine if the effect obtained in
this experiment is reproducible or occurs in other varieties of Dracaena. The practical
uses for these results would be limited, though, because most commercially grown
varieties of Dracaena have very high rooting percentages. In addition, D. xmasseffiana
is not a widely grown variety, so a stimulatory effect for rooting would not have much
commercial use.
The estimation of the LDso values for the root and shoot formation of the four
varieties of Dracaena treated in this experiment could aid in the development of new
varieties (Table 9). Large numbers of cuttings could now be irradiated at the LDso
dosages to induce desirable mutations. In addition, the LDso dosages provide a useful
estimate for optimal irradiation dosages of other species and varieties of Dracaena not
used in this experiment.
61
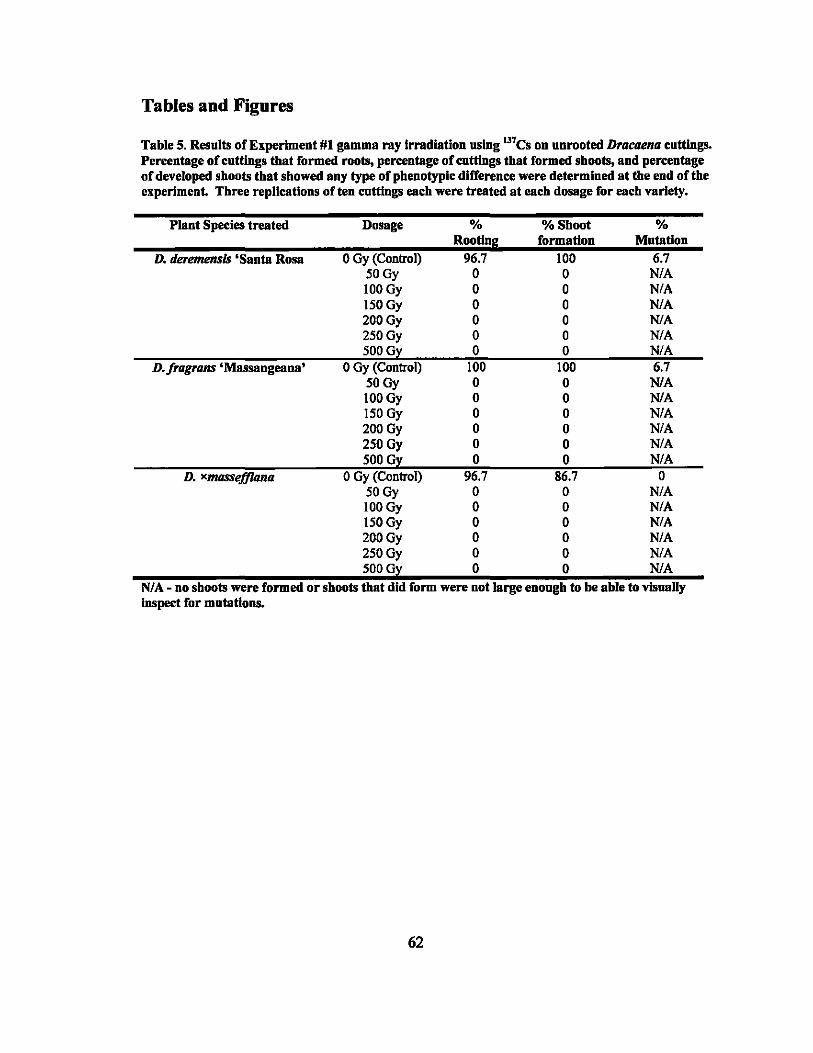
Tables and Figures
Table 5. Results of Experiment #1 gamma ray Irradiation using Il7Cs on nnrooted Dracaena cuttings. Percentage of cnttings tbat formed roots, percentage of CIIttings that formed shoots, and percentage of developed shoots that showed any type of phenotypic difference were determined at the end of the experiment. Three replications of ten cuttings each were treated at each dosage for each variety.
Plant Species treated Dosage % % Shoot % Rooting formation Mutation
D. tleremensis 'Santa Rosa 0 Gy (Control) 96.7 100 6.7 ~~ 0 0 ~ 100~ 0 0 ~ I~~ 0 0 ~ 200Gy 0 0 N/A 250Gy 0 0 N/A 500Gy 0 0 N/A
D.fragrans'Massangeana' 0 Gy (Control) 100 100 6.7 ~~ 0 0 ~ 100Gy 0 0 NlA 150 Gy 0 0 N/A 200 Gy 0 0 N/A 250 Gy 0 0 N/A 500 Gy 0 0 N/A
D. xl1IIJSSef.flona 0 Gy (Control) 96.7 86.7 0 ~~ 0 0 ~ 100~ 0 0 N/A ISOGy 0 0 N/A 200Gy 0 0 N/A 250Gy 0 0 N/A SOOGy 0 0 N/A
NI A - no shoots were formed or shoots that did form were not large enough to be able to visually luspect for mutations.
62
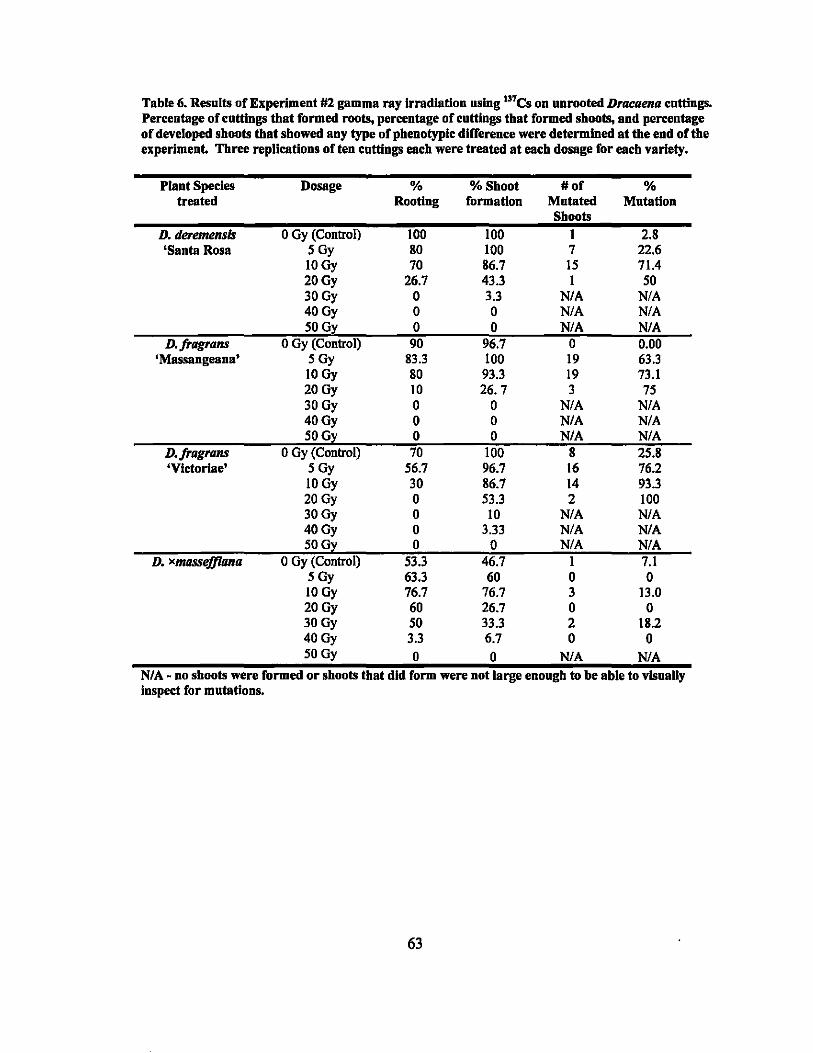
Table 6. Results of Experlmeut #2 gamma ray Irradlatiou uslug l37es on unrooted DrflCllena cuttiugs. Percentage of cuttiugs tbat formed roots, percentage of cnttiugs tbat formed sboots, and percentage of developed sboots tbat sbowed any type of pbenotyplc difference were determlued at tbe end of tbe experiment. Three replications of ten cuttiugs eacb were treated at eacb dosage for each variety.
Plant Species Dosage % %Sboot #of % treated Rooting formation Mutated Mutation
Sboots D. deretlll!nsis o Gy (Control) 100 100 I 2.8 'Santa Rosa 5Gy 80 100 7 22.6
lOGy 70 86.7 15 71.4 20Gy 26.7 43.3 I 50 30Gy 0 3.3 N/A N/A 40Gy 0 0 N/A N/A 50G:l: 0 0 N/A N/A
D·fragrans o Gy (Control) 90 96.7 0 0.00 'Massangeana' 5Gy 83.3 100 19 63.3
10 Gy 80 93.3 19 73.1 20Gy 10 26. 7 3 75 30 Gy 0 0 N/A N/A 40Gy 0 0 N/A N/A 50G~ 0 0 N/A N/A
D·fragrans o Gy (Control) 70 100 8 25.8 'Vlctorlae' 5Gy 56.7 96.7 16 76.2
lOGy 30 86.7 14 93.3 20Gy 0 53.3 2 100 30Gy 0 10 N/A N/A 40Gy 0 3.33 N/A N/A 50G~ 0 0 N/A N/A
D. xlIIIlSSejJlana o Gy (Control) 53.3 46.7 I 7.1 5Gy 63.3 60 0 0 lOGy 76.7 76.7 3 13.0 20Gy 60 26.7 0 0 30Gy 50 33.3 2 182 40Gy 3.3 6.7 0 0 50Gy 0 0 N/A NlA
NI A - no sboots were formed or sboots tbat did form were not large enougb to be able to vlsuaUy Inspeet for mutations.
63
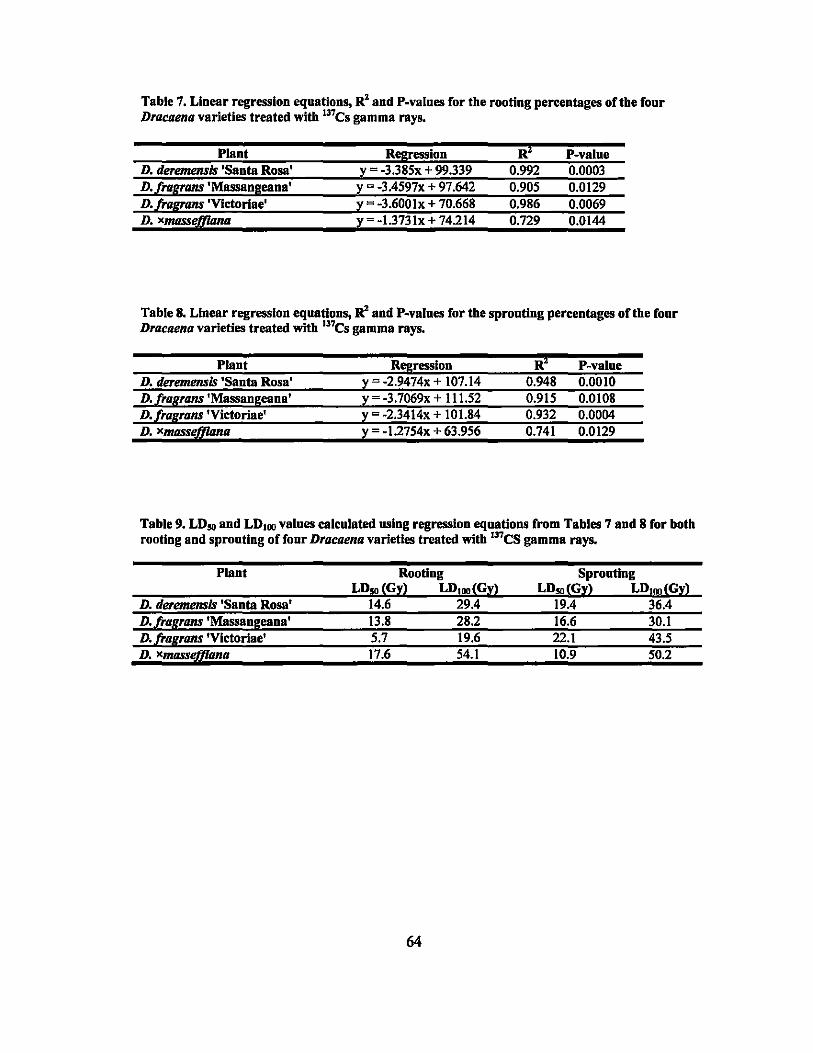
Table 7. Linear regression equations, RZ and P-values for the rooting percentages of the fonr Dracaena varieties treated witb 137CS gamma rays.
Plant Regression Ri P-value D. deremensls 'Santa Rosa' y -3.385x + 99.339 0.992 0.0003 D./ragrans'Massangeana' y - -3.4597x + 97.642 0.905 0.0129 D. /rarrans 'Victoriae' y = -3.600Ix + 70.668 0.986 0.0069 D. xmasseJ[il1na y -1.3731x + 74.214 0.729 0.0144
Table 8. Linear regression equations, RZ and P-valnes for tbe spronting percentages of tbe fonr Dracaena varieties treated with I37CS gamma rays.
Plant Regression Ri P-va1ue D. deremensls 'Santa Rosa' y - -2.9474x + 107.14 0.948 0.0010 D. /ragrans 'Massangeana' y -3.7069x + 111.52 0.915 0.0108 D. /ragrans 'Vlctoriae' y - -2.3414x + 101.84 0.932 0.0004 D. xlI/IJSSeJ[il1na y - -1.2754x + 63.956 0.741 0.0129
Table 9. LD .. and LDIIMl valnes calculated using regression equatious from Tables 7 and 8 for botb rooting and sprouting of fonr Dracaena varieties treated with Il7CS gamma rays.
Plant Rooting Sprouting LD .. ~Gll LDIIMl~Gll LD .. ~Gl~ LDIIMl~Gll
D. deremensls 'Santa Rosa' 14.6 29.4 19.4 36.4 D.l!.aflrans'Massangeana' 13.8 28.2 16.6 30.1 D. f!.aflrans'Vlctoriae' 5.7 19.6 22.1 43.5 D. xlI/IJSSt1JfIma 17.6 54.1 10.9 50.2
64
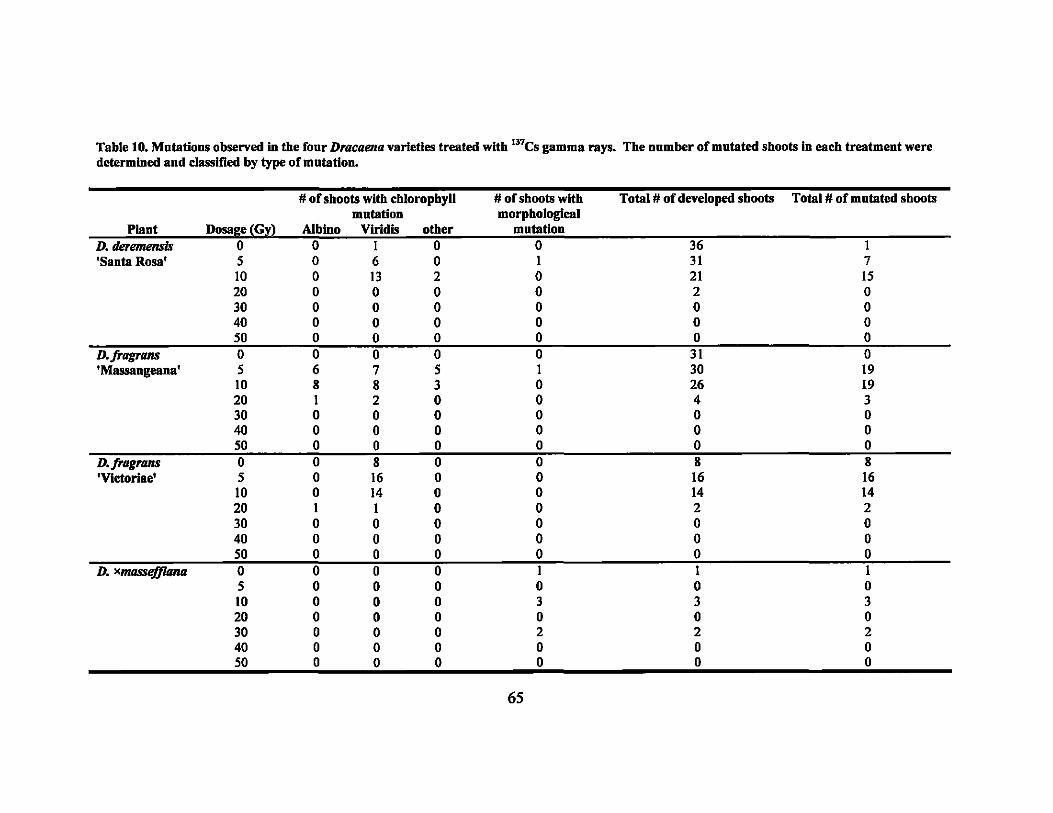
Table 10. Mutatious observed in the four D,acaena varieties treated wltb 137CS gamma rays. Tbe number of mutated sboots in eacb treatment were determined aud classified by type of mutation.
# of sboots with cbloropbyU # of sboots witb Total # of developed sboots Total # of mutated sboots mutation morpbological
Plant DosaGe~Gl) Albino Viridis otber mutation D.deremensis 0 0 1 0 0 36 1 'Santa Rosa' 5 0 6 0 1 31 7
10 0 13 2 0 21 15 20 0 0 0 0 2 0 30 0 0 0 0 0 0 40 0 0 0 0 0 0 50 0 0 0 0 0 0
D·fragrans 0 0 0 0 0 31 0 'Massangeana' 5 6 7 5 I 30 19
10 8 8 3 0 26 19 20 1 2 0 0 4 3 30 0 0 0 0 0 0 40 0 0 0 0 0 0 50 0 0 0 0 0 0
D·frag,ans 0 0 8 0 0 8 8 'Vlctorlae' 5 0 16 0 0 16 16
\0 0 14 0 0 14 14 20 I 1 0 0 2 2 30 0 0 0 0 0 0 40 0 0 0 0 0 0 50 0 0 0 0 0 0
D. Xt1IIJSSejJUuuI 0 0 0 0 I I 1 5 0 0 0 0 0 0 \0 0 0 0 3 3 3 20 0 0 0 0 0 0 30 0 0 0 2 2 2 40 0 0 0 0 0 0 50 0 0 0 0 0 0
65
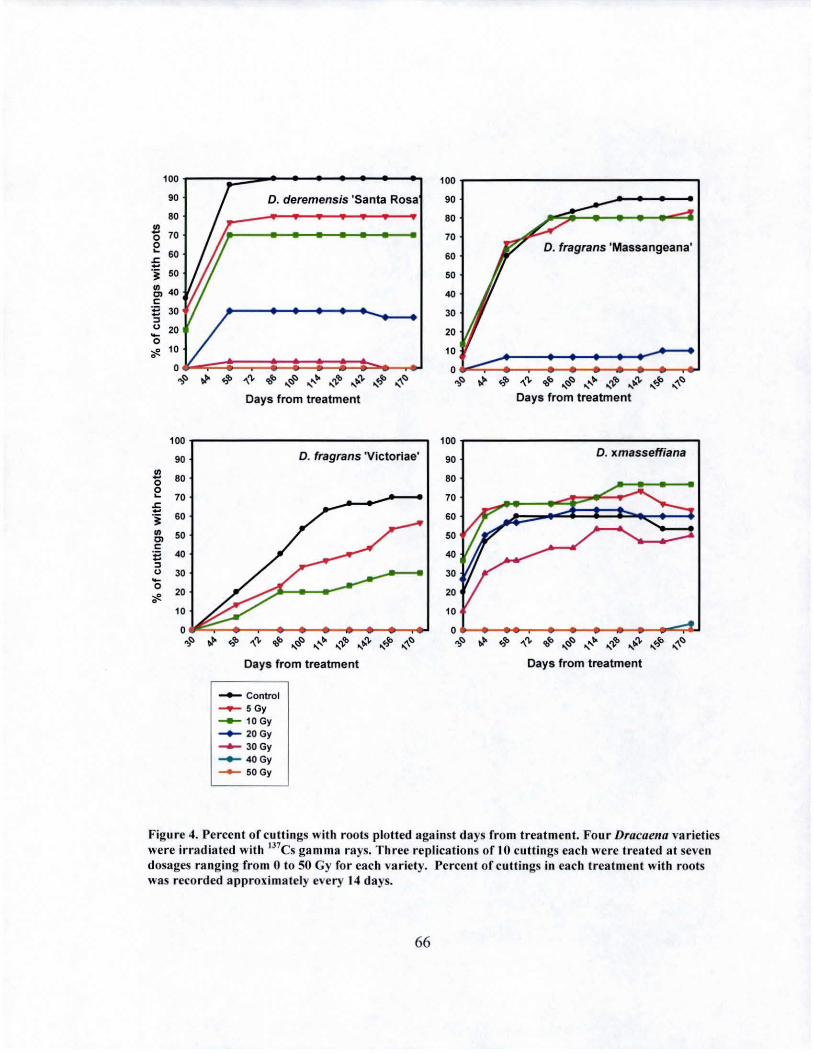
100r---~===-"~~~-4--~~--"
90
80 !l o 70 ~
'" -60
.§ 50
~ 40 c: E 30 ::l
~ 20 o
D. deremensis 'Santa Rosa
';!. 10
~::::=;::::::::::==:::::--....I
100
90
!l 80 0 0 ~ 70 .t: -.~ 60 .,
50 '" c: 'il 40 ::l
" 30 -0 20
~ • 10
0 ~~
~ ~ '\t'L. 'bra ... ~t:J ...... '00 ,,'),.#b ,,'I;.'),. ... +,1'0 ~t::J
Days from treatment
D. fragrans 'Victoriae'
.,. 4' "", COlO ... t::J~ ....... bo ... ~ ... '1;." ... ,,'0 ~~
Days from treatment
--- Control ......- SGy -..- 10 Gy ~ 20Gy ___ 30Gy ___ 40Gy
--- 50Gy
100r----------------------------,
90
80
70
60
50
40
30
20
D. fragrans 'Massangeana'
': ~~::::::::~~~::::::::::::=::J ~t::J .» ':Jib "\'\. t:t;,fo ,,()f::) ........ ...~ .... "" .... ~ ~()
Days from treatment
100 r---------------.., 90
80
70
60
50
40
30
20
D. xmasseffiana
10
o ~._~~~~._~_.~~~~
,.,(;) ~ 4' '\'\. 'bra ... ()() ....... tt. ... ",'b ...... '" "tJ' ~()
Days from treatment
Figure 4. Percent of cuttings with roots plotted against days from treatment. Four Dracaellu varieties were irradiated with 1J7CS ga mm a rays. Three replications of 10 cuttings each were treated at seven dosages ranging from 0 to 50 Gy for each variety. Percent of cuttings in each treatment with roots was recorded approximately every 14 days.
66
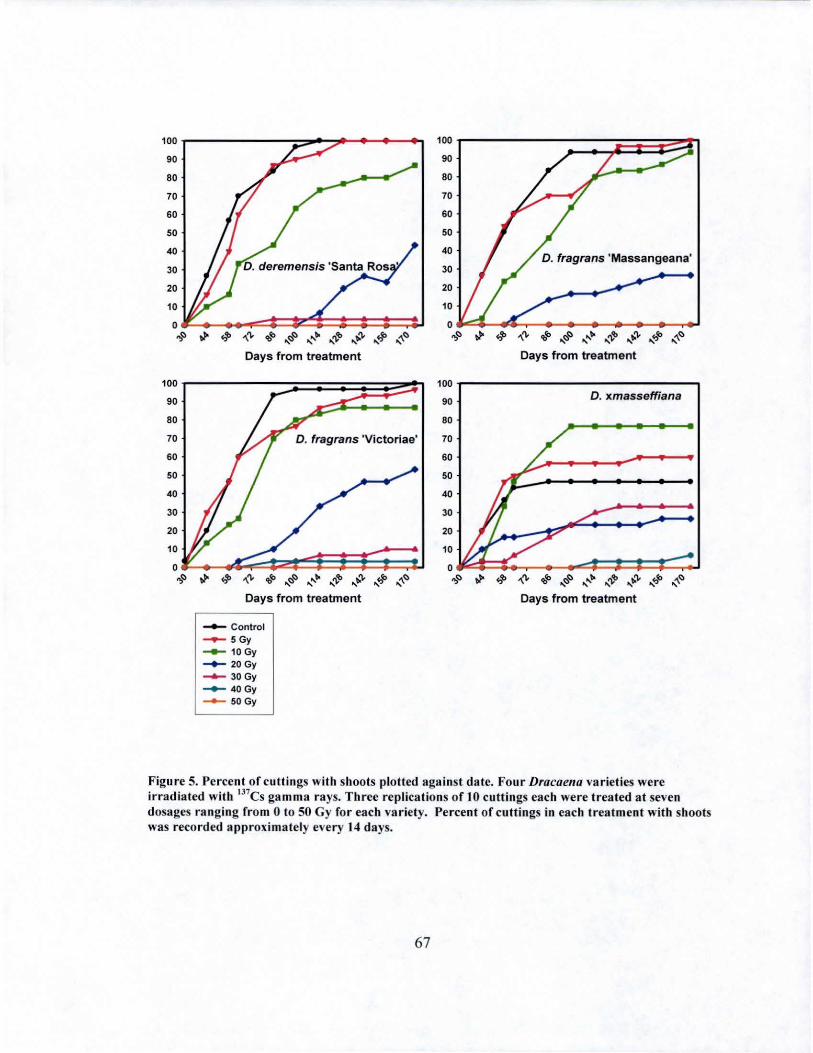
100 100
90 90
80 80
70 70
60 60
50 50
40 40
30 30
20 20
10 10
0
.". 4> ~'" .... ....~() "' .... ~ .... "'fb ~'" .... /P ~() .,~ .". 4> ~'" 'bt'o .... r:§' .... ,,~ ,~ .'" , .," , ~~ , Days from treatment Days f rom treatment
100 100
90 90 D. xmasseffiana
80 80
70 70
60 60
'0 50
40 40
30 30
20 20
10 10
0 .". 4> ~'" 'bfo .... ~() ........ bo ,,,," ~'" , .... t!P .... '\.() .,~ .". 4> ~'" 'bro .... r:::,() ........ fI< .... "''b ...... '" .," , ~~ ,
Days from treatment Days from treatment
___ Control
.......- 5Gy ___ 10 Gy
-.- 20Gy ...... 30Gy ___ 40Gy
--- 50 Gy
Figure 5. Percent of cuttings wit h shoots plotted against date. Four Dracaella varieties were irradiated with 1l7Cs gamma rays. Three replications of 10 cuttings each were treated at seven dosages ranging from 0 to 50 Gy for each variety. Percent of cuttings in each t reatment with shoots was recorded approximately every 14 days.
67
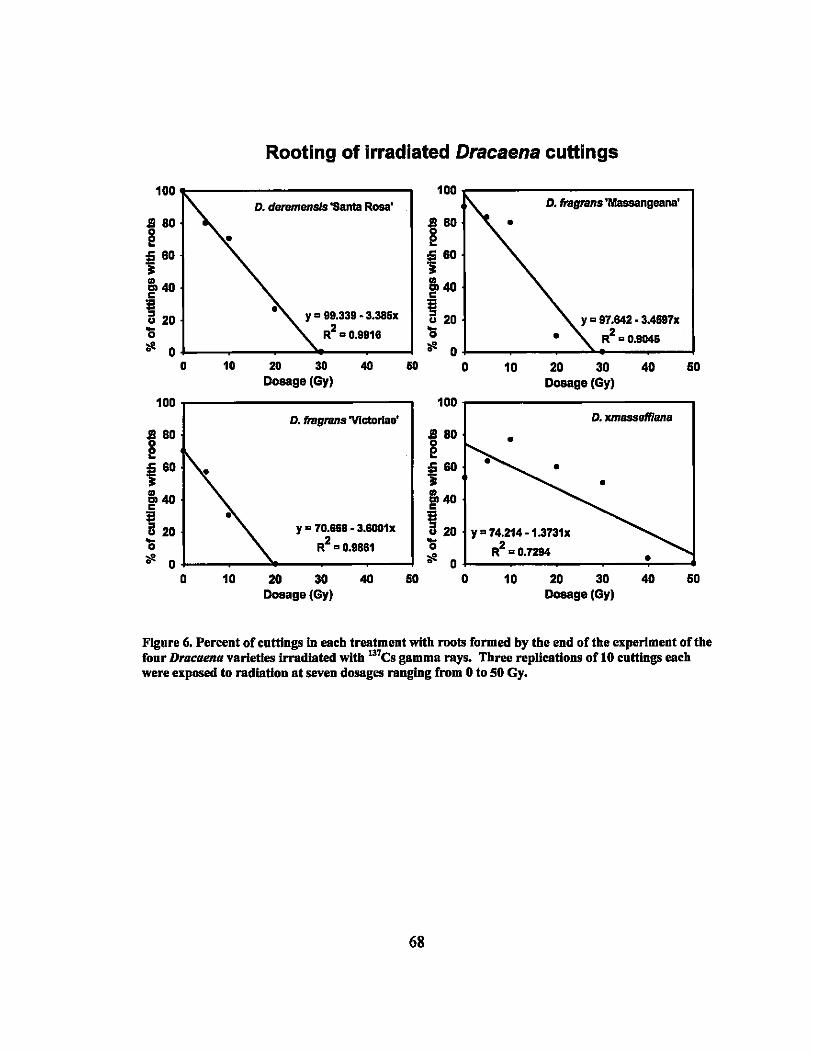
Rooting of irradiated Dracaena cuttings
100 100 D. dBtBnlensis 'Santa Rosa' D. fragrans'Massangeana'
.$!I 80
~ 180 •
~ aD ~ 60
&40 c &40 c
~ 20 Y = 99.339 -3.385x ~ 20 Y = 97.842 • 3.4997x '0 R2 = 0.9916 '0 • R2 = 0.9046 :.!! 0 :.!! 0
0 10 20 30 40 &0 0 10 20 30 40 50 Dosage(Gy) Dosage (Gy)
100 100 D. fragrans'VIctorias' D. xmesseff/ena
.$!I 80
~ 180 •
~ 60 ~60 • •
&40 c ~4O
~ 20 Y a 70.668 • 3.5001x ~ 20 ya 74.214 -1.3731x '0 R2 a 0.9661 '0 2 :.!! :.!! R =0.7284 • 0
10 20 30 40 50 0 10 20 30 40 50 Dosage (Gy) Dosage (Gy)
Figure 6, Percent of cnttings In each treatment with roots formed by the end of the experiment of the four Dracaena varieties irradiated with 13'es gamma rays. Three replications of 10 cuttings each were exposed to radiation at seven dosages ranging from 0 to 50 Gy.
68
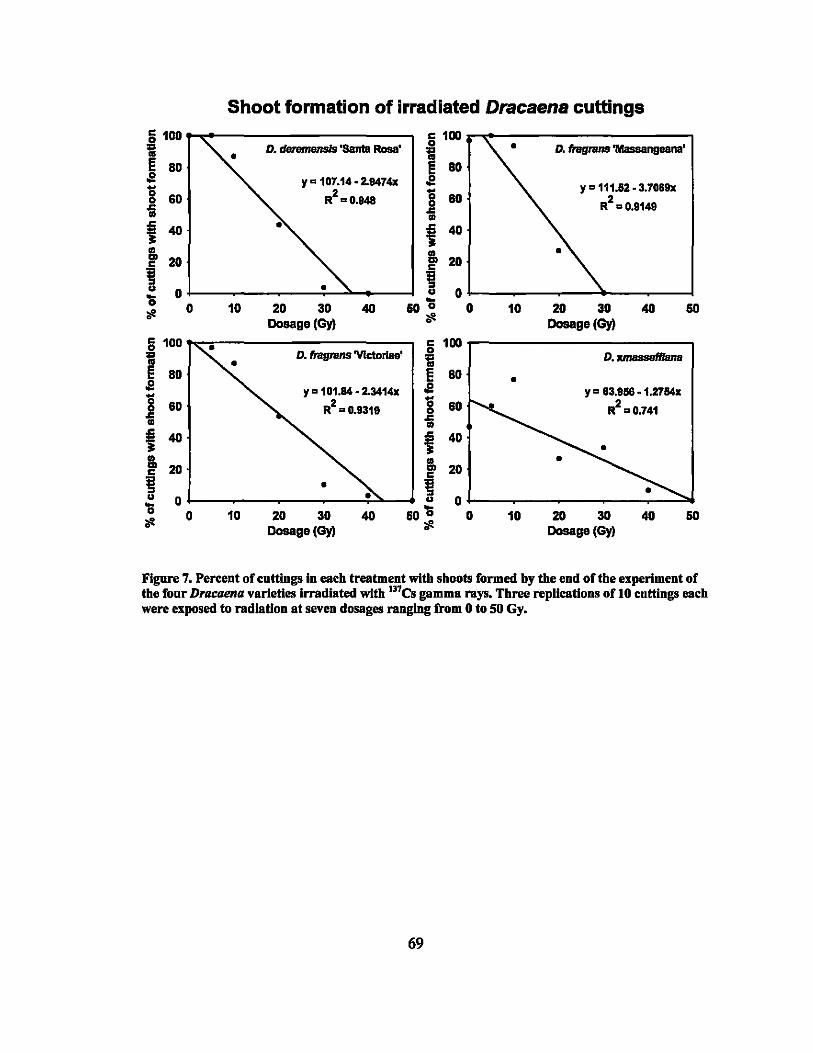
Shoot formation of irradiated Dracaena cuttings
~ 100 lji100 D. _ 'Santa Rosa' 0 D. fragnms'Masaangeana'
0 i ~ 80 80 .e Y" 107.14·2.9474x .e g R2" 0.848 g Y " 111.62 • 3.70&9x 60 60 2
.c .c R "0.9149 CD CD
t 40 t 40
~ 20 I!L 20 c ~ ~ .. 0 .. 0 '0 ... 111 0 10 20 30 40 60 0 0 10 20 30 40 60
Dosage (Gy) 111 Dosage(Gy) c 100 c 100 0 0 l: D. fragnms'VIctoriae' l: D. xmssseIllsna
~ 80 ~ 80 0 .e Y" 101.94· 2.3414x .e Y" 83.958 ·l.2794x g 60 2 g 60 2 R "0.9319 R "0.741 .c .c CD CD
t 40 t 40 CD I!L 0 ... 20 20 c c ~ 0 ~ .. 0 .. 0 '0 ... 111 0 10 20 30 40 60 0 0 10 20 30 40 60
Dosage(Gy) 111 Dosage (Gy)
Figure 7, Percent of cnttlngs In each treatment with shoots formed by the end of the experiment of the foor D,acaena varieties irradiated with l31es gumma rays, Three replications of 10 cuttings each were exposed to radiation at seven dosages ranging from 0 to 50 Gy,
69
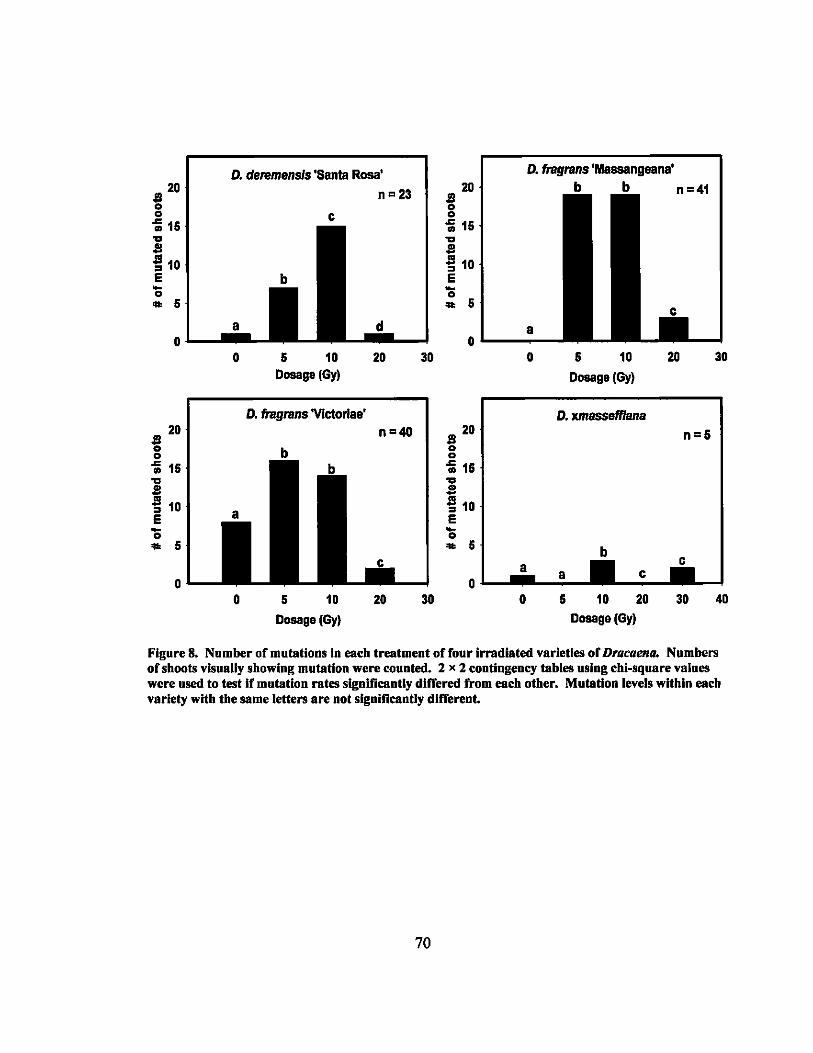
~ 20 o -; 16 ... S ~ 10 E -o .. 6
D. deremens/s 'Santa Rosa'
n=23
c
a o.L.--_.-
~ 20 o -; 15
~ ~ 10 E ~ .. 5
o
o
o
5 10 Dosage (Gy)
D. fragrans 'VIctorias'
5 10
Dosage (Gy)
20
n=4O
20
30
30
D. fragrans 'Massangeana'
20 ~ o -; 16 ... S ~ 10 E ~ .. 6
a 0'-----
~ 20 o -; 16 ... S ~ 10 E
'It 5
o & 10
Dosage(Gy)
D. xmassefflana
a OL-_ ..... _ a ..... _
o 5 10 20 Dosage(Gy)
20
n e &
30
30
40
Figure 8. Number of mutations in each treatment of four irradiated varieties of Dracaena. Numbers of shoots visually showing mutation were counted. 2 x 2 contingency tahies using chi-squnre vaiues were used to test if mutation rates significantly differed from each other. Mutation levels within each variety with the same letters are not significantly dlffereut.
70

Normal D.fral!rans 'Massangeana'
D. fral!rans 'Massangeana' chlorophyll mutants
Figure 9. Comparison of normal D./ragrtIllS 'Massa ngeana ' shoots and chlorophyll mutants obtained from irradiation of unrooted cuttings with 137CS ga mma rays
71

Control
Figure 10. Irradiated D. x masse/floI/o cuttings showing hormetic erfects. Larger percentages of cuttings in 5 Gy and lOGy treatmcnts formed roots and shoots than in the control treatment.
72

CHAPTER 6. POL VPLOIDIZATION
Introduction
Polyploid induction is a useful nontraditional breeding method because polyploid
forms of plants are known to have desirable characteristics such as being more robust and
sturdy, having thicker leaves and stems, a deeper green color, an increased width-to
length ratio of leaves, larger flowers, a longer or later flowering period, or a more
compact growth habit (Gao et al., 1996).
A variety of methods can be used to induce polyploids. Mutagens that have
shown effectiveness include colchicine and oryzalin, with oryzalin effective at much
lower concentrations and less phytotoxic. Accordingly, it has recently gained popularity
in polyploid induction experiments. Many plant parts have been treated with these
compounds to induce polyploids, but treatments of newly forming buds or of callus in
tissue culture seem to be the most effective methods.
Oryzalin is clearly an effective chemical when the aim is to double chromosome
number, but most studies test effectiveness by using wide ranges of concentrations, an
impractical method unless a large amount of plant material is available for the
experiment. If large amounts are unavailable, it is best to use concentrations that have
been successful on closely related plants. An experiment on Lilium longijlorum tested
the effect of oryzalin at concentrations of 0.001 % to 0.005% to treat in vitro cultures.
The optimal concentration was found to be 0.003%, which induced over 60% polyploids
with minimal phytotoxicity (Takamura et al., 2002). Another study on Lilium
longijlorum involving in vitro chromosome doubling used concentrations of oryzalin of
73

0.005% and 0,01 % and was able to restore fertility to a sterile hybrid by inducing
polyploidy (Tuyl et aI., I 992). No studies attempting to double the chromosome number
of any Dracaena species were found in publication. Therefore, studies on related plants
such as Lilium were used as guidelines for conducting the experiments on Dracaena. The
objective of this experiment is to produce tetraploid plants of Dracaena species using
oryzalin both in vivo by treating buds and in vitro by treating callus tissue.
Materials and Methods
Experiment #1 - In vivo treatment
Field grown plants of D. deremensis 'Santa Rosa' approximately 1 to 2 m tall at
the Waimanalo Experimental Station of the University of Hawaii were used. The tips of
the plants were cut off so that plants were approximately 0.6 m tall. When the new buds
reached 1 cm in size, the tip of the bud was cut off and the oryzalin solution applied
(Figure II). A cotton ball soaked in the solution was placed over the bud to ensure
continued exposure to the chemical and the entire area was covered in plastic wrap. After
the duration of the treatment (24 or 48 h), the plastic wrap and cotton ball were removed,
and the bud was rinsed with water to remove any lingering solution. On each plant, two
buds were treated and the experiment was replicated three times.
A completely randomized design was set up with a 3 x 2 factorial treatment
design for a total of six treatments. The treatments consisted of three concentrations of
oryzalin at two different durations and the experiment was replicated three times.
l. Control, 24 hrs 2. Control, 48 hrs 3. 0.005% oryzalin, 24 hrs 4. 0.005% oryza1in, 48 hrs 5.0.01% oryzalin, 24 hrs
74

6.0.01% oryzalin, 48 hrs
Controls consisted of only the carrier solution ofDMSO (dimethylsulfoxide) and water.
The oryzalin (phytotechnology Laboratories, Shawnee Mission, KS) solution was
prepared by dissolving in DMSO and then diluting with distilled water to the desired
volume with the final solution being 2% DMSO.
The number of buds regenerated and observations on abnormal growth
morphology were recorded. After 2 to 3 months of additional growth, flow cytometry
was used to determine ploidy with the Cystain PI Absolute P (partec, MUnster, Germany)
kit on leaf samples. A 1 cm2 piece of tissue from a new leaf was chopped with a double
edged razor blade (Gillette Super Stainless Super Inoxydable Blades, Proctor and
Gamble, Cincinnati, OII) for 30 to 60 s in 500J.1L of ice-cold nuclei extraction buffer and
incubated for another 30 to 60 s. It is essential that the blade used is sharp to obtain good
samples. Each edge on the blade was used for two samples and then discarded. The
suspension was then poured through a CellTrics 50 J.1m nylon filter (partee, MUnster,
Germany) to remove the large pieces of tissue. Two mL of ice cold staining buffer
containing propidium iodide stain was then added and the sample was incubated on ice
for at least 1 h. The analysis was done using a Beckman-Coulter Altra flow cytometer
(Miami, Florida) using the 488 nm line of a Coherent I90C argon ion laser set at 200
m W. The linear, log and peak fluorescence signals of the propidium iodide stained nuclei
were collected, along with forward and side scatter signals. At least 5000 nuclei were
analyzed for each sample. Control samples of untreated plants were regularly run during
analyses to check the stability of the peak position of the diploid level. Histograms of
75

linear DNA fluorescence were analyzed using FlowJo (v. 6.3.4, Treestar Inc.), and the
mean fluorescence and coefficient of variation for each peak were calculated.
Experiment #2 - In vitro treatment
D. deremensis 'Lisa' shoot tip explants were placed onto callus-inducing medium
(MS basal medium supplemented with 30 gIL sucrose, 3 gIL gellan gum, 0.5 mgIL 2,4 D,
and 15% (v/v) coconut water adjusted to pH 6.0). Explants were approximately two to
three em tall and were obtained from Alpha Genesis laboratories in Hilo, Hawaii. The
formulation of the medium was modified and adapted from Chua et al. (1981). Cultures
were grown in 16 h of light at 26 ±1 °C, and spherical yellowish callus was observed on
the base of stems after 4 to 8 weeks (Figure 12). When calli reached approximately 1 cm
in diameter, they were separated from the shoots and subcultured on the same medium
for 16 weeks, but placed in the dark to multiply. After a sufficient number of calli was
obtained they were placed in the oryzalln solution for treatment
A completely randomized design was set up with a 3 x 2 factorial treatment
design. The treatments consisted of three concentrations of oryzalin at two different
durations and the experiment was replicated three times.
1. Control, 24 hrs 2. Control, 48 hrs 3. 0.005% oryzalin, 24 hrs 4. 0.005% oryzalin, 48 hrs 5.0.01% oryzalin, 24 hrs 6. 0.01% oryzalin, 48 hrs
Oryzalin dissolved in DMSO was added to the callus inducing medium without agar to a
final volume of2% DMSO and filter sterilized. Controls consisted of only the carrier
solution ofDMSO and the liquid callus inducing medium. The callus pieces were placed
76

in the solutions for the specified durations and then rinsed in sterile water. After
treatment, calli were subcultured for 10 weeks on the same medium and then moved onto
shoot inducing medium (MS basal medium supplemented with 30 gIL sucrose, 3 gIL
gellan gum, 1 mgIL kinetin, and 15% (v/v) coconut water adjusted to pH 6.0). After 4 to
5 weeks on this medium the calli showed no signs of shoot formation, so the shoot
induction medium formulation was modified. The new shoot induction strategy had two
stages.
First, the calli were placed on shoot inducing medium (MS basal medium
supplemented with 30 gIL sucrose, 2.75 gIL gellan gum, 5 mgIL kinetin, 0.2 mgIL
Indole-3-Acetic Acid (lAA), 80 mg!L adenine sulfate, 140 mgIL sodium phosphate
monobasic, 0.001 % vitamin solution (MS Vitamin solution, Sigma Aldrich M7150, St.
Louis, MO) and 0.0005% PPM (Preservative for Plant Tissue Culture Media, Plant Cell
Technology, Washington, D.C.) adjusted to pH 5.6). When the calli started to show signs
of shoot formation in the form of bud primordia, they were transferred to a new medium
(MS basal medium supplemented with 30 gIL sucrose, 2.75 gIL gellan gum, 2 mgIL
kinetin, 0.2 mgIL Indole-3-Acetic Acid (lAA), 80 mgIL adenine sulfate, 100 mgIL
sodium phosphate monobasic, 0.001 % vitamin solution and 0.0005% PPM adjusted to
pH 5.6). This medium differed from the shoot inducing medium only in that the
concentration of kinetin and sodium phosphate monobasic were lower. When buds fully
developed into shoots, they were transferred onto medium without hormones (MS basal
medium supplemented with 30 gIL sucrose, 2.75 gIL gellan gum, 0.001 % vitamin
solution, 100 mgIL glutamine, and 0.0005% PPM adjusted to pH 5.6).
77

Flow cytometry was used to screen for possible polyploids using the same
procedure as the in vivo experiments described earlier (p. 75-76).
Results
Experiment #1 - In vivo polyploid induetion
The percentage of buds that grew back after treatment with oryzalin ranged from
64.3% to 88.9% (Table 11). No abnormal morphology was observed in the new growth
that developed. One leaf from each developed shoot was used to analyze for conversion
using flow cytometry. Only one mixoploid shoot was identified from the treated buds.
This shoot was treated with 0.01 % oryzalin for 24 h but did not show any morphological
difference in growth from untreated shoots. However, the flow cytometric analysis
showed two peaks, one at the diploid level and one at the tetraploid level (Figure 13). All
the other histograms obtained showed fluorescence levels near the diploid controls.
Experiment #2 - In vitro polyploid induetion
The first formulation used for the shoot regeneration media was unsuccessful in
inducing shoots from the callus. Therefore, the formulation was changed, and after this,
the first shoots started emerging from the callus in 3 weeks. Calli were transferred to
new media every 7 to 14 days for 16 weeks to initiate more shoots. At the end of the
experiment (approximately 6 months after treatment), 36 regenerated plantlets were large
enough to be analyzed by flow cytometry.
Flow cytometric analysis revealed one tetraploid and one mixoploid plantlet
(Table 12). The mixoploid plantlet resulted from treatment with 0.01 % oryzalin for 48 h.
The histogram obtained showed two peaks, one at the diploid level and one at the
tetraploid level (Figure 14). This mixoploid plantlet is not morphologically different from
78

untreated control plants. The tetraploid plantlet resulted from treatment with 0.005%
oryzalin for 48 h. The histogram showed one peak at a fluorescence level approximately
double of that shown in untreated control plants (Figure 15). The regenerated plantlet
shows signs of being a polyploid such as very short internodes and more compact leaves
(Figure 16).
Discussion and Conclusions
The in vivo oryzalin treatment was unsuccessful in creating a tetraploid of D.
deremensis 'Santa Rosa'. Past experiments using in vivo treatments for polyploidization
have had mixed results. Treatments on buds of coffee and camellia were successful while
treatments of kiwi were not (Ackerman and Dermen, 1996; Berthou, 1975; Chalak and
Legave, 1996). The successful experiments had the common factor of isolating or
targeting the meristem for exposure to the colchicine or oryzalin solutions. Meristems
are desirable targets because of the higher mitotic activity, which improves the chances
the anti-mitotic chemicals will be effective. Several steps were taken in this experiment
to target the meristem and increase its exposure to the solution. Tips of the buds were cut
off to expose the meristems. Also, DMSO was used as the carrier solution for oryzalin.
OryzaIin doesn't dissolve well in water but it readily dissolves in DMSO, which has the
added benefit of being a good penetrant. It enhances the permeability of cell membranes
and therefore is rapidly absorbed by cells. As it is absorbed, it carries other substances
with it. Subsequently, using it as the carrier solution should have helped the oryzaIin
penetrate into the cells. Another step taken was continuously exposing the buds to the
oryzalin solution using the cotton ball and plastic wrapping. However, it seems these
steps were not sufficient to ensure the success of the treatment.
79

It was expected that the mnnber of buds killed during in vivo treatments would
increase with increased concentration of oryzalin and increased duration of treatment, but
the results did not show any clear trend (Table 11). The buds were probably not killed by
the oryzalin and instead by other causes such as the imprecise procedure of cutting the
bud to expose the meristem. The goal was to expose the meristem as much as possible,
but in doing so, the meristems could easily have been damaged as it was very difficult to
know how much of the newly growing bud to cut
The shoot identified as a mixoploid from the in vivo treatment of D. deremensis
'Santa Rosa' buds does not exhibit any abnormal morphological characteristics. It would
be interesting to count the chromosomes of this shoot to see if it is different from the
normal diploid number of2n = 38 (Mathew and Vijayavalli, 1989). However, this was
not possible as the shoot was not yet large enough to make a cutting to form roots, which
are needed for chromosome counts. When it is large enough, roots should be induced to
count the chromosomes. The shoot will continue to be monitored for any changes in
morphology or variegation.
Generally, previous polyploidization experiments using in vitro methods have had
higher levels of success in obtaining tetraploids than this experiment on Dracaena. The
treatment concentrations used were based on experiments on Lilium, in which lily scales
were treated instead of callus and 33.7% of regenerated plants were tetraploids (Tuyl et
al., 1992). The lower success rates in our experiments on Dracaena could have been due
to several different factors. Different kinds of obstacles were encountered during the in
vitro experiment than during the in vivo experiment, mostly related to the plant culture
media formulations. Oryzalin acts on mitotic cells and if the cells are not rapidly
80

dividing, the effect is lessened. Callus was used as the treated material in our
experiments for two main reasons. Regenerating plants from induced callus often gives
rise to new variation, even if no mutagenic treatment is made. Also, callus by definition
is in a high state of division, and this is the best kind of tissue to treat with antimitotic
chemicals. In our experiments, the callus growth after treatment was not as rapid as
desired. A different formulation for callus inducing media which results in faster callus
growth would be desirable for maximum polyploidization
Regeneration of shoots from callus was also problematic. As previously
mentioned, the first shoot induction medium was unsuccessful in regenerating shoots and
a new formulation had to be used that contained higher levels of kinetin and also
contained IAA for a very high cytokinin:auxin ratio of 25: 1. High cytokinin:auxin ratios
in plant culture media have been shown to stimulate shoot elongation and was also
reported to stimulate shoot growth in another D. deremensis variety (Debergh, 1975).
However, the loss of activity in the cultures before using the new formulation could have
negatively affected the results. Future experiments could prevent this by using high
cytokinin:auxin ratios for shoot induction.
One mixoploid and one tetraploid plantlet were obtained from the in vitro
experiments. The mixoploid does not exhibit any unusual characteristics, but the
tetraploid does have shorter internodes, which may translate into a more compact growth
habit. It also has shorter, more compact leaves. The chromosome count of this plant
would be useful and should be attempted in the future. The tetraploid plant will be
further evaluated to determine if it will be suitable as a new cultivar. It may also be useful
in breeding to create seedling variability and triploid progeny.
81

Overall, the objective of creating a polyploid of Dracaena was met, but the
success was limited as only one tetraploid and two mixoploids were obtained. However,
much information was learned about what procedures are most effective. The problems
encountered during the in vivo experiments were isolating the meristern and penetrating
the tissue. Higher concentrations of oryzalin or longer durations of treatment could be
used, but similar problems may be encountered. Since in vitro methods seem to be more
successful, perhaps future efforts should concentrate on improving this protocol. An
improved callus inducing media which stimulates more rapid growth should be tested.
Future experiments could also test colchicine to determine if it may be more effective on
Dracaena. Although oryzalin has recently proven effective in different crops for
polyploidization, the long history of success using colchicine cannot be ignored.
Different types of cultures should also be used for treatments, such as very young shoot
tips. Also, higher concentrations and increased durations of the oryzalin could be tested
for their effectiveness. With an improved procedure that overcomes the problems
encountered during this experiment, future efforts should yield larger numbers of
converted tetraploids.
82
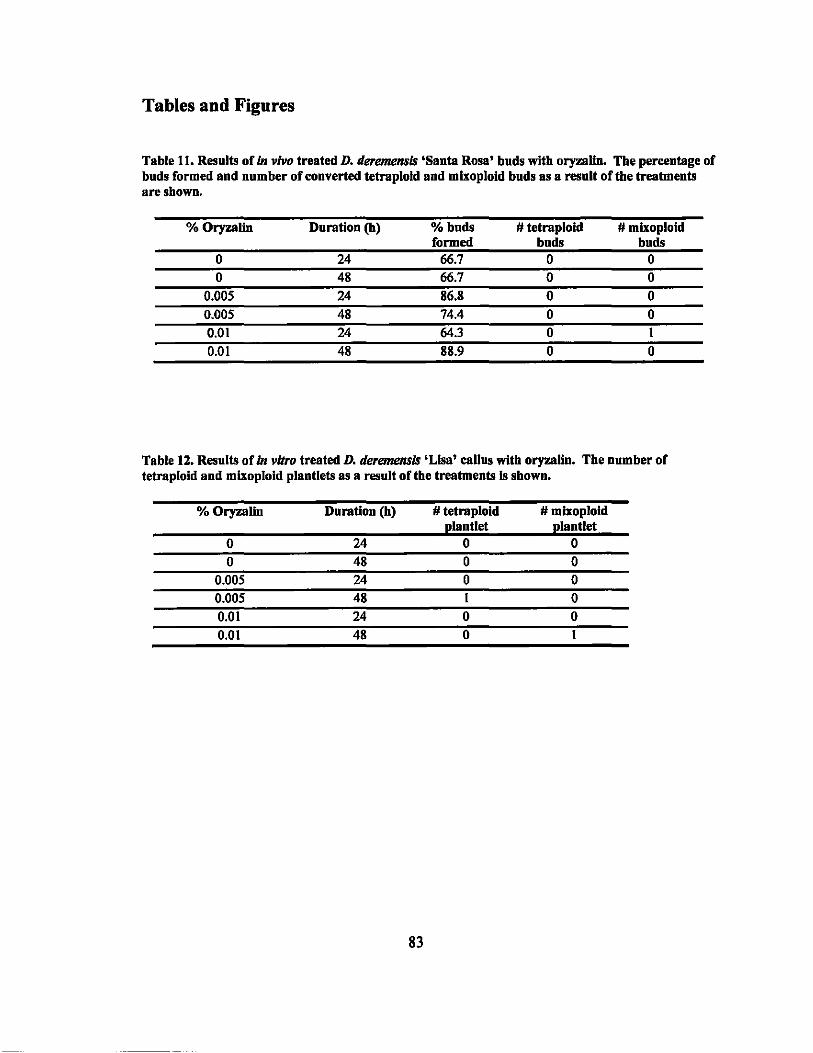
Tables and Figures
Table 11. Results of in vivo treated D. deremensls 'Santa Rosa' buds with oryzalln. The percentage of buds formed and number of converted tetraploid and mixoploid buds as a resnlt of tbe treatments aresbown.
%Oryzalln Duration (h) %bnds 1# tetraploid 1# mlxoploid formed bnds buds
0 24 66.7 0 0 0 48 66.7 0 0
0.005 24 86.8 0 0 0.005 48 74.4 0 0 0.01 24 64.3 0 1
0.01 48 88.9 0 0
Table 12. Results of in vitro treated D. deremensls 'Lisa' callns witb oryzalln. The number of tetraploid and mlxoplold plantlets as a result oUbe treatments Is shown.
%Oryzalln Duration (h) 1# tetraploid 1# mlxoploid planttet plantlet
0 24 0 0 0 48 0 0
0.005 24 0 0 0.005 48 1 0 0.01 24 0 0 0.01 48 0 1
83

Figure 11.1" vivo oryzalin treatment procedures of D. deremellsi~' 'Santa Rosa' buds. a. Size of buds treated. b. Tip of bud cut off to expose meriste",. c. Bud covered in oryzalin soaked cotton and plastic during treatment.
Figure 12. III vitro oryzalin treatment procedures of D. deremellsis 'Lisa '. a. Callus formation at base of shoot explant. b. Calills being treated with oryzalin solution. c. Callus on shoot regeneration media with bud primordia developing. d. Developing regenerated shoot.
84
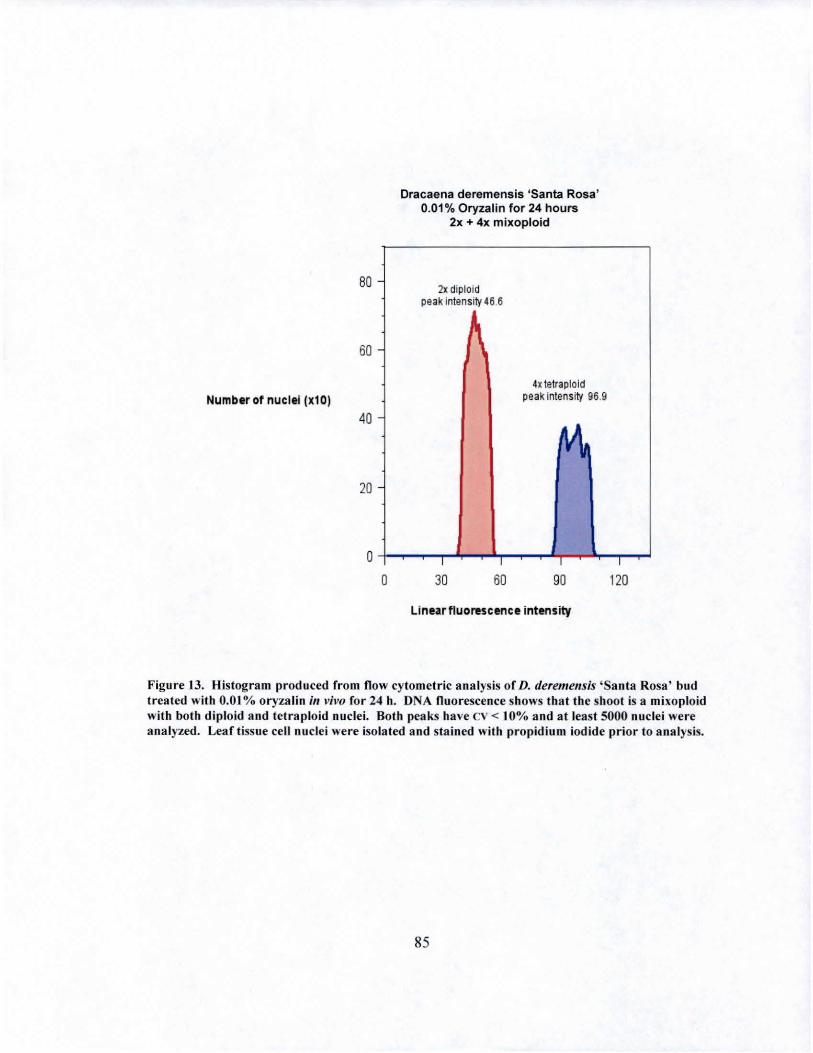
80 -
60 -
Number of nuclei (xl0)
40
20
o o
Dracaena deremensis ISanta Rosa ' 0.01 % Oryzalin for 24 hours
2x + 4x mixoploid
2x diploid peak i nten S~4 6.6
4xletrap loid peak inlens ity 96.9
~
I I I
30 60 90
Linearfluorescence intensity
I
120
Figure 13. Histogram produced from n ow cytometric a na lys is of D. deremensis ' Sa nta Rosa ' bud treated with 0.0 1 % oryzalin ill vivo for 24 h. DNA nuorescence shows that the shoot is a mixoploid with both diploid and tet ra ploid nuclei. Both peaks have cv < 10% a nd a t leas t 5000 nuclei were analyzed. Leaf tissue cell nuclei were isolated and stained with propidium iod ide prior to analys is.
85
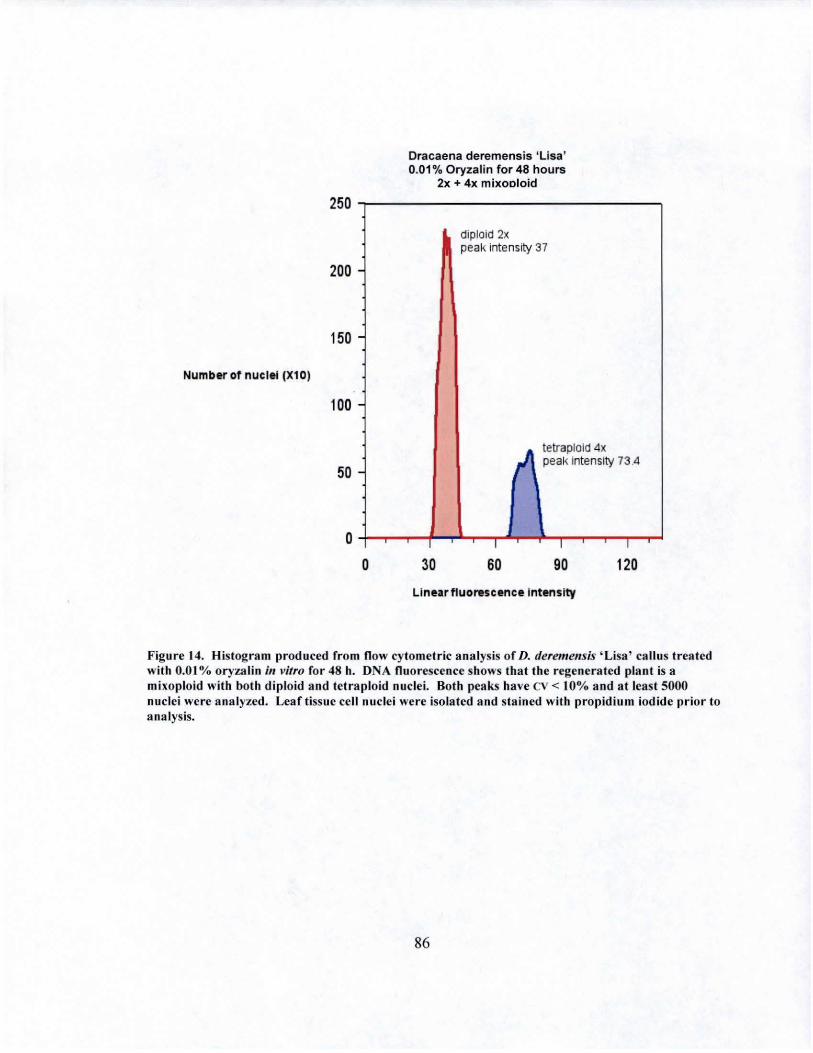
Numb.rot nuclei (Xl0)
250
200
150 -
100
50 -
Dracaena deremensis 'Lisa' 0.01 % Oryzalin for 48 hours
2x + 4x mixooloid
diploid 2x peak intensity 37
tetraploid 4x peak Intensity 73 .4
O +-~~-+~~~~I ~~-r-I ~~-r-I ~
o 30 60 90 120
Figure 14. Histogram produced from now cytometric analysis of D. fleremells;s 'Lisa' callus treated with 0.01 % oryzalin ill vitro for 48 h. DNA flu orescence shows that the regenerated plant is a mixoploid with both diploid and tetraploid nuclei. Both peaks have cv < 10% and at least 5000 nuclei were analyzed. Leaf tisslie cell nuclei were isolated and stained with propidium iodide prior to analysis.
86
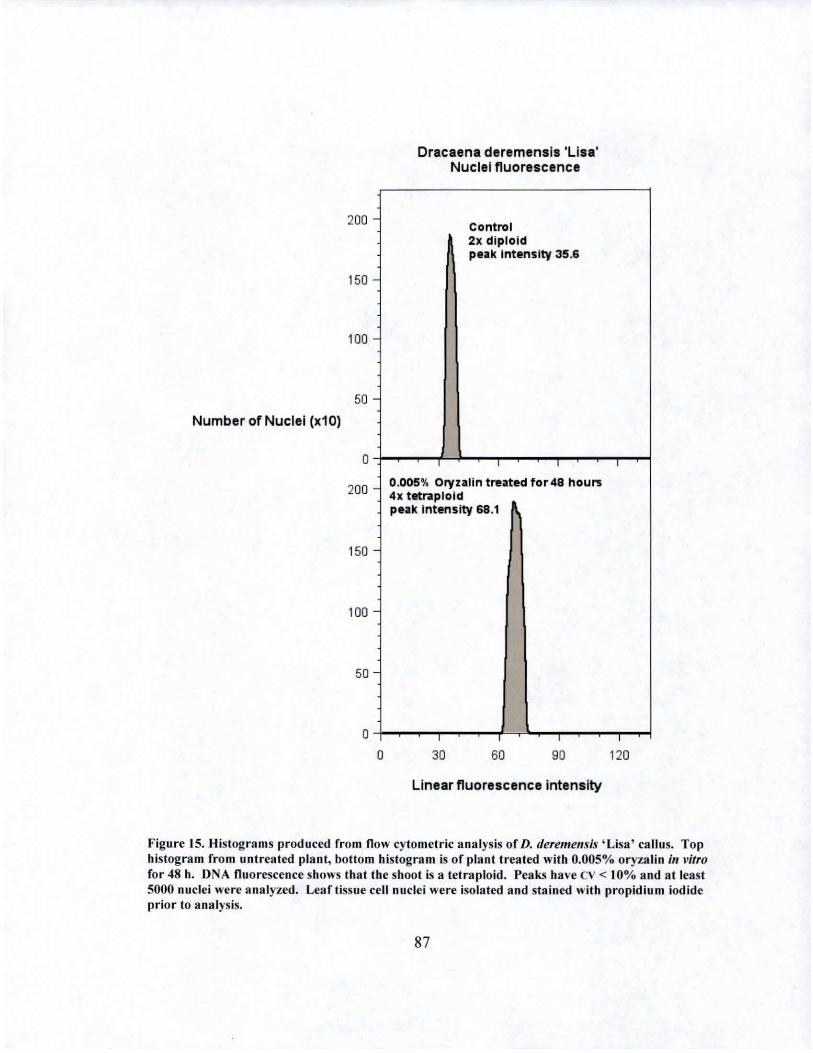
200 -
150 -
100 -
50 -Number of Nuclei (x10)
o
200 -
150
100 -
50
o o
Dracaena deremensis 'Lisa' Nuclei fluorescence
Cont ro l 2x diploid peak Intensity 35.6
0.005% Oryzalin treated for48 hours 4x tetraploid p eak Intensity 68 .1
I
30 60 90
Linear fluorescence intensity
I
120
Figu re 15. Histograms produced rrom flow cyto mctric analysis of D. deremellsis 'Lisa' callus. Top histogram from untreated plant, bottom histogram is of plant t reated wit h 0.005% oryza lin in vitro for 48 h. D A nuorcscence shows that the shoot is a tetra ploid. Peaks have cv < 10% and at least 5000 nuclei were analyzed. Leaf tissue cell nuciei were isolated and stained with propidium iod ide prior to ana lysis.
87

D. deremensis ' Lisa'
Diploid Tetraploid
Figure 16. Comparison of diploid and tetraploid D. deremensis ' Lisa' ill vitro plantlels. Tetraploid plantlet obtained from callus treated with 0.005% oryzalin for 48 h has shorter internodes and more compact leaves than its diploid counterpart.
88
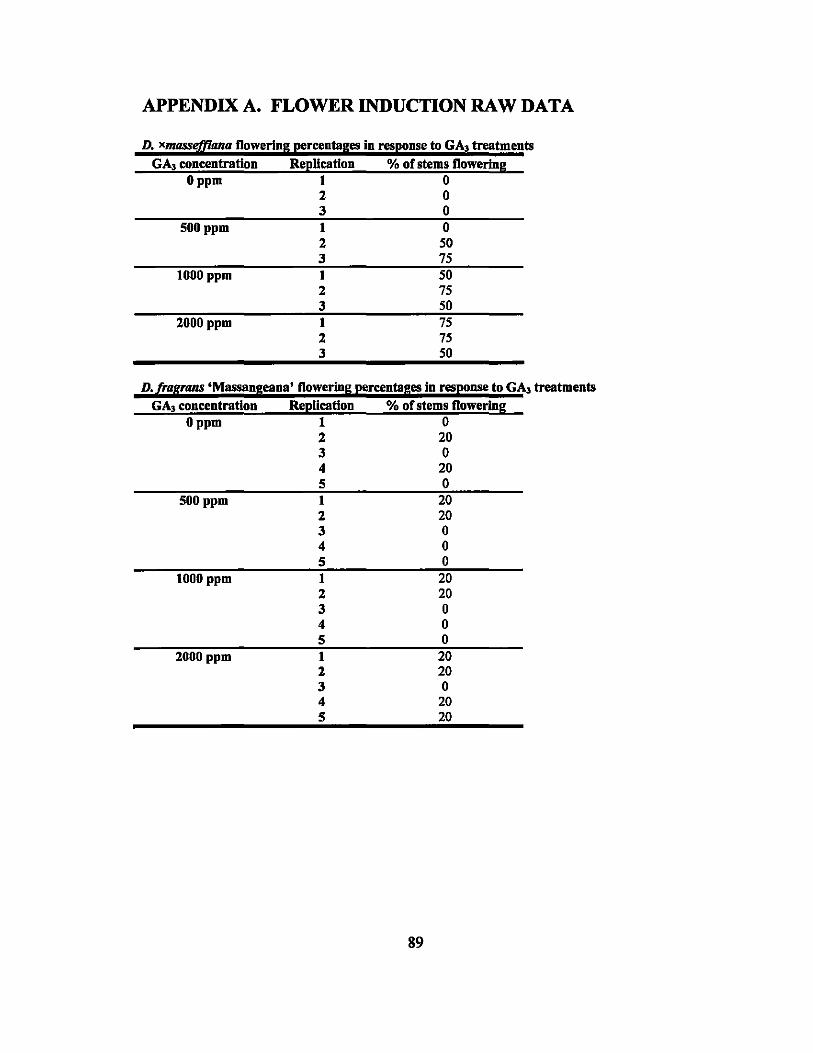
APPENDIX A. FLOWER INDUCTION RAW DATA
D. xmasstifflana flowering percentages in response to GA. treatments GA, concentration
o ppm
SOOppm
1000 ppm
2000 ppm
Replication 1 2 3 1 2 3 1 2 3 I 2 3
% of stems flowering o o o o
SO 75 50 75 50 75 75 SO
D.[rapans 'Massangeana' flowering percentages in response to GA. treatments GA. concentration
o ppm
500 ppm
1000 ppm
2000 ppm
Replication I 2 3 4 5 1 2 3 4 5 I 2 3 4 5 1 2 3 4 5
% of stems flowering
89
o 20 o 20 o
20 20 o o o 20 20 o o o
20 20 o 20 20
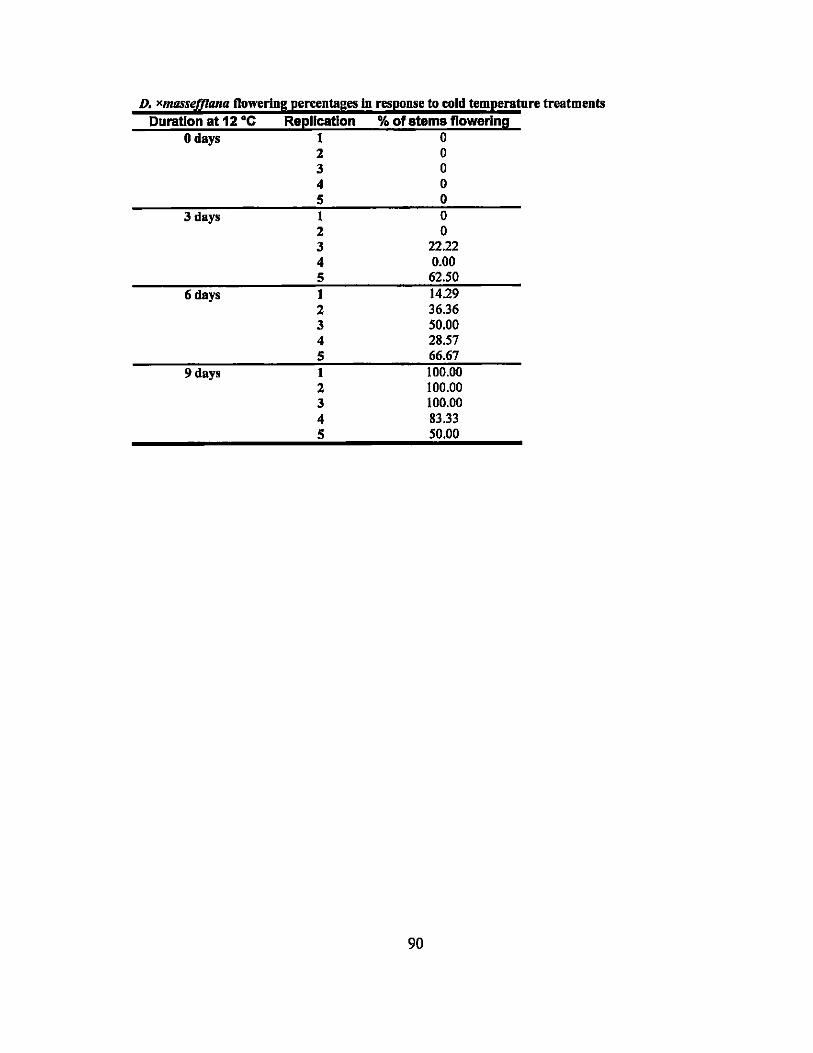
D. xlltIlSSefflanu Dowering percentages In response to cold temperatnre treatments Duration at 12 ·C ReplIcation % ofatems flowering
o days 1 0 2 0 3 0 4 0 5 0
3 days I 0 2 0 3 22.22 4 0.00 5 62.50
6 days 1 14.29 2 36.36 3 50.00 4 28.57 5 66.67
9 days I 100.00 2 100.00 3 100.00 4 83.33 5 50.00
90
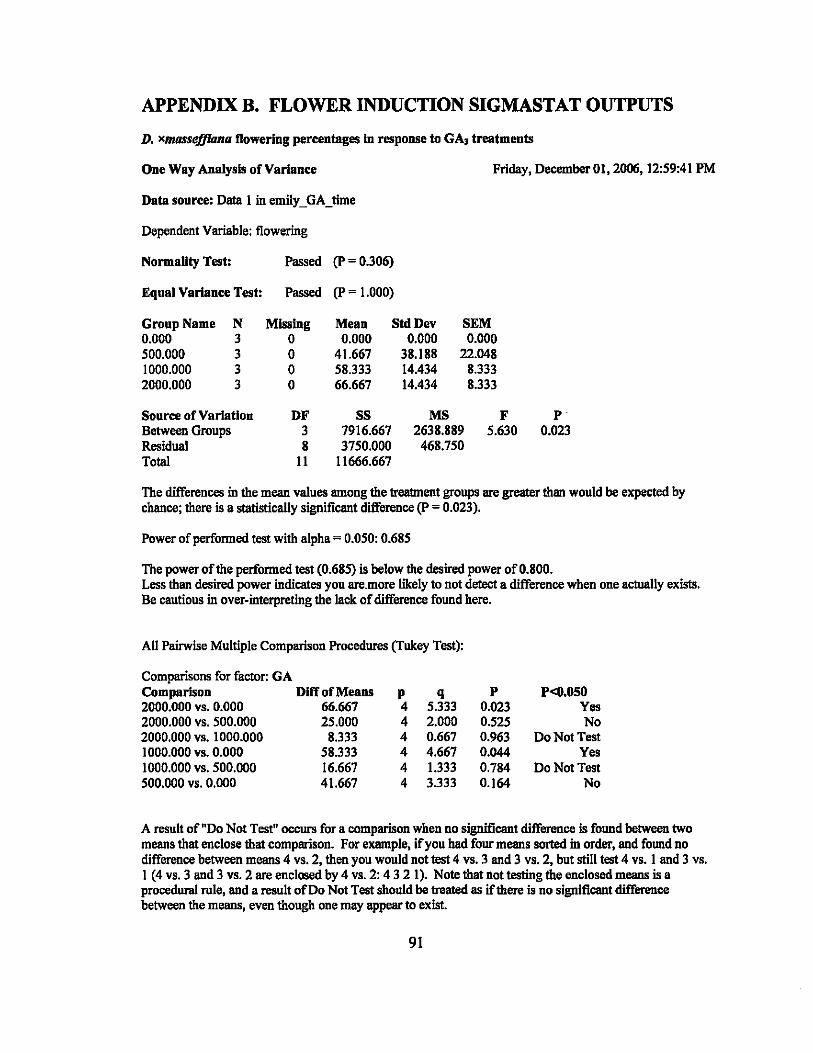
APPENDIX B. FLOWER INDUCTION SIGMASTAT OUTPUTS
D. X1IIIJSSejJIIlna flowering percentages in response to GA. treatments
One Way Analysis of Vartance Friday, December 01,2006, 12:59:41 PM
Dependent Variable: flowering
Normality Test: Passed (P= 0.306)
Equal Vartance Test: Passed (P = 1.000)
Group Name N Missing Mean StdDev SEM 0.000 3 0 0.000 0.000 0.000 500.000 3 0 41.667 38.188 22.048 1000.000 3 0 58.333 14.434 8.333 2000.000 3 0 66.667 14.434 8.333
Source of Vartation DF SS MS F P Between Groups 3 7916.667 2638.889 5.630 0.023 Residual 8 3750.000 468.750 Total II 11666.667
The differences in the mean values among the treatment groups are greater thao would be expected by chance; there is a statistically significant difference (P = 0.023).
Power of performed test with alpha = 0.050: 0.685
The power of the performed test (0.685) is below the desired power of 0.800. Less than desired power indicates you are.more likely to not detect a difference when one actually exists. Be cautious in over-interpreting the lack of difference found here.
All Pairwise Multiple Comparison Procedures (Tukey Test):
Comparisons for factor: GA Comparison Diff of Means p q 2000.000 vs. 0.000 66.667 4 5.333 2000.000 vs. 500.000 25.000 4 2.000 2000.000 vs. 1000.000 8.333 4 0.667 1000.000 vs. 0.000 58.333 4 4.667 1000.000 vs. 500.000 16.667 4 1.333 500.000 vs. 0.000 41.667 4 3.333
P 0.023 0.525 0.963 0.044 0.784 0.164
P<O.OSO Yes No
Do Not Test Yes
Do Not Test No
A result of "Do Not Test" occurs for a comparison when no significant difference is found between two means that enclose that comparison. For example, if you had four means sorted in order, and found no difference between means 4 vs. 2, then you would not test 4 vs. 3 and 3 vs. 2, but still test 4 vs. I and 3 vs. 1 (4 vs. 3 and 3 vs. 2 are enclosed by 4 vs. 2: 432 I). Note that not testing the enclosed means is a procedural rule, and a result of Do Not Test should be treated as if there is no significant difference between the means, even though one may appear to exist.
91
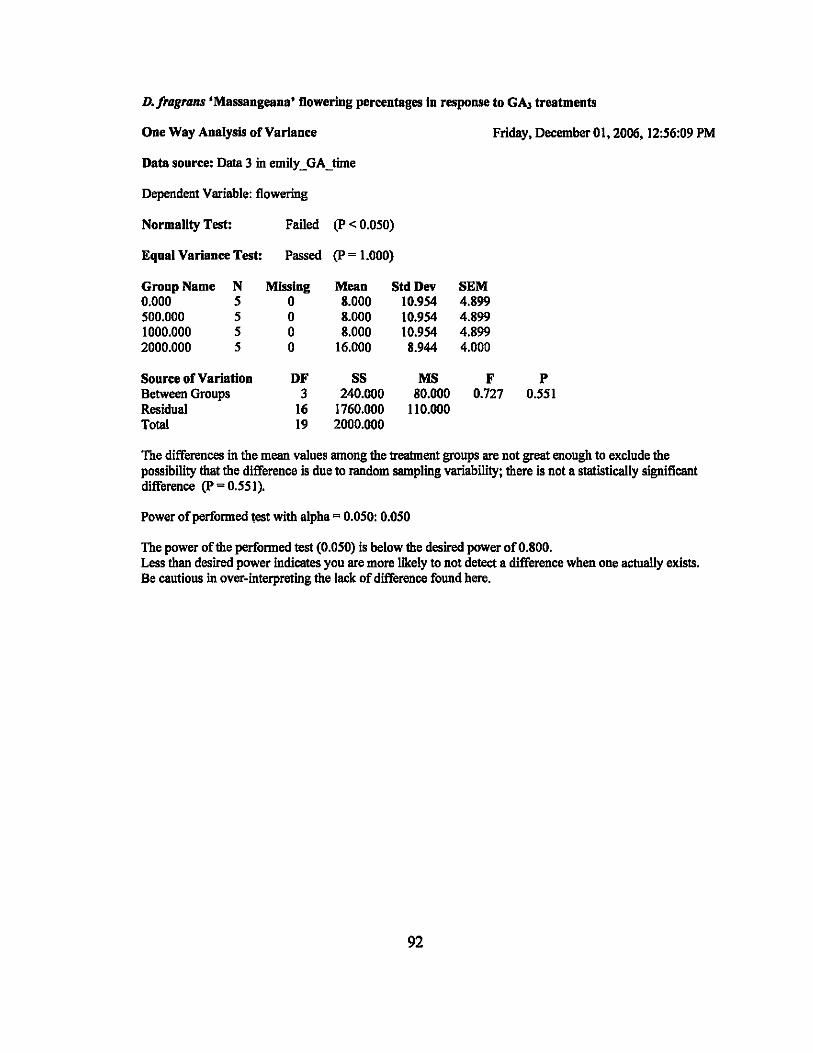
D./ragrans 'Massangeana' flowering percentages In response to GA3 treatments
One Way Analysis of Variance Friday, December 01, 2006, 12:56:09 PM
Data source: Data 3 in emilLGA_time
Dependent Variable: flowering
Normality Test: Failed (P < 0.050)
Equal Variance Test: Passed (P = 1.000)
Group Name N Missing Mean Std Dev SEM 0.000 5 0 8.000 10.954 4.899 500.000 5 0 8.000 10.954 4.899 1000.000 5 a 8.000 10.954 4.899 2000.000 5 a 16.000 8.944 4.000
Source of Variation DF SS MS F P Between Groups 3 240.000 80.000 0.727 0.551 Residual 16 1760.000 110.000 Total 19 2000.000
The differences in the mean values among the treatment groups are not great enough to exclude the possibility that the difference is due to random sampling variability; there is not a statistically significant difference (P=0.551).
Power of performed ~ with alpha = 0.050: 0.050
The power of the performed test (0.050) is below the desired power of 0.800. Less than desired power indicates you are more likely to not detect a difference when one actually exists. Be cautious in over-interpreting the lack of difference found here.
92
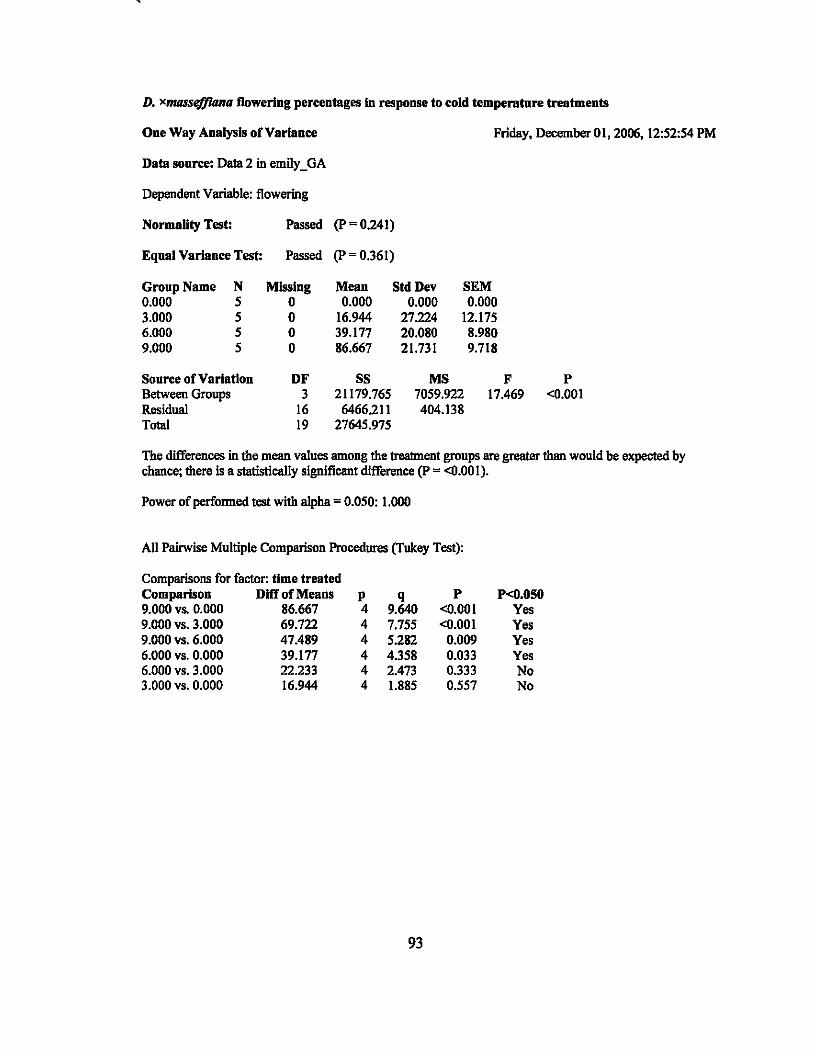
D. xmassefl1ana flowering percentages In response to cold temperatnre treatments
One Way Analysis of Variance Friday, December 01, 2006,12:52:54 PM
Data sonrce: Data 2 in emily _ GA
Dependent Variable: flowering
Normality Test: Passed (p=0241)
Eqnal Variance Test: Passed (p=0.361)
Group Name N Missing Meau Std Dev SEM 0.000 5 0 0.000 0.000 0.000 3.000 5 0 16.944 27224 12.175 6.000 5 0 39.177 20.080 8.980 9.000 5 0 86.667 21.731 9.718
Source of Variation DF SS MS F P Between Groups 3 21179.765 7059.922 17.469 <0.001 Residual 16 6466211 404.138 Total 19 27645.975
The differences in the mean values among the treatment groups are greater than would be expected by chance; there is a statistieally significant difference (p = <0.001).
Power ofperfonned test with alpha = 0.050: 1.000
All Pairwise Multiple Comparison Procedures (Tukey Test):
Comparisons for factor: time treated Comparison Dlff of Means p 9.000 vs. 0.000 86.667 4 9.000 vs. 3.000 69.722 4 9.000 vs. 6.000 47.489 4 6.000 vs. 0.000 39.177 4 6.000 vs. 3.000 22.233 4 3.000 vs. 0.000 16.944 4
q 9.640 7.755 5.282 4.358 2.473 1.885
93
P <0.001 <0.001
0.009 0.033 0.333 0.557
P<O.OSO Yes Yes Yes Yes No No
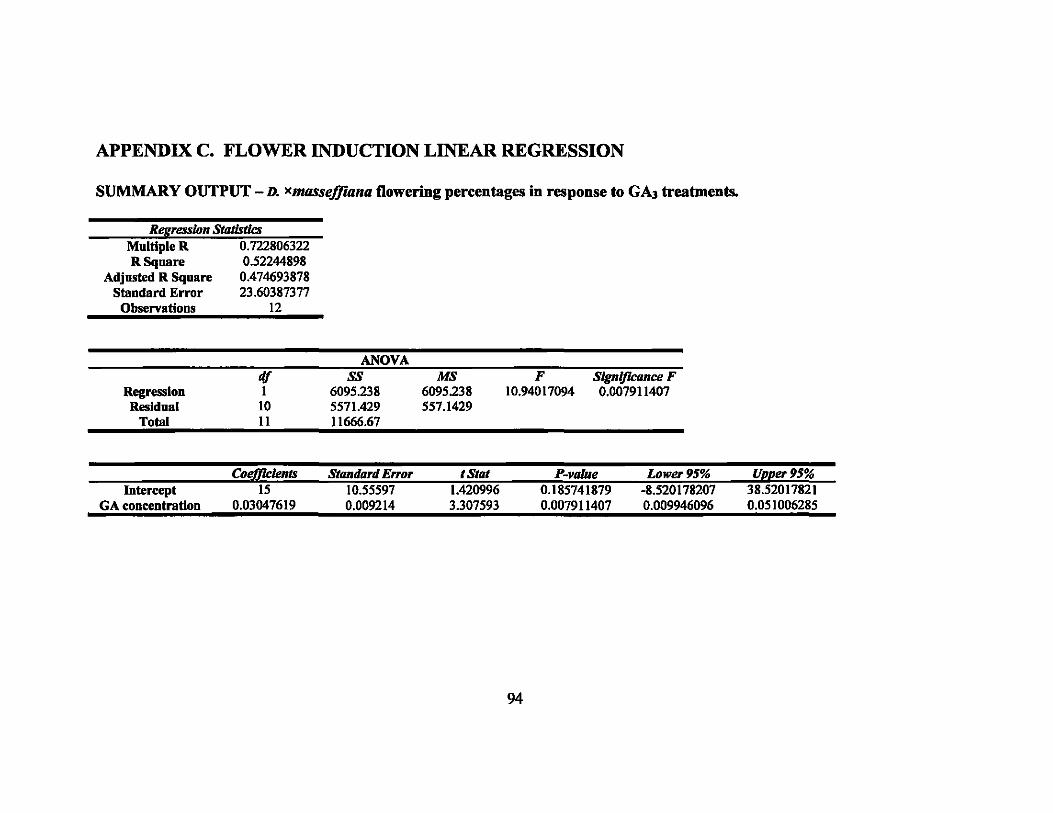
APPENDIX C. FLOWER INDUCTION LINEAR REGRESSION
SUMMARY OUTPUT - D. xlIIIlSseffwna flowering percentages in response to GA3 treatments.
Regression StIlIistks Multiple R 0.722806322 R Square 0.52244898
Adjusted R Square 0.474693878 Standard Error 23.60387377
Observatious 12
Regression Residual
Total
GA concentration
df 1
10 11
0.03047619
ANOVA ss
6095238 5571.429 11666.67
0.009214
MS 6095238 557.1429
3.307593
F 10.94017094
Slgnljlcance F 0.007911407
0.007911407 0.009946096
94
0.051006285
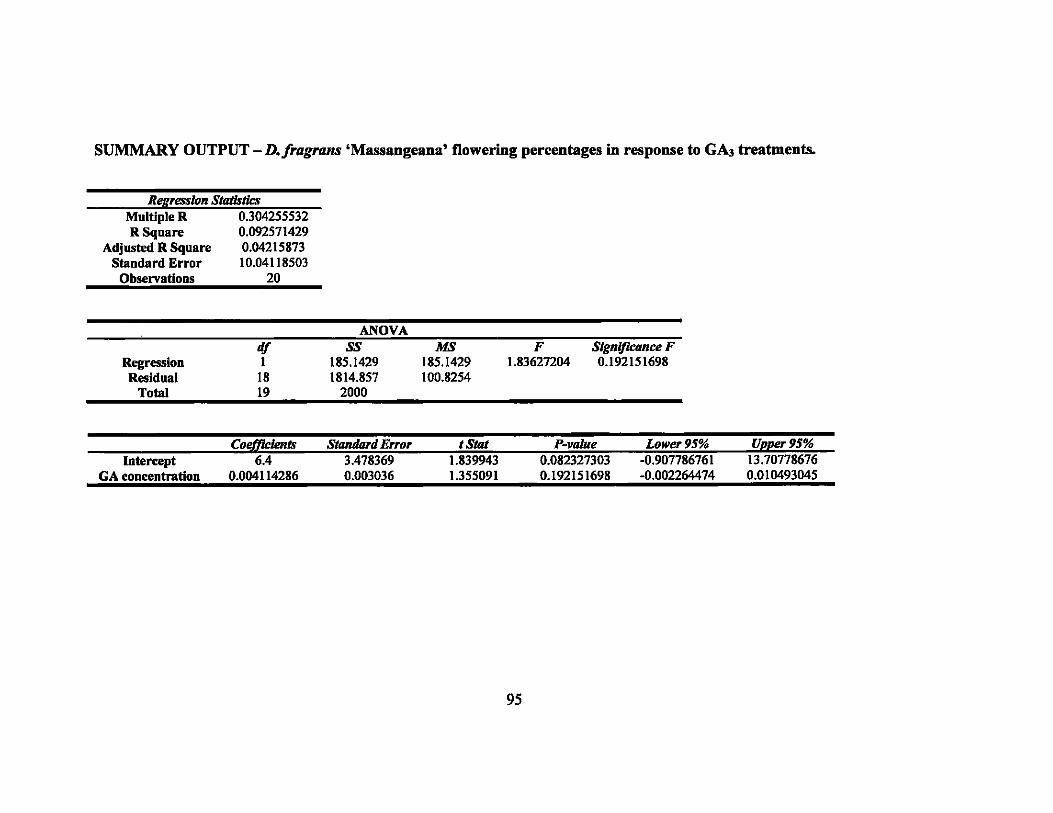
SUMMARY OUTPUT - D./ragrans 'Massangeana' flowering percentages in response to GAJ treatments.
Regression StatlstJcs Multiple R 0.304255532 R Square 0.092571429
Adjusted R Square 0.04215873 Standard Error 10.04118503
Observations 20
Regression Residual
Total
df I
18 19
ANOVA ss
185.1429 1814.857
2000
MS 185.1429 100.8254
F 1.83627204
Slgnljlcance F 0.192151698
Col!fficlents Stamlard E"or t Stot P-vaJue Lower 95% JfJJJ!er 95% Intercept 6.4 3.478369 1.839943 0.082327303 -0.907786761 13.70778676
GA concentration 0.004114286 0.003036 1.355091 0.192151698 -0.002264474 0.010493045
95
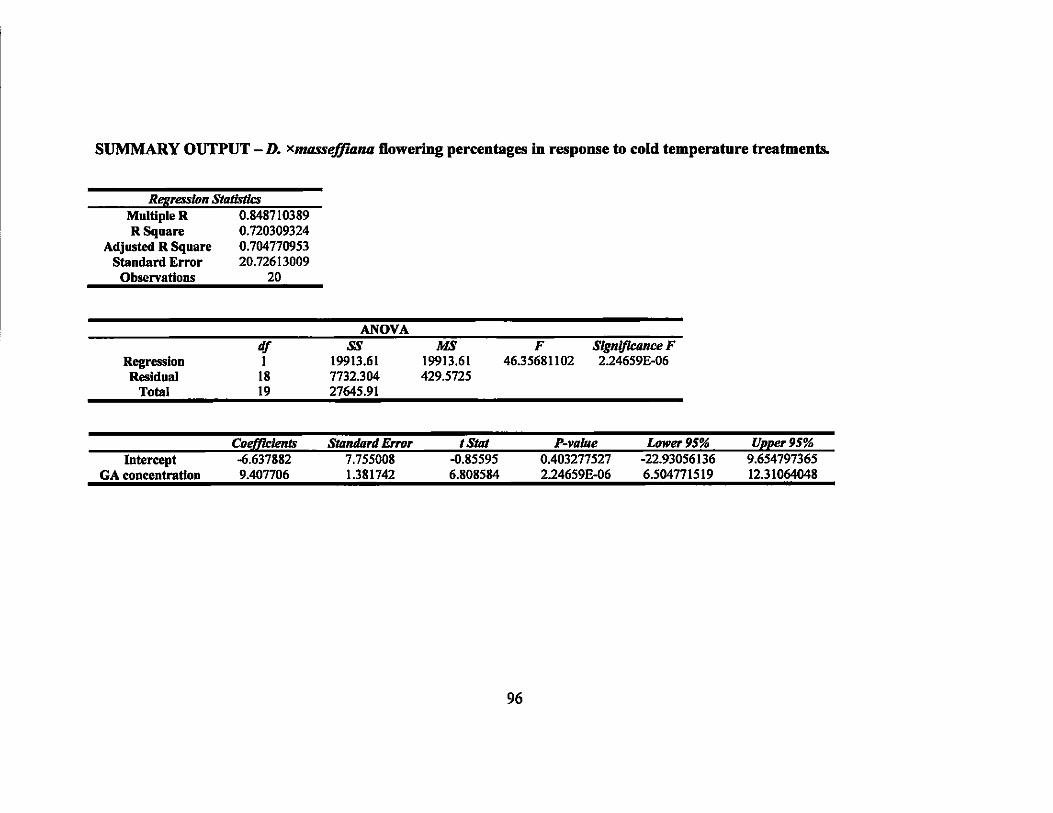
SUMMARY OUTPUT - D. XmtlSse!flllna Dowering percentages in response to cold temperature treatments.
Regression Statistics Multiple R 0.848710389 R Square 0.720309324
Adjusted R Square 0.704770953 Staudard Error 20.72613009
Observations 20
Regression Residual
Total
GA concentration
df 1
18 19
9.407706
ANOVA ss
19913.61 7732304 27645.91
1.381742
MS 19913.61 429.5725
6.808584
F 46.35681102
Slgnlfkance F 2.24659E-06
2.24659E·06 6.504771519
96
12.31064048
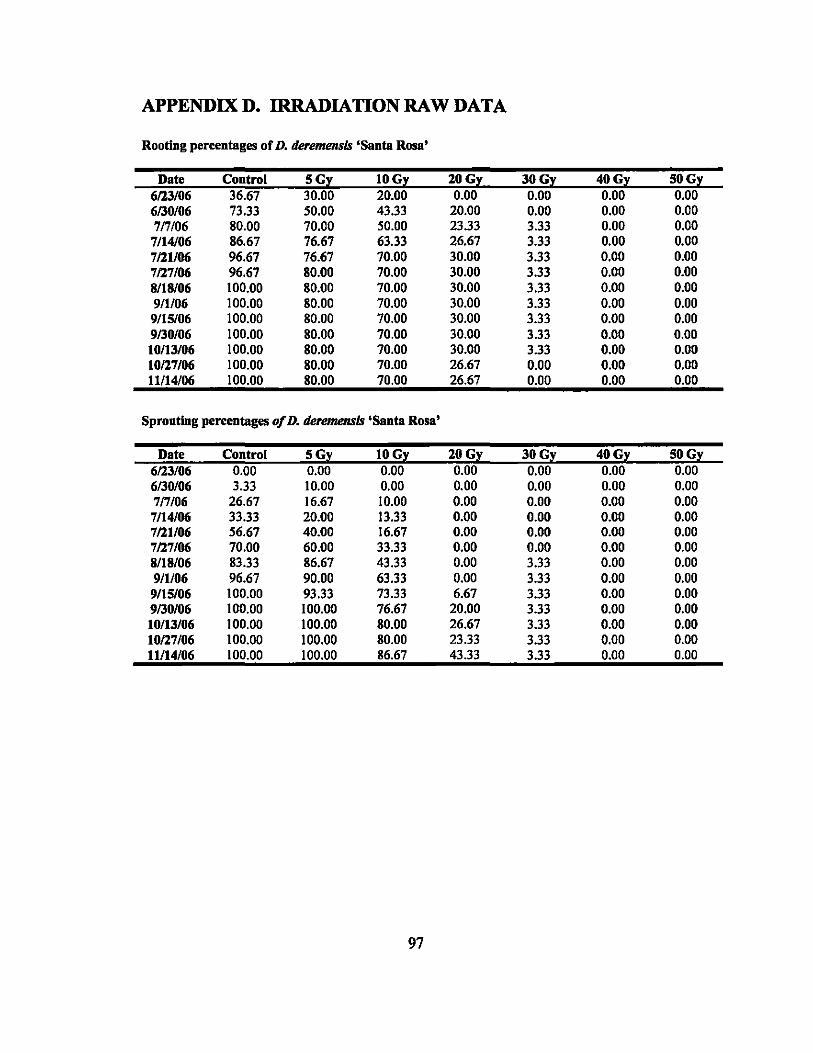
APPENDIXD. IRRADIATIONRAWDATA
Rooting percentages of D. derenumsls 'Santa Rosa'
Date Control SGy lOGy lOGy 30Gy 40Gy 50Gy 6123/06 36.67 30.00 20.00 0.00 0.00 0.00 0.00 6130/06 73.33 50.00 43.33 20.00 0.00 0.00 0.00 7n/06 80.00 70.00 50.00 23.33 3.33 0.00 0.00 7/14/06 86.67 76.67 63.33 26.67 3.33 0.00 0.00 7121/06 96.67 76.67 70.00 30.00 3.33 0.00 0.00 7127/06 96.67 80.00 70.00 30.00 3.33 0.00 0.00 8118/06 100.00 80.00 70.00 30.00 3.33 0.00 0.00 9/1106 100.00 80.00 70.00 30.00 3.33 0.00 0.00 9/15/06 100.00 80.00 70.00 30.00 3.33 0.00 0.00 9130/06 100.00 80.00 70.00 30.00 3.33 0.00 0.00 10/13/06 100.00 80.00 70.00 30.00 3.33 0.00 0.00 10127/06 100.00 80.00 70.00 26.67 0.00 0.00 0.00 11114/06 100.00 80.00 70.00 26.67 0.00 0.00 0.00
Spronting percentages of D. derenumsls 'Santa Rosa'
Date Control SGy lOGy lOGy 30Gy 40Gy 50Gy 6123/06 0.00 0.00 0.00 0.00 0.00 0.00 0.00 6130/06 3.33 10.00 0.00 0.00 0.00 0.00 0.00 7n/06 26.67 16.67 10.00 0.00 0.00 0.00 0.00 7/14/06 33.33 20.00 13.33 0.00 0.00 0.00 0.00 7121/06 56.67 40.00 16.67 0.00 0.00 0.00 0.00 7127/06 70.00 60.00 33.33 0.00 0.00 0.00 0.00 8118/06 83.33 86.67 43.33 0.00 3.33 0.00 0.00 9/1/06 96.67 90.00 63.33 0.00 3.33 0.00 0.00 9/15/06 100.00 93.33 73.33 6.67 3.33 0.00 0.00 9130/06 100.00 100.00 76.67 20.00 3.33 0.00 0.00 10/13/06 100.00 100.00 80.00 26.67 3.33 0.00 0.00 10127/06 100.00 100.00 80.00 23.33 3.33 0.00 0.00 11114/06 100.00 100.00 86.67 43.33 3.33 0.00 0.00
97
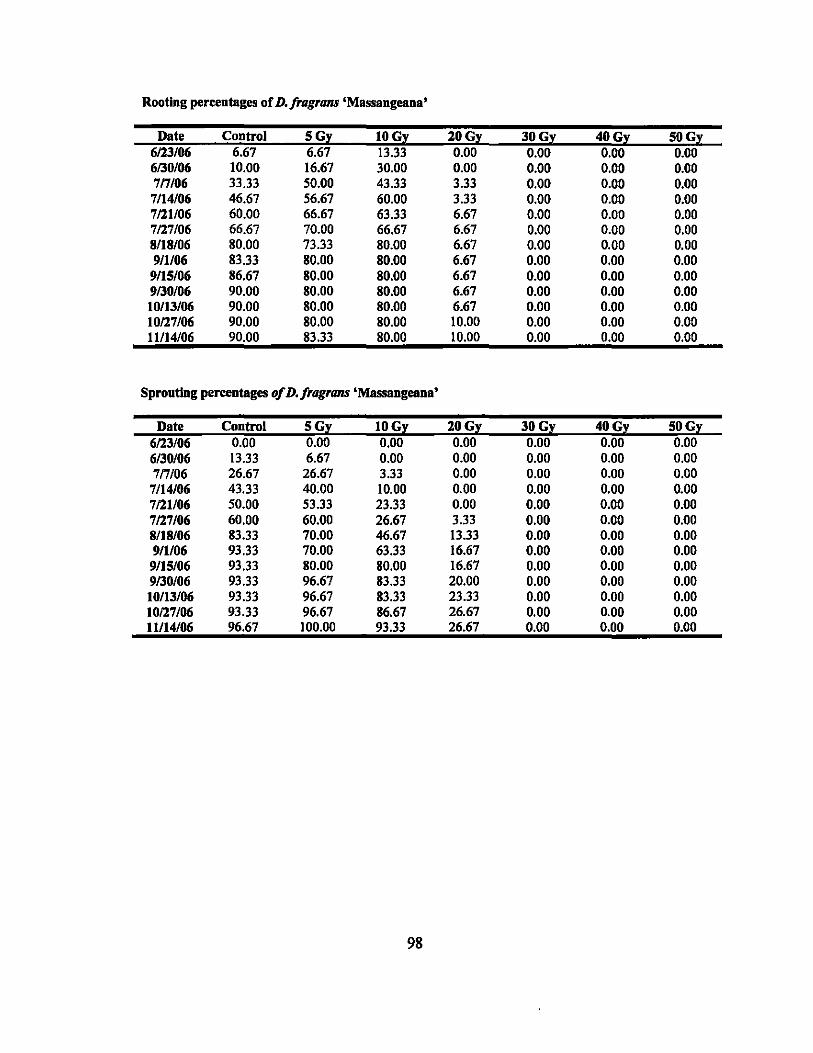
Rooting percentages of D. fragrans 'Massangeana'
Date Control 5Gy lOGy 20Gy 30Gy 40Gy 50Gy 6123/06 6.67 6.67 13.33 0.00 0.00 0.00 0.00 6130/06 10.00 16.67 30.00 0.00 0.00 0.00 0.00 7n/06 33.33 50.00 43.33 3.33 0.00 0.00 0.00 7/14/06 46.67 56.67 60.00 3.33 0.00 0.00 0.00 7121106 60.00 66.67 63.33 6.67 0.00 0.00 0.00 7127/06 66.67 70.00 66.67 6.67 0.00 0.00 0.00 8/18/06 80.00 73.33 80.00 6.67 0.00 0.00 0.00 9/1/06 83.33 80.00 80.00 6.67 0.00 0.00 0.00 9/15/06 86.67 80.00 80.00 6.67 0.00 0.00 0.00 9130/06 90.00 80.00 80.00 6.67 0.00 0.00 0.00 10/13/06 90.00 80.00 80.00 6.67 0.00 0.00 0.00 10127/06 90.00 80.00 80.00 10.00 0.00 0.00 0.00 11114/06 90.00 83.33 80.00 10.00 0.00 0.00 0.00
Sprouttag percentages ofD.fragrans 'Massangeaua'
Date Control 5Gy lOGy 20Gy 30Gy 40Gy 50Gy 6123/06 0.00 0.00 0.00 0.00 0.00 0.00 0.00 6130/06 13.33 6.67 0.00 0.00 0.00 0.00 0.00 7n/06 26.67 26.67 3.33 0.00 0.00 0.00 0.00 7/14/06 43.33 40.00 10.00 0.00 0.00 0.00 0.00 7121/06 50.00 53.33 23.33 0.00 0.00 0.00 0.00 7127/06 60.00 60.00 26.67 3.33 0.00 0.00 0.00 8118/06 83.33 70.00 46.67 13.33 0.00 0.00 0.00 9/1/06 93.33 70.00 63.33 16.67 0.00 0.00 0.00 9/15/06 93.33 80.00 80.00 16.67 0.00 0.00 0.00 9130/06 93.33 96.67 83.33 20.00 0.00 0.00 0.00 10/13/06 93.33 96.67 83.33 23.33 0.00 0.00 0.00 10127106 93.33 96.67 86.67 26.67 0.00 0.00 0.00 11114/06 96.67 100.00 93.33 26.67 0.00 0.00 0.00
98
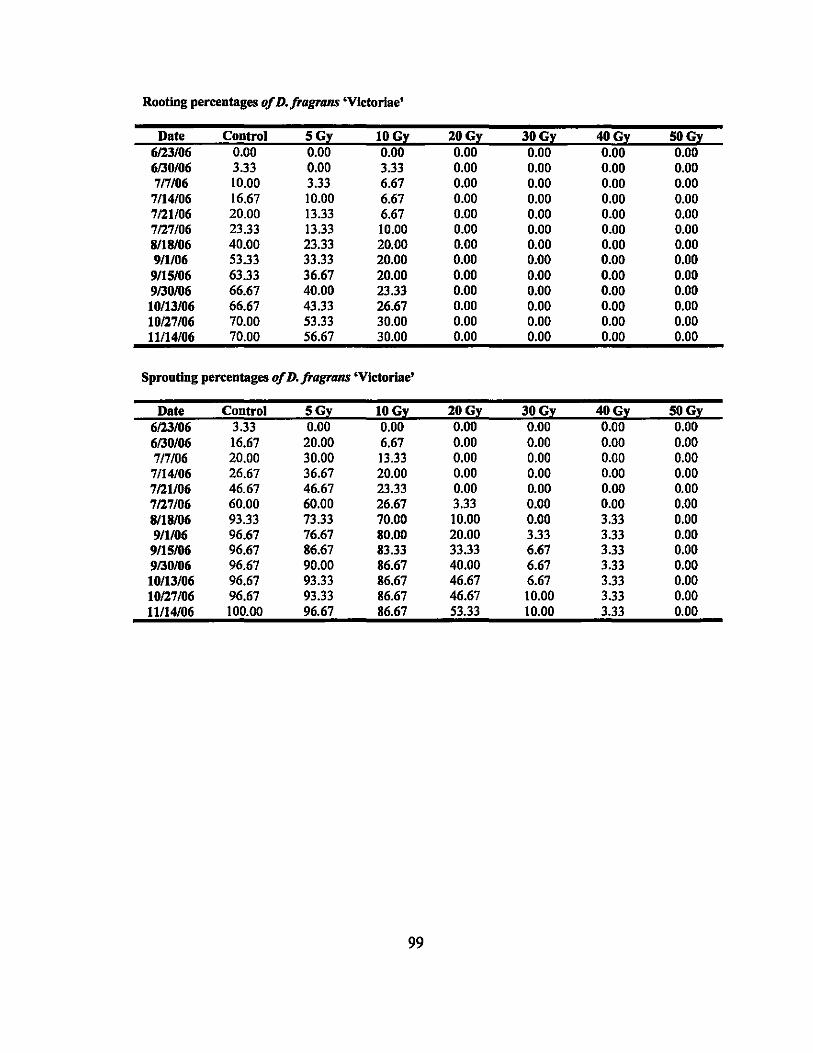
Rooting percentages of D. fragrans 'Vlctorlae'
Date Control 5Gy lOGy 20Gy 30Gy 40Gy SUGy 6123/06 0.00 0.00 0.00 0.00 0.00 0.00 0.00 6130/06 3.33 0.00 3.33 0.00 0.00 0.00 0.00 717106 10.00 3.33 6.67 0.00 0.00 0.00 0.00
7114/06 16.67 10.00 6.67 0.00 0.00 0.00 0.00 7121106 20.00 13.33 6.67 0.00 0.00 0.00 0.00 7127/06 23.33 13.33 10.00 0.00 0.00 0.00 0.00 8/18106 40.00 23.33 20.00 0.00 0.00 0.00 0.00 9/1/06 53.33 33.33 20.00 0.00 0.00 0.00 0.00 9/15/06 63.33 36.67 20.00 0.00 0.00 0.00 0.00 9130106 66.67 40.00 23.33 0.00 0.00 0.00 0.00 10/13/06 66.67 43.33 26.67 0.00 0.00 0.00 0.00 10127106 70.00 53.33 30.00 0.00 0.00 0.00 0.00 11114/06 70.00 56.67 30.00 0.00 0.00 0.00 0.00
Sprouting percentages of D. fragrans 'Victorlae'
Date Control 5Gy lOGy 20Gy 30Gy 40Gy 50Gy 6123/06 3.33 0.00 0.00 0.00 0.00 0.00 0.00 6130/06 16.67 20.00 6.67 0.00 0.00 0.00 0.00 7n/06 20.00 30.00 13.33 0.00 0.00 0.00 0.00 7114/06 26.67 36.67 20.00 0.00 0.00 0.00 0.00 7121106 46.67 46.67 23.33 0.00 0.00 0.00 0.00 7127106 60.00 60.00 26.67 3.33 0.00 0.00 0.00 8/18106 93.33 73.33 70.00 10.00 0.00 3.33 0.00 9/1/06 96.67 76.67 80.00 20.00 3.33 3.33 0.00
9115/06 96.67 86.67 83.33 33.33 6.67 3.33 0.00 9130/06 96.67 90.00 86.67 40.00 6.67 3.33 0.00 10/13/06 96.67 93.33 86.67 46.67 6.67 3.33 0.00 10127106 96.67 93.33 86.67 46.67 10.00 3.33 0.00 11114/06 100.00 96.67 86.67 53.33 10.00 3.33 0.00
99
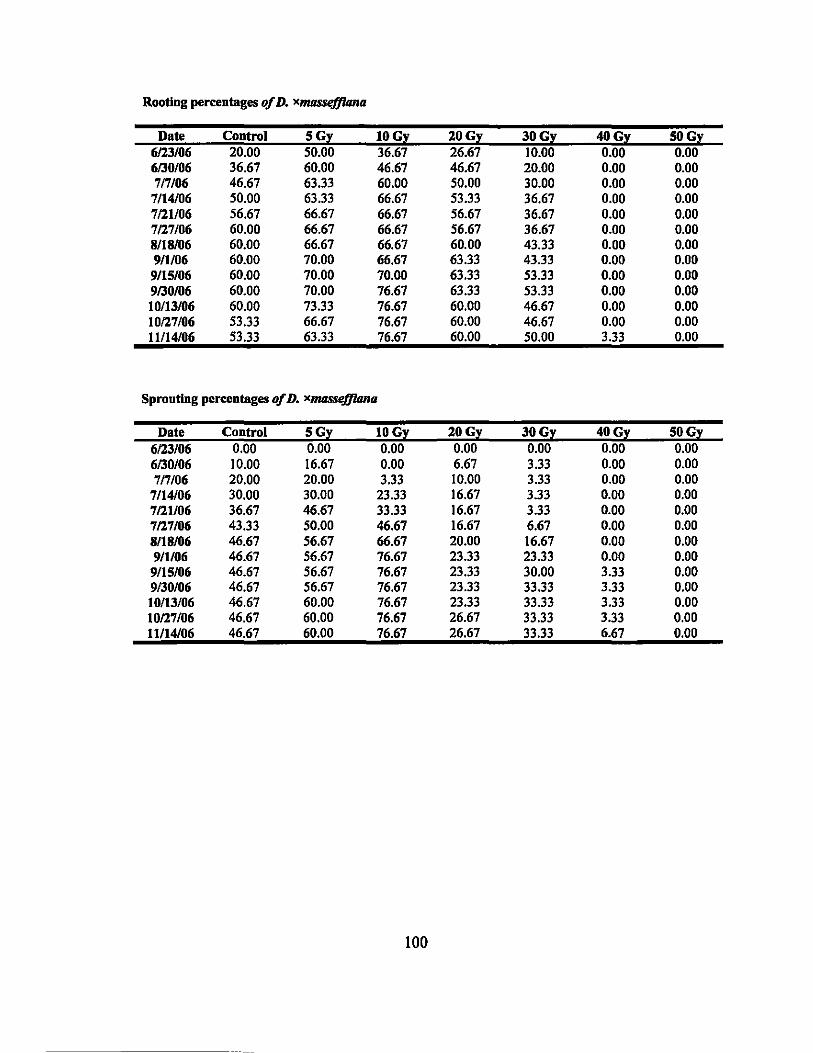
Rooting percentages of D. )(massejJUma
Date Control SGy lOGy lOGy 30Gy 40Gy SOGy 6123/06 20.00 50.00 36.67 26.67 10.00 0.00 0.00 6/30/06 36.67 60.00 46.67 46.67 20.00 0.00 0.00 717106 46.67 63.33 60.00 50.00 30.00 0.00 0.00
7114/06 50.00 63.33 66.67 53.33 36.67 0.00 0.00 7121106 56.67 66.67 66.67 56.67 36.67 0.00 0.00 7127106 60.00 66.67 66.67 56.67 36.67 0.00 0.00 8/18106 60.00 66.67 66.67 60.00 43.33 0.00 0.00 9/1/06 60.00 70.00 66.67 63.33 43.33 0.00 0.00 9/15/06 60.00 70.00 70.00 63.33 53.33 0.00 0.00 9130/06 60.00 70.00 76.67 63.33 53.33 0.00 0.00 10/13/06 60.00 73.33 76.67 60.00 46.67 0.00 0.00 10127106 53.33 66.67 76.67 60.00 46.67 0.00 0.00 1lI14/06 53.33 63.33 76.67 60.00 50.00 3.33 0.00
Sprouting percentages of D. )(massejJUma
Date Control SGy lOGy lOGy 30Gy 40Gy SOGy 6123/06 0.00 0.00 0.00 0.00 0.00 0.00 0.00 6130/06 10.00 16.67 0.00 6.67 3.33 0.00 0.00 717106 20.00 20.00 3.33 10.00 3.33 0.00 0.00
7114/06 30.00 30.00 23.33 16.67 3.33 0.00 0.00 7121/06 36.67 46.67 33.33 16.67 3.33 0.00 0.00 7127106 43.33 50.00 46.67 16.67 6.67 0.00 0.00 8118106 46.67 56.67 66.67 20.00 16.67 0.00 0.00 9/1/06 46.67 56.67 76.67 23.33 23.33 0.00 0.00
9/15/06 46.67 56.67 76.67 23.33 30.00 3.33 0.00 9130/06 46.67 56.67 76.67 23.33 33.33 3.33 0.00 10/13/06 46.67 60.00 76.67 23.33 33.33 3.33 0.00 10127106 46.67 60.00 76.67 26.67 33.33 3.33 0.00 11114/06 46.67 60.00 76.67 26.67 33.33 6.67 0.00
100
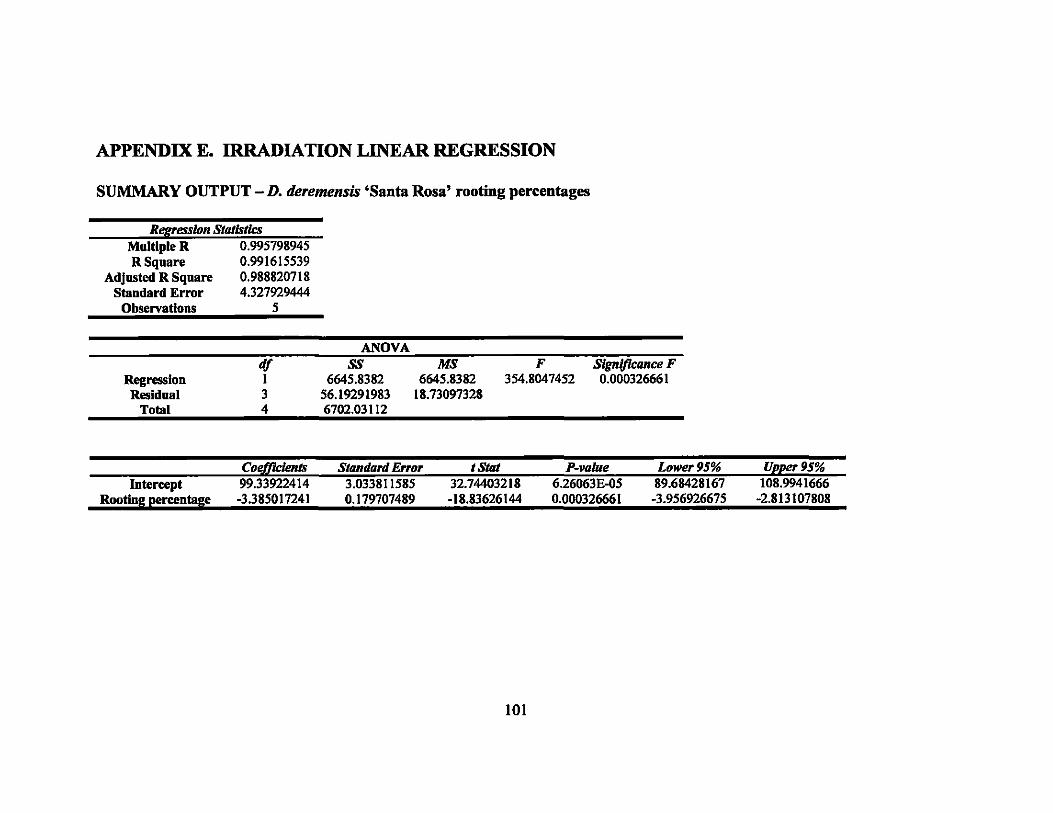
APPENDIX E. IRRADIATION LINEAR REGRESSION
SUMMARY OUTPUT - D. deremensis 'Santa Rosa' rooting percentages
Regression StatIstics Multiple R 0.995798945 R Square 0.991615539
Adjusted R Square 0.988820718 Standard Error 4.327929444
Observations 5
Regresslou Residual
Total
df I 3 4
ANOVA SS
6645.8382 56.19291983 6702.03112
MS 6645.8382
18.73097328
F 354.8047452
SlgnJfkance F 0.000326661
Coeflklents Standard E"or t Stat P-vaJue Lower 95% Upper 95% Intercept 99.33922414 3.033811585 32.74403218 6.26063E-05 89.68428167 108.9941666
Rooting percentage -3.385017241 0.179707489 -18.83626144 0.000326661 -3.956926675 -2.813107808
101
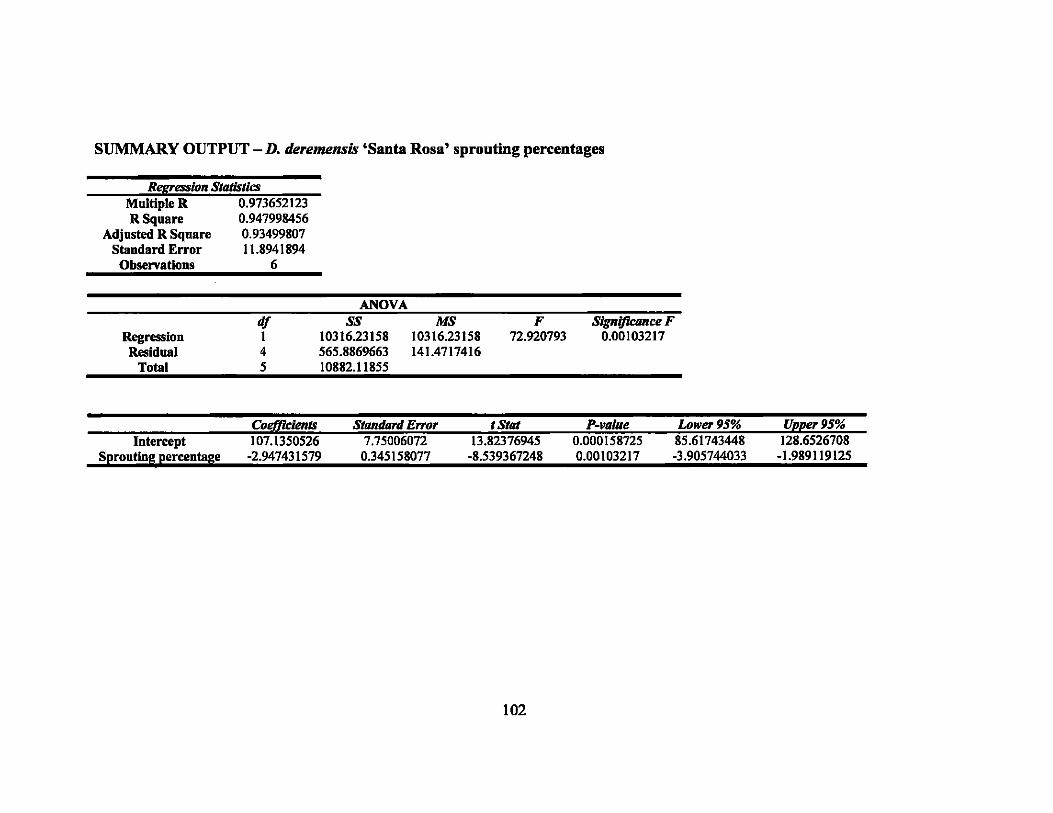
SUMMARY OUTPUT - D. deTemensis 'Santa Rosa' sprouting percentages
Regression Statistics Multiple R 0.973652123 R Square 0.947998456
Adjusted R Square 0.93499807 Standard Error 11.8941894
Observations 6
Regression Residual
Total
df 1 4 5
ANOVA ss
1031623158 565.8869663 10882.11855
MS 10316.23158 141.4717416
F 72.920793
Slgnijkance F 0.00103217
Cod/1cients StondJlrd E"or t Stat P-vallle Lower 95% Upper 95% Intercept 107.1350526 7.75006072 13.82376945 0.000158725 85.61743448 128.6526708
SprontingpercentaGe -2.947431579 0.345158077 ·8.539367248 0.00103217 -3.905744033 -1.989119125
102
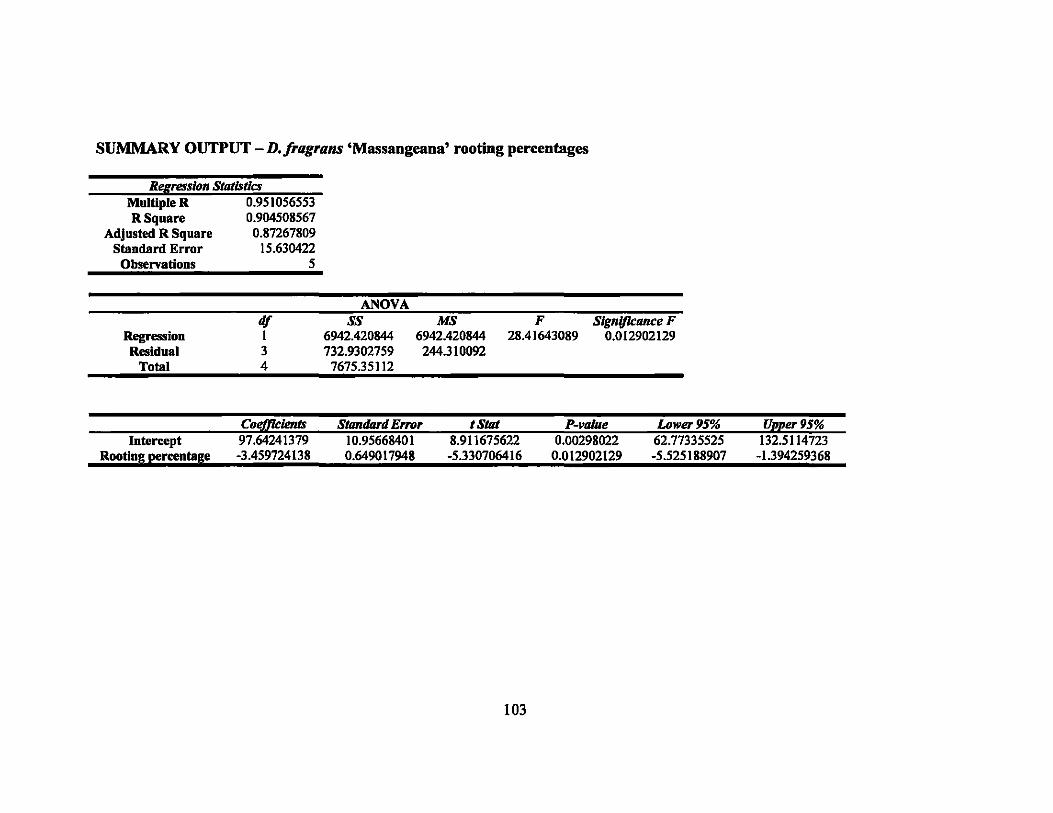
SUMMARY OUTPUT - D. fragrans 'Massangeana' rooting percentages
Regression Statistics Multiple R 0.951056553 R Square 0.904508567
Adjusted R Square 0.87267809 Standard Error 15.630422
Observations 5
Regression Residual
Total
Intercept
df 1 3 4
Rooting percentage -3.459724138
ANOVA SS
6942.420844 732.9302759
7675.35112
0.649017948
MS 6942.420844 244.310092
F 28.41643089
Slgnljkance F 0.012902129
-5.330706416 0.012902129 -5.525188907
103
-1.394259368
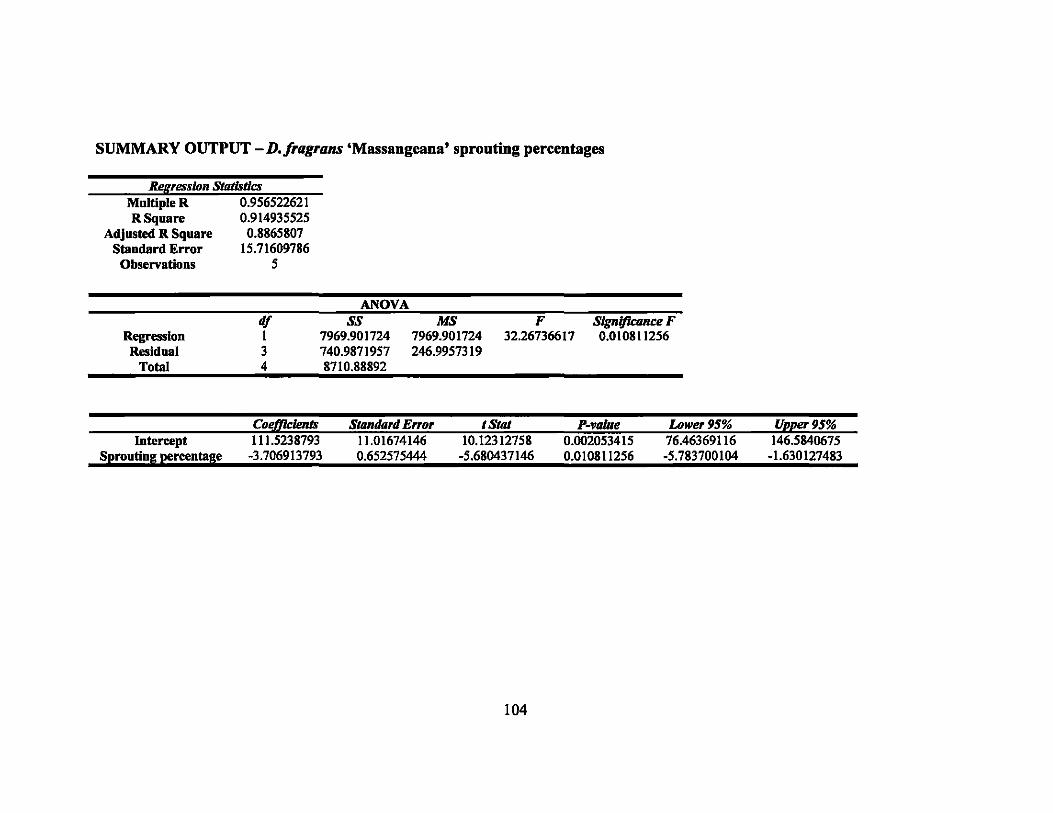
SUMMARY OUTPUT - D.fragrans 'Massangeana' spronting percentages
R£gression Statistics Multiple R 0.956522621 R Square 0.914935525
Adjusted R Square 0.8865807 Standard Error 15.71609786
Observations 5
Regression Residual
Total
Intercept
d/ 1 3 4
ANOVA SS
7969.901724 740.9871957 8710.88892
MS 7969.901724 246.9957319
F 32.26736617
Significance F 0.010811256
Sprouting percentage -3.706913793 1 0.652575444 -5.680437146
0.002053415 0.010811256 -5.783700104
104
-1.630127483
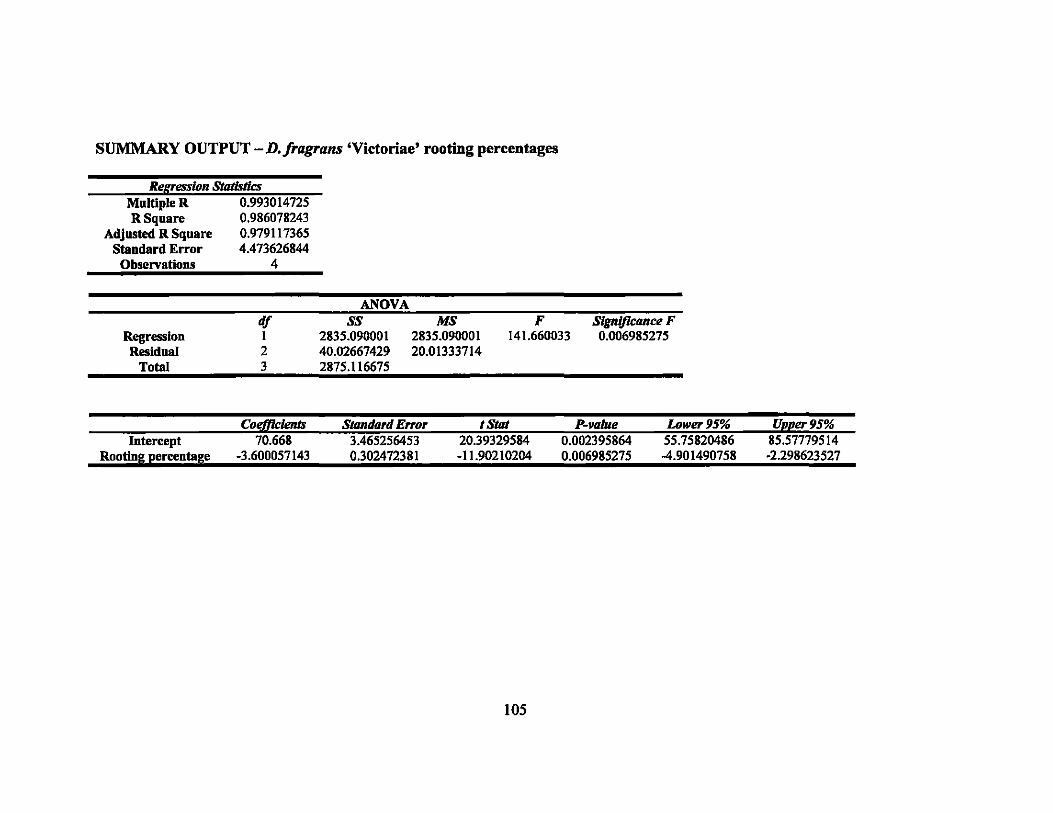
SUMMARY OUTPUT -D.fragrans 'Victoriae' rooting percentages
Regression StatIstics Multiple R 0.993014725 R Square 0.986078243
Adjusted R Square 0.979117365 Staudard Error 4.473626844
Observations 4
Regression Residual
Total
Intercept
df 1 2 3
Rootlnll percentalle -3.600057143
ANOVA ss
2835.090001 40.02667429 2875.116675
MS 2835.090001 20.01333714
F 141.660033
SlgnlJkance F 0.006985275
0.302472381 -11.90210204 0.006985275 -4.901490758
105
-2.298623527
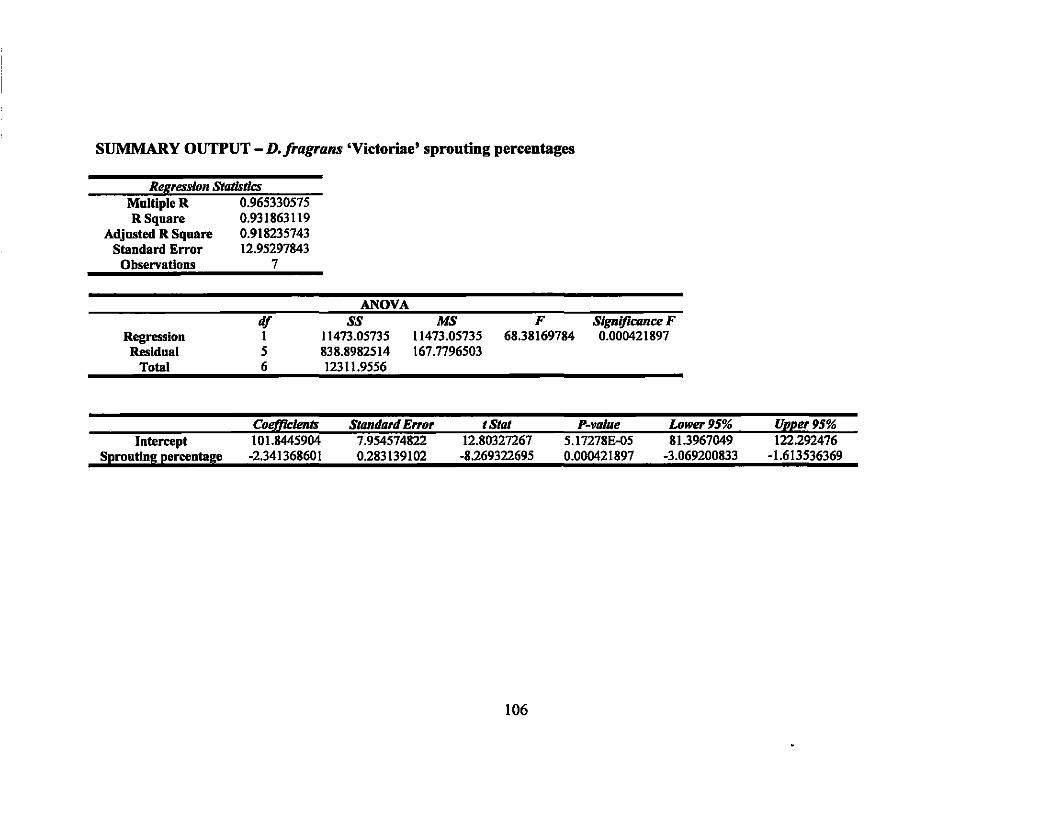
SUMMARY OUTPUT - D. fragrans 'Victoriae' sprouting percentages
Regresaion Statistics Multiple R 0.965330575 R Square 0.931863119
Adjusted R Square 0.918235743 Standard Error 12.95297843
Observatious 7
Regression Residual
Total
df 1 5 6
ANOVA SS
11473.05735 838.8982514 12311.9556
MS 11473.05735 167.7796503
F 68.38169784
Signijicance F 0.000421897
CoeJJkienls Sttmdard E"or t Stal P-vaJue Lower 95% Upper 95% Intercept 101.8445904 7.954574822 12.80327267 5.17278E-05 81.3967049 122.292476
Sprouting percentage -2.341368601 0.283139102 -8.269322695 0.000421897 -3.069200833 -1.613536369
106
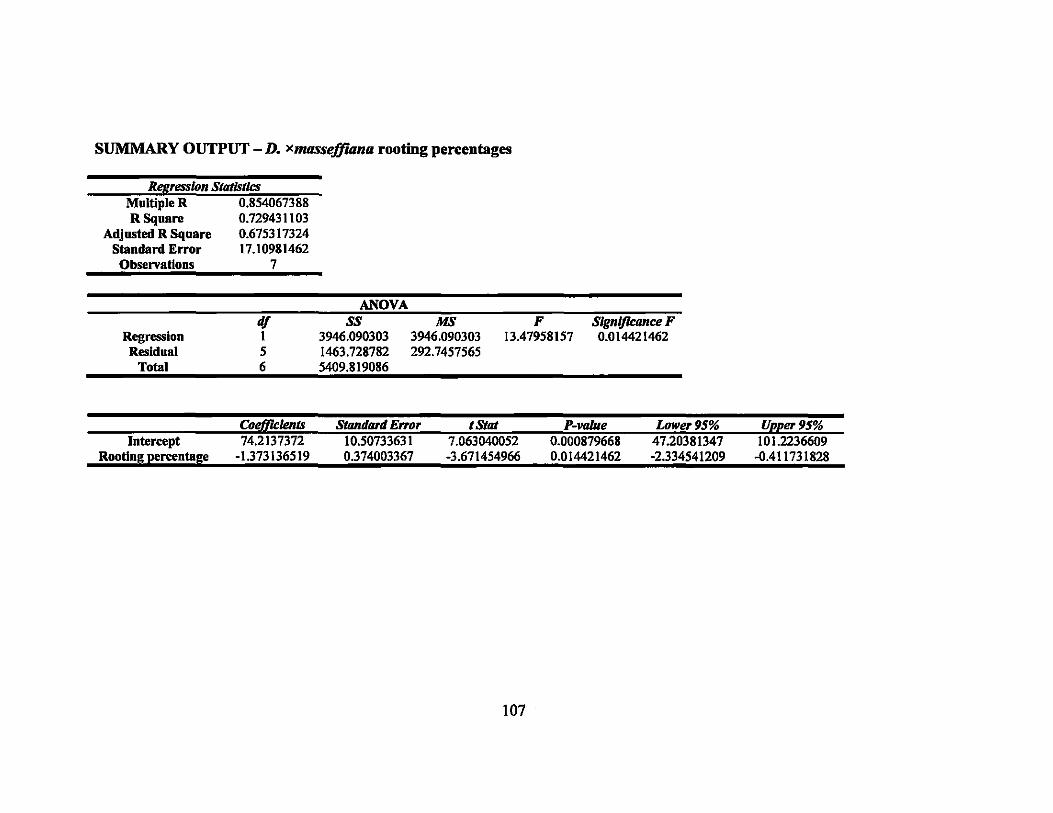
SUMMARY OUTPUT - D. x11UlSseffiana rooting percentages
Regression StatistiJ:s Multiple R 0.854067388 R Square 0.7294311 03
Adjusted R Square 0.675317324 Standard Error 17.10981462
Observations 7
Regression Residual
Total
dl I 5 6
ANOYA SS
3946.090303 1463.728782 5409.819086
MS 3946.090303 292.7457565
F 13.47958157
Slgnlflcance F 0.014421462
Co~1ents Standard E"or t StIlt P-vaJue Lower 95% Upper 95% Intercept 74.2137372 10.50733631 7.063040052 0.000879668 47.20381347 101.2236609
Rooting percentage ·1.373136519 0.374003367 ·3.671454966 0.014421462 ·2.334541209 -0.411731828
107
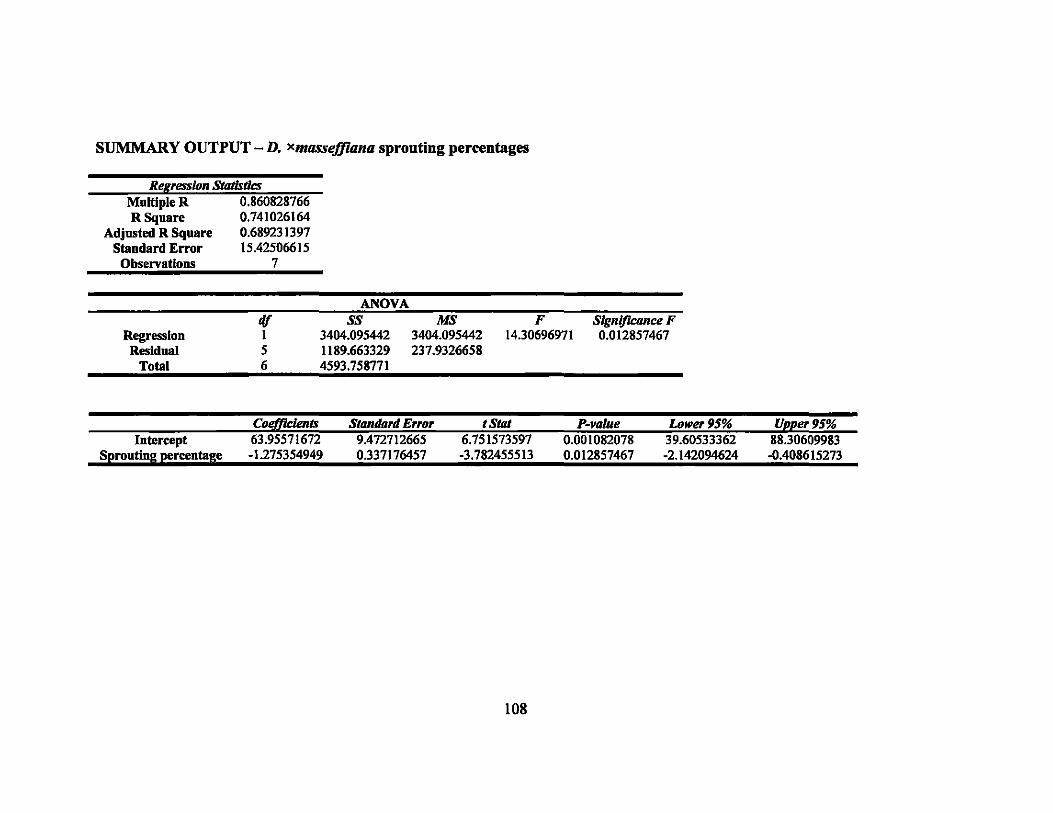
SUMMARY OUTPUT - D. x11UISseffiana sprouting percentages
Rl!f(1'ession StotlstJcs Multiple R 0.860828766 R Square 0.741026164
Adjnsted R Square 0.689231397 Standard Error 15.42506615
Observations 7
Regresslou Residual
Total
df 1 5 6
ANOVA SS
3404.095442 1189.663329 4593.758771
MS 3404,095442 237.9326658
F 14.30696971
Signlj1cance F 0.012857467
Cotdfidenls Standard E"or t Stat P-vaJue Lower 95% Upper 95% Intercept 63.95571672 9.472712665 6.751573597 0.001082078 39.60533362 88.30609983
Sprouting percentaGe -1275354949 0.337176457 -3.782455513 0.012857467 -2.142094624 -0.408615273
108
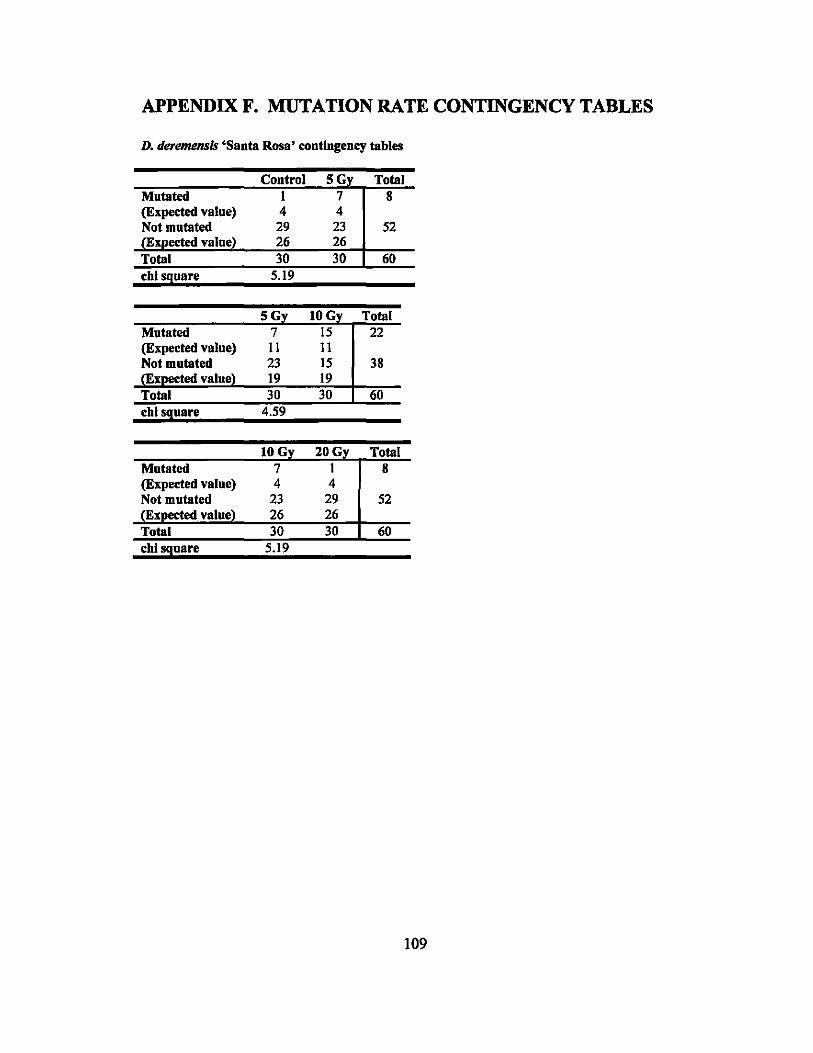
APPENDIX F. MUTATION RATE CONTINGENCY TABLES
D. deremensfs 'Santa Rosa' contingency tables
Control 5G Total Mutated I 7 8 (Expected value) 4 4 Not mutated 29 23 52
26 26 30 30 60
chi !9uare 5.19
5G Total Mutated 7 15 22 (Expected value) 11 11 Not mutated 23 15 38
19 19 30 30 60
chi !9uare 4.59
lOG Total Mutated 7 8 (Expected value) 4 Not mutated 23 52
26 30 30 60
chi !9nare 5.19
109
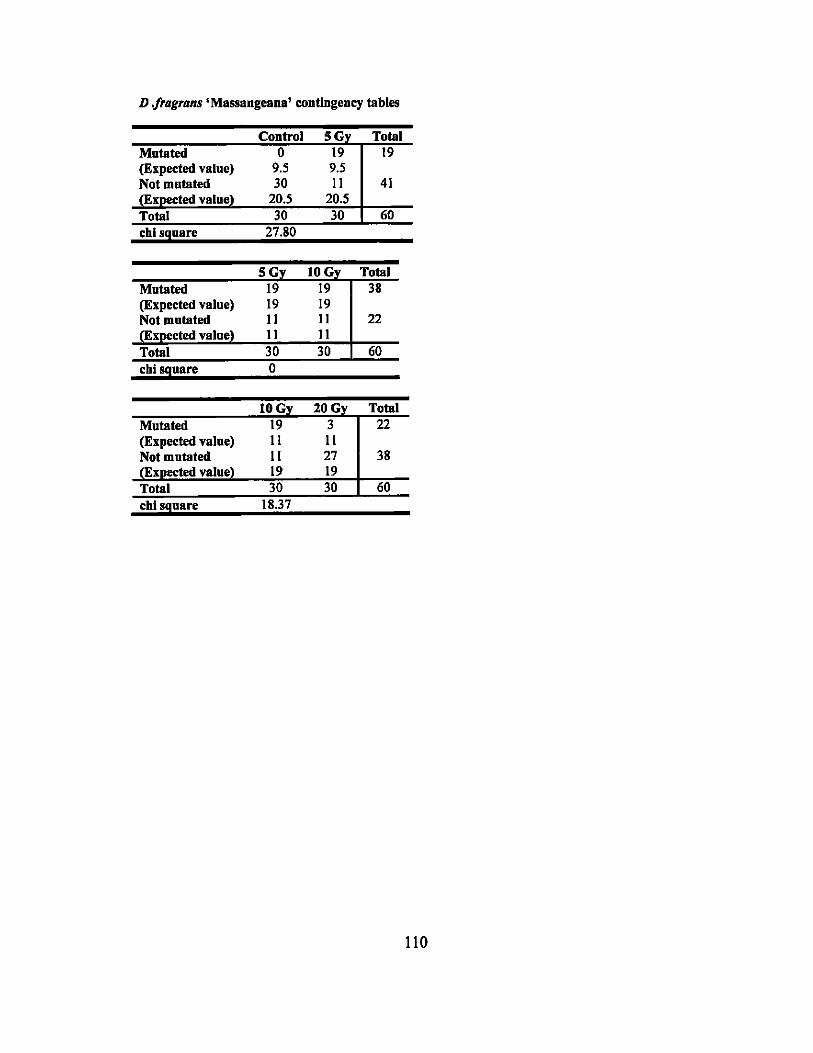
D "ragrans • Massangeana' contingency tables
Control SG Total Mntated 0 19 19 (Expected valne) 9.5 9.5 Not mutated 30 II 41
20.5 20.5 30 30 60
chls9uare 27.80
SG lOG Total Mutated 19 19 38 (Expected value) 19 19 Not mutated II II 22
II II 30 30 60
chl~nare 0
lOG 20G Total Mntated 19 3 22 (Expected value) II II Not mutated II 27 38
19 19 30 30 60
chi~uare 18.37
110
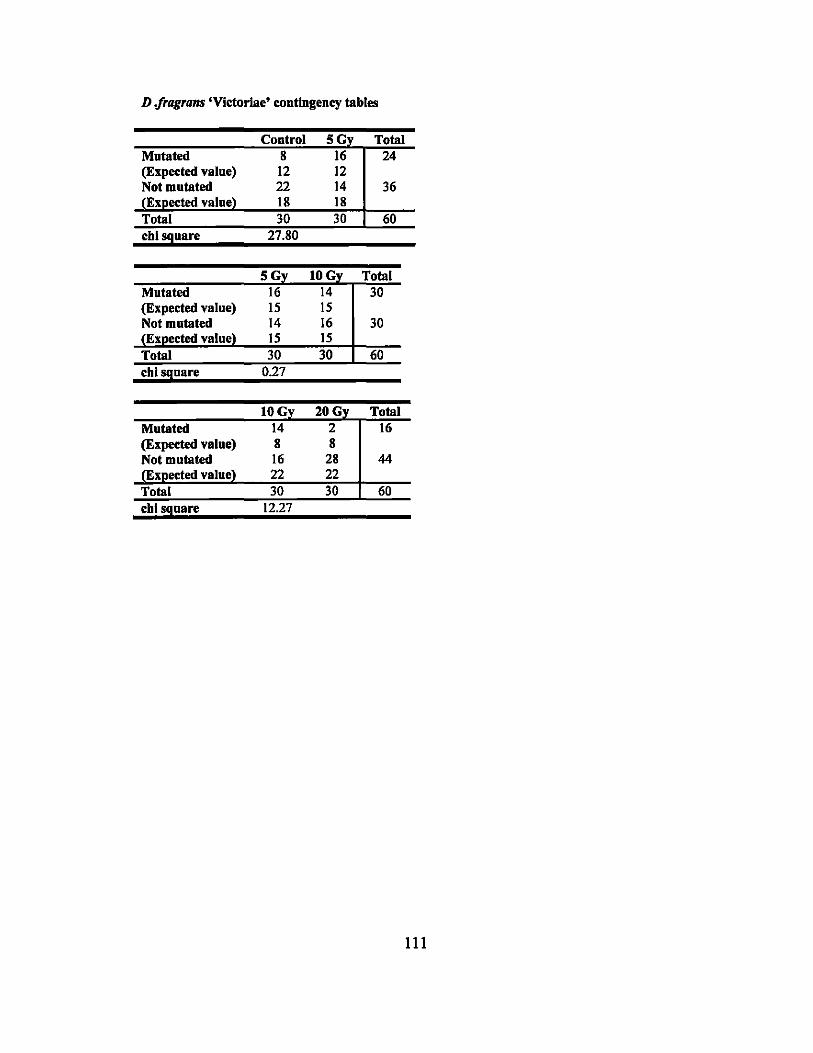
D .jragrans 'Victoriae' contingency tables
Control 5G Total Mutated 8 16 24 (Expected value) 12 12 Not mutated 22 14 36
18 18 30 30 60
cbl s9uare 27.80
5G Total Mutated 16 14 30 (Expected value) 15 15 Not mutated 14 16 30
15 15 30 30 60
cbi;:9uare 0.27
lOG Total Mutated 14 2 16 (Expected value) 8 8 Not mutated 16 28 44
22 22 30 30 60
cbls9uare 12.27
111
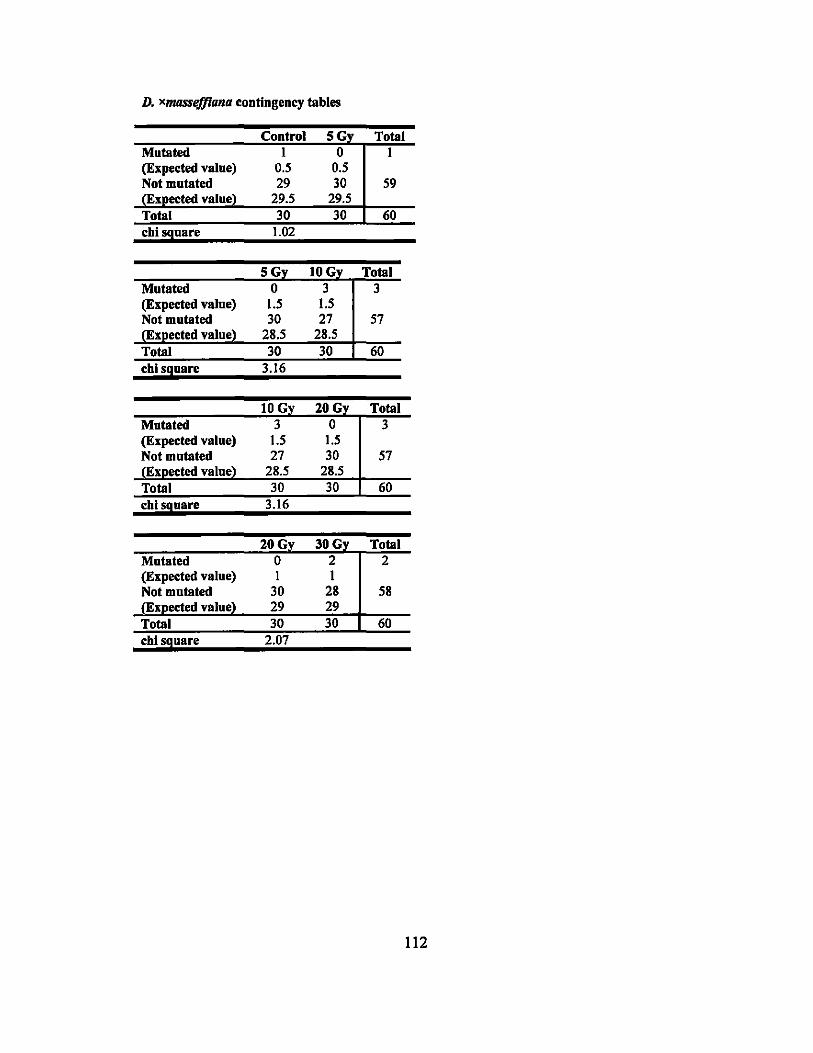
D. xmasselfiana contingency tables
Control SG Total Mntated I 0 I (Expected value) 0.5 0.5 Not mutated 29 30 59
29.5 29.5 30 30 60
chi !9uare 1.02
SG Total Mutated 0 3 (Expected value) 1.5 Not mutated 30 57
28.5 30 60
chi s9uare 3.16
10 G Total Mutated 3 3 (Expected value) 1.5 Not mutated 27 57
value 28.5 30 60
chi !9uare 3.16
lOG 30G Total Mutated 0 2 2 (Expected value) I I Not mutated 30 28 58
29 29 30 30 60
chi s9uare 2.07
112

LITERATURE CITED
Ackerman, W. L. and H. Dermen. 1996. A fertile colchiploid from a sterile interspecific Camellia hybrid. J. Hered. 63 (2):55-59.
Alamn, S. and C. R. McDavid. 1978. Promotion of flowering in edible aroids by gibberellic acid. Trop. Agr. 55 (1 ):81-86.
Ashikari, M. and M. Matsuoka. 2002. Application of rice genomics to plant biology and breeding. Bot Bul. Acad. Sinica. 43 (1): 1-11.
Banerji, B. K. and S. K. Datta. 1986. Induction of single flower mutant in Hibiscus cv. 'Alipur Beauty'. J. Nucl. Agr. BioI. 15 (4):237-240.
Banerji, B. K. and S. K. Datta. 1987. Gamma my induced mutation in double bmcted Bougainvillea cv. 'Los Banos Beauty'. J. Nucl. Agr. BioI. 16:48-50.
Banerji, B. K. and S. K. Datta. 1990. Induction of somatic mutation in Chrysanthemum cultivar 'Anupam'. J. Nucl. Agr. BioI. 19:252-256.
Banerji, B. K., A. K. Dwivedi and S. K. Datta. 1996. Gamma irmdiation studies on rose and chrysanthemum. J. Nucl. Agr. BioI. 25 (2):63-67.
Banerji, B. K., P. Nath and S. K. Datta. 1987. Mutation breeding in double-bracted Bougainvillea cv. 'Roseville's Delight'. 1. Nucl. Agr. BioI. 16:45-47.
Barker, R. E., 1. A. Kilgore, R. L. Cook and A. E. Garay. 1998. Validity of flow cytometry for testing ploidy level in ryegmsses. <http://cropandsoil.oregonstate.edulseed-extlPub/1998/SR9919.pdf>.
Basiran, M. N. and S. Ariffin. 2002. The progress and potentials of mutation induction in vegetatively propagated plants in Malaysia. 06 March 2006. <http://www.fuca.jp/englishle 0ldl2 totuzenheni/3/2002ws/04/04malaysialmain.html>.
Beattie, D. J. and E. J. Holcomb. 1983. Effects of chilling and photoperiod on forcing Astilbe. HortScience. 18 (4):449-450.
BerthoD, F. 1975. Method of obtaining polyploids in the genns Coffia by localized colchicine treatments of the buds. Cafe caeoa the. 19 (3):197-202.
Biswas, A. K. and N. K. Bhattacharyya. 1971. Induced polyploidy in legumes I. Cyamopsis psoaloides DC. Cytologia. 36:469-479.
Bos, J. J. 1984. Dmcaena in west Africa. Agr. Univ. Wageningen Papers. 84 (1):1-126.
113

Broertjes, C., P. Koene and T. H. Pronk. 1983. Radiation-induced low-temperature tolerant cultivars of Chrysanthemum morifolium Ram. Euphytica 32:97-101.
Broertjes, C., P. Koene and J. W. H. v. Veem. 1980. A mutant of a mutant of a mutant of .... :lrradiation of progressive radiation-induced mutants in a mutation-breeding programme with Chrysanthemum morifolium Ram. Euphytica 29:525-530.
Calabrese, E. J. and L. A. Baldwin. 2000. Radiation hormesis: its historical foundations as a biological hypothesis. Human Expt Toxico!. 19:41-75.
Casarett, A. P. 1968. Radiation Biology. Prentice-Hall, Inc., Englewood Cliffs, N.J.
Chalak, L. and J. M. Legave. 1996. Oryzalin combined with adventitious regeneration for an efficient chromosome doubling oftrihaploid kiwifruit. Plant Cell. Rpt. 16:97-100.
Chen, J., R. J. Henny and D. B. McConnell 2002. Development of new foliage plant cultivars. p.466-472. In: Janick, J. a. A. W. (eds). Trends in New Crops and New Uses. ASHS Press. Alexandria, VA.
Chua, B. M., J. T. Kunisaki and Y. Sagawa. 1981. In vitro propagation of Dracaena marginata'Tricolor'. HortScience. 16 (4):494.
Cialone, J. 1984. Developments in dracaena production. Combined Proc. - Inti. Plant Prop. Soc. 34:491-494.
Corr, B. E. and R. E. Widmer. 1987. Gibberellic acid increases flower number in Zantedeschia elliottiana and Z. rehmannii. HortScience. 22 (4):605-607.
Cuevas-Ruiz, J. 1973. Effect of gamma irradiation on sweet potato stern cuttings. J. Agr. Univ. P.R. 57 (4):351-354.
Debergh, P. 1975. Intensified vegetative multiplication of Dracaena deremensis. Acta Hort. 54:83-92.
Dewey, D. R. 1980. Some applications and misapplications of induced polyploidy to plant breeding. p.445-470. In: Lewis, W. H. (eds). Polyploidy: Biological Relevance. Plenum Press. New York. .
Dnyansagar, V. R. and R. S. Nadkarni. 1983. Induced tetraploidy in Crotolariajuncea Linn. Cytologia. 48:483-489.
Dolozel, J. 1991. Flow cytometric analysis of nuclear DNA content in higher plants. Phytochem. Anal. 2:143-154.
114

Donini, P. and A. Sonnino 1998. Induced mutation in plant breeding: current status and future outlook. p.255-291. In: Jain, S. M., D.S. Brar, and B.S. Ahloowalia (eds). Somaclonal Variation and Induced Mutations in Crop Improvement. KIuwer Academic Publishers. Dordrecht.
Doorenbos, J. and J. J. Karper. 1975. X-ray induced mutations in Begonia x Hiemalis. Euphytica 24:13-19.
Ehrenberg,1. 1955. Factors influencing radiation induced lethality, sterility, and mutations in barley. Hereditas. 41:123-146.
Evans, M. R., H. F. Wilkins and W. P. Hackett. 1992. Gibberellins and temperature influence long-day floral initiation in Poinsettia J. Amer. Soc. Hort. Sci. 117 (6):966-971.
FAO and IAEA. 1977. Manual on Mutation Breeding, Second Edition; Technical Reports Series No. 119. International Atomic Energy Agency, Vienna.
Fereol, L., S. Louis and 1. Luce. 1996. Effects of gamma radiation on in vitro plantlets ofAlpiniapupurata. Journal ofHorticultura1 Science. 71 (2):243-247.
Fisher, J. B. 1980. Gibberellin-induced flowering in Cordyline (Agavaceae). J. Expt. Bot. 31 (122):731-735.
French, C. J. and J. Alsbury. 1988. Effect of pre-force storage conditions on early flowering of Rhododendron. HortScience. 23 (2):356-358.
Fulton, T. A., A. J. Hall and J. L. Catley. 2001. Chilling requirements of Paeonia cultivars. Scientia Hort. 89:237-248.
Gao, S. 1., D. N. Zhu and D. R. Xu. 1996. Autotetraploid plants from colchicine-treated bud culture of Salvia miltio"hiza Bge. Plant Cell, Tissue and Organ Cult. 47:73-77.
Gupta, M. N., V. Laxmi, B. S. Dixit and S. N. Srivastava. 1982. Gamma ray induced variability in Costus speciosus. Progressive Hort. 14 (4):193-197.
Gupta, R. and P. K. Gupta. 1975. Induced polyploidy in Crotolaria 1. I. C. Juncea and C. Retusa. J. Indian Bot. Soc. 54:175-182.
Han, L., G. Zhao, Y. Li, H. Zhang and X. Fan. 1998. Study on inducing polyploid of Actinidia sp. J. Fruit Sci. 15 (3):273-276.
Hancock. J. F. 1997. The colchicine story. HortScience. 32 (6):1011-1012.
115

Hansen, N. J. P. and S. B. Andersen. 1996. In vitro chromosome doubling potential of colchicine, oryzalin, triflura1in and APM in Brassica napus microspore culture. Euphytica. 88:159-164.
Harbaugh, B. K. and G. J. Wilfret. 1979. Gibberellic acid (GA3) stimulates flowering in Caladium hortulanum Birdsey. HortScience. 14 (1):72-73.
Hartmann, H. T., D. E. Kester, F. T. Davies and R. L. Geneve. 2001. Hartmann and Kester's Plant Propagation: Principles and Practices. Prentice Hall, Englewood Cliffs, NJ.
Hawaii Department of Agriculture. 2006. Hawaii Flowers and Nursery Products Annual Summary. United States Department of Agriculture National Agricultural Statistics Service, Washington, D.C.
Henny, R. J. 1980. Gibberellic acid (GAl) induces flowering in Dieffenbachia maculata 'Perfection'. HortScience. 15 (5):596-597.
Henny, R. J. 1981. Promotion of flowering in Spathiphyllum 'Mauna Loa' with gibberellic acid. Hereditas. 16 (4):554-555.
Henny, R. J. 1983. Flowering of Aglaonema commutatum 'Treubii' following treatment with gibberellic acid. HortScience. 18 (3):374.
Henny, R. J. 1988. Inducing flowering of Homalomena lindenii (Rodigas) Ridley with gibberellic acid. HortScience. 23 (4):711-712.
Henny, R. J. 1991. Flowering of anthurium following treatment with gibberellic acid. HortScience. 27 (12):1328.
Henny, R. J. and J. Chen. 2003. Cultivar development of ornamental foliage plants. Plant Breeding Rev. 23:245-291.
Henny, R. J. and R. L. Hamilton. 1992. Flowering of Anthurium following treatment with gibberellic acid. HortScience. 27 (12):1328.
Henny, R. J., D. J. Norman and M. E. Kane. 1999. Gibberellic acid-induced flowering of Syngonium podophyllum Schott 'White Butterfly'. HortScience. 34 (4):676-677.
Jambhale, N. D. a. Y. S. N. 1983. Induction of polyploidy in the Abelmoschus species hybrids. Progressive Hort. 15:157-158.
Jones, R. L. 1973. Gibberel1ins: Their physiological role. Annu. Rev. Plant Physio!. 24:571-578.
116

Kamenetsky, R., A. Barzilay, A. Erez, and A.H. Halevy. 2003. Temperature requirements for floral development of herbaceous peony cv. 'Sarah Bernhardt'. Scientia Hort. 97:309-320.
Kane, M. E., G. 1. Davis, R. D. Hoffuer and R. J. Henny. 1995. Gibherellins promote flowering in two Cryptocoryne species. HortScience. 30 (2):380.
King, R. W. 2003. Unlocking the hormonal and molecular controls of flowering. Acta Hort. 624:139-146.
Lapade, A. G., A. M. S. Veluz, 1. J. Marhella, A. C. Barrida and M. G. Rama. 2002. Status of mutation breeding in vegetatively propagated crops in the Philippines. 06 March 2006. <http://www.fnca.jp/englisb/eoldl2totuzenheni/3/2002wsl04/05philippineimain.htm1>.
Larkin, P. 1998. Introduction. p.3-13. In: Jain, S. M., D.S. Brar, and B.S. Ahloowa1ia (eds). Somaclonal Variation and Induced Mutations in Crop Improvement. Kluwer Academic Publishers. Dordrecht.
Lu, W. 2002. Direct regeneration of inflorescence from callus in Dracaena fragrans cv. Massangeana Hort. Acta Botanica Sinica. 44 (1):113-116.
Lu, W. 2003. Control of in vitro regeneration of individual reproductive and vegetative organs in Dracaenafragrans cv. massangeana Hort. - regularities of the direct regeneration of individual organs in vitro. Acta Botanica Sinica. 45 (12):1453-1464.
Mathew, P. M. and B. Vijayavalli. 1989. Cytology of species of Cordy line and Dracaena from South India. Cytologia. 54:573-579.
Menninger, E. D. 1965. The culture of miniature-flowered cymbidiums. Amer. Orchid Soc. Bull. 34:196-199.
Miller, M. W. and W. M. Miller. 1987. Radiation hormesis in plants. Health Physics. 52 (5):607-616.
Miyahara, K. 1997. Mutation induction in rice by soft x-ray irradiation. Technical News -Institute of Radiation Breeding. 58:2.
Nair, M. K. and M. J. Ratnambal. 1974. Colchicine induced tetraploids in the Areca palm Areca catechu 1. J. Plantation Crops. 2 (1):7-9.
Nell, T. A., J. W.H. Bodnaruk, J. N. Joiner and T. J. Sheehan. 1983. Ethylene evolution and flowering of cold- and GA-treated'Redwing' azaleas. HortScience. 18 (4):454-455.
117

Nybom, N. 1961. The use of induced mutations for the improvement of vegetatively propagated plants. p. 253-294. In: (eds). Symposium on Mutation and Plant Breeding Pub. 891. National Academy of Sciences-National Research Council. Washington, D.C.
Pearson, S., A. Parker, P. Hadley and H. M. Kitchener. 1995. The effect of photoperiod and temperature on reproductive development of Cape Daisy (Osteospermum Jucundum cv. 'Pink Whirls'). Scientia Hort. 62:225-235.
Petersen, K. K., P. Hagberg and K. Kristiansen. 2003. Colchicine and oryza1in mediated chromosome doubling in different genotypes of Miscanthus sinensis. Plant Cell, Tissue and Organ Cult. 73:137-146.
Ponnuswami, V. 1991. Sensitivity of Muscat grape cuttings to gamma irradiation. South Indian Hort. 39 (5):317-318.
Rao, D. V. R. 1999. Effect of gamma irradiation on growth, yield and quality of turmeric. Adv. Hort. For. 6:107-110.
Rood, S. B., T. 1. Blake and R. P. Pharis. 1983. Gibbere11ins and heterosis in maize. II. Response to gibberellic acid and metabolism of [3H] gibberellin A20. Plant Physio!. 71 (3):645-651.
Rose, J. B., J. Kubba and K. R Tobutt. 2001. Induction oftetraploids for breeding hardy ornsrnentals. Acta Hort. 560: 1 09-1I2.
Rnnk1e, E. S., R. D. Heins, A. C. Cameron and W. H. Carlson. 1999. Cold treatment modifies the photoperiodic flowering response of Lobelia xspeciosa. Scientia Hort. 80:247-258.
Sadasivam, A. and o. P. Saxena. 1998. Gibberellic acid-like substances and heterosis in pearl millet. Indian J. Environ. Toxico!. 8 (2):66-68 (abstr.).
Sagan, L. A. 1987. What is hormesis and why haven't we heard about it before? Health Physics. 52 (5):521-525.
Sangowawa, B. B. 1994. Effect of colchicine in a non-tuberous Solanum species. The Nucleus. 37:6-10.
Schum, A. 2003. Mutation breeding in ornsrnentals: an efficient breeding method? Acta Hort. 612:47-53.
Schum, A. and W. Preil 1998. Induced mutations in ornamental plants. p. 333-366. In: Jain, S. M., D. S. Brar and B. S. Ahloowalia (eds). Somaclonal Variation and Induced Mutations in Crop Improvement. Kluwer Academic Publishers. Dordrecht, England.
1I8

Sklar, L. A. 2005. Flow Cytometry for Biotechnology. Oxford University Press, New York.
Skok, J., W. Chorney and J. E.1. Rakosnik. 1965. An examination of stimulatory effects of ionizing radiation in plants. Radiation Bot. 5:281-292.
Sparrow, A. H. 1961. Types of ionizing radiation and their cytogenetic effects. p. 55-119. In: (eds). Symposium on Mutation and Plant Breeding, Pub. 891. National Academy of Sciences-National Research Council. Washington, D.C.
Sparrow, A. H., A. F. Rogers and S. S. Schwemmer. 1968. Radiosensitivity studies with woody plants - I. Acute gamma irradiation survival data for 28 species and predictions for 190 species. Radiation Bot. 8:149-186.
Sparrow, A. H., L. A. Schairer and R. C. Sparrow. 1963. Relationship between nuclear volumes, chromosome numbers, and relative radiosensitivities. Science. 141 (3576):163-166.
Staples, G. W. and D. R. Herbst. 2005. A Tropical Garden Flora: Plants Cultivated in the Hawaiian Islands and other Tropical Places. Bishop Museum Press, Honolulu.
Strachan, S. D. and F. D. Hess. 1983. The biochemica1mechanism of action of the dinitroaniline herbicide oryzalin. Pesticide Biochem. Physiol. 20: 141-150.
Takamura, T., K. B. Lim and 1. M. v. Tuyl. 2002. Effect of a new compound on the mitotic polyploidization of Lilium longijlorum and oriental hybrid lilies. Acta Hort. 572:37-42.
Thao, N. T. P., K. Ureshino, I. Miyajima, Y. Ozaki and H. Okubo. 2003. Induction of tetraploids in ornamental Alocasia through colchicine and oryza1in treatments. Plant Cell, Tissue and Organ Cult. 72: 19-25.
Tosca, A., R. Pandolfi, S. Citterio, A. Fasoli and S. Sgorbati. 1995. Determination by flow cytometry of the chromosome doubling capacity of colchicine and oryzalin in gynogenetic haploids of Gerbera. Plant Cell. Rpt. 14:455-458.
Tuyl, J. M. v., B. Meifer and M. P. v. Dien. 1992. The use of oryzalin as an alternative for colchicine in in vitro chromosome doubling of Lilium and Nerine. Acta Hort. 325:625-630.
Vainola, A. 2000. Polyploidization and early screening of Rhododendron hybrids. Euphytica. 112:239-244.
Vince-Prue, D. 1975. Photoperiodism in Plants. McGraw-Hill, London.
119

Wright, R. D. and D. C. Milbocker. 1979. Chilling requirements for flowering of Rex cornuta Lind!. Paxt. cv. Burfordii. HortScience. 14 (5):624-625.
Zeevart, J. A. D. 1976. Physiology of flowering formation. Annu. Rev. PlantPhysiol. 27:321-348.
120





























