Embed Size (px)
Citation preview

INFECTiON AND IMMUNITY, May 1994, p. 1726-1732 Vol. 62, No. 50019-9567/94/$04.00+0Copyright (© 1994, American Society for Microbiology
Structural and Functional Characterization of the HumanFormyl Peptide Receptor Ligand-Binding Region
STEPHEN J. RADEL,* ROBERT J. GENCO, AND ERNESTO DE NARDINDepartment of Oral Biology, State University ofNew York at Buffalo, Buffalo, New York 14214
Received 12 October 1993/Returned for modification 9 December 1993/Accepted 17 February 1994
The formyl peptide (N-formyl-1-methionyl-1-leucyl-1-phenylalanine [FMLP]) receptor is involved in theactivation of neutrophils and their subsequent response to chemotactic N-formylated peptides. Recently, wefound that the first extracellular loop closest to the N-terminal end of the FMLP receptor exhibited thestrongest ligand binding compared with that shown by other extracellular regions. By constructing amino acidsubstitutional variants of this domain, we have determined that residues Arg-84 and Lys-85 on this loop playmajor roles in ligand-binding activity. Furthermore, random rearrangement of the residues of this receptorregion demonstrated that the position of these charged amino acids did not affect their involvement in ligandbinding, although their presence was essential for this binding to occur. We propose that the portion of the firstN-terminal extracellular loop of the FMLP receptor containing residues Arg-84 and Lys-85 contributessignificantly to the active site in ligand-receptor binding. We further propose that this binding is not dependenton defined structure but rather that these charged moieties may function as important "contacts" inreceptor-ligand interactions.
Polymorphonuclear leukocytes (neutrophils) provide an im-portant host defensive mechanism against bacterial and fungalinfections. Neutrophils respond to inflammation both in vitroand in vivo through a chemotactic pathway of directed loco-motion towards a stimulus (22). N-formylated peptides such asN-formyl-1-methionyl-1-leucyl-1-phenylalanine (FMLP), be-lieved to be structural analogs of bacterial metabolic products,are potent chemoattractants for neutrophils (26, 33). In addi-tion to chemotaxis, a number of neutrophil functions, includingaggregation, phagocytosis, generation of cytotoxic superoxideanions, production of bioactive arachidonate metabolites, andproteolytic enzyme release, can be elicited by potent chemoat-tractants such as FMLP (3, 6, 23, 28, 32). Through the use ofradioiodinated cross-linking reagents and photoaffinity label-ing techniques, the human neutrophil FMLP receptor has beenreported to consist of three components, the predominantglycoprotein having a molecular weight of 55,000 to 70,000 (12,17, 25). Functional responses of neutrophils are the result ofligand binding to specific plasma membrane receptors andappear to be mediated through a transduction mechanisminvolving the activation of phospholipase C and specific gua-nine nucleotide-binding proteins (18), which are involved inthe receptor-mediated control of neutrophil function (19).Recently, the human FMLP receptor has been proposed to bea seven-transmembrane-domain structure characteristic of theG-protein-coupled receptor superfamily (7, 8).The production of synthetic peptides from the known amino
acid sequence of a biologically active protein allows thecharacterization of the immunological and functional domainsof that protein. This technique has been successfully utilized inthe identification of functional domains of receptors such asthe phosphorylcholine-binding C-reactive protein (38) and thefibronectin receptor (35). Synthetic peptides have also beenused for the functional mapping of the hamster 3-adrenergicreceptor (5), one of the G-protein-coupled receptors of the
* Corresponding author. Mailing address: Department of OralBiology, 210 Foster Hall, State University of New York at Buffalo,Buffalo, NY 14214. Phone: (716) 829-3518. Fax: (716) 829-3942.
rhodopsin family which is similar in structure to the formylpeptide receptor. Using synthetic peptides, we found that theprimary ligand-binding site of the FMLP receptor was local-ized to a 17-amino-acid (aa) stretch on the first extracellularloop of the N-terminal end of the receptor protein (31),although other portions of the molecule may contribute toligand binding. Recently, studies in which FMLP receptorchimeras were made also found that the major structuraldeterminant for high-affinity ligand binding was localized tothe first extracellular loop of the FMLP receptor (29, 30).
In this study, in order to pinpoint the amino acid residuesnecessary to maintain ligand-receptor binding, we have synthe-sized different peptides corresponding to amino acid variationsin this extracellular loop and have tested them for theirinvolvement in ligand binding. We provide evidence thatresidues 84 and 85, Arg-84 and Lys-85, of this first extracellularloop of the FMLP receptor molecule contribute to the activesite in formylated peptide binding to the receptor.
MATERIALS AND METHODS
Isolation of human neutrophils. Normal human neutrophilswere isolated from fresh buffy coats according to the methoddescribed by De Nardin et al. (12). The neutrophil yield fromone buffy coat (-400 ml) was 1.3 x 109 to 2.8 x 109 cells, witha viability >96% as assessed by the exclusion of trypan bluestain. Immunofluorescence staining, analyzed by flow cytom-etry with a FACScan (Becton Dickinson), demonstrated a cellpopulation that was greater than 99% neutrophils.
Preparation of neutrophil plasma membranes. Neutrophilplasma membranes were prepared by the methodology de-scribed by De Nardin and Genco (11), with modifications.Neutrophils (2.0 x 109 to 2.5 x 109) were suspended in aphosphate-buffered saline lysing buffer (pH 7.2) containing 2.5mM MgCl2, 1 mg of ovalbumin per ml, 2 mM phenylmethyl-sulfonyl fluoride, and 4 mM Not-p-tosyl-L-lysine chloromethylketone (TLCK) hydrochloride and then processed as describedby Goetzl et al. (17). The final membrane preparation wasresuspended in chromatography buffer containing 1% CHAPSdetergent (20 mM HEPES [N-2-hydroxyethylpiperazine-N'-2-
1726
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 24
Dec
embe
r 20
21 b
y 18
3.91
.203
.37.

NEUTROPHIL FMLP RECEPTOR LIGAND-BINDING REGION 1727
ethanesulfonic acid], 100 mM NaCl [pH 7.2]). To detectpotential proteolysis by neutrophil enzymes, '4C-bovine serum
albumin ("5 x 10" cpm) was added to a sample aliquot ofcells and subjected to the entire plasma membrane isolationprocedure at both 4°C and room temperature. No proteolysiswas observed at any of the steps.
Peptide synthesis. The peptides were synthesized by a
solid-phase peptide synthesis methodology on an AppliedBiosystems model 431A synthesizer, as described by Radel et
al. (31). The purity of the peptides was checked by amino acidanalysis and protein sequencing, and purification was by high-performance liquid chromatography (HPLC).
Purification of peptides, amino acid analysis, and sequenc-
ing. Synthetic peptides were purified by HPLC (Rainin Rabbit-HP) and then analyzed for amino acid composition (BeckmanSystem 6300 amino acid analyzer) and sequence (AppliedBiosystems model 471A sequencer) according to the methoddescribed by Radel et al. (31).
Photoaffinity ligand conjugation and radioiodination. Astable photoaffinity label (PAL) was prepared by conjugatingthe acetate form of the peptide FMLP-l-lysine (Sigma Chem-ical Co.) to the photoreactive cross-linker sulfosuccinimidyl-2-(p-azidosalicylamido)ethyl-1,3-dithiopropionate (Pierce Chemi-cal Co.) (2). Conjugation was carried out in aqueous medium bythe method described by Wollenweber and Morrison (39).Radioiodination of the PAL preparation was carried out by a
modification of the method described by Allen et al. (2).Briefly, 100 p.g of IODO-GEN (Pierce Chemical Co.) was
dissolved in 100 [l of chloroform and dried to a thin film undernitrogen in an amber vial. The PAL preparation was thenadded to the amber vial with 1 mCi (10 pLI) of carrier-freeNa-'251 (Amersham International), and the reaction was al-lowed to proceed for 30 min at room temperature. The samplewas purified on a Bio-Gel P-2 column (1.3 by 27 cm; 200 to 400mesh) by elution with 20 mM NaOH buffer at a rate of 0.2 to0.3 ml/min. Aliquots of 1.0-ml fractions were collected andanalyzed for activity. All procedures involving the synthesisand purification of the PAL were performed under light-reduced conditions.
Photoaffinity labeling of peptides and human neutrophilmembranes. Photoaffinity labeling of the synthetic peptidesand neutrophil membrane preparations were performed by themethod described by Radel et al. (31). Briefly, photoaffinitylabeling was carried out at room temperature for 30 min in thedark, using 5 to 10 pmol (_10" cpm) of '25I-PAL. Covalentbonding of the label was initiated by photolysis for 5 min underlong-wave UV light (Black-Ray; U-V Products Inc.). Thespecificity of labeling was assessed by studies of the inhibitionof binding of 125I-PAL to a separate aliquot of neutrophilmembrane by preincubation with unlabeled PAL or FMLP.Also, as a control, bovine serum albumin was incubated with'25I-PAL to assess nonspecific binding. No binding of '25I-PALto bovine serum albumin was observed. The reaction mixtureswere then analyzed by sodium dodecyl sulfate-polyacrylamidegel electrophoresis (SDS-PAGE) and autoradiography.PAGE. SDS-PAGE was carried out by the method of
Laemmli (20) in 10, 15, or 20% acrylamide "mini" slab gels(Hoefer Scientific). Separated protein components were visu-alized by Coomassie brilliant blue staining and autoradiogra-phy.
Densitometer analysis. Densitometer analysis was per-
formed on autoradiographs of the SDS-PAGE gels, using an
LKB Ultroscan laser densitometer (Pharmacia LKB). Ligandbinding was quantitated by using the Gel Scan XL softwareprogram (Pharmacia LKB), which calculated the area under
TABLE 1. Peptide inhibition of ligand ('125I-PAL) binding to the68-kDa membrane component from normal human neutrophils"
Amino acidsPeptide
Position Residues" Inhibition
2 84-100 RKAMGGHWPFGWFLCKF 812a 88-100 GGGGGGHWPFGWFLCKF 222b 84-95 RKAMGGHWPFGWGGGGG 692c 84-100 RKAMGGGWPFGWFLCKF 792d -C GFWLKWMRPKFAHFGGC 942e 51-62 AGFRMTHTVYTI 02f 72-83 FCFTSTLPFFMV 02g 85-100 GKAMGGHWPFGWFLCKF 602h 86-100 GGAMGGHWPFGWFLCKF 182i GFWLKWMGPKFAHFGGC 832j GFWLGWMGPGFAHFGGC 0
" Synthetic peptides at 200-p.M concentrations were incubated with 301-p.gsamples of solubilized neutrophil membranes, photolabeled with '251-PAL, andsubjected to SDS-PAGE and autoradiography. Densitometric analysis wasperformed on autoradiographs of the gels, using an LKB Ultroscan liaserdensitometer. The intensity of ligand binding to the 68-kDa neutrophil inem-brane protein was quantitated with the Gel Scan XL software program. Percentbinding inhibition was then calculated by comparing the strength of ligandbinding in the presence of the peptides versus that in their absence."Underlined G residues indicate where glycine substitutions were madc.-, random arrangement of amino acid residues of pcptide 2.
the peak generated from the intensity of the autoradiographicsignal.
RESULTS
Synthetic peptides. A number of peptides (Table 1) wereinvestigated for the ability to bind ligand directly and to inhibitthe binding of ligand to neutrophil membrane preparations.Most of these peptides represented amino acid residue varia-tions in the first extracellular loop near the N-terminal end ofthe receptor (residues 84 to 100, referred to as peptide 2),which has been shown to possess the major ligand-bindingportion of the FMLP receptor molecule (31). Also includedwere peptides which represented intracellular (peptide 2e) andtransmembrane (peptide 2f) regions, as well as a randomizedarrangement of the amino acid residues that make up peptide2 (peptide 2d).
Ligand-peptide inhibition studies. Studies were performedto determine if ligand binding to the 68-kDa neutrophilmembrane component as well as to peptide 2 was specific.Thirty-microgram samples of solubilized neutrophil mem-branes were photolabeled with '25I-PAL in the presence ofincreasing concentrations of peptide 2 and subjected to SDS-PAGE and autoradiography. Results after laser scanning den-sitometer analysis (Fig. IA) showed that the inhibition ofligand binding to the 68-kDa protein by peptide 2 responded ina concentration-dependent fashion (complete inhibition ofbinding by -10 mM peptide 2). In addition, the direct bindingof '25I-PAL to peptide 2 was examined by photolabeling 10-,ugsamples of peptide 2 to '25I-PAL in the presence of increasingconcentrations of nonradiolabeled ligand (PAL). Densitomet-ric analysis demonstrated that '25I-PAL binding to peptide 2was inhibited almost 90% by - 12 ,uM unlabeled ligand (Fig.1B). Results for both of the interactions indicated that theprocess was specific and saturable. The high affinity of thechemotactic peptides for their receptor has been documentedbefore, especially with photoactivatable reagents as cross-linkers (1).
Inhibition of ligand binding to neutrophil membrane com-ponents. Previously, we have shown strong incorporation of
VOL. 62, 1994
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 24
Dec
embe
r 20
21 b
y 18
3.91
.203
.37.
1728 RADEL ET AL.
c0
C0
01A.1
A
97 kDa-
68 kDa _66 kDa-
45 kDa-
c 80-
._
- 60-C
c 40-20
20
4 6 8 10 12 14 16Unlabeled Ligand (gM)
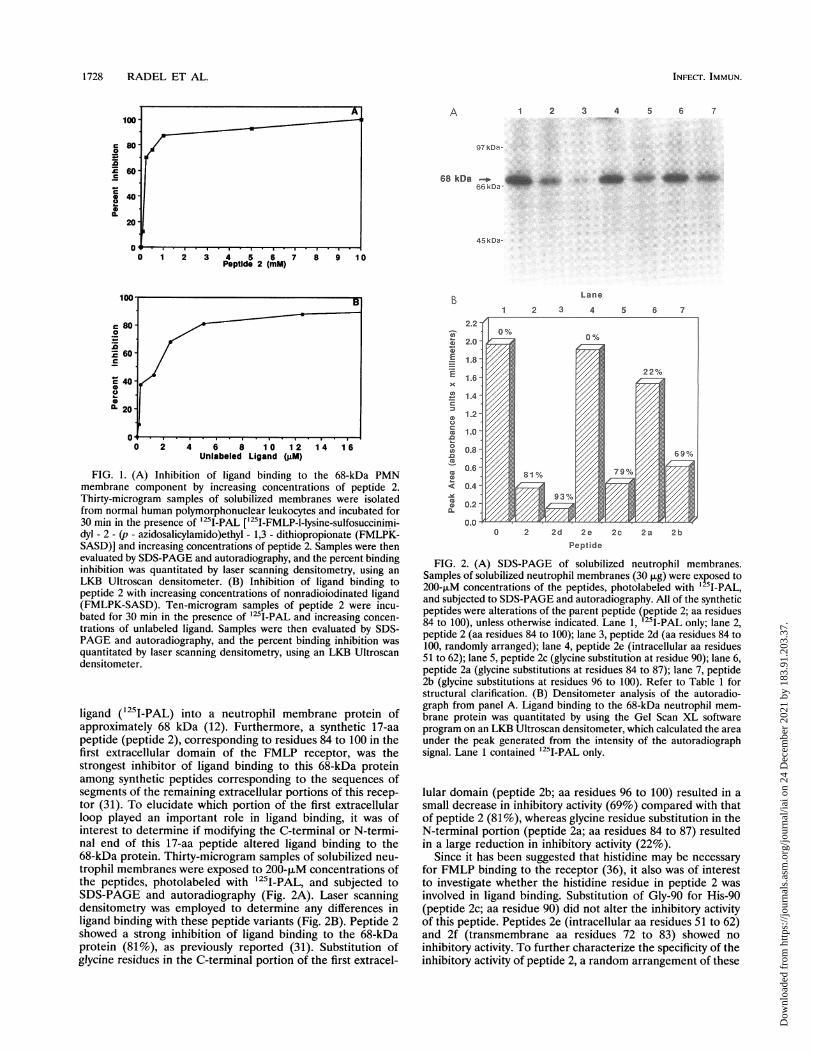
FIG. 1. (A) Inhibition of ligand binding to the 68-kDa PMNmembrane component by increasing concentrations of peptide 2.Thirty-microgram samples of solubilized membranes were isolatedfrom normal human polymorphonuclear leukocytes and incubated for30 min in the presence of '25I-PAL [125I-FMLP-l-lysine-sulfosuccinimi-dyl - 2 - (p - azidosalicylamido)ethyl - 1,3 - dithiopropionate (FMLPK-SASD)] and increasing concentrations of peptide 2. Samples were thenevaluated by SDS-PAGE and autoradiography, and the percent bindinginhibition was quantitated by laser scanning densitometry, using anLKB Ultroscan densitometer. (B) Inhibition of ligand binding topeptide 2 with increasing concentrations of nonradioiodinated ligand(FMLPK-SASD). Ten-microgram samples of peptide 2 were incu-bated for 30 min in the presence of 125I-PAL and increasing concen-trations of unlabeled ligand. Samples were then evaluated by SDS-PAGE and autoradiography, and the percent binding inhibition wasquantitated by laser scanning densitometry, using an LKB Ultroscandensitometer.
ligand (125I-PAL) into a neutrophil membrane protein ofapproximately 68 kDa (12). Furthermore, a synthetic 17-aapeptide (peptide 2), corresponding to residues 84 to 100 in thefirst extracellular domain of the FMLP receptor, was thestrongest inhibitor of ligand binding to this 68-kDa proteinamong synthetic peptides corresponding to the sequences ofsegments of the remaining extracellular portions of this recep-tor (31). To elucidate which portion of the first extracellularloop played an important role in ligand binding, it was ofinterest to determine if modifying the C-terminal or N-termi-nal end of this 17-aa peptide altered ligand binding to the68-kDa protein. Thirty-microgram samples of solubilized neu-trophil membranes were exposed to 200-piM concentrations ofthe peptides, photolabeled with 125I-PAL, and subjected toSDS-PAGE and autoradiography (Fig. 2A). Laser scanningdensitometry was employed to determine any differences inligand binding with these peptide variants (Fig. 2B). Peptide 2showed a strong inhibition of ligand binding to the 68-kDaprotein (81%), as previously reported (31). Substitution ofglycine residues in the C-terminal portion of the first extracel-
B Lane1 2 3 4 5 6 7
2.2-0% 0
0~~~~~01.8
01.6d 2e 2c 2a2%
* 1.4C
201.20
0.
0.6
0 2 2d 2e 2c 2a 2bPeptide
FIG. 2. (A) SDS-PAGE of solubilized neutrophil membranes.Samples of solubilized neutrophil membranes (30 V.g) were exposed to200-,uM concentrations of the peptides, photolabeled with '25I-PAL,and subjected to SDS-PAGE and autoradiography. All of the syntheticpeptides were alterations of the parent peptide (peptide 2; aa residues84 to 100), unless otherwise indicated. Lane 1, 125I-PAL only; lane 2,peptide 2 (aa residues 84 to 100); lane 3, peptide 2d (aa residues 84 to100, randomly arranged); lane 4, peptide 2e (intracellular aa residues51 to 62); lane 5, peptide 2c (glycine substitution at residue 90); lane 6,peptide 2a (glycine substitutions at residues 84 to 87); lane 7, peptide2b (glycine substitutions at residues 96 to 100). Refer to Table 1 forstructural clarification. (B) Densitometer analysis of the autoradio-graph from panel A. Ligand binding to the 68-kDa neutrophil mem-brane protein was quantitated by using the Gel Scan XL softwareprogram on an LKB Ultroscan densitometer, which calculated the areaunder the peak generated from the intensity of the autoradiographsignal. Lane 1 contained '25I-PAL only.
lular domain (peptide 2b; aa residues 96 to 100) resulted in asmall decrease in inhibitory activity (69%) compared with thatof peptide 2 (81%), whereas glycine residue substitution in theN-terminal portion (peptide 2a; aa residues 84 to 87) resultedin a large reduction in inhibitory activity (22%).
Since it has been suggested that histidine may be necessaryfor FMLP binding to the receptor (36), it also was of interestto investigate whether the histidine residue in peptide 2 wasinvolved in ligand binding. Substitution of Gly-90 for His-90(peptide 2c; aa residue 90) did not alter the inhibitory activityof this peptide. Peptides 2e (intracellular aa residues 51 to 62)and 2f (transmembrane aa residues 72 to 83) showed noinhibitory activity. To further characterize the specificity of theinhibitory activity of peptide 2, a random arrangement of these
INFECT. IMMUN.
I ---
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 24
Dec
embe
r 20
21 b
y 18
3.91
.203
.37.

NEUTROPHIL FMLP RECEPTOR LIGAND-BINDING REGION
amino acid residues (peptide 2d) was tested as a negativecontrol. Surprisingly, peptide 2d demonstrated an inhibitoryactivity greater than that shown by peptide 2 (93 and 81%,respectively).
Studies have suggested that FMLP binds to a predominantlyhydrophobic pocket by interacting at separate points withpositively charged amino acid residues (4, 16). Our studiesshowed that the strongest ligand-binding activity was associ-ated with the N-terminal portion of the first extracellular loop,of which the first two residues were charged moieties (Arg-84and Lys-85). Furthermore, a random arrangement of peptide 2residues (peptide 2d) unexpectedly showed a greater inhibitoryactivity than the parent peptide. Therefore, it was of interest toexamine the contribution of charged residues to ligand bind-ing, not only with the parent peptide sequence but also with therandom peptide. Thirty-microgram samples of solubilized neu-trophil membranes were again exposed to the peptides at200-pLM concentrations, photolabeled with 125I-PAL, and sub-jected to SDS-PAGE and autoradiography (Fig. 3A). Densi-tometric analysis (Fig. 3B) showed that substitution of Gly-84for Arg-84 (peptide 2g; aa residue 84) resulted in a smalldecrease in inhibitory activity (60%) compared with the de-crease when parent peptide 2 was used (78%). Glycine substi-tutions for both Arg-84 and Lys-85 (peptide 2h; aa residues 84to 85) resulted in a large decrease in inhibitory activity (18%).A synthetic peptide that contained substitutions for all of thecharged residues (Arg-84, Lys-85, and Lys-100) resulted intotal abrogation of inhibitory activity of ligand binding to theFMLP receptor (results not shown). Furthermore, no effect onligand binding was observed with polyamino acid controls(poly-L-arginine, poly-L-lysine, and polyglycine) at concentra-tions as high as 1 mM. Densitometric analysis examiningglycine substitutions for the charged residues in the randompeptide (Fig. 4) showed that a single substitution of a glycinefor the arginine residue (peptide 2i) resulted in a decrease ininhibitory activity (83%) compared with that in peptide 2d(94%). Inhibitory activity was completely lost when glycineswere substituted for the arginine and both lysines (peptide 2j).The same effects seen with the random peptide were foundwhen peptide 2 residues were arbitrarily arranged in a second"random" peptide (results not shown).
Direct ligand binding to the synthetic peptides. Variationsin peptide amino acid composition of the first extracellularloop of the FMLP receptor were also examined in order todetermine which changes had the greatest effect on directligand binding. Ten-microgram samples of the peptide variantswere incubated with '25I-PAL and subjected to SDS-PAGEand autoradiography. Densitometric analysis results of themembrane studies with the synthetic peptide variants of pep-tide 2 correlated with the direct peptide-ligand binding studies:i.e., the greater the binding inhibition of ligand to the 68-kDaneutrophil membrane component, the stronger the peptide-ligand binding (results not shown).
DISCUSSION
In this study, we wanted to determine the portions of theproposed first extracellular loop of the N-terminal end of theFMLP receptor which were important in ligand-binding func-tion. Previously, we had shown that this 17-aa segment, resi-dues 84 to 100 referred to as peptide 2, bound ligand thestrongest compared with binding by the other two proposedextracellular loops of this receptor. Peptide 2 also demon-strated the strongest ligand-binding inhibition to the 68-kDaFMLP receptor component. A recent study with chimericmolecules of the FMLP and CSa receptors also found that the
A
97kDa-
68 kDa.6k
45kDa-
B
1 2
Lane
U)a0E
'Ea)
0W11
0
.0
a.
0 2 2g 2hPeptide
FIG. 3. (A) SDS-PAGE of solubilized neutrophil membranes.Samples of solubilized neutrophil membranes (30 ,Ug) were exposed to200-,uM concentrations of the peptides, photolabeled with 25I-PAL,and subjected to SDS-PAGE and autoradiography. All of the syntheticpeptides were alterations of the parent peptide (peptide 2; aa residues84 to 100), unless otherwise indicated. Lane 1, '25I-PAL only; lane 2,peptide 2 (aa residues 84 to 100); lane 3, peptide 2g (glycine substitu-tion at residue 84); lane 4. peptide 2h (glycine substitutions at residues84 to 85). Refer to Table I for structural clarification. (B) Densitom-eter analysis of the autoradiograph from panel A. Ligand binding tothe 68-kDa neutrophil membrane protein was quantitated by using theGel Scan XL software program on an LKB Ultroscan densitometer,which calculated the area under the peak generated from the intensityof the autoradiograph signal. Lane 1 contained '25I-PAL only.
first extracellular loop of the FMLP receptor played an impor-tant role in ligand binding (29). Since this receptor domainappeared to play a critical role in ligand binding, it was ofinterest to determine if changes in the structural compositionof the C-terminal or N-terminal end of this peptide alteredligand binding to the 68-kDa protein. By SDS-PAGE, autora-diography, and laser densitometric analysis, each of the pep-tides was tested for the ability to inhibit binding of radiolabeledphotoaffinity-conjugated FMLP (125I-PAL) to solubilized neu-trophil membranes (major protein species of 68 kDa). Aspreviously reported, peptide 2 exhibited the strongest inhibi-tion (81%) of ligand binding to the 68-kDa protein (31).Substitution of glycine residues in the C-terminal portion ofpeptide 2 (peptide 2b) resulted in a 69% inhibition of ligandbinding to the 68-kDa protein, whereas glycine residue substi-tutions in the N-terminal portion (peptide 2a) showed a largereduction in inhibitory activity (22%). Peptides 2e (intracellu-
VC)L. 62, 1994 1 729
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 24
Dec
embe
r 20
21 b
y 18
3.91
.203
.37.

1730 RADEL ET AL.
Lane2 3 5
-
0E
a
0-
e
eD
D
0
0 2 2d 21 2jPeptide
FIG. 4. Densitometer analysis of the autoradiograph from an SDS-10% PAGE gel which examined ligand binding to the 68-kDa neutro-phil membrane protein in the presence of synthetic peptides. Samplesof solubilized neutrophil membranes (30 jxg) were exposed to 200-,uMconcentrations of the peptides, photolabeled with 1251I-PAL, subjectedto SDS-PAGE and autoradiography, and then analyzed on an LKBUltroscan densitometer. Ligand binding was quantitated by using theGel Scan XL software program, which calculated the area under thepeak generated from the intensity of the autoradiograph signal. Lane1, '25I-PAL only; lane 2, peptide 2 (aa residues 84 to 100); lane 3,peptide 2d (aa residues 84 to 100, randomly arranged); lane 4, peptide2i (peptide 2d amino acid sequence with a glycine substitution for thearginine residue); lane 5, peptide 2j (peptide 2d amino acid sequencewith glycine substitutions for the arginine and both lysine residues).Refer to Table 1 for structural clarification.
lar aa residues 51 to 62) and 2f (transmembrane aa residues 72to 83) showed no measurable ability to inhibit ligand binding tothe 68-kDa protein. These results suggested that the N-terminal end of the first extracellular loop is a major contrib-utor to ligand binding.
After localizing ligand-binding activity to the N-terminalregion of the first extracellular loop of the FMLP receptor, itwas of interest to further characterize the specificity of theactivity of peptide 2 (the original 17-aa residues making up thefirst extracellular domain of the FMLP receptor). To accom-plish this, we synthesized a peptide (peptide 2d) that consistedof a random rearrangement of peptide 2 residues which shouldact as a negative control. However, this randomly arrangedpeptide showed inhibition of ligand binding to the 68-kDaprotein that was actually greater than that shown by peptide 2.This was surprising since it was hypothesized that a randompeptide would have no activity. It has been suggested thatFMLP binds to a pocket in the receptor which is predomi-nantly hydrophobic and that FMLP "contacts" certain aminoacid residues in the hydrophobic pocket (4, 16). These contactsinclude a positively charged region, presumably interactingwith a methionine residue, and a hydrophobic area likely to beinteracting with the phenylalanine aromatic ring. Furthermore,numerous studies have implicated charged residues, particu-larly arginine, as playing a critical role in receptor-ligandbinding (9, 10, 24). Since peptide 2 contained a number ofcharged residues, this led us to investigate whether histidine,arginine, and/or lysine played a role(s) in ligand binding to theFMLP receptor. This was accomplished by making glycinesubstitutions for the individual charged moieties in peptide 2,
as well as in the random peptide (peptide 2d). ExaminingHis-90 in the first extracellular loop was interesting for tworeasons. Not only is this charged residue the only histidine onthe extracellular portion of the receptor, but also it has beenreported that blocking these residues on neutrophil mem-branes with diethylpyrocarbonate could inhibit the binding ofFMLP to such membranes, thus suggesting that histidine maybe a necessary residue for FMLP binding to the receptor (36).To investigate this issue, we synthesized a peptide (peptide 2c)which was identical to peptide 2 except for the replacement ofHis-90 with Gly-90. This substitution had no effect on ligandbinding; i.e., the inhibitory activity of this peptide to the68-kDa neutrophil protein as well as its direct ligand-bindingcharacteristics remained unchanged regardless of whetherthere was a histidine or a glycine residue at position 90. Sincethis is the only histidine residue in the extracellular portion ofthe receptor molecule, it is possible that, although such aresidue may not play a direct role in ligand binding, agentswhich react with it may indirectly affect this extracellular loop,thus interfering with binding.
Substitution of Gly-84 for Arg-84 resulted in a decrease inthe inhibitory activity of peptide 2. When both Arg-84 andLys-85 were replaced by glycine residues, a large reduction inpeptide 2 inhibitory activity occurred. A similar result wasfound for both of the random peptides; i.e., substituting for thecharged arginine and lysine residues in the randomly arrangedpeptide resulted in a total abrogation of inhibitory activity. Thefact that inhibitory activity was lost even when the arginine andlysine residues in the random peptide were replaced suggeststhat the charged moieties on the first extracellular loop of theFMLP receptor may function as important contacts in recep-tor-ligand interactions, regardless of their orientation. Thiseffect cannot be attributed solely to the effects of polycationicsubstances since poly-L-arginine and poly-L-lysine had no effecton ligand binding to the FMLP receptor component.The seven-transmembrane-domain structure proposed for
the FMLP receptor is similar to the structure of the ,-adren-ergic, G-protein-coupled receptor (5, 27). It has been reportedthat ligand binding to the P-adrenergic receptor occurs in theregion of the second and third transmembrane regions (14,37).Our previous studies have shown that the FMLP ligand bindsstrongest directly to peptide 2 (31), which is the extracellularloop (aa residues 84 to 100) between the second and thirdtransmembrane domains of the FMLP receptor (no bindingwas observed for synthetic peptides to transmembrane regionsin this area). Concomitantly, the strongest inhibition of ligandbinding (81%) to neutrophil membrane proteins occurs withthe same peptide. Our synthetic peptide studies which alteredthe C-terminal end of this loop, leaving the N-terminal endintact, showed a small decrease (69%) in inhibitory activity ofligand binding to the 68-kDa protein compared with thedecrease seen with peptide 2 (81%). Concomitantly, alter-ations in the N-terminal end of this loop, leaving the C-terminal end intact, demonstrated a large decrease (22%) ininhibitory activity. Furthermore, glycine substitutions for argi-nine and lysine residues on the original, intact, first extracel-lular loop peptide and the randomly arranged peptide of thisregion implicated these residues as playing critical roles inbinding. The first two residues (Arg-84 and Lys-85) of theN-terminal portion of the first extracellular loop are chargedresidues. This area not only has been implicated in ligandbinding as a result of our N-terminal substitution studies butalso contains the necessary features needed for ligand bindingas proposed by Freer et al. (16). Recently, Quehenberger et al.(30) reported a large decrease in ligand-binding affinity when achimeric molecule was made in which the first extracellular
INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 24
Dec
embe
r 20
21 b
y 18
3.91
.203
.37.

NEUTROPHIL FMLP RECEPTOR LIGAND-BINDING REGION 1731
loop of the FMLP receptor was replaced with the correspond-ing region from a 69% homologous granulocyte receptor(FPR2). The FPR2 receptor contains substitutions of nonpolaramino acids for the charged residues Arg-84 and Lys-85.Furthermore, a recombinant truncated FMLP receptor in anEscherichia coli expression system which lacked the first 86residues of the full-length molecule was constructed. Thismutant construct, which did not contain Arg-84 and Lys-85,failed to demonstrate specific ligand binding, whereas thefull-length receptor protein did (21). Our study indicates thatthe charged moieties on the N-terminal end of the firstextracellular loop of the FMLP receptor protein, specifically,Arg-84 and Lys-85, contains or contributes significantly toligand-receptor interaction. These charged residues could actas important contacts in binding ligand; i.e., these receptormoieties on the first extracellular loop will bind ligand regard-less of their position.
By definition, the FMLP receptor has been classically iden-tified as binding a tripeptide (FMLP). Most receptors havebeen characterized as binding a single ligand. For any receptor,the possibility exists of two (or more) structurally distinctclasses of receptors which bind the same ligand but possessseparate affinities (15). However, a single FMLP receptor hasbeen reported to bind numerous oligopeptide chemotacticfactors with varying degrees of neutrophil activation depen-dent on the structure of these oligopeptide FMLP derivatives(13, 34). It seems odd that a receptor would have such a varietyof distinct ligands even though it has been assumed that theFMLP receptor in nature is there primarily to bind FMLP. Itis interesting to note that the random peptide inhibited ligandbinding in our studies to a greater degree than the peptide asit exists in nature (the peptide corresponding to the firstextracellular loop receptor domain). These results suggestedthat the charged residues in the first extracellular domain wereresponsible for ligand interaction. This evidence can explainwhy the FMLP receptor can bind such a wide assortment ofligands and suggests that FMLP may not be the best ligandmodel for this receptor. Rather, the binding of FMLP may bean indirect and nonspecific phenomenon functioning by inter-acting with the charged moieties in the first extracellular loopof the FMLP receptor regardless of the orientation of theresidues. We are currently investigating the contribution ofindividual charged residues of this first extracellular loop of theFMLP receptor through site-directed mutagenesis studies, aswell as examining the peptide variants to determine if the levelof their ligand-binding inhibitory activity actually correlates toa functionally reduced state of neutrophil activation.
ACKNOWLEDGMENTS
We thank James Bergey and Gary Haraszthy for expert technicalassistance and informative discussions.
This work was supported in part by Public Health Service grantsDE-07926 and DE-04898 from the National Institute of DentalResearch.
REFERENCESI. Allen, R. A., A. J. Jesaitis, and C. G. Cochrane. 1987. Photoaffinity
labelling of N-formyl peptide receptors. Pharmacol. Ther. 33:333-348.
2. Allen, R. A., J. 0. Tolley, and A. J. Jesaitis. 1986. Preparation andproperties of an improved photoaffinity ligand of the N-formylpeptide receptor. Biochim. Biophys. Acta 882:271-280.
3. Aswanikumar, S., B. A. Corcoran, E. Schiffmann, A. R. Day, R. J.Freer, H. J. Showell, E. L. Becker, and C. B. Pert. 1977. Demon-stration of a receptor on rabbit neutrophils for chemotacticpeptides. Biochem. Biophys. Res. Commun. 74:810-817.
4. Becker, E. L. 1987. The formylpeptide receptor of the neutrophil:
a search and conserve operation. Am. J. Pathol. 129:16-24.5. Benovic, J. L., J. Onorato, M. J. Lohse, H. G. Dohiman, C.
Staniszewski, M. G. Caron, and R. J. Lefkowitz. 1990. Syntheticpeptides of the hamster beta 2-adrenoceptor as substrates andinhibitors of the beta-adrenoceptor kinase. Br. J. Clin. Pharmacol.30:3S-12S.
6. Bokoch, G. M., and P. W. Reed. 1980. Stimulation of arachidonicacid metabolism in the polymorphonuclear leukocyte by an N-formylated peptide: comparison with ionophore A23187. J. Biol.Chem. 255:10223-10226.
7. Boulay, F., M. Tardif, L. Brouchon, and P. Vignais. 1990. Synthesisand use of a novel N-formyl peptide derivative to isolate a humanN-formyl peptide receptor cDNA. Biochem. Biophys. Res. Com-mun. 168:1103-1109.
8. Boulay, F., M. Tardif, L. Brouchon, and P. Vignais. 1990. Thehuman N-formylpeptide receptor. Characterization of two cDNAisolates and evidence for a new subfamily of G-protein-coupledreceptors. Biochemistry 29:11123-11133.
9. Chen, F., and D. Puet. 1991. Contributions of arginines-43 and -94of human choriogonadotropin beta to receptor binding and acti-vation as determined by oligonucleotide-based mutagenesis. Bio-chemistry 30:10171-10175.
10. Clements, J. M., L. J. Bawden, R. E. Bloxidge, G. Catlin, A. L.Cook, S. Craig, A. H. Drummond, R. M. Edwards, A. Fallon, D. R.Green, P. G. Hellewell, P. M. Kirwin, P. D. Nayee, S. J. Richard-son, D. Brown, S. B. Chahwala, M. Snarey, and D. Winslow. 1991.Two PDGF-, chain residues, arginine 27 and isoleucine 30,mediate receptor binding and activation. EMBO J. 10:4113-4120.
11. De Nardin, E., and R. J. Genco. 1991. Immunochemical charac-terization of the formyl peptide receptor moieties on humanneutrophils. Hybridoma 10:49-63.
12. De Nardin, E., S. J. Radel, and R. J. Genco. 1991. Isolation andpartial characterization of the formyl peptide receptor compo-nents on human neutrophils. Biochem. Biophys. Res. Commun.174:84-89.
13. Dentino, A. R., P. A. Raj, K. K. Bhandary, M. E. Wilson, and M. J.Levine. 1991. Role of peptide backbone conformation on biolog-ical activity of chemotactic peptides. J. Biol. Chem. 266:18460-18468.
14. Dohlman, H. G., M. G. Caron, C. D. Strader, N. Amlaiky, and R. J.Lefkowitz. 1988. Identification and sequence of a binding sitepeptide of the P.-adrenergic receptor. Biochemistry 27:1813-1817.
15. Feltner, D. E., and W. A. Marasco. 1989. Regulation of the formylpeptide receptor binding to rabbit neutrophil plasma membranes.Use of monovalent cations, guanine nucleotides, and bacterialtoxins to discriminate among different states of the receptor. J.Immunol. 142:3963-3970.
16. Freer, R. J., A. R. Day, N. Muthukumaraswamy, D. Pinon, A. Wu,H. J. Showell, and E. L. Becker. 1982. Formyl peptide chemoat-tractants: a model of the receptor on rabbit neutrophils. Biochem-istry 21:257-263.
17. Goetzl, E. J., D. W. Foster, and D. W. Goldman. 1981. Isolationand partial characterization of membrane protein constituents ofhuman neutrophil receptors for chemotactic formylmethionylpeptides. Biochemistry 20:5717-5722.
18. Koo, C., R. J. Lefkowitz, and R. Snyderman. 1983. Guaninenucleotides modulate the binding affinity of the oligopeptidechemoattractant receptor of human polymorphonuclear leuko-cytes. J. Clin. Invest. 72:748-753.
19. Lad, P. M., M. M. Glovsky, J. H. Richards, D. B. Learn, D. M.Reisinger, and P. A. Smiley. 1984. Identification of receptorregulatory proteins, membrane glycoproteins, and functional char-acteristics of adenylate cyclase in vesicles derived from the humanneutrophil. Mol. Immunol. 21:627-639.
20. Laemmli, U. K. 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature (London)227:680-685.
21. Lala, A., A. Sharma, H. Sojar, S. J. Radel, R. J. Genco, and E. DeNardin. 1993. Recombinant expression and partial characteriza-tion of the human formyl peptide receptor. Biochim. Biophys.Acta 1178:3t)2-306.
22. Malech, H. L., and J. I. Gallin. 1987. Current concepts: immunol-ogy. Neutrophils in human disease. N. Engl. J. Med. 317:687-694.
VC)L. 62, 1994
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 24
Dec
embe
r 20
21 b
y 18
3.91
.203
.37.

1732 RADEL ET AL.
23. Naccache, P. H., H. J. Showell, E. L. Becker, and R. I. Sha'afi.1977. Transport of sodium, potassium and calcium across rabbitpolymorphonuclear leukocyte membranes. J. Cell Biol. 73:428-444.
24. Nanduri, V. B., J. D. Hulmes, Y. C. Pan, P. L. Kilian, and A. S.Stern. 1991. The role of arginine residues in interleukin 1 receptorbinding. Biochim. Biophys. Acta 1133:160-166.
25. Niedel, J., J. Davis, and P. Cuatrecasas. 1980. Covalent affinitylabeling of the formyl peptide chemotactic receptor. J. Biol. Chem.255:7063-7066.
26. Niedel, J., S. Wilkinson, and P. Cuatrecasas. 1979. Receptor-mediated uptake and degradation of '251-chemotactic peptide byhuman neutrophils. J. Biol. Chem. 254:10700-10706.
27. O'Dowd, B. F., R. J. Lefkowitz, and M. G. Caron. 1989. Structureof the adrenergic and related receptors. Annu. Rev. Neurosci.12:67-83.
28. Painter, R. G., A. J. Jesaitis, and L. A. Sklar. 1984. Leukocytechemotaxis, p. 43-75. In E. Elson, E. W. Frazier, and L. Glaser(ed.), Cell membranes, vol. 2. Plenum Press, New York.
29. Perez, H. D., R. Holmes, L. R. Vilander, R. R. Adams, W.Manzana, D. Jolley, and W. H. Andrews. 1993. Formyl peptidereceptor chimeras define domains involved in ligand binding. J.Biol. Chem. 268:2292-2295.
30. Quehenberger, O., E. R. Prossnitz, S. L. Cavanagh, C. G. Coch-rane, and R. D. Ye. 1993. Multiple domains of the N-formylpeptide receptor are required for high-affinity ligand binding. J.Biol. Chem. 268:18167-18175.
31. Radel, S. J., R. J. Genco, and E. De Nardin. 1991. Localization ofligand binding regions of the human formyl peptide receptor.Biochem. Int. 25:745-753.
32. Rubin, R. P., L. E. Sink, and R. J. Freer. 1981. On the relationship
between formylmethionyl-leucyl-phenylalanine stimulation ofarachidonyl phosphatidylinositol turnover and lysosomal enzymesecretion by rabbit neutrophils. Mol. Pharmacol. 19:31-37.
33. Schiffmann, E., B. A. Corcoran, and S. M. Wahl. 1975. N-Formylmethionyl peptides as chemoattractants for leukocytes.Proc. Natl. Acad. Sci. USA 72:1059-1062.
34. Showell, H. J., R. J. Freer, S. H. Zigmond, E. Schiffmann, S.Aswanikumar, B. Corcoran, and E. L. Becker. 1976. The structure-activity relations of synthetic peptides as chemotactic factors andinducers of lysosomal enzyme secretion for neutrophils. J. Exp.Med. 143:1154-1169.
35. Signas, C., G. Raucci, K. Jonsson, P. Lindgren, G. M. Ananthara-maiah, M. Hook, and M. Lindberg. 1989. Nucleotide sequence ofthe gene for a fibronectin-binding protein from Staphylococcusalurelis: use of this peptide sequence in the synthesis of biologicallyactive peptides. Proc. Nat]. Acad. Sci. USA 86:699-703.
36. Spilberg, I., J. Mehta, and K. Hellenga. 1986. Chemical modifica-tion of human neutrophil membrane proteins: effect on fMet-Leu-Phe binding and function. J. Leukocyte Biol. 39:645-655.
37. Strader, C. D., I. S. Sigal, R. B. Register, M. R. Candelore, E.Rands, and R. A. F. Dixon. 1987. Identification of residuesrequired for ligand binding to the 3-adrenergic receptor. Proc.Natl. Acad. Sci. USA 84:4384-4388.
38. Swanson, S. J., and R. F. Mortensen. 1990. Binding and immuno-logical properties of a synthetic peptide corresponding to thephosphorylcholine-binding region of C-reactive protein. Mol. Im-munol. 27:679-687.
39. Wollenweber, H. W., and D. C. Morrison. 1985. Synthesis andbiochemical characterization of a photoactivatable, iodinatable,cleavable bacterial lipopolysaccharide derivative. J. Biol. Chem.260:15068-15074.
INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 24
Dec
embe
r 20
21 b
y 18
3.91
.203
.37.

















![SESSION 1 Identification of novel peptide ligand-receptor ... · Luis Cárdenas, Carmen Quinto, Ana Velarde-Buendia, Rosana Sanchez, Hen-Ming Wu, Alice Cheung [10’+5’] Presenter](https://img.pdfslide.us/doc/110x75/5f395132778e016080717ca0/session-1-identification-of-novel-peptide-ligand-receptor-luis-crdenas-carmen.jpg)



![SESSION 1 Identification of novel peptide ligand-receptor ... program.pdf · Identification of novel peptide ligand-receptor pairs in plants Yoshikatsu Matsubayashi [35’+10’]](https://img.pdfslide.us/doc/110x75/5c72dd7b09d3f2b92e8c5403/session-1-identification-of-novel-peptide-ligand-receptor-identification.jpg)







