Embed Size (px)
Citation preview

481
Biochimica et Biophysics Acta, 530 (1978) 487-502 @ Elsevier/North-Holland Biomedical Press
BBA 57236
FORMATION OF 3-~EXAPRENYL-4-HYDROXYBENZOATE BY MATRIX-FREE MITOCH~NDRIAL MEMBR~E-RICH PREPARATIONS OF YEAST
JOHN CASEY and DAVID R. THRELFALL
Department of Plant Biology, University of Hull, Hull, HUG 7RX (U.K.)
(Received February 6th, 1978)
Summary
It has been shown that a 10 000 X g matrix-free mitochondrial membrane- rich preparation from commercial bakers’ yeast is able to synthesize 3a&truns- hexaprenyl-4-hydroxybenzoate from 4-hydroxybenzoate and isopentenyl pyrophosphate. The synthesis is Mg2+ dependent and is stimulated markedly by the p$mer for polyprenylpyrophosphate synthesis, 3,3-dimethylallyl pyrophos- phate. At the optimal MgZf concentration for the synthesis of 3-hexaprenyl-4- hydroxybenzoate from 4-hydroxybenzoate, isopentenyl pyrophosphate and 3,3dimethylallyl pyrophosphate the priming function of 3,3-dimethylallyl pyrophosphate can be performed by either geranyl pyrophosphate (most effi- cient) or farnesyl pyrophosphate. At high Mg2+ concentrations, however, geranyl pyrophosphate and farnesyl pyrophosphate act mainly as sources of preformed side chains and Sdiprenyl- and 3-triprenyl-4-hydroxybenzoate, respectively, are produced. In the presence of a source of preformed polyprenyl pyrophosphates the membrane preparation catalysed the polyprenylation of methyl-4-hydrox- ybenzoate, 4_hydroxybenzaldehyde, 4-hydroxybenzylalcohol and 4-hydrox- ycinnamate. No evidence was obtained for the involvement of either 4-hydrox- ybenzoyl CoA or 4hydroxybenzoyl-S-protein in the formation of 3-poly- prenyl-4-hydroxybenzoates.
Introduction
It is well established that in many prokaryotic cells the first steps on the bio- synthetic pathway leading from 4-hydroxybenzoate to ubiquinone are as fol- lows: 4-hydroxybenzoate + 3-polyprenyl-4-hydroxybenzoate -+ Z-polyprenyl- phenol - -+ 6-methoxy-2-polyprenylphenol- - + ubiquinone [ 11. In eukaryotic cells, however, there is still speculation as to the nature of the first steps on the pathway leading from 4-hydroxybenzoate to ubiquinone (see below and Discussion).

488
Momose and Rudney [2], as part of an investigation into the nature of the biosynthetic pathway leading from 4-hydroxybenzoate to ubiquinone is mam- mals, demonstrated that the inner membranes of rat liver mitochondria are able to form 14C-labelled compounds with the thin layer chromatographic properties expected of 3-nona- and 3-decaprenyl-4-hydroxybenzoate from 4-hydroxy- [ i4C] benzoate and isopentenyl pyrophosphate. They also presented some evidence of a precursor-product relationship between 3-nonaprenyl-4-hydrox- ybenzoate and ubiquinone-9 in rat liver mince. In a similar series of studies, Thomas and Threlfall [3,4] were unable to demonstrate the synthesis of 3- polyprenyl-4-hydroxybenzoates from 4-hydroxybenzoate and isopentenyl pyrophosphate in mitochondrial membrane preparations from yeast, broad bean seeds and Euglena gracilis. More recently, however, we have found that intact yeast mitochondria are able to synthesize a i4C!-labelled compound with the thin layer chromatographic properties of 3-hexaprenyl-4-hydroxybenzoate from 4-hydroxy [ i4C] benzoate and isopentenyl pyrophosphate, which they then convert into [ 14C] ubiquinone-6 [ 51.
In the present paper we report on (a) the development of a set of incubation conditions under which it is possible to demonstrate the synthesis of a com- pound with the chromatographic properties expected of 3-hexaprenyl-4- hydroxybenzoate, from 4-hydroxybenzoate and isopentenyl pyrophosphate in matrix-free mitochondrial membrane-rich preparations of bakers’ yeast and, (b) the characterization of this compound as 3aZl-trans-hexaprenyl-4-hydrox- ybenzoate.
Materials and Methods
Radiochemicals 4-Hydroxy[ 7-14C] benzoic acid (55 Ci * mol-‘) was purchased from Schwarz-
Mann, Orangeburgh, New York (U.S.A.). L-[U-‘4C]Tyrosine hydrochloride (10 Ci * mol-‘), L-[2,3,5,6-3H4]tyrosine (78 Ci . mmoll’), L-[3,5-3Hz]tyrosine (53 Ci * mmol-‘), triammonium [ 1-14C]isopentenyl pyrophosphate (60 Ci * mol“), 3 RS-[ 2-3H]mevalonic acid lactone (82 Ci * mol-‘) and [3R, 4R-4-3H + 3S, 4S- 4-3H]mevalonic acid lactone (1.8 Ci * mmol-‘) were purchased from the Radio- chemical Centre, Amersham (U.K.).
(a). 14C- or 3H-labelled 4-hydroxy benzaldehydes. These were prepared by a procedure similar to that described by Parson and Rudney [6]. The principal modifications made to the procedure were that 14C- or 3H-labelled 4-hydroxy phenylpyruvate was prepared by incubating the appropriate i4C- or 3H-labelled tyrosine with L-amino acid oxidase [ 71; the 14C- or 3H-labelled 4-hydroxy benzaldehyde was purified by thin layer chromatography on either Silica gel G developed with diethyl ether/light petroleum/methanol (6 : 6 : 1, v/v) (RF = 0.47) or microgranular cellulose developed with aq. 2% (v/v) acetic acid (RF = 0.72). The yield of pure product was about 20%.
(b). 14C- or 3H-labelled 4-hydroxy benzoates. 5 /_Ei of 14C- or 3H-labelled 4- hydroxy-benzaldehyde was dissolved in 0.5 ml 0.05 M phosphate buffer (pH 7.0) and added to 2 ml of the same buffer containing 25 I.tmol MgClz and 10 mg yeast mitochondrial membrane. The mixture was incubated for 1 h at 30°C with agitation. The pH of the incubation mixture was then adjusted to 2 and

489
the 14C- or 3H-labelled 4-hydroxy-benzoate extracted with diethyl ether and purified by thin layer chromatography in the systems described under (a) (RF values 0.12 and 0.68, respectively). The yield of pure product was about 95%.
(c). Methyl-4-hydroxy[7-14C]benzoate. 4-Hydroxy[7-14C]benzoate (2 PCi; 7.6 Ci * mol-‘) was dissolved in 5 ml methylating reagent [ 91 and the mixture refluxed for 2 h. The methyl-4-hydroxy[7-14C]benzoate was then extracted with diethyl ether and purified by thin-layer chromatography in the first system described under (a) (RF = 0.54). The yield of radio-gas-liquid chro- matography pure product was about 80%.
cd). 4-Hydroxy-[ 7-14C] benzylalcohol. Methyl-4-hydroxy [ 7-14C] benzoate (0.5 /-AX; 7.6 Ci . mol-‘) was dissolved in dry peroxide-free diethyl ether and treated with an excess amount of sodium dihydro-bis(2-methoxyethoxy) aluminate. The reaction mixture was allowed to stand for 15 min, treated with 10% HCl and washed with Hz0 until free of acid. The alcohol was then separated from any unreacted ester by thin-layer chromatography in the first system described under (a) (RF 0.25). The yield of pure product was about 15%.
(e). 4-Hydroxy[U-14C!]cinnamate. L [U-14C]Tyrosine hydrochloride (3 PCi, 10 Ci * mol-‘) was incubated with 3 ml 0.1 M borate buffer (pH 8.8) (contain- ing 0.2 g of an acetone powder of 7-day-old etiolated barley shoots) for 3 h at 40°C. The reaction mixture was then diluted with 7 ml HzO, filtered and the filtrate adjusted to pH 2. 4-Hydroxy[ U-14C]cinnamate was extracted from the acidified filtrate with diethyl ether and purified by thin-layer chromatography in the second system described under (a) (RF = 0.50). The yield of pure prod- uct was 10%.
(f). [4-3H]- and [2S-3H]labelled isopentenyl pyrophoiphate. 3RS-[2-3H]- mevalonic acid lactone (50 PCi; 82 Ci - mol-‘) or [3R,4R-4-3H + 3S, 4S-4-3H]- mevalonic acid (10 &i; 1.8 Ci . mmolF1) was converted to [3H]mevalonic acid by treatment with NaHCO, and then incubated with a suitably supplemented [9] undialysed 30 000 X g supernatant fraction from yeast cells for 10 min at 30°C. The reaction was terminated by boiling the mixture for 3 min. 5 ml ethanol was then added and the precipitated protein removed by centrifuga- tion. The ethanolic supernatant was taken to dryness in a rotary evaporator and the [ 3H] isopentenyl pyrophosphate and unreacted 3S- [ 3H] mevalonic acid taken up in a small volume on n-propanol/NH3/H,0 (6 : 3 : 1, v/v) and chro-
.matographed on thin layers of Silica gel H developed with the same solvent. The [3H]isopentenyl pyrophosphate (RF = 0.35) was eluted from the gel with developing solvent. The yield of triammonium [ ‘H] isopentenyl pyrophosphate was about 80% (after allowance for the biologically inactive isomer).
Chemicals. Trilithium isopentenyl pyrophosphate was synthesized from 3- methyl-3-buten-l-01, by the method of Donninger and Popjik [lo]. Tri- ammonium salts of 3,3-dimethylallyl pyrophosphate, geranyl pyrophosphate and farnesyl pyrophosphate were synthesized from 3-methyl-2-buten-l-01, trans- geraniol and farnesol (trans, trans, 38%; cis, trans, 49%; cis,cis, 12%) respectively, by the method of Cramer and Bohm [ll] as described by Cornforth and Popjik [ 121. The pyrophosphates were purified by repeated chromatography in the thin layer system used for the purification of [3H]isopentenyl pyrophos- phate (3,3dimethylallyl pyrophosphate, R F = 0.38; geranyl pyrophosphate, RF = 0.42; farnesyl pyrosphosphate, RF = 0.46).

490
The free acids of the Cs-pyrophosphates were prepared by passing solutions of the salts through small columns of Dowex 50 in the H’ form [l].
All other chemicals were of the best commercial grades. Biological material. Blocks of compressed bakers’ yeast (Saccharomyces cere-
visiae) (N.G. and SF., Fermentation Products, Dock Road, Felixstowe, U.K.) were purchased from a local distributor and stored at 3°C until required.
Preparation of yeast mitochondrial membrane-rich fractions. In the first series of experiments (Table I), one batch of 10 000 X g particulate fraction was prepared from cells of bakers’ yeast that had been broken by mechanical disruption in a Mickle High Speed Tissue Disintegrator [ 31.
The rest of the work was carried out with membranes from yeast cells that had been disrupted by one passage through a French Pressure Cell. 25-30 g yeast cells were suspended in 45 ml 0.3 M mannitol/0.05 M phosphate buffer (pH 7.5) and subjected to one passage through a pressure cell at 667 bar. The resultant homogenate was collected in 30 ml 0.3 M mannitol/0.05 M phosphate buffer (pH 7.5) and the mixture centrifuged at 1500 Xg for 10 mm. The 1500 X g supernatant fraction was then centrifuged at 10 000 X g for 15 min to sediment mitochondrial membranes. The membrane preparation was washed 3 times by resuspension in 30 ml mannitol/phosphate buffer followed by resedimentation at 10 000 X g for 15 min, and finally resuspended in 2 ml 0.05 M phosphate buffer (pH 7.5). This preparation (15-20 mg protein * ml-‘) was rich in mitochondrial membranes (50-60% of total succinate dehydrogenase activity) but was deficient in matrix enzymes (O-5% of total fumarase activ- ity).
In the intracellular distribution study (Fig. 1) the cells were ruptured at a pressure of 200 bar and then fractionated by centrifugation at 1000 X g for 10 min, 5000 X g for 15 min, 30 000 X g for 20 min and 100 000 X g for 45 min. The sedimented membranes were washed twice by resuspension in mannitol/ phosphate buffer followed by resedimentation at the appropriate g values.
Assay of marker enzyme activities. Succinate dehydrogenase (EC 1.3.99.1), fumarase (EC 4.2.1.2) and NADPH-cytochrome c reductase (EC 1.6.2.4) activi- ties were measured spectroscopically by the methods described by Lord and Merrett [ 131, Collins and Merrett [ 141 and Haas [ 151, respectively.
Assay of prenyltransferase activity. This assay was based on the diprenyl transferase assay described by Popjak [16]. 1.35 ml 0.05 M phosphate buffer (pH 7.5)/5 pmol MgClJ0.05 pmol triammonium [1-14C]isopentenyl pyrophos- phate (10 Ci . mol-‘)/O.l pmol triammonium 3,3-dimethylallyl pyrophosphate/ 0.1-0.5 mg protein was incubated for 30 min at 30°C. The reaction was stopped by the rapid addition of 1 ml aq. 80% (v/v) ethanol/2 M HCl and incubated for 30 min at 37°C. The mixture was then adjusted to pH 10 with 10 M NaOH and extracted with light petroleum (b.p. 40-60°C). The prenyltrans- ferase activity is expressed as the amount of radioactivity that was extracted into light petroleum.
The nature of the i4C-labelled compounds extracted by light petroleum was investigated by thin-layer chromatography (a) on either Rhodamine 6G- impregnated Silica gel G or Silica gel G developed with ethyl acetate-benzene (3 : 22, v/v) (squalene, RF = 0.75; nerolidol, RF = 0.55; hexaprenol, RF = 0.48; farnesol, RF = 0.39) and (b) on paraffin-impregnated silica gel G developed

491
with aq. 75% (v/v) acetone (squalene, R F = 0, nerolidol, RF = 0.55; hexaprenol, RF = 0.20; farnesol, RF = 0.65). The marker compounds were located either by ex~ination of the plate under ultraviolet light or by staining with I2 vapour.
Assay of 3-polyprenyl-4-hydroxybenzoate formation. This was assayed by measuring the incorporation of radioactivity from 14C- or 14C!, 3H-labelled 4- hydroxy benzoate into the CHCl,-soluble extracts of the incubation mixtures. In those incubations supplemented with Micrococcus Zuteus extracts that had been preincubated with isopentenyl pyrophosphate, the M. Euteus extract was prepared and preincubated as described by Raman et al. 1171. The mixtures were incubated in 25-ml conical flasks in air for the appropriate period of time at 30°C with gentle agitation. The reaction was stopped by the rapid addition of 5 ~01s. CHC13/CH30H (1 : 2, v/v) and, after allowing the mixture to stand for 2 h, the CHCIJ-soluble lipids were extracted and assayed for 14C activity [ 3]. In all of the experiemts reported in this paper, apart from those involving [ 14C] - and [ 3H] isopentenyl pyrophosphates and [ 14C] -1abelled $-hydroxy- benzoate analogues, the 14C activity was associated entirely with 3-polyprenyl- 4-hydroxybenzoates.
The purification and determination of the chain lengths of the 3-polyprenyl- 4-hydroxybenzoates was carried out by adsorptive thin-layer chromatography on silica gel H developed with acetone/light petroleum (3 : 7, v/v) followed by reversed-phase thin-layer chromatography on paraffin-imprecated Silica gel G developed with aq. 95% (v/v) acetone. The RF values of the di, tri and hexa through to deca homologues in the two systems are 0.16, 0.21, 0.33, 0.36, 0.42, 0.46 and 0.50, and 0.81, 0.73, 0.52, 0.46, 0.39, 0.33 and 0.27, respec- tively. The radioactive compounds were located by means of either a Panax TL Chromato~am Scanner or autoradio~aphy.
Radioassay. Samples were assayed for radioactivity in a Beckman Liquid Scintillation Spectrometer [ 31. 14C : 3H ratios were determined by the discrim- inator-ratio method [17]. Thin-layer chromatograms were scanned for radio- activity in a Panax TL Chromatogram Scanner. Detection of 14C-labelled com- pounds on chromato~ams was also carried out by autoradiography using Induxtrex C X-ray film (Kodak Ltd.) 133.
Radio-gas-liquid chromatography. This was carried out in a Pye Unicam Series 104 GC equipped with a flame ionization detector and linked to a Panax Radiogas Detector System. The conditions were: 2.1 m X 2 mm (inner diam- eter) glass column packed with 5% OV-25 coated Gas-Chrom Q; column tem- perature, 135°C; argon flow rate 60 ml . min-‘; eluate split 99 : 1 in favour of passage through the Radiogas Detector System; response of radiogas detector, Rt + 0.5 min. The I4 C-labelled phenol was chromatographed as its trimethyl- silyl derivative (prepared by the variation of method 6 [19]. The R, values of the trimethylsilyl derivatives of 4-hydroxybenzylalcohol, methyl-4-hydrox- ybenzoate and 4-hydroxybenzoate were 7.0,10.25 and 14.4 min, respectively.
Protein estimation. Protein in the isolated cell fractions was precipitated with 10% trichloroacetic acid, washed free of lipid with aq. 95% (v/v) ethanol and estimated by the Biuret reaction [ZO] with bovine serum albumin fraction V as standard.

Results
3-Hexaprenyl-4-hydroxybenzoate formation by 10 000 X g mitochondrial membrane-rich fractions of yeast cells. A preliminary series of experiments established that small amounts of CHCl,-soluble 14C-labelled compounds with the thin-layer chromatographic properties expected of 2-hexaprenyl-4-hydrox- ybenzoate (major component, >90% of the mixture) and 2-heptaprenyl-4- hydroxybenzoate are synthesized from 4-hydroxybenzoate (55 Ci - mol-‘) and isopentenyl pyrophosphate in 10 000 X g mitochondrial membrane-rich frac- tions prepared from yeast cells that have been disrupted either by shaking with Ballotini beads in a Mickle Mill or by passage through a Pressure Cell (Table I). They also established that the rates of synthesis of these compounds are increased dramatically by the inclusion in the incubation mixtures of 3,3- dimethylallyl pyrophosphate (Table I), the primer for polyprenyl pyrophos- phate formation. Confirmation that the major component produced under these conditions is 3-hexaprenyl-4-hydroxybenzoate was provided by experi- ments to be described later in this paper.
As part of this series of experiments, it was demonstrated that a CHC13- soluble 14C-labelled compound with the thin-layer chromatographic properties of 3decaprenyl-4-hydroxybenzoate is synthesized from 4-hydroxy[ 7-14C]- benzoate and isopentenyl pyrophosphate in a 10 000 X g membrane prepara- tion from soaked bean seeds (Table I), and that the rates of synthesis of both
TABLE I
EFFECT OF 3.3-DIMETHYLALLYL PYROPHOSPHATE ON THE SYNTHESIS OF 3-POLYPRENYL-4-
HYDROXYBENZOATES FROM 4-HYDROXY[7-‘4CIBENZOATE AND ISOPENTENYL PYROPHOS- PHATE BY MITOCHONDRIAL PREPARATIONS
Yeast and broad-bean seed mitochondriai membrane preparations and rat liver mitochondria were prepared as described in Materials and Methods (Mickle Mill method), and refs. 3 and 2. respectively. Incubations l-4 consisted of 5 ml 0.05 M phosphate buffer (PH 7.2). 7.3 nmol 4-hydroxy[7-14CJbenzo-
ate (55 Ci . moT1). 7 pmol trilithium isopentenyl pyrophosphate. ? 3 ~~rnol triammonium 3.3-dimethyl- ally1 pyrophosphate, 100 wnol MgClZ and either 3 mg yeast mitochondrial protein or 12 mg broad-bean mitochondrial protein. The mixtures were incubated for 2 h at 30°C. Incubations 5 and 6 consisted of 1
ml 0.05 M phosphate buffer (pH 7.2). 2.1 nmol 4-hydroxy[7-‘4 Clbenzoate (55 Ci . mcC1). 1 pm01 tri- lithium isopentenyl pyrophosphate, tl pm01 triammonium 3,3-dimethyl~dlyl pyrophosphate. 10 ~mol
MgClZ, 10 ~101 KF, 200 fig bovine serum albumin and 5.2 mg mitochondrial protein. The mixtures were incubated for 1 h at 37%.
Source of
preparation Presence or
absence of 3,3-dimethylallyl pyrophosphate
Yeast ** yeast Broad bean
Broad bean Rat Rat
- +
+ - +
3-Polyprenyl-4-hydroxybenzoates formed
(pm01 h-l . mgl protein) *
hexa
1.5 48.5
hepta
n.e. n.e.
-
- 5.8
-
nona
-
-
n.e. 12.3
deca
1.3
3.8
n.e.
* Not formed in the appropriate control incubations. ** Similar results were obtained for preparations from yeast cells that had been broken with a pressure
cell. n.e., present (<lo% of major homologue). but not estimated.

493
Fig. 1. Intracellular distribution of fdi)prenyltransferase activity. A 1000 Xg homogenate from yeast cells that had been broken by one passage through a pressure cell at 200 bar was subjected to a scheme of dif- ferential centrifugation and the resultant 5000 Xg (3 mg protein). 30 000 Xg (1.9 mg protein) and 100 000 Xg (0.72 mg protein) membrane preparations and 100 000 Xg supernatant fraction (34.1 rng’ protein) assayed for their succinate dehydrogenase (o----- 0: 100% = 0.576 pro01 substrate consumed n~in-~), fumarase f v-0; 100% = 3.6 pm01 substrate consumed . min-I), NADFH-cytochrom~ i reductase (~1 -------A; 100% = 8.2 ~.trnol substrate consumed . min-*) and fdi)prenyltransferase (*-0; 100% = 2.4 firno [l-14Clisopentenyl pyrophosphate incorporated . 0.5 h-l) activities.
this compound and of the 14C-labelled compound with the chromatographic properties expected of 3-nonaprenyl-4-hydroxybenzoate formed in rat liver mitochondria [Z] are stimulated by the inclusion of 3,3-dimethyl~lyl pyro- phosphate in the incubation mixtures (Table I).
Subcellular distribution of prenyl transferase activity. The experiments just described provided evidence that yeast mitochondrial membranes are capable of the synthesis of the hexaprenyl pyrophosphate required for the formation of 3-hexaprenyl-4-hydroxybenzoate. To investigate this possib~ity further, a homogenate prepared by passage of yeast cells through the pressure cell was subjected to a scheme of differential centrifugation, and the various fractions assayed for protein content, and succinate dehydrogenase, NADPH-cytochrome c reductase, fumarase and prenyl transferase activities (Fig. 1).
It was found, as expected in view of the high rate of sterol synthesis which is observed in aerobic yeast cells, that most of the prenyl transferase activity was present in the supernatant fraction (Fig. 1). Examination of the light petro- leum-soluble products from the acid-hydrolized supernatant assay mixtures by thin-layer chromatography showed that most of the 14C activity was present in farnesol and nerolidol in the proportions 1 : 4. Thus demonstrating that farnesyl pyrophosphate is the principal product of the supernatant prenyl transferase activity [ 16 1.
The prenyl transferase activity associated with the other fractions was dis- tributed between them in much the same way as the mitochondrial membrane marker enzyme (succinate dehydrogenase) and did not follow the distribution of either the mitochondrial matrix marker enzyme (fumarase) or the micro- somal marker enzyme (NADPH-~yto~hrome c reductase) [21] (Fig. 1). Analy-

494
sis of the light petroleum-soluble products from the acid-hydrolized membrane assay mixtures by thin-layer chromatography showed that in each case the 14C activity was associated with farnesol and merolidol (1 : 4) and two ‘high RF’ compounds, the highest of which migrated with the same RF as squalene and the major acid-hydrolysis product of presqualene pyrophosphate [22]. No radioactivity migrated with the RF value expected of either hexaprenol or farnesylnerolidol.
In the experiments reported in the rest of the paper, the 10 000 X g mito- chondrial membrane-rich fractions were obtained from homogenate of cells that had ruptured by one passage at 667 bar through the pressure cell. The distribution of the marker enzymes for such a preparation are given in Materials and Methods.
Conditions for optimal rate of synthesis of 3-hexaprenyl-4-hydroxybenzoate. It was established that the optimum conditions for the synthesis of 3-hexa- prenyl-4-hydroxybenzoate from 1 nmol 4-hydroxy [ 7-14C] benzoate (55 Ci * mol-‘) by a 10 000 X g matrix-free mitochondrial membrane-rich fraction con- taining 2-10 mg protein are: 675 pmol phosphate buffer (pH 7.5), 1 nmol 4- hydroxy[7-14C]benzoate (55 /.Xi . mol-‘), 0.5 pmol trilithium isopentenyl pyrophosphate, 1 nmol triammonium 3,3-dimethylallyl pyrophosphate, 5 E.tmol MgClz and 2-10 mg protein in a total volume of 1.35 ml. The rate of synthesis under these conditions was 5060 pmol 3-hexaprenyl-4-hydroxybenzoate * h-’ - mg-’ protein and this rate was maintained for periods of up to 120 min. The rate was extremely pH dependent and reduced markedly if the Mg” (there is an absolute requirement for this metal ion), trilithium isopentenyl pyrophos- phate or triammonium 3,3dimethylallyl pyrophosphate concentrations were altered. It was, however, unaffected by (a) replacement of the salts of C5-pyro- phosphates by their acid forms, (b) addition of phosphatase inhibitors (KF and bovine serum albumin), (c) replacement of the phosphate buffer, which has been reported to be an inhibitor of prenylpyrophosphate synthesis [ 161, by Tris - HCl buffer, and (d) washing of the preparation. Furthermore, the addi- tion of Mn*+, the metal ion required for maximal isopentenyl pyrophosphate- isomerase activity [ 161, did not dispense with the requirement for 3,3-dimeth- ylallyl pyrophosphate.
On replacement of the 4-hydroxy [ 7-14C] benzoate with 4-hydroxy [ U-i4C] - benzoate (55 Ci * mol-’ ), 3-hexaprenyl-4-hydroxy [ 14C] benzoate was still the only radioactive CHC13-soluble compound produced by the incubation mixture.
Utilization of geranyl pyrophosphate or farnesyl pyrophosphate in place of 3,3-dimethylallyl pyrophosphate. At the optimal Mg*+ concentration (3.7 mM) for the synthesis of 3-hexaprenyl-4-hydroxybenzoate from 4_hydroxybenzoate, isopentenyl pyrophosphate and 3,3-dimethylallyl pyrophosphate, geranyl pyro- phosphate and, to a lesser extent, farnesyl pyrophosphate can be used as pri- mers in place of 3,3dimethylallyl pyrophosphate (Table II). Geranyl pyrophos- phate, in addition to its use as a primer, is also used as a source of side chain for the synthesis of 3diprenyl4-hydroxybenzoate (Table II). At the optimal Mgz+ concentration (33.3 mM) for the synthesis of 3-triprenyl-4-hydroxybenzoate from 4-hydroxybenzoate and farnesyl pyrophosphate [ 31, farnesyl pyrophos- phate is used only as a source of side chain for the formation of 3-triprenyl-4- hydroxybenzoate, whereas geranyl pyrophosphate is still used both as a primer

495
TABLE II
EFFECT OF Mg*+ CONCENTRATION ON THE USE OF 3.3-DIMETHYLALLYL PYROPHOSPHATE. GERANYL PYROPHOSPHATE AND FARNESYL PYROPHOSPHATE EITHER AS PRIMERS FOR SIDE CHAIN SYNTHESIS OR AS NUCLEAR SUBSTITUENTS
Incubation mixtures consisted of 1.35 ml 0.05 M phosphate buffer (PH 7.5), 2.1 nmol 4-hydroxy-[7-
14Clbenzoate (55 Ci . mol-1). +l pmol trilithium isopentenyl pyrophosphate, 1 pmol triammonium primer, 3 mg yeast mitochondrial membrane protein and either 45 wmol MgCl2 or 5 pmol MgC12. The
mixtures were incubated for 1 h at 3O’C.
Side chain precursor(s) Mg*+
(mM)
3-Polyprenyl-4-hydroxy- benzoate synthesis
(% of incubation 1)
di tri hexa
3,3-Dimethylallyl pyrophosphate, isopentenyl pyrophosphate 3,3-Dimethylallyl pyrophosphate Isopentenyl pyrophosphate
3,3-Dimethylallyl pyrophosphate, isopentenyl pyrophosphate 3,3-Dimethylallyl pyrophosphate Isopentenyl pyrophosphate
Geranyl pyrophosphate Geranyl pyrophosphate, isopentenyl pyrophosphate
Geranyl pyrophosphate Geranyl pyrophosphate, isopentenyl pyrophosphate
Famesyl pyrophosphate Famesyl pyrophosphate, isopentenyl pyrophosphate
Famesyl pyrophosphate Famesyl pyrophospbate, isopentenyl pyrophosphate
3.7 3.7 3.7
33.3 33.3 33.3
3.7
3.7
33.3 33.3
3.7 3.7
33.3 33.3
- No products
-
No products -
45 -
29 -
86 87 -
4 0
37 - 45
100 *
13
30
2
0
115
0 37
0 32
0 0
* 100% = 127 pmol . h-l. mgl protein.
and as a source of side chain, although the relative importance of the two func- tions is now reversed (Table II). At both Mg*+ concentrations, 3,3_dimethylallyl pyrophosphate functions only as a primer.
Characterization of 3-hexaprenyl-4-hydroxybenzoate. The fact that only small amounts of 3-hexaprenyl-4-hydroxybenzoate were produced in the incubations meant that it could not be characterized by conventional chemical procedures.
Preliminary evidence that the CHCl,-soluble I4 C-labelled compound syn- thesized from 4-hydroxy [ 14C] benzoate, isopentenyl pyrophosphate and a primer is 3-hexaprenyl4-hydroxybenzoate was provided by the facts that it was chloroform soluble, that its thin-layer chromatographic properties (adsorp- tive and reversed phase) were those expected for 3-hexaprenyl-4-hydrox- ybenzoate, that it could be labelled with radioactivity from 4-hydroxy [ 7-14C] - benzoate, 4-hydroxy[U-14C]benzoate and [l-‘4C]isopentenyl pyrophosphate and that it was synthesized in a membrane preparation which contains ubi- quinone-6.
The unsaturated nature of the side chain and the nuclear substitution pattern were established by the following two experiments. In the first experiment, it was demonstrated that 3-hexaprenyl-4-hydroxy [ 7-14C] benzoate, as expected for a compound with an unsaturated side chain, an -OH group and a -COOH

group, can be sequentially hydrogenated using PtO*/H, (conditions as in ref. 23), methylated by refluxing with a methyl esterification reagent [8] and tri- methylsilylated (method 6, ref. 19) to form in quantitative yield progressively less polar “C-labelled compounds with the thin-layer chromato~aphic proper- ties expected of 3-perhydrohexaprenyl-4-hydroxybenzoate (Silica gel H/ acetone/light petroleum (3 : 7, v/v), RF = 0.65), 3_perhydroxyhexaprenyl- methyl-4-hydroxybenzoate (System just described, RF = 0.92; Silica gel G: benzene/light petroleum (2 : 3, v/v), RF = 0.28) and 3-perhydrohexaprenyl- methyl-4-0-trimethylsilylbenzoate (Silica gel G: benzen/light petroleum (2 : 3, v/v), RF = 0.42) (Fig. 2). In the second experiment, the substitution of the ring at position 3 was confirmed by the demonstration that 67% and 43% of the 3H is retained in the 3-hexaprenyl-4-hydroxy [ 14C, 3H] benzoate species biosyn- thesized from 4-hydroxy[LJ-14C, 2,3,5,6-3H4]benzoate and 4-hydroxy[7-14C, 3,5-3H2 Jbenzoate, respectively (Table III). In addition, it was shown that
TABLE III
STEREOCHEMISTRY OF THE SIDE CHAIN AND NUCLEAR SUBSTITUTION PATTERN OF 3-HEXAPRENYL-4-HYDROXYBENZOATE
Incubation 1 and 2 consisted of 1.35 ml 0.05 M phosphate buffer (PH 7.5). 1 wnol 4-hydroxybenzoate, 0.89 nmol triammonium Cl- I 4C, 2%3Hlisopentenyl pyrophosphate, (’ 4C, 60 Ci . mol-1: ) 4C : 3H ratio
1 : 3.77), f10 nmol triammonium 3,3_dimethylailyl pyrophosphate, 0.3 mg KF, 3 mg bovine serum albu- mm, 5 ~.lmol MgCl2 and 2 mg yeast mitochondrial membrane protein. Incubations 3-10 each consisted
of 1.35 mI phosphate buffer, (pH 7.5). 0.45 nmol 4-hydroxy17-14C,3,5-3H21benzoate f’ 4C. 5.5 Ci . mol-I ; 14C : 3H ratio, 1 : 7.13 or 1 : 6.90) or94 pmol 4-hydroxy[U-i4C,2,3.5,6-3H4]benzoate (14C, 400 Ci . mol-1; 14C : 3H ratio, 1 : 8.13). 5 nmol (incubations 3 and 6) or 45 nmol MgClz, 1 nmol amounts of the appropriate alcohol pyrophosphate(s) (see Table) or 0.25 ml M. luteus extract that had been preincu- bated with tritithium isopentenyl pyrophosphate (this preparation serves as a source of preformed POIY-
prenyl pyrophosphates 13,171) and 3 mg yeast mitoehonrial membrane protein. The mixtures were incu- bated for 1 h at 3O’C.
Radioactive substrate and source of side chain
Purified 3-polyprenyl-4-hydroxybenzoate(s) --
Homologue(s) 14C! activity Atomic r4C : 3H ratio
isolated tdpm) Corrected Expected *
fl-14C, 2.~-~HIIsopentenyt pyrophos- 1:l
phate 14C, 3H substrate hexa 130 1:l. 1:l ‘4C. 3H substrate, 3,3_dimethyl- hexa 450 1:l 1:l
ally1 pyrophosphate
4-Hydroxy[7-14C,3,5-3H2]benzoate - 1:2 Isopentenyl pyrophosphate, 3,3- hexa 1460 1 : 0.85 1:l
dimethylahyl pyrophosphate
Geranyl pyrophosphate di 1131 1 : 0.78 1:1 M. luteus preparation mixture 6956 1 : 0.87 1:l
4-Hydroxy[U-‘4C,2,3,5,6-3H4)benzoate 7:4
Isopentenyl pyrosphosphate,3,3,- hexa 12 067 7 : 2.9 7:3 dimethylaIly1 pyrophosphate
Famesyl pyrophosphate tri 8343 7 : 2.8 7:3 Geranyl pyrophosphate di 6866 7 : 2.9 7:3 IV. tuteus preparation mixture 10 259 7 : 2.9 7:3
* Expected ratios if the hexaprenyl side chain is biosynthetically all-frons and I-hydroxybenzoate is poly- prenylated at position 3.

497
3-ALL-TRANS - HEXAPRENYL 2.ALL-TRANS -HEXAPRENYL - -L-HYIJROXYBENZOATE PHENOL
OH OSi(CH3J3
3-PERHYDROHEXAPRENYL -I.-HYDROXYBENZOATE
3-PERHYDROHEXAPRENYL -METHYL - L - HYDROXYBENZOATE
3-PERHYDROHEXAPRENYL -METHYL-L-O-TRI - METHYLSILYLBENZOATE
Fig. 2. Chemical and enzymatic reactions used in the characterization of 3-hexaprenyl-4-hydroxybenzo- ate.
similar losses of 3H occur in the formation of 3-polyprenyl-4-hydroxybenzoates from preformed polyprenylpyrophosphates (Table III).
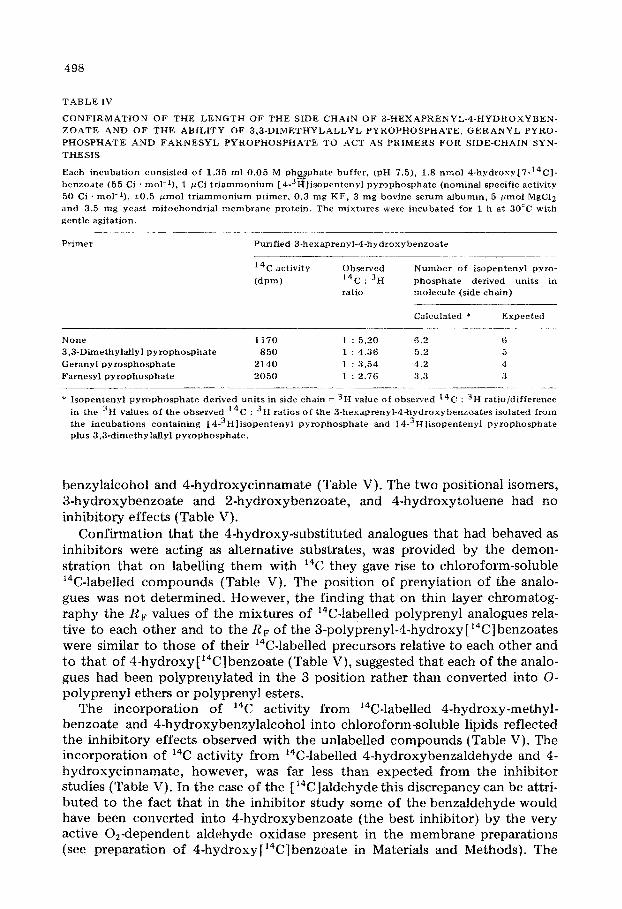
The length of the 3-polyprenyl residue was determined by measuring the dif- ferences in the 14C : 3H ratios of the 14C, 3H-labelled 3-hexaprenyl-4-hydrox- ybenxoates synthesized from 4-hydroxy [ 7-14C] benzoate and [ 4-3H]isopentenyl pyrophosphate in the presence or absence or primers (Table IV). The results of this experiment established that the side chain is 6 prenyl units in length and that 3,3_dimethylallyl pyrophosphate, geranyl pyrophosphate and farnesyl pyrophosphate do act as primers for the synthesis of the hexaprenyl unit (Table IV).
Finally, the hexaprenyl unit was shown to be made up of biogenetically all- truns unsaturated isoprene units by the demonstrations that 3-hexaprenyl-4- hydroxy[ U-14C] benzoate was decarboxylated to a 14C-labelled compound having the thin layer chromatographic properties (adsorptive and reverse phase) expected of 2-hexaprenylphenol [24] (Fig. 2) by a cell-free preparation of Rhodospirillum rubrum (conditions as described in ref. 17, except that the substrate was added in methanol) and that [l-14C, 2S3H]isopentenyl pyro- phosphate was incorporated into 3-hexaprenyl-4-hydroxybenzoate without loss of 3H (Table III).
Ability to use 4-hydroxybenzoate analogues. The effect of modifying or moving the C-l substituent of 4-hydroxybenzoate was investigated. A prelimi- nary study established that the most effective inhibitor of 4-hydroxybenzoate- polyprenyltransferase activity was 4-hydroxybenzoate followed by 4hydrox- ybenzaldehyde, 4-aminobenzoate, methyl-4-hydroxybenzoate, 4-hydroxy-
498
TABLE IV
CONFIRMATION OF THE LENGTH OF THE SIDE CHAIN OF 3-HEXAPRENYL-4-HYDROXYBEN-
ZOATE AND OF THE ABILITY OF 3,3-DI~~THYLALLYL PYROPHOSPHATE, GERANYL PYRO-
PHOSPHATE AND FARNESYL PYROPHOSP~IATE TO ACT AS PRIMERS FOR SIDE-CHAIN SYN-
THESIS
Each incubation consisted of 1.35 ml 0.05 M phmphate buffer, (pH 7.5), 1.8 nmol 4-hydroxy[7-14C1-
benzoate (55 Ci . mol-I), 1 @Ci triammonium [4-3ftilisopentenyl pyrophosphate (nominal specific activity
50 Ci . mol-‘1, kO.5 wnol triammonium pkner, 0.3 mg KF, 3 mg bovine serum albumin, 5 umol MgClZ
and 3.5 mg yeast mitochondrial membrane protein. The mixtures were incubated for 1 h at 30cC with
gentle agitation.
Primer Purified 3-hexaprenyl-4hydroxybenzoate
’ 4C activity
(dpm)
Observed
‘4c : 3I-I ratio
Number of isopentenyl pyro-
phosphate derived units in
molecule (side chain)
Cafcufated * Expected
._____
N0Ile 1170 1 : 5.20 6.2 6
3,3-Dimethylallyl pyrophosphate 850 1 : 4.36 5.2 5
Geranyl pyrosphosphate 2140 1 : 3.54 4.2 4
Farnesyl pyrovhosphate 2050 1 : 2.76 3.3 3
* Isopentenyl pyrophosphate derived units in side chain = 3H value of observed 14C : 3H ratio/difference
in the 3H values of the observed I4 3 C . H ratios of the 3-hexaprenyl-4hydroxybenzoates isolated from
the incubations containing [4-3H]isopentenyl pyrophosphate and 14-3Hlisopentenyl pyrophosphate
plus 3,3-dimethylallyl pyrophosphate.
benzylalcohol and 4-hydroxycinnamate (Table V). The two positional isomers, 3-hydroxybenzoate and Z-hydroxyb~nzoate, and 4-hydroxytoluene had no inhibitory effects (Table V).
Confirmation that the 4-hydroxy-substituted analogues that had behaved as inhibitors were acting as alternative substrates, was provided by the demon- stration that on labelling them with 14C they gave rise to chloroform-soluble “C-labelled compounds (Table V). The position of prenylation of the analo- gues was not determined. However, the finding that on thin layer chromatog- raphy the RF values of the mixtures of 14C-labelled polyprenyl analogues rela- tive to each other and to the RF of the 3-polyprenyl-4-hydroxy[ 14C] benzoates were similar to those of their 14C-labelled precursors relative to each other and to that of 4-hydroxy[‘4C]benzoate (Table V), suggested that each of the analo- gues had been polyprenyIated in the 3 position rather than converted into O- polyprenyl ethers or polyprenyl esters.
The incorporation of 14C activity from 14C-labelled 4-hydroxy-methyl- benzoate and 4-hydroxybenzylalcohol into chloroform-soluble lipids reflected the inhibitory effects observed with the unlabelled compounds (Table V). The incorporation of 14C activity from “C-labelled 4-hydroxybenzaldehyde and 4- hydroxycinnamate, however, was far less than expected from the inhibitor studies (Table V). In the case of the [‘4C]aldehyde this discrepancy can be attri- buted to the fact that in the inhibitor study some of the benzaldehyde would have been converted into 4-hydroxybenzoate (the best inhibitor) by the very active O,-dependent aldehyde oxidase present in the membrane preparations (see preparation of 4-hydroxy[14C]benzoate in Materials and Methods). The

499
TABLE V
INHIBITION OF 4-HYDROXYBENZOATE-POLYPRENYLTRANSFERASE ACTIVITY BY ANAL- OGUES OF 4-HYDROXYBENZOATE. AND POLYPRENYLATION OF 14C ANALOGUES
In the inhibition studies the incubations consisted of 0.95 ml 0.05 M phosphate buffer (pH 7.5). 3.1 nmoi
4hydroxy[7-14CJbenzoate (55 Ci . moth), i450 nmol analogue, 25 firno MgC12, 3 mg yeast mitochon- drial membrane protein and 0.2 ml M. luteus extract that had been preincubated with trilithium isopen- tenyl pyrophosphate (this preparation serves as a source of preformed polyprenylpyrophosphates 13,171). The mixtures were incubated for 30 min at 30°C with gentle agitation. The incubations which contained 14C anslogues consisted of 2.5 ml 0.05 M phosphate buffer (PH 7.5). 7.7 nmol 7-14C- or U-i4C-labelled analogue (7.6 Ci . mol-I). 50 /.nnol MgCl2, 8 mg yeast mitochondrial membrane protein and 0.5 ml M. luteus extract that had been prelncubated with isopentenyl pyrophosphate. The mixtures were incubated for 45 min at 30°C under an atmosphere of Np and were then analysed for the presence of CHC13soluble “C-labelled products. Control incubations from which mitochondrial protein was omitted produced llOlll2.
Dlluent or ’ 4 C-&belled
analogur
Inhibition of Incorporation of l 4C- RF values of ‘4C-
3-polyprenyl-4- labelled analogues into labelled compounds * hydroxybenzoate CHCls-soluble compounds formation
(%) pmol . h-1. % Control Analogue Products rng’ protein
None 0 **
4-Hydroxybenzoate (control) 98 4-Hydroxybenzaldehyde 86 4-Aminobenzoate 65 Methyl-4-hydroxybenzoate 50 4-Hydroxybenzylalcohol 30
4-Hydroxycinnamate 25 3-Hydroxybenzoate 16 2-Hydroxybenzoate 8 4-Hydroxytoluene 3
- - - - 967 x00 0.12 0.35
373 39 0.47 0.47 n.t. I_ - -
527 54 0.54 0.55 250 26 0.25 0.29
83 9 0.08 0.33 n.t. . - - n.t. - - n.t. - - -
* Dn thin layers of Silica gel G developed with diethyl ether/light petroleum/methanol (8 : tj : 1. ,s,v). The use of Silica gel G mead that each of the mixtures of homologues produced under these mcubs- tion conditions 131 behaved as a single component.
** 0% = 487 pm01 h-1. mg-1 protein. wt., not tested.
poor incorporation of 4-hydroxy[U-*4C]cinnamate was at first thought to be due to an effect similar to that just described, since it had been shown that cumcumber mitochondria contain a 4-hydroxybenzoate synthetase which c0nvert.s 4-hydroxycinnamate into 4-hydroxybenzoate [ 251. However, in a series of experiments designed to test this possibility no evidence was obtained that this conversion can occur in yeast mitochondrial membranes (unpublished data).
bob-inuoZ~e~en~ of -SH esters in the formation of 3-hexaprenyl-4-hydroxy- bemoate. It had been proposed that the CoASH esters of 4-hydroxybenzoate and 3-nonaprenyl-4-hydroxybenzoate are the true intermediates in the biosyn- thesis of ubiquinone-9 in the rat, and that 3-nonaprenyl-4-hydroxybenzoate is produced in a side reaction as the result of the hydrolysis of 3-nonaprenyl-4- hydroxybenzoyl CoA by a non-specific deacylase [ 261. The possibility that the formation of 3-hexaprenyl-4-hydroxybenzoate in yeast membranes involved the intermediacy of CoASH esters was difficult to accept, since the well washed matrix-free mitochondrial-rich membrane fractions used for this study would contain no CoASH or ATP (needed for thioester formation). Nevertheless, to examine the remote possibility that CoASH or protein-SH esters might be

500
involved in the formation of 3-hexaprenyl-4-hydroxybenzoate, a series of incubations were set up in which mitochondrial membrane preparations were incubated with 4-hydroxy [ 7-14C] benzoate (55 Ci . mol-‘), isopentenyl pyro- phosphate, 3,3dimethylallyl pyrophosphate, CoASH, ATP and the -SH ester- trapping reagent hydroxylamine. At the end of the incubation period, the reac- tion mixtures were extracted with acetone [27] and the extracts examined by thin-layer chromatography for the presence of hydroxamates of 4-hydroxy- benzoate and 3-hexaprenyl-4-hydroxybenzoate. In none of the incubations per- formed were any compounds other than 4-hydroxybenzoate and 3-hexaprenyl- 4-hydroxybenzoate found to be present in the acetone extracts, and it was concluded that CoASH esters or protein-SH esters are not intermediates in the formation of 3-hexaprenyl-4-hydroxybenzoate.
Discussion
It has been established that a 10 000 X g matrix-free mitochondrial mem- brane-rich preparation from commercial bakers’ yeast is able to synthesize 3- all-trans-hexaprenyl-4-hydroxybenzoate from 4-hydroxybenzoate and iso- pentenyl pyrophosphate (Tables I-IV). The synthesis is Mg2+ dependent and is stimulated markedly by the primer for polyprenylpyrophosphate synthesis, 3,3-dimethylallyl pyrophosphate (Tables I and II). At the optimal Mg2+ concen- tration (3.7 mM) for the synthesis of 3-hexaprenyl-4-hydroxybenzoate from 4- hydroxybenzoate, isopentenyl pyrophosphate and 3,3-dimethylalIy1 pyro- phosphate the priming function of 3,3-dimethylallyl pyrophosphate can be performed by either geranyl pyrophosphate (most efficient) or farnesyl pyro- phosphate (Table II). At a high Mg*+ concentration (33.3 mM), however, geranyl pyrophosphate and farnesyl pyrophosphate act mainly as sources of preformed side chains and 3-diprenyl- and 3-triprenyl-4-hydroxybenzoate,
.respectively are produced (Table II). No evidence was obtained to support the proposal that 3-polyprenyl-4-hydroxybenzoates are produced as the result of the action of non-specific CoA lyases on 3-polyprenyl-4-hydroxybenzoyl CoAs formed from 4-hydroxybenzoyl CoA [ 261.
In the light of the above results the failure to demonstrate the formation of 3~~~-~~~~s-hexaprenyl-4-hydroxybenzoate by yeast mitochondrial membranes in a previous study [ 31 is attributable to the use of a 4-hydroxy]J4C] benzoate species with a low specific activity and to the poor rate of synthesis of 3-hexa- prenyl-4-hydroxybenzoate at high Mg2+ concentrations and in the absence of a primer for side-chain formation.
The demonstration that 3-hexaprenyl-4-hydroxybenzoate is an efficient precursor of 5-demethyoxyubiquinone-6 and ubiquinone-6 in whole mitochon- dria of yeast would seem to establish it as the first intermediate on the pathway leading from 4-hydroxybenzoate to ubiquinone6 in yeast mitochondria [ 51. If this is the case then the next two intermediates on the pathway are likefy to be the 3-hexaprenyl-4,5dihydroxybenzoate and 3-hexaprenyl-4-hydroxy-5 methoxybenzoate recently isolated from ubiquinone-deficient mutants of yeast [28,29]. Indeed, the group who isolated these two compounds have been led to suggest that the biosynthesis of ubiquinone in all eukaryotic ceils proceeds via the general pathway: 4-hydroxybenzoate + 3-polyprenyl-4-hydroxybenzoate -+

501
3-polyprenyl-4,5_dihydroxybenzoate + 3-polyprenyl-4-hydroxy-4-methoxy- benzoate -+ 3-polyprenyl-4-hydroxy-5-methoxybenzoate + 2-polyprenyl-6- methoxyphenol-+ ubiquinone. However, the facts that membrane preparations of yeast are able to catalyse the polyprenylation of five analogues of 4-hydroxy- benzoate (Table V) and that rat liver and heart mitochondrial preparations catalyse the polyprenylation of 4-aminobenzoate, 3,4dihydroxybenzoate and 3- hydroxy-4-methoxybenzoate [ 30,311, raises the possibility that some of the mitochondrial biosynthesis of ubiquinone may proceed by alternative pathways to the one involving the formation of 3-polyprenyl-4-hydroxybenzoate from 4- hydroxybenzoate and an appropriate polyprenylpyrophosphate [ 5,281 e.g. 4- hydroxybenzoate + 3,4_dihydroxybenzoate + 3,4-dihydroxy-5-polyprenyl- benzoate-+ ubiquinone [ 311.
In rat liver the isopentenyl pyrophosphate isomerase, diprenyltransferase and 4-hydroxybenzoate-polyprenyl-transferase activities needed to make the 14C- labelled compounds with the chromatographic properties of 3-polyprenyl-4- hydroxybenzoates from 4-hydroxy[ 14C] benzoate and isopentenyl pyrophos- phate are associated with the inner membranes of the mitochondrial [ 21, and it is known from previous studies that 4-hydroxybenzoate-polyprenyltransferase activity is also associated with yeast mitochondrial membranes [3]. In the present study it was found that whilst yeast membrane fractions, as judged by their ability to form 3-hexaprenyl-4-hydroxybenzoate from 4-hydroxybenzoate and isopentenyl pyrophosphate (Tables I--IV), must also posses isopentenyl pyrophosphate isomerase and hexaprenylpyrophosphate synthetase activities, it is necessary to add 3,3_dimethylallyl pyrophosphate to obtain good rates of synthesis of 3-hexaprenyl-4-hydroxybenzoate (Table I). The latter finding sug- gests that the isomerase is a mitochondrial matrix enzyme. Support for this proposal is provided by the finding that the synthesis of 3-hexaprenyl-4- hydroxybenzoate by whole mitochondria of yeast is independent of the presence of 3,3dimethylallyl pyrophosphate in the incubation mixture [5]. Further evidence for the localisation of the hexaprenylpyrophosphate syn- thetase activity on the mitochondrial membrane was provided by the demon- strations that the rate of synthesis of 3-hexaprenyl-4-hydroxybenzoate from 4- hydroxybenzoate, isopentenyl pyrophosphate and dimethylallyl pyrophos- phate is unaffected by repeated washing of the membrane preparation and that the distribution of the small but measurable amounts of membrane- bound (di)prenyl transferase activity paralleled the distribution of the succinate dehydrogenase acitivty (Fig. 1). The acid hydrolysis products of this activity, however, were found to be farnesol and nerolidol (formed by the hydrol- ysis of farnesyl pyrophosphate) and two hydrocarbons (one of which ran with the RF of squalene and the acid hydrolysis product of presqualene pyro- phosphate) and not the expected hexaprenol and its tertiary isomer. A possible explanation for this finding is that under the conditions of the assay [‘“Cl- farnesyl pyrophosphate produced by the hexaprenylpyrophosphate synthetase is converted into presqualene pyrophosphate (by the very active squalene syn- thetase present in the microsomes which contaminate the preparation) before it can be used as a substrate for further chain elongation reactions. In this respect it is of interest that mitochondrial preparations from germinating castor bean synthesize farnesyl pyrophosphate and geranylgeranyl pyrophosphate from iso-

502
pentenyl pyrophosphate, and not the expected decaprenyl pyrophosphate [23]. It is also worth pointing out that in the studies with rat liver mitochon- drial membrane preparations the identities of the compounds formed from iso- pentenyl pyrophosphate in the (di)prenyl transferase assays were not deter- mined [2]. In vitro the hexaprenyl pyrophosphate synthetase (measured indirectly) is not restricted to the use of 3,3-dimethylallyl pyrophosphate as a primer and can make use of either geranyl pyrophosphate or farnesyl pyrophos- phate (Tables II and IV). Indeed, in view of the recent demonstration that rat liver mitochondria are able to use solanesyl (nonaprenyl) pyrophosphate as a ‘primer’ for the synthesis of the side chain of 3-decaprenyl-4-hydroxy- benzoate [33], it is to be expected that in vitro the yeast synthetase will be able to make use of geranylgeranyl pyrophosphate and pentaprenyl pyrophos- phate as ‘primers’ for the synthesis of hexaprenyl pyrophosphate.
Acknowledgements
This work was supported by the Science Research Council. We thank Mrs. Susan Swetez for expert technical assistance, Mr. C.F.G. Jones for the gas- liquid chromatography determination and Miss Eileen Sharpe for typing the manuscript.
References
1 Leppik. R.A., Stroobant, P.. Shineberg. B., Young, LG. and Gibson, F. (1976) Biochim. Biophys.
Acta 428. 146-156
2 Momose. K. and Rudney, H. (1972) J. Biol. Chem. 247, 3930-3940
3 Thomas. G. and Threlfall, D.R. (1974) Phytochem. 13.1825-1833
4 Thomas, G. and Threlfall, D.R. (1975) Phytochem. 14, 2607-2615
5 Casey, J. and Threlfall, D.R. (1978) FEBS Lett. 85. 249-253
6 Parson, W.W. and Rudney, H. (1965) J. Biol. Chem. 240. 1855-1863
7 Whistance, G.R. and Threlfall. D.R. (1970) Biochem. J. 117, 593-600
8 Hartman. L. and Law, R.C.A. (1973) Lab. Practice 22, 475-477
9 Green, T.R. and B&ted. D.J. (1972) Biochem. J. 130, 983-995
10 Donninger, C. and Popj&, G. (1967) Biochem. J. 105, 505-547
11 Cramer, F. and Bohm, W. (1959) Angew Chem. 71, 775-787
12 Cornforth. R.H. and PopjLk. G. (1969) Methods Enzymol. 15. 359-390
13 Lord, J.M. and Merrett, M.J. (1971) Biochem. .J. 124, 275-281
14 Collins, N. and Men‘ett, M.J. (1975) Biochem. J. 148. 321.-328
15 Haas, E. (1966) Methods Enzymol. 2, 712-719
16 Popjgk, G. (1969) Methods Enzymol. 15. 393-454
17 Raman, T.S., Rudney. H. and Buzelli, N.K. (1969) Arch. Biochem. Biophys. 130, 164-174
18 Okita, G.T., Kabara, J.J., Richardson, F. and LeRoy, G.V. (1957) Nucleonics 15 (No. 6). 111-114
19 Pierce Handbook and General Catalogue (1977-1978) p. 234, Pierce and Warriner (U.K.) Ltd.
20 Plummer, D.T. (1971) An Introduction to Practical Biochemistry, pp. l-369, McGraw-Hill, U.K.
21 Bandlow, W. and Baeur, P. (1976) Methods Cell. Biol. 12, 311-335
22 Popjik, G.. Edmond, J., Clifford, K. and Williams, V. (1969) J. Biol. Chem. 244. 1897-1918
23 Whistance, G.R. and Threlfall. D.R. (1970) Phytochem. 2, 213-219
24 Spiller. G.H., Threlfall, D.R. and Whistance. G.R. (1968) Arch. Biochem. Biophys. 125, 786-796
25 Hagel, P. and Kindle, H. (1975) FEBS Lett. 59, 120-124
26 Trumpower, B.L., Housser. R.M. and Olson, R.E. (1974) J. Biol. Chem. 249, 3041-3048
27 Hahlbrock, K. and Grisebach, H. (1970) FEBS Lett. 11, 6244
28 Goewert, R.R.. Sippel, J. and Olson. R.E. (1977) Biochem. Biophys. Res. Commun. 76, 282-287
29 Goewert, R.R., Sippel, J.. Grimm, M.F. and Olson, R.E. (1978) FEBS Lett. 87, 219-221
30 Alam. S.S., Nambudiri. A.M.D. and Rudney, H. (1975) Arch. Biochem. Biophys. 171,183-190
31 Nambudiri, A.M.D., Brockman. D., Alam, S.S. and Rudney, H. (1977) Biochem. Biophys. Res. Com-
mun. 76,282-287
32 Green, T.R., Dennis, A.T. and West, C.A. (1975) Biochem. Biophys. Res. Commun. 64. 976-982
33 Nishino. T. and Rudney, H. (1977) Biochemistry 16. 605-609





























