Embed Size (px)
Citation preview

Forest Seedling Establishment in Neotropical Savannas: Transplant Experiments with Xylopiafrutescens and Calophyllum brasilienseAuthor(s): Martin KellmanSource: Journal of Biogeography, Vol. 12, No. 4 (Jul., 1985), pp. 373-379Published by: WileyStable URL: http://www.jstor.org/stable/2844868 .
Accessed: 19/09/2013 14:40
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Wiley is collaborating with JSTOR to digitize, preserve and extend access to Journal of Biogeography.
http://www.jstor.org
This content downloaded from 129.173.72.87 on Thu, 19 Sep 2013 14:40:44 PMAll use subject to JSTOR Terms and Conditions

Journal of Biogeography (1985) 12, 373-379
Forest seedling establishment in Neotropical savannas: transplant experiments with Xylopiafrutescens and Calophyllumn brasiliense
MARTIN KELLMAN Department of Geography, York University, Downsview, Ontario, Canada M3J 1 P3
ABSTRACT. Observations on the survivorship, growth and tissue nutrient content of seedlings of two forest tree species have been used to test the hypo- thesis that trees and shrubs of Neotropical savannas facilitate the establishment of forest trees. Xylopia frutescens was grown from seed planted beneath and beyond the cover of five savanna trees and shrubs, and observed for 5 years. Calophyllum brasiliense seedlings were transplanted from forest to sites beneath and beyond Byrsonima crassifolia trees and observed for 3 years. In both species survivorship and growth of seedlings was superior beneath tree covers and it was unlikely that any open-grown plant would survive to reproduce. Xylopia seedling growth beneath Miconia albicans was also superior to that beneath any other cover type. Despite these differences in performance, an analysis of mineral nutrient content of tissues of seedlings from covered and open sites failed to show dramatic differences, although there was evidence of lower K contents in poorly performing Xylopia seedlings and of lower Ca concentrations in open- grown Calophyllum seedlings. The results confirm the facilitative role that can be played by woody savanna plants during forest invasion of savannas, and it is suggested that a closed thicket of these plants is a probable early stage in the transformation process.
Introduction
In an earlier paper (Kellman, 1979), the soils beneath woody dicot trees and shrubs in a Neotropical savanna in Belize were shown to be preferentially enriched with mineral nutrients, and these sites were postulated to be the nuclei of tree seedling establishment during forest invasions of savanna. In a sub- sequent experiment this hypothesis was tested in a preliminary way by planting seed of several forest tree species beneath and beyond woody plants in the savanna (Kellman & Miyanishi, 1982). Low germination rates in this experiment resulted in data being available on the performance of only two species: Swietenia macro phylla King and Xylopia frutescens Aubl. Swietenia failed ultimately at
24
all sites and a growth chamber experiment im- plicated K deficiency as the likely cause. How- ever, Xylopia showed much greater survivor- ship over 2 years, and performance of these seedlings over 3 further years is reported here. Because of the low germination rates achieved in this work, a further experiment was initiated using bare-root transplantation of seedlings of Calophyllum brasiliense Camb. var. Rekoi Standl. Three years' results from this trans- plantation experiment were also reported here.
Xylopia frutescens is a small tree that is found commonly in low woodland and savanna-edge thickets throughout Central America. In the Mountain Pine Ridge savanna, where the experiments took place, seedlings and saplings of Xylopia are widely scattered within areas that have, not been burned for
373
This content downloaded from 129.173.72.87 on Thu, 19 Sep 2013 14:40:44 PMAll use subject to JSTOR Terms and Conditions

374 Martin Kellman
several decades. Calophyllum brasiliense var. Rekoi is a large rain forest tree common in forests throughout Central America. Occa- sional saplings of this species are also found in parts of the Mountain Pine Ridge savanna that have not been burned for several decades.
The experiments took place in the Mountain Pine Ridge savanna, Belize (lat. 170N, long. 890W) near the settlement of Augustine. The area is a granite plateau with a surface eleva- tion of approximately 500 m and receives an annual rainfall of 1560 mm with a pronounced dry season from February to April (Walker, 1973). In the western part of the savanna, where the experiments took place, wildfires have been suppressed for approximately 30 years and today much of the area comprises open stands of Pinus caribaea Morelet, within which occur occasional patches of treeless grassland and thickets of woody dicot trees and shrubs. The soils of the area are intensely weathered Ultisols with coarse sandy loam topsoils of variable depth overlying a clay-rich B horizon. Surface soil beneath treeless grass- land is acid (pH 5.0), of low cation exchange capacity (- 21 mequiv. per 100 g), low base saturation (; 2.5%) and low in available phos- phorus (; 1 ppm) and total nitrogen (- 0.05%). Further details of the soils are provided in Kellman (1979), Kellman & Hudson (1982) and Kellman & Sanmugadas (1985).
Methods
Xylopia seed was planted on 26 and 27 May 1979 beneath the following savanna trees and shrubs: Byrsonima crassifolia (L.) Kunth, Quercus oleoides Cham & Schlecht., Q. schippii Standl., Miconia albicans (Sw.) Triana and Clethra hondurensis Britton, and in treeless areas beyond each cover type. Two seeds were planted at ten points beneath and beyond each tree. Survivorship and height of seedlings were measured at 9, 19, 24, 35, 48 and 60 months after planting, and leaflet numbers counted in the last three observation periods. The Clethra site was inadvertently burned in early 1982 and survivorship data are collated excluding seedlings at this site. However, the data on seedling heights at this site for the first three observation periods are used in calculating means.
Two hundred and ten Calophyllum seed- lings were transplanted between 16 and 23 June 1981 from riparian forest on alluvium to sites beneath and beyond four Byrsonima trees within the savanna. Seedlings appeared to be all of a single cohort less than 1 year old, and many had seed cases still attached. Seedlings were exhumed with a shovelful of forest soil, placed in a plastic sack for transport, and kept moist until transplantation. At the transplant sites forest soil was removed from seedlings' roots and each was planted 50 cm apart along permanently marked lines in equal sets of 105 seedlings beneath and beyond the Byrsonima canopies. Seedlings were planted in spade slits that were then tamped firmly closed. The height and leaf number of each seedling were then recorded. Seedlings at open sites suffered high mortality during the first week after transplantation, apparently as a result of sun- scorch. Consequently eighty-nine dead and dying seedlings were replaced with new trans- plants on 29 and 30 June 1981 and all seed- lings at open sites were covered with a thin layer of grass and pine needle litter. This litter cover was removed 1 month later before leaving the field area, and dead seedlings recorded. These dead seedlings were excluded from the starting cohort, leaving steventy-six seedlings in open sites and 105 at covered sites. Survivorship, height and leaf number of each seedling were recorded at 1, 2 and 3 years after transplanting.
Tissue subsamples for chemical analysis were collected from both Xylopia and Calophy- llum seedlings at the time of final observations in May 1984. Leaves of ten randomly-selected Calophyllum seedlings in open and tree-covered sites were collected for analysis of Ca, Mg, K, Na and P content. Two further randomly- selected sets of leaf tissue were collected for analysis of N content. In each set, where in- sufficient leaf tissue for analysis was available on one seedling, that of the closest unsampled seedling was added to make a composite sample. The small size and low numbers of Xylopia seedlings surviving to 1984 precluded the analysis of leaf tissues only, or of seedlings separately. Consequently all seedings were harvested at ground level and the stem and leaf tissue was bulked into five composite samples, one for each cover type plus a single open class. Insufficient material was available
This content downloaded from 129.173.72.87 on Thu, 19 Sep 2013 14:40:44 PMAll use subject to JSTOR Terms and Conditions
100 ( a) Survivorship 26 (b) Height (covered vs. open) 26 (c) Height (Miconia -covered vs other 80-
26- 26- ~~~~~~~~~~~~~~~~~~~~~~~~~~~~tree covers)
24- 1 + Covered sites 24 Miconia-covered sites
|0- > _ _22 4 Open sites 22 4- Other tree covers
201 204
18- 18-
16- 16- E -
C: ~~~~~~~~~~~14- 14-
12 - 12-
10- 10-
8 8-
* Covered sites 6y 6-
o Opn sites 4* 4*
2- 2- NS ~ * NS NS NS N SN
I I I ~~~~~0 0 10 20 30 40 50 60 10 20 30 40 50 60 10 20 30 40 50 6
Months Months Months
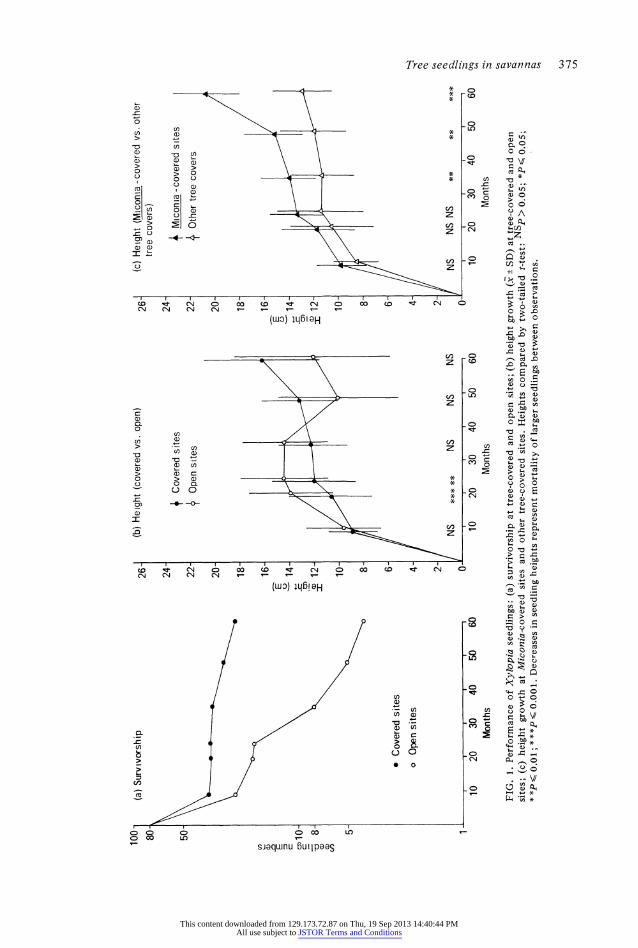
FIG. 1. Performance of Xylopia seedlings: (a) survivorship at tree-covered and open sites; (b) height growth (x ? SD) at tree-covered and open sites; (c) height growth at Miconia-covered sites and other tree-covered sites. Heights compared by two-tailed t-test: NSp > 0.05; *J1< 0.05; **P 0.01 ; **P< 0.001. Decreases in seedling heights represent mortality of larger seedlings between observations. Z
This content downloaded from 129.173.72.87 on Thu, 19 Sep 2013 14:40:44 PMAll use subject to JSTOR Terms and Conditions

376 AMartin Kellman
for an analysis of N content of Xylopia tissues. Tissue samples for cation and P analysis
were dry ashed after grinding in a Wiley mill (No. 40 mesh screen) and the ash taken up in 1N HCI. Cations in solution were determined by atomic absorption spectrophotometry and phosphorus concentration by the molyb- dophosphoric blue method using an auto- analyser. Extracts for N determination were prepared by wet digestion (Issac & Johnson, 1976) and concentrations determined colori- metrically using an autoanalyser.
Results
Xylopia
Survivorship of Xylopia seedlings over 5 years is shown in Fig. l(a). Both seedling sets have been subject to relatively constant mor- tality since the first observation period, but with rates at the open sites being much higher than at tree-covered sites. Should the prevailing mortality rates continue, no seedlings would remain alive at the open sites after 8 years, while some plants could persist at the tree- covered sites for approximately 30 years. Despite having a higher mortality rate in open sites, Xylopia seedlings grew more rapidly there for the first 2 years (Fig. lb). However, by 3 years, mean heights and leaf numbers in open sites had dropped below those at covered sites, although the differences were not signifi- cant due to large variances. Pooling data for Xylopia seedlings at all tree-covered sites ob- scured a major dichotomy in seedling perfor- mance: seedlings planted beneath Miconia were distinctly less chlorotic, and apparently growing more vigorously, than those beneath any other tree cover throughout the experi- ment. A comparison of seedling growth at this site with that at other tree-covered sites showed that both heights and leaflet numbers were significantly greater there by 3 years,
TABLE 1. Leaflet numbers on Xylopia seedlings at 3, 4 and 5 years after planting (mean ? SD)
Year Open Miconia-covered Other tree- sites sites covered sites
1 982 5.20 ? 0.75 1 8.45 ? 6.42 8.30 ? 4.83 1 983 5.20 ? 2.28 24.10 ? 8.47 5.83 ? 3.20 1984 4.25 ? 2.06 37.30 ? 12.67 7.14 ? 3.70
and that the difference became more pronoun- ced in each of the two subsequent years (Fig. I c, Table 1).
The chemical analyses of above-ground Xylopia tissues are of limited value because of the absence of replication. The analyses of bulked seedling samples (Table 2) showed that plants at open sites had slightly lower concen- tration of P, and a substantially lower concen- tration of K, in their tissues than plants at any tree-covered site. In contrast, plants beneath Miconia, where growth was most rapid, had higher concentrations of K in their tissues than those of any other site, but rather low concentrations of P. The data thus provide some suggestion that the availability of K may have been involved in the differences in per- formance observed.
Calophyllum
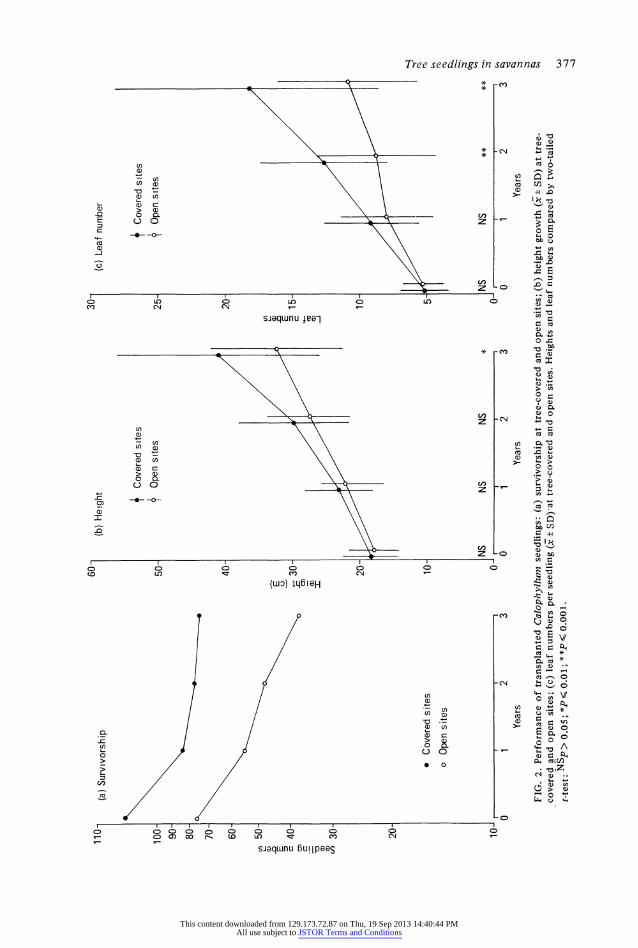
The difference in survivorship of Calophy- llum seedlings at open and tree-covered sites over three years (Fig. 2a) were less dramatic than those observed in Xylopia seedlings. However, while mortality rates declined at the tree-covered sites over the 3-year period, those at the open sites were both higher and rela- tively constant. Many open-growth seedlings were also chlorotic at each observation period, while those beneath trees were generally healthy looking. Heights and leaf numbers of seedlings transplanted to open and tree- covered sites were not significantly different at the time of transplantation, or 1 year later (Fig. 2b, c). However, leaf numbers per seed-
TABLE 2. Macro-nutrient content (%) of bulked leaf and stem tissues of Xylopia seedlings harvested at 5 years
Cover Ca Mg K Na P
Byrsonima crassifolia 0.56 9 0.142 0.308 0.036 0.039 Quercus oleoides 0.271 0.218 0.372 0.051 0.043 Quercus schippii 0.526 0.202 0.351 0.073 0.054 Miconia albicans 0.579 0.210 0.434 0.020 0.027 Open 0.651 0.159 0.244 0.053 0.024
This content downloaded from 129.173.72.87 on Thu, 19 Sep 2013 14:40:44 PMAll use subject to JSTOR Terms and Conditions
110 (a) Survivorship 60- (b) Height 30 (c) Leaf number
Covered sites | Covered sites
100 50- ' Open sites 25- Open sites 90-
80-
70 1
60 40 20/
u)50-
-E O E o ~~~~~~~~~~~~~~~~~~~~~~~E 40 r 30, , 15,
c 30-
20 101
20-
Yea rs Years Yea rs >
FIG. 2. Performance of transplanted Calophyllum seedlings: (a) survivorship at tree-covered and open sites; (b) height growth (x ? SD) at tree- covered and open sites; (c) leaf numbers per seedling +SD)lat tree-covered and open sites. Heights and leaf numbers compared by two-tailed t-test:NNSp> N 05; *PN 0.01; **P AN 0.001. *
This content downloaded from 129.173.72.87 on Thu, 19 Sep 2013 14:40:44 PMAll use subject to JSTOR Terms and Conditions

3 78 Martin Kellman
TABLE 3. Macro-nutrient content of Calophyllum seedling leaf tissues harvested 3 years after transplantation (mean % ? SD; z = 1O). Comparison by one-tailed t-test.
Cover Ca Mg K Na P N
Byrsonima 0.83 5 0.118 0.743 0.040 0.032 0.798 crassifolia ? 0.171 ?0.033 ?0.120 ?0.012 ?0.004 ?0.082
Open 0.667 0.105 0.793 0.049 0.034 0.805 ?0.200 ?0.025 ?0.101 ?0.012 ?0.005 ? 0.119
t 2.02* 0.98NS -1.01NS -1.63NS -1.28NS -0.16NS
NSp> o0o5; *p60.05.
ling were significantly greater at tree-covered sites after 2 years, with the difference be9oming more pronounced after 3 years. Height growth of seedlings was relatively constant at the open sites (Fig. 2b) but was accelerating at the tree-covered sites, where seedlings were signi- ficantly taller at the final observation in 1984 (Fig. 2b).
Despite the inferior appearance and perfor- mance of open-grown Calophyllum seedlings, chemical analysis of their leaf tissues did not show consistently lower concentrations of macro-nutrients than in the leaves of tree- covered seedlings (Table 3). Only Ca concen- trations were significantly lower in open- grown than in tree-covered leaf tissues.
Discussion
The results confirm the superior performance of seedlings of both species when growing beneath savanna trees as opposed to open grassland. Although young plants of both species are now establishing in fire-protected savanna, and so appear to be tolerant of savanna conditions in general, neither seems capable of successful establishment and growth to a reproductive stage in treeless grassland. The savanna trees thus appear to play a facilitative role (sensu Connell & Slatyer, 1977) in the transformation of grassland to forest.
Despite the differences in performance of seedlings at open and tree-covered sites, chemical analyses of tissues showed fewer dif- ferences in concentrations of macro-nutrients than had been anticipated. Several potential explanations exist for this discrepancy. (i) The differences in nutrient element concentration,
while small and selective, may have been suffi- cient to account for the observed differences in performance. (ii) Alternatively, significant differences may have existed in micro-nutrients that were not analysed. (iii) A third possibility is that differences in performance at the two sites may have been associated with some non- nutrient effect, such as intolerance of high light or grass competition at the open sites. However, this appears to be an unlikely explanation for differences in performance of Xylopia seedlings, whose height growth in open areas was superior to that at tree-covered sites over the first 2 years of observation (Fig. 1 b). (iv) A final explanation for the discrepancy may be a response to nutrient deficiency in both species that involved overall reductions in growth rate without the earlier appearance of lowered tissue concentrations.
Slow growth appears to be an important adaptation of plants occupying infertile soils (Chapin, 1980), and is probably crucial to the success of young forest trees establishing in savanna, where net nutrient accumulation must derive from atmospheric sources (Kellman & Miyanishi, 1982; Kellman & Hudson, 1982; Kellman, Hudson & Sanmugadas, 1982). The high mortality rate in the larger open-grown Xylopia seedlings that resulted in actual decreases in mean seedling heights at 4 and 5 years (Fig. I b), illustrates the disadvantages of rapid growth in this environment. It is there- fore possible that the slower growth rates imposed by low light levels beneath tree canopies may have partially alleviated soil nutrient deficiencies and contributed to the greater survivorship of seedlings beneath them.
Results of the Xylopia experiment suggest that the facilitative effect of different tree covers vary. It is possible that these differences
This content downloaded from 129.173.72.87 on Thu, 19 Sep 2013 14:40:44 PMAll use subject to JSTOR Terms and Conditions

reflect not only differing degrees of soil en- richment beneath different cover types, but also differences in the availability of mycor- rhizal fungal associates there. The superior performance of Xylopia seedlings beneath the shrub Miconia albicans may also have derived in part from the overstorey of Pinus caribaea that existed at this site. The canopy of this species appears to act as a filter of aerosols and can increase the input of nutrient elements from atmospheric souces (Kellman, unpub- lished).
Irrespective of the mechanisms involved the results of these transplant experiments strongly suggest that the establishment of forest seedlings in savanna requires the pre- existence of woody savanna plants to act as 'nurse' trees. Observation in both the Mountain Pine Ridge savanna and the Lanos savannas of Venezuela indicate that populations of woody savanna plants can expand rapidly once fire is suppressed (Miyanishi, 1984; San Jose & Farirnas, 1983), and pollen records from Neo- tropical savannas record periodic increases in the pollen of woody taxa such as Byrsonima and Curatella (Wymstra & van der Hammen, 1966). The establishment of relatively con- tinuous woody thickets in savanna also reduces the flammability of fuels and the potential for spread of fire (Kellman, 1 984), and is probably a normal precursor to forest development in such areas. After 30 years of fire suppression in the Mountain Pine Ridge savanna, widely scattered, but apparently successfully estab- lished, young plants of several rain forest tree species can be seen. In addition to Calophy- llum, those observed include Vochysia hon- durensis Sprague, Term inalia obovata (R. & P.) Steud, Bursera simaruba (L.) Sarg. and Sima- ruba glauca D.C. In contrast, other locally common rain forest species, such as Swietenia macrophylla, Cedrela mexicana Roem., Ceiba pen tandra (L.) Gaertn. and Achras sapote L. show no sign of invading the savanna, sugges- ting that the first generation forest developing on such sites will comprise a distinct subset of the regional forest flora.
Tree seedlings in savannas 379
Acknowledgments
I am grateful to Kiyoko Miyanishi for assis- tance in the field and to Kelly Robertson for laboratory assistance. The cooperation of the Belize Forestry Department has been much appreciated. Financial support was provided by a Guggenheim Fellowship and an operating grant from the Natural Sciences and En- gineering Research Council of Canada.
References Chapin, F.S. (1980) The mineral nutrition of wild
plants. Ann. Rev. Ecol. Syst. 11, 233-260. Connell, J.H. & Slatyer, R.O. (1977) Mechanisms of
succession in natural communities and their role in community stability and organization. Amer. Nat. 111, 1119-1144.
Issac, R.A. & Johnson, W.C. (1976) Determination of total nitrogen in plant tissue using a block digester. J. Ass. offic. analyt. Chem. 59, 98-100.
Kellman, M. (1979) Soil enrichment by Neotropical savanna trees. J. Ecol. 67, 56 5-577.
Kellman, M. (1 984) Synergistic relationships between fire and low soil fertility in Neotropical savannas: a hypothesis. Biotropica, 16, 1 5 8-160.
Kellman, M. & Hudson, J. (1982) Nutrition of Pinus caribaea in its native savanna habitat. Plant & Soil, 64, 381-391.
Kellman, M., Hudson, J. & Sanmugadas, K. (1982) Temporal variability in atmospheric nutrient in- flux to a tropical ecosystem. Biotropica, 14, 1 -9.
Kellman, M. & Miyanishi, K. (1982) Forest seedling establishment in Neotropical savannas: observa- tions and experiments in the Mountain Pine Ridge savanna, Belize. J. Biogeogr. 9, 193-206.
Kellman, M. & Sanmugadas, K. (1985) Nutrient retention by savanna ecosystems. I. Retention in the absence of fire. J. Ecol. 73, (in press).
Miyanishi, K. (1984) The effects of prescribed burning on the population dynamics of Miconia albicans and Clidemia sericea. Ph.D. thesis, York Univer- sity, Toronto.
San Jose, J.J. & Farinias, M.R. (1 983) Changes in tree density and species composition in a protected Trachypogon savanna, Venezuela. Ecology, 64, 447-453.
Walker, S.H. (1973) Summary of climatic records for Belize. Supp. Rept. No. 3, Land Resources Divi- sion, Surbiton, Surrey.
Wymstra, T.A. & van der Hammen, T. (1966) Palyno- logical data on the history of tropical savannas in northern South America. Leid. geol. Meded. 38, 71-90.
This content downloaded from 129.173.72.87 on Thu, 19 Sep 2013 14:40:44 PMAll use subject to JSTOR Terms and Conditions













![Chemical Constituents of Methanol Fruit Extract of Xylopia ...inadequate[9]. Scientific evaluation of ethno pharmacological information from medicinal plants is necessary for the development](https://img.pdfslide.us/doc/110x75/5f07c78c7e708231d41eb43e/chemical-constituents-of-methanol-fruit-extract-of-xylopia-inadequate9-scientific.jpg)















