Embed Size (px)
Citation preview

BACTERIOLOGICAL REVIEWS, Sept. 1973, p. 343-370Copyright © 1973 American Society for Microbiology
Vol. 37, No. 3Printed in U.S.A.
Cyanophages-Viruses Attacking Blue-GreenAlgae
ETANA PADAN AND MOSHE SHILODepartment of Microbiological Chemistry, The Hebrew University-Hadassah Medical School,
Jerusalem, Israel
INTRODUCTION .................................. 343CHARACTERISTICS OF CYANOPHAGES .................................. 344Typology .................................. 344Host Range.............................................................. 345Morphology.......... 346Composition.......... 348Nucleic acids ..........348Proteins.......... 348
Physicochemical Characteristics ..... .......................................... 349GROWTH CYCLE OF CYANOPHAGES ........... ............................... 349Phage Development in Filamentous Blue-Green Algae ....... .................... 349Sequence of Synthesis of Cyanophage Macromolecules ....... ................... 351DNA.................................................................... 351Protein synthesis ............................................................. 352
Morphological Aspects of the Cyanophage Infective Process ...... ............... 352INTERACTION BETWEEN CYANOPHAGE DEVELOPMENT AND THE PHO-TOAUTOTROPHIC METABOLISM OF THE BLUE-GREEN ALGAL HOST..... 356
Effect of Cyanophage Development on Host Metabolism.358Photosynthesis during infection ............. .................................. 358Host DNA and protein synthesis during infection ........ ...................... 359
Dependence of Cyanophage Development on the Photoautotrophic Metabolism ofthe Host ................................................................... 359
Photosynthesis and respiration ............. .................................. 359Building blocks ............................................................... 361
ECOLOGICAL CONSIDERATIONS ............. ................................. 362GENETICS OF CYANOPHAGES ............... .................................. 365CONCLUDING REMARKS....................................................... 365ACKNOWLEDGMENTS......................................................... 365LITERATURE CITED ............................................................. 366
INTRODUCTION
Hardly a decade has passed since Saffermanand Morris (106) reported the discovery of aviral agent which attacks and lyses severalspecies of blue-green algae. These agents, nowknown to attack a wide range of blue-greenalgae, have been variously called phycoviruses(113), algophages (42), blue-green algal viruses(77, 107), and cyanophages (77, 94). The latterdesignation seems most apt because of theirclose analogies to bacteriophages and the nowwidely accepted close affinity of the Cyano-phyta to the bacteria (143).The significance and implications of the cya-
nophages have been faithfully reflected in thevolume and range, both geographical and disci-plinary, of the investigations undertaken of thealga-cyanophage system. In undertaking this
review of cyanophages, we are well aware of thegreat many questions still open to inquiry andhave endeavored to indicate them in the light ofthe results summarized from published andsome as yet unpublished data.The interest in the cyanophages stems in no
small measure from the importance, both theo-retical and practical, of their hosts. The blue-green algae occupy a special position in thebiological world. They resemble bacteria intheir prokaryotic cellular structure. However,the photoautotrophic metabolism of the blue-green algae resembles that of eukaryotic plants.The blue-green algae are among the most an-cient organisms on earth, appearing in MiddlePrecambrian rock and, perhaps, earlier (115).Nevertheless, a monophyletic origin and limitedgenetic diversification during evolution of thefilamentous blue-green algae are suggested by
343
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

PADAN AND SHILO
their uniform deoxyribonucleic acid (DNA)base ratio (31).
Until the discovery of the cyanophages, pho-toautotrophic metabolism under viral infectioncould only be studied in tissues of higher plants.Although not isolated, viruses attacking eukar-yotic algae have been indicated in severalworks, as mentioned in a recent review on algalviruses (18). Only recently a virus attacking aphotosynthetic bacterium has been isolated(34). Data collected from studies of higher plantviruses on their interaction with the host werecontradictory and could not be interpreted in aconsistent fashion. This is because the plantconsists of different tissue systems, which aredifficult to isolate in tissue cultures and do notallow for homogenous infection. Cyanophagesoffer an advantage by providing a system thatresembles that of bacteriophages. Furthermore,because the photosynthesis of blue-green algaeis like that of higher plants, the alga-cyano-phage system can be adopted as a model forstudying plant photosynthesis under viral infec-tion.
Blue-green algae are widespread in theaquatic environment and often occur in massblooms related to eutrophication of waterbodies. The economic and nuisance effects ofblue-green algae on water quality and in caus-ing fish intoxications have been amply docu-mented (33, 122, 152). The probable influence ofcyanophages on the geographical and seasonaldistribution, as well as population dynamics, ofblue-green algae, and their possible use inbiological control of blue-green algae, added afurther impetus to investigations of the cyano-phages.
CHARACTERISTICS OF CYANOPHAGES
TypologyThe cyanophages isolated so far have been
named according to their known hosts. Accord-ingly, cyanophage groups were designated bythe initials of the generic names of the hosts, towhich arabic numerals have been added todesignate serological subgroups (111); differentisolates of similar types were marked by a suffixconsisting of the first letter of the provenance(26, 37, 111).
It is accepted that the criterion of hostspecificity is less valid for classification ofviruses than that of serological or morphologi-cal properties (82). The hazard in the hostspecificity criterion is obvious, because certainproperties of bacteriophage systems, such ashost range and types of host responses, are more
subject to mutational changes than others; suchfrequent mutational changes occur in the cya-nophage systems as well (44, 95, 131). Anadditional difficulty is the confusing state of thetaxonomy of most blue-green algae, whose clas-sification has been mainly determined throughmicroscopic examination of field collections andeven fixed specimens. Only recently, R. Y.Stanier and his co-workers proposed an inte-grated classification of unicellular blue-greenalgae based on morphological and physiologi-cal-biochemical properties of axenic cultures(142) and, in a like manner, they have begun toclassify the filamentous blue-green algae (59).In fact, it is conceivable that cyanophages couldaid in this classification of the blue-green algae(discussed below under "Host Range"). Accord-ingly, the presently-used nomenclature of cya-nophages may be misleading. For example, thecyanophage called "SM-1" (112) was namedafter Synechococcus elongatus and Microcystisaeruginosa (strain NRC-1); it now seems ques-tionable whether M. aeruginosa is a host sincethe strain NRC-1 is most probably a misclassi-fied Synechococcus (142). Thus, the presentcyanophage nomenclature should be regardedsimply as code names without taxonomic orevolutionary significance.
Serological specificities and morphologicalproperties (Fig. 1) of the cyanophages will beregarded here as the chief criteria for thegroupings of the four main typological groupsthat may be defined at present (Table 1).The LPP group is subdivided into two sero-
logically distinct subgroups, LPP-1 and LPP-2(111). This accords with the finding that onlysogeny of Plectonema boryanum with LPP-2SPI, the alga was found to be immune only tocyanophages of the LPP-2 group and not to anyof the LPP-1 strains (96). Because only onevirus is known for each of the SM, N, and ASgroups, they have been designated SM-1, N-1,and AS-1, respectively.A long-tailed virus A-1, active against Anaba-
ena variabilis, was observed by Kozjakov, Gro-mov, and Khudiakov of Leningrad University(64). A case has been reported of a cyanophageattacking Anabaenopsis raciborskii (130, 132),which may resemble phage trapping in bacterialspores (140, 149, 162). Reports of discoveries ofother cyanophages have been published (41-43,104, 130, 131, 150). These data were not suffi-cient to allow typological descriptions.
Because cyanophages are very similar to bac-teriophages both in structure and infectioncycle (see below), it is suggested that theyshould be considered a subgroup of the bacterio-phages.
344 BACTERIOL. REV.
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from
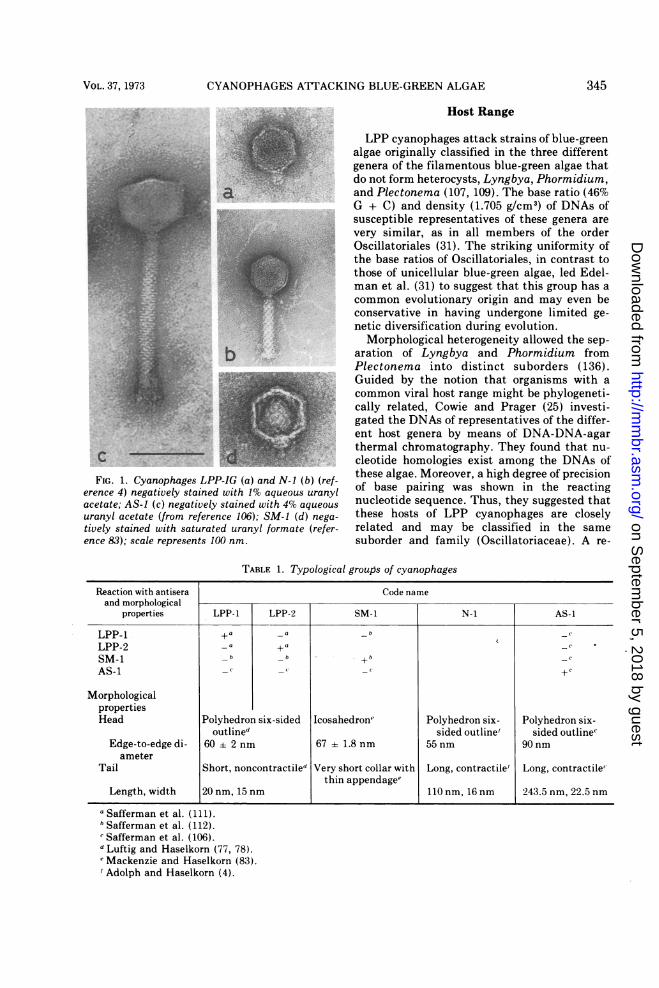
CYANOPHAGES ATTACKING BLUE-GREEN ALGAE
FIG. 1. Cyanophages LPP-IGerence 4) negatively stained witacetate; AS-1 (c) negatively staiuranyl acetate (from reference Jtively stained with saturated u
ence 83); scale represents 100 nr
Host Range
LPP cyanophages attack strains of blue-greenalgae originally classified in the three different
_* 0 genera of the filamentous blue-green algae thatdo not form heterocysts, Lyngbya, Phormidium,
and Plectonema (107, 109). The base ratio (46%G + C) and density (1.705 g/cm3) of DNAs ofsusceptible representatives of these genera are
very similar, as in all members of the orderOscillatoriales (31). The striking uniformity ofthe base ratios of Oscillatoriales, in contrast to
those of unicellular blue-green algae, led Edel-man et al. (31) to suggest that this group has a
common evolutionary origin and may even beconservative in having undergone limited ge-
netic diversification during evolution.-'; Morphological heterogeneity allowed the sep-
aration of Lyngbya and Phormidium fromPlectonema into distinct suborders (136).
Guided by the notion that organisms with a
common viral host range might be phylogeneti-cally related, Cowie and Prager (25) investi-
gated the DNAs of representatives of the differ-ent host genera by means of DNA-DNA-agarthermal chromatography. They found that nu-
cleotide homologies exist among the DNAs ofa() nd N-1 (b) (ref- these algae. Moreover, a high degree of precision1% aqueous uranyl of base pairing was shown in the reacting
urnylnucleotide sequence. Thus, they suggested thattned with 4% aqueous106); SM-1 (d) nega- these hosts of LPP cyanophages are closelyranyl formate (refer- related and may be classified in the samem. suborder and family (Oscillatoriaceae). A re-
TABLE 1. Typological groups of cyanophages
Reaction with antisera Code nameand morphological
properties LPP-1 LPP-2 SM-1 N-1 AS-1
LPP-1 + a _a _ C
LPP-2 _a +a
SM-1 _bb_ c
AS-i - c -C-
MorphologicalpropertiesHead Polyhedron six-sided Icosahedrone Polyhedron six- Polyhedron six-
outlined sided outline' sided outlinecEdge-to-edge di- 60 ± 2 nm 67 ± 1.8 nm 55 nm 90 nm
ameterTail Short, noncontractiled Very short collar with Long, contractile' Long, contractile"
thin appendage"Length, width 20 nm, 15 nm 110 nm, 16 nm 243.5 nm, 22.5 nm
a Safferman et al. (111).b Safferman et al. (112).c Safferman et al. (106).d Luftig and Haselkorn (77, 78).e'Mackenzie and Haselkorn (83).' Adolph and Haselkorn (4).
345VOL. 37, 1973
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

PADAN AND SHILO
classification of members of this family groupsthese algae into a single species, Schizotrixcalcicola (28). Moreover, the new classificationof the filamentous blue-green algae (59) basedon physiological and morphological criteria alsoplaces these genera close to one another.The blue-green alga designated Lyngbya sp.
strain 621 is neither sensitive to LPP- 1 nor doesits DNA show homology with DNAs of otherLPP-1 hosts (25). It is most probable that thisstrain might have been misclassified. Addi-tional isolates (named either Plectonema, Lyng-bya, or Phormidium) in different algal collec-tions were unaffected by LPP-1 (26, 107, 109).These algae might also have been misclassified.However, it is possible that they are differentvariants of the host (such as resistant mutantsand lysogenic strains) which are not susceptibleto the virus.Cyanophage SM-1 attacks S. elongatus (112),
a unicellular blue-green alga that may formshort filaments in different culture conditionsand growth stages. The DNA of this alga has 66to 67% G + C content (83) and a density of 1.73g/cm3 (142), which are the highest among theChroococcales. Similar to other unicellular or-ganisms, the Chroococcales exhibit a wide rangeof DNA base composition (35 to 70% G + C),demonstrating the heterogeneity of this groupand suggesting a polyphyletic origin and/orintensive genetic diversification during evolu-tion (31, 142).Cyanophage AS-1 attacks strains designated
Anacystis nidulans and Synechococcus ce-droum (106). Although classified within twodistinct genera, these algal species show markedmorphological, biochemical, and physiologicalsimilarity and were classified in one typologicalgroup (142). This accords with their susceptibil-ity to AS-1.Cyanophage N-1 attacks the filamentous
strain Nostoc muscorum (4) having typicaldifferentiated cells, the heterocysts, among thevegetative cells. Other isolates assigned to N.muscorum are insensitive to N-1, but againthese may represent either misclassification,different Nostoc species, or resistant variants.
It appears evident that the several cyano-phage groups isolated so far each have rathernarrow host ranges. Taking into account theexistence of nonsusceptible variants and host-range mutants of the viruses, this specificity ofthe cyanophages could aid in typing algalstrains and in reorganization of the blue-greenalgal systematics, in the same way as staphylo-coccus phages have been used for typing strainsof Staphylococcus aureus and other relatedspecies (157).
MorphologyCyanophages of the LPP group isolated so far
are virions of the head-tail type and are identi-cal in structure to one another (6, 26, 77-79, 95,111). A typical representative of the LPP group
is shown in Fig. la. The morphology of LPP-ihas been the most extensively studied, both on
intact particles and on separate head and tailpreparations. A model of LPP-1 suggested byLuftig and Haselkorn (78) is shown in Fig. 2.The viral head capsid is a polyhedron, appear-ing hexagonal in projection. The edge-to-edgedistance of the viral head is 58.6 2.1 nm (1).The detailed structure of the head has recentlybeen studied by Adolph and Haselkorn (6) whosuggested an icosahedral form and models forits construction from the capsomers. A short tail(20 nm long, 15 nm wide) is attached to one ofthe vertices of the head (78). Transverse andside views of isolated tails led to the suggestionthat the tail is a hollow cylinder with sixfoldradial symmetry made up of several (two tofour) rings of six subunits. However, the orien-tation of the rings with respect to each otheralong the tail is still unknown. If the rules ofsymmetry apply, as in bacteriophage structure(16), then the observed sixfold radial symmetryof the tail requires similar symmetry in the head
6151
IOne Linsor DNA DupblexXof Length 13 a,g801~~~~~~~100 A
Tail Capital 1
TaI(Sheath)_
Tail Cavity 200- 225x
~-150A--
250A--^
End Viewof TO I I
FIG. 2. Model of LPP-1 proposed by Luftig andHaselkorn (78).
346 BACTERIOL. REV.
Coat)0A
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from
CYANOPHAGES ATTACKING BLUE-GREEN ALGAE
or, as suggested by Moody (86) with reference tothe T-even phages, an intermediate structureadapting the unlike head and tail symmetries.At present it is impossible to choose betweenthese alternatives. Another unsolved problem isthe role of the tail capital that protrudes intothe head capsid at the head-tail junction (78).This may be either a simple mechanical joint tofix the tail to the head and/or the asymmetryadaptor suggested above.Specimens of either LPP-1 or LPP-1G ob-
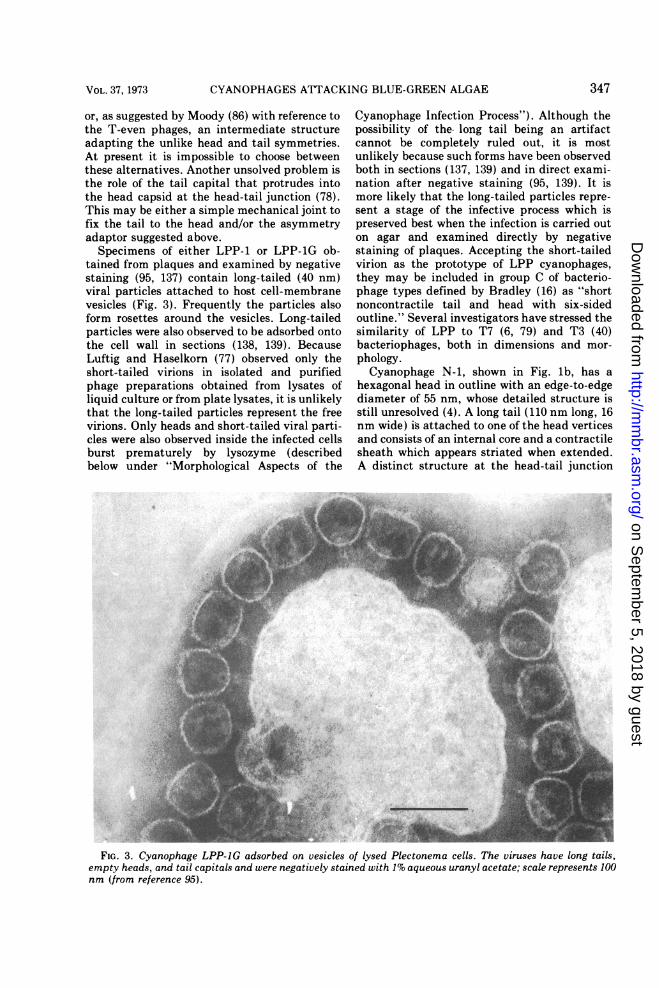
tained from plaques and examined by negativestaining (95, 137) contain long-tailed (40 nm)viral particles attached to host cell-membranevesicles (Fig. 3). Frequently the particles alsoform rosettes around the vesicles. Long-tailedparticles were also observed to be adsorbed ontothe cell wall in sections (138, 139). BecauseLuftig and Haselkorn (77) observed only theshort-tailed virions in isolated and purifiedphage preparations obtained from lysates ofliquid culture or from plate lysates, it is unlikelythat the long-tailed particles represent the freevirions. Only heads and short-tailed viral parti-cles were also observed inside the infected cellsburst prematurely by lysozyme (describedbelow under "Morphological Aspects of the
Cyanophage Infection Process"). Although thepossibility of the. long tail being an artifactcannot be completely ruled out, it is mostunlikely because such forms have been observedboth in sections (137, 139) and in direct exami-nation after negative staining (95, 139). It ismore likely that the long-tailed particles repre-sent a stage of the infective process which ispreserved best when the infection is carried outon agar and examined directly by negativestaining of plaques. Accepting the short-tailedvirion as the prototype of LPP cyanophages,they may be included in group C of bacterio-phage types defined by Bradley (16) as "shortnoncontractile tail and head with six-sidedoutline." Several investigators have stressed thesimilarity of LPP to T7 (6, 79) and T3 (40)bacteriophages, both in dimensions and mor-phology.Cyanophage N-1, shown in Fig. lb, has a
hexagonal head in outline with an edge-to-edgediameter of 55 nm, whose detailed structure isstill unresolved (4). A long tail (110 nm long, 16nm wide) is attached to one of the head verticesand consists of an internal core and a contractilesheath which appears striated when extended.A distinct structure at the head-tail junction
FIG. 3. Cyanophage LPP-JG adsorbed on vesicles of lysed Plectonema cells. The viruses have long tails,empty heads, and tail capitals and were negatively stained with 1% aqueous uranyl acetate; scale represents 100nm (from reference 95).
347VOL. 37, 1973
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from
PADAN AND SHILO
(head capital) was also observed. The structureof N-1 is very similar to the basic morphologicalbacteriophage type A designated by Bradley(16) as "contractile tail with six-sided outlinedhead."The largest cyanophage yet observed is AS-1,
shown in Fig. ic. The head is hexagonal inoutline with an edge-to-edge diameter of 90 nm.A long tail (243.5 nm long, 22.5 nm wide),attached to one of the head vertices, consists ofa base plate to which short pins are attached, acontractile sheath, and an internal core (106).Cyanophage SM-1 (Fig. id) has an icosahe-
dral head capsid with an edge-to-edge diameterof 67 + 1.8 nm. A very short collar is attached tothe head, from which a thin appendage, whichcould be a tail, protrudes (83). Particles dam-aged by phosphotungstic acid appear to have athin inner membrane, in addition to the princi-pal capsid shell (83). SM-1 particles attachedto the cell wall appear to have longer tails thanthose of the free phage (84), similar to LPP-1Gdescribed above.
CompositionNucleic acids. All nucleic acids of cyano-
phages analyzed so far are linear double-stranded DNAs. The physicochemical charac-teristics of the DNAs of cyanophages and theirhosts are summarized in Table 2.Because there is a good agreement in the % G
+ C content values calculated from buoyantdensity and melting points of DNA in LPP-1(77), N-1 (4), and SM-1 (83), these virusesprobably contain no odd bases as major constit-uents. Resolution of DNAs of host and virusduring infection has already been achieved for
the LPP-1-Plectonema system using the equi-librium density gradients (118). The differencein buoyant density values of N-1 and Nostoc (cf.Table 2) suggests that equilibrium density gra-
dients can also be employed for resolution oftheir DNAs during infection. The very highdensity of Synechococcus DNA, similar to thatof SM-1 DNA, suggests that this is not possiblein this case (83).The presence of double-stranded DNA in
LPP-1, N-1, AS-1, and SM-1 conforms withBradley's (16) prediction that head-tail bacteri-ophages contain only this type of nucleic acid.Thermally denatured DNA of LPP-1 exhibits
a single band in CsCl at a density 0.012 g/mlhigher than the native DNA. Unfortunately no
separation of the strands could be achieved bymixing the denatured DNA with poly IG (77).This behavior contrasts with T7 or A DNA forwhich poly IG selectively shifts one of thestrands to a higher density.
In a search for common evolutionary patternsamong cyanophages and their hosts, Cowie andPrager (25) found no homology between DNAsof LPP-1 and its hosts (when using DNA-DNA-agar thermal chromatography). A similar con-
clusion was reached indirectly by Luftig andHaselkorn (77). It would be most interesting tostudy homology in the lysogenic blue-greenalgae systems (21, 96). No homology was evi-dent when the DNA of LPP-1 was reacted withthe DNAs of A, 434 hybrid, 080, P22 (25), andT7 (79).
Proteins. The major proteins of LPP-1 were
characterized by sodium dodecyl sulfate-gelelectrophoresis of bulk viral proteins and sepa-rate head-tail preparations (78, 119). The major
TABLE 2. Physicochemical characteristics of the DNA of the cyanophages and typical hosts
Physicochemical Plecto- Nostoc Synecho- Synecho-charactcherlcal LPP-1 N-Pa SM-1h AS-i nema muscor coccus coccuscharacter boryanum muscorum elongatus cedrorum
Molecular weight 27 x 106ld 38 x 106 56-62 x 106 2.109tContour length (nm) 13.2e 17.2 24.3Buoyant density in CsCl 1.714 at 1.696 at 1.725 1.706 at 1.702 at 1.73 at 1.71k
(g/cm3) 25 Cd e 20 C 25 C' 25 Ca 25 C9Sedimentation coefficient 33.4" 38.7 48.0
(S2a, -.)G + C content (%) 53"' X 37 66-67 53-54 48h. i 43h 71.4k 55.69
a All results from Adolph and Haselkorn (4).All results from Mackenzie and Haselkorn (83).Safferman et al. (106).
d Goldstein and Bendet (39).e Luftig and Haselkorn (77).' Sherman and Haselkorn (118).9 Stanier et al. (142).h Edelman et al. (31).i Kaye et al. (56).
348 BACTERIOL. REV.
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from
CYANOPHAGES ATTACKING BLUE-GREEN ALGAE
head proteins are the 39,000 and 13,000 molecu-lar weight species. The major tail protein has a
molecular weight of 80,000. No internal proteinswere detected. The sum of molecular weightsaccounts for about 35% of the coding capacity ofLPP-1 DNA.The major structural proteins of N-1, simi-
larly characterized, are the 37,000 and 14,000molecular weight species, whose sum accountsfor 34% of the coding capacity of the DNA (4).Mature SM-1 virions contain a dozen pro-
teins, of which two (40,000 and 25,000 molecularweight) must be the major components of thecapsid shell and contain over 80% of the sulfuratoms. The sum of the molecular weights ofSM-1 structural proteins requires a coding ca-pacity of 16% of the entire chromosome, a
considerably smaller fraction than is found inviruses such as LPP-1 and T7 (83).
Physicochemical CharacteristicsTable 3 summarizes physicochemical param-
eters of certain cyanophages. LPP-1 and, possi-bly, N-1 require Mg2+ for their stability. Luftigand Haselkorn (78) showed that LPP-1 tailsseparate from the heads after ethylenediamine-tetraacetate DNAse treatment which burststhe head and releases DNA. Goldstein et al.(40) related this to the relatively high DNA con-
tent of these cyanophages. In agents such as
T3 and T7, the Mg2+ requirement was relatedto diffusion of the polyamines out of the headduring purification and their replacement bythe cations, which neutralize phosphate groupsof the viral DNA to maintain structural in-tegrity (7).Compared with bacterial viruses, which are
generally stable from pH 5 to 8 (3), both LPP-1
(108) and SM-1 (112) are remarkably stable inthe alkaline range (pH 7 to 11). This accordswith the pH range of the blue-green algal host(see below, "ECOLOGICAL CONSIDERA-TIONS"). AS-1 (106) shows great stability athigh pHs (up to pH 10), but can also toleratepHs as low as 4. It would be interesting todetermine the pH range of N-1 and the optimalpHs for the cyanophage growth cycles.
GROWTH CYCLE OF CYANOPHAGES
Phage Development in FilamentousBlue-green Algae
Elucidation of the growth cycle of some
cyanophages is complicated by the fact thatmany of the algal hosts are filamentous. Mosttechniques and statistical analyses used in thestudies of phage infection systems in unicellularhosts cannot be strictly applied in the case ofmulticellular hosts. Furthermore, filamentousorganization may influence the pattern of viralreproduction. Cyanophage growth might bemodified both by the number and character ofcells in the trichome (for instance, in heterocystor spore-containing filaments). The indicationsthat plasmadesmata link algal cells (71) andthat substrates are transferred from cell to cellin trichomes (158) seem to support this assump-tion. Viruses in higher plants have been indi-cated to spread from cell to cell via plasmade-smata (61, 72). The peculiar slow and nonlinearadsorption kinetics of LPP-1 on Plectonemafound by Goldstein et al. (40) was relatedmainly to the variability in the host filamentlength.To compare viral growth in filaments of
different lengths, a short trichome (1 to 6 cells)
TABLE 3. Physicochemical characteristics of cyanophage particles
Cyanophage groupPhysicochemical parameter
LPP-1 N-1 SM-1 AS-la
Sedimentation coefficient (s20, ) 555-548b' 539d 1.021, 1.029eBuoyant density in CsCl (g/cm3) 1.48f 1.489Mg2+ requirement (M) 0.001b, h. i Not requirede Not requiredTemperature of inactivation (C) 55i 55e 60Temperature range of greatest stability 4-40i 4-40epH range of greatest stability 5-11, 5-11e 4-10
a All results from Safferman et al. (106).Goldstein et al. (40).Schneider et al. (113).
d Adolph and Haselkorn (4).e Safferman et al. (112).' Luftig and Haselkorn (79).g Mackenzie and Haselkorn (83).Luftig and Haselkorn (77).Safferman and Morris (108).
349VOL. 37, 1973
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from
PADAN AND SHILO
mutant of P. boryanum was selected from awild-type population consisting of trichomes of50 cells or more (94). The growth rate of themutant is identical to that of the wild type, andLPP-1G grows on both strains with equal effi-ciency.The adsorption kinetics of LPP-1G on the
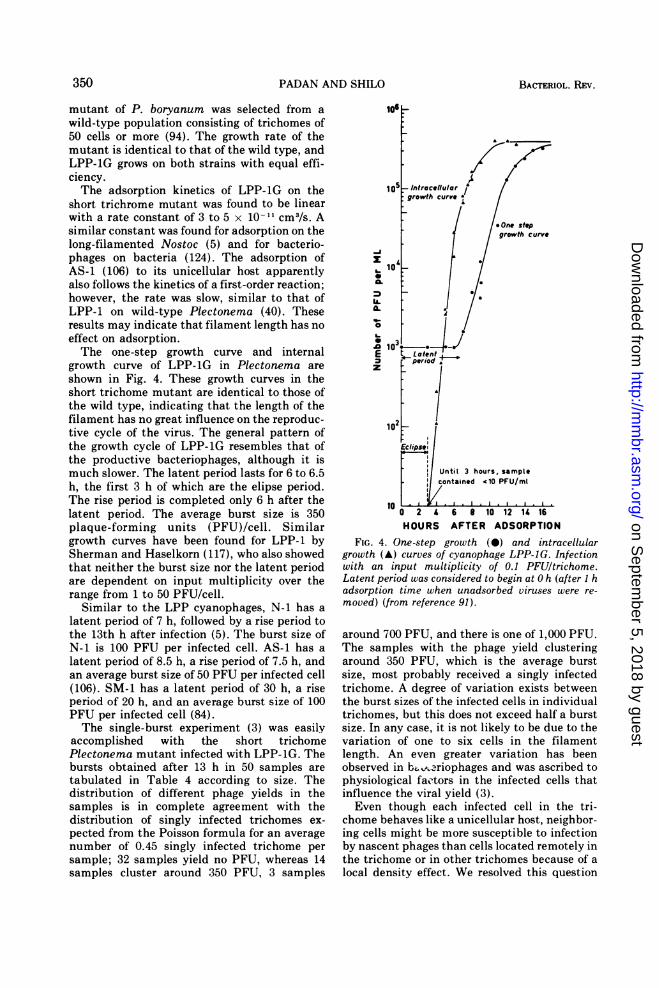
short trichrome mutant was found to be linearwith a rate constant of 3 to 5 x 10-11 cm3/s. Asimilar constant was found for adsorption on thelong-filamented Nostoc (5) and for bacterio-phages on bacteria (124). The adsorption ofAS-1 (106) to its unicellular host apparentlyalso follows the kinetics of a first-order reaction;however, the rate was slow, similar to that ofLPP-1 on wild-type Plectonema (40). Theseresults may indicate that filament length has noeffect on adsorption.The one-step growth curve and internal
growth curve of LPP-1G in Plectonema areshown in Fig. 4. These growth curves in theshort trichome mutant are identical to those ofthe wild type, indicating that the length of thefilament has no great influence on the reproduc-tive cycle of the virus. The general pattern ofthe growth cycle of LPP-1G resembles that ofthe productive bacteriophages, although it ismuch slower. The latent period lasts for 6 to 6.5h, the first 3 h of which are the elipse period.The rise period is completed only 6 h after thelatent period. The average burst size is 350plaque-forming units (PFU)/cell. Similargrowth curves have been found for LPP-1 bySherman and Haselkorn (117), who also showedthat neither the burst size nor the latent periodare dependent on input multiplicity over therange from 1 to 50 PFU/cell.
Similar to the LPP cyanophages, N-1 has alatent period of 7 h, followed by a rise period tothe 13th h after infection (5). The burst size ofN-1 is 100 PFU per infected cell. AS-1 has alatent period of 8.5 h, a rise period of 7.5 h, andan average burst size of 50 PFU per infected cell(106). SM-1 has a latent period of 30 h, a riseperiod of 20 h, and an average burst size of 100PFU per infected cell (84).The single-burst experiment (3) was easily
accomplished with the short trichomePlectonema mutant infected with LPP-1G. Thebursts obtained after 13 h in 50 samples aretabulated in Table 4 according to size. Thedistribution of different phage yields in thesamples is in complete agreement with thedistribution of singly infected trichomes ex-pected from the Poisson formula for an averagenumber of 0.45 singly infected trichome persample; 32 samples yield no PFU, whereas 14samples cluster around 350 PFU, 3 samples
~1
E Fa. S
Z
~. Laentd
Eclipse:Sl
Until 3 hours, samplecontained '10 PFU/ml
11/
O 2 4 6 8 10 12 14 16HOURS AFTER ADSORPTION
FIG. 4. One-step growth (*) and intracellulargrowth (A) curves of cyanophage LPP-JG. Infectionwith an input multiplicity of 0.1 PFUltrichome.Latent period was considered to begin at 0 h (after I hadsorption time when unadsorbed viruses were re-moved) (from reference 91).
around 700 PFU, and there is one of 1,000 PFU.The samples with the phage yield clusteringaround 350 PFU, which is the average burstsize, most probably received a singly infectedtrichome. A degree of variation exists betweenthe burst sizes of the infected cells in individualtrichomes, but this does not exceed half a burstsize. In any case, it is not likely to be due to thevariation of one to six cells in the filamentlength. An even greater variation has beenobserved in bz. -2riophages and was ascribed tophysiological factors in the infected cells thatinfluence the viral yield (3).Even though each infected cell in the tri-
chome behaves like a unicellular host, neighbor-ing cells might be more susceptible to infectionby nascent phages than cells located remotely inthe trichome or in other trichomes because of alocal density effect. We resolved this question
350 BACTERIOL. REV.
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from
CYANOPHAGES ATTACKING BLUE-GREEN ALGAE
by incubating the highly diluted single-burstexperimental systems for an additional growthcycle period of 13 h. The number of samplesshowing a second-cycle yield of 700 to 1,200PFUs was within the range expected for singlyinfected trichomes. This is what might beexpected from infection of neighboring cells atthe moment of the original burst. Thus thejuxtaposition of the algal cells in the trichomeand not the plasmodesmata expedites thespread of a single infection along the wholelength of the filament.
Sequence of Synthesis of CyanophageMacromolecules
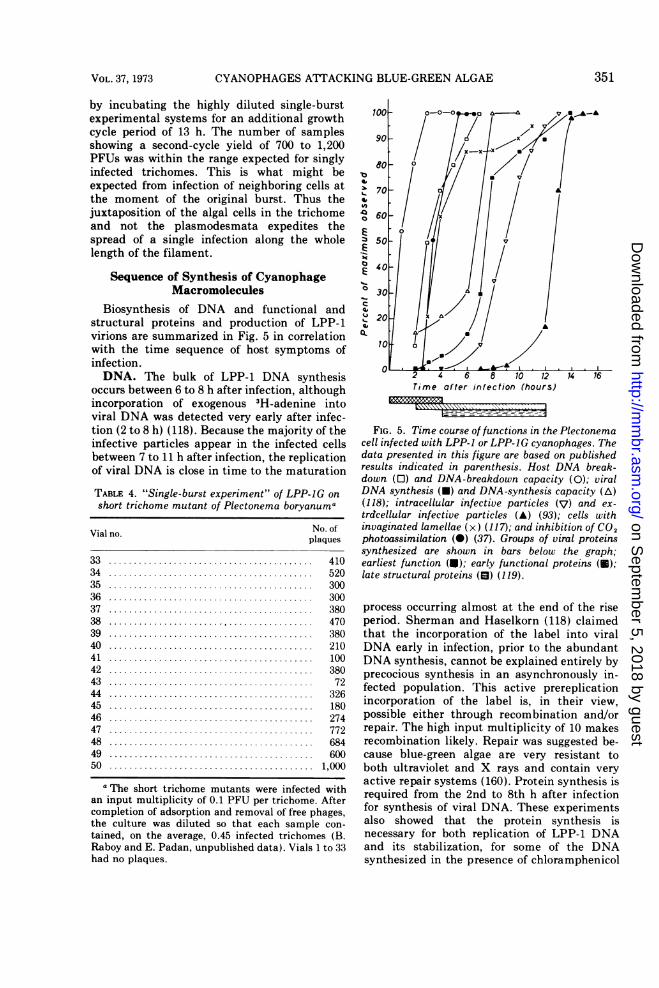
Biosynthesis of DNA and functional andstructural proteins and production of LPP-1virions are summarized in Fig. 5 in correlationwith the time sequence of host symptoms ofinfection.DNA. The bulk of LPP-1 DNA synthesis
occurs between 6 to 8 h after infection, althoughincorporation of exogenous 3H-adenine intoviral DNA was detected very early after infec-tion (2 to 8 h) (118). Because the majority of theinfective particles appear in the infected cellsbetween 7 to 11 h after infection, the replicationof viral DNA is close in time to the maturation
TABLE 4. "Single-burst experiment" of LPP-JG on
short trichome mutant of Plectonema boryanuma
No. ofplaques
333435363738394041424344454647484950
41052030030038047038021010038072
326180274772684600
1,000
a The short trichome mutants were infected withan input multiplicity of 0.1 PFU per trichome. Aftercompletion of adsorption and removal of free phages,the culture was diluted so that each sample con-
tained, on the average, 0.45 infected trichomes (B.Raboy and E. Padan, unpublished data). Vials 1 to 33had no plaques.
4,460 U/!/(70 A
a4~ 60
E /0Z3 50- 0)3
Em
40//-030-20
10
2 4 6 8 10 12 14 16Time after infection (hours)
FIG. 5. Time course of functions in the Plectonemacell infected with LPP-1 or LPP-JG cyanophages. Thedata presented in this figure are based on publishedresults indicated in parenthesis. Host DNA break-down (0) and DNA-breakdown capacity (0); viralDNA synthesis (U) and DNA-synthesis capacity (A)(118); intracellular infective particles (V) and ex-trdcellular infective particles (A) (93); cells withinvaginated lamellae (x) (117); and inhibition of CO2photoassimilation (0) (37). Groups of viral proteinssynthesized are shown in bars below the graph;earliest function (U); early functional proteins (U);late structural proteins (El) (119).
process occurring almost at the end of the riseperiod. Sherman and Haselkorn (118) claimedthat the incorporation of the label into viralDNA early in infection, prior to the abundantDNA synthesis, cannot be explained entirely byprecocious synthesis in an asynchronously in-fected population. This active prereplicationincorporation of the label is, in their view,possible either through recombination and/orrepair. The high input multiplicity of 10 makesrecombination likely. Repair was suggested be-cause blue-green algae are very resistant toboth ultraviolet and X rays and contain veryactive repair systems (160). Protein synthesis isrequired from the 2nd to 8th h after infectionfor synthesis of viral DNA. These experimentsalso showed that the protein synthesis isnecessary for both replication of LPP-1 DNAand its stabilization, for some of the DNAsynthesized in the presence of chloramphenicol
351VOL. 37, 1973
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

PADAN AND SHILO
is labile. This probably reflects the fate of un-
packaged viral DNA (118).Assuming that mitomycin C inhibits DNA
replication in blue-green algae as in bacteria, A.Rimon in our laboratory tested its effect on theLPP-1G infection cycle. Because the inhibitor islight sensitive, the infective process was carriedout in the dark with Plectonema cells prein-cubated in the light to enhance viral synthesis(see "Photosynthesis and respiration" below). Itwas found that mitomycin C (2 ,g/ml) inhibitscyanophage development when administered atany time during the growth cycle (103). Hence,DNA synthesis is required throughout the cya-
nophage growth cycle for full yield. This accordswith the findings that LPP-1 DNA synthesiscontinues up to the end of the cycle (118).
Protein synthesis. Sherman and Haselkorn(119) characterized the virus-induced proteinsand followed the course of their synthesis duringLPP-1 infection. Three classes of viral proteinswere distinguished (cf. Fig. 5). Synthesis of theearliest class (three proteins) begins soon afterinfection and is shut off by the 4th h. The earlyclass (another three proteins) begins to formaround the 2nd h and continues forming untillysis, but is not found in the mature particles.Finally, the late class includes viral structuralproteins whose synthesis begins at about the 4thh and continues until lysis. It is significant thatthe synthesis of structural proteins begins 2 hbefore the bulk synthesis of viral DNA. A totalof 17 distinct, viral-induced proteins were de-tected after infection. They account for about65% of the coding capacity of the viral genome.Sherman and Haselkorn found no evidence forthe synthesis of large precursors of any struc-tural proteins during LPP-1 infection, such as
noted for T4 (70) and poliovirus (146). Thepattern of viral protein synthesis in the LPP-1-Plectonema system is very similar to that of theT7-Escherichia coli system (119).
At any time during infection, chlorampheni-col (25 gig/ml) and erythromycin (5 ,ug/ml)inhibit the formation of new, infective viralparticles (103). Hence, protein synthesis is re-
quired throughout the infective process, cor-
roborating the conclusion of Sherman and Ha-selkorn (119).Assuming that actinomycin D and rifampin
inhibit DNA-dependent ribonucleic acid (RNA)synthesis, as they do in bacteria, they were usedto determine the sequence of RNA synthesis inLPP-1G infection. As these inhibitors are sensi-tive to light, the dark reaction system was
employed (see below under "Photosynthesisand respiration"). Rifampin (50 jig/ml) andactinomycin D (30 gg/ml) inhibit cyanophage
development until the end of the infectioncycle, implying that mRNA synthesis requiredfor cyanophage development continues up tothe end of the infection process.
It is apparent that the pattern of the infectioncycle of the cyanophage is very similar to that ofbacteriophages. However, it is much slower,lasting 13 h at 26 C in LPP-1G or 50 h in SM-1,whereas most bacteriophages complete theirreproduction in tens of minutes. This may berelated to the slower generation time of blue-green algae as compared with most bacteria.
Morphological Aspects of the CyanophageInfective Process
The cell in which the cyanophage develops isa highly organized prokaryont (71). In manyblue-green algae, such as Plectonema, the pho-tosynthetic lamellae are organized in sheaths atthe periphery and compartmentalize the cell toa certain degree (117, 137). The DNA is locatedin the central cytoplasm (nucleoplasm) wherepolyphosphate granules and the enigmatic po-lyhedral bodies are also found. Additional inclu-sion bodies, such as a- or polyglucoside granules(22), fl-granules which are probably lipids (71),and peptide-containing cyanophycin granules(127), are seen mainly between or outside thelamellae (156). Thus, the topology of the cyano-phage development cycle is no less interestingthan its morphological aspects.The cyanophage LPP-1 adsorbs to the algal
cell wall by the distal end of the tail (138). Theexact depth to which the tail penetrates has notbeen determined, although Smith et al. (138,139) found that it appears to penetrate the cellwall. The cyanophage injects its DNA into thehost cell, most probably through its tail assuggested for bacteriophages, and empty ghostsadhere to the host cell. It must be noted,however, that the classical experiment of Her-shey and Chase (47) to demonstrate DNAinjection has not yet been conducted in thissystem. The existence of a resistant Plectonemamutant (95) that does not adsorb the cyano-phage LPP-1G indicates that specific receptorsites exist on the surface of susceptible hostcells, such as those shown in bacteria (154).By studying sections of Plectonema
boryanum cells infected with LPP-1, Smith andhis co-workers (19, 137) observed that the firstsign of infection is invagination of the photosyn-thetic lamellae. The space between the foldedlamellae and the plasma membrane in whichviral particles later appear was called the viro-genic stroma. Similar symptoms appear inother hosts of LPP-1 (139). Sherman and Ha-
352 BACTERIOL. REV.
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from
CYANOPHAGES ATTACKING BLUE-GREEN ALGAE
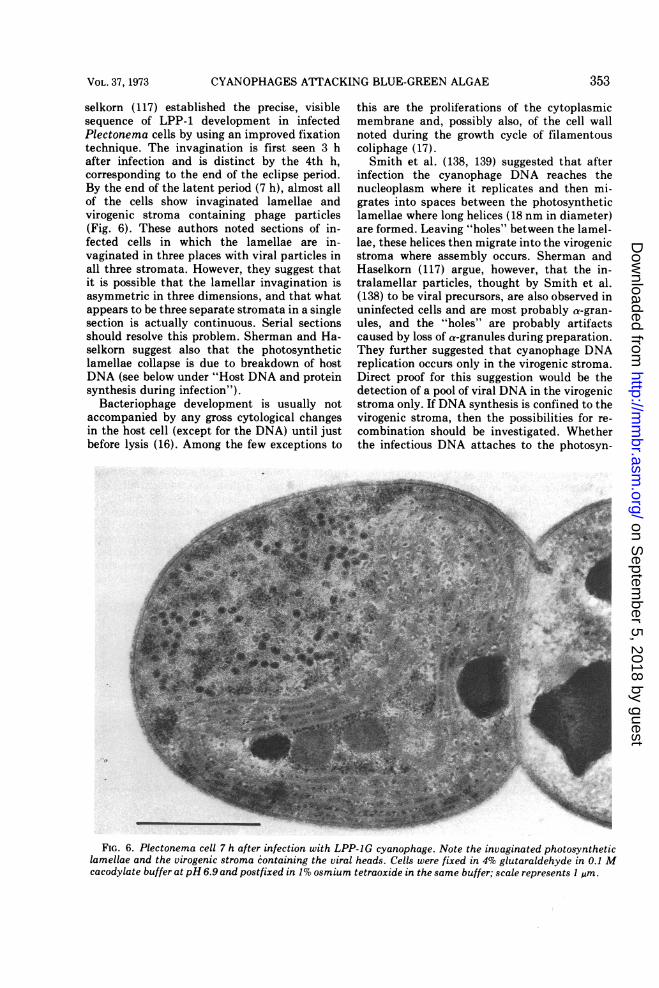
selkorn (117) established the precise, visiblesequence of LPP-1 development in infectedPlectonema cells by using an improved fixationtechnique. The invagination is first seen 3 hafter infection and is distinct by the 4th h,corresponding to the end of the eclipse period.By the end of the latent period (7 h), almost allof the cells show invaginated lamellae andvirogenic stroma containing phage particles(Fig. 6). These authors noted sections of in-fected cells in which the lamellae are in-vaginated in three places with viral particles inall three stromata. However, they suggest thatit is possible that the lamellar invagination isasymmetric in three dimensions, and that whatappears to be three separate stromata in a singlesection is actually continuous. Serial sectionsshould resolve this problem. Sherman and Ha-selkorn suggest also that the photosyntheticlamellae collapse is due to breakdown of hostDNA (see below under "Host DNA and proteinsynthesis during infection").
Bacteriophage development is usually notaccompanied by any gross cytological changesin the host cell (except for the DNA) until justbefore lysis (16). Among the few exceptions to
this are the proliferations of the cytoplasmicmembrane and, possibly also, of the cell wallnoted during the growth cycle of filamentouscoliphage (17).Smith et al. (138, 139) suggested that after
infection the cyanophage DNA reaches thenucleoplasm where it replicates and then mi-grates into spaces between the photosyntheticlamellae where long helices (18 nm in diameter)are formed. Leaving "holes" between the lamel-lae, these helices then migrate into the virogenicstroma where assembly occurs. Sherman andHaselkorn (117) argue, however, that the in-tralamellar particles, thought by Smith et al.(138) to be viral precursors, are also observed inuninfected cells and are most probably a-gran-ules, and the "holes" are probably artifactscaused by loss of a-granules during preparation.They further suggested that cyanophage DNAreplication occurs only in the virogenic stroma.Direct proof for this suggestion would be thedetection of a pool of viral DNA in the virogenicstroma only. If DNA synthesis is confined to thevirogenic stroma, then the possibilities for re-combination should be investigated. Whetherthe infectious DNA attaches to the photosyn-
- p
FIG. 6. Plectonema cell 7 h after infection with LPP-1G cyanophage. Note the invaginated photosyntheticlamellae and the virogenic stroma containing the viral heads. Cells were fixed in 4% glutaraldehyde in 0.1 Mcacodylate buffer at pH 6.9 and postfixed in 1% osmium tetraoxide in the same buffer; scale represents I g.sm.
353VOL. 37, 1973
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from
PADAN AND SHILO
thetic lamellae and/or to the cell membrane alsoremains to be investigated.
In preliminary work in our laboratory (35) onthe localization of protein synthesis and viralassembly, attempts were made to locate theviral antigens in sections of infected cells byusing ferritin-labeled antibodies preparedagainst the purified, whole-virus particle. Thesephage antigens appear to be localized only inthe virogenic stroma similar to the viral heads(117). Therefore, cyanophage particles seem toassemble only in a virogenic stroma separatedfrom the nucleoplasm by the photosyntheticlamellae.The time of appearance of the viral heads, as
seen by electron microscope (35, 117), is similarto the rate of appearance of infective viralparticles in infected cells (cf. Fig. 5). Until 4 hafter infection (end of eclipse period), viralheads were very rarely seen in the virogenicstroma. By the end of the latent period (7 h)many cells contain viral heads; by 10 h viralassembly appears to have begun in almost everycell.Sherman and Haselkorn (117) noted viral
heads with various configurations of DNA to-gether with heads which were empty and inter-preted this inhomogeneity as a glutaraldehyde-fixation artifact. Similar observations in T7 ledZagury et al. (163) to suggest inhomogenouspacking of the DNA. Such particles were de-tected in bacteria infected with other phages(126).
Further investigations of the morphologicalaspect of cyanophage development were under-taken in our laboratory (35). For this purpose,
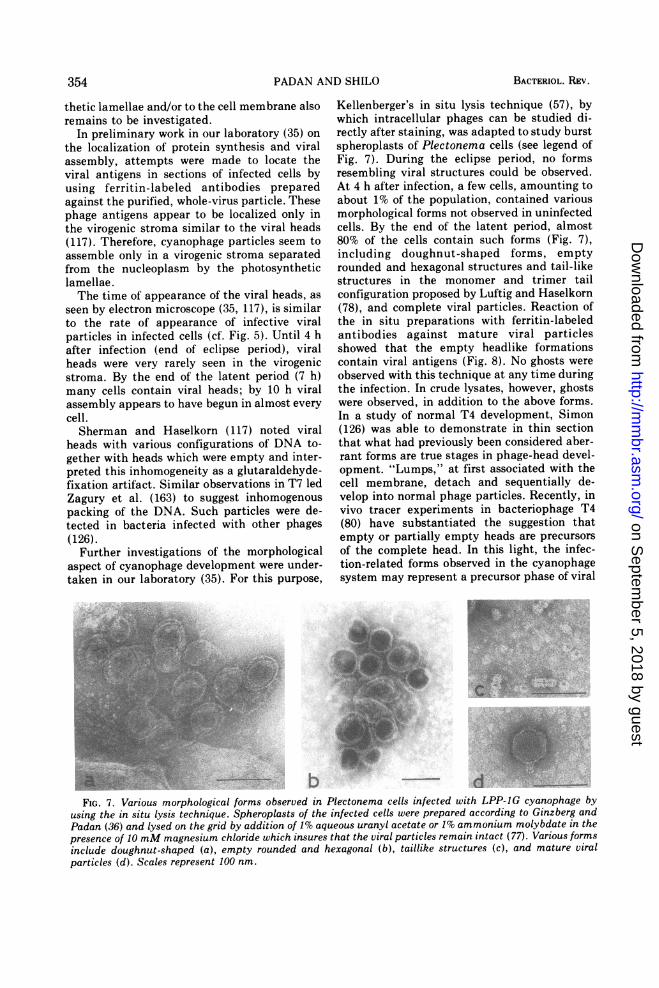
Kellenberger's in situ lysis technique (57), bywhich intracellular phages can be studied di-rectly after staining, was adapted to study burstspheroplasts of Plectonema cells (see legend ofFig. 7). During the eclipse period, no formsresembling viral structures could be observed.At 4 h after infection, a few cells, amounting toabout 1% of the population, contained variousmorphological forms not observed in uninfectedcells. By the end of the latent period, almost80% of the cells contain such forms (Fig. 7),including doughnut-shaped forms, emptyrounded and hexagonal structures and tail-likestructures in the monomer and trimer tailconfiguration proposed by Luftig and Haselkorn(78), and complete viral particles. Reaction ofthe in situ preparations with ferritin-labeledantibodies against mature viral particlesshowed that the empty headlike formationscontain viral antigens (Fig. 8). No ghosts wereobserved with this technique at any time duringthe infection. In crude lysates, however, ghostswere observed, in addition to the above forms.In a study of normal T4 development, Simon(126) was able to demonstrate in thin sectionthat what had previously been considered aber-rant forms are true stages in phage-head devel-opment. "Lumps," at first associated with thecell membrane, detach and sequentially de-velop into normal phage particles. Recently, invivo tracer experiments in bacteriophage T4(80) have substantiated the suggestion thatempty or partially empty heads are precursorsof the complete head. In this light, the infec-tion-related forms observed in the cyanophagesystem may represent a precursor phase of viral
FIG. 7. Various morphological forms observed in Plectonema cells infected with LPP-IG cyanophage byusing the in situ lysis technique. Spheroplasts of the infected cells were prepared according to Ginzberg andPadan (36) and lysed on the grid by addition of 1% aqueous uranyl acetate or 1% ammonium molybdate in thepresence of 10 mM magnesium chloride which insures that the viral particles remain intact (77). Various formsinclude doughnut-shaped (a), empty rounded and hexagonal (b), taillike structures (c), and mature viralparticles (d). Scales represent 100 nm.
354 BACTERIOL. REV.
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from
CYANOPHAGES ATTACKING BLUE-GREEN ALGAE
W-4-1*P~~ ~ ~ ~ ~ - 11.l He
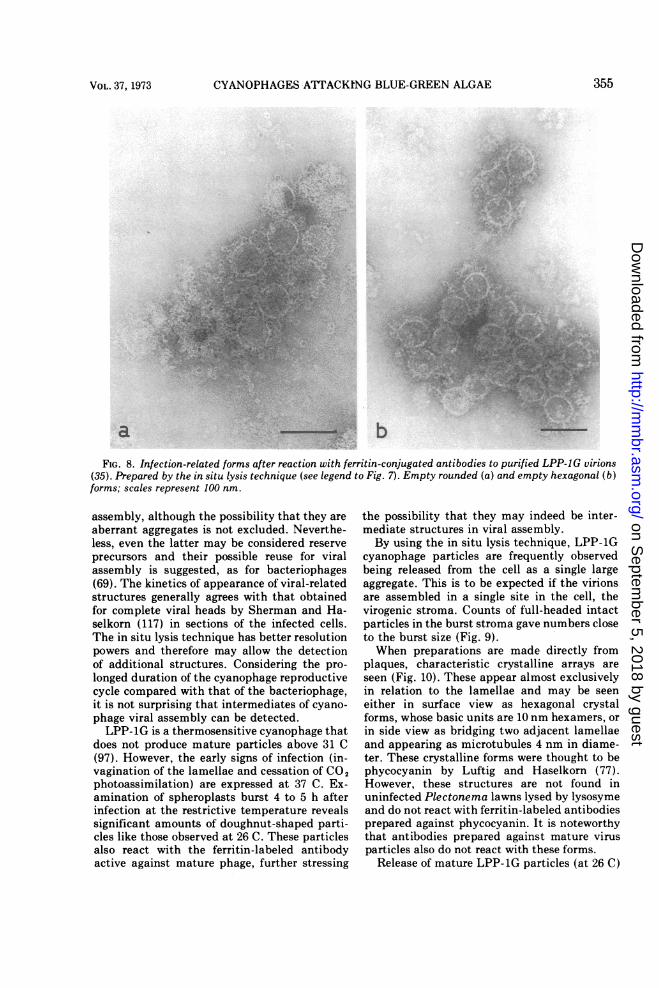
FIG. 8. Infection-related forms after reaction with ferritin-conjugated antibodies to purified LPP-1G virions(35). Prepared by the in situ lysis technique (see legend to Fig. 7). Empty rounded (a) and empty hexagonal (b)forms; scales represent 100 nm.
assembly, although the possibility that they areaberrant aggregates is not excluded. Neverthe-less, even the latter may be considered reserveprecursors and their possible reuse for viralassembly is suggested, as for bacteriophages(69). The kinetics of appearance of viral-relatedstructures generally agrees with that obtainedfor complete viral heads by Sherman and Ha-selkorn (117) in sections of the infected cells.The in situ lysis technique has better resolutionpowers and therefore may allow the detectionof additional structures. Considering the pro-longed duration of the cyanophage reproductivecycle compared with that of the bacteriophage,it is not surprising that intermediates of cyano-phage viral assembly can be detected.LPP-1G is a thermosensitive cyanophage that
does not produce mature particles above 31 C(97). However, the early signs of infection (in-vagination of the lamellae and cessation of CO2photoassimilation) are expressed at 37 C. Ex-amination of spheroplasts burst 4 to 5 h afterinfection at the restrictive temperature revealssignificant amounts of doughnut-shaped parti-cles like those observed at 26 C. These particlesalso react with the ferritin-labeled antibodyactive against mature phage, further stressing
the possibility that they may indeed be inter-mediate structures in viral assembly.By using the in situ lysis technique, LPP-1G
cyanophage particles are frequently observedbeing released from the cell as a single largeaggregate. This is to be expected if the virionsare assembled in a single site in the cell, thevirogenic stroma. Counts of full-headed intactparticles in the burst stroma gave numbers closeto the burst size (Fig. 9).When preparations are made directly from
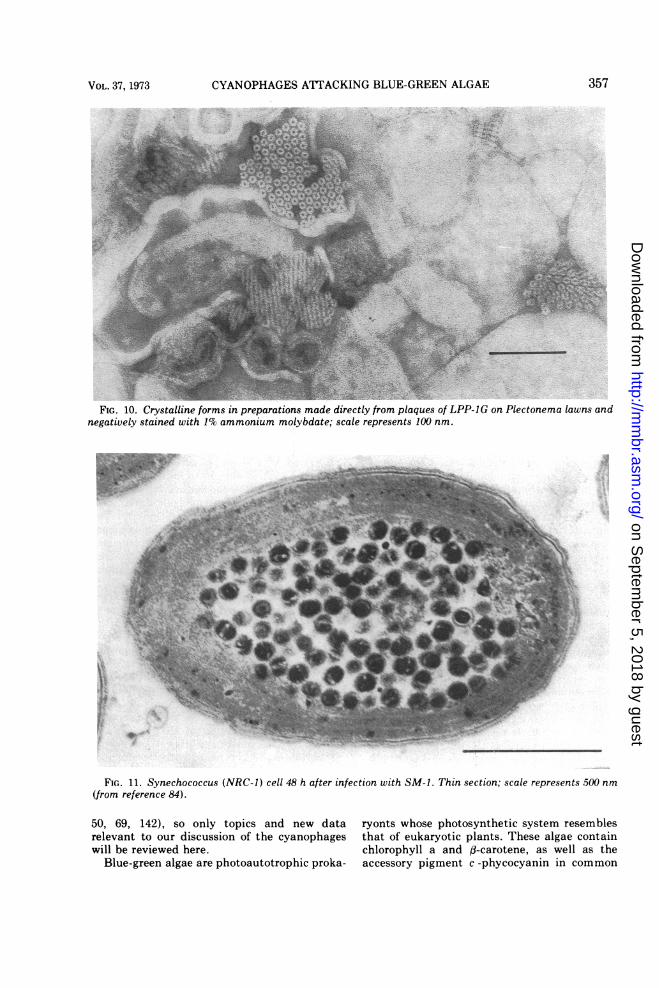
plaques, characteristic crystalline arrays areseen (Fig. 10). These appear almost exclusivelyin relation to the lamellae and may be seeneither in surface view as hexagonal crystalforms, whose basic units are 10 nm hexamers, orin side view as bridging two adjacent lamellaeand appearing as microtubules 4 nm in diame-ter. These crystalline forms were thought to bephycocyanin by Luftig and Haselkorn (77).However, these structures are not found inuninfected Plectonema lawns lysed by lysosymeand do not react with ferritin-labeled antibodiesprepared against phycocyanin. It is noteworthythat antibodies prepared against mature virusparticles also do not react with these forms.
Release of mature LPP-1G particles (at 26 C)
355VOL. 37, 1973
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

PADAN AND SHILO
FIG. 9. Aggregate of LPP-IG viral particles released from burst Plectonema cell. Prepared by in situ lysistechnique (see legend to Fig. 7); scale represents 1 gm.
is accomplished by cell lysis. Cell debris re-maining after lysis includes remnants of iso-lated vesicles of different size and shape whichmay be torn lamellae, cell membranes, andeven cell walls (95, 137, 139). Viral particleswere seen both in sections and with negativestaining, either embedded in or adsorbed onthese structures, by the distal end of their longtails (cf. Fig. 3). The attachment of the cyano-phages to all the different structures remainingafter lysis might well reflect a common compo-nent in the membranes acting as viral receptor.The presence of related antigen on the photo-synthetic vesicles and on the cell wall has, infact, been demonstrated in Rhodospirillumrubrum by Newton (88).
Lysis of various phage-infected bacteria isaccompanied by the formation of numerousvesicles consisting of cell membrane and wallmaterial (16). Different bacteriophages adsorbonto such structures after lysis (89).The morphology of unicellular blue-green
algae (Synechococcus) infected with virus SM-1has been studied in thin sections prepared atvarious times during the 50 h required for onecycle of virus growth (84). In contrast to LPPsystems, no changes in the photosynthetic la-mellae are seen prior to lysis. Furthermore,
SM-1 virions accumulate in the nucleoplasm.After 24 h, mature virions start appearing, andby 32 h the entire space interior to the photosyn-thetic lamellae is packed with virus particles(Fig. 11). Similar to our results with LPP-1G,forms that could be either precursors of SM-1particles or artifacts have been identified (84);these include apparently empty heads and con-densed "cores."The morphological aspects of N-1 and AS-1
infection cycles have not yet been studied. Inview of the localization of viral assembly proc-esses in a special site in the cell, the virogenicstroma, in LPP, but not in SM-1, such investi-gations would be most interesting, especiallybecause the assembled virions in most bacteriainfected with DNA-bacteriophages are evenlydistributed throughout the cell or are formed inthe nucleoplasm itself (16).
INTERACTION BETWEENCYANOPHAGE DEVELOPMENT AND
THE PHOTOAUTOTROPHICMETABOLISM OF THE BLUE-GREEN
ALGAL HOSTThe biochemistry and metabolism of blue-
green algae have been recently summarized (30,
356 BACTERIOL. REV.
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from
CYANOPHAGES ATTACKING BLUE-GREEN ALGAE
FIG. 10. Crystalline forms in preparations made directly from plaques of LPP-JG on Plectonema lawns andnegatively stained with 1% ammonium molybdate; scale represents 100 nm.
FIG. 11. Synechococcus (NRC-I) cell 48 h after infection with SM-i. Thin section; scale represents 500 nm(from reference 84).
50, 69, 142), so only topics and new data ryonts whose photosynthetic system resemblesrelevant to our discussion of the cyanophages that of eukaryotic plants. These algae containwill be reviewed here. chlorophyll a and d-carotene, as well as the
Blue-green algae are photoautotrophic proka- accessory pigment c -phycocyanin in common
357VOL. 37, 1973
I - I
I
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

PADAN AND SHILO
with some eukaryotic algae (30). Both photosys-tems I and II and related reactions, includingthe oxygen-evolution mechanism (14, 29, 147),are operative in blue-green algae, yielding elec-tron flow from H2O to triphosphopyridine nu-cleotide (oxidized form) and the accompanyingnoncyclic photophosphorylation. It is possibleto inhibit photosystem II by 3-(3, 4-dichlorophe-nyl)-1, 1-dimethylurea (DCMU) in blue-greenalgal cell extracts and to obtain only cyclicphotophosphorylation (147). This reaction hasbeen inferred indirectly in intact blue-greenalgal cells (91) and may perhaps be identicalwith that found in plants (148).Many strains of blue-green algae, considered
obligate autotrophs, were recently shown tohave heterotrophic metabolism (23, 32, 48, 49,59, 60, 62, 73, 98, 101, 134, 151, 153). Thisheterotrophic potency is also manifested in lightgrowth of these strains in the presence ofDCMU (59).
Effect of Cyanophage Development on HostMetabolism
Photosynthesis during infection. The studyof the effect of viral infection on systems supply-ing energy and metabolic intermediates in thehost has been almost neglected. Such a study inheterotrophic bacteria was conducted by Cohenand Anderson (24), who found that the respira-tion rate of E. coli cells infected with T2 isconstant throughout the latent period, an;oxygen uptake only stops at the time of celllysis. It was mainly from this work that thenotion emerged that phages do not directlyinfluence the energetic and intermediary me-tabolism of the host cell. However, it was foundrecently that the bacteriophage directly affectsthe cell membrane, thereby inhibiting bacterialrespiration at the end of the latent period andbefore lysis (55, 87).
Investigation of the energetic and intermedi-ary metabolism during viral infection in auto-trophic cells was restricted to studies of tissuesof higher plants until the isolation of the cyano-phages. Although respiration generally in-creased after infection (27), the study of photo-synthesis in infected plants yielded contradic-tory results. In chloroplasts isolated from Chi-nese cabbage infected with turnip yellow mosaicvirus (38), an increase in photophosphorylationwas observed. In tobacco leaves infected withtobacco mosaic virus, a decrease in photo-phosphorylation occurred 1 h after inoculation(90). Zaitlin and Jagendorf (164), however,found that both photophosphorylation and the
Hill reaction are normal in chloroplasts isolatedfrom the tobacco mosaic virus-infected plantif the nitrogen supply is sufficient. Contradic-tory results have also been obtained concern-ing CO2 photoassimilation and other metabolicpathways (12). The confusing findings may stemfrom experimental difficulties due to the compli-cated structure of higher plants and the almostcomplete impossibility of homogenic infection.The behavior of the photosynthetic system of
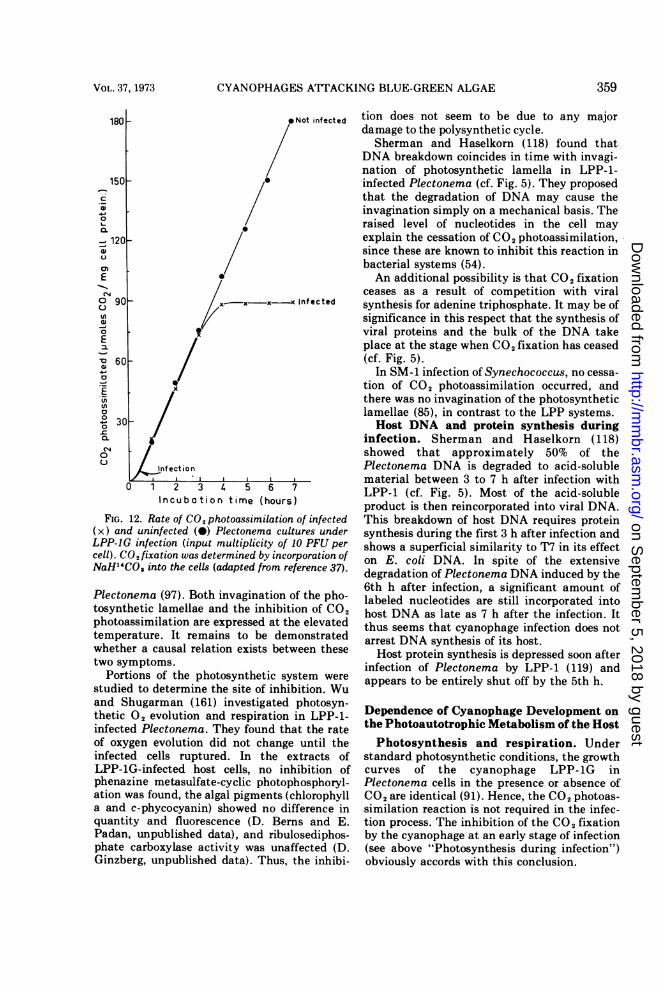
Plectonema during LPP-1G infection was stud-ied by following the CO2-fixation reaction (37).It was found that cyanophage infection effects arapid and complete cessation of CO2 photoas-similation. The cyanophage-induced inhibitionstarts 2.5 to 3 h after infection and becomescomplete by the 5th h (Fig. 12), which isdistinctly before lysis of the cells (7 to 13 h).These results were later confirmed for LPP-1 aswell (120).Adolph and Haselkorn (5) recently noted
similar inhibition of CO2 photoassimilation byNostoc cells after infection with N-1. Althoughthe effect is less immediate than that foundwith LPP-lG-infected Plectonema, it precedeslysis of the host cells.
Infection with a single LPP-1G virion perPlectonema cell is enough to inhibit CO2 fixa-tion completely (37), for neither the onset northe magnitude of the cyanophage-induced inhi-bition depend on the input multiplicity. Theeffect of nitrogen on photoassimilation in higherplants, mentioned above (164), was excluded forthe LPP-1G infection system, because nitrogensupplementation both before and during infec-tion did not influence the inhibition (37).
Light is required during the eclipse period forthe completion of the cyanophage developmentcycle at maximal burst size and during theentire latent period for completion of the cycleat the maximal rate (91). By excluding light fordifferent time periods and at different stages, itwas established that a fixed period of illumina-tion (1.5 to 2 h) after adsorption is required forthe cyanophage-induced inhibition (37). Thisimplies that an active process requiring energyand leading to inhibition of CO2 fixation isinduced by the cyanophage after attachment.
Invagination of the photosynthetic lamellaeof Plectonema cells appearing from 3 to 8 h afterLPP-1 infection requires protein synthesis dur-ing the first 3 h of infection (117). The coinci-dence in time of invagination and cessation ofthe CO2 photoassimilation is evident (cf. Fig.5). The relationship between these symptoms isfurther emphasized in the infection cycle at 35C of the temperature-sensitive strain LPP-1G in
358 BACTERIOL. REV.
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from
CYANOPHAGES ATTACKING BLUE-GREEN ALGAE
C
0
0.
/
_ 120
0~~~~E
a
0go
eifcto
0
0
0 -
0
-.3 0
Incubation time (hours
FIG. 12. Rate of CO2 photoassimilation
(x) and uninfected (0) Plectonema cullLPP-IG infection (input multiplicity of.cell). CO2 fixation was determined by incosNaH14CO3 into the cells (adapted from re
Plecton~ema (97). Both invaginationctosynthetic lamellae and the inhibiti
photoassimilation are expressed at tIhtemperature. It remains to be den
whether a causal relation exists bets
two symptoms.
Portions of the photosynthetic sylstudied to determine the site of inhil
and Shugarman (161) investigated
thetic 02 evolution and respiration
infected Plectonema. They found theof oxygen evolution did not change
infected cells ruptured. In the e
LPP-iG-infected host cells, no in}phenazine metasulfate-cyclic photop
ation was found, the algal pigments (ca and c-phycocyanin) showed no dii
quantity and fluorescence (D. Ben]Padan, unpublished data), and ribul
phate carboxylase activity was unal
Ginzberg, unpublished data). Thus,
oNot infected tion does not seem to be due to any majordamage to the polysynthetic cycle.Sherman and Haselkorn (118) found that
DNA breakdown coincides in time with invagi-nation of photosynthetic lamella in LPP-1-infected Plectonema (cf. Fig. 5). They proposedthat the degradation of DNA may cause theinvagination simply on a mechanical basis. Theraised level of nucleotides in the cell mayexplain the cessation of CO2 photoassimilation,since these are known to inhibit this reaction inbacterial systems (54).An additional possibility is that CO2 fixation
ceases as a result of competition with viralKinfected synthesis for adenine triphosphate. It may be of
significance in this respect that the synthesis ofviral proteins and the bulk of the DNA takeplace at the stage when CO2 fixation has ceased(cf. Fig. 5).
In SM-1 infection of Synechococcus, no cessa-tion of CO2 photoassimilation occurred, andthere was no invagination of the photosyntheticlamellae (85), in contrast to the LPP systems.Host DNA and protein synthesis during
infection. Sherman and Haselkorn (118)showed that approximately 50% of thePlectonema DNA is degraded to acid-solublematerial between 3 to 7 h after infection withLPP-1 (cf. Fig. 5). Most of the acid-solubleproduct is then reincorporated into viral DNA.
of infected This breakdown of host DNA requires proteintures under synthesis during the first 3 h after infection and10 PFU per shows a superficial similarity to T7 in its effectrporation of on E. coli DNA. In spite of the extensiveerence 37). degradation of Plectonema DNA induced by the
6th h after infection, a significant amount ofof the pho- labeled nucleotides are still incorporated intoon of CO2 host DNA as late as 7 h after the infection. Ite elevated thus seems that cyanophage infection does notnonstrated arrest DNA synthesis of its host.ween these Host protein synthesis is depressed soon after
infection of Plectonema by LPP-1 (119) andition. Wu appears to be entirely shut off by the 5th h.
photosyn- Dependence of Cyanophage Development onat the rate the Photoautotrophic Metabolism of the Hostuntil the Photosynthesis and respiration. Under
xtracts of standard photosynthetic conditions, the growthhibition of curves of the cyanophage LPP-1G inihosphoryl- Plectonema cells in the presence or absence ofchlorophyll CO2 are identical (91). Hence, the CO2 photoas-fference in similation reaction is not required in the infec-ns and E. tion process. The inhibition of the CO2 fixationlosediphos- by the cyanophage at an early stage of infectionffected (D. (see above "Photosynthesis during infection")the inhibi- obviously accords with this conclusion.
359VOL. 37, 1973
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

PADAN AND SHILO
In the presence of DCMU, the burst size andrate of virion release were the same as in normalinfection. Only the eclipse period is somewhatprolonged. Thus, it appears that neither noncy-clic photophosphorylation nor photoreductionof triphosphopyridine nucleotide (oxidizedform) are absolute requirements during theinfective process of the virus and that they onlyaccelerate the eclipse processes. Similar resultswere obtained in another LPP-1 strain (120);the slightly decreased burst size in the latterexperiments may be due to the exceedingly highconcentration of DCMU used.Because darkness depresses the burst size by
90% (Table 5), most of the adenosine triphos-phate required for viral synthesis under theseconditions must be derived from photophospho-rylation. Moreover, it is evident that cyclicphotophosphorylation can supply (in the pres-ence of DCMU) all the adenosine triphosphaterequirements of the virus. Accordingly, theuncoupler of phosphorylation at the electrontransfer level, carbonyl cyanide m-chloro-phenylhydrazone (CCCP) (147), was found toinhibit viral synthesis completely. Thus, sub-strate level phosphorylation or the existence ofan intracellular adenosine triphosphate pool donot seem to be significant under autotrophicconditions. Furthermore, anaerobiosis in thelight does not seem to influence the cyanophageinfection cycle (cf. Table 5), indicating thatoxidative phosphorylation under these condi-tions is quantitatively insignificant.
TABLE 5. Yield of cyanophages in Plectonema cellsincubated under different experimental conditions"
Percent ofYield' maximal
Conditios.(PFU/ burst sizeinfected under
cell) standardconditions
DarkAerobic 36 10Anaerobic 8 2Aerobic + CCCP (10-4 M) 0 0
LightAerobic 300-400 100Anaerobic 300-400 100Aerobic + DCMU (10 - 6 M) 300-400 100Anaerobic + DCMU (10-6 M) 300-400 100Aerobic + CCCP (10-4 M) 0 0
aTable reproduced from reference 93. CCCP, Car-bonyl cyanide m-chlorophenylhydrazone; DCMU,3-(3,4-dichlorophenyl)-1, 1-dimethylurea.
b Yield of cyanophage was determined from intra-cellular growth curves for each experimental condi-tion.
To determine when photophosphorylation isrequired in the cyanophage infection cycle, theinfection cycle of LPP- 1G was followed inPlectonema cells illuminated for different pe-riods of time from the beginning of the latentperiod (91). Illumination during the entireeclipse period is sufficient to produce the maxi-mal yield, although at a slower rate. Shorterillumination periods within the eclipse periodcaused decreases in yield and rate of formationof the infective particles in the cells. Continua-tion of illumination beyond the eclipse periodup to the end of the latent period progressivelyaccelerated the rate of intracellular synthesis ofthe infective virions. Light must also affectrelease of the viral particles, for only part of thechange in the rate of release can be explained byits effect on intracellular synthesis. Illumina-tion after free phages begin to appear does notaffect the cycle.The dependence of the cyanophage LPP-1
development cycle on photophosphorylation isnot absolute (92, 161). We have seen that 10% ofthe viral synthesis proceeds in the dark (cf.Table 5). No virus is produced in the dark underanaerobic conditions or in the presence of CCCP(92). Thus, viral multiplication in the darkmust depend for its energy on oxidative phos-phorylation. Indeed, oxidative phosphorylationhas been demonstrated in intact cells andextracts of several blue-green algae (15, 74, 75,114). Although the endogenous respiration ofblue-green algae preincubated in standard lightconditions is low, it can be enhanced progres-sively by increasing the period of preincubationin optimal photoassimilation conditions (highlight intensities and bicarbonate medium). Thisenhancement of endogenous respiration in in-fected cells is fully correlated with an increasedburst size up to 50% of light yield and rate ofviral synthesis (92). Similar cyanophage yieldsare obtained when Plectonema is grown hetero-trophically (on glucose substrate) and infectedin the dark (D. Ginzberg and B. Raboy, unpub-lished data).Sherman and Haselkorn (120) recently meas-
ured the infection cycle of LPP-1 in Plectonemacells kept in the dark for various times beforeand after infection. Although maintenance ofcells in the dark for 10 h after infection delayedthe viral synthesis, the final yield was abouttwice that of the control, and the rate of releaseof virions was accelerated. These authors as-sumed that incomplete viral particles accumu-lating in the postinfection dark period arerapidly completed upon illumination. They fur-ther inferred that early viral functions proceedas efficiently in the dark as in the light. How-
360 BACTERIOL. REV.
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

CYANOPHAGES ATTACKING BLUE-GREEN ALGAE
ever, two early functions, the capacities tosynthesize DNA and degrade host DNA, are
delayed in the dark. It seems equally plausiblethat the algal cells must be in a specificphysiological state for replication of the cyano-phage. Synchronization of the algal population(by introduction of dark periods) could contrib-ute to more efficient infection cycles, as was
recently demonstrated in another phage system(155). Synchrony of the infection seems to bethe explanation for the finding that dark pe-riods before and after infection markedly accel-erated phage-induced inhibition of CO2 pho-toassimilation (37).
In general, the dependence of the N-1 infec-tion cycle on the energy metabolism of N.muscorum resembles the situation found inLPP-1- and LPP-lG-infected Plectonema, al-though some differences may be noted (5).DCMU reduces yields in N-1 infections to 25%of the control, whereas it had no effect on LPP-1yields. As expected, addition of CCCP in thelight drastically reduces the viral yield to 2% ofthe control. Because N-1 multiplication is notcompletely abolished in the presence of thisinhibitor, it may be inferred that limited adeno-sine triphosphate pools or substrate level phos-phorylation may be present in Nostoc cells andcontribute in a limited way to N-1 multiplica-tion in the light. N-1 appears to be more
dependent on photosynthesis than LPP (5).However, this finding should be reevaluated inthe light of our results that the respiration isclosely linked to the growth history and physio-logical stage of the algae.
In contrast to LPP and N-1 cyanophages inwhich the dependence on photosynthesis is notabsolute, SM-1 development in the infectedSynechococcus is completely stopped byDCMU, darkness, or absence of CO2. SM-1 isconsiderably more dependent upon host photo-synthetic metabolism than any previously stud-ied cyanophage (85). In SM-1 infection, CO2photoassimilation is not inhibited, photosyn-thetic lamellae do not invaginate, and virusesdevelop in the nucleoplasm.Building blocks. Sherman and Haselkorn
(118) showed that approximately 50% of thehost DNA is degraded 6 to 7 h after LPP-1infection, and most of the acid-soluble productsare reincorporated into viral DNA.
Preliminary results of A. Rimon in our labora-tory indicate that proteins of the algal host,synthesized before the infection, contribute tothe synthesis of cyanophage proteins (103). Inan experiment in which sulfate-starvedPlectonema cells were uniformly labeled with35S, by the method of Sherman and Haselkorn
(119), about 90% of the host label was recoveredin proteins; the residual 10% was probablybound to polysaccharides. Because almost nocold-acid-soluble label was recovered after sul-fate labeling, it was inferred that the sulfur-amino acid pool in the cells is very small. Thelabeled cells were washed, infected, and dividedinto the three experimental systems describedin Table 6. After completion of the infectioncycle, the cyanophages were purified, and theirradioactivity was determined. The phage pro-teins produced in the control system becamehighly labeled, and their computed specificactivity reached that of the host proteins. Thissimilarity of specific activities indicates thatalgal proteins are the major precursors of phageproteins and that CO2 photoassimilation doesnot contribute significantly, even before it isinhibited in the infected cell (see above "Photo-synthesis during infection"). This is also sup-ported by similar specific activities of cyano-phage proteins produced in the presence orabsence (due to DCMU addition) of CO2 pho-toassimilation. When the experiment was con-ducted with methionine and Na2SO,, the com-puted specific activity of cyanophage proteinsdecreased to 15% of that of the host proteins.After infection, proteins of the host cells proba-bly undergo degradation to amino acids whichare then reincorporated into the phage proteins.The residual labeling of the cyanbophage in thepresence of the methionine chase may be due tolabeled sulfate-containing polysaccharides thatalso may contribute to phage proteins. There
TABLE 6. Contribution of host proteins to cyanophageLPP-lG proteinsa
Specific Ratio of35S label Cyano- activity specific
Experimental of cyano- phage (counts activitiessystem phage yield per min of viral
(counts/ (PFU) permg of proteins tomin) viral pro- host pro-
teins)" teins (%)
NaSO4 + 5,790 2.73 x 109 4.25 x 107 15.0methionine
DCMU 50,800 4.44 x 109 2.45 x 108 91.0Control 30,260 2.92 x 109 2.14 x 108 79.2
a Proteins of Plectonema cells were labeled with 35Saccording to Sherman and Haselkorn (119) and incubatedunder the standard experimental infective conditions (91) atan input multiplicity of 10 PFU/cell in the above systems.After a 9-h incubation, the cells were ruptured, virions werepurified by differential centrifugation and in CsCl gradients(103), and the amount of isotopes in the cyanophages wasdetermined. DCMU, 3-(3, 4-Dichlorophenyl)-1, 1-dime-thylurea.
' For determination of viral specific activity, 52% of thevirus particle weight was assumed to be protein with amolecular weight of 30 x 106 (40).
361VOL. 37, 1973
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

PADAN AND SHILO
are indications that polysaccharides breakdown during infection (118).
In the light of these preliminary indications,several interesting problems remain to be ex-amined. One problem is whether all or onlyselected host proteins are utilized by the cyano-phage. Another is whether mainly host proteinsare required for cyanophage protein synthesis orwhether other cellular constituents can contrib-ute building blocks. Finally, there is the ques-tion of how different physiological conditions,such as starvation in the dark or loading thecells with reserve materials, affect the cyano-phage protein synthesis.
Since cyanophage (LPP-1G) can multiplywithout an exogeneous carbon source and mostprobably without external nitrogen as well, hostcomponents must supply all the building blocksfor viral synthesis. Furthermore, growth of thehost cells is not required for LPP multiplica-tion; this differs from that of most bacterio-phage which only develop efficiently in growingcells. Synthesis of bacteriophages attackingheterotrophic hosts depends on exogeneous sub-stances (125), except for host DNA in somecases (82).The unique dependence of the LPP-1G cya-
nophage on host cellular constituents for build-ing blocks represents an extreme degree ofhost-parasite interaction. Such extreme para-sitism could be visualized in a photoautotrophichost in which the photosynthetic system sup-plies adenosine triphosphate, and minimalbreakdown of host proteins together with hostDNA utilization suffice to satisfy phage synthe-sis. This extreme dependence even seems tooffer selective advantages, because the infectiveprocess would thus be less dependent on envi-ronmental factors. The lack of strict control ofenzyme synthesis found for blue-green algae(51, 52, 76, 99, 101) may allow a spontaneousflood of intermediates without disturbing thestate of the cell. Heterotrophic bacteria, on theother hand, have biosynthetic and energy me-tabolism depending on substrate supply, and astrict control mechanism governs the steadystates of the cell. In such hosts, the agent thatdisturbs the host metabolism least is the onewith the selective advantage. A study of othercyanophage systems and other viral infectionsof other photoautotrophic and chemoauto-trophic prokaryonts, such as Rhodopseu-domonas spheroides (34) and Hydrogenomonas(63), is needed to evaluate these suggestions.It would also be interesting to identify theprecursors of LPP synthesis in dark conditionsas compared with those in photosynthesizing
cells, as well as those of SM-1 which showsextreme dependence on CO2 photoassimilation.
ECOLOGICAL CONSIDERATIONSThe hosts of cyanophages, the blue-green
algae, are widely spread in the aquatic environ-ment, although some terrestrial species are alsoknown (50). In freshwaters, the blue-green algaeexhibit characteristic cyclic growth; they bloomin large masses usually during the summer, andthen often spontaneously die off and lyse (122).Although this decomposition causes enormouseconomic damage by deteriorating water qual-ity and by causing mass mortality of fishpopulations, neither the causes of the extensivegrowth of the blue-green algae nor the factorsleading to their rapid decomposition are thor-oughly understood.
Ever since the discovery by Safferman andMorris of a virus attacking blue-green algae,particular attention was paid to the ecology ofthe cyanophages and to studies of their relation-ships with the algal host in the ecosystem andtheir possible role in the fluctuations of blue-green algae populations (109, 123).
Systematic searches for cyanophages weretherefore undertaken in all parts of the world.Safferman and his group showed that the LPPcyanophages represent a steady population inwaste stabilization ponds (110). Strains ofLPP-1 and LPP-2 were found in all seasons in awide variety of locations in the United States(111). Shane (116) initially screened the Chris-tina River area of the Delaware River and foundLPP-type cyanophages in all waters tested (in-cluding natural lakes, quarries, farm and resi-dential ponds, industrial storage tanks, stabili-zation lagoons, and streams and rivers). Waterareas near urban settlements gave higher viralcounts. Routine collections from sewage oxida-tion ponds gave the highest viral concentra-tions. Safferman analyzed these isolates ofShane and found them all to be identical orsimilar to LPP-1.A variety of LPP types, including LPP-1 and
LPP-2 types, and a temperature-sensitive strain(97) have been isolated in Israel from brackishand freshwater ponds and from Lake Kinneret(93).Viruses attacking Plectonema, Phormidium,
and Lyngbya were also isolated in a number ofsewage ponds in Scotland (26). One isolate wasshown to be serologically identical with LPP-1.These isolates showed some variability inplaque size, indicating that different viralstrains are present. The cyanophages were con-sistently detected throughout the year in one of
362 BACTERIOL. REV.
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

CYANOPHAGES ATTACKING BLUE-GREEN ALGAE
the ponds. Findings of LPP-type cyanophagesalso have been reported from India in sewageponds (130) and rice fields (128) and in theSoviet Union (44).
Other types of cyanophages have also beenisolated in very divergent geographical andhabitational locations. Cyanophages activeagainst vegetative cells of A. raciborskii andCylindrospermum were reported from India(130). SM-1 cyanophage was isolated from awaste stabilization pond in Indiana (112), andAS-1 was also reported from the United States(106). A group of Swedish investigators (43) anda group in the Soviet Union (64) describedviruses active on A. variabilis. Two cyano-phages active against different species ofMicro-cystis were reported by a Kiev-based group ofinvestigators (41, 42). Cyanophage N-1 wasisolated from Lake Mendota in Wisconsin (4).
It is now clearly evident that cyanophages areubiquitously distributed in freshwater bodies.Systematic screenings of Israeli fishponds (93)and oxidation ponds in the United States (110)for cyanophages attacking Plectonema yieldedcyanophages in 70 to 90% of the samples.Safferman et al. (110) found numbers of 4 to 270PFU per ml throughout the year. The cyano-phage yields obtained are comparable to bacte-riophage titers recorded in sewage (8). Numbersof these viruses in selected fishponds in Israelover a year generally ranged between a few toseveral tens of PFU per liter; a drastic increasein number to several thousands occasionallyoccurs during the summer when intensive blue-green algal blooms are noted (93). The fact thatLPP cyanophages maintain their numbers inhabitats with appreciable flow rates (up to 1.5x 106 gallons/day) indicates that the virus isbeing replicated rapidly in nature (26).An instructive observation, made by Saffer-
man and Morris (110) and in our laboratory(93), was the fact that the susceptible algalgenera were never dominant in ponds where theappropriate cyanophages appeared in signifi-cant amounts. This possibly may be due tocontinuous lysis of sensitive algae by prolifera-ting cyanophages, so that population equilib-rium among them is established. The worldwidedistribution of LPP-type cyanophages and thefact that Lyngbya, Phormidium, andPlectonema are not known to form bloomssuggest that such an equilibrium indeed existsin many natural water bodies.The predominance of finds of LPP-type cya-
nophages the world-over is striking, but notsurprising if one considers that their host generawere described by Drouet (28) as ecophenes of asingle species, Schizotrix calcicola, which seems
to be "the most widely distributed and mostfrequently encountered blue-green alga onearth." In addition, the hosts of LPP viruses (inmost cases, P. boryanum) have been the speciesmost widely used in isolation by direct platingof samples (110) or of concentrates (93), or havebeen the host organism for selective enrich-ments (93, 110) and baiting of the cyanophagesin situ (26). The limited success in isolationmay also be due to the use of blue-green algalstrains kept for years in culture collections andreadily available to the investigators, but notnecessarily the most suitable hosts for thecyanophages from the natural environment. Atpresent, fresh isolation of prevalent blue-greenalgal forms and their use as hosts are underwayin our laboratory.The interaction of cyanophage with its host is
markedly influenced by environmental condi-tions. The requirement for high (0.001 M)magnesium ion concentration in the milieu formaintenance of activity of LPP-1 (40, 113) andpossibly N-1 (4) has been demonstrated. On theother hand, AS-1 (106) and SM-1 (112) cyano-phages show stable activity in water lacking thecation. Although the salinity range of cyano-phages has not been rigidly determined, LPPphages have been isolated from brackish fishponds with salinities reaching 2,000 mg ofCl-/liter (110). Interestingly, NaCl exerts astabilizing effect on attachment in the AS-1system (106).The tolerance of the cyanophages (see "Phys-
icochemical Characteristics") to alkalinity ac-cords with the alkaline range (7 to 11) ofPlectonema, which shows little or no growthbelow neutrality. Most other blue-green algaeprefer the alkaline range, which prevails inaquatic systems where these algae thrive (illus-trated in the extreme in several African equato-rial lakes, such as Lake Aranguadi, the "greenlake" near Debra Zet in Ethiopia, which havepermanently high alkaline pHs and continuousblooms of Spirulina platensis). Survival of cya-nophages at high pHs must have great ecologi-cal significance as a selective factor in condi-tions common to a great many blue-green algalblooms.
Additional environmental factors influencingthe host-phage interaction are temperature ef-fects on the survival of free phage and oncyanophage development in hosts. LPP andSM-1 cyanophages show similar temperaturesensitivity: 85% of the free phage particlesremain infective at temperatures up to 40 C,whereas only 55% attack at 45 C, and less than0.001% at 50 C (108, 112). Two LPP isolatesfrom nature (LPP-1 and LPP-1G) are tempera-
363VOL. 37, 1973
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

PADAN AND SHILO
ture sensitive (97). They multiply normally inthe 26 to 29 C range, but produce only earlysymptoms of infection in hosts (invagination ofthe photosynthetic lamellae and cessation ofCO2 photoassimilation) above 31 C. The stabil-ity of the different cyanophages to temperaturesup to 50 C is also consistent with the blue-greenalgal temperature range in nature which isbetween 20 to 50 C, and is even higher forspecial thermophilic types.Provided that adenosine triphosphate supply
is ensured, certain cyanophages (LPP) can growand replicate in the dark and under conditionsinhibiting host growth (such as lack of anyexternal carbon sources and presence ofDCMU), whereas bacteriophages require ac-tively multiplying hosts. This wide tolerance ofthe LPP cyanophages makes them quite suitedto succeed in surviving and reproducing innatural conditions. The natural life cycle of avirus whose replication demands photosyn-thetic CO2 fixation must be extremely hazard-ous. Incubation of SM-1 for 12 h in the dark isenough for irreversible lose of 20% of plated,infectious centers. Mackenzie and Haselkorn(85) expect to find rather striking circadianphenomena characteristic of SM-1 populationsin nature.
All of the cyanophages isolated until recentlyshowed characteristics of virulent phages. How-ever, LPP-1D, LPP-2SPI, and other cyano-phages were shown to be temperate phages (20,21, 96, 132). Attempts at induction of the SPIprophage with ultraviolet, X rays, or mitomycinC proved relatively unsuccesful. However, whena thermosensitive mutant of SPI was prQphage,its induction was accomplished by transferringthe lysogenic culture from 25 to 35 C (96).Factors, such as temperature, ultraviolet, orunbalanced growth conditions, which areknown to influence formation of lysogenic cul-tures and the lytic cycle, might have considera-ble significance on the algal population balancein nature. The lytic cycle could be induced if athermosensitive, lysogenic algal population innature is exposed to elevated temperatures, aswas seen experimentally.The role of cyanophages in determining the
cyclic blooms of blue-green algae in nature hasbeen suggested ever since these viruses werediscovered. Daft et al. (26) consider it possiblethat viral and microbial pathogens of algae maybe even as important in determining bloomformation and disappearance in nature as areconventional factors such as light intensity andnutrient level. Indeed, an effect on certainspecies is indicated (as illustrated by the factthat Plectonema blooms never form in nature).
However, it is likely that blue-green algal popu-lation fluctuations cannot be solely related tocyanophage attack. Die-off has been observed inmany different blue-green algal species, some ofwhich are not yet known to be attacked bycyanophages. In certain other cases, sensitiza-tion of blue-green algal populations to photooxi-dation in conditions of CO2 limitation (1, 2)seems to be the cause of the algal die-offs.The possible use of cyanophages for biological
control has been suggested (105). By simulatingconditions of a Plectonema bloom in the labora-tory, Safferman et al. (109) were able to provethe feasibility of bloom control with LPP cyano-phages. Jackson and Sladecek (53) studied theinteraction of LPP-1 with its host P. boryanumon a large scale in 5,000-gallon tanks. Theyconcluded that algal viruses can prevent growthof their blue-green algal hosts under naturalconditions, because all attempts to grow thealgae in effluents of a sewage treatment plantfailed, but excellent growth of Plectonema wassupported by autoclaved samples of the sewagewater. Russian investigators have claimed suc-cess in using the virus to clear Microcystisblooms in an experiment in a large waterreservoir in the Ukraine (L. A. Sirenko, personalcommunication). These experiments werebased on observations of V. Goriushyn andothers (42, 104) that the Kremenchug reservoirhad clear areas even at the height of aMicrocystis bloom. He found that within theseclear patches the algae had been destroyed by avirus which had not previously been character-ized.An important consideration in establishing
the role of cyanophages and their potential ascontrol agents in nature is the rapid appearanceof resistant host mutants. Algal mutants resist-ant to LPP-1 cyanophages were found at afrequency of 107. HoWever, such resistant hostsare still susceptible to siable host-range phagemutants (44, 95). One of the resistant algalclones isolated by Gromov and Kozykov (44)appeared to be refractive in all their tests.Resistance to phages among the algal mutantstested by us seems to involve a change in thealgal cell envelope which prevents phage ad-sorption (95).The high degree of host specificity, the selec-
tion of resistant host mutants, and the depend-ence on environmental factors indicate thecomplexity of the alga, cyanophage interaction,whose outcome depends not only on inherentproperties of both phage and host, but onfluctuations in external conditions as well. Thelength of the phage developmental cycle whichvaries over a large range is still another, perhaps
364 BACTERIOL. REV.
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

CYANOPHAGES ATTACKING BLUE-GREEN ALGAE
critical, factor in the choice of the phage prom-ising for control. Deliberate manipulation, bothof viral genetic material and of external condi-tions, could serve as the key to prospectivebiological control of undesirable blue-greenalgal blooms.
GENETICS OF CYANOPHAGESApart from several spontaneous mutants,
neither mutagenesis nor recombination havebeen reported so far for cyanophages, althoughcomplementation and recombination in thetemperature-sensitive mutants (LPP-2-SPI)have been indicated in the most recent work ofA. Rimon in our laboratory. The spontaneousmutations, reported for LPP cyanophages, in-clude acriflavine resistance (131) and rapid lysisrate (108, 131). Host-range mutants of LPPcyanophages have also been isolated (95, 131).
Recently Cannon et al. (20, 21) reported onthe lysogeny of P. boryanum. Addition of mi-tomycin C induced a 100-fold increase in virusproduction between 4 and 5 h after treatment.When the lysogenic cultures were grown withantivirus serum, all free virus was eliminated. Alysogenic strain of Anabaenopsis has also beenindicated (132). We have shown that LPP-2-SPIis a temperate cyanophage (96). Immunity israpidly conferred on Plectonema cells 90 minafter infection with LPP-2-SPI. About 80% ofthe Plectonema cells become lysogenic; culturesof such cells always contain virulent phages,probably produced by occasional induction.The frequency of induction can be increasedonly fivefold after ultraviolet or X-ray irradia-tion. When a temperature-sensitive mutant ispresent as a prophage, efficient induction canbe accomplished by heat treatment of thelysogenic culture. The mechanism of lysogeni-zation of Plectonema by LPP-2-SPI thus seemssimilar to that of lysogenic bacteriophages andconsists of a proteinous repressor. The lysogenicstrain P1(SPI) is also immune to infection by allother phages of the LPP-2 group, indicatingthat all LPP-2 strains are homoimmune (96).This further justifies the serological classifica-tion of LPP cyanophages into LPP-1 and LPP-2groups (111). The ecological importance of lyso-genic strains has been discussed.The genetics of blue-green algae is poorly
known. Some cases of mutagenesis have beenreported (9-11, 66-68, 129, 131, 141, 144, 145).Recombination has been claimed by some au-thors (13, 65, 133), while these results have beenquestioned by others (102). Recently, transfor-mation was also reported (45, 46, 121). Thetemperate cyanophages may allow for transduc-tion studies, which can lead to extensive genetic
investigations of the blue-green algae and cya-nophages. Searches are currently underway inour laboratory for LPP-2-SPI-mediated trans-duction. Singh et al. (132) claim transduction ofstreptomycin resistance in P. boryanum byLPP-1.
CONCLUDING REMARKSA general conception of the sequence of the
developmental cycle of several groups of cyano-phages is now available to us. Nevertheless,details of phage synthesis, assembly, and thenature of the interaction with the host systemand of regulatory processes involved at themolecular level have yet to be unraveled.The absolute dependence of SM-1 cyano-
phage on the photosynthetic activity of itsunicellular host is in sharp contrast to thewell-documented action of LPP and N-1 cyano-phages on their filamentous hosts. The infectioncycle of the latter groups of cyanophages notonly can proceed independently of host photo-synthesis, but disrupts (structurally and physi-ologically) the host's photosynthetic apparatus.These different patterns of interaction suggestdivergent evolutionary trends in the develop-ment of these blue-green algae. In the light oftheir unique position, further studies of theinfection cycle may aid in clarifying the phylo-genetic development of this ancient group.Use of phage material to induce transduction
should become a valuable tool in opening up thefield of genetical studies of the blue-green algae.Among the more promising lines of study wouldbe a genetical approach to studies of the struc-ture and function of the cyanophytic photosyn-thetic system.The ubiquitous distribution of the LPP cya-
nophages and the fact that their host algae,though widespread, never form blooms serve asindications of the importance of cyanophages indetermining the dynamics of algal populationsin nature. Nevertheless, much has yet to belearned of the role of the cyanophages in differ-ent ecosystems. Finally, the use of cyanophagesin manipulating and controlling algal bloomsremains an open but still promising question.
ACKNOWLEDGMENTSThe preparation of this study was supported
through the Special Foreign Currency Program of theNational Library of Medicine, Public Health Service,under an agreement with the Israel Journal of Medi-cal Scences, Jerusalem, Israel.We are indebted to the publishers of Virology and
the authors for permission to reproduce Fig. lb, c, andd, 2, 3, 4, and 11.The original results of Dalia Ginzberg, Avraham
365VOL. 37, 1973
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

PADAN AND SHILO
Rimon, and Bilha Raboy of our laboratory have beenpresented here and are acknowledged with our appre-ciation.
Special thanks are due to Martin Kessel for theoriginal electron micrographs (Fig. la, 6-10) and forhis participation in writing the section on morphol-ogy, to Binah Golek for aid in preparing the manu-script, and to Muriel Isaacs for devotedly typing it.Finally we should like to thank Donald Berns, Morde-chai Avron, and Amos Oppenheim for their criticalreading.
LITERATURE CITED1. Abeliovich, A., and M. Shilo. 1972. Photooxida-
tive reactions of c-phycocyanin. Biochim. Bio-phys. Acta 203:483-491.
2. Abeliovich, A., and M. Shilo. 1972. Photooxida-tive death in blue-green algae. J. Bacteriol.111:682-689.
3. Adams, M. H. 1959. Bacteriophages. Inter-science Publishers, New York.
4. Adolph, K. W., and R. H. Haselkorn. 1971.Isolation and characterization of a virus in-fecting the blue-green alga Nostoc muscorum.Virology 46:200-208.
5. Adolph, K. W., and R. H. Haselkorn. 1972.Photosynthesis and the development of blue-green algal virus N-1. Virology 47:370-374.
6. Adolph, K. W., and R. H. Haselkorn. 1972.Comparison of the structures of blue-greenalgal viruses LPP-1M and LPP-2 and bacteri-ophage T7. Virology 47:701-710.
7. Ames, B. N., and D. T. Dubin. 1960. The role ofpolyamines in the neutralization of bacterio-phage deoxyribonucleic acid. J. Biol. Chem.235:769-775.
8. Anderson, E. S. 1957. The relations of bacterio-phages to bacterial ecology, p. 189-217. Symp.Soc. Gen. Microbiol. 7th
9. Asato, Y. 1972. Isolation and characterization ofultraviolet light-sensitive mutants of the blue-green alga Anacystis nidulans. J. Bacteriol.110:1058-1064.
10. Asato, Y., and C. E. Folsome. 1969. Mutagenesisof Anacystis nidulans N-nitro-N-nitroso-guanidine and UV irradiation. Mutat. Res. 8:531-536.
11. Asato, Y., and C. E. Folsome. 1970. Temporalgenetic mapping of the blue-green alga Ana-cystis nidulans. Genetics 65:407-419.
12. Baur, J. R., B. Richardson, R. S. Halliwell, andR. Langston. 1967. Effect of tobacco mosaicvirus infection on glucose metabolism in Nico-tiana tabacum var. Samsun. Virology32:580-588.
13. Bazin, M. J. 1968. Sexuality in a blue-greenalga: genetic recombination in Anacystisnidulans. Nature (London) 218:282-283.
14. Biggins, J. 1967. Photosynthetic reactions bylysed protoplasts and particle preparationsfrom the blue-green alga, Phormidiumluridum. Plant Physiol. 42:1447-1456.
15. Biggins, J. 1969. Respiration in bfue-green algae.J. Bacteriol. 99:570-575.
16. Bradley, D. E. 1967. Ultrastructure of bacterio-phages and bacteriocins. Bacteriol. Rev.31:230-314.
17. Bradley, D. E., and C. A. Dewar. 1967. Intracell-ular changes in cells of Escherichia coli in-fected with a filamentous bacteriophage. J.Gen. Virol. 1:179-188.
18. Brown, R. M., Jr. 1972. Algal viruses. Advan.Virus Res. 17:243-277.
19. Brown, R. M., Jr., K. M. Smith, and P. L.Walne. 1966. Replication cycle of the blue-green algal virus LPP-1. Nature (London)212:729-730.
20. Cannon, R. E., and M. S. Shane. 1972. Theeffect of antibiotic stress on protein synthesisin the establishment of lysogeny of Plec-tonema boryanum. Virology 49: 130-133.
21. Cannon, R. E., M. S. Shane, and V. N. Bush.1971. Lysogeny of a blue-green alga, Plec-tonema boryanum. Virology 45:149-153.
22. Chao, L., and C. C. Bowen. 1971. Purificationand properties of glycogen isolated from ablue-green alga, Nostoc muscorum. J. Bacte-riol. 105:331-338.
23. Cheung, W. Y., and M. Gibbs. 1966. Dark andphotometabolism of sugars by a blue-greenalga: Tolypothrix tenuis. Plant Physiol.41:731-737.
24. Cohen, S. S., and T. F. Anderson. 1946. Chemi-cal studies of host-virus interactions. I. Theeffect of bacteriophage adsorption on the mul-tiplication of its host Escherichia coli B. J.Exp. Med. 84:511-521.
25. Cowie, D. B., and L. Prager. 1969. Cyanophytaand their viruses, p. 391-397. In CarnegieInstitution of Washington Year Book, 1968-69.Carnegie Institution of Washington, Washing-ton, D.C.
26. Daft, M. J., J. Begg, and W. D. P. Stewart. 1970.A virus of blue-green algae from fresh waterhabitats in Scotland. New Phytol.69:1029-1038.
27. Diener, T. 0. 1963. Physiology of virus-infectedplants. Annu. Rev. Phytopathol. 1:197-218.
28. Drouet, F. 1963. Ecophenes of Schizothrix cal-cicola (oscillatoriaceae). Proc. Nat. Acad. Sci.U.S.A. 115:261-281.
29. Duane, W. C., M. C. Hohl, and D. W. Krogman.1965. Photophosphorylation activity in cell-free preparations of a blue-green alga. Bio-chim. Biophys. Acta 109:108-116.
30. Echlin, P., and I. Morris. 1965. The relationshipbetween blue-green algae and bacteria. Biol.Rev. (Cambridge) 40:143-187.
31. Edelman, M., D. Swinton, J. A. Schiff, H. T.Epstein, and B. Zeldin. 1967. Deoxyribonu-cleic acid of the blue-green algae (cyano-phyta). Bacteriol. Rev. 31:315-331.
32. Fay, P. 1965. Heterotrophy and nitrogen fixationin Chlorogloea fritschii. J. Gen. Microbiol.39:11-20.
33. Fogg, G. E. 1969. The physiology of an algalnuisance. Proc. Roy. Soc. Ser. B. Biol. Sci.173:175-189.
34. Freund-Molbert, E., G. Drews, K. Bosecker, and
366 BACTERIOL. REV.
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

CYANOPHAGES ATTACKING BLUE-GREEN ALGAE
B. Schubel. 1968. Morphologie und Wirstkreiseines neu isolierten Rhodopseudomonaspalustris-phagen. Arch. Mikrobiol. 64:1-8.
35. Ginzberg, D. 1973. The interaction betweencyanophage LPP-1G and the photosyntheticsystem of the blue-green alga Plectonemaboryanum. Ph.D. thesis, Hebrew University,Jerusalem.
36. Ginzberg, D., and E. Padan. 1972. Light effecton the internal osmotic pressure of the blue-green algae Plectonema boryanum and Phor-midium luridum. Arch. Mikrobiol.87:181-183.
37. Ginzburg, D., E. Padan, and M. Shilo. 1968.Effect of cyanophage infection on CO2 pho-toassimilation in Plectonema boryanum. J.Virol. 2:695-701.
38. Goffeau, A., and J. M. Bove. 1965. Virus infec-tion and photosynthesis. 1. Increased photo-phosphorylation by chloroplasts from Chinesecabbage infected with turnip yellow mosaicvirus. Virology 27:243-252.
39. Goldstein, D. A., and I. J. Bendet. 1967. Physicalproperties of the DNA from the blue-greenalgal virus LPP-1. Virology 32:614-618.
40. Goldstein, D. A., I. J. Bendet, M. A. Lauffer,and K. M. Smith. 1967. Some biological andphysicochemical properties of blue-green algalvirus LPP-1. Virology 32:601-613.
41. Goriushyn, V. A., and S. M. Chaplinskyaya.1966. Existence of viruses of blue-green algae.Mykrobiol. Zhurn. (Kiev) 28:94-97.
42. Goriushyn, V. A., and S. M. Chaplinskyaya.1968. The discovery of viruses lysing blue-green algae in the Dneprovsk reservoirs.Tsvetenie Vody, Dumka, Kiev.
43. Granhall, U., and A. V. Hofsten. 1969. Theultrastructure of a cyanophage attack on Ana-baena variabilis. Physiol. Plant. 22:713-722.
44. Gromov, B. V., and S. Y. Koz'yakov. 1970.Study of the properties of the interrelationshipbetween populations of the blue-green algaPlectonema boryanum GOM, and the cyano-phage LPP-1. Vestn. Leningrad Univ. Ser.Biol. 1:128-135.
45. Herdman, M., and N. G. Carr. 1971. Recombi-nation in Anacystis nidulans mediated by anextracellular DNA/RNA complex. J. Gen. Mi-crobiol. 68:14.
46. Herdman, M., and N. G. Carr. 1972. The isola-tion and characterization of mutant strains ofthe blue-green alga, Anacystis nidulans. J.Gen. Microbiol. 70:213-220.
47. Hershey, A. D., and M. Chase. 1952. Independ-ent function of viral protein and nucleic acidin growth of bacteriophage. J. Gen. Physiol.36:39.
48. Hoare, D. S., S. L. Hoare, and A. J. Smith. 1969.Assimilation of organic compounds by blue-green algae and photosynthetic bacteria, p.1540-1573. In H. Metzner (ed.), Progress inphotosynthetic research III.
49. Hoare, D. S., L. 0. Ingram, E. L. Thurston, andR. Walkup. 1971. Dark heterotrophic growthof an endophytic blue-green alga. Arch. Mik-
robiol. 78:310-321.50. Holm-Hansen, 0. 1968. Ecology, physiology and
biochemistry of blue-green algae. Annu. Rev.Microbiol. 22:47-70.
51. Hood, W., and N. G. Carr. 1971. Apparent lackof control by repression of arginine metabo-lism in blue-green algae. J. Bacteriol.107:365-367.
52. Hood, W., A. G. Leaver, and N. G. Carr. 1969.Extracellular nitrogen and the control of argi-nine biosynthesis in Anabaena variabilis. Bio-chem. J. 114:12P.
53. Jackson, D. F., and Sladecek, V. 1970. Algalviruses-eutrophication control potential.Yale Sci. Mag. 44:16-22.
54. Johnson, E. J. 1966. Occurrence of the adenosinemonophosphate inhibition of carbon dioxidefixation in photosynthetic and chemosyn-thetic autrophs. Arch. Biochem. Biophys.114:178-183.
55. Josslin, R. 1970. The lysis mechanism of phageT4: mutants affecting lysis. Virology40:719-726.
56. Kaye, A. M., R. Salomon, and B. Fridlander.1967. Base composition and presence of meth-ylated bases in DNA from a blue-green alga,Plectonema boryanum. J. Mol. Biol.24:479-483.
57. Kellenberger, E., F. A. Eiserling, and E. Boy dela Tour. 1968. Studies on the morphopoiesis ofthe head of phage T-even. III. The cores ofhead-related structures. J. Ultrastruct. Res.21:335-360.
58. Kellenberger, E., J. Sechaud, and A. Ryter.1959. Electron microscopical studies of phagemultiplication. IV. The establishment of theDNA pool of vegetative phage and the matu-ration of phage particles. Virology 8:478-498.
59. Kenyon, C. N., R. Rippka, and R. Y. Stanier.1972. Fatty acid composition and physiologi-cal properties of some filamentous blue-greenalgae. Arch. Mikrobiol. 83:216-236.
60. Khoja, T., and B. A. Whitton. 1971. Hetero-trophic growth of blue-green algae. Arch. Mik-robiol. 79:280-282.
61. Kitajima, E. W., and J. A. Lauritis. 1969. Plantvirions in plasmodesmata. Virology37:681-684.
62. Kiyohara, T., Y. Fujita, A. Hattori, and A.Watanabe. 1960. Heterotrophic culture of ablue-green alga, Tolypothrix tenuis I. J. Gen.Appl. Microbiol. 6:176-182.
63. Korant, B. D., and F. P. Pootjes. 1968. Bacterio-phage producing by autotrophically growingHydrogenomonas facilis. J. Virol. 2:87-88.
64. Kozjakov, S. Y., B. V. Gromov, and I. Y.Khudiakov. 1972. A-1 (L)-cyanophage of theblue-green alga Anabaena variabilis. Mikrobi-ologiya 41:555-559.
65. Kumar, H. D. 1970. Apparent genetic recombi-nation in a blue-green alga. Nature (London)196:1121-1122.
66. Kumar, H. D. 1963. Effects of radiations onblue-green algae. I. The production and char-acterization of a strain of Anacystis nidulans
367VOL. 37, 1973
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

PADAN AND SHILO
resistant to ultraviolet radiation. Ann. Bot.27:723-733.
67. Kumar, H. D. 1966. Some experiments on theinduction of nutritionally deficient mutants inthe blue-green alga, Anacystis nidulans. Ad-van. Front. Plant Sci. 14:133-142.
68. Kunisawa, R., and G. Cohen-Bazire. 1970. Mu-tations of Anacystis nidulans that affect celldivision. Arch. Mikrobiol. 71:49-59.
69. Kushner, D. J. 1969. Self-assembly of biologicalstructure. Bacteriol. Rev. 33:302-345.
70. Laemmli, U. K. 1970. Cleavage of structuralproteins during the assembly of the head ofbacteriophage T4. Nature (London)227:680-685.
71. Lang, N. J. 1968. The fine structure of blue-green algae. Annu. Rev. Microbiol. 22:15-46.
72. Lawson, R. H., and H. Hearon. 1970. Subcellularlocalization of chrysanthemum aspermy virusin tobacco and chrysanthemum leaf tissue.Virology 41:30-37.
73. Lazaroff, N., and W. Vishniac. 1964. The rela-tionship of cellular differentiation to colonialmorphogenesis of the blue-green alga, Nostocmuscorum A. J. Gen. Microbiol. 35:447-457.
74. Leach, C. K., and N. G. Carr. 1969. Oxidativephosphorylation in an extract of Anabaenavariabilis. Biochem. J. 112:125-126.
75. Leach, C. K., and N. G. Carr. 1970. Electrontransport and oxidative phosphorylation inthe blue-green alga Anabaena variabilis. J.Gen. Microbiol. 64:55-70.
76. Leach, C. K., J. M. Old, and N. G. Carr. 1971.Aspects of macromolecular synthesis in theblue-green alga Anabaena variabilis. J. Gen.Microbiol. 68:14.
77. Luftig, R., and R. Haselkorn. 1967. Morphologyof a virus of blue-green algae and properties ofits deoxyribonucleic acid. J. Virol. 1:344-361.
78. Luftig, R., and R. Haselkorn. 1968. Studies onthe structure of blue-green algae virus LPP-1.Virology 34::644-674.
79. Luftig, R., and R. Haselkorn. 1968. Comparisonof blue-green algae virus LPP-1 and the mor-phologically related viruses G111 and coliphageT7. Virology 34:675-678.
80. Luftig, R. B., W. B. Wood, and R. Okinaka.1971. Bacteriophage T4 head morphogenesison the nature of gene 49-defective heads andtheir role as intermediates. J. Mol. Biol.57:555-573.
81. Lund, J. W. G. 1965. Ecology of freshwaterphytoplankton. Biol. Rev. 40:231-293.
82. Luria, S. E., and J. E. Darnell, Jr. 1967. Generalvirology. John Wiley & Sons, Inc., New York.
83. Mackenzie, J. J., and R. Haselkorn. 1972. Physi-cal properties of blue-green algal virus SM-1and its DNA. Virology 49:497-504.
84. Mackenzie, J. J., and R. Haselkorn. 1972. Anelectron microscope study of infection by theblue-green algal virus SM-1. Virology49:505-516.
85. Mackenzie, J. J., and R. Haselkorn. 1972. Photo-synthesis and the development of blue-greenalgal virus SM-1. Virology 49:517-521.
86. Moody, M. F. 1965. The shape of the T-evenbacteriophage head. Virology 26:567-576.
87. Mukai, F., G. Streisinger, and B. Miller. 1967.The mechanism of lysis in phage T4-infectedcells. Virology 33:398-404.
88. Newton, J. W. 1960. Macromolecular variationin the chromatophores of the photosyntheticbacterium Rhodospirillum rubrum. Biochim.Biophys. Acta 42:34-43.
89. Niblack, J. F., R. W. McDaniel, P. Killmer, andI. C. Gunsalus. 1966. Density heterogeneity inPseudomonas putida bacteriophage prepara-tions. J. Bacteriol. 92:1254-1255.
90. Owen, P. C. 1957. The effects of infection withtobacco mosaic virus on the photosynthesis oftobacco leaves. Ann. Appl. Biol. 45:456-461.
91. Padan, E., D. Ginzburg, and M. Shilo. 1970. Thereproductive cycle of cyanophage LPP1-G inPlectonema boryanum and its dependence onphotosynthetic and respiratory system. Virol-ogy 40:514-521.
92. Padan, E., B. Raboy, and M. Shilo. 1971.Endogenous dark respiration of the blue-greenalga, Plectonema boryanum. J. Bacteriol.106:45-50.
93. Padan, E., and M. Shilo. 1969. Distribution ofcyanophages in natural habitats. Verh. Int.Verein. Limnol. 17:747-751.
94. Padan, E., and M. Shilo. 1969. Short trichomemutant of Plectonema boryanum. J. Bacte-riol. 97:975-976.
95. Padan, E., M. Shilo, and N. Kislev. 1967.Isolation of "cyanophages" from freshwaterponds and their interaction with Plectonemaboryanum. Virology 32:234-246.
96. Padan, E., M. Shilo, and A. B. Oppenheim.1972. Lysogeny of the blue-green alga Plec-tonema boryanum by LPP2-SPI cyanophage.Virology 47:525-526.
97. Padan, E., A. Rimon, D. Ginzberg, and M.Shilo. 1971. A thermosensitive cyanophage(LPP1-G) attacking the blue-green alga Plec-tonema boryanum. Virology 45:773-776.
98. Pan, P., and W. W. Umbreit. 1972. Growth ofmixed cultures of autotrophic and hetero-trophic organisms. Can. J. Microbiol.18:153-156.
99. Pearce, J., and N. G. Carr. 1969. The incorpora-tion and metabolism of glucose by Anabaenavariabilis. J. Gen. Microbiol. 54:451-462.
100. Pearce, J., C. K. Leach, and N. G. Carr. 1969.The incomplete tricarboxylic acid cycle in theblue-green alga Anabaena variabilis. J. Gen.Microbiol. 55:371-378.
101. Pearce, J., and N. G. Carr. 1967. The metabo-lism of acetate by the blue-green algae Anaba-ena variabilis and Anacystis nidulans. J. Gen.Microbiol. 49:301-313.
102. Pikalek, P. 1967. Attempt to find genetic recom-bination in Anacystis nidulans. Nature (Lon-don) 215:666-667.
103. Rimon, A. 1971. The source of the LPPIGcyanophage proteins in the blue-green algaPlectonema boryanum and the study of therelations between events in the infected host
368 BACTrERIOL. REV.
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

CYANOPHAGES AFTACKING BLUE-GREEN ALGAE
and the multiplication of the phage. M. S.thesis, Hebrew University, Jerusalem.
104. Rubenchik, L. I., 0. I. Bershova, N. S. Novi-kova, and Z. P. Koptyeva. 1966. Lysis of blue-green alga Microcystis pulvera. Mikrobiol. Zh.Akad. Nauk. Ukr. RSR. 28:88-91.
105. Safferman, R. S. 1968. Virus diseases in blue-green algae. In Daniel F. Jackson (ed.), Algae,man and the environment. Syracuse Univer-sity Press, New York.
106. Safferman, R. S., T. 0. Diener, P. R. Desjardins,and M. E. Morris. 1972. Isolation and charac-terization of AS-1, a phycovirus infecting theblue-green algae Anacystis nidulans andSynechococcus cedrorum. Virology 47:105-113.
107. Safferman, R. S., and M. E. Morris. 1963. Algalvirus: isolation. Science 140:679-680.
108. Safferman, R. S., and M. E. Morris. 1964.Growth characteristics of the blue-green algalvirus LPP-1. J. Bacteriol. 88:771-775.
109. Safferman, R. S., and M. E. Morris. 1964.Control of algae with viruses. J. Amer. WaterWorks Ass. 56:1217-1224.
110. Safferman, R. S., and M. E. Morris. 1967.Observations on the occurrence, distribution,and seasonal incidence of blue-green algalviruses. Appl. Microbiol. 15:1219-1222.
111. Safferman, R. S., M. E. Morris, L. A. Sherman,and R. Haselkorn. 1969. Serological and elec-tron microscopic characterization of a newgroup of blue-green algal viruses (LPP-2).Virology 39:775-780.
112. Safferman, R. S., I. R. Schneider, R. L. Steere,M. E. Morris, and T. 0. Diener. 1969. Phyco-virus SM-1: a virus infecting unicellular blue-green algae. Virology 37:386-395.
113. Schneider, I. R., T. 0. Diener, and R. S. Saffer-man. 1964. Blue-green algal virus LPP-1:purification and partial characterization.Science 144:1127-1130.
114. Scholes, P., P. Mitchell, and J. Moyle. 1969. Thepolarity of proton translocation in some photo-synthetic microorganisms. Eur. J. Biochem.8:450-454.
115. Schopf, J. W. 1970. Precambrian microorganismsand evolutionary events prior to the origin ofvascular plants. Biol. Rev. 45:319-352.
116. Shane, M. S. 1971. Distribution of blue-greenalgal viruses in various types of natural wa-ters. Water Res. 5:711-716.
117. Sherman, L. A., and R. Haselkorn. 1970. LPP-1infection of the blue-green alga Plectonemaboryanum. I. Electron microscopy. J. Virol.6:820-833.
118. Sherman, L. A., and R. Haselkom. 1970. LPP-1infection of the blue-green alga Plectonemaboryanum. II. Viral deoxyribonucleic acid syn-thesis and host deoxyribonucleic acid break-down. J. Virol. 6:834-840.
119. Sherman, L. A., and R. Haselkorn. 1970. LPP-1infection of the blue-green alga Plectonemaboryanum. III. Protein synthesis. J. Virol.6:841-846.
120. Sherman, L. A., and R. Haselkorn. 1971. Growth
of the blue-green algae virus LPP-1 underconditions which impair photosynthesis. Vi-rology 45:739-746.
121. Shestakou, S. U., and N. T. Khyon. 1970.Evidence for genetic transformation in blue-green alga Anacystis nidulans. Mol. Gen.Genet. 107:372-375.
122. Shilo, M. 1967. Formation and mode of action ofalgal toxins. Bacteriol. Rev. 31:180-193.
123. Shilo, M. 1971. The ecology of cyanophages.Vehr. Int. Verein. Limnol. 21:(in press).
124. Shlesinger, M. 1932. Adsorption of bacterio-phages to homologous bacteria. II. Quantita-tive investigations of adsorption velocity andsaturation. In G. S. Stent (ed.), Papers onbacterial viruses. Little, Brown and Co., Bos-ton.
125. Siddiqi, M. S., L. M. Kozloff, F. W. Putnam,and E. A. Evans, Jr. 1952. Biochemical studiesof virus reproduction. IX. Nature of the hostcell contributions. J. Biol. Chem. 199:165-176.
126. Simon, L. D. 1972. Infection of Escherichia coliby T2 and T4 bacteriophages as seen in theelectron microscope: T4 head morphogenesis.Proc. Nat. Acad. Sci. U.S.A. 69:907-911.
127. Simon, R. D. 1971. Cyanophycin granules fromthe blue-green alga Anabaena cylindrica: areserve material consisting of copolymers ofaspartic acid and arginine. Proc. Nat. Acad.Sci. U.S.A. 68:265-267.
128. Singh, P. K. 1973. Occurrence and distributionof cyanophages in ponds, sewage and ricefields. Arch. Mikrobiol. 89:169-72.
129. Singh, R. N., and H. N. Singh. 1964. Ultra-violetinduced mutants of blue-green algae. II. Ana-baena doliolum Bharadwaja. Arch. Mikrobiol.48:118-121.
130. Singh, R. N., and P. K. Singh. 1967. Isolation ofcyanophages from India. Nature (London)216: 1020-1021.
131. Singh, R. N., P. K. Singh, A. K. Kashyap, T. A.Sarma, B. Dhar, I. J. Chaubey, and K. I. D.Choudhury. 1971. Isolation and characteriza-tion of new cyanophages and mutations ofLPP-1 and host alga Plectonema boryanum.12th Ann. Congr. Ass. Microbiol. India.
132. Singh, R. N., P. K. Singh, and P. K. Varanasi.1969. Lysogeny and induction of lysis in blue-green algae and their viruses, p. 272. Proc.56th Int. Sci. Congr.
133. Singh, R. N., and R. Sinha. 1965. Geneticrecombination in blue-green alga, Cylindros-permum majus Kuetz. Nature (London)207:782-783.
134. Smith, A. J., J. London, and R. Y. Stanier. 1967.Biochemical basis of obligate autotrophy inblue-green algae and thiobacilli. J. Bacteriol.94:972-983.
135. Smith, A. J., and C. Lucas. 1971. Propionateresistance in blue-green algae. Biochem. J.124:23.
136. Smith, G. 1950. Freshwater algae of the UnitedStates. McGraw-Hill Book Co. Inc., NewYork.
137. Smith, K. M., R. M. Brown, Jr., D. A. Goldstein,
369VOL. 37, 1973
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

PADAN AND SHILO
and P. L. Walne. 1966. Culture methods forthe blue-green alga Plectonema boryanumand its virus, with an electron microscopestudy of virus-infected cells. Virology28:580-591.
138. Smith, K. M., R. M. Brown, Jr., and P. L.Walne. 1967. Ultrastructural and time-lapsestudies on the replication cycle of the blue-green algal virus LPP-1. Virology 31:329-337.
139. Smith, K. M., R. M. Brown, Jr., P. L. Walne,and D. A. Goldstein. 1966. Electron micros-copy of the infection process of the blue-greenalga virus. Virology 30:182-192.
140. Sonenshein, A. L., and D. H. Roscoe. 1969. Thecourse of phage Oe infection in sporulatingcells of Bacillus subtilis strain 3610. Virology39:265-276.
141. Srivastava, B. S. 1969. Ultraviolet induced mu-tations to growth factor requirement and peni-cillin resistance in a blue-green alga. Arch.Mikrobiol. 66:234-238.
142. Stanier, R. Y., R. Kunisawa, M. Mandel, and G.Cohen-Bazire. 1971. Purification and proper-ties of unicellular blue-green algae. (orderChroococcales). Bacteriol. Rev. 35:171-205.
143. Stanier, R. Y., and C. B. Van-Niel. 1962. Theconcept of a bacterium. Arch. Mikrobiol.42:17-35.
144. Stevens, S. E., and C. Van Baalen. 1969. N-methyl-N-nitro-N-nitrosoguanidine as a mu-tagen for blue-green algae: evidence for repair.J. Phycol. 5:136-139.
145. Stevens, S. E., Jr., and C. Van Baalen. 1970.Growth characteristics of selected mutants ofa coccoid blue-green alga. Arch. Mikrobiol.72:1-8.
146. Summers, D. F., and J. V. Maizel. 1968. Evi-dence for large precursor proteins in poliovirussynthesis. Proc. Nat. Acad. Sci. U.S.A.59:966-971.
147. Susor, W. A., and D. W. Krogmann. 1964. Hillactivity in cell-free preparations of a blue-green alga. Biochim. Biophys. Acta 88:11-19.
148. Tagawa, K., H. Y. Tsujimoto, and D. I. Arnon.1963. Role of chloroplast ferredoxin in theenergy conversion process of photosynthesis.Proc. Nat. Acad. Sci. U.S.A. 49:567-572.
149. Takahashi, I. 1964. Incorporation of bacterio-phages genome by spores of Bacillus subtilis.J. Bacteriol. 87: 1499-1502.
150. Ueda, K. 1965. Virus-like structures in the cellsof the blue-green alga, Oscillatoria princeps.Exp. Cell Res. 40:671-673.
151. Van Baalen, C., D. S. Horre, and E. Brandt.1971. Heterotrophic growth of blue-greenalgae in dim light. J. Bacteriol. 105:685-689.
152. Walsby, A. E. 1970. The nuisance algae: curiosi-ties in the biology of planktonic blue-greenalgae. Water Treat. Exam., no. 4.
153. Watanabe, A., and Y. Yamamoto. 1967. Hetero-trophic nitrogen fixadtipm:by the blue-greenalga Anabaenopsis clfcularis. Nature (Lon-don) 214:738.
154. Weidel, W., and E. Kellenberger. 1955. The E.coli B receptor for the phage T,. II. Electronmicroscopic studies. Biochim. Biophys. Acta17:1-9.
155. Weppelman, R. M., and C. C. Brinton. 1971.The infection of Pseudomonas aeruginosa byRNA pilus phage PP7: the adsorption or-ganelle and the relationship between phagesensitivity and the division cycle. Virology44: 1-17.
156. Whitton, B. A., N. G. Carr, and I. W. Craig.1971. A comparison of the fine structure andnucleic acid biochemistry of chloroplasts andblue-green algae. Protoplasma 72:325-357.
157. Williams, R. E. O., and J. E. Rippon. 1953.Bacteriophage typing of strains of Staph-lyococcus aureus from various sources. Lan-cet 264(1):510-514.
158. Wolk, C. P. 1968. Movement of carbon fromvegetative cells to heterocysts in Anabaenacylindrica. J. Bacteriol. 96:2138-2143.
159. Wu, J. H., G. L. Choules, and R. A. Lewin. 1968.Early stages of the infection process in ablue-green algal virus system, as affected byKCN and light, p. 153-160. In Biochemicalregulation in diseased plants or injury. Phyto-pathological Society of Japan, Tokyo.
160. Wu, J. H., R. A. Lewin, and H. Werbin. 1967.Photoreactivation of UV irradiated blue-greenalgal virus LPP-1. Virology 31:657-664.
161. Wu, J. H., and P. M. Shugarman. 1967. Effect ofvirus infection on rate of photosynthesis andrespiration of a blue-green alga, Plectonemaboryanum. Virology 32:166-167.
162. Yehle, C. O., and R. H. Doi. 1967. Differentialexpression of bacteriophage genomes in vege-tative and sporulating cells of Bacillus sub-tilis. J. Virol. 1:935-947.
163. Zagury, D., P. Model, and G. D. Pappas. 1967. Asimplified technique for preparing thin sec-tions of bacterial and viral suspensions: ap-plication to Escherichia coli and bacterio-phage T2. Virology 33:347-352.
164. Zaitlin, M., and A. T. Jagendorf. 1960. Photo-synthetic phosphorylation and Hill reactionactivities of chloroplasts isolated from plantsinfected with tobacco mosaic virus. Virology12:477-486.
370 BACTERIOL. REV.
on Septem
ber 5, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from





























