Embed Size (px)
Citation preview

P1: SFK/UKS P2: SFK
BLBS102-c15 BLBS102-Simpson March 21, 2012 13:21 Trim: 276mm X 219mm Printer Name: Yet to Come
Part 3Meat, Poultry and Seafoods
285

P1: SFK/UKS P2: SFK
BLBS102-c15 BLBS102-Simpson March 21, 2012 13:21 Trim: 276mm X 219mm Printer Name: Yet to Come
15Biochemistry of Raw Meat and Poultry
Fidel Toldra and Milagro Reig
Background InformationStructure of MuscleMuscle Composition
Muscle ProteinsMyofibrillar ProteinsSarcoplasmic ProteinsConnective Tissue ProteinsEnzymes
Non-protein CompoundsFree Amino AcidsDipeptides
Muscle and Adipose Tissue LipidsTriacylglycerolsPhospholipids
Conversion of Muscle to MeatFactors Affecting Biochemical Characteristics
Effect of GeneticsGenetic TypeGenes
Incidence of Exudative MeatsEffect of the Age and SexEffect of the Type of Feed
Carcass ClassificationCurrent Grading SystemsNew Grading Systems
Physical TechniquesBiochemical Assay Techniques
Bioactive CompoundsReferences
Abstract: The consumers’ concern for safety and health is drivingthe consumption patterns in accordance to meat composition, espe-cially for some key substances. In addition, a significant percentageof meat is used as raw material for further processing into differentproducts such as cooked, fermented, and dry-cured meats and oneof the most relevant problems of the meat industry is that genet-ics, age and sex, intensive or extensive production systems, type offeeding and slaughter procedures including preslaughter handling,
stunning methods, and postmortem treatment have an influence onimportant biochemical traits with a direct effect on the meat qualityand its aptitude as raw material. All these factors are described inthis chapter.
BACKGROUND INFORMATIONIn recent years, there has been a decline in consumption of beef,accompanied by a slight increase in pork and an increased de-mand for chicken. There are many reasons behind these changesincluding the consumer concern for safety and health, changesin demographic characteristics, changes in consumer lifestyles,availability and convenience, price, and so on (Resurreccion2003). The quality perception of beef changes depending on thecountry. So, Americans are concerned with cholesterol, caloriecontent, artificial ingredients, convenience characteristics, andprice (Resurreccion 2003). In the United States, this changingdemand has influenced the other meat markets. Beef has beengradually losing market share to pork and, especially, chicken(Grunert 1997).
But a significant percentage of meat is used as raw material forfurther processing into different products such as cooked, fer-mented, and dry-cured meats. Some of the most well-knownmeat products are bacon, cooked ham, fermented and dry-fermented sausages, and dry-cured ham (Flores and Toldra 1993,Toldra 2002).
The processing meat industry faces various problems, but oneof the most important is the variability in the quality of meatas a raw material. Genetics, age and sex, intensive or extensiveproduction systems, type of feeding and slaughter proceduresincluding pre-slaughter handling, stunning methods, and post-mortem treatment have an influence on important biochemicaltraits with a direct effect on the meat quality and its aptitude asraw material.
Food Biochemistry and Food Processing, Second Edition. Edited by Benjamin K. Simpson, Leo M.L. Nollet, Fidel Toldra, Soottawat Benjakul, Gopinadhan Paliyath and Y.H. Hui.C© 2012 John Wiley & Sons, Inc. Published 2012 by John Wiley & Sons, Inc.
287

P1: SFK/UKS P2: SFK
BLBS102-c15 BLBS102-Simpson March 21, 2012 13:21 Trim: 276mm X 219mm Printer Name: Yet to Come
288 Part 3: Meat, Poultry and Seafoods
STRUCTURE OF MUSCLEA good knowledge of the structure of muscle is essential forthe use of muscle as meat. The structure is important for theproperties of the muscle, and its changes during postmortemevents influence the quality properties of the meat.
Muscle has different colors within the range of white tored, depending on the proportion of fibers. There are differentclassifications for fibers. On the basis of color, they can beclassed as red, white, or intermediate (Moody and Cassens1968): (1) red fibers are characterized by a higher content ofmyoglobin and higher numbers of capillaries and mitochondria,and they exhibit oxidative metabolism; (2) white fibers containlow amounts of myoglobin and exhibit glycolytic metabolism;and (3) intermediate fibers exhibit intermediate properties. Redmuscles contain a high proportion of red fibers and are mostlyrelated to locomotion, while white muscles contain a higherproportion of white fibers and are engaged in support tasks(Urich 1994). Other classifications are based on the speed ofcontraction (Pearson and Young 1989): Type I for slow-twitchoxidative fibers, type IIA for fast-twitch oxidative fibers, andIIB for fast-twitch glycolytic fibers.
The skeletal muscle contains a great number of fibers. Eachfiber, which is surrounded by connective tissue, contains around1000 myofibrils, all of them arranged in a parallel way and re-sponsible for contraction and relaxation. They are embedded ina liquid known as sarcoplasm, which contains the sarcoplas-mic (water-soluble) proteins. Each myofibril contains clear darklines, known as Z-lines, regularly located along the myofibril(see Fig. 15.1). The distance between two consecutive Z-lines is
Figure 15.1. Structure of a myofibril with details of main filaments inthe sarcomere.
known as a sarcomere. In addition, myofibrils contain thick andthin filaments, partly overlapped and giving rise to alternatingdark (A band) and light (I band) areas. The thin filaments extendinto the Z-line that serves as linkage between consecutive sar-comeres. All these filaments are composed of proteins known asmyofibrillar proteins.
Muscle, fat, bones, and skin constitute the main componentsof carcasses; muscle is the major compound and is, furthermore,associated to the term of meat. The average percentage of musclein relation to live weight varies depending on the species, degreeof fatness, and dressing method: 35% for beef, 32% for veal,36% for pork, 25% for lamb, 50% for turkey, and 39% forbroiler chicken. The muscle to bone ratio is also an importantparameter representative of muscling: 3.5 for beef, 2.1 for veal,4.0 for pork, 2.5 for lamb, 2.9 for turkey, and 1.8 for poultry(Kauffman 2001).
MUSCLE COMPOSITIONEssentially, meat is basically composed of water, protein, lipids,minerals, and carbohydrates. Lean muscle tissue contains ap-proximately 72–74% moisture, 20–22% protein, 3–5% fat, 1%ash, and 0.5% of carbohydrate. An example of typical porkmeat composition is shown in Table 15.1. These proportions arelargely variable, especially in the lipid content, which dependson species, amount of fattening, inclusion of the adipose tissue,and so on. There is an inverse relationship between the percent-ages of protein and moisture and the percentage of fat so that
Table 15.1. Example of the Approximate Compositionof Pork Muscle Longissimus Dorsi. Expressed asMeans and Standard Deviation (SD)
Units Mean SD
Gross compositionMoisture g/100g 74.5 0.5Protein g/100g 21.4 2.1Lipid g/100g 2.7 0.3Carbohydrate g/100g 0.5 0.1Ash g/100g 0.9 0.1
ProteinsMyofibrillar g/100g 9.5 0.8Sarcoplasmic g/100g 9.1 0.6Connective g/100g 3.0 0.4
LipidsPhospholipids g/100g 0.586 0.040Triglycerides g/100g 2.12 0.22Free fatty acids g/100g 0.025 0.005
Some minor compoundsCholesterol mg/100g 46.1 6.1Haem content mg/100g 400 30Dipeptides mg/100g 347.6 35.6Free amino acids mg/100g 90.2 5.8
Source:Aristoy and Toldra 1998, Hernandez et al. 1998, Toldra 1999,Unpublished.

P1: SFK/UKS P2: SFK
BLBS102-c15 BLBS102-Simpson March 21, 2012 13:21 Trim: 276mm X 219mm Printer Name: Yet to Come
15 Biochemistry of Raw Meat and Poultry 289
Table 15.2. Type, Localization and Main Role of Major Muscle Proteins
CellularLocalization Main Proteins Main Role in Muscle
Myofibrillar Myosin and actin Cytoskeletal proteins providing support to the myofibril andresponsible for contraction–relaxation of the muscle.
Tropomyosin Regulatory protein associated to troponins than cause its movementtoward the F-actin helix to permit contraction.
Troponins T, C, I Regulatory proteins. Tn-T binds to tropomyosin, Tn-C binds Ca2+and initiates the contractile process and Tn-I inhibits theactin–myosin interaction in conjunction with tropomyosin andTn-T and Tn-C.
Titin and nebulin Large proteins located between Z-lines and thin filaments. Theyprovide myofibrils with resistance and elasticity.
α-, β-, γ -, and eu-actinin Proteins regulating the physical state of actin. α-actinin acts as acementing substance in the Z-line. γ -actinin inhibits thepolymerization of actin at the nucleation step. β-actinin is locatedat the free end of actin filaments, preventing them to bind eachother. Eu-actinin interacts with both actin and α-actinin.
Filamin, synemin, vinculin, zeugmatin,Z-nin, C, H, X, F, I proteins
Proteins located in the Z-line that contribute to its high density.
Desmin Protein that links adjacent myofibrils through Z-lines.Myomesin, creatine kinase and
M-proteinProteins located in the center of the sarcomere forming part of the
M-line.Sarcoplasm Mitochondrial enzymes Enzymes involved in the respiratory chain.
Lysosomal enzymes Digestive hydrolases very active at acid pH (cathepsins, lipase,phospholipase, peptidases, glucohydrolases, etc.).
Other cytosolic enzymes Neutral proteases, lipases, glucohydrolases, ATPases, etc.Myoglobin Natural pigment of meat.Hemoglobin Protein present from remaining blood within the muscle.
Connectivetissue
Collagen Protein giving support, strength, and shape to the fibers,
Elastin Protein that gives elasticity to tissues like capillaries, nerves,tendons, etc.
Source: Bandman 1987, Pearson and Young 1989.
meats with high content of fat have lower content of moistureand proteins. A brief description of proteins and lipids as majorcomponents of meat is given later.
Muscle Proteins
Proteins constitute the major compounds in the muscle, approx-imately 15–22%, and have important roles for the structure, nor-mal function, and integrity of the muscle. Proteins experienceimportant changes during the conversion of muscle to meat thatmainly affect tenderness; and additional changes occur duringfurther processing, through the generation of peptides and freeamino acids as a result of the proteolytic enzymatic chain. Thereare three main groups of proteins in the muscle (see Table 15.2):myofibrillar proteins, sarcoplasmic proteins, and connective tis-sue proteins.
Myofibrillar Proteins
These proteins are responsible for the basic myofibrillar struc-ture, and thus they contribute to the continuity and strength of
the muscle fibers. They are soluble in high ionic strength buffers,and the most important of them are listed in Table 15.2. Myosinand actin are by far the most abundant and form part of the struc-tural backbone of the myofibril. Tropomyosin and troponins C,T, and I are considered as regulatory proteins because they playan important role in muscle contraction and relaxation (Pearson1987). There are many proteins in the Z-line region (althoughin a low percentage) that serve as bridges between the thin fil-aments of adjacent sarcomeres. Titin and nebulin are two verylarge proteins, present in a significant proportion, that are lo-cated in the void space between the filaments and the Z-lineand contribute to the integrity of the muscle cells (Robson et al.1997). Desmin is located on the external area of the Z-line andconnects adjacent myofibrils at the level of the Z-line.
Sarcoplasmic Proteins
These are water-soluble proteins, comprising about 30–35%of the total protein in muscle. Sarcoplasmic proteins containa high diversity of proteins (summarized in Table 15.2), mainly

P1: SFK/UKS P2: SFK
BLBS102-c15 BLBS102-Simpson March 21, 2012 13:21 Trim: 276mm X 219mm Printer Name: Yet to Come
290 Part 3: Meat, Poultry and Seafoods
metabolic enzymes (mitochondrial, lysosomal, microsomal, nu-clear, or free in the cytosol) and myoglobin. Some of these en-zymes play a very important role in postmortem meat and duringfurther processing, as described in Chapter 16. Minor amountsof hemoglobin may be found in the muscle if blood has not beendrained properly. Myoglobin is the main sarcoplasmic protein,responsible for the red color of meat as well as the typical pinkcolor of drippings. The amount of myoglobin depends on manyfactors. Red fibers contain higher amounts of myoglobin thanwhite fibers. The species is very important, and thus beef andlamb contain more myoglobin than pork and poultry. For a givenspecies, the myoglobin content in the muscle increases with theage of the animal.
Connective Tissue Proteins
Collagen, reticulin, and elastin constitute the main stromal pro-teins in connective tissue. There are several types (I–V) of colla-gen containing different polypeptide chains (up to ten α-chains).Type I collagen is the major component of the epimysium andperimysium that surround the muscles. Types III, IV, and V col-lagen are found in the endomysium, which provides support tothe muscle fiber (Eskin 1990). There are a high number of cross-linkages in the collagen fibers that increase with age, and thisis why meat tougher in older animals. Elastin is found in loweramounts, usually in capillaries, tendons, nerves, and ligaments.
Enzymes
Muscle contains a large number of enzymes, mainly peptidaseslike calpains, cathepsins, proteasome, tri- and dipeptidylpep-tidases, aminopeptidases, carboxypeptidases, and dipeptidases,but also lipases, glycohydrolase, nucleotidases, and so on (Toldra2007). All these enzymes have a special relevance for meat pro-cessing and they are covered in-depth in Chapter 16.
Non-protein Compounds
Free Amino Acids
The action of muscle aminopeptidases contributes to the gener-ation of free amino acids in living muscle. An example of thetypical content of free amino acids, at less than 45 minutes post-mortem, in glycolytic and oxidative porcine muscles is shownin Table 15.3. It can be observed that most of the amino acidsare present in significantly higher amounts in the oxidative mus-cle (Aristoy and Toldra 1998). The free amino acid content isrelatively low just postmortem, but it is substantially increasedduring postmortem storage due to the action of the proteolyticchain, which is very active and stable during meat aging.
Dipeptides
Muscle contains three natural dipeptides: carnosine (β-alanyl-l-histidine), anserine (β-alanyl-l-1-methylhistidine), and balenine(β-alanyl-l-3-methylhistidine). These dipeptides perform somephysiological functions in muscle, for example, as buffers, an-
Table 15.3. Example of the Composition in Free AminoAcids (Expressed as mg/100g of Muscle) of theGlycolytic Muscle Longissimus Dorsi and OxidativeMuscle Trapezius.
Amino Acids
MuscleLongissimus
dorsiMuscle
Trapezius
EssentialHistidine 2.90 4.12Threonine 2.86 4.30Valine 2.78 2.09Methionine 0.90 1.01Isoleucine 1.52 1.11Leucine 2.43 1.82Phenylalanine 1.51 1.25Lysine 2.57 0.22NonessentialAspartic acid 0.39 0.74Glutamic acid 2.03 5.97Serine 2.02 4.43Asparagine 0.91 1.63Glycine 6.01 12.48Glutamine 38.88 161.81Alanine 11.29 26.17Arginine 5.19 5.51Proline 2.83 4.45Tyrosine 2.11 1.63Ornithine 0.83 0.83
Source: Aristoy and Toldra 1998.
tioxidants, neurotransmitters, and modulators of enzyme action(Chan and Decker 1994, Gianelli et al. 2000). Dipeptide contentis especially higher in muscles with glycolytic metabolism (seeTable 15.4), but it varies with the animal species, age, and diet(Aristoy and Toldra 1998, Aristoy et al. 2004). Beef and porkhave a higher content of carnosine and are lower in anserine,lamb has similar amounts of carnosine and anserine, and poultryis very rich in anserine (see Table 15.3). Balenine is present inminor amounts in pork muscle but at very low concentrationsin other animal muscle, except in marine mammalians such asdolphins and whales (Aristoy and Toldra 2004).
Muscle and Adipose Tissue Lipids
Skeletal muscle contains a variable amount of lipids, between1% and 13%. Lipid content mainly depends on the degree offattening and the amount of adipose tissue. Lipids can be foundwithin the muscle (intramuscular), between muscles (intermus-cular), and in adipose tissue. Intramuscular lipids are mainlycomposed of triacylglycerols, which are stored in fat cells, andphospholipids, which are located in cell membranes. The amountof cholesterol in lean meat is around 50–70 mg/100 g. Intermus-cular and adipose tissue lipids are mainly composed of tria-cylglycerols and small amounts of cholesterol, around 40–60mg/100g (Toldra and Flores 2004).

P1: SFK/UKS P2: SFK
BLBS102-c15 BLBS102-Simpson March 21, 2012 13:21 Trim: 276mm X 219mm Printer Name: Yet to Come
15 Biochemistry of Raw Meat and Poultry 291
Table 15.4. Example of the Composition in Dipeptides(Expressed as mg/100g of Muscle) of the PorcineGlycolytic Muscle Longissimus Dorsi and OxidativeMuscle Trapezius.
Carnosine Anserine
Effect of muscle metabolismGlycolytic (muscle Longissimus dorsi) 313 14.6Oxidative (muscle. Trapezius) 181.0 10.7
Animal speciesPork (loin) 313.0 14.5Beef (top loin) 372.5 59.7Lamb (neck) 94.2 119.5Chicken (pectoral) 180.0 772.2
Source: Aristoy and Toldra 1998, Aristoy and Toldra 2004.
Triacylglycerols
Triacylglycerols are the major constituents of fat, as shown inTable 15.1. The fatty acid content mainly depends on age, pro-duction system, type of feed, and environment (Toldra et al.1996b). Monogastric animals such as swine and poultry tend toreflect the fatty acid composition of the feed in their fat. In thecase of ruminants, the nutrients and fatty acid composition aresomehow standardized due to biohydrogenation by the microbialpopulation of the rumen (Jakobsen 1999). The properties of thefat will depend on its fatty acid composition. A great percentageof the triacylglycerols are esterified to saturated and monounsat-urated fatty acids (see neutral muscle fraction and adipose tissue
data in Table 15.5). When triacylglycerols are rich in polyunsat-urated fatty acids (PUFA) such as linoleic and linolenic acids,fats tend to be softer and prone to oxidation. These fats mayeven have an oily appearance when kept at room temperature.
Phospholipids
These compounds are present in cell membranes, and althoughpresent in minor amounts (see Table 15.1), they have a strongrelevance to flavor development due to their relatively highproportion of PUFA (see polar fraction in Table 15.5). Ma-jor constituents are phosphatidylcholine (lecithin) and phos-phatidylethanolamine. The phospholipid content may vary de-pending on the genetic type of the animal and the anatomicallocation of the muscle (Hernandez et al. 1998, Armero et al.2002). For instance, red oxidative muscles have a higher amountof phospholipids than white glycolytic muscles.
CONVERSION OF MUSCLE TO MEATA great number of chemical and biochemical reactions take placein living muscle. Some of these reactions continue, while othersare altered due to changes in pH, the presence of inhibitory com-pounds, the release of ions into the sarcoplasm, and so on duringthe early postmortem time. In a few hours, these reactions areresponsible for the conversion of muscle to meat; this process isbasically schematized in Figure 15.2 and consists of the follow-ing steps: Once the animal is slaughtered, the blood circulationis stopped, and the importation of nutrients and the removal ofmetabolites to the muscle cease. This fact has very important
Table 15.5. Example of Fatty Acid Composition (Expressed as Percentage of Total Fatty Acids) of MuscleLongissimus Dorsi and Adipose Tissue in Pigs Feeded with a Highly Unsaturated Feed. Neutral and Polar Fractionsof Muscle Lipids are also Included
Fatty Acid TotalMuscleNeutral Polar
AdiposeTissue
Myristic acid (C 14:0) 1.55 1.97 0.32 1.40Palmitic acid (C 16:0) 25.10 26.19 22.10 23.78Stearic acid (C 18:0) 12.62 11.91 14.49 11.67Palmitoleic acid (C 16:1) 2.79 3.49 0.69 1.71Oleic acid (C 18:1) 36.47 42.35 11.45 31.64C 20:1 0.47 0.52 0.15 0.45Linoleic acid (C 18:2) 16.49 11.38 37.37 25.39C 20:2 0.49 0.43 0.66 0.78Linolenic acid (C 18:3) 1.14 1.17 0.97 2.64C 20:3 0.30 0.10 1.04 0.10Arachidonic acid (C 20:4) 2.18 0.25 9.83 0.19C 22:4 0.25 0.08 0.84 0.07Total SFA 39.42 40.23 37.03 37.02Total MUFA 39.74 46.36 12.26 33.81Total PUFA 20.84 13.41 50.70 29.17Ratio MUFA/SFA 1.01 1.15 0.33 0.91Ratio PUFA/SFA 0.53 0.33 1.37 0.79
SFA, saturated fatty acids; MUFA, monosaturated fatty acids; PUFA, polysaturated fatty acid.

P1: SFK/UKS P2: SFK
BLBS102-c15 BLBS102-Simpson March 21, 2012 13:21 Trim: 276mm X 219mm Printer Name: Yet to Come
292 Part 3: Meat, Poultry and Seafoods
Muscle
Slaughter: Blood circulation is stopped
Very fast decrease of oxygen concentration in the muscle
ppg
Lack of available oxygen
Redox potential decreasesdown to –50 mV Cell respiration stops Cease of the activity
of the mitochondrial system
Lactic acid is generated The enzymatic generation of ATP is reduced
Glycolysis
and accumulated
pH drops down to around 5.6 Actomyosin is formed
ATP consumptionEnzyme inhibition
Proteins are denaturated Decrease in water-binding capacity
Contraction
Reduction in red color
Release of water and soluble nutrients
Rigor mortis
Figure 15.2. Summary of main changes during conversion of muscle to meat.
and drastic consequences. The first consequence is the reduc-tion of the oxygen concentration within the muscle cell becausethe oxygen supply has stopped. An immediate consequence is areduction in mitochondrial activity and cell respiration (Pearson1987). Under normal aerobic values (see an example of restingmuscle in Fig. 15.3), the muscle is able to produce 12 moles ofadenosine triphosphate (ATP) per mole of glucose, and thus theATP content is kept around 5–8 µmol/g of muscle (Greser 1986).ATP constitutes the main source of energy for the contractionand relaxation of the muscle structures as well as other biochem-
muscleStressed muscleResting
GlycogenGlucoseBlood
12ATP(TCA cycle)
Glucose
2ATP(anaerobic)
O2
gy
Glycogen
Energy-requiring processesas creatin phosphatein
mitochondria
Energy-requiringprocesses
Lactic acid CO2
Blood
Figure 15.3. Comparison of energy generation between resting andstressed muscles.
ical reactions in postmortem muscle. As the redox potential isreduced toward anaerobic values, ATP generation is more costly.So, only 2 moles of ATP are produced per mole of glucose underanaerobic conditions (an example of a stressed muscle is shownin Fig. 15.3). The extent of anaerobic glycolysis depends on thereserves of glycogen in the muscle (Greaser 1986). Glycogenis converted to dextrins, maltose, and finally, glucose througha phosphorolytic pathway; glucose is then converted into lacticacid with the synthesis of 2 moles of ATP (Eskin 1990). In addi-tion, the enzyme creatine kinase may generate some additionalATP from adenosine diphosphate (ADP) and creatine phosphateat very early postmortem times, but only while creatine phos-phate remains. The contents of creatine have been reported tovary depending on the type of muscle (Mora et al. 2008). Themain steps in glycolysis are schematized in Figure 15.4.
The generation of ATP is strictly necessary in the muscle tosupply the required energy for muscle contraction and relaxationand to drive the sodium-potassium pump of the membranes andthe calcium pump in the sarcoplasmic reticulum. The initialsituation in postmortem muscle is rather similar to that in thestressed muscle, but with an important change: the absenceof blood circulation. Thus, there is a lack of nutrient supplyand waste removal (see Fig. 15.5). Initially, the ATP content inpostmortem muscle does not drop substantially because someATP may be formed from ceratin phosphate through the actionof the enzyme creatine kinase and through anaerobic glycolysis.As mentioned earlier, once creatine phosphate and glycogenare exhausted, ATP drops within a few hours to negligible
P1: SFK/UKS P2: SFK
BLBS102-c15 BLBS102-Simpson March 21, 2012 13:21 Trim: 276mm X 219mm Printer Name: Yet to Come
15 Biochemistry of Raw Meat and Poultry 293
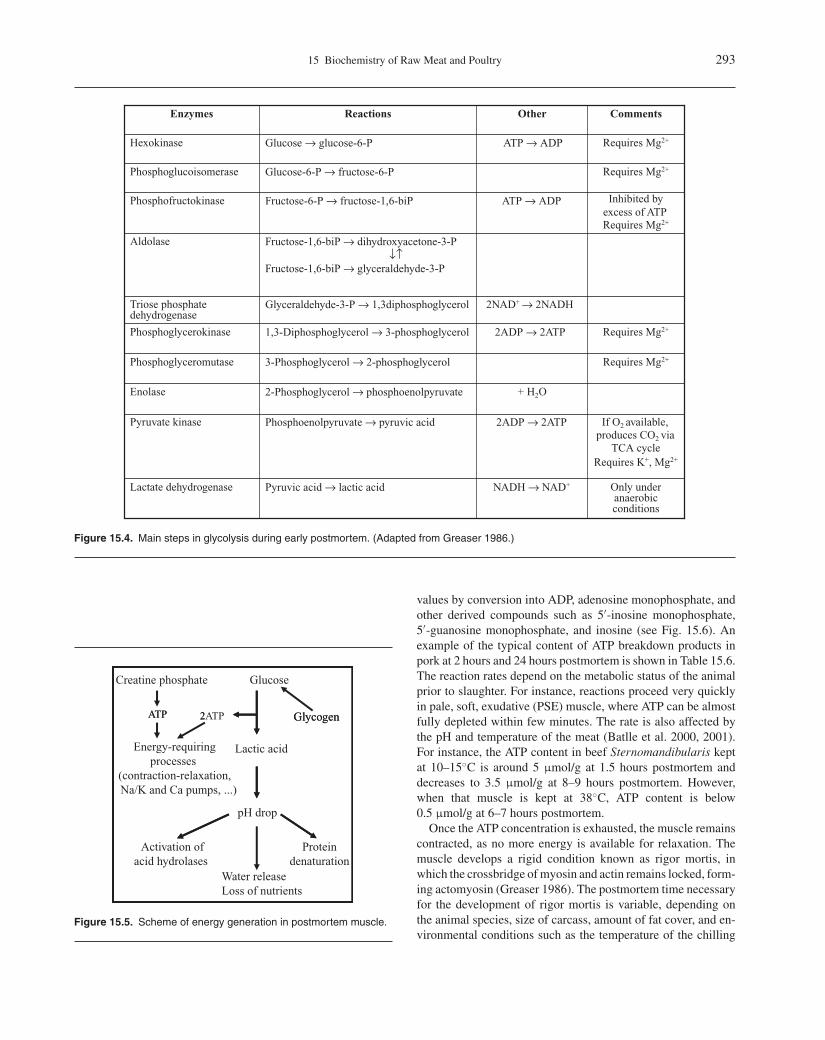
NADH → NAD+
2ADP → 2ATP
+ H2O
2ADP → 2ATP
2NAD+ → 2NADH
ATP → ADP
ATP → ADP
Other
Only under anaerobic conditions
Pyruvic acid → lactic acid Lactate dehydrogenase
If O2 available, produces CO2 via
TCA cycleRequires K+, Mg2+
Phosphoenolpyruvate → pyruvic acidPyruvate kinase
2-Phosphoglycerol → phosphoenolpyruvateEnolase
Requires Mg2+3-Phosphoglycerol → 2-phosphoglycerolPhosphoglyceromutase
Requires Mg2+1,3-Diphosphoglycerol → 3-phosphoglycerolPhosphoglycerokinase
Glyceraldehyde-3-P → 1,3diphosphoglycerolTriose phosphate dehydrogenase
Fructose-1,6-biP → dihydroxyacetone-3-P↓↑
Fructose-1,6-biP → glyceraldehyde-3-P
Aldolase
Inhibited by excess of ATPRequires Mg2+
Fructose-6-P → fructose-1,6-biPPhosphofructokinase
Requires Mg2+Glucose-6-P → fructose-6-PPhosphoglucoisomerase
Requires Mg2+Glucose → glucose-6-P Hexokinase
CommentsReactionsEnzymes
Figure 15.4. Main steps in glycolysis during early postmortem. (Adapted from Greaser 1986.)
Creatine phosphate Glucose
2ATP GlycogenATP
Lactic acid
2 Glycogen
Energy-requiringprocesses
ATP
pH drop
(contraction-relaxation,Na/K and Ca pumps, ...)
Proteindenaturation
Activation ofacid hydrolases
Water releaseLoss of nutrients
Figure 15.5. Scheme of energy generation in postmortem muscle.
values by conversion into ADP, adenosine monophosphate, andother derived compounds such as 5′-inosine monophosphate,5′-guanosine monophosphate, and inosine (see Fig. 15.6). Anexample of the typical content of ATP breakdown products inpork at 2 hours and 24 hours postmortem is shown in Table 15.6.The reaction rates depend on the metabolic status of the animalprior to slaughter. For instance, reactions proceed very quicklyin pale, soft, exudative (PSE) muscle, where ATP can be almostfully depleted within few minutes. The rate is also affected bythe pH and temperature of the meat (Batlle et al. 2000, 2001).For instance, the ATP content in beef Sternomandibularis keptat 10–15◦C is around 5 µmol/g at 1.5 hours postmortem anddecreases to 3.5 µmol/g at 8–9 hours postmortem. However,when that muscle is kept at 38◦C, ATP content is below0.5 µmol/g at 6–7 hours postmortem.
Once the ATP concentration is exhausted, the muscle remainscontracted, as no more energy is available for relaxation. Themuscle develops a rigid condition known as rigor mortis, inwhich the crossbridge of myosin and actin remains locked, form-ing actomyosin (Greaser 1986). The postmortem time necessaryfor the development of rigor mortis is variable, depending onthe animal species, size of carcass, amount of fat cover, and en-vironmental conditions such as the temperature of the chilling

P1: SFK/UKS P2: SFK
BLBS102-c15 BLBS102-Simpson March 21, 2012 13:21 Trim: 276mm X 219mm Printer Name: Yet to Come
294 Part 3: Meat, Poultry and Seafoods
Adenosine triphosphate (ATP)Creatine phosphateCreatin kinase
Adenosine diphosphate (ADP)
ATPase
Adenosine monophosphate (AMP)
Myokinase
Inosine monophosphate (IMP)Guanosine monophosphate (GMP)
AMP deaminase
Inosineguanosine
Hypoxanthine
Nucleoside phosphorylase
Figure 15.6. Main adenosine triphosphate (ATP) breakdown reactions in early postmortem muscle.
tunnel and the air velocity (see pork pieces after cutting in aslaughterhouse in Fig. 15.7). The rates of enzymatic reactionsare strongly affected by temperature. In this sense, the carcass-cooling rate will affect glycolysis rate, pH drop rate, and thetime course of rigor onset (Faustman 1994). The animal speciesand size of carcass have a great influence on the cooling rate
Table 15.6. Example of Nucleotides and NucleosidesContent (Expressed as µmol/g Muscle) in PorkPostmortem Muscle at 2 hours and 24 hours
Compound2 h Postmortem
Time24 h Postmortem
Time
ATP 4.39 −ADP 1.08 0.25AMP 0.14 0.20ITP + GTP 0.18 −IMP 0.62 6.80Inosine 0.15 1.30Hypoxanthine 0.05 0.32
Source: Batlle et al. 2001.ATP, adenosine triphosphate; ADP, adenosine diphosphate; AMP,adenosine monophosphate; ITP, inosine triphosphate; GMP,guanosine monophosphate; IMP, inosine monophosphate.
of the carcass. Furthermore, the location in the carcass is alsoimportant because surface muscles cool more rapidly than deepmuscles (Greaser 2001). So, when carcasses are kept at 15◦C,the time required for rigor mortis development may be about2–4 hours in poultry, 4–18 hours in pork, and 10–24 hours inbeef (Toldra 2006).
Muscle glycolytic enzymes hydrolyze the glucose to lacticacid, which is accumulated in the muscle because muscle wastesubstances cannot be eliminated due to the absence of bloodcirculation. This lactic acid accumulation produces a relativelyrapid (in a few hours) pH drop to values of about 5.6–5.8. The pHdrop rate depends on the glucose concentration, the temperatureof the muscle, and the metabolic status of the animal previous toslaughter. Water binding decreases with pH drop because of thechange in the protein’s charge. Then, some water is released outof the muscle as a drip loss (DL). The amount of released waterdepends on the extent and rate of pH drop. Soluble compoundssuch as sarcoplasmic proteins, peptides, free amino acids, nu-cleotides, nucleosides, B vitamins, and minerals may be partlylost in the drippings, affecting nutritional quality (Toldra andFlores 2004).
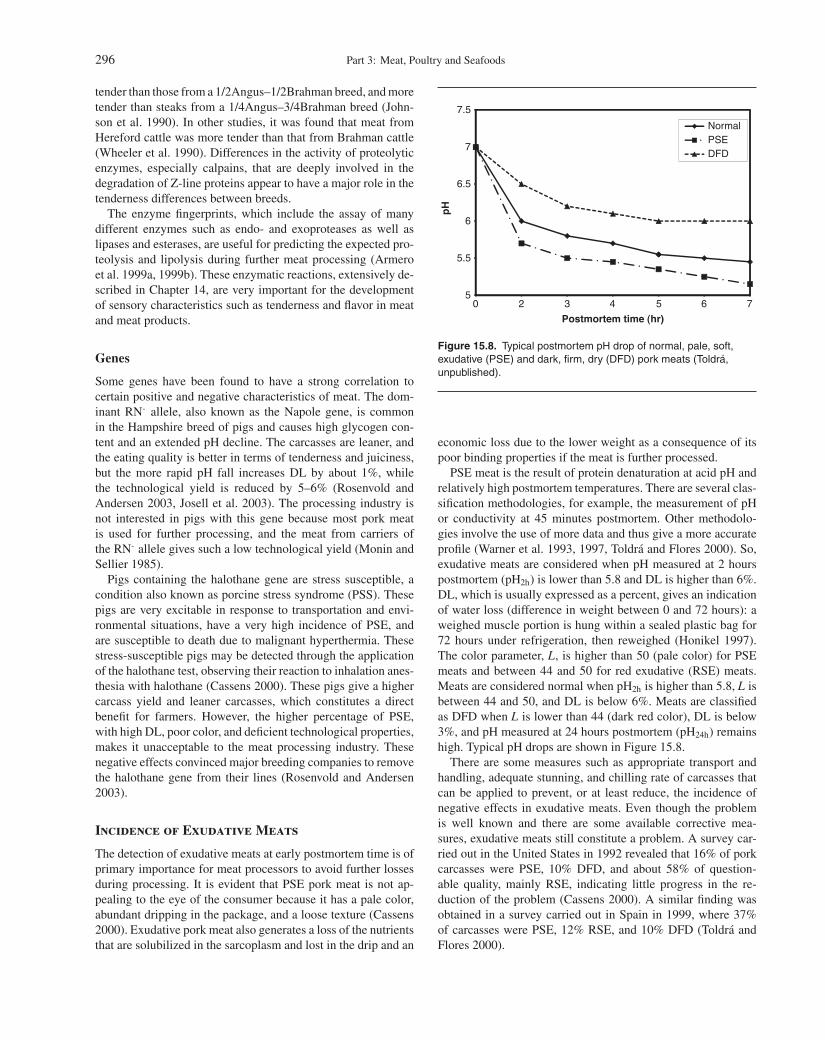
The pH drop during early postmortem has a great influenceon the quality of pork and poultry meats. The pH decrease isvery fast, below 5.8 after 2 hours postmortem, in muscles fromanimals with accelerated metabolism. This is the case of the PSEpork meats and red, soft, exudative pork meats. ATP breakdown

P1: SFK/UKS P2: SFK
BLBS102-c15 BLBS102-Simpson March 21, 2012 13:21 Trim: 276mm X 219mm Printer Name: Yet to Come
15 Biochemistry of Raw Meat and Poultry 295
Figure 15.7. Pork hams after cutting in a slaughterhouse, ready for submission to a processing plant. (Courtesy of Industrias CarnicasVaquero SA, Madrid, Spain.)
also proceeds very quickly in these types of meats, with almostfull ATP disappearance in less than 2 hours (Batlle et al. 2001).Red, firm, normal meat experiences a progressive pH drop downto values around 5.8–6.0 at 2 hours postmortem. In this meat,full ATP breakdown may take up to 8 hours. Finally, the dark,firm, dry pork meat (DFD) and dark cutting beef meat are pro-duced when the carbohydrates in the animal are exhausted frombefore slaughter, and thus almost no lactic acid can be generatedduring early postmortem due to the lack of a substrate. Very lowor almost negligible glycolysis is produced, and the pH remainshigh in these meats, which constitutes a risk from the microbio-logical point of view. These meats constitute a risk because theyare prone to contamination by foodborne pathogens and mustbe carefully processed, with extreme attention to good hygienicpractices.
Protein oxidation is another relevant change during post-mortem aging. Some amino acid residues may be convertedinto carbonyl derivatives and cause the formation of inter- andintraprotein disulfide links that can reduce the functionality ofproteins (Huff-Lonergan 2010).
FACTORS AFFECTING BIOCHEMICALCHARACTERISTICSEffect of Genetics
Genetic Type
The genetic type has an important relevance for quality, not onlydue to differences among breeds, but also to differences amonganimals within the same breed. Breeding strategies have been
focused toward increased growth rate and lean meat contentand decreased backfat thickness. Although grading traits arereally improved, poorer meat quality is sometimes obtained.Usually, large ranges are found for genetic correlations betweenproduction and meat quality traits, probably due to the reducednumber of samples when analyzing the full quality of meat, or toa large number of samples but with few determinations of qualityparameters. This variability makes it necessary to combine theresults from different research groups to obtain a full scope(Hovenier et al. 1992).
Current pig breeding schemes are usually based on a backcrossor on a three- or four-way cross. For instance, a common crossin the European Union is a three-way cross, where the sow is aLandrace × Large White (LR × LW) crossbreed. The terminalsire is chosen depending on the desired profitability per animal,and there is a wide range of possibilities. For instance, the Durocterminal sire grows faster and shows a better food conversionratio but accumulates an excess of fat; Belgian Landrace andPietrain are heavily muscled but have high susceptibility to stressand thus usually present a high percentage of exudative meats;or a combination of Belgian Landrace × Landrace gives goodconformation and meat quality (Toldra 2002).
Differences in tenderness between cattle breeds have also beenobserved. Brahman cattle is used extensively in the southwestof the United States since it is tolerant to adverse environmentalconditions but may give some tenderness issues (Brewer 2010).Even toughness was associated to an increased amount of calpas-tatin, the endogenous muscle calpain inhibitor (Ibrahim 2008).Studies have been performed for cattle breeding. For instance,after 10 days of aging, the steaks from an Angus breed were more
P1: SFK/UKS P2: SFK
BLBS102-c15 BLBS102-Simpson March 21, 2012 13:21 Trim: 276mm X 219mm Printer Name: Yet to Come
296 Part 3: Meat, Poultry and Seafoods
tender than those from a 1/2Angus–1/2Brahman breed, and moretender than steaks from a 1/4Angus–3/4Brahman breed (John-son et al. 1990). In other studies, it was found that meat fromHereford cattle was more tender than that from Brahman cattle(Wheeler et al. 1990). Differences in the activity of proteolyticenzymes, especially calpains, that are deeply involved in thedegradation of Z-line proteins appear to have a major role in thetenderness differences between breeds.
The enzyme fingerprints, which include the assay of manydifferent enzymes such as endo- and exoproteases as well aslipases and esterases, are useful for predicting the expected pro-teolysis and lipolysis during further meat processing (Armeroet al. 1999a, 1999b). These enzymatic reactions, extensively de-scribed in Chapter 14, are very important for the developmentof sensory characteristics such as tenderness and flavor in meatand meat products.
Genes
Some genes have been found to have a strong correlation tocertain positive and negative characteristics of meat. The dom-inant RN- allele, also known as the Napole gene, is commonin the Hampshire breed of pigs and causes high glycogen con-tent and an extended pH decline. The carcasses are leaner, andthe eating quality is better in terms of tenderness and juiciness,but the more rapid pH fall increases DL by about 1%, whilethe technological yield is reduced by 5–6% (Rosenvold andAndersen 2003, Josell et al. 2003). The processing industry isnot interested in pigs with this gene because most pork meatis used for further processing, and the meat from carriers ofthe RN- allele gives such a low technological yield (Monin andSellier 1985).
Pigs containing the halothane gene are stress susceptible, acondition also known as porcine stress syndrome (PSS). Thesepigs are very excitable in response to transportation and envi-ronmental situations, have a very high incidence of PSE, andare susceptible to death due to malignant hyperthermia. Thesestress-susceptible pigs may be detected through the applicationof the halothane test, observing their reaction to inhalation anes-thesia with halothane (Cassens 2000). These pigs give a highercarcass yield and leaner carcasses, which constitutes a directbenefit for farmers. However, the higher percentage of PSE,with high DL, poor color, and deficient technological properties,makes it unacceptable to the meat processing industry. Thesenegative effects convinced major breeding companies to removethe halothane gene from their lines (Rosenvold and Andersen2003).
Incidence of Exudative Meats
The detection of exudative meats at early postmortem time is ofprimary importance for meat processors to avoid further lossesduring processing. It is evident that PSE pork meat is not ap-pealing to the eye of the consumer because it has a pale color,abundant dripping in the package, and a loose texture (Cassens2000). Exudative pork meat also generates a loss of the nutrientsthat are solubilized in the sarcoplasm and lost in the drip and an
6
6.5
7
7.5
pH
NormalPSEDFD
5
5.5
7654320
Postmortem time (hr)
Figure 15.8. Typical postmortem pH drop of normal, pale, soft,exudative (PSE) and dark, firm, dry (DFD) pork meats (Toldra,unpublished).
economic loss due to the lower weight as a consequence of itspoor binding properties if the meat is further processed.
PSE meat is the result of protein denaturation at acid pH andrelatively high postmortem temperatures. There are several clas-sification methodologies, for example, the measurement of pHor conductivity at 45 minutes postmortem. Other methodolo-gies involve the use of more data and thus give a more accurateprofile (Warner et al. 1993, 1997, Toldra and Flores 2000). So,exudative meats are considered when pH measured at 2 hourspostmortem (pH2h) is lower than 5.8 and DL is higher than 6%.DL, which is usually expressed as a percent, gives an indicationof water loss (difference in weight between 0 and 72 hours): aweighed muscle portion is hung within a sealed plastic bag for72 hours under refrigeration, then reweighed (Honikel 1997).The color parameter, L, is higher than 50 (pale color) for PSEmeats and between 44 and 50 for red exudative (RSE) meats.Meats are considered normal when pH2h is higher than 5.8, L isbetween 44 and 50, and DL is below 6%. Meats are classifiedas DFD when L is lower than 44 (dark red color), DL is below3%, and pH measured at 24 hours postmortem (pH24h) remainshigh. Typical pH drops are shown in Figure 15.8.
There are some measures such as appropriate transport andhandling, adequate stunning, and chilling rate of carcasses thatcan be applied to prevent, or at least reduce, the incidence ofnegative effects in exudative meats. Even though the problemis well known and there are some available corrective mea-sures, exudative meats still constitute a problem. A survey car-ried out in the United States in 1992 revealed that 16% of porkcarcasses were PSE, 10% DFD, and about 58% of question-able quality, mainly RSE, indicating little progress in the re-duction of the problem (Cassens 2000). A similar finding wasobtained in a survey carried out in Spain in 1999, where 37%of carcasses were PSE, 12% RSE, and 10% DFD (Toldra andFlores 2000).

P1: SFK/UKS P2: SFK
BLBS102-c15 BLBS102-Simpson March 21, 2012 13:21 Trim: 276mm X 219mm Printer Name: Yet to Come
15 Biochemistry of Raw Meat and Poultry 297
Effect of the Age and Sex
The content in intramuscular fat content increases with the ageof the animal. In addition, the meat tends to be more flavor-ful and colorful, due to an increased concentration of volatilesand myoglobin, respectively (Armero et al. 1999a). Some of themuscle proteolytic and lipolytic enzymes are affected by age.Muscles from heavy pigs (11 months old) are characterized by agreater peptidase to proteinase ratio and a higher lipase, dipep-tidylpeptidase IV, and pyroglutamyl aminopeptidase activity. Onthe other hand, the enzyme activity in light pigs (7–8 monthsold) shows two groups. The larger one is higher in moisture con-tent and cathepsins B and B+L and low in peptidase activity,while the minor one is intermediate in cathepsin B activity andhigh in peptidase activity (Toldra et al. 1996a). In general, thereis a correlation between the moisture content and the activityof cathepsins B and B+L (Parolari et al. 1994). So, muscleswith higher moisture content show higher levels of cathepsinsB and B+L activity. This higher cathepsin activity may producean excess of proteolysis in processed meat products with longprocessing times (Toldra 2002).
A minor effect of sex is observed. Meats from barrows containmore fat than those from gilts. They present higher marbling,and the subcutaneous fat layer is thicker (Armero et al. 1999a).In the case of muscle enzymes, only very minor differences havebeen found. Sometimes, meats from entire males may give somesexual odor problems due to high contents of androstenone orescatol.
Effect of the Type of Feed
A great research effort has been exerted since the 1980s for themanipulation of the fatty acid composition of meat, to achievenutritional recommendations, especially an increase in the ra-tio between PUFA and saturated fatty acids (SFA) (PUFA:SFAratio). More recently, nutritionists recommend that PUFA com-position should be manipulated toward a lower n-6:n-3 ratio.
Fats with a higher content of PUFA have lower melting pointsthat affect the fat firmness. Softer fats may raise important prob-lems during processing if the integrity of the muscle is disruptedby any mechanical treatment (chopping, mincing, stuffing, etc.).The major troubles are related to oxidation and generation ofoff-flavors (rancid aromas) and color deterioration (trend towardyellowness in the fat) (Toldra and Flores 2004).
Pigs and poultry are monogastric animals that incorporate partof the dietary fatty acids practically unchanged into the adiposetissue and cellular membranes, where desaturation and chainelongation processes may occur (Toldra et al. 1996b, Jakobsen1999). The extent of incorporation may vary depending on thespecific fatty acid and the type of feed. Different types of cerealsas well as dietary oils and their effects on the proportions in fattyacid composition have been studied. The use of canola or linseedoils produces a substantial increase in the content of linolenicacid (C 18:3), which is an n-3 fatty acid (Jimenez-Colmeneroet al. 2006). In this way, the n-6:n-3 ratio can be reduced from 9to 5 (Enser et al. 2000). Other dietary oils such as soy, peanut,corn, and sunflower increase the content of linoleic acid (C
18:2), an n-6 fatty acid. Although it increases the total PUFAcontent, this fatty acid does not contribute to decrease the n-6:n-3 ratio, just the reverse. A similar trend is observed in the caseof poultry, where the feeds with a high content of linoleic acidsuch as grain, corn, plant seeds, or oils also increase the n-6:n-3ratio (Jakobsen 1999). As in the case of pork, the use of feedscontaining fish oils or algae, enriched in n-3 fatty acids such aseicosapentaenoic (C 22:5 n-3) and docosahexanoic (C 22:6 n-3)acids, can enrich the poultry meat in n-3 fatty acids and reducethe n-6:n-3 ratio from around 8.4 to 1.7 (Jakobsen 1999).
The main problem arises from oxidation during heating, be-cause some volatile compounds such as hexanal are typicallygenerated, producing rancid aromas (Larick et al. 1992). Therate and extent of oxidation of muscle foods mainly dependson the level of PUFA, but they are also influenced by earlypostmortem events such as pH drop, carcass temperature, ag-ing, and other factors. It must be pointed out that the increasedlinoleic acid content is replacing the oleic acid to a large ex-tent (Monahan et al. 1992). Feeds rich in saturated fats such astallow yield the highest levels of palmitic, palmitoleic, stearic,and oleic acids in pork loin (Morgan et al. 1992). Linoleic andlinolenic acid content may vary as much as 40% between theleanest and the fattest animals (Enser et al. 1988). An exampleof the effect of feed type on the fatty acid composition of sub-cutaneous adipose tissue of pigs is shown in Table 15.7. ThePUFA content is especially high in phospholipids, located insubcellular membranes such as mitochondria, microsomes, andso on, making them vulnerable to peroxidation because of theproximity of a range of pro-oxidants such as myoglobin, cy-tochromes, nonheme iron, and trace elements (Buckley et al.1995). Muscle contains several antioxidant systems, for exam-ple, those of superoxide dismutase and glutathione peroxidase,and ceruloplasmin and transferrin, although they are weakenedduring postmortem storage.
An alternative for effective protection against oxidation con-sists of the addition of natural antioxidants like vitamin E (alpha-tocopheryl acetate); this has constituted a common practice in thelast decade. This compound is added in the feed as an antioxidantand is accumulated by animals in tissues and subcellular struc-tures, including membranes, substantially increasing its effect.The concentration and time of supplementation are important.Usual levels are around 100–200 mg/kg in the feed for severalweeks prior to slaughter. The distribution of vitamin E in theorganism is variable, being higher in the muscles of the thoraciclimb, neck, and thorax and lower in the muscles of the pelvic limband back (O’Sullivan et al. 1997). Dietary supplementation withthis lipid-soluble antioxidant improves the oxidative stability ofthe meat. Color stability in beef, pork, and poultry is improvedby protection of myoglobin against oxidation (Houben et al.1998, Mercier et al. 1998). The water-holding capacity in porkis improved by protecting the membrane phospholipids againstoxidation (Cheah et al. 1995, Dirinck et al. 1996). The reductionin DL by vitamin E is observed even in frozen pork meat, uponthawing. Oxidation of membrane phospholipids causes a lossin membrane integrity and affects its function as a semiperme-able barrier. As a consequence, there is an increased passage ofsarcoplasmic fluid through the membrane, known as DL.

P1: SFK/UKS P2: SFK
BLBS102-c15 BLBS102-Simpson March 21, 2012 13:21 Trim: 276mm X 219mm Printer Name: Yet to Come
298 Part 3: Meat, Poultry and Seafoods
Table 15.7. Effect of Type of Feed on Total Fatty Acid Composition (Expressed as Percentage of Total Fatty Acids)of Pork muscle Lipids
Fatty Acid\Feed EnrichedBarley +
Soybean MealaSafflower
OilbTallowDietc
High OleicSunflower Oild
CanolaOile
Myristic acid (C 14:0) − − 1.37 0.05 1.6Palmitic acid (C 16:0) 23.86 27.82 24.15 6.35 20.6Stearic acid (C 18:0) 10.16 12.53 11.73 4.53 9.8Palmitoleic acid (C 16:1) 3 3.56 3.63 0.45 3.6Oleic acid (C 18:1) 39.06 37.81 46.22 71.70 45.9C 20:1 − 0.01 0.29 0.26Linoleic acid (C 18:2) 17.15 14.60 8.95 15.96 12.3C 20:2 − 0.01 0.44 − 0.4Linolenic acid (C 18:3) 0.91 0.01 0.26 0.71 3.0C 20:3 0.21 0.01 0.25 − 0.1Arachidonic acid (C 20:4) 4.26 2.14 2.13 − 0.74Eicosapentaenoic acid (C 22:5) 0.64 0.01 − − −Hexadecanoic acid (C22:6) 0.75 0.01 − − −Total SFA 34.02 40.35 37.83 10.93 33.6Total MUFA 42.06 42.38 50.26 72.41 49.5Total PUFA 23.92 16.79 11.91 16.67 16.6Ratio MUFA/SFA 1.24 1.05 1.33 6.62 1.47Ratio PUFA/SFA 0.70 0.42 0.32 1.53 0.49
Sources: aMorgan et al. (1992), bLarick et al. (1992), cLeszczynski et al. (1992), dRhee et al. (1988), eMiller et al. (1990).SFA, saturated fatty acids; MUFA, monosaturated fatty acids; PUFA, polysaturated fatty acid.
The fatty acid profile in ruminants is more saturated than inpigs, and thus the fat is firmer (Wood et al. 2003). The ma-nipulation of fatty acids in beef is more difficult due to therumen biohydrogenation. More than 90% of the PUFA are hy-drogenated, leaving a low margin for action to increase thePUFA:SFA ratio above 0.1. However, meats from ruminants arerich in conjugated linoleic acid (CLA), mainly 9-cis,11-trans-octadecadienoic acid, which exerts important health-promotingbiological activity (Belury 2002).
In general, a good level of nutrition increases the amount ofintramuscular fat. On the other hand, food deprivation may resultin an induced lipolysis that can be rapidly detected (in just 72hours) through a higher content of free fatty acids and monoa-cylglycerols, especially in glycolytic muscles (Fernandez et al.1995). Fasting within 12–15 hours preslaughter is usual to re-duce the risk of microbial cross-contamination during slaughter.
CARCASS CLASSIFICATIONCurrent Grading Systems
Meat grading constitutes a valuable tool for the classification ofa large number of carcasses into classes and grades with similarcharacteristics such as quality and yield. The final purpose isto evaluate specific characteristics to determine carcass retailvalue. In addition, the weight and category of the carcass areuseful for establishing the final price to be paid to the farmer.Carcasses are usually evaluated for conformation, carcass length,and back-fat thickness. The carcass yields vary depending onthe degree of fatness and the degree of muscling. The grade
is determined based on both degrees. The grading system is,thus, giving information on quality traits of the carcass that helpproducers, processors, retailers, and consumers.
Official grading systems are based on conformation, quality,and yield. Yield grades indicate the quantity of edible meat in acarcass. In the United States, beef carcasses receive a grade forquality (prime, choice, good, standard, commercial, utility, andcutter) and a grade for predicted yield of edible meat (1–5). Thereare four grades for pork carcasses (US No. 1 to US No. 4) basedon back-fat thickness and expected lean yield. The lean yield ispredicted by a combination of back-fat thickness measured atthe last rib and the subjective estimation of the muscling degree.In the case of poultry, there are three grades (A–C) based onthe bilateral symmetry of the sternum, the lateral convexity anddistal extension of the pectoral muscles, and fat cover on thepectoral muscles (Swatland 1994).
In Europe, beef, pork, and lamb carcasses are classified ac-cording to the EUROP scheme (Council regulation 1208/81,Commission directives 2930/81, 2137/92, and 461/93). TheseEuropean Union Directives are compulsory for all the mem-ber states. Carcass classification is based on the conformationaccording to profiles, muscle development, and fat level. Eachcarcass is classified by visual inspection, based on photos cor-responding to each grade (see Fig. 15.9). The six conformationclassification ratings are S (superior), E (excellent), U (verygood), R (good), O (fair), and P (poor). S represents the highestquality level and must not present any defect in the main pieces.The five classification ratings for fat level are 1 (low), 2 (slight),3 (average), 4 (high), and 5 (very high). The grading system foreach carcass consists in a letter for conformation, which is given
P1: SFK/UKS P2: SFK
BLBS102-c15 BLBS102-Simpson March 21, 2012 13:21 Trim: 276mm X 219mm Printer Name: Yet to Come
15 Biochemistry of Raw Meat and Poultry 299
Figure 15.9. Classification of pork carcasses in the slaughterhouse.(Courtesy of Industrias Carnicas Vaquero SA, Madrid, Spain.)
first, and a number for the fat level. In some countries, additionalgrading systems may be added. For instance, in France, the coloris measured and rated 1 to 4 from white to red color.
New Grading Systems
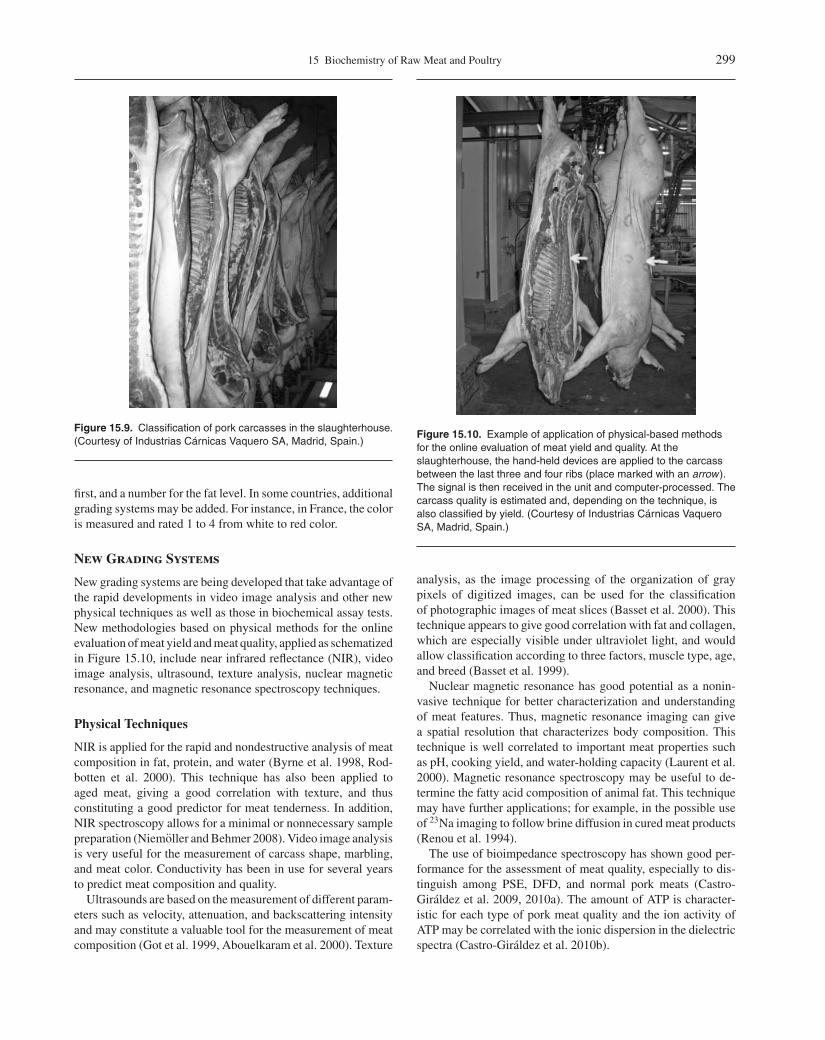
New grading systems are being developed that take advantage ofthe rapid developments in video image analysis and other newphysical techniques as well as those in biochemical assay tests.New methodologies based on physical methods for the onlineevaluation of meat yield and meat quality, applied as schematizedin Figure 15.10, include near infrared reflectance (NIR), videoimage analysis, ultrasound, texture analysis, nuclear magneticresonance, and magnetic resonance spectroscopy techniques.
Physical Techniques
NIR is applied for the rapid and nondestructive analysis of meatcomposition in fat, protein, and water (Byrne et al. 1998, Rod-botten et al. 2000). This technique has also been applied toaged meat, giving a good correlation with texture, and thusconstituting a good predictor for meat tenderness. In addition,NIR spectroscopy allows for a minimal or nonnecessary samplepreparation (Niemoller and Behmer 2008). Video image analysisis very useful for the measurement of carcass shape, marbling,and meat color. Conductivity has been in use for several yearsto predict meat composition and quality.
Ultrasounds are based on the measurement of different param-eters such as velocity, attenuation, and backscattering intensityand may constitute a valuable tool for the measurement of meatcomposition (Got et al. 1999, Abouelkaram et al. 2000). Texture
Figure 15.10. Example of application of physical-based methodsfor the online evaluation of meat yield and quality. At theslaughterhouse, the hand-held devices are applied to the carcassbetween the last three and four ribs (place marked with an arrow).The signal is then received in the unit and computer-processed. Thecarcass quality is estimated and, depending on the technique, isalso classified by yield. (Courtesy of Industrias Carnicas VaqueroSA, Madrid, Spain.)
analysis, as the image processing of the organization of graypixels of digitized images, can be used for the classificationof photographic images of meat slices (Basset et al. 2000). Thistechnique appears to give good correlation with fat and collagen,which are especially visible under ultraviolet light, and wouldallow classification according to three factors, muscle type, age,and breed (Basset et al. 1999).
Nuclear magnetic resonance has good potential as a nonin-vasive technique for better characterization and understandingof meat features. Thus, magnetic resonance imaging can givea spatial resolution that characterizes body composition. Thistechnique is well correlated to important meat properties suchas pH, cooking yield, and water-holding capacity (Laurent et al.2000). Magnetic resonance spectroscopy may be useful to de-termine the fatty acid composition of animal fat. This techniquemay have further applications; for example, in the possible useof 23Na imaging to follow brine diffusion in cured meat products(Renou et al. 1994).
The use of bioimpedance spectroscopy has shown good per-formance for the assessment of meat quality, especially to dis-tinguish among PSE, DFD, and normal pork meats (Castro-Giraldez et al. 2009, 2010a). The amount of ATP is character-istic for each type of pork meat quality and the ion activity ofATP may be correlated with the ionic dispersion in the dielectricspectra (Castro-Giraldez et al. 2010b).

P1: SFK/UKS P2: SFK
BLBS102-c15 BLBS102-Simpson March 21, 2012 13:21 Trim: 276mm X 219mm Printer Name: Yet to Come
300 Part 3: Meat, Poultry and Seafoods
3Mixing2
Additionof buffer
andenzyme
1Muscle sample
from carcass extraction
45Enzyme extract and substrate
added to
Incubationand
lecture in
6Computer
reaction buffer in multiwell plates
spectrofluorimeterrecording
Figure 15.11. Example of application of biochemical-basedmethods for evaluation of meat quality. A sample of the carcass(1) is mixed with buffer (2) for enzyme extraction andhomogenization (3). Enzyme extracts are placed in the wells of amultiwell plate and synthetic substrates, previously dissolved inreaction buffer, are added (4). The released fluorescence, which isproportional to the enzyme activity, is read by a multiwell platespectrofluorometer (5) and computer-recorded (6).
Biochemical Assay Techniques
Biochemical assay methodologies are based on biochemicalcompounds that can be used as markers of meat quality. Themode of operation is essentially schematized in Figure 15.11.Some of the most promising techniques include assay of pro-teolytic muscle enzymes and use of peptides as biochemicalmarkers.
The assay of certain proteolytic muscle enzymes such as cal-pain I, alanyl aminopeptidase, or dipeptidylpeptidase IV at just 2hours postmortem, has shown good ability to predict the water-holding capacity of the meat (Toldra and Flores 2000). Modifiedassay procedures, based on the use of synthetic fluorescent sub-strates, have been developed to allow relatively fast and simplemeasurements of enzyme activity with enough sensitivity.
Some peptides have been proposed as biochemical markersfor meat tenderness, which is particularly important in beef.These are peptides with molecular masses ranging from 1282 to5712 kDa, generated from sarcoplasmic and myofibrillar pro-teins (Stoeva et al. 2000). The isolation and identification ofthese peptides are tedious and time consuming, but once thefull sequence is known, Enzyme-linked immunosorbent assay(ELISA) test kits can be developed, and this would allow rapidassay and online detection at the slaughterhouse.
BIOACTIVE COMPOUNDSIt is well known that meat contains a large amount of proteinswith high biological value. However, in recent years, the bioac-tivities of certain compounds that are present in meat, eventhough in minor amounts, have received increased attention be-cause they may have an important nutritional role.
CLA is a group of isomers of octadecadienoic acid that isabundant in the fat of ruminants like cattle and sheep. The con-tent may change depending on the breed, feed, and age. CLA hasbeen reported to reduce the risk for certain types of cancer likethe colorectal cancer as well as having other activities like an-tiartherioesclerotic, antioxidative, and also playing certain rolein controlling obesity (Arihara and Motoko 2010).
Some of these substances are small peptides, which are presentin meat. Dipeptides carnosine and anserine are present in meatalthough their content varies depending on the animal speciesand the type of muscle metabolism (Aristoy et al. 2004, Aristoyand Toldra 2004, Mora et al. 2008). Both peptides have showngood resistance to proteases and a good antioxidant activity.Other bioactive peptides are derived from meat proteins after di-gestion. These peptides exert an antihypertensive effect throughthe inhibitory action on the angiotensin I converting enzyme. So,several bioactive peptides have been identified and sequencedafter simulated in vitro gastrointestinal digestion of pork meatproteins (Escudero et al. 2010a, 2010b).
In summary, there are many biochemical and chemical re-actions of interest in raw meats that contribute to importantchanges in meat and affect its quality. Most of these changes, asdescribed in this chapter, have an important role in defining theaptitude of the meat as raw material for further processing.
REFERENCES
Abouelkaram S et al. 2000. Effects of muscle texture on ultrasonicmeasurements. Food Chem 69: 447–455.
Arihara K, Motoko O. 2010. Functional meat products. In: F Toldra(ed.) Handbook of Meat Processing. Wiley-Blackwell, Ames, IA,pp. 423–439.
Aristoy MC, Toldra F. 1998. Concentration of free amino acids anddipeptides in porcine skeletal muscles with different oxidativepatterns. Meat Sci 50: 327–332.
Aristoy MC, Toldra F. 2004. Histidine dipeptides HPLC-based testfor the detection of mammalian origin proteins in feeds for rumi-nants. Meat Sci 67: 211–217.
Aristoy MC et al. 2004. A simple, fast and reliable methodologyfor the analysis of histidine dipeptides as markers of the presenceof animal origin proteins in feeds for ruminants. Food Chem 94:485–491
Armero E et al. 1999a. Effect of the terminal sire and sex onpork muscle cathepsin (B, B+L and H), cysteine proteinaseinhibitors and lipolytic enzyme activities. Meat Sci 51: 185–189.
Armero E et al. 1999b. Effects of pig sire types and sex on carcasstraits, meat quality and sensory quality of dry-cured ham. J SciFood Agric 79: 1147–1154.
Armero E et al. 2002. Lipid composition of pork muscle as affectedby sire genetic type. J Food Biochem 26: 91–102.
Bandman E. 1987. Chemistry of animal tissues. Part 1-Proteins.In: JF Price, BS Schweigert (eds.) The Science of Meat andMeat Products. Food and Nutrition Press, Inc, Trumbull, CT,pp. 61–102.
Basset O et al. 1999. Texture image analysis: application to theclassification of bovine muscles from meat slice images. OpticalEng 38: 1956–1959.

P1: SFK/UKS P2: SFK
BLBS102-c15 BLBS102-Simpson March 21, 2012 13:21 Trim: 276mm X 219mm Printer Name: Yet to Come
15 Biochemistry of Raw Meat and Poultry 301
Basset O et al. 2000. Application of texture image analysis for theclassification of bovine meat. Food Chem 69: 437–445.
Batlle N et al. 2000. Early postmortem detection of exudative porkmeat based on nucleotide content. J Food Sci 65: 413–416.
Batlle N et al. 2001. ATP metabolites during aging of exudative andnon exudative pork meats. J Food Sci 66: 68–71.
Belury, MA. 2002. Dietary conjugated linoleic acid in health: phys-iological effects and mechanisms of action. Ann Rev Nutr 22:505–531.
Brewer S. 2010. Technological quality of meat for processing. In:F Toldra (ed.) Handbook of Meat Processing. Wiley-Blackwell,Ames, IA, pp. 25–42.
Buckely DJ et al. 1995. Influence of dietary vitamin E on the oxida-tive stability and quality of pig meat. J Anim Sci 73: 3122–3130.
Byrne CE et al. 1998. Non-destructive prediction of selected qual-ity attributes of beef by near infrared reflectance spectroscopybetween 750 and 1098 nm. Meat Sci 49: 399–409.
Cassens RG. 2000. Historical perspectives and current aspects ofpork meat quality in the USA. Food Chem 69: 357–363.
Castro-Giraldez M et al. 2009. Physical sensors and techniques. In:LML Nollet, F Toldra (eds.) Handbook of Processed Meats andPoultry Analysis. CRC Press, Boca Raton, FL, pp. 7–34.
Castro-Giraldez M et al. 2010a. Physical sensors for quality controlduring processing. In: F Toldra (ed.) Handbook of Meat Process-ing. Wiley-Blackwell, Ames, IA, pp. 443–456.
Castro-Giraldez M et al. 2010b. Low-frequency dielectric spectrumto determine pork meat quality. Innov. Food Sci. Emerging Tech-nol 11: 376–386.
Chan KM, Decker EA. 1994. Endogenous skeletal muscle antioxi-dants. Crit Rev Food Sci Nutr 34: 403–426.
Cheah KS et al. 1995. Effect of dietary supplementation of vitaminE on pig meat quality. Meat Sci 39: 255–264.
Dirinck P et al. 1996. Studies on vitamin E and meat quality: 1. Effectof feeding high vitamin E levels on time-related pork quality. JAgric Food Chem 44: 65–68.
Enser M et al. 1998. Fatty acid content and composition of UK beefand lamb muscle in relation to production system and implica-tions for human nutrition. Meat Sci 49: 329–341.
Enser M et al. 2000. Feeding linseed to increase the n-3 PUFA ofpork: fatty acid composition of muscle, adipose tissue, liver andsausages. Meat Sci 55: 201–212.
Escudero E et al. 2010a. Angiotensin I converting enzyme inhibitorypeptides generated from in vitro gastrointestinal digestion of porkmeat. J Agric Food Chem 58: 2895–2901.
Escudero E et al. 2010b. Characterization of Peptides Released byin vitro Digestion of Pork Meat. J Agric Food Chem in press.
Eskin NAM. 1990. Biochemical changes in raw foods: meat andfish. In: Biochemistry of Foods, 2nd edn. Academic Press, SanDiego, pp. 3–68
Faustman LC. 1994. Postmortem changes in muscle foods. In: DMKinsman, AW Kotula, BC Breidenstein (eds.) Muscle Foods.Chapman and Hall, New York, pp. 63–78.
Fernandez X et al. 1995. Effect of muscle type and food deprivationfor 24 hours on the composition of the lipid fraction in musclesof large white pigs. Meat Sci 41: 335–343.
Flores J, Toldra F. 1993. Curing: processes and applications.In: R MacCrae et al. (eds.) Encyclopedia of Food Science,Food Technology and Nutrition. Academic Press, London,pp. 1277–1282.
Gianelli MP et al. 2000. Effect of carnosine, anserine and otherendogenous skeletal peptides on the activity of porcine mus-cle alanyl and arginyl aminopeptidases. J Food Biochem 24:69–78.
Got F et al. 1999. Effects of high intensity high frequency ultra-sound on aging rate, ultrastructure and some physico-chemicalproperties of beef. Meat Sci 51: 35–42.
Greaser ML. 1986. Conversion of muscle to meat. In: PJ Bechtel(ed.) Muscle as Food. Academic Press, Orlando, pp. 37–102.
Greaser ML. 2001. Postmortem muscle chemistry. In: YH Hui, et al.(eds.) Meat Science and Applications. Marcel Dekker, New York,pp. 21–37.
Grunert KG. 1997. What’s in a steak? A cross-cultural study on thequality perception of beef. Food Qual Pref 8: 157–174.
Hernandez P et al. 1998. Lipid composition and lipolytic enzymeactivities in porcine skeletal muscles with different oxidative pat-tern. Meat Sci 49: 1–10.
Honikel KO. 1997. Reference methods supported by OECD andtheir use in Mediterranean meat products. Food Chem 59:573–582.
Houben JH et al. 1998. Effect of the dietary supplementation withvitamin E on colour stability and lipid oxidation in packaged,minced pork. Meat Sci 48: 265–273.
Hovenier R et al. 1992. Genetic parameters of pig meat quality traitsin a halothane negative population. Meat Sci 32: 309–321.
Huff-Lonergan E. 2010. Chemistry and Biochemistry of meat. In:F Toldra, (ed.), Handbook of Meat Processing. Wiley-Blackwell,Ames, IA, pp 5–24.
Ibrahim RM et al. 2008. Effect of two dietary concentrate levels ontenderness, calpain and calpastatin activities and carcass merit inWaguli and Brahman sterrs. J Anim Sci 86: 1426–1433.
Jakobsen K. 1999. Dietary modifications of animal fats: status andfuture perspectives. Fett/Lipid 101: 475–483.
Jimenez Colmenero F et al. 2006. New approaches for the devel-opment of functional meat products. In: LML Nollet, F Toldra(eds.) Advanced technologies for meat processing. CRC Press,Boca Raton, FL, pp. 275–308.
Johnson MH et al. 1990. Differences in cathepsin B+L and calcium-dependent protease activities among breed type and their relation-ship to beef tenderness. J Anim Sci 68: 2371–2379.
Josell A et al. 2003. Sensory and meat quality traits of pork inrelation to post-slaughter treatment and RN genotype. Meat Sci66: 113–124.
Kauffman RG. 2001. Meat composition. In: YH Hui, WK Nip, RWRogers, OA Young, (eds.) Meat Science and Applications. MarcelDekker, New York, pp. 1–19.
Larick DK et al. 1992. Volatile compound contents and fatty acidcomposition of pork as influenced by linoleic acid content of thediet. J Anim Sci 70: 1397–1403.
Laurent W et al. 2000. Muscle characterization by NMR imagingand spectroscopic techniques. Food Chem 69: 419–426.
Leszczynski et al. 1992. Characterization of lipid in loin and baconfrom finishing pigs fed full-fat soybeans or tallow. J Anim Sci 70:2175–2181.
Mercier Y et al. 1998. Effect of dietary fat and vitamin E on colourstability and on lipid and protein oxidation in turkey meat duringstorage. Meat Sci 48: 301–318.
Miller MF et al. 1990. Determination of the alteration in fatty acidprofiles, sensory characteristics and carcass traits of swine fed

P1: SFK/UKS P2: SFK
BLBS102-c15 BLBS102-Simpson March 21, 2012 13:21 Trim: 276mm X 219mm Printer Name: Yet to Come
302 Part 3: Meat, Poultry and Seafoods
elevated levels of monounsaturated fats in the diet. J Anim Sci68: 1624–1631.
Monahan FJ et al. 1992. Influence of dietary vitamin E (alphatocopherol) on the color stability of pork chops. Proc 38thInt Congress of Meat Science and Technology, pp. 543–544,Clermont-Ferrand, France, August 1992.
Monin G, Sellier P. 1985. Pork of low technological quality witha normal rate of muscle pH fall in the immediate post-mortemperiod: the case of the Hampshire breed. Meat Sci 13: 49–63.
Moody WG, Cassens RG. 1968. Histochemical differentiation ofred and white muscle fibers. J Anim Sci 27: 961–966.
Mora L et al. 2008. Contents of creatine, creatinine and carno-sine in pork muscles of different metabolic type. Meat Sci 79:709–715.
Morgan CA et al. 1992. Manipulation of the fatty acid compositionof pig meat lipids by dietary means. J Sci Food Agric 58: 357–368.
Niemoller A, Behmer D. 2008. Use of near infrared spectroscopyin the food industry. In J Irudayaraj, C Reh (eds.) Nondestruc-tive Testing of Food Quality. Blackwell Publishing, Ames, IA,pp. 67–118.
O’Sullivan MG. 1997. The distribution of dietary vitamin E in themuscles of the porcine carcass. Meat Sci 45: 297–305.
Parolari G et al. 1994. Relationship between cathepsin B activityand compositional parameters in dry-cured ham of normal anddefective texture. Meat Sci 38: 117–122.
Pearson AM. 1987. Muscle function and postmortem changes. In:JF Price, BS Schweigert (eds.) The Science of Meat and MeatProducts. Food and Nutrition Press, Westport, CT, pp. 155–191.
Pearson AM, Young RB. 1989. Muscle and Meat Biochemistry.Academic Press, San Diego, pp. 1–261.
Renou JP et al. 1994. 23Na magnetic resonance imaging: distribu-tion of brine in muscle. Magn Reson Imaging 12: 131–137.
Resurreccion AVA. 2003. Sensory aspects of consumer choices formeat and meat products. Meat Sci 66: 11–20.
Rhee KS et al. 1988. Effect of dietary high-oleic sunflower oil onpork carcass traits and fatty acid profiles of raw tissues. Meat Sci24: 249–260.
Robson RM et al. 1997. Postmortem changes in the myofibrillarand other cytoskeletal proteins in muscle. Proceedings of the50th Annual Reciprocal Conference, Ames, IA. 50: 43–52.
Rodbotten R et al. 2000. Prediction of beef quality attributes fromearly post mortem near infrared reflectance spectra. Food Chem69: 427–436.
Rosenvold A, Andersen HJ. 2003. Factors of significance for porkquality: A review. Meat Sci 64: 219–237.
Stoeva S et al. 2000. Isolation and identification of proteolytic frag-ments from TCA soluble extracts of bovine M. Longissimus dorsi.Food Chem 69: 365–370.
Swatland HJ. 1994. Structure and Development of Meat Animalsand Poultry. Technomic Publishing, Lancaster, pp. 143–199.
Toldra F. 1992. The enzymology of dry-curing of meat products.In: FJM Smulders, et al. (eds.) New Technologies for Meat andMeat Products. Audet, Nijmegen, pp. 209–231.
Toldra F. 2002. Dry-Cured Meat Products. Food and Nutrition Press,Trumbull, CT, pp. 1–238.
Toldra F. 2006. Meat: chemistry and biochemistry. In: YH Hui et al.(eds.) Handbook of Food Science, Technology and Engineering,Vol 1. CRC Press, Boca Raton, FL, pp. 28–1 a 28–18.
Toldra F. 2007. Biochemistry of muscle and fat. In: F Toldra et al.(eds.) Handbook of Fermented Meat and Poultry. Blackwell Pub-lishing, Ames, IA, pp. 51–58.
Toldra F, Flores, M. 2000. The use of muscle enzymes as predictorsof pork meat quality. Food Chem 69: 387–395.
Toldra F, Flores M. 2004. Analysis of meat quality. In: LML Nollet(ed.) Handbook of Food Analysis. Marcel Dekker, New York,pp. 1961–1977.
Toldra F et al. 1996a. Pattern of muscle proteolytic and lipolyticenzymes from light and heavy pigs. J Sci Food Agric 71: 124–128.
Toldra F et al. 1996b. Lipids from pork meat as related to a healthydiet. Recent Res Devel Nutr 1: 79–86.
Urich K. 1994. Comparative Animal Biochemistry. Springer, Berlin,pp. 526–623.
Warner RD et al. 1993. Quality attributes of major porcine mus-cles: a comparison with the Longissimus lumborum. Meat Sci 33:359–372.
Warner RD et al. 1997. Muscle protein changes post mortem inrelation to pork quality traits. Meat Sci 45: 339–372.
Wheeler TL et al. 1990. Mechanisms associated with the variationin tenderness of meat from Brahman and Hereford cattle. J AnimSci 68: 4206–4220.
Wood JD et al. 2003. Effects of fatty acids on meat quality. MeatSci 66: 21–23.





























