Embed Size (px)
Citation preview

Foetal Regression in Rabbits; Experimental Studies of Histolysis and PhagocytosisAuthor(s): Megan HendersonSource: Proceedings of the Royal Society of London. Series B, Biological Sciences, Vol. 142, No.906 (Feb. 18, 1954), pp. 88-112Published by: The Royal SocietyStable URL: http://www.jstor.org/stable/82738 .
Accessed: 07/05/2014 13:58
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
The Royal Society is collaborating with JSTOR to digitize, preserve and extend access to Proceedings of theRoyal Society of London. Series B, Biological Sciences.
http://www.jstor.org
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions

Foetal regression in rabbits; experimental studies of
histolysis and phagocytosis
By Megan Henderson
Department of Zoology, University College of North Wales, Bangor
{Communicated by F. W. R. Brambell, F.R.S.?Received 27 May 1953?
Revised 7 August 1953)
[Plates 8 to 11]
The processes resulting in the removal of the embryos and the regeneration of the uterine tissues after foetal death are described in detail. The embryos were killed either by admini? stration of stilboestrol to the mother, or by direct mechanical damage, at ages of 11 days or 16 days or 19 to 20 days post-coitum. Specimens, at known intervals thereafter, were preserved and sectioned. Regression of the embryos in situ was followed until they had disappeared and the uterine tissues had been restored to normal. Sometimes this process of gradual removal was curtailed by abortion. The various tissues involved became necrotic in a definite sequence which was affected by the way in which the embryos were killed. During regression the uterine epithelium and submucosa became eroded antimesometrially and the decidua fragmented mesometrially, when all the embryos were dead. When any embryos survived the erosion of the antimesometrial uterine wall in the vicinity of the dead embryos did not occur and the fragmentation of the decidua was postponed until parturition. Marked leucocytosis of the uterine tissues and lumen occurred when all the embryos were dead, but such inflammatory changes were in abeyance when any embryos survived. The majority of these leucocytes appeared to be discharged gradually with the tissue debris through the vagina, but it was not possible to determine by examination alone whether any such leucocytes returned from the uterine lumen into the maternal system. The use of marker substances injected into the uterine lumen during resorption threw further light on the phago- cytic activity of the leucocytes and their subsequent movements. Leucocytes laden with marker were found in all the uterine tissues and congregated in lymphoid tissue situated antimesometrially within the serous coat of the uterus.
The possible significance of the retreat of laden leucocytes from a resorbing conceptus into the maternal system is discussed in relation to the problem of the sensitization of the mother to incompatible foetal antigens.
1. Introduction
Haemolytic disease of the new-born has drawn attention recently to the occurrence
of immunological incompatibility between mother and foetus. How the foetal
antigens reach the mother is a problem of great importance. During normal
gestation the primary function of the placenta is obviously to provide a physio?
logical barrier between mother and foetus and it is difficult to regard leakage
through it of antigens as other than accidental. It is of course possible that such
accidental leakage may occur sufficiently often and in adequate amounts to account
for the occurrence of immunization of the mother to the foetus. Nevertheless, the
processes of involution in the uterus that normally follow parturition, when foetal
tissue debris may be present in considerable quantity and still more those which
follow death of the embryo within the uterus and its regression in situ, would
appear to offer much more favourable opportunity for sensitization of the mother
to foetal antigens. Bruner, Hull & Doll (1948) consider that haemolytic jaundice
[ 88 ]
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions

Foetal regression in rabbits 89
in foals results from sensitization of the mare during pregnancy through the
absorption of the products of the breakdown of necrotic areas such as are often
found in the foetal membranes. The purpose of this investigation was to examine
the evidence that foetal tissue products may be absorbed into the maternal system
during resorption of dead foetuses. For this purpose the histological changes
following experimental destruction of the embryos in the uterus of rabbits have
been studied in detail. The material employed was obtained from a series of
experiments described elsewhere (Brambell, Henderson & Mills 1948) that were
designed to determine the time relations of the processes of retrogression and
abortion. These studies have been supplemented by a further series of experiments in which suspensions of inert particles of graphite and of carmine were injected into the uterus during resorption as markers of phagocytic activity.
Prenatal death and the removal of dead embryos by regression is well known
in many mammals. Studies of the process of regression of embryos which had
died in the uterus are, however, few in number. Meyer (1917) has described
regression in the guinea-pig, Fortuyn (1920) in the mouse, and Castle (1928),
dealing with a special case, in the rabbit. Fortuyn (1920) describes eosinophilous
phagocytes and giant cells as being the active elements in the removal of the
decidua during regression in the mouse, following natural foetal death. Kerr (1947), also working with the mouse, studied regression following experimental killing of
the embryos and found that the leucocytes may aid the breakdown of coherent
necrotic tissue and may be phagocytic. The order in which the various tissues
regress is significant, as has been shown by Long & Parkes (1924), who studied
the autolysis of embryos in vitro. Doubt still exists as to whether the process of
removal of dead embryos properly can be called reabsorption, as evidence of the
passage of embryonic material into the maternal tissue is lacking. As is well known retrogression in utero is not the only means of removing dead
embryos. Abortion results in the uterus being completely cleared, and is the usual
method of removal in monotocous animals, while in polytocous animals retro?
gression is more usual, thus avoiding the sacrifice of the whole litter. The dis?
tinction between resorption and abortion is not clear-cut and depends on the
extent to which autolysis of the foetal remains has progressed before the products are discharged from the uterus.
2. Material and technique
The material for ? 3 was obtained from experiments described elsewhere (Bram? bell et al. 1948) and is summarized in table 1. Details of the experimental technique need not be repeated here beyond stating that where the embryos were killed by
stilboestrol, a uniform dose of 5 mg. in olive oil was injected subcutaneously into
the mother. In the case of mechanical damage the uterus was exposed at laparotomy and the embryo pierced with a needle several times. Both methods were used at
16 and at 19 to 20 days post-coitum, and the first method only at 11 days post- coitum. At autopsy some of the conceptuses were opened under saline and the
remainder were left intact, the whole uterus being fixed in aqueous Bouin.
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions

90 Megan Henderson
Ehrlich's haematoxylin and aqueous eosin, Pasini's tricolour stain, Leishman's
stain and Giemsa's azure-eosin were used for staining the sections; of these
haematoxylin and eosin was found to be the most useful. Dekanski's (1949)
aqueous safranin was used for staining macrophages. Lendrum's method (1944) was used to stain eosinophil leucocytes. Carbol fuchsin and methylene blue were
used where bacterial infection was suspected.
time of
autopsy after
injection or
operation (days)
1 2 3
3+
Table 1
no. of days post-coitum at which stilboestrol injected
no. of days post-coitum at which
operated
11
E52/4L E53/1L E47/1R E54/2R E70/2R E71/1L E50/1R E49/1R E58/1R
E50/1R E60/1L
16
E66/1L E62/1L E48/2L
E64/1R E65/1L E67/4L E56/2L E63/1R E68/1R
19-20 16* 16 19-20
? E84/4L
E91/1R
W14/2R
E122/2L
E72/3L E57/1R
E85/1R
E107/2L 1R
E123/1R
E110/1L
E115/1L
E114/1L 1R
E120/1R E121/1R
10
11 12 13
E73/2R E69/1R E113/3R ?
E 92/4L 2L
E74/1L ? ? E98/2L E86/3R 2R
? ? ? ? E83/1L
? ? ? E82/2R E89/1R E81/1L
* Only some embryos in each litter killed.
The further experiments described in ?4 were also divided into two series in
which the embryos were killed by the means described above. Both methods were
used at 16 days post-coitum. The marker substance was injected into the uterus
at laparotomy 3 days after the death of the embryos. One or both the uterine
horns were used. Carmine, graphite and' Aquadag' were used as markers. Carmine
(Messrs Gurr) as specified by Carleton (1931) was finely ground and made up to
a 5% suspension in normal saline. The suspension was sterilized by boiling for
a few minutes. Graphite made up to a 5 % suspension was used in one experiment. Commercial cAquadag', a form of colloidal graphite, was used at a dilution of
1:10 parts of sterile normal saline. The quantities injected were 0*2 to 1 -0 ml./embryo. The injections were made at the tubal end, and the puncture in the uterine wall
was washed with saline. In one case where the embryos were killed surgically,
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions

Foetal regression in rabbits 91
subsequent leakage of carmine into the peritoneum was detected. There was little
risk of such leakage after killing with stilboestrol, since the only puncture in the
uterine wall was that made by the needle at the tubal end, and the conceptuses used for section were selected from the cervical end.
Vaginal smears were taken once or twice daily from one day after foetal death
until autopsy. The smears were stained with Ehrlich's haematoxylin and aqueous eosin.
The animals were killed 4 to 9 days after foetal death and the reproductive tract
fixed in 5 % formalin.
Sections were stained with Ehrlich's haematoxylin and either aqueous eosin or
light green. Sections of liver, spleen and mesenteric lymph glands, smears of bone
marrow and of lymph, and pieces of uterine mesentery were examined also.
Material employed is listed in table 2.
time of
autopsy after
injection or
operation (days)
4 5 5 6 7 7 7 8 8 9
no. of animal
360 618 226 219 248 158 448 813 276 246
Table 2
method of
killing stilboestrol stilboestrol
surgical stilboestrol stilboestrol
surgical surgical stilboestrol
surgical stilboestrol
marker
injected
graphite ' Aquadag'
carmine carmine carmine carmine carmine ' Aquadag'
carmine carmine
quantity of marker
(ml./embryo) ca. 0-7 ca. 0-2 ca. 0-5 ca. 0-3 ca. 1-0 ca. 0-3 ca. 1-0 ca. 0-2 ca. 1-0 ca. 1-0
3. Description of regression
Accounts of the embryology of the rabbit are available in the literature (Marshall
1893; Minot & Taylor 1905). Duval (1892) and Mossman (1926) have dealt with
the structure of the placenta in much detail.
Series I. Embryos killed by injection of stilboestrol at 11 days post-coitum
A section of a normal conceptus at 11 days post-coitum (figure 11) shows a com?
plete bilaminar omphalopleur, in contact with the uterine epithelium (b.o. and
u.e.). The yolk-sac splanchnopleur or dorsal wall of the yolk-sac is being pushed
ventralwards, so that the yolk-sac cavity (s.y.), which reaches its maximum size at
about 9j days, is decreasing. Uninucleate cells make up the bulk of the maternal
placenta, the former glandular region being occupied by multinucleate cells, which
separate the ectoplacenta (ex.) and uninucleate cell region? The most prominent
vessels seen are the sinus-like vessels in the uninucleate tissue, which are almost
completely filled with polymorphonuclear leucocytes. Radially arranged lobules
of the ectoplacenta are distinguishable together with the separating vascular villi
from the allantois. The embryo (e.), lying in a depression of the upper yolk-sac
wall, is approximately 8 mm. in length.
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions

92 Megan Henderson
Uterine wall
Throughout this series the embryos died at approximately an 11 J-day stage. The
first noticeable changes are in the bilaminar omphalopleur and uterine epithelium. As early as 24 h after the injection of stilboestrol the yolk-sac wall is necrotic and
the uterine epithelium is incomplete antimesometrially. The exposed submucosa
and giant cells soon become necrotic, the latter leaving naked nuclei. Frequently the uterine circular muscle is exposed to the uterine lumen following the break?
down and removal of the submucosa. At 3 to 3| days after injection destruction
of the uterine wall is at a maximum. This complete removal of the uterine epi? thelium is shown well in figure 13, plate 8, with the remains of the bilaminar
omphalopleur. Regeneration of the mucosa soon begins periplacentally, gradually
spreading antimesometrially. A small naked area in contact with the necrotic
decidua remains until approximately 7 days after injection, when the uterine
epithelium of the periplacental and antimesometrial regions is once more complete.
Mesometrially, the uterine epithelium is not completely regenerated until 10 days after injection. A uterus at this stage shows two well-defined mesometrial lobes,
separated from three small antimesometrial lobes by two blunt lobes. Numerous
small uterine glands are present. This uterus differs only from that typical of
oestrus, in that the mucosa is thrown into three lobes antimesometrially instead
of one.
Ectoplacenta
Necrosis of the ectoplacenta is evident at 3 days after injection, the changes which are then apparent being foreshadowed a little earlier. The boundaries of
the foetal vessels are indistinct, and the maternal surface sinuses are greatly
enlarged (figure 1). Enlargement of the surface sinuses normally occurs at the
12th day of gestation (Mossman 1926), but not to the extent noted here. Necrosis
of the trophoblastic partitions of the placenta, followed by their fragmentation, not only enlarges the surface sinuses, but also the deeper maternal vessels. The
foetal blood corpuscles fragment, forming deeply staining granules which can be
um.+f.
V.S.+/
Figtjbe 1. Area of ectoplacenta in transverse section 3 days after induction of resorption by stilboestrol at 11 days post-coitum. ( x 35.)
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions

Foetal regression in rabbits 93
distinguished in the allantoic mesoderm. The maternal vessels contain a fibrin
network. At 4 days after injection the two cotyledons of the ectoplacenta are
displaced so as to lie facing each other. Displacement is due to the decrease in
volume of the uterine conceptus, and to the almost complete detachment of the
ectoplacenta, following the breakdown of the multinucleate cells at 3J days after
injection. The ectoplacenta is almost in contact with the antimesometrial uterine
wall in some instances. Fragmentation follows, which may be complete at 5 days after injection, leaving naked decidual remains.
Decidua
Slight changes are apparent in the decidua concurrently with the destruction of
the uterine epithelium. Soon a slight breakdown of the multinucleate decidual
cells is noticeable in the periplacental position. A fibrin network intermingled with
the disintegrating tissue can be distinguished. By the second day after injection necrosis of the multinucleate decidual cells is more widespread, with clumping of
nuclei, and general discontinuity of tissues. Finally, at 3 days after injection,
yellow granules can be distinguished scattered throughout the necrotic mass,
indicating the remains of blood. Necrosis of the multinucleate decidual cells is so
advanced by now that fragmentation occurs in the periplacental region, causing cell debris to collect in the uterine lumen. Simultaneously the uninucleate decidual
cells become divided into two regions. One region, nearest the circular muscle,
appearing normal and the other region, in contact with the necrotic multinucleate
cells, contains crushed cells, with slightly pycnotic nuclei, suggesting a spreading of necrosis outwards from the multinucleate cell region. At 3 J days after injection three distinct regions can be recognized in the decidua. Figure 2 shows an outer
region of apparently normal, healthy uninucleate cells, a region of slightly necrotic
tissue still maintaining its continuity, and a fragmenting region of necrotic uni?
nucleate and multinucleate cells. Most of the vessels (v.c.) in the necrotic uninucleate
cell region are flattened in a direction perpendicular to that of the uterine circular
muscle, in contrast to the flattening in the general direction of the encircling uterine musculature of a normal uterus. The vessels in the healthy uninucleate
region are so crushed as to make it impossible to distinguish their lumina. This
flattening of the vessels is no doubt due to pressure on the decidual tissue resulting from the rapid decrease in size of the conceptus of which, at 3| days after injection, the placenta occupies approximately half the volume. The position of vessels in
the necrotic multinucleate cell region are marked by well-defined clots of fibrin.
As necrosis of the decidua spreads, fragmentation in the periplacental region
proceeds, and breaks become evident in the mass of multinuclear cells abutting on
the ectoplacenta. The ectoplacenta is completely removed by 5 days after injection, as mentioned previously, together with the multinucleate and some uninucleate
decidual tissue, thus exposing to the uterine lumen the remainder of the necrotic
decidua. The rapid decrease in volume of the conceptus has two effects. First the
remaining naked decidual tissue comes into contact with the antimesometrial uterine
wall, the uterine epithelium of which then disappears. Secondly, the ring of uterine
muscle is crenated, suggesting that the uneven shrinkage of the uterine content
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions
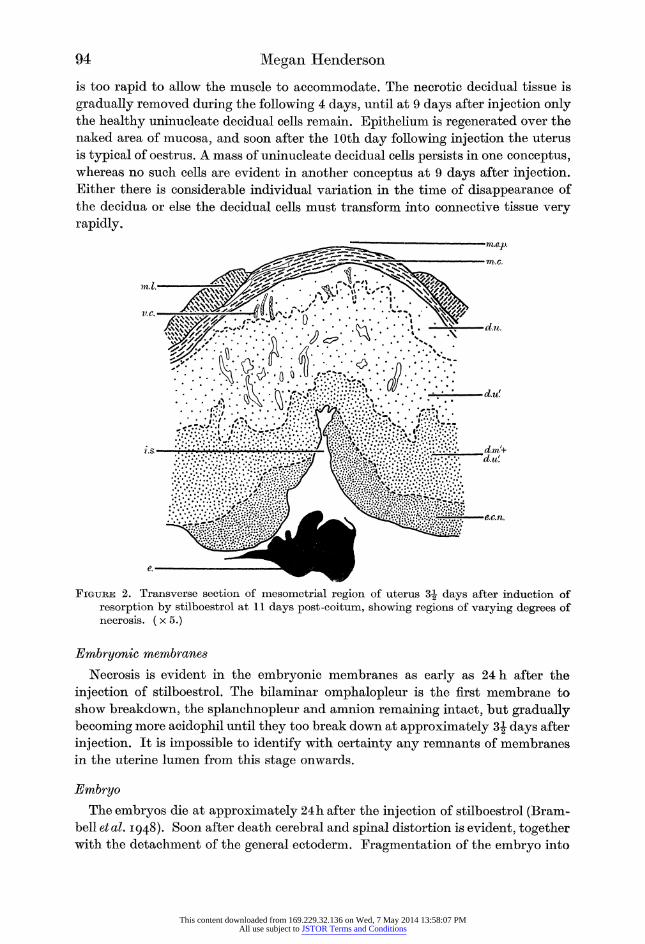
94 Megan Henderson
is too rapid to allow the muscle to accommodate. The necrotic decidual tissue is
gradually removed during the following 4 days, until at 9 days after injection only the healthy uninucleate decidual cells remain. Epithelium is regenerated over the
naked area of mucosa, and soon after the 10th day following injection the uterus
is typical of oestrus. A mass of uninucleate decidual cells persists in one conceptus, whereas no such cells are evident in another conceptus at 9 days after injection. Either there is considerable individual variation in the time of disappearance of
the decidua or else the decidual cells must transform into connective tissue very
rapidly.
^ '^<$?;>? ?")}?? ??? ?*?'?.()???:???'???
Figure 2. Transverse section of mesometrial region of uterus 3| days after induction of resorption by stilboestrol at 11 days post-coitum, showing regions of varying degrees of necrosis. (x 5.)
Embryonic membranes
Necrosis is evident in the embryonic membranes as early as 24 h after the
injection of stilboestrol. The bilaminar omphalopleur is the first membrane to
show breakdown, the splanchnopleur and amnion remaining intact, but gradually
becoming more acidophil until they too break down at approximately 3| days after
injection. It is impossible to identify with certainty any remnants of membranes
in the uterine lumen from this stage onwards.
Embryo
The embryos die at approximately 24h after the injection of stilboestrol (Bram? bell etal. 1948). Soon after death cerebral and spinal distortion is evident, together with the detachment of the general ectoderm. Fragmentation of the embryo into
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions

Foetal regression in rabbits 95
the amniotic cavity begins at 2 days after injection, the neural canal being obliterated previously by the disintegration of the spinal cord. Complete frag? mentation follows breakdown of the embryonic membranes by 3| days after
injection. The cell debris, which is highly acidophil, remains in the uterine lumen
until approximately 4 days after injection.
Leucocytosis
Leucocytes are present in the exposed submucosa of the antimesometrial uterine
wall at 2 days after injection, becoming very numerous as the epithelial destruction
increases. They are seen in the circular and longitudinal uterine muscles in some
cases. Figure 14, plate 8, shows a polymorphonuclear leucocyte passing through the endothelial lining of a uterine vessel and several other polymorphs in the
uterine circular muscle. Leucocytes appear first in the uterine lumen very soon
after the appearance of decidual cell debris in the periplacental region. They are
not detectable in the decidua, ectoplacenta or embryo while these are still in place. Numerous leucocytes appear in the debris when the embryo and ectoplacenta
fragment. They are abundant also in the naked margin of the decidua which is
then exposed. Throughout, leucocytes are present in the submucosa, being quite numerous even after the reformation of the uterine epithelium. The predominant
type, both in the tissue and free in the uterine lumen, undoubtedly is the poly?
morphonuclear leucocyte. Small numbers of eosinophil leucocytes are seen in the
submucosa, but are not detectable in the lumen. It should be pointed out that the
identification of leucocytes in the lumen is made difficult by the extremely acidophil nature of the pycnotic cell debris.
Summary
The antimesometrial uterine wall is the first region to show necrosis and frag? mentation of the tissue, similar changes occurring in the decidua, embryonic
membranes, embryo and ectoplacenta at a slightly later stage. The order in which
the tissues are given here corresponds with the order in which they fragment and
are removed, except for the decidua which begins to break down very early, but is
the last tissue to be completely removed. Polymorphonuclear leucocytes pass from the uterine tissue into the lumen, finally invading the necrotic decidua which
still remains attached to the uterine wall. Although polymorphs are present in the
submucosa after the reformation of the epithelium, there is no evidence to show
that they have returned there from the uterine lumen, as they may never have
left the uterine tissue.
Series II. Embryos killed by injection of stilboestrol at 16 days post-coitum
Figures 3 and 12 show the condition of the membranes and the placenta in
a normal 16-day conceptus. Comparison with figure 11 shows that the bilaminar
wall of the yolk-sac is absent, and the splanchnopleur (s.p.) is now almost in
contact with the uterine wall. The placenta shows three well-defined regions,
namely, a region of uninucleate decidual cells, a region of multinucleate decidual
cells often called the glycogenous cells, and the ectoplacenta. The uninucleate cells
are divided into two regions (not shown in figures 3 and 12), a narrow outer region
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions
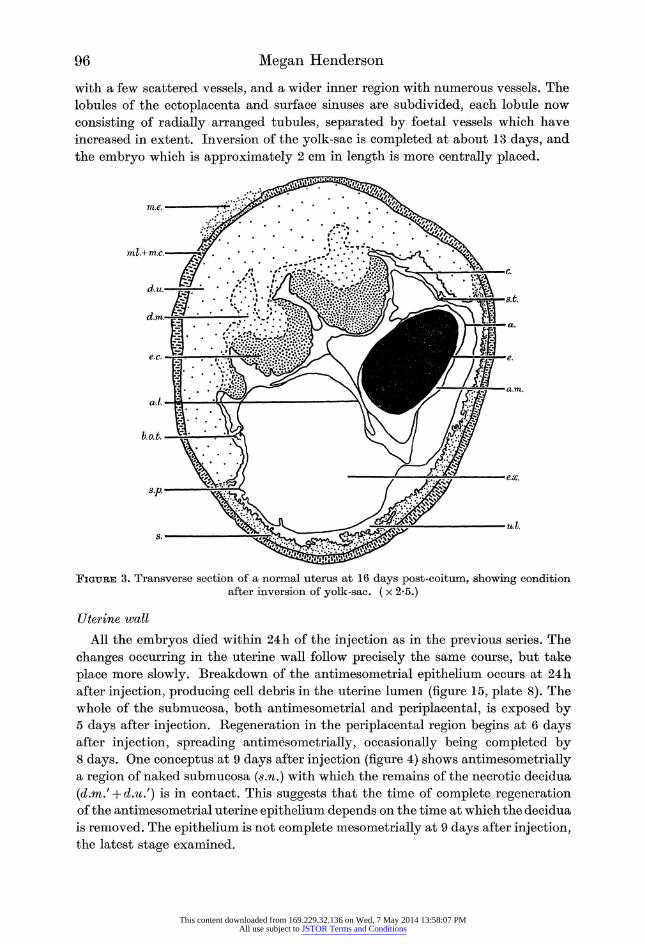
96 Megan Henderson
with a few scattered vessels, and a wider inner region with numerous vessels. The
lobules of the ectoplacenta and surface sinuses are subdivided, each lobule now
consisting of radially arranged tubules, separated by foetal vessels which have
increased in extent. Inversion of the yolk-sac is completed at about 13 days, and
the embryo which is approximately 2 cm in length is more centrally placed.
ml.+m.c.
b.o.t.
Figure 3. Transverse section of a normal uterus at 16 days post-coitum, showing condition after inversion of yolk-sac. ( x 2-5.)
Uterine wall
All the embryos died within 24h of the injection as in the previous series. The
changes occurring in the uterine wall follow precisely the same course, but take
place more slowly. Breakdown of the antimesometrial epithelium occurs at 24 h
after injection, producing cell debris in the uterine lumen (figure 15, plate 8). The
whole of the submucosa, both antimesometrial and periplacental, is exposed by 5 days after injection. Regeneration in the periplacental region begins at 6 days after injection, spreading antimesometrially, occasionally being completed by 8 days. One conceptus at 9 days after injection (figure 4) shows antimesometrially a region of naked submucosa (s.n.) with which the remains of the necrotic decidua
(d.m/ + d.u/) is in contact. This suggests that the time of complete regeneration of the antimesometrial uterine epithelium depends on the time at which the decidua
is removed. The epithelium is not complete mesometrially at 9 days after injection, the latest stage examined.
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions
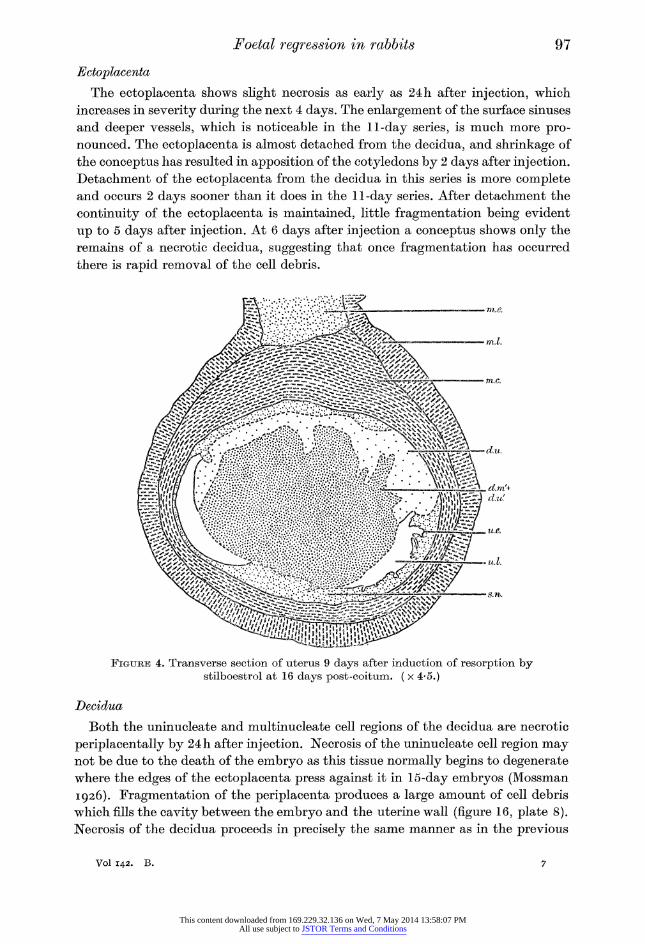
Foetal regression in rabbits 97
Ectoplacenta
The ectoplacenta shows slight necrosis as early as 24h after injection, which
increases in severity during the next 4 days. The enlargement of the surface sinuses
and deeper vessels, which is noticeable in the 11-day series, is much more pro? nounced. The ectoplacenta is almost detached from the decidua, and shrinkage of
the conceptus has resulted in apposition of the cotyledons by 2 days after injection. Detachment of the ectoplacenta from the decidua in this series is more complete and occurs 2 days sooner than it does in the 11-day series. After detachment the
continuity of the ectoplacenta is maintained, little fragmentation being evident
up to 5 days after injection. At 6 days after injection a conceptus shows only the
remains of a necrotic decidua, suggesting that once fragmentation has occurred
there is rapid removal of the cell debris.
{^\x+y>:^}t^ie?
Figure 4. Transverse section of uterus 9 days after induction of resorption by stilboestrol at 16 days post-coitum. ( x 4-5.)
Decidua
Both the uninucleate and multinucleate cell regions of the decidua are necrotic
periplacentally by 24 h after injection. Necrosis of the uninucleate cell region may not be due to the death of the embryo as this tissue normally begins to degenerate where the edges of the ectoplacenta press against it in 15-day embryos (Mossman
1926). Fragmentation of the periplacenta produces a large amount of cell debris
which fills the cavity between the embryo and the uterine wall (figure 16, plate 8). Necrosis of the decidua proceeds in precisely the same manner as in the previous
Vol 142. B.
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions

98 Megan Henderson
series, but degeneration of the multinucleate tissue is more rapid and the ecto?
placenta is detached at an earlier stage. Contraction of the conceptus produces
septa of crushed decidual tissue not noted before. Complete exposure of the
decidua, through detachment of the ectoplacenta, occurs at approximately 6 days after injection, and the necrotic decidual remains thereafter are gradually removed.
The time taken for this removal varies, as a conceptus at 8 days after injection shows little necrotic material, yet another at 9 days shows a large mass in contact
with the antimesometrial submucosa. Since no animals were killed later than 9 days after injection, a completely healed uterus was not obtained. Judging from the
condition at 8 and 9 days, the removal of the decidua and reformation of the
epithelium might be expected to be complete by approximately 13 days after
injection.
Embryonic membranes
The yolk-sac splanchnopleur and amnion rapidly become necrotic and the extra-
embryonic coelom almost completely obliterated by the close apposition of these
membranes (cf. figure 15, plate 8 and figure 3). This condition is evident at 1 day after injection and persists until 4 days, the membranes becoming more necrotic
and more closely applied to the embryo. Cell debris collects in the amnion and
extra-embryonic coelom, and the membranes rupture completely by 5 days after
injection.
Embryo
Immediately after the embryonic membranes have ruptured the embryo frag?
ments, filling the uterine lumen with cell debris. The changes that occur in the
embryo prior to fragmentation are similar to those occurring in the 11-day series, but extend over 5 days instead of 3 days. Distortion of the spinal cord is evident
1 day after injection (figure 15, plate 8). The early displacement of the ectoplacenta causes the embryo to be crushed against the antimesometrial uterine wall, a state
which does not arise in the previous series as the embryo has fragmented before
the displacement of the ectoplacenta. All trace of the embryo has disappeared from the uterus within a day of fragmentation, that is, by 6 days after injection.
Leucocytosis
Leucocytes are evident in the exposed submucosa at 24h after injection; then
they appear in the lumen amongst the cell debris and, finally, in the exposed decidua. In the previous series leucocytes are not detectable in the decidua while
the ectoplacenta remains in position, but in this series leucocytes are seen in the
periplacental decidual tissue at 3 days after injection (figure 16, plate 8) following
fragmentation of the multinucleate decidual cells at 2 days. Figure 17, plate 9, shows leucocytes within the amnion at 4 days after injection, together with some
embryonic cell debris. The amniotic membrane and yolk-sac splanchnopleur are
still recognizable but are very necrotic. Fragmentation of the membranes and
embryo occurs at 5 days. Up to 9 days after injection leucocytes are still evident
in the decidua and antimesometrial submucosa. Polymorphonuclear leucocytes far outnumber any other type, although eosinophil leucocytes and macrophages fl.rp rlftt,ftp,t,p,blft in small nnmWs
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions
Foetal regression in rabbits 99
Summary
Necrosis and fragmentation of the various tissues occur mainly in the same order
as in the previous series and leucocytes are similarly distributed. The chief
differences are the slower removal of the various tissues and the displacement of
the ectoplacenta before fragmentation of the embryo in the present series.
Series III. Embryos killed by injection of stilboestrol at
19 to 20 days post-coitum
The incompleteness of the antimesometrial uterine epithelium is similar to that
in the two previous series. All conceptuses examined show either a small area of
exposed submucosa (figure 7) or a region of regenerated epithelium devoid of
glands (figure 5). The order in which the tissues are broken down is similar also.
Leucocytes are evident as in the other series, figure 21, plate 9, showing poly?
morphonuclear leucocytes in the periplacental uterine wall. Polymorphs are again
predominant, although eosinophil leucocytes, macrophages and lymphocytes are
present also in the submucosa.
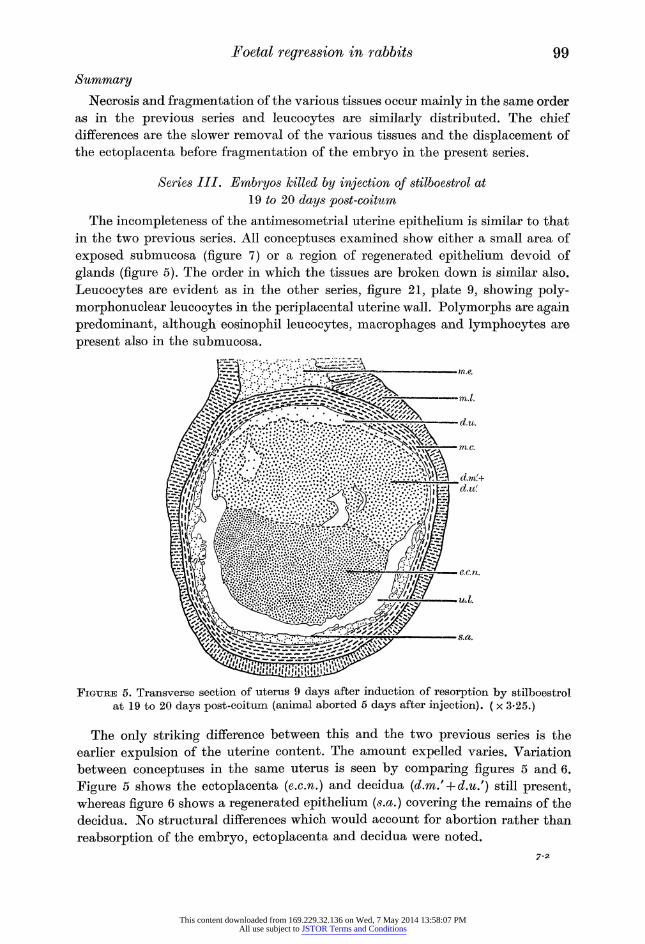
Figure 5. Transverse section of uterus 9 days after induction of resorption by stilboestrol at 19 to 20 days post-coitum (animal aborted 5 days after injection). ( x 3-25.)
The only striking difference between this and the two previous series is the
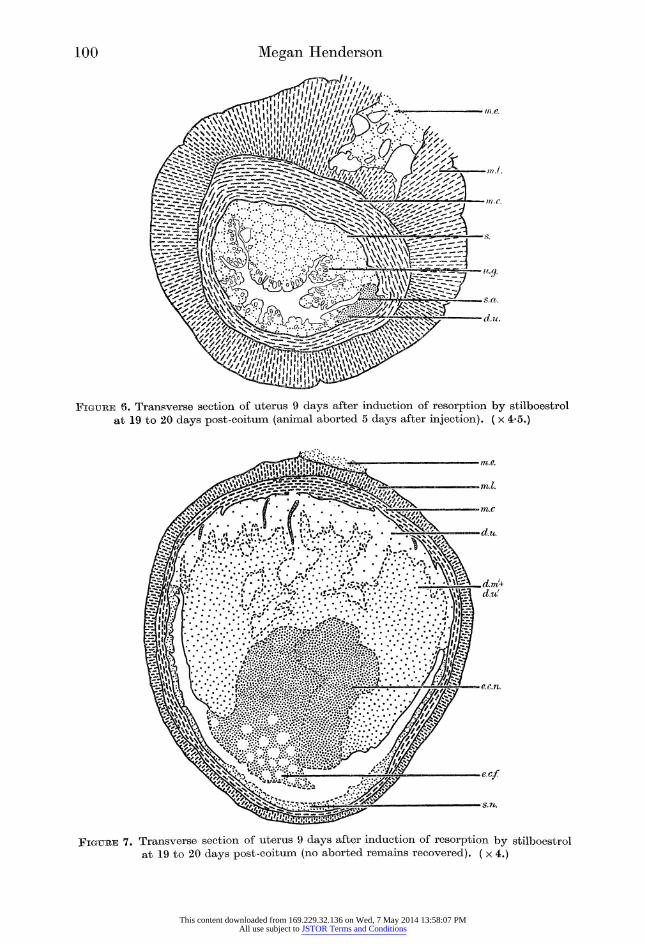
earlier expulsion of the uterine content. The amount expelled varies. Variation
between conceptuses in the same uterus is seen by comparing figures 5 and 6.
Figure 5 shows the ectoplacenta (e.c.n.) and decidua (d.m/ + d.u/) still present, whereas figure 6 shows a regenerated epithelium (s.a.) covering the remains of the
decidua. No structural differences which would account for abortion rather than
reabsorption of the embryo, ectoplacenta and decidua were noted.
7-a
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions
100 Megan Henderson
Figure 6. Transverse section of uterus 9 days after induction of resorption by stilboestrol at 19 to 20 days post-coitum (animal aborted 5 days after injection). ( x 4-5.)
Figure 7. Transverse section of uterus 9 days after induction of resorption by stilboestrol at 19 to 20 days post-coitum (no aborted remains recovered). ( x 4.)
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions
Foetal regression in rabbits 101
Series IV. Some but not all embryos killed surgically at
16 days post-coitum
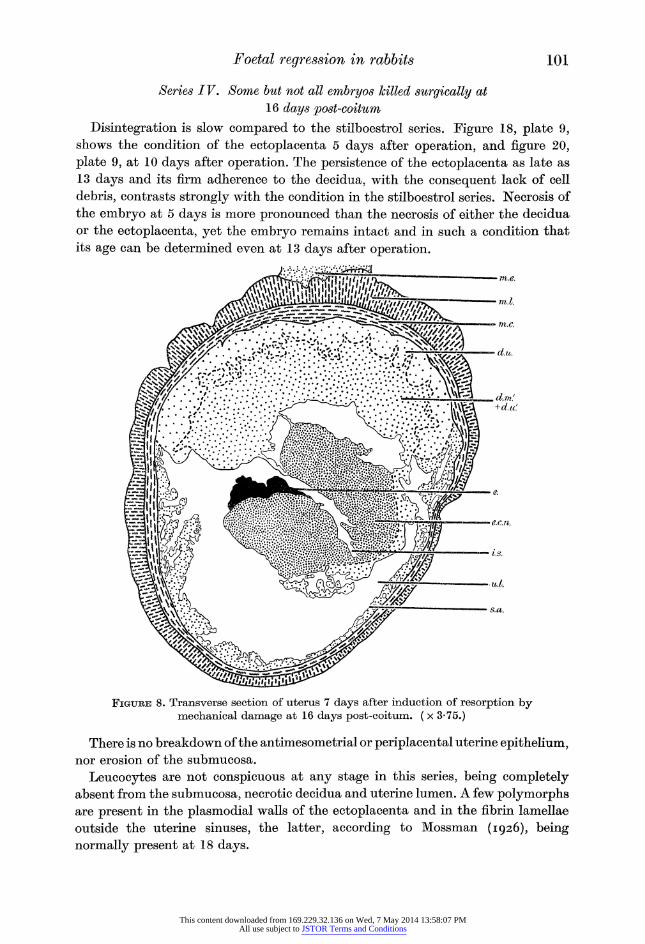
Disintegration is slow compared to the stilboestrol series. Figure 18, plate 9, shows the condition of the ectoplacenta 5 days after operation, and figure 20,
plate 9, at 10 days after operation. The persistence of the ectoplacenta as late as
13 days and its firm adherence to the decidua, with the consequent lack of cell
debris, contrasts strongly with the condition in the stilboestrol series. Necrosis of
the embryo at 5 days is more pronounced than the necrosis of either the decidua
or the ectoplacenta, yet the embryo remains intact and in such a condition that
its age can be determined even at 13 days after operation.
Figure 8. Transverse section of uterus 7 days after induction of resorption by mechanical damage at 16 days post-coitum. ( x 3-75.)
There is no breakdown of the antimesometrial or periplacental uterine epithelium, nor erosion of the submucosa.
Leucocytes are not conspicuous at any stage in this series, being completely absent from the submucosa, necrotic decidua and uterine lumen. A few polymorphs are present in the plasmodial walls of the ectoplacenta and in the fibrin lamellae
outside the uterine sinuses, the latter, according to Mossman (1926), being
normally present at 18 days.
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions
102 Megan Henderson
A few naturally occurring reabsorptions of odd embryos in otherwise healthy litters were available and closely resemble the experimentally induced reabsorp? tions in this series.
Series V. All embryos killed surgically at 16 days post-coitum
The process of disintegration up to 7 days follows the pattern of the previous
surgical series, but thereafter it is curtailed by abortion.
It is clear that disintegration is much slower than after treatment with
stilboestrol.
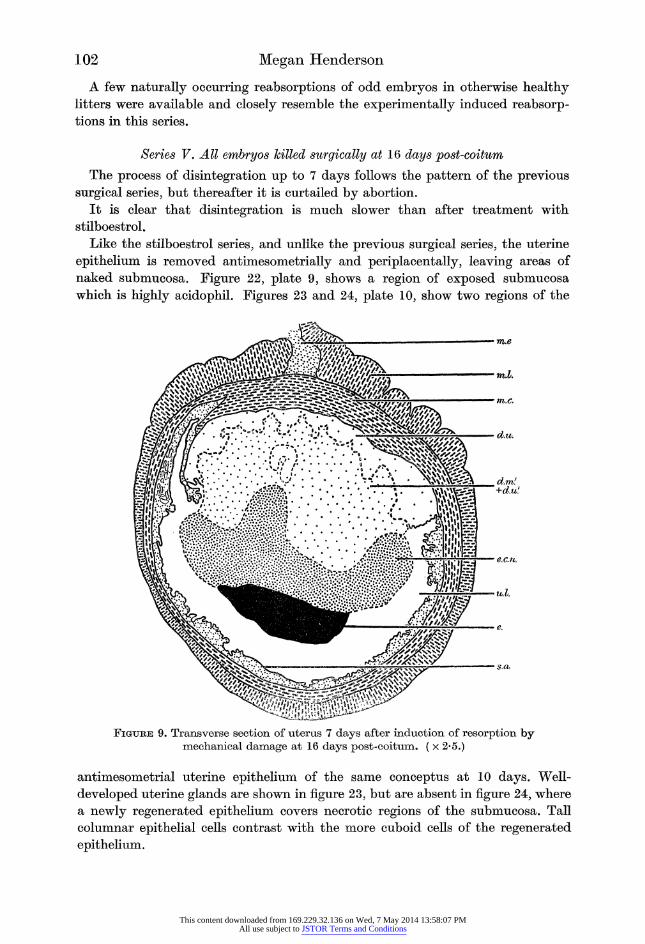
Like the stilboestrol series, and unlike the previous surgical series, the uterine
epithelium is removed antimesometrially and periplacentally, leaving areas of
naked submucosa. Figure 22, plate 9, shows a region of exposed submucosa
which is highly acidophil. Figures 23 and 24, plate 10, show two regions of the
agSlfev
mm
*rrue
-m.l.
mm.c.
' //' V ?' ?' ? ' ? V ''-%J*' /'? .'
' V '. 4 ? ,#J\ . ^S^wS^Ci.
?u.l.
Figure 9. Transverse section of uterus 7 days after induction of resorption by mechanical damage at 16 days post-coitum. ( x 2*5.)
antimesometrial uterine epithelium of the same conceptus at 10 days. Well-
developed uterine glands are shown in figure 23, but are absent in figure 24, where
a newly regenerated epithelium covers necrotic regions of the submucosa. Tall
columnar epithelial cells contrast with the more cuboid cells of the regenerated
epithelium.
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions
Foetal regression in rabbits 103
The decidua fragments, as in the stilboestrol series. The amount of fragmenta? tion varies, as can be seen by comparing figures 8 and 9, of two conceptuses from
the same animal, killed at 7 days.
Leucocytosis occurs, but somewhat later than in the corresponding stilboestrol
series, and reaches a maximum at 10 days (figure 26, plate 10). Polymorphs are
conspicuous in the plasmodial walls of the tubules of the ectoplacenta (figure 19,
plate 9). They are also present in the uterine lumen and naked submucosa (figure
22, plate 9; figure 24, plate 10). Polymorphs, eosinophils and some macrophages are seen at 8 days in the submucosa (figure 25, plate 10).
Series VI. All embryos killed surgically at 19 to 20 days post-coitum
Death of the litter, brought about surgically at 20 days, usually results in
abortion, as after stilboestrol treatment at the same age. The time of abortion
varies. Whereas one animal aborted at 3 days, another probably aborted between
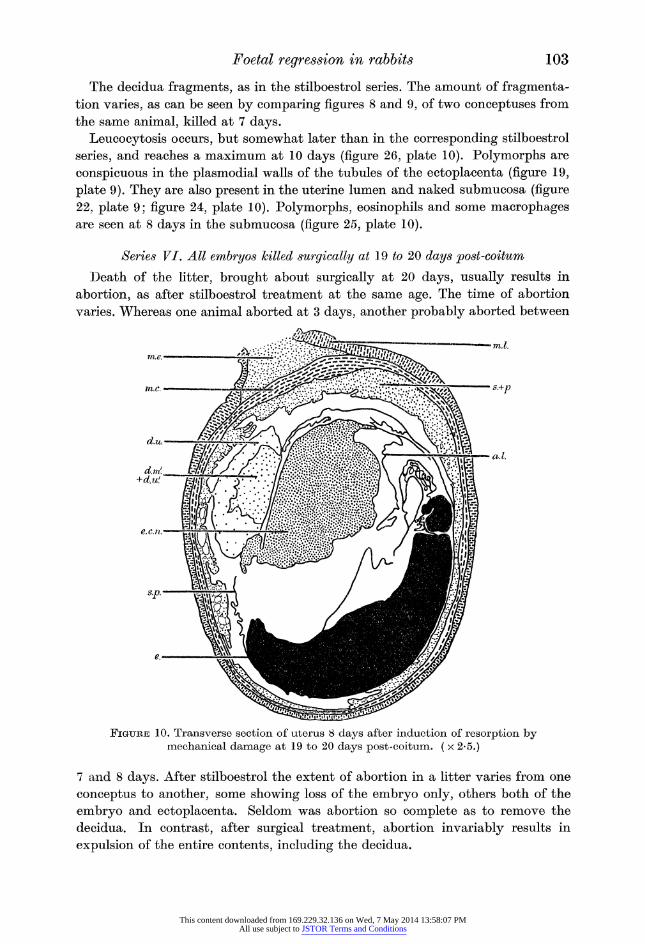
Figure 10. Transverse section of uterus 8 days after induction of resorption by mechanical damage at 19 to 20 days post-coitum. ( x 2*5.)
7 and 8 days. After stilboestrol the extent of abortion in a litter varies from one
conceptus to another, some showing loss of the embryo only, others both of the
embryo and ectoplacenta. Seldom was abortion so complete as to remove the
decidua. In contrast, after surgical treatment, abortion invariably results in
expulsion of the entire contents, including the decidua.
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions

104 Megan Henderson
The rate of autolysis is slower than after stilboestrol treatment.
Polymorphonuclear leucocytes are evident in all conceptuses being distributed
as in series V. Figure 28, plate 11, shows polymorphs in the exposed margin of the
decidua. The embryo lies against the antimesometrial uterine wall (figure 10).
Figure 27, plate 11, shows leucocytes making their way from the submucosa,
through the epithelium, with a large number in the uterine lumen in close contact
with the splanchnopleur.
4. Phagocytosis
Examination of the material from these 16-day series of experiments indicates
that the method and timing of reabsorption follows the course already described.
Stilboestrol treatment always results in complete reabsorption in situ, while
surgical treatment results in disintegration in situ followed by abortion about the
7th or 8th day. Vaginal smears
Vaginal smears on the first and second days after the injection of stilboestrol
contain epithelial cells and little else. Subsequently polymorphs appear in increasing numbers up to the 5th day, and thereafter decline gradually. Leucocytes appear in the smears 2 or 3 days later after surgical treatment. Cell debris increases at the
same time as the leucocytes, suggesting that fragmentation of the uterine content
may be facilitated by their action. A few macrophages can be recognized in most
of the smears. Free graphite, 'Aquadag' and carmine are detectable in these
smears a few hours after injection. These marker substances could not be detected
in the smears in polymorphs but were visible in macrophages.
Sections
The suspension of marker substance is distributed throughout the uterine lumen
soon after injection at the tubal end. The marker accumulates in two main regions,
namely, between the ectoplacenta and decidua, and in the antimesometrial uterine
wall. Both these regions are characterized by early and rapid necrosis. A cleft
between the edge of the decidua and the ectoplacenta, which soon appears, forms
a pocket which traps the marker, at least for a time. The tissues become loose and
spongy as necrosis progresses, allowing the marker to penetrate further. This
concentration of marker is dispersed throughout the uterine lumen with the dis?
integration of the decidua and ectoplacenta. There is no evidence that the marker
actually enters cells of the decidua or ectoplacenta. The early breakdown of the
antimesometrial uterine wall after the death of the embryo exposes a naked
necrotic submucosa. Granules of marker collect in clumps in this, apparently in intercellular spaces. These clumps remain and are still evident long after the
formation of a covering epithelium (figure 29, plate 11). In this way the marker
is enclosed in the regenerating uterine wall beneath the epithelium. Granules of marker are evident in polymorphs and macrophages, but not in any
other type of cell. Polymorphs and macrophages are found scattered throughout the uterine lumen amongst the cell debris, the former far outnumbering the latter.
Macrophages predominate amongst the leucocytes containing the marker, and are
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions
Foetal regression in rabbits 105
rmal
5 days
10 days
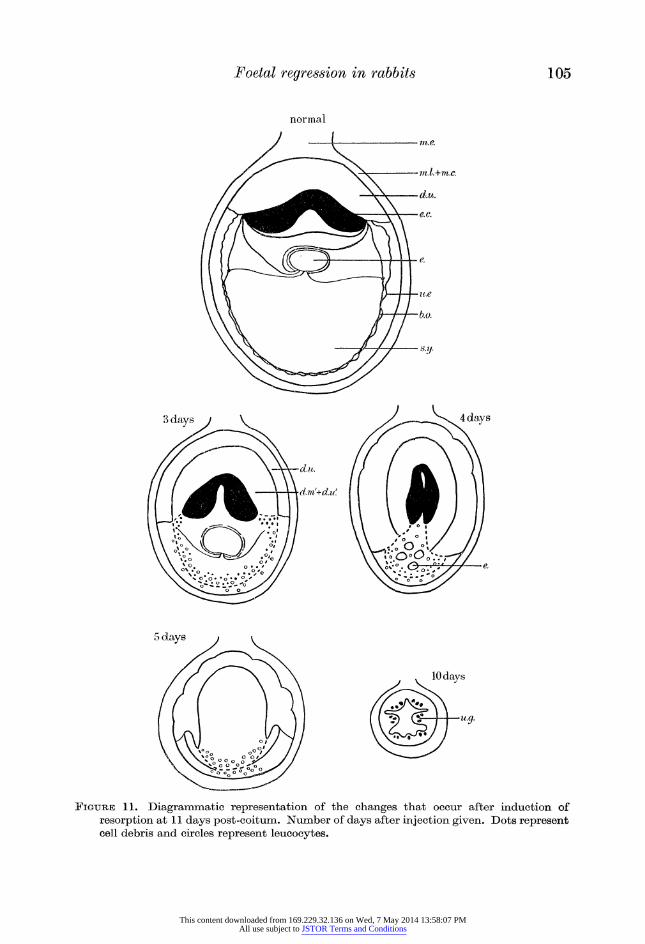
Figure 11. Diagrammatic representation of the changes that occur after induction of resorption at 11 days post-coitum. Number of days after injection given. Dots represent cell debris and circles represent leucocytes.
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions
106 Megan Henderson
stilboestrol injection
3 days
mechanical damage of all embryos
mechanical damage of some embryos
5 days
13 days
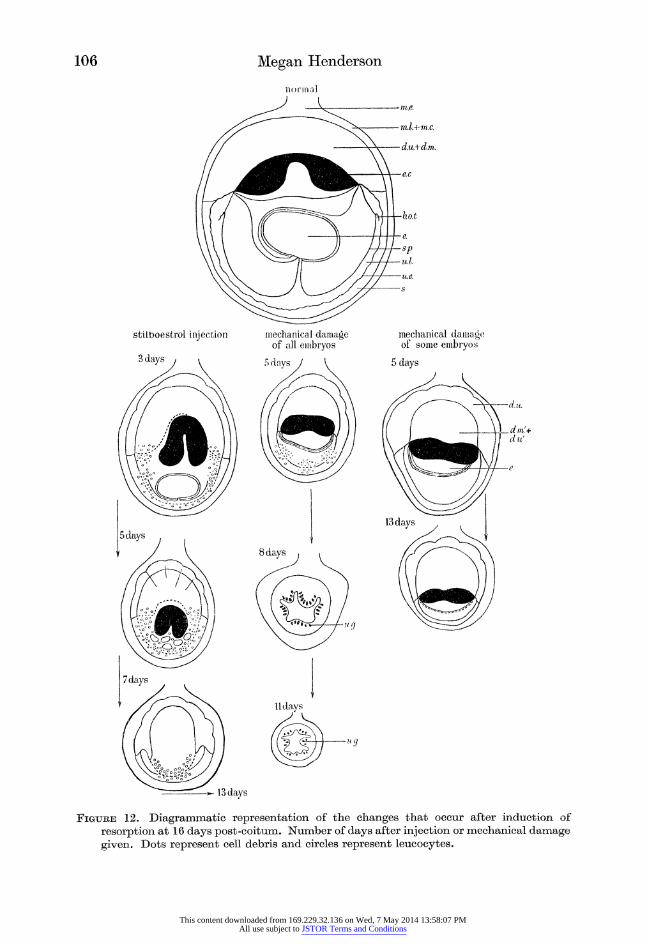
Figure 12. Diagrammatic representation of the changes that occur after induction of
resorption at 16 days post-coitum. Number of days after injection or mechanical damage given. Dots represent cell debris and circles represent leucocytes.
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions

Foetal regression in rabbits 107
much more heavily laden with it than the polymorphs. Laden leucocytes are
detectable in the exposed antimesometrial submucosa and periplacentally where
the epithelium is still intact. They also accumulate in the naked decidual remains.
Even late in reabsorption when the uterine epithelium is complete, leucocytes are
still evident in the submucosa beneath it. Macrophages containing carmine are
present occasionally in both the circular and longitudinal muscle layers antimeso?
metrially. Sometimes they occur in the connective tissue separating the longitudinal muscle bundles.
Invariably the serous coat of the uterus is thickened antimesometrially. This is
shown in figure 30, plate 11, of an animal 6 days after stilboestrol treatment. This
thickening reaches a maximum when the naked decidua comes in contact with the
antimesometrial submucosa and decreases as the uterus returns to normal. It
contains numerous lymphocytes and laden macrophages and is obviously lymphoid tissue. Small clumps of leucocytes laden with marker are present in lymph spaces in the mesometrium, and both laden macrophages and polymorphs occur scattered
in the tissues of the mesometrium. Laden macrophages were found in the serous
coat of the vagina in one animal.
Laden leucocytes were not identified either in sections or smears from other
regions. The spleen, liver, mesentery and lymph nodes were examined in sections,
and smears were made of lymph and bone marrow, without result.
Discussion
Both the methods employed to kill the embryos, stilboestrol or surgical, were
efficient. The changes that occur in the uterus subsequently depend, first, on
whether some embryos survive and the pregnancy is maintained or whether all the
embryos die, with consequent termination of pregnancy, secondly, on the method
of killing the embryos, and thirdly, on the age at which the embryos died.
The changes observed in series IV, in which some only of the embryos were
retrogressing and pregnancy was maintained, differ strikingly from those in the
other series, in all of which pregnancy was terminated by the death of all the
embryos. Retrogression of a conceptus in a pregnant uterus is characterized by
delayed fragmentation, the foetus remaining intact and recognizable until full
term, by the antimesometrial uterine mucosa remaining intact and by the absence
of marked leucocytosis. The changes observed can be assigned to necrosis of the
foetal tissues without the intervention of maternal inflammatory processes or even
of repair of the placental site, since the placenta remains in position. The uterus
does not appear to react to the presence of a dead foetus so long as pregnancy
persists. The method of killing affects the order in which the embryonic and placental
tissues degenerate, the time of onset of the inflammatory and repair processes and
the stage at which the uterine contents are expelled. After stilboestrol the decidua
and ectoplacenta become necrotic before the embryo, whereas after surgical treat1
ment the embryonic tissues are affected before the decidua and ectoplacenta. It
is well known that oestrogens are injurious to the placenta, and hence it is not
surprising that the placenta is the first region to be affected by stilboestrol. Several
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions

1AQ Morra/n R ^nri^T^cirkn
authors have claimed that the placenta may survive and continue to grow after
the death of the embryo. This was reported by Newton (1935) in the mouse, by van Wagenen & Newton (1943) in the monkey (Macaca mulatta), and by Huggett & Pritchard (1945) in the rat. This has not been found to occur in the rabbit,
although the trophoblast of the plasmodial walls of the tubules in the ectoplacenta
may survive for a few days (see figures 18 and 19, plate 9).
Leucocytosis occurs both when death is induced by stilboestrol and by surgical
treatment, but it is more rapid in onset and more pronounced after the former
method of killing. This may be a direct effect of stilboestrol in the one case, and an
effect of natural oestrogens reappearing, after the suppression of pregnancy, in the
other. This would account for the difference both in intensity and timing of the
inflammatory processes according to the method of killing. Kerr (1947), dealing with the effect of colchicine treatment of mouse embryos, notes the presence of
acidophil leucocytes which, he states, do not resemble the ' eosinophilous phago?
cytes' figured by Fortuyn (1920). Marchand (1895), dealing with the human
embryo, spoke of an infiltration of the conceptus by leucocytes, as did Daels (1908), who states that maternal leucocytes can penetrate a normal syncytium to reach
a necrotic area beyond. It is interesting to note in this connexion that Rowson,
Lamming & Fry (1953) have shown that the uterus of the cow is susceptible to
bacterial infection during the luteal phase of the cycle, but is highly insusceptible
during oestrus, and that stilboestrol injection of ovariectomized cows confers a
similar insusceptibility.
Disintegration takes place more rapidly after stilboestrol than after surgical
treatment, as is seen by comparing series II and V. Uterine contractility is no
doubt an important factor in promoting disintegration as well as expulsion. Abortion does not appear to occur in the rabbit before the 19 to 20th day post-
coitum; thereafter uteri in which all the embryos were killed surgically at 16 or
20 days post-coitum aborted completely. After stilboestrol at 16 days, abortion
did not occur, probably because the rapid onset of autolysis and fragmentation had reduced the uterine content sufficiently before the 20th day. Abortion did
occur after stilboestrol at 20 days, but was incomplete. It may be that the
oestrogenic action of stilboestrol upsets the normal endocrine sequence which leads
to parturition. The destruction of the uterine mucosa antimesometrially and periplacentally is
one of the most remarkable phenomena observed. It occurs in all uteri in which
there were no surviving embryos. The uterine epithelium disintegrates and the
underlying tissues become necrotic, leaving the submucosa exposed. These regions of the mucosa do not contain decidual tissue in the rabbit, and they do not break
down after parturition, nor do they do so in retrogressing conceptuses in otherwise
pregnant uteri. The breakdown occurs only in retrogressing conceptuses in uteri
with no surviving embryos and in those regions where the mucosa is in contact
with decidual debris. It is difficult to avoid the assumption that the destruction
of the mucosa is brought about by this contact.
The time at which the placenta comes away from the mesometrial wall of the
uterus and the completeness of the separation varies according to whether it
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions

Foetal regression in rabbits 109
results from reabsorption, abortion or parturition. Fragments of necrotic decidual
tissue remain attached to the margin of the healthy uninucleate cell region what?
ever the method of removal of the placenta, but are large after reabsorption, and
small after abortion or parturition. Most of these fragments are lost before the
placental site heals. The regeneration of uterine mucosa over the placental site, once this has been denuded, is similar after reabsorption to that after abortion or
parturition. The evidence available in the literature as to whether all the products of retro?
gression of the dead foetuses are ultimately expelled from the uterus or whether
some part of these are reabsorbed into the maternal system has been conflicting. If foetal material is reabsorbed, then the process may provide a means by which
foetal antigens can reach the maternal system. It is well known that, both in man
and in the horse, the mother does in certain circumstances become immunized to
foetal antigens during pregnancy. It was a main purpose of this research to
determine if the processes of reabsorption in the rabbit provide an avenue for such
passage of foetal material, but the question of whether that material is antigenic is obviously beyond the scope of the present work. Since histological examination
of the problem was indecisive, experiments involving the injection of suspensions of marker substances into the uterine lumen were performed.
Leucocytes laden with marker are present in the uterine tissues, macrophages
being more numerous and more heavily laden than polymorphs. Laden leucocytes
penetrate into the uterine tissues while the submucosa is naked and are then
covered by the regenerated epithelium, yet they are apparently capable of
penetrating after the complete regeneration of the epithelium. Macrophages are
known to ingest inert particles in particular, so that the presence in them of
marker is not necessarily a guide to the amount of foetal debris which they would
engulf. Marker lodges readily in crevices and clefts in the exposed necrotic submucosa
antimesometrially and becomes enclosed in the regenerating uterine wall by the
overgrowth of the epithelium. It thus becomes shut into the tissues, ultimately
being removed by leucocytes. Laden cells are evident in the tissues surrounding such areas. Doubtless foetal cell debris can become enclosed in the tissues in
a similar manner and likewise would be removed by the leucocytes. Laden
leucocytes are most prominent antimesometrially where they tend to accumulate
in lymphoid tissues below the serous coat, and also in the connective tissue of the
mesometrium itself. The accumulation of leucocytes beneath the serous coat anti?
mesometrially is as marked in conceptuses at the cervical end of the uterus as in
those near the point of injection at the tubal end. Hence, it is unlikely that they are laden with marker that has leaked into the abdominal cavity, since this would
have led to a local accumulation near the point of injection at the tubal end. Their
even distribution in a distinct area antimesometrially on each conceptus is very
strong evidence that they have come from the uterine lumen where the marker is
evenly distributed.
Carleton (1931) showed non-ciliated columnar cells of the vagina of rabbits to
be phagocytic, but found no definite instance of phagocytosis by the uterine
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions

110 Megan Henderson
epithelium. In the present work no evidence was gained of phagocytosis by either
the vaginal or uterine epithelium.
Although the evidence that laden leucocytes retreat from the lumen into the
uterine tissues appears conclusive, it is nevertheless important to realize that these
form only a small proportion of those in the lumen. Examination of vaginal smears
shows beyond question that the bulk of the leucocytes in the lumen together with
the products of tissue disintegration are lost through the vagina. Yet, should only a very small quantity of foetal tissue products be carried into the maternal system,
they would be sufficient, if antigenic, to bring about immunological sensitization
of the mother.
It is concluded, therefore, that the process of reabsorption in the rabbit provides a particularly favourable opportunity for the transfer of foetal antigens, should
these be present, to the mother, and it seems that this may be found to be true of
other species also.
The author is greatly indebted to Professor F. W. Rogers Brambell, F.R.S., for
his advice and encouragement at all times, and to Mr R. A. Lansdowne for pre?
paring the photomicrographs. The work was financed by a grant from the
Agricultural Research Council to Professor Brambell, for which the author wishes
to express her thanks.
Refebences
Brambell, F. W. R., Henderson, M. & Mills, I. H. 1948 J. Exp. Biol. 25, 209-218. Bruner, D. W., Hull, E. F. & Doll, E. R. 1948 Amer. J. Vet. Res. 9, 237-242. Carleton, H. M. 1931 Proc. Roy. Soc. B, 108, 1-10. Castle, W. E. 1928 Amer. J. Anat. 42, 399-401. Daels, F. 1908 Au sujet de l'etiologie de la mole hydatique. Ann. Soc. Med. Gand (cited by
Mall, F. P. & Meyer, A. W. (1921)). Dekanski, J. 1949 J. Physiol. 108, 233-244. Duval, M. 1889-1892. J. Anat., Paris, 25, 309-342, 573-627; 26, 1-48, 273-344.
Fortuyn, A. B. D. 1920 Arch. Biol., Paris, 30, 323-357.
Huggett, A. St G. & Pritchard, J. J. 1945 Proc. Roy. Soc. Med. 6, 261-266. Kerr, T. 1947 Proc. Zool. Soc, Lond. 116, 551-564. Lendrum, A. C. 1944 J. Path. Bact. 56, 441.
Long, C. N. H. & Parkes, A. S. 1924 Biochem. J. 18, 800-805. Mall, F. P. & Meyer, A. W. 1921 Contr. Embryol. Garneg. Instn, 275, 1-364. Marchand, F. 1895 Mschr. Geb. Gyndk. p. 132 (cited by Mall, F. P. & Meyer, A. W. (1921)). Marshall, A. M. 1893 Vertebrate embryology, 1st ed., pp. 341-447. London: Smith, Elder
and Co.
Meyer, A. W. 1917 Anat. Rec. 12, 293-307. Minot, C. S. & Taylor, E. 1905 Normentaf. Wirbelt. 5. Mossman, H. W. 1926 Amer. J. Anat. 37, 433-497. Newton, W. H. 1935 J. Physiol. 84, 196-207. Rowson, L. E., Lamming, G. E. & Fry, R. M. 1953 Nature, Lond., 171, 749-750. van Wagenen, G. & Newton, W. H. 1943 Surg. Gynec. Obstet. 77, 539-543.
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions

Foetal regression in rabbits \\\
Abbreviations used in figiibes and plates
a. amniotic membrane m. macrophage a.l. allantoic membrane m.c. uterine circular muscle a.l.m. allantoic mesoderm m.e. mesometrium a.m. amniotic cavity m.e.p. position of mesometrium b.o. bilaminar omphalopleur m.e.s. mesoderm. b.o.t. bilaminar omphalopleur tag m.l. uterine longitudinal muscle c. chorion n. nuclei of plasmodial walls of tubules ca. carmine p. polymorphonuclear leucocytes cd. cell debris r.c maternal erythrocytes ct. connective tissue s. submucosa d.m. normal multinucleate decidual cells s.a. newly regenerated epithelium d.m. necrotic multinucleate decidual cells s.c distorted spinal cord d.u. normal uninucleate decidual cells s.n. submucosa devoid of epithelium d.u. necrotic uninucleate decidual cells s.p. splanchnopleur e. embryo s.t. sinus terminalis e.c normal ectoplacenta s.y. yolk sac
e.c.f. fragmenting ectoplacenta t.n. necrotic trophoblast e.cn. extremely necrotic ectoplacenta t.n.f. fragmenting trophoblast e.l. eosinophil leucocyte u.e. uterine epithelium e.x. exocoel u.g. uterine glands /. fibrin clot u.l. uterine lumen g.c giant cell v.c crushed vessels g.c' necrotic giant cell v.f. foetal vessels i.s. intercotyledonary space v.m. maternal vessels I. lymphoid tissue v.s. surface sinus
Descbiption of plates 8 TO 11
Plate 8
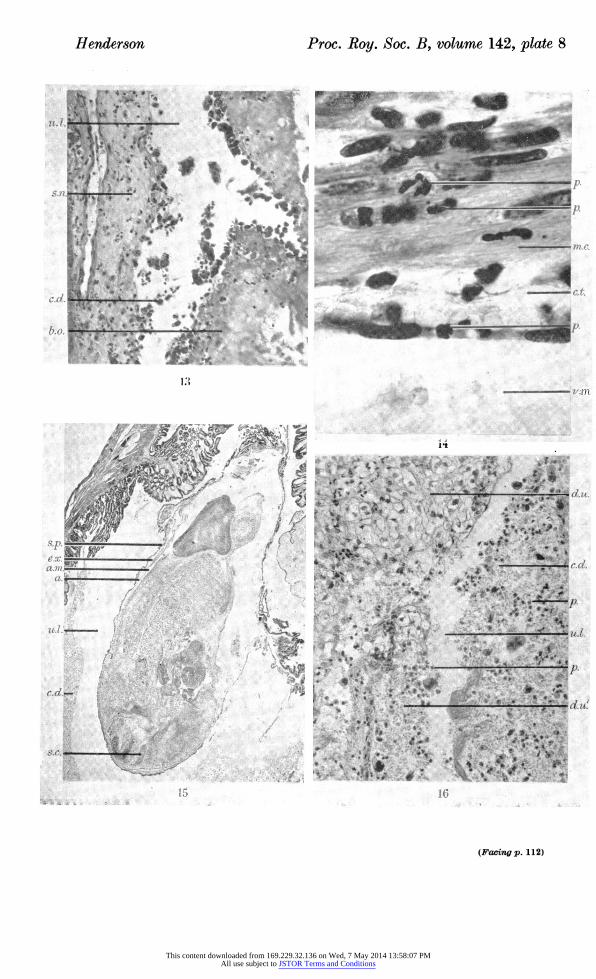
Figure 13. Photomicrograph of area of antimesometrial uterine wall in transverse section, 3 days after induction of resorption by stilboestrol at 11 days post-coitum, showing removal of uterine epithelium and necrosis of submucosa. ( x 170.)
Figure 14. Photomicrograph of area of antimesometrial uterine wall in transverse section, 2 days after induction of resorption by stilboestrol at 11 days post-coitum. A leucocyte is shown traversing the lining of a vessel, with several others in the circular muscle. ( x 933.)
Figure 15. Photomicrograph of embryo and membranes in transverse section, 1 day after induction of resorption by stilboestrol at 16 days post-coitum, showing apposition of splanchnopleur and amniotic membrane. ( x 11.)
Figure 16. Photomicrograph of area of periplacental decidua in transverse section, 3 days after induction of resorption by stilboestrol at 16 days post-coitum, showing fragmenta? tion into the uterine lumen. ( x 227.)
Plate 9
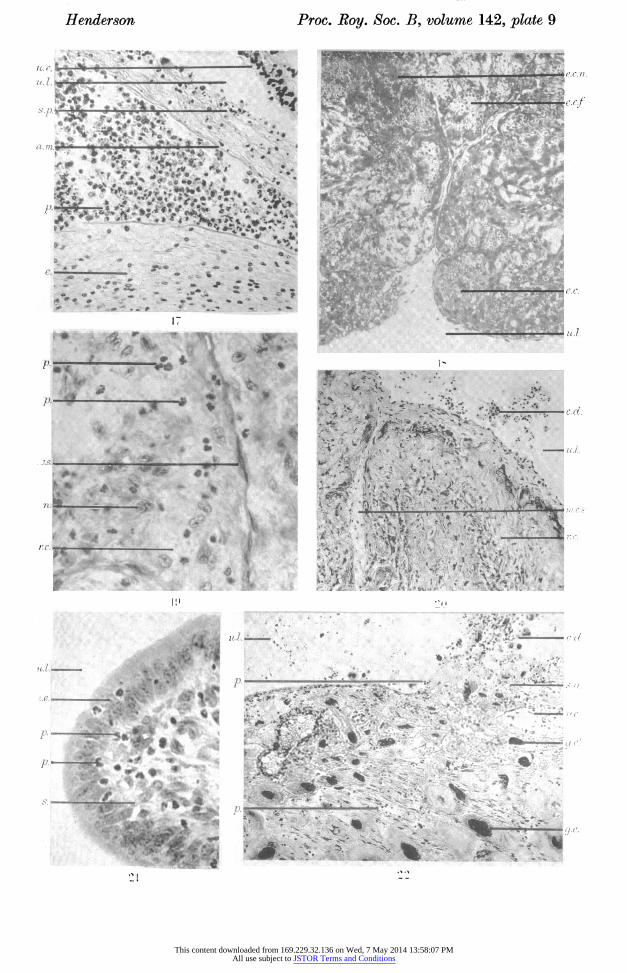
Figure 17. Photomicrograph of area of embryo and membranes in transverse section, 4 days after induction of resorption by stilboestrol at 16 days post-coitum, showing leucocytes within the amnion. ( x 227.)
Figure 18. Photomicrograph of area of ectoplacenta in transverse section, 5 days after induction of resorption by mechanical damage at 16 days post-coitum, showing regions of necrosis. ( x 123.)
Figure 19. Photomicrograph of area of ectoplacenta in transverse section, 6 days after induction of resorption by mechanical damage at 16 days post-coitum, showing oblitera? tion of foetal vessels and distribution of leucocytes. ( x 493.)
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions

112 Megan Henderson
Figure 20. Photomicrograph of area of ectoplacenta in transverse section, 10 days after induction of resorption by mechanical damage at 16 days post-coitum, showing general necrosis. ( x 123.)
Figure 21. Photomicrograph of lobe of periplacental uterine wall in transverse section, 9 days after induction of resorption by stilboestrol at 19 to 20 days post-coitum, showing- leucocytes (animal aborted). ( x 466.)
Figure 22. Photomicrograph of area of antimesometrial uterine wall in transverse section, 5 days after induction of resorption by mechanical damage at 16 days post-coitum, showing breakdown of uterine epithelium and exposure of necrotic submucosa. ( x 123.)
Plate 10
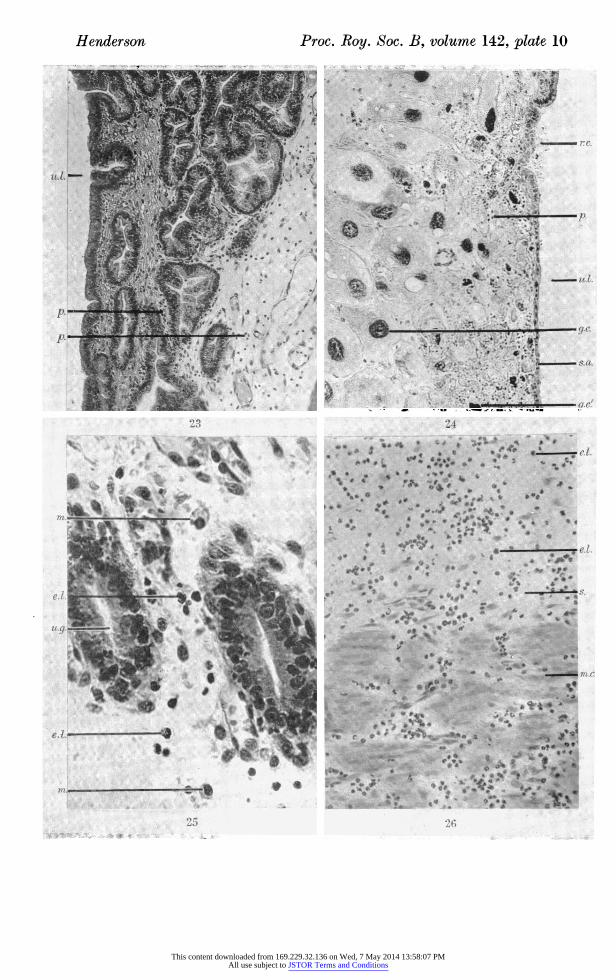
Figure 23. Photomicrograph of area of antimesometrial uterine wall in transverse section, 10 days after induction of resorption by mechanical damage at 16 days post-coitum. Area shows unaffected uterine epithelium and glands. ( x 123.)
Figure 24. Photomicrograph of area of antimesometrial uterine wall in transverse section, 10 days after induction of resorption by mechanical damage at 16 days post-coitum. Area shows reformation of epithelium and absence of glands. ( x 123.)
Figure 25. Photomicrograph of area of submucosa in transverse section, 8 days after induction of resorption by mechanical damage at 16 days post-coitum, showing macro?
phages and eosinophil leucocytes. ( x 493.) Figure 26. Photomicrograph of area of periplacental uterine wall in transverse section,
10 days after induction of resorption by mechanical damage at 16 days post-coitum, showing numerous polymorphonuclear leucocytes and 2 eosinophil leucocytes. ( x 227.)
Plate 11
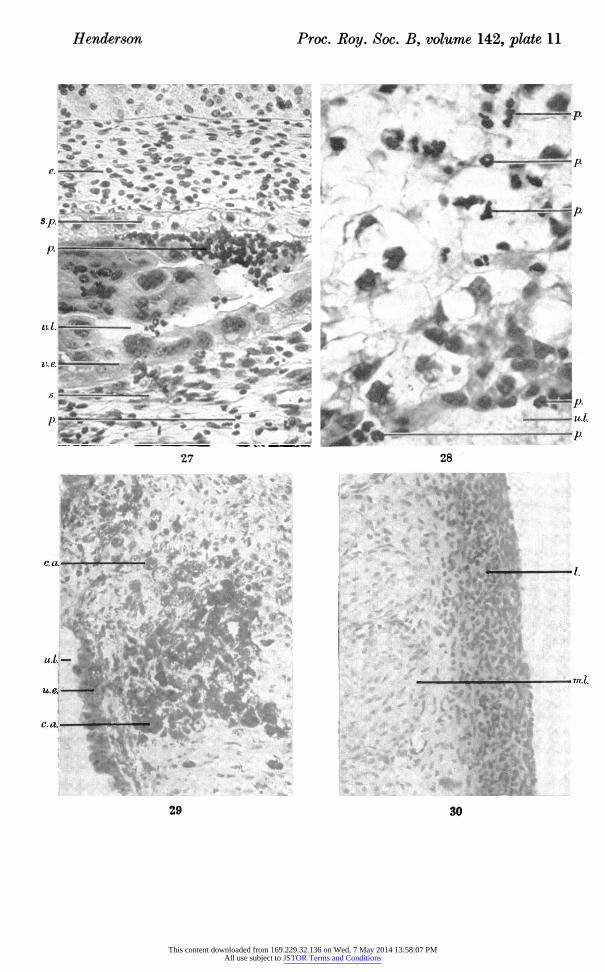
Figure 27. Photomicrograph of area of antimesometrial uterine wall and embryo in trans? verse section, at 8 days after induction of resorption by mechanical damage at 19 to 20 days post-coitum, showing distribution of leucocytes. ( x 360.)
Figure 28. Photomicrograph of area of decidua in transverse section, 8 days after induction of resorption by mechanical damage at 19 to 20 days post-coitum. Polymorphonuclear leucocytes are shown in remains of decidua. (Animal aborted.) ( x 733.)
Figure 29. Photomicrograph of area of submucosa in transverse section, 7 days after induction of resorption by stilboestrol at 16 days post-coitum, and 4 days after injection of carmine into the uterine lumen. Carmine is shown within the tissues, covered by a
complete epithelium. ( x 246.) Figure 30. Photomicrograph of area of antimesometrial uterine wall in transverse section,
6 days after induction of resorption by stilboestrol at 16 days post-coitum, showing lymphoid tissue within the serous coat. ( x 246.)
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions
Henderson Proc. Roy. Soc. B, volume 142, plate 8
(Facing p. 112)
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions
Henderson Proc. Roy. Soc. B, volume 142, plate 9
erf
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions
Henderson Proc. Roy. Soc. B, volume 142, plate 10
gfJlPiifigf p|H!? =!??*????
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions
Henderson Proc. Roy. Soc. B, volume 142, plate 11
30
This content downloaded from 169.229.32.136 on Wed, 7 May 2014 13:58:07 PMAll use subject to JSTOR Terms and Conditions