Embed Size (px)
Citation preview

Flow-Dependent Remodeling in the Carotid Artery ofFibroblast Growth Factor-2 Knockout Mice
Chris J. Sullivan, James B. Hoying
Objective—Fibroblast growth factor-2 (FGF2) has been implicated as a mediator in the structural remodeling of arteries.Chronic changes in blood flow are known to cause reorganization of the vessel wall, resulting in permanent changes inartery size (flow-dependent remodeling). Using FGF2 knockout (Fgf22/2) mice, we tested the hypothesis that FGF2 isrequired during flow-dependent remodeling of the carotid arteries.
Methods and Results—All branches originating from the left common carotid artery (LCCA), except for the left thyroidartery, were ligated to reduce flow in the LCCA and increase flow in the contralateral right common carotid artery(RCCA). Age- and sex-matched control animals did not undergo ligation of the LCCA branches. Morphometric analysisshowed that by day 7, vessel diameter was significantly greater in the high-flow RCCA of FGF2 wild-type (Fgf21/1)andFgf22/2 mice versus the respective control RCCA, demonstrating outward remodeling. In contrast, vessel diameterwas decreased by day 7 in the low-flow LCCA of both genotypes compared with the control LCCA, showing inwardremodeling. No differences were observed betweenFgf21/1 and Fgf22/2 mice in either high-flow or low-flowremodeling.
Conclusions—Given these results, we demonstrate that FGF2 is not essential for flow-dependent remodeling of the carotidarteries.(Arterioscler Thromb Vasc Biol. 2002;22:●●●-●●●.)
Key Words: arterial remodelingn basic fibroblast growth factorn knockout micen flow-dependent remodelingnfibroblast growth factor-2
Vascular remodeling is the structural reorganization of avessel involving a variety of cell activities, including
proliferation, apoptosis, migration, and extracellular matrixrestructuring.1–3 Remodeling of the arterial wall occurs afterchronic changes in blood pressure and blood flow and inresponse to vessel injury.4–7 Arterial remodeling due tochanges in blood flow (flow-dependent remodeling) occurs inphysiological1,8 and pathological situations.9–12 In patholog-ical settings, such as atherosclerosis and angioplasty, arterialremodeling plays a critical role in the degree of vesselnarrowing during plaque or lesion progression.5,13–16
The molecular mediators of vessel remodeling are stillunclear. Fibroblast growth factor (FGF)2 is a molecule that isstrongly implicated in flow-dependent remodeling. FGF2mRNA expression is sensitive to alterations in fluid flow andshear stress,17 and FGF2 protein expression increases in thevascular wall during flow-induced arterial enlargement.18 Inaddition, antibody neutralization of endogenous FGF2 hasbeen shown to reduce inward remodeling in a mouse modelof carotid artery flow cessation.19 The specific function ofFGF2 during these remodeling events is not clear. Previousstudies suggest that FGF2 could possibly be affecting vascu-lar cell turnover, gene expression, or matrix restructuring inthe adapting vessel.20–25
We used a novel mouse model of vessel remodeling, FGF2knockout (Fgf22/2) mice, to test the hypothesis that FGF2 isrequired during flow-dependent arterial remodeling. Themodel induces inward (low flow–induced) and outward (highflow–induced) remodeling in the left and right carotid arter-ies, respectively.
MethodsAn expanded Methods section is available online athttp://atvb.ahajournals.org.
Experimental AnimalsMale FGF2 wild-type (Fgf21/1) and Fgf22/2 mice26 (50% BlackSwiss and 50% 129 SV) were used for all experiments in accordancewith University of Arizona Institutional Animal Care and Use–approved procedures. All mice were genotyped by polymerase chainreaction by using primers specific for the FGF2 wild-type andknockout alleles, after collection of genomic DNA. Procedures weretimed so that all mice were 8 weeks of age (64 days) at the time ofeuthanasia. Mice were anesthetized with 2.5% Avertin (Aldrich) at adose of 0.15 mL/10 g body wt injected intraperitoneally.
Surgery to Induce Flow-DependentCarotid RemodelingThe mouse model presented is a modification of procedures previ-ously published for use in the rat.27,28All branches originating from
Received March 11, 2002; revision accepted May 8, 2002.From the Physiological Sciences (C.J.S., J.B.H.) and Biomedical Engineering (J.B.H.) Programs, University of Arizona, Tucson.Correspondence to James B. Hoying, PhD, Arizona Health Sciences Center, Room 5328, 1501 N Campbell, PO Box 245084, Tucson, AZ 85724.
E-mail [email protected]© 2002 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol.is available at http://www.atvbaha.org DOI: 10.1161/01.ATV.0000023230.17493.E3
1
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from

the left common carotid artery (LCCA), except for the left thyroidartery, were ligated (6.0 silk) to reduce flow in the LCCA andincrease flow in the contralateral right common carotid artery(RCCA). Mice were euthanized at days 4, 7, and 28 after surgicalligation. Age- and sex-matched control animals were euthanizedwithout having undergone ligation.
Carotid Artery Blood FlowCarotid artery (RCCA and LCCA) blood flow was measured with theuse of an ultrasonic transit-time flowmeter (Transonic Systems) witha 0.5-V series probe as described previously.29 Blood flow wasevaluated in mice (n53 per genotype) before ligation, immediatelyafter ligation, and again at day 14 after ligation.
MorphometryControl (n54 mice per genotype), day 7 (n56 mice per genotype),and day 28 (n57Fgf21/1 mice and n56Fgf22/2 mice) wereperfusion-fixed at constant pressure (90 to 100 mm Hg) with 10%formalin through a polyethylene catheter placed in the left ventricle.The neck, between the clavicle and mandible, was isolated, placed infixative overnight, and then decalcified by using Decalcifier I and II(Surgipath) for 24 hours each. After paraffin embedding, serial crosssections were cut (8mm) and stained with hematoxylin. Morpho-metric analysis30,31 was performed on the carotid arteries from 2whole-neck sections for each animal that were cut 160 to 200mmapart and located at approximately the midportion of the commoncarotid artery.
AngiographyFor angiography, the arterial circulation was perfused (constantpressure of 90 to 100 mm Hg) with PBS containing 131025 mol/Lsodium nitroprusside, followed by filling with contrast agent (210%
[wt/vol] barium sulfate, Liqui-Coat, Lafayette Pharmaceuticals)through a catheter inserted into the left ventricle. Angiograms of thehead and neck region were obtained with the use of a high-definitionx-ray cabinet system (Faxitron).
Vascular Cell Proliferation and ApoptosisTo examine proliferation, animals (n53 per genotype) were injectedintraperitoneally with bromodeoxyuridine (BrdU, 30 mg/kg body wt;Sigma Chemical Co) at 24 hours and 12 hours before euthanasia onday 4 after LCCA surgery. Mice were perfusion-fixed, and thevertebrae were removed by careful dissection before paraffin em-bedding. BrdU incorporation into the nuclei of proliferating cells wasidentified on 6-mm sections32 with the use of a peroxidase-conjugated sheep anti-BrdU antibody (Biodesign International).BrdU-positive nuclei were counted per 2 whole-vessel transversesections from each artery. Apoptotic cells were identified by using aBoehringer-Mannheim In Situ Cell Death Detection Kit.
Statistical AnalysisValues are presented as mean6SEM. Comparison between 2 meanswas accomplished by using the Student unpairedt test. Multiplegroups were compared by 1-way ANOVA with a Student-Newman-Keuls test. Comparison of carotid artery blood flow, within agenotype, before and after ligation was accomplished by using a1-way repeated-measures ANOVA with a Student-Newman-Keulstest. Statistical significance was set atP,0.05.
ResultsCarotid Artery Blood FlowAverage blood flow was evaluated in the LCCA and theRCCA (Figure 1). Acutely, the procedure significantly re-
Figure 1. A, Photograph of LCCA and its branches, including the external carotid (EC), internal carotid (IC), occipital (OCC), and leftthyroid (T) arteries. The EC, IC, and OCC were permanently ligated at positions indicated by an asterisk. The arterial circulation wasfilled with a white contrast agent (barium sulfate) to improve visualization for this photo. B, Average blood flow (mL/min) in the LCCAand RCCA of Fgf21/1 (n53) and Fgf22/2 (n53) mice before ligation of LCCA branches, immediately after the ligations (day 0), and againat 14 days after the ligations (day 14). *P,0.05 and #P,0.01 vs respective values before ligation; P5NS for Fgf21/1 vs Fgf22/2.
2 Arterioscler Thromb Vasc Biol. July 2002
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
duced flow in the LCCA (decreased by.80%) and signifi-cantly increased flow in the contralateral RCCA (increasedby .40%). These changes persisted by day 14 after theLCCA surgery. Blood flow values were not significantlydifferent betweenFgf21/1 and Fgf22/2 mice. Similar topreviously published data,29 ligation of just the left externalcarotid artery caused only a modest decrease in LCCA bloodflow ('30% reduction) and resulted in no change in bloodflow within the contralateral RCCA (data not shown).
Flow-Dependent Arterial RemodelingAt day 7 and day 28 after surgery, inFgf21/1 and inFgf22/2
mice, the low-flow LCCA was inwardly remodeled, whereas
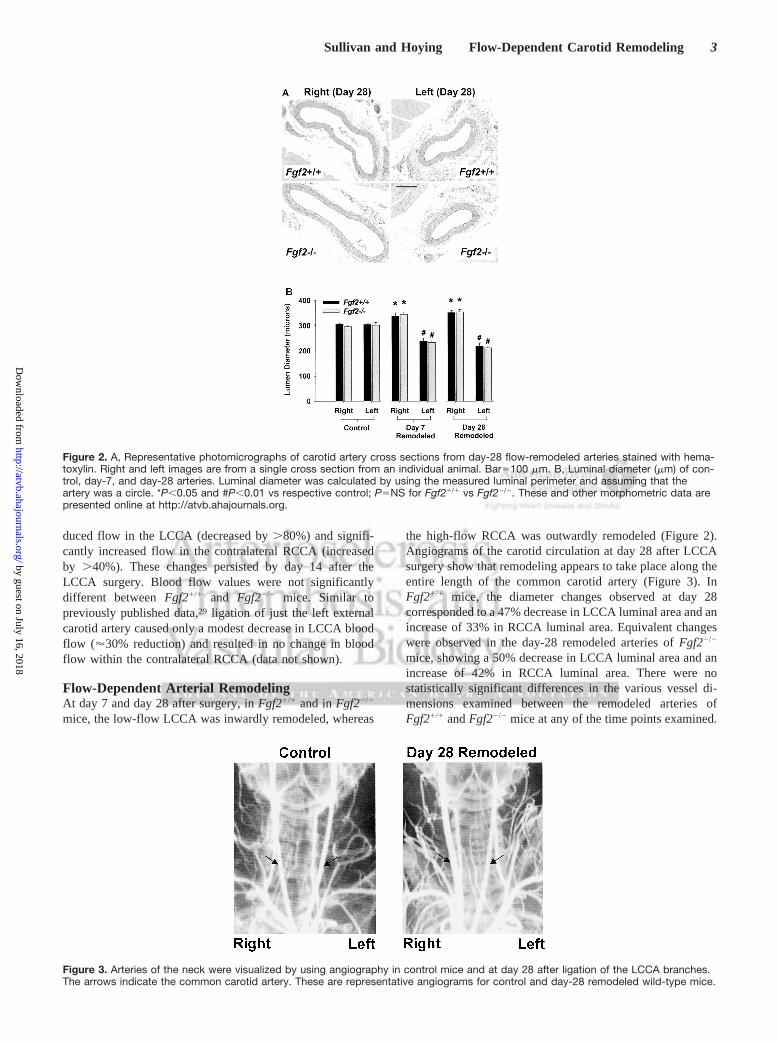
the high-flow RCCA was outwardly remodeled (Figure 2).Angiograms of the carotid circulation at day 28 after LCCAsurgery show that remodeling appears to take place along theentire length of the common carotid artery (Figure 3). InFgf21/1 mice, the diameter changes observed at day 28corresponded to a 47% decrease in LCCA luminal area and anincrease of 33% in RCCA luminal area. Equivalent changeswere observed in the day-28 remodeled arteries ofFgf22/2
mice, showing a 50% decrease in LCCA luminal area and anincrease of 42% in RCCA luminal area. There were nostatistically significant differences in the various vessel di-mensions examined between the remodeled arteries ofFgf21/1 andFgf22/2 mice at any of the time points examined.
Figure 2. A, Representative photomicrographs of carotid artery cross sections from day-28 flow-remodeled arteries stained with hema-toxylin. Right and left images are from a single cross section from an individual animal. Bar5100 mm. B, Luminal diameter (mm) of con-trol, day-7, and day-28 arteries. Luminal diameter was calculated by using the measured luminal perimeter and assuming that theartery was a circle. *P,0.05 and #P,0.01 vs respective control; P5NS for Fgf21/1 vs Fgf22/2. These and other morphometric data arepresented online at http://atvb.ahajournals.org.
Figure 3. Arteries of the neck were visualized by using angiography in control mice and at day 28 after ligation of the LCCA branches.The arrows indicate the common carotid artery. These are representative angiograms for control and day-28 remodeled wild-type mice.
Sullivan and Hoying Flow-Dependent Carotid Remodeling 3
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
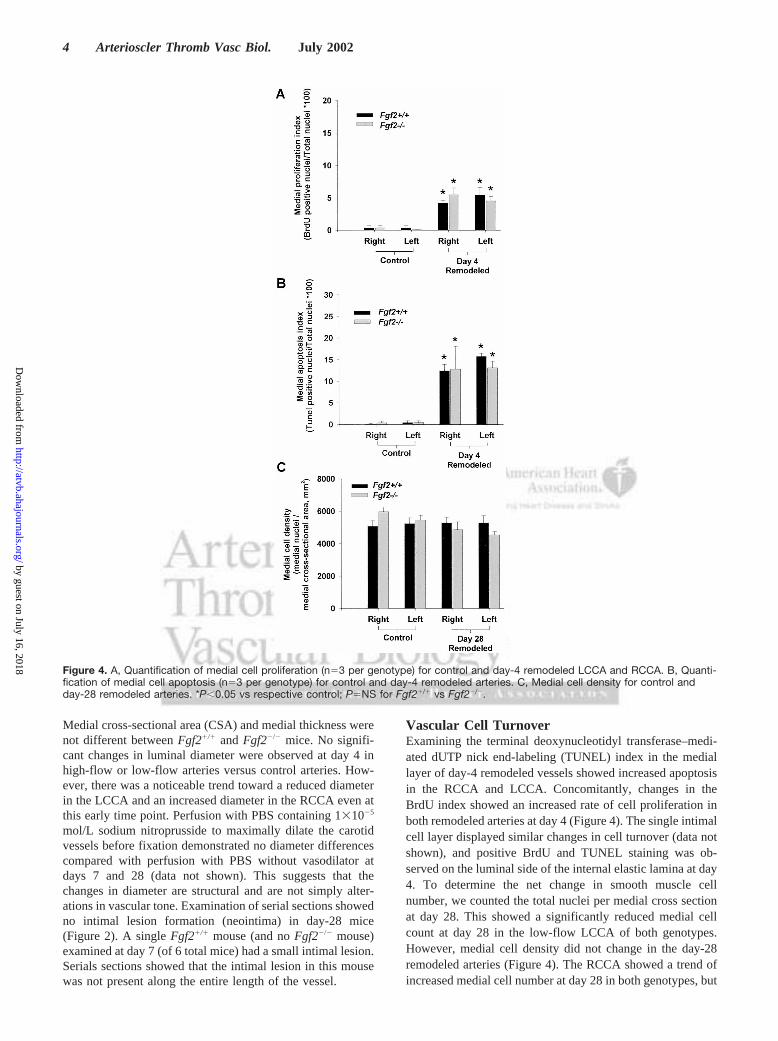
Medial cross-sectional area (CSA) and medial thickness werenot different betweenFgf21/1 andFgf22/2 mice. No signifi-cant changes in luminal diameter were observed at day 4 inhigh-flow or low-flow arteries versus control arteries. How-ever, there was a noticeable trend toward a reduced diameterin the LCCA and an increased diameter in the RCCA even atthis early time point. Perfusion with PBS containing 131025
mol/L sodium nitroprusside to maximally dilate the carotidvessels before fixation demonstrated no diameter differencescompared with perfusion with PBS without vasodilator atdays 7 and 28 (data not shown). This suggests that thechanges in diameter are structural and are not simply alter-ations in vascular tone. Examination of serial sections showedno intimal lesion formation (neointima) in day-28 mice(Figure 2). A singleFgf21/1 mouse (and noFgf22/2 mouse)examined at day 7 (of 6 total mice) had a small intimal lesion.Serials sections showed that the intimal lesion in this mousewas not present along the entire length of the vessel.
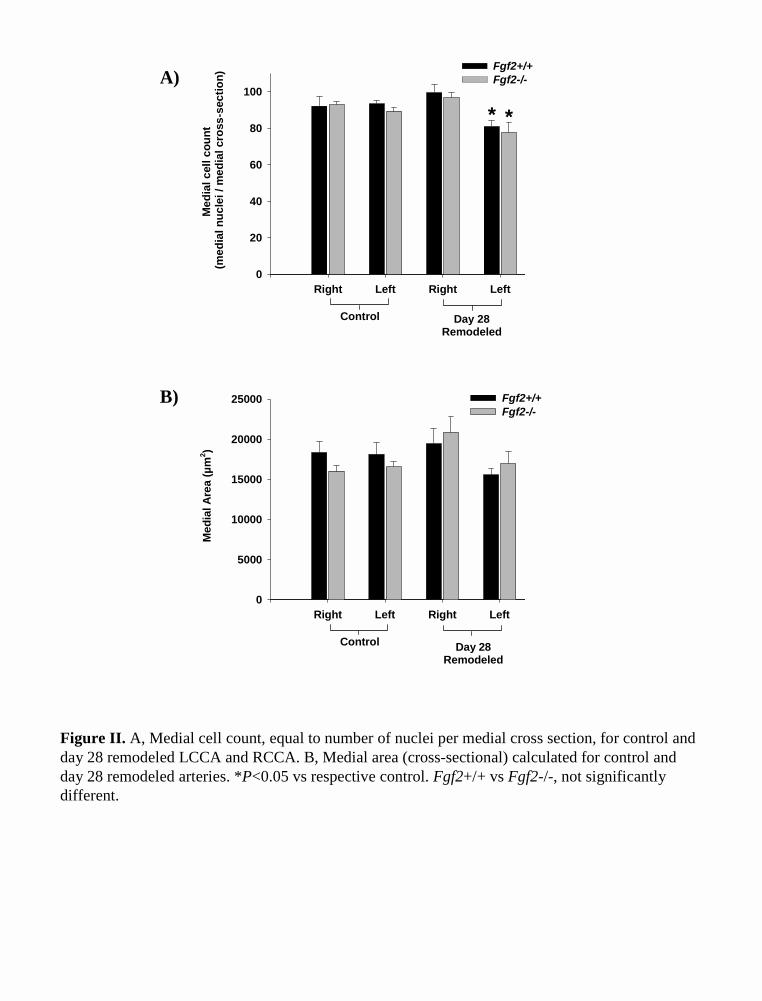
Vascular Cell TurnoverExamining the terminal deoxynucleotidyl transferase–medi-ated dUTP nick end-labeling (TUNEL) index in the mediallayer of day-4 remodeled vessels showed increased apoptosisin the RCCA and LCCA. Concomitantly, changes in theBrdU index showed an increased rate of cell proliferation inboth remodeled arteries at day 4 (Figure 4). The single intimalcell layer displayed similar changes in cell turnover (data notshown), and positive BrdU and TUNEL staining was ob-served on the luminal side of the internal elastic lamina at day4. To determine the net change in smooth muscle cellnumber, we counted the total nuclei per medial cross sectionat day 28. This showed a significantly reduced medial cellcount at day 28 in the low-flow LCCA of both genotypes.However, medial cell density did not change in the day-28remodeled arteries (Figure 4). The RCCA showed a trend ofincreased medial cell number at day 28 in both genotypes, but
Figure 4. A, Quantification of medial cell proliferation (n53 per genotype) for control and day-4 remodeled LCCA and RCCA. B, Quanti-fication of medial cell apoptosis (n53 per genotype) for control and day-4 remodeled arteries. C, Medial cell density for control andday-28 remodeled arteries. *P,0.05 vs respective control; P5NS for Fgf21/1 vs Fgf22/2.
4 Arterioscler Thromb Vasc Biol. July 2002
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from

this difference was not significant. No differences wereobserved betweenFgf21/1 andFgf22/2 mice.
DiscussionPrevious studies have implicated FGF2 in a wide variety ofvascular cell-signaling processes, including proliferation, dif-ferentiation, and migration (see review33). Specific to flow-dependent remodeling, arterial endothelial cells upregulateFgf2 mRNA levels in response to fluid shear stress,17 andFGF2 expression increases in the vascular wall during flow-induced arterial enlargement.18 In addition, FGF2 regulatesthe expression of molecules involved in extracellular matrixremodeling,24 which is an important component of arterialwall reorganization.1,2 Furthermore, FGF2 is also thought tobe an important regulator of endothelial cell and smoothmuscle cell proliferation22,34–36and apoptosis.20,21,37The mostdirect evidence to date supporting a role for FGF2 in arterialremodeling is the attenuation of inward remodeling byantibody neutralization of endogenous FGF2 in a mousemodel of carotid artery flow cessation.19 Despite this largebody of evidence, we did not to observe an essential role ofFGF2 in flow-dependent carotid artery remodeling, given theequivalent remodeling responses ofFgf21/1 and Fgf22/2
mice. Thus, the results of the present study do not support thehypothesis that FGF2 is required for large-artery restructuringin response to chronically decreased or increased blood flow.
The apparent contradiction between our results and thoseof the previous study in which antibodies to FGF2 attenuatedinward remodeling after complete LCCA ligation in mice19
may simply be due to the differences between the 2 models(flow-cessation versus low-flow remodeling) and the distinctstimuli present in each model. In the flow-cessation model,originally published by Kumar and Lindner,30 net forwardblood flow was completely interrupted, resulting in bloodstasis in the LCCA. By contrast, our procedures causesubstantial blood flow reduction, but forward flow is main-tained within the LCCA via the patent thyroid artery. Aftercomplete ligation, a gradient of increased intimal lesionformation is observed toward the clotted ligation site.30
Depending on the strain used, varying amounts of neointimallesion formation and/or inward remodeling are observed inthe ligated artery.38 Also, it has been reported that theendothelial layer, although intact, detaches from the internalelastic lamina in the ligated LCCA.30 This exposes the highlythrombogenic extracellular matrix and may increase theactivation of blood components, such as platelets.38 Thepossibility exists that additional factors, not present in thelow-flow situation, contribute to the carotid artery responsesinduced by complete flow cessation. Platelet activation,hypoxia, metabolite accumulation, and/or inflammation couldpotentially influence the remodeling response in the com-pletely ligated artery.38–40Such a stimulus, possibly unique tothe no-flow condition, may require FGF2 signaling to induceinward remodeling. Alternatively, animals with a chronicgene ablation (eg, knockout mice) and animals with an acuteloss of a gene product (eg, antibody neutralization) maysimply have different responses to a given stimulus. Last, it ispossible that FGF2-neutralizing antibodies are cross-reacting
with other FGFs, given that there are at least 23 known FGFfamily members.41
The changes in flow and the resulting carotid remodelingobserved in the present study in the mouse are comparable tothose in prior studies in the rat27,28 but different from aprevious study in the mouse in which blood flow was onlymoderately reduced and the LCCA diameter was decreasedby just 8% to 10%.29,31 The LCCA branch ligations, per-formed in the present study, reduced flow in the LCCA by'80% while increasing the contralateral RCCA flow by 40%.The comparable procedures in the rat reduced LCCA flow by90% and increased RCCA flow by 45%.27 These flowchanges in the rat caused a 16% reduction in LCCA outerdiameter and an 11% increase in RCCA outer diameter after4 weeks of remodeling. In comparison, wild-type mice in thepresent study showed an'23% reduction in LCCA vesseldiameter and a 13% increase in RCCA vessel diameter at day28. In rabbits, ligation of the left external carotid arterydecreased LCCA blood flow by 70%, causing a 21% reduc-tion of LCCA luminal diameter after 2 weeks.6 Overall, ourresults in the mouse are consistent with previous studies offlow-dependent remodeling in other species, demonstratingthat chronically increased blood flow leads to arterial enlarge-ment (outward remodeling), whereas blood flow reductionresults in arterial narrowing (inward remodeling).27,42–44
Associated with structural remodeling in this model isincreased vascular cell turnover, as indicated by increasedapoptosis and proliferation in the low-flow LCCA and thehigh-flow RCCA early in the remodeling process. Previousinvestigators have observed increased BrdU labeling andincreased apoptosis in vessels after chronic flow reduc-tion.28,29,44 Similar to others, we demonstrate a net loss ofvascular cells after chronic flow reduction in the mouse.31
Specifically, we observed a reduced medial smooth musclecell count in the low-flow LCCA at day 28. However, medialsmooth muscle density remained unchanged because of thenoticeable trend toward reduced medial CSA in the LCCA. Inthe high-flow RCCA, apoptosis and proliferation increased,but there was not a significant change in medial cell count,density, or CSA compared with control conditions. A study offlow remodeling in the rat mesentery showed that increasedapoptosis and proliferation occurred simultaneously in high-flow exposed resistance arteries and that this was coupled toan increase in medial CSA of high-flow arteries.44 Miyashiroet al27 showed no change in medial CSA in the low-flowcarotid artery of juvenile rats, whereas the high-flow carotidartery had increased medial CSA. Taken as a whole, thepresent results are in agreement with the concept that chronicchanges in blood flow result in dynamic changes in vascularcell turnover,1,27 although we observed a constancy in medialcell density in both flow conditions.
The specific molecules regulating vascular cell growth orapoptosis during flow remodeling are largely unknown.Carotid arteries of endothelial NO synthase (eNOS) knockout(eNOS2/2) mice subjected to chronically reduced flowshowed increased vascular cell proliferation and cell numbercompared with carotid arteries of eNOS wild-type (eNOS1/1)mice. Thus, endothelium-derived NO may be an essentialcontroller of vascular cell turnover during flow-dependent
Sullivan and Hoying Flow-Dependent Carotid Remodeling 5
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from

carotid remodeling. Previous studies show that FGF2 canmediate endothelial cell and smooth muscle cell prolifera-tion22,34–36and apoptosis.20,21,37Interestingly, FGF2 has beenshown to stimulate eNOS mRNA expression and eNOSprotein production in cultured endothelial cells.23,45 Also, ithas been shown that NO promotes proliferation of in vitroendothelial cells through endogenous FGF2.24 However,there was no difference betweenFgf21/1 andFgf22/2 mice inthe present study that would indicate that vascular cellturnover was affected by lack of FGF2. Thus, FGF2 does notappear to be an essential mediator acting upstream or down-stream of NO signaling in this model. In terms of vascularcell proliferation, it has previously been shown that thecarotid arteries ofFgf22/2 mice undergo a normal hyperplas-tic response after intra-arterial mechanical injury.26 Also,intimal area and cellularity were not affected by FGF2antibody in the ligated mouse carotid artery, suggesting thatsmooth muscle proliferation was not altered by FGF2neutralization.19
Overall, the apparently normal remodeling responses ob-served inFgf22/2 mice may reflect compensation for the lossof FGF2 by another gene product. There are numerous FGFfamily members, and these proteins bind to a common groupof receptors, although with differing affinities.33 Thus, it ispossible that$1 FGF protein could be compensating for thedisruption of theFgf2 gene. Recently, a double knockout ofFGF1 and FGF2 was shown to have the same phenotype asFgf22/2 mice.46 This suggests that FGF1, the FGF familymember most closely related to FGF2, is not compensatingfor the loss of FGF2 in situations such as development andwound healing.46 On the other hand, it is possible that there isnot compensation and that other growth factors or moleculesmay be the actual endogenous mediators of processes cur-rently ascribed to FGF2 (eg, flow-dependent remodeling). Inthis regard, changes in FGF2 expression may be mediatingsome other event during arterial remodeling that is eitherunrelated or not critical for structural changes in the artery.
It is important to note that considerable strain variability inthe vascular responses of mice to various challenges has beendescribed.38,47 More specifically, Harmon et al,38 using theLCCA flow-cessation model in mice, demonstrated a largedegree of strain-dependent variability in carotid remodelingof the ligated LCCA. Additionally, they showed that not allstrains displayed significant outward remodeling of the con-tralateral RCCA. Others using the flow-cessation modelobserved no RCCA enlargement despite measuring a neardoubling of RCCA blood flow in 129 SV mice.48 Thus, it isreasonable to expect that there might be strain-specificdifferences in the extent and character of vessel remodelingwhen the model presented in the present study is used. Inpreliminary experiments, we noted that the LCCA and RCCAof FVB/NJ and C57BL/6J mice had been remodeled to aextent similar to that observed inFgf21/1 andFgf22/2 mice,which are on a mixed background of 50% Black Swiss and50% 129 SV (data not shown). These other strains showedinward remodeling with minimal neointimal lesion formationin the LCCA. When a neointima was observed in these mice,it was typically only 2 or 3 cell layers thick (data not shown).
We also noticed strain-dependent variations in the carotidartery architecture (ie, position of branching vessels).
In conclusion, we describe a model of bilateral carotidremodeling in the mouse. In a single mouse, the simultaneousreduction in blood flow in the LCCA and increase in bloodflow in the RCCA provide a powerful research tool toeffectively examine the molecular mechanisms of arteryremodeling. With this model, we show that lack of FGF2does not affect structural remodeling of large arteries inresponse to chronically altered blood flow. FGF2 appearsdispensable during flow-dependent remodeling of the arterywall and does not significantly regulate vascular cell turnoverin this model.
AcknowledgmentsThis study was supported by an American Heart Association,Desert/Mountain Affiliate, Predoctoral Fellowship (No. 9910147Z toC.J.S.) and National Institutes of Health grant HL-63732 (J.B.H.).We thank Kim Heiman for her assistance with data collection andanalysis.
References1. Langille BL. Remodeling of developing and mature arteries: endotheli-
um, smooth muscle, and matrix.J Cardiovasc Pharmacol. 1993;21(suppl1):S11–S17.
2. Gibbons GH, Dzau VJ. The emerging concept of vascular remodeling.N Engl J Med. 1994;330:1431–1438.
3. Ward MR, Pasterkamp G, Yeung AC, Borst C. Arterial remodeling:mechanisms and clinical implications.Circulation. 2000;102:1186–1191.
4. Korsgaard N, Aalkjaer C, Heagerty AM, Izzard AS, Mulvany MJ. His-tology of subcutaneous small arteries from patients with essential hyper-tension.Hypertension. 1993;22:523–526.
5. Kakuta T, Currier JW, Haudenschild CC, Ryan TJ, Faxon DP. Dif-ferences in compensatory vessel enlargement, not intimal formation,account for restenosis after angioplasty in the hypercholesterolemic rabbitmodel.Circulation. 1994;89:2809–2815.
6. Langille BL, O’Donnell F. Reductions in arterial diameter produced bychronic decreases in blood flow are endothelium-dependent.Science.1986;231:405–407.
7. Guyton JR, Hartley CJ. Flow restriction of one carotid artery in juvenilerats inhibits growth of arterial diameter.Am J Physiol. 1985;248:H540–H546.
8. Miyachi M, Tanaka H, Yamamoto K, Yoshioka A, Takahashi K, OnoderaS. Effects of one-legged endurance training on femoral arterial andvenous size in healthy humans.J Appl Physiol. 2001;90:2439–2444.
9. Krams R, Wentzel JJ, Oomen JA, Schuurbiers JC, Andhyiswara I, KloetJ, Post M, de Smet B, Borst C, Slager CJ, Serruys PW. Shear stress inatherosclerosis, and vascular remodelling.Semin Interv Cardiol. 1998;3:39–44.
10. Ward MR, Jeremias A, Huegel H, Fitzgerald PJ, Yeung AC. Accentuatedremodeling on the upstream side of atherosclerotic lesions.Am J Cardiol.2000;85:523–526.
11. Ward MR, Tsao PS, Agrotis A, Dilley RJ, Jennings GL, Bobik A. Lowblood flow after angioplasty augments mechanisms of restenosis: inwardvessel remodeling, cell migration, and activity of genes regulatingmigration.Arterioscler Thromb Vasc Biol. 2001;21:208–213.
12. Wentzel JJ, Kloet J, Andhyiswara I, Oomen JA, Schuurbiers JC, de SmetBJ, Post MJ, de Kleijn D, Pasterkamp G, Borst C, Slager CJ, Krams R.Shear-stress and wall-stress regulation of vascular remodeling afterballoon angioplasty: effect of matrix metalloproteinase inhibition.Circu-lation. 2001;104:91–96.
13. Glagov S, Weisenberg E, Zarins CK, Stankunavicius R, KolettisGJ. Compensatory enlargement of human atherosclerotic coronaryarteries.N Engl J Med. 1987;316:1371–1375.
14. Pasterkamp G, Wensing PJ, Post MJ, Hillen B, Mali WP, Borst C.Paradoxical arterial wall shrinkage may contribute to luminal narrowingof human atherosclerotic femoral arteries.Circulation. 1995;91:1444–1449.
15. Pasterkamp G, Schoneveld AH, Hijnen DJ, de Kleijn DP, Teepen H, vander Wal AC, Borst C. Atherosclerotic arterial remodeling and the local-
6 Arterioscler Thromb Vasc Biol. July 2002
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from

ization of macrophages and matrix metalloproteases 1, 2 and 9 in thehuman coronary artery.Atherosclerosis. 2000;150:245–253.
16. Mintz GS, Popma JJ, Pichard AD, Kent KM, Satler LF, Wong C, HongMK, Kovach JA, Leon MB. Arterial remodeling after coronary angio-plasty: a serial intravascular ultrasound study.Circulation. 1996;94:35–43.
17. Malek A, Gibbons G, Dzau V, Izumo S. Fluid shear stress differentiallymodulates expression of genes encoding basic fibroblast growth factorand platelet-derived growth factor B chain in vascular endothelium.J ClinInvest. 1993;92:2013–2021.
18. Singh T, Abe K, Sasaki T, Zhuang Y, Masuda H, Zarins C. Basicfibroblast growth factor expression precedes flow-induced arterialenlargement.J Surg Res. 1998;77:165–173.
19. Bryant SR, Bjercke RJ, Erichsen DA, Rege A, Lindner V. Vascularremodeling in response to altered blood flow is mediated by fibroblastgrowth factor-2.Circ Res. 1999;84:323–328.
20. Fox JC, Shanley JR. Antisense inhibition of basic fibroblast growth factorinduces apoptosis in vascular smooth muscle cells.J Biol Chem. 1996;271:12578–12584.
21. Karsan A, Yee E, Poirier GG, Zhou P, Craig R, Harlan JM. Fibroblastgrowth factor-2 inhibits endothelial cell apoptosis by Bcl-2-dependentand independent mechanisms.Am J Pathol. 1997;151:1775–1784.
22. Lindner V, Reidy MA. Proliferation of smooth muscle cells after vascularinjury is inhibited by an antibody against basic fibroblast growth factor.Proc Natl Acad Sci U S A. 1991;88:3739–3743.
23. Kostyk SK, Kourembanas S, Wheeler EL, Medeiros D, McQuillan LP,D’Amore PA, Braunhut SJ. Basic fibroblast growth factor increases nitricoxide synthase production in bovine endothelial cells.Am J Physiol.1995;269:H1583–H1589.
24. Ziche M, Parenti A, Ledda F, Dell’Era P, Granger HJ, Maggi CA, PrestaM. Nitric oxide promotes proliferation and plasminogen activator pro-duction by coronary venular endothelium through endogenous bFGF.Circ Res. 1997;80:845–852.
25. Cai W, Vosschulte R, Afsah-Hedjri A, Koltai S, Kocsis E, Scholz D,Kostin S, Schaper W, Schaper J. Altered balance between extracellularproteolysis and antiproteolysis is associated with adaptive coronary arte-riogenesis.J Mol Cell Cardiol. 2000;32:997–1011.
26. Zhou M, Sutliff RL, Paul RJ, Lorenz JN, Hoying JB, Haudenschild CC,Yin M, Coffin JD, Kong L, Kranias EG, Luo W, Boivin GP, Duffy JJ,Pawlowski SA, Doetschman T. Fibroblast growth factor 2 control ofvascular tone.Nat Med. 1998;4:201–207.
27. Miyashiro JK, Poppa V, Berk BC. Flow-induced vascular remodeling inthe rat carotid artery diminishes with age.Circ Res. 1997;81:311–319.
28. Mondy JS, Lindner V, Miyashiro JK, Berk BC, Dean RH, Geary RL.Platelet-derived growth factor ligand and receptor expression in responseto altered blood flow in vivo.Circ Res. 1997;81:320–327.
29. Rudic RD, Bucci M, Fulton D, Segal SS, Sessa WC. Temporal eventsunderlying arterial remodeling after chronic flow reduction in mice:correlation of structural changes with a deficit in basal nitric oxidesynthesis.Circ Res. 2000;86:1160–1166.
30. Kumar A, Lindner V. Remodeling with neointima formation in the mousecarotid artery after cessation of blood flow.Arterioscler Thromb VascBiol. 1997;17:2238–2244.
31. Rudic RD, Shesely EG, Maeda N, Smithies O, Segal SS, Sessa WC.Direct evidence for the importance of endothelium-derived nitric oxide invascular remodeling.J Clin Invest. 1998;101:731–736.
32. Couffinhal T, Silver M, Zheng LP, Kearney M, Witzenbichler B, IsnerJM. Mouse model of angiogenesis.Am J Pathol. 1998;152:1667–1679.
33. Bikfalvi A, Klein S, Pintucci G, Rifkin D. Biological roles of fibroblastgrowth factor-2.Endocr Rev. 1997;18:26–45.
34. Lindner V, Majack RA, Reidy MA. Basic fibroblast growth factor stim-ulates endothelial regrowth and proliferation in denuded arteries.J ClinInvest. 1990;85:2004–2008.
35. Gospodarowicz D, Ferrara N, Haaparanta T, Neufeld G. Basic fibroblastgrowth factor: expression in cultured bovine vascular smooth musclecells.Eur J Cell Biol. 1988;46:144–151.
36. Itoh H, Mukoyama M, Pratt RE, Dzau VJ. Specific blockade of basicfibroblast growth factor gene expression in endothelial cells by antisenseoligonucleotide.Biochem Biophys Res Commun. 1992;188:1205–1213.
37. Kondo S, Yin D, Aoki T, Takahashi JA, Morimura T, Takeuchi J. bcl-2gene prevents apoptosis of basic fibroblast growth factor-deprived murineaortic endothelial cells.Exp Cell Res. 1994;213:428–432.
38. Harmon KJ, Couper LL, Lindner V. Strain-dependent vascularremodeling phenotypes in inbred mice.Am J Pathol. 2000;156:1741–1748.
39. Kumar A, Hoover JL, Simmons CA, Lindner V, Shebuski RJ.Remodeling and neointimal formation in the carotid artery of normal andP-selectin-deficient mice.Circulation. 1997;96:4333–4342.
40. Kawasaki T, Dewerchin M, Lijnen HR, Vreys I, Vermylen J, HoylaertsMF. Mouse carotid artery ligation induces platelet-leukocyte–dependentluminal fibrin, required for neointima development.Circ Res. 2001;88:159–166.
41. Yamashita T, Yoshioka M, Itoh N. Identification of a novel fibroblastgrowth factor, FGF-23, preferentially expressed in the ventrolateralthalamic nucleus of the brain.Biochem Biophys Res Commun. 2000;277:494–498.
42. Kamiya A, Togawa T. Adaptive regulation of wall shear stress to flowchange in the canine carotid artery.Am J Physiol. 1980;239:H14–H21.
43. Langille BL, Bendeck MP, Keeley FW. Adaptations of carotid arteries ofyoung and mature rabbits to reduced carotid blood flow.Am J Physiol.1989;256:H931–H939.
44. Buus CL, Pourageaud F, Fazzi GE, Janssen G, Mulvany MJ, De Mey JG.Smooth muscle cell changes during flow-related remodeling of rat mes-enteric resistance arteries.Circ Res. 2001;89:180–186.
45. Babaei S, Teichert-Kuliszewska K, Monge JC, Mohamed F, BendeckMP, Stewart DJ. Role of nitric oxide in the angiogenic response in vitroto basic fibroblast growth factor.Circ Res. 1998;82:1007–1015.
46. Miller DL, Ortega S, Bashayan O, Basch R, Basilico C. Compensation byfibroblast growth factor 1 (FGF1) does not account for the mild phe-notypic defects observed in FGF2 null mice.Mol Cell Biol. 2000;20:2260–2268.
47. Rohan RM, Fernandez A, Udagawa T, Yuan J, D’Amato RJ. Geneticheterogeneity of angiogenesis in mice.FASEB J. 2000;14:871–876.
48. Schiffers PM, Henrion D, Boulanger CM, Colucci-Guyon E,Langa-Vuves F, van Essen H, Fazzi GE, Levy BI, De Mey JG. Alteredflow-induced arterial remodeling in vimentin-deficient mice.ArteriosclerThromb Vasc Biol. 2000;20:611–616.
Sullivan and Hoying Flow-Dependent Carotid Remodeling 7
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from

Chris J. Sullivan and James B. HoyingKnockout Mice
Flow-Dependent Remodeling in the Carotid Artery of Fibroblast Growth Factor-2
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 2002 American Heart Association, Inc. All rights reserved.
Greenville Avenue, Dallas, TX 75231is published by the American Heart Association, 7272Arteriosclerosis, Thrombosis, and Vascular Biology published online May 23, 2002;Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/early/2002/05/23/01.ATV.0000023230.17493.E3.citationWorld Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org/content/suppl/2002/07/11/22.7.1100.DC1Data Supplement (unedited) at:
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document. Question and AnswerPermissions and Rightspage under Services. Further information about this process is available in the
which permission is being requested is located, click Request Permissions in the middle column of the WebCopyright Clearance Center, not the Editorial Office. Once the online version of the published article for
can be obtained via RightsLink, a service of theArteriosclerosis, Thrombosis, and Vascular Biologyin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from

Online Only Data Supplement (Tables I, II, & III)
Table I. Blood flow changes before and after ligation of LCCA branches in Fgf2+/+ and Fgf2-/- mice.
Average blood flow (ml/min) in the LCCA (left) and RCCA (right) of Fgf2+/+ and Fgf2-/- mice
before ligation of LCCA branches, immediately after the ligations (day 0), and again at 14 days
after the ligations (day 14). Values are mean ± SEM *P<0.05 vs respective control (before)
values. Fgf2+/+ vs Fgf2-/-, not significantly different.
Control Day 0 Day 14
Right Left Right Left Right Left
Fgf2+/+ (n)
0.77 ± 0.02 (3)
0.74 ± 0.04 (3)
1.11 ± 0.05* (3)
0.06 ± 0.02* (3)
1.33 ± 0.07* (3)
0.16 ± 0.03* (3)
Fgf2-/- (n)
0.73 ± 0.05 (3)
0.73 ± 0.07 (3)
1.03 ± 0.01* (3)
0.12 ± 0.01* (3)
1.34 ± 0.03* (3)
0.12 ± 0.02* (3)
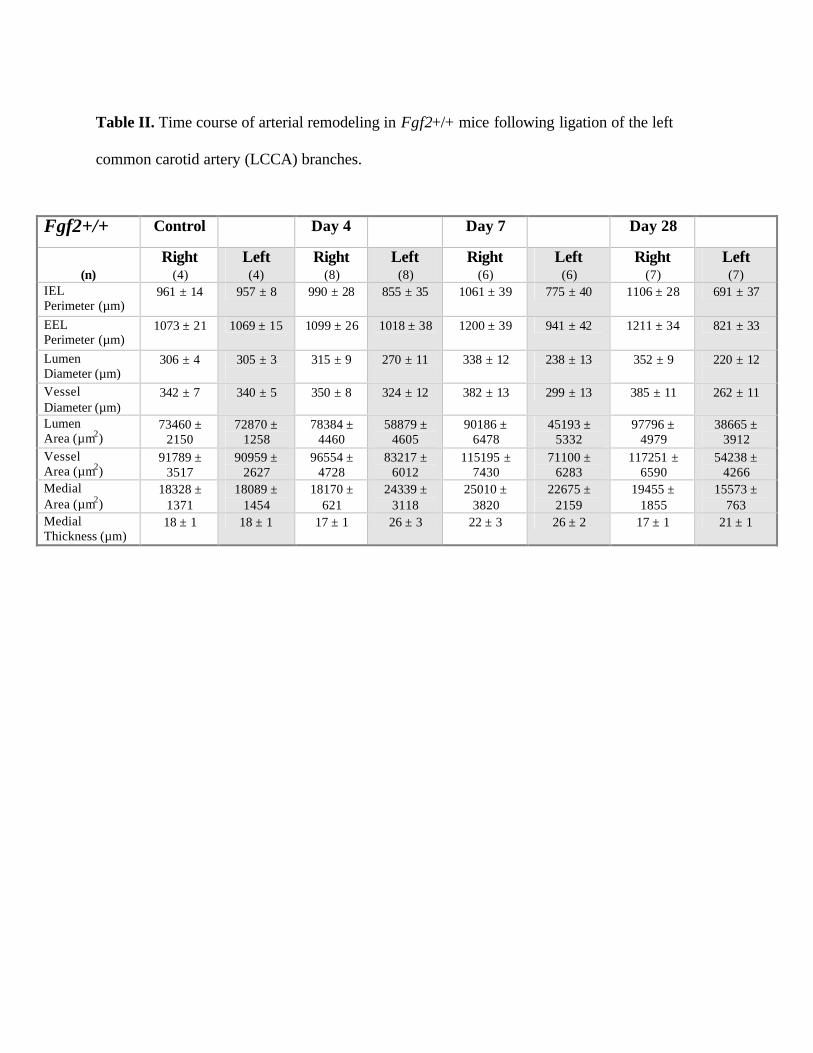
Table II. Time course of arterial remodeling in Fgf2+/+ mice following ligation of the left
common carotid artery (LCCA) branches.
Fgf2+/+ Control Day 4 Day 7 Day 28
(n)
Right (4)
Left (4)
Right (8)
Left (8)
Right (6)
Left (6)
Right (7)
Left (7)
IEL Perimeter (µm)
961 ± 14 957 ± 8 990 ± 28 855 ± 35 1061 ± 39 775 ± 40 1106 ± 28 691 ± 37
EEL Perimeter (µm)
1073 ± 21 1069 ± 15 1099 ± 26 1018 ± 38 1200 ± 39 941 ± 42 1211 ± 34 821 ± 33
Lumen Diameter (µm)
306 ± 4
305 ± 3
315 ± 9
270 ± 11
338 ± 12
238 ± 13
352 ± 9
220 ± 12
Vessel Diameter (µm)
342 ± 7 340 ± 5 350 ± 8 324 ± 12 382 ± 13 299 ± 13 385 ± 11 262 ± 11
Lumen Area (µm2)
73460 ± 2150
72870 ± 1258
78384 ± 4460
58879 ± 4605
90186 ± 6478
45193 ± 5332
97796 ± 4979
38665 ± 3912
Vessel Area (µm2)
91789 ± 3517
90959 ± 2627
96554 ± 4728
83217 ± 6012
115195 ± 7430
71100 ± 6283
117251 ± 6590
54238 ± 4266
Medial Area (µm2)
18328 ± 1371
18089 ± 1454
18170 ± 621
24339 ± 3118
25010 ± 3820
22675 ± 2159
19455 ± 1855
15573 ± 763
Medial Thickness (µm)
18 ± 1 18 ± 1 17 ± 1 26 ± 3 22 ± 3 26 ± 2 17 ± 1 21 ± 1
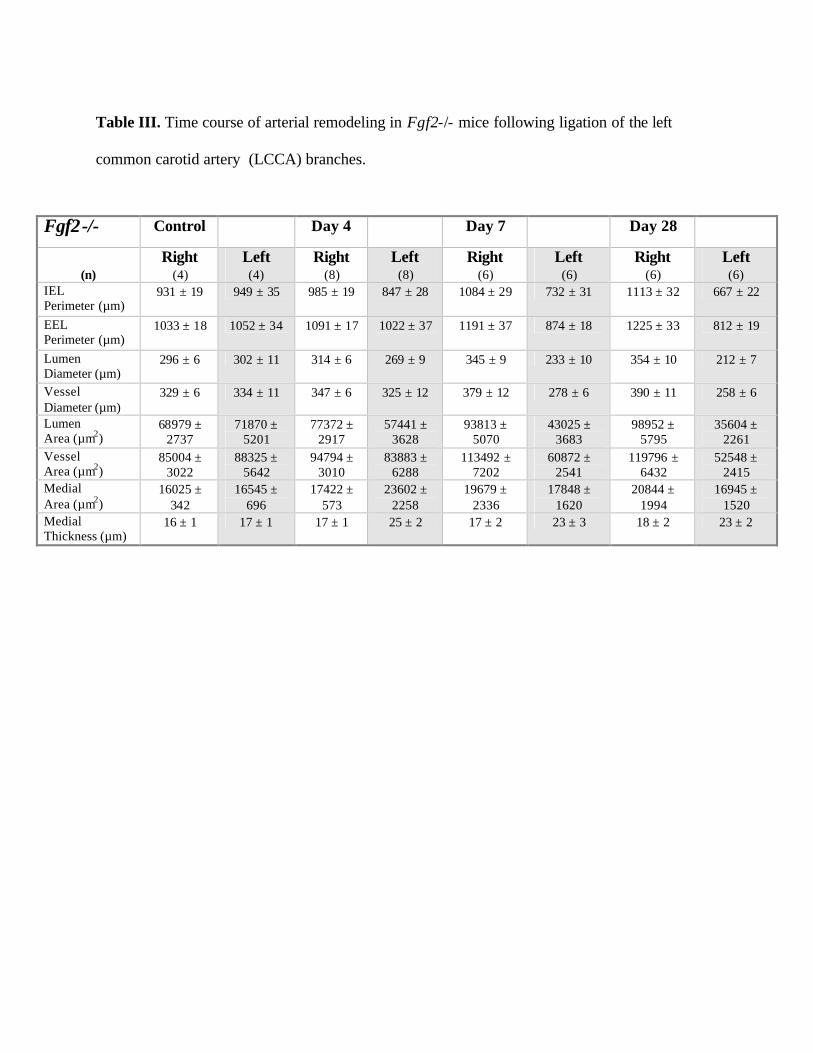
Table III. Time course of arterial remodeling in Fgf2-/- mice following ligation of the left
common carotid artery (LCCA) branches.
Fgf2-/- Control Day 4 Day 7 Day 28
(n)
Right (4)
Left (4)
Right (8)
Left (8)
Right (6)
Left (6)
Right (6)
Left (6)
IEL Perimeter (µm)
931 ± 19 949 ± 35 985 ± 19 847 ± 28 1084 ± 29 732 ± 31 1113 ± 32 667 ± 22
EEL Perimeter (µm)
1033 ± 18 1052 ± 34 1091 ± 17 1022 ± 37 1191 ± 37 874 ± 18 1225 ± 33 812 ± 19
Lumen Diameter (µm)
296 ± 6 302 ± 11 314 ± 6 269 ± 9 345 ± 9 233 ± 10 354 ± 10 212 ± 7
Vessel Diameter (µm)
329 ± 6 334 ± 11 347 ± 6 325 ± 12 379 ± 12 278 ± 6 390 ± 11 258 ± 6
Lumen Area (µm2)
68979 ± 2737
71870 ± 5201
77372 ± 2917
57441 ± 3628
93813 ± 5070
43025 ± 3683
98952 ± 5795
35604 ± 2261
Vessel Area (µm2)
85004 ± 3022
88325 ± 5642
94794 ± 3010
83883 ± 6288
113492 ± 7202
60872 ± 2541
119796 ± 6432
52548 ± 2415
Medial Area (µm2)
16025 ± 342
16545 ± 696
17422 ± 573
23602 ± 2258
19679 ± 2336
17848 ± 1620
20844 ± 1994
16945 ± 1520
Medial Thickness (µm)
16 ± 1 17 ± 1 17 ± 1 25 ± 2 17 ± 2 23 ± 3 18 ± 2 23 ± 2
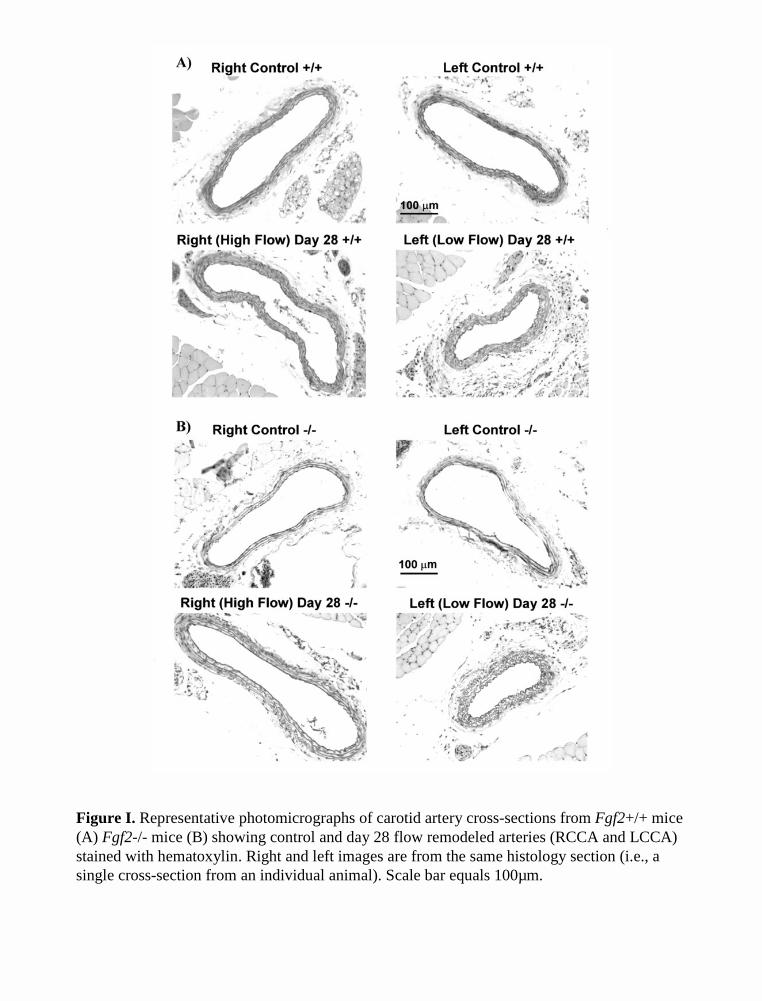
Figure I. Representative photomicrographs of carotid artery cross-sections from Fgf2+/+ mice (A) Fgf2-/- mice (B) showing control and day 28 flow remodeled arteries (RCCA and LCCA) stained with hematoxylin. Right and left images are from the same histology section (i.e., a single cross-section from an individual animal). Scale bar equals 100µm.
A)
B)
Right Left Right Left
Med
ial c
ell c
ount
(m
edia
l nuc
lei /
med
ial c
ross
-sec
tion)
0
20
40
60
80
100
Day 28Remodeled
Control
**
Fgf2+/+Fgf2-/-
Right Left Right Left
Med
ial A
rea
(µm
2 )
0
5000
10000
15000
20000
25000 Fgf2+/+Fgf2-/-
Control Day 28Remodeled
Figure II. A, Medial cell count, equal to number of nuclei per medial cross section, for control and day 28 remodeled LCCA and RCCA. B, Medial area (cross-sectional) calculated for control and day 28 remodeled arteries. *P<0.05 vs respective control. Fgf2+/+ vs Fgf2-/-, not significantly different.
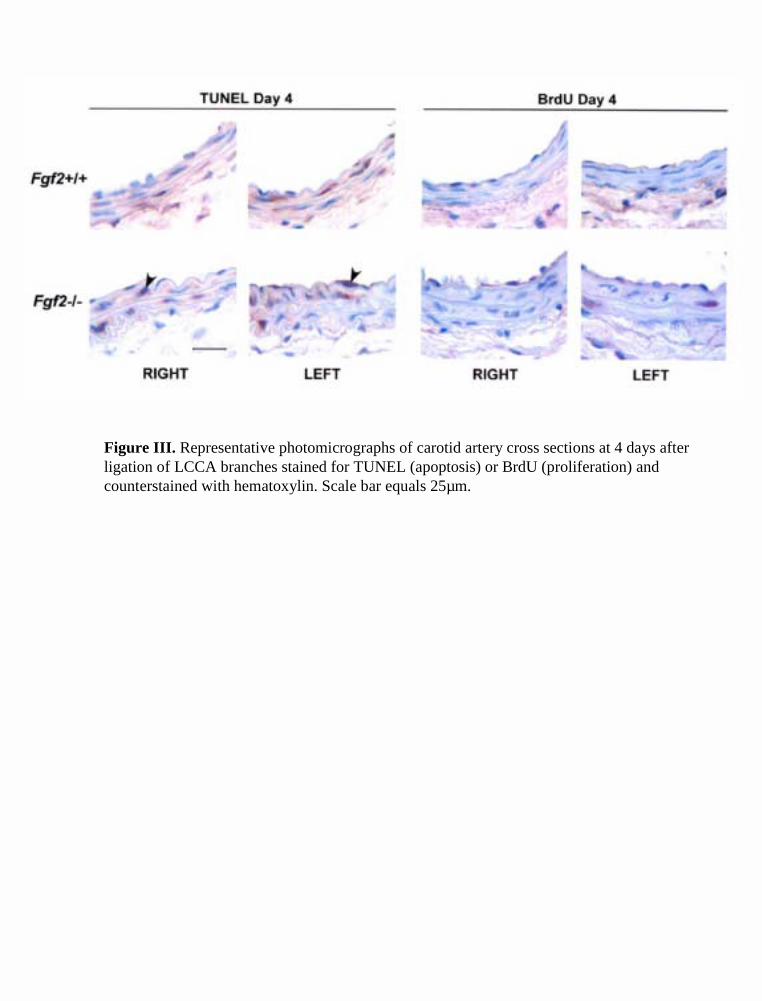
Figure III. Representative photomicrographs of carotid artery cross sections at 4 days after ligation of LCCA branches stained for TUNEL (apoptosis) or BrdU (proliferation) and counterstained with hematoxylin. Scale bar equals 25µm.

MATERIALS AND METHODS (Online Supplement)
Experimental animals
Male Fgf2+/+ and Fgf2-/- mice1 (50% Black Swiss and 50% 129 SV) were used for all
experiments according to the University of Arizona IACUC approved procedures. All mice were
genotyped by PCR using primers specific for the Fgf2 wildtype allele (forward, 5’–
GCTGTACACTCAAGGGGCTC –3’; reverse, 5’– CGCCGTTCTTGCAGTAGAG –3’) and the
Fgf2 knockout allele (forward, 5’– TCCAAAGCCTGACTTGATCC –3’; reverse, 5’–
CTGACTAGGGGAGGAGTAGAAGG –3’), following collection of genomic DNA from tail
clips. Procedures were timed so that all mice were 8 weeks of age (± 4 days) at the time of
sacrifice. Mice were anesthetized with 2.5% Avertin (2.5% 2,2,2-tribromoethanol, 2.5% tert-
amyl alcohol in PBS: Aldrich) at a dose of 0.15 ml per 10 gram body weight injected
intraperitoneally.
Surgery to induce flow-dependent carotid remodeling
The mouse model presented is a modification of procedures previously published for use in the
rat.2,3 Mice were anesthetized and a midline incision was made along the neck. All branches
originating from the left common carotid artery (LCCA), except for the left thyroid artery, were
ligated (6.0 silk) in order to reduce flow in the LCCA and increase flow in the contralateral right
common carotid artery (RCCA). Specifically, the distal portion of the left external carotid was
ligated where it bifurcates while the internal carotid and occipital arteries were ligated with a
single suture at their origin (Figure 1). Mice were sacrificed at day 4, 7, and 28 following

2
surgical ligation. Age and sex matched control animals were sacrificed without having
undergone ligation.
Carotid artery blood flow
Carotid artery (RCCA and LCCA) blood flow was measured using an ultrasonic transit-time
flowmeter (Transonic Systems, Inc.) with a 0.5 V-series probe as described previously.4 Blood
flow was evaluated in mice (n=3 per genotype) before ligation, immediately after ligation (day
0), and again at day 14 after ligation. Average volume flow (ml/min) was recorded for 5 min in
each artery. The values presented are the average for the 5 min data acquisition period. After
ligation of the LCCA branches, blood flow measurements were repeated after a stabilization
period of at least 15 min. The incision was closed and mice were allowed to recover. After 14
days, the mice were reanesthetized and blood flow was evaluated for a final time.
Morphometry
Control (n=4 per genotype), day 7 (n=6 per genotype), and day 28 (+/+ n=7; -/- n=6) animals
were perfuse fixed through a polyethylene catheter placed in the left ventricle. Animals were
perfused at constant pressure (90-100 mmHg) with 20 ml of heparinized PBS followed by 10 ml
of 10% phosphate buffered formalin. The neck, between the clavicle and mandible, was isolated
and placed in fixative overnight. Neck sections were then decalcified using Decalcifier I and II
(Surgipath) for 24 hr each. Decalcified necks were processed, paraffin embedded, and serial
sections cut (8 µm) for morphometric analysis. Cross sections of the entire neck were stained
using hematoxylin. Morphometric analysis was carried out on the RCCA and LCCA of each
animal from 2 whole neck cross sections approximately 160-200 µm apart, cut at approximately

3
the mid-portion of the common carotid artery. Digitized images were analyzed using image
analysis software (Scionimage 4.0). The perimeter (length) of the lumen, internal elastic lamina
(IEL), and external elastic lamina (EEL) were measured and these values were used to calculate
various vessel dimensions, assuming the artery was a perfect circle.5,6
Angiography
Mice were anesthetized and subsequently overdosed with 2.5% Avertin following exposure of
the thoracic cavity and heart. The arterial circulation was perfused (constant pressure of 90-100
mmHg) with PBS containing 1x10-5 mol/L sodium nitroprusside through a tapered polyethylene
catheter (PE-90) inserted into the left ventricle followed by filling with contrast agent (barium
sulfate 210% w/v, Liqui-Coat, Lafayette Pharmaceuticals, Inc). Angiograms of the head and
neck region were obtained using a high-definition x-ray cabinet system (Faxitron).
Vascular cell proliferation and apoptosis
To examine proliferation, animals (n=3 per genotype) were injected with bromodeoxyuridine
(BrdU: 30mg/kg body weight: Sigma Chemical Co., St.Louis, MO) i.p. at 24 h and 12 h prior to
sacrifice on day 4 following LCCA surgery. Decalcification interfered with BrdU and TUNEL
staining. So, mice were perfuse fixed and the vertebrae along with all other bones were removed
by careful dissection prior to paraffin embedding. BrdU incorporation into the nuclei of
proliferating cells was identified on 6-µm sections7 using a peroxidase conjugated sheep anti-
BrdU antibody (Biodesign International, Kennebunk, ME). BrdU-positive nuclei were counted
per two whole vessel transverse sections from each artery. Proliferation is expressed as the
percentage of the total nuclei counted in the vessel cross-section that stain positive for BrdU.

4
Apoptotic cells were identified using Boehringer-Mannheim's In Situ Cell Death Detection Kit.
Apoptosis in each vessel was expressed as a percentage of total nuclei per vessel cross-section
that are labeled as apoptotic. Calculations (e.g., proliferation index) were performed for the
media (between the IEL and EEL), intima (lumen side of the IEL), and the whole vessel (intima
+ media).
Statistical analysis
Values are presented as the mean±SEM. Comparison between two means was done using
Student’s unpaired t-Test. Multiple groups were compared by One Way ANOVA with a Student-
Newman-Keuls Test. Comparison of carotid artery blood flow, within a genotype, before and
after ligation was done using a One Way Repeated Measures ANOVA with a Student-Newman-
Keuls Test. Statistical significance was set at P < 0.05.
REFERENCES
1. Zhou M, Sutliff RL, Paul RJ, Lorenz JN, Hoying JB, Haudenschild CC, Yin M, Coffin JD, Kong L, Kranias EG, Luo W, Boivin GP, Duffy JJ, Pawlowski SA, Doetschman T. Fibroblast growth factor 2 control of vascular tone. Nat Med. 1998;4(2):201-207.
2. Miyashiro JK, Poppa V, Berk BC. Flow-induced vascular remodeling in the rat carotid artery diminishes with age. Circ Res. 1997;81(3):311-319.
3. Mondy JS, Lindner V, Miyashiro JK, Berk BC, Dean RH, Geary RL. Platelet-derived growth factor ligand and receptor expression in response to altered blood flow in vivo. Circ Res. 1997;81(3):320-327.
4. Rudic RD, Bucci M, Fulton D, Segal SS, Sessa WC. Temporal events underlying arterial remodeling after chronic flow reduction in mice: correlation of structural changes with a deficit in basal nitric oxide synthesis. Circ Res. 2000;86(11):1160-1166.
5. Kumar A, Lindner V. Remodeling with neointima formation in the mouse carotid artery after cessation of blood flow. Arterioscler Thromb Vasc Biol. 1997;17(10):2238-2244.
6. Rudic RD, Shesely EG, Maeda N, Smithies O, Segal SS, Sessa WC. Direct evidence for the importance of endothelium-derived nitric oxide in vascular remodeling. J Clin Invest. 1998;101(4):731-736.
7. Couffinhal T, Silver M, Zheng LP, Kearney M, Witzenbichler B, Isner JM. Mouse model of angiogenesis. Am J Pathol. 1998;152(6):1667-1679.