Embed Size (px)
Citation preview

Bulletin 593 March 1955
Floral Morphology
of White Oal~
rhe Pennsylvania State University, College of Agriculture
Agricultural Experiment Station, University Park, I)crinsylvania

Digest
Both staminate and pistillate flowers of Q. IIllin are functionally IIlonosporangiate and have their origin during' vegetative growth of the year preceding the maturation of the fruit.
The development of the staminate inflorescence and the flower is initiated in late June. By October, it has developed to the overwintering condition. Sporogenous tissne is first noticeable in April of the year following its initiation and pollen is shed during' the last week of May.
The over-wintering condition of the staminate Hower would seem to preclude any low temperature damage, except as it might occur as a result of late frosts in the spring.
The pistillate inHorescence has its origin during the vegetative growth period of late SUlIlmer. However, because of the similarities to vegetative development, the primordium of the pistillate iunor· escence is hardly discernible until early October and may even be in doubt at that time.
Growth and differentiation are resumed early in l\Iay. At about the time of pollination, the pistillate Hower is only partly developed. Subseqpent to pollination, the development of the ovules, megasporo. genesis and gametogenesiB are cause for delay of syngamy for at least a month following' the dehiscence of the anthers.
Although not observed, fertilization must (l(:cur late in JUllC or early July. Embryo development was seen as early as Junc !) in one instance. This would indicate that considerable variation might well exist in the time schedule of development as set forth above.
The formation of an abscission layer at the base of the OV<Ll'Y, between its tissues and those of the involucre, was noted as early as the time of pollination. Even superficial observation of the shedding of immature oak fruits leaves little doubt that early abscission layer forination is cause fm' a relatively small and rather unpredictable mast crop.
The developing embryo sac follows the Adoxa type, with consider. able breakdown of the nucellar tissues and inner integument.

Floral Morphology of White Oak* HAROLD S. TURKIo;I" ALU:N L. REBUCK, ALVIN R. GROVE, JR.t
THAT THERE IS CONSIDERABLE VARIATION in the produc-tivity of white oak acorns has long been recognized. For any year
in which a substantial crop is produced, there are others in which there is little or no yield. This is, of course, not unique in white oak. But because of the relatively wide distribution and the potential value of white oak mast, the interest of botanists, foresters, and wildlife specialists has been stimulated.
An attempt has been made here to relate the development of the inllorescenccs and flowers to specific time periods. This fundamental information is prerequisite (.0 any additional exploration of the causes for failure of the crop.
The reasons which are generally offered for failure of fruit developmel,It :lppeal' to be largely a matter of conjecture. There is little, if any, baSIS III fact for them and certainly no proof.
I t has often been assumed that temperature might be the causative agent of low productivity. Low temperatures early in the spring might well be an important consideration. High temperatures during some critical period of development later in the summer-perhaps at the time of pollination 01' between the time of pollination and syngamymight be significant.
By relating-l1oral development to relatively specific times, it is hoped lhat additional investigations at critical periods might offer an acceptable reason or reasons for poor fruit set.
'M ost of the previous morphological interest in the genus QuerClls has centered on the nature of the group, its phylogenetic relationship to other genera of the Amentiferae, and its position in the Angiosperm classifIcation. For this reason, investigation largely has been concerned with the relationships of such characteristics as seemed important to
establish t.he separate lines of development of the several sub-groups of the Amentiferae. Detailed morphological studies of particular genera or species have been neglected. The genus Quercus has received little attention of an ontogenetic nature (2,8).
Conrad (3), working' wi til black oak ((2.uercus -uelll tirw), made it partial ontogcnetic study oj' the staminate and pistillate llowers, detailing the lalter through sporogenesis, gametogenesis, and early embryonic development. It is of intcrest that many of the observations made by Conrad are similar to those reported' here for Q. allJa except for the
• Allthori""d [01' puhllC'IliOll Januar)' II. 1051\. Contrihlltion No, I!II [rolll the Department oC llolilny and Plnnt Pathology. 'r Graduate Assistant, and A,.ocl"t~ I'ro(es.,or of :Botany respectively.
4$.7S3 I'lflllliflllfllmli(l.Ifillllrillllilll1
I~RI

Digest
Both staminate and pistillate tlowers of Q. alba are functionally monosporangiate and have their origin during vegetative growth of the year preceding the maturation of the fruit.
The development of the staminate inflorescence and the flower is initiated in late June. I~y October, it has developed to the overwintering condition. Sporogenous tissue is first noticeable in April of the year following- its iniliation and pollen is shed during the last week of May.
The over-wintering condition of the staminate Hower would seem to preclude any low temperature uamage, except as it might occur as a result of late frosts in the spring.
The pistillate inflorescence has its origin during the veg'etative growth period of late summer. However, because of the similarities to vegetative development, the primordium of the pistillate innorescence is hardly discernible until early October and may even be in doubt at that time.
Growth and differentiation are resumed early in l\Iay. At about the time of pollination, the pistillate flower is only partly developed. Subsequent to pollination, the development of the ovules, megasporogenesis and gametogenesis are cause for delay of syngamy (or at least a month following the dehiscence of the anthers.
Although not observed, fertilization must occur late in .June or ~arly July. Embryo development was seen as early as June 9 in one l11stance. This would indicate that considerable variation might well exist in the time schedule of development as set forth above.
The formation of an abscission layer at the base oC the ovary, between its tissues and those oE the involucre, was noted as early as the time of pollination. Even superficial obsel"vation of the shedding of immature oak fruits leaves little uoubt that early abscission layer formation is cause for a relatively small and rather unpredictable mast crop.
The developing embryo sac follows the Adoxa type, with considerable breakdown of the nucellar tissues and inner integument.

Floral Morphology of White Oak* HAROLD S. TURKEL, ALLEN L. REBUCK, ALVIN R. GROVE, .TR.t
THAT THERE IS CONSIDERABLE VARIATION in the produc-tivity of white oak acorns has long heen recognized. For any year
in which a substantial crop is produced, there are others in which there is little or no yield. This is, of course, not unique in white oak. But because of the relatively wide distribu don and the potential value of white oak mast, the interest of botanists, foresters, and wildlife specialists has been stimulated.
An attempt has been made here to relate the development of the inflorescences and flowers to specific time periods. This fundamental information is prerequisite to any additional exploration of the causes for failure of the crop.
The reasons which are generally oIfered for failure of fruit development appear to be largely a matter of conjecture. There is little, if any, basis in fact for them and certainly no proof.
It has often been assumed that temperature might be the c;msative agent of low productivity. Low temperatures early ill the spring' might well be an important consideration. High temperatures during some critical period of development later in the summer-perhaps at the time of pollination or between the time of pollination and syngamymight be significant.
By relating Horal development to relatively specific times, it is hoped that additional investigations at critical periods mig'ht oJfer an acceptable reason or reasons for poor fruit set.
Most of the previous morphological interest in the genus (~ueH'lIs has centered on the nature of the group, its phylogenetic relationship to other genera of the Amentiferae, and its position in the Ang'iosperm classification. For this reason, investigation largely has been concerned with the relationships of such characteristics as seemed important to establish the separate lines of development of the several sub-groups of the Amentiferae. Detailed morphological studies of particular genera or species have been neglected. The genus QlIeftllS has received little attention of an ontogenetic nature (2,8).
Conrad (3), working with hlack oak (Quercus "lleiutina), made a partial ontogenetic: study of the staminate and pistillate flowers, detailing the latter through sporogenesis, gametogenesis, and early embryonic development. It is of interest that many of the observations made by Conrad are similar to those reported' here [or (2: alba except for the
• Authorized [or publlcnrion J:lllU"rl' II, I !I!i!i. Contriilutioll No. I !ll fl"lllll lhe Department. of lI()~any and Plant l'athology.
t Graduate Assistants ~n(! Associ"'" l'rn[esHor of Botany respectively.

2 PENNSYLVANIA AGRICULTURAL EXPERIMENT STATION
fact that two years are necessary for the maturation of the fruit of (2,. velutina.
Langdon (7) investigated the various groups comprising the Amentiferae, including reel oak (Q. rubm). Her work was primarily concerned with Horal morphology.
Hjelmqvist (5), in a relatively recent study, investigated and interpreted the variolls structural modifications uf the flower and inflorescence of the Arnentiferae but did not report any observations concerned with sporogenesis or gametogenesis.
Reece (11) reported on the anatomical variations and tendencies in the Fagaceae. I t is of interest to note that an interpretation of the vascular anatomy of the pistillate flower of the Fagaceae supports the appendicular theory of carpel development.
Benson (1) studied the embryology of numerOus representative species of the Amentiferae, and Quercus was among those investigated. But, finding little distinctive difference between it and Castanea, she felt it unnecessary to report her findings. Conrad also reported observations concerning the embtyo of Q. velulina) which since has been placed under the Trifolium variation of the Onograd type (9).
PROCEDURE Collections of both pistillate and staminate material of white oak
were made from May 1949 until July 1952. Samples were taken from two trees on the campus of The Pennsylvania State University at weekly intervals, except at those times of development when three or fOllr collections a week were deemed necessary.
More than 100 staminate and 200 pistillate flowers were sectioned and they form the basis of this study. Additional series of duplicated material were discarded as unnecessary.
All material was killed and fixed in FOflnalin-Aceto-Alcohol, dehydrated in N-butyl alcohol, and sectioned in paraffin.
All buds were peeled or trimmed to remove as much of the UIl·
necessary material as possible. In some instances, it was necessary to treat the buds with 3 I)er cent KOH for several days before sectioning.
Numerous capitate hairs and stone cells made sectioning relatively difficult but most sections were cut at 10 to 15 mici'a. Sections were stained with safranin 0 and fast green.
STAMINATE STRUCTURES The staminate infiorescence is a catkin in which the individual
florets are functionally mOllosporangiate. Its initial development occurs in the young bud during' the latter part of June.
Early stages of development resemble those of the vegetative stem tip, fig. 1. Multiplaned divisions in the region of the hypodermis cause

BULLETIN 593-FLORAL MORPHOLOGY OJ' "VHlTE OAK .3
Fig. I.-A longitudinal section of the vegetative apex.
Fig. 2.-A longitudinal sectioll of the staminate primordiulll.
Fig. 3.-A longitudinal section of the young staminate prilllordiulIl.
Fig. -t.-Face view of the staminate Howe!" prilllordiul11.

4 FENNSYLVANIA AC;RICULTLJRAL EXPERIMENT STATION
bulging to take place, which makes it possible to distinguish this primordiulll from the vegetative one.
Development proceeds in this manner unlil the ce11lral axis primordium is approximately 27 micra in length. At this time, growth becomes apical, 'with the organization of a tunica-corpus stralification, fig. 2. Further evidence of its floral nature is obviolls as continued elongation takes place without the formation 01 additional vegetative clive} gences.
About the middle of July, localized, hypodermal, lllerislelllatic patches appear along the length of the young axis. These areas soon become raised and delimited as discrete islands surrounding the maturing tissues o[ the central axis, fig. 3. As a result of lllutual growth pressures, these individual Hower primordia assume the shape, in facc view, of a compacted diamond, pentagon, or hexagon, ftg. 4.
Coincident with the formation of the Hower primordiulll, a bract, associated with each staminate nO'weI', begins its development from adjacent tissues of the central axis. I nili'll growth is relatively rapid, and the bract is quite prominent at the tillle of stamen origination. However, gTovvth soon slackens and the rate of development is similar to other parts of the Bower.
By the latter part of July, recognition of the individual IllcmlJers o( the staminate flower is possible. Development is acropetal and the perianth or.igina tees before stamen developmen t. The sepals arisc as a common structure. However, as growth continues and clongation takes place, they are diverged at their tips. The segments of the pcrianth vary in number but six sepals were most commonly obscrved.
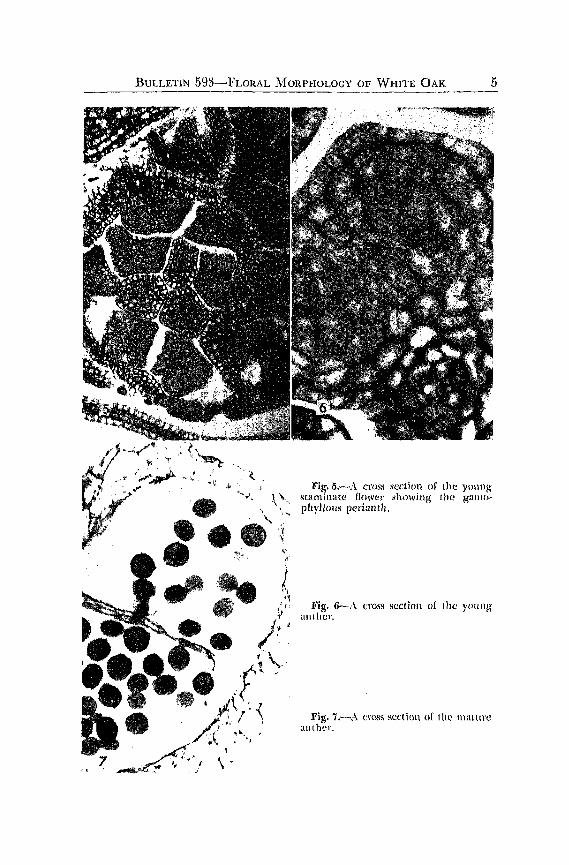
Stamen primordia originate in the typical eusporangiatc manner, from localized hypodermal groups of meristematic cells. The stamen initials are easily recognized by the end o[ July and the ftlaments are free. Continued development of the anther is slow. Althoug'h it is structurally mature by early fall (October), the resting' condition is histologically ,mnwture, fig. 5.
The anther, in the over-wintering condition, consists of a homogeneous parenchymatous mass, with the vascular clements identifi.ed as procambiul11. Anther insertion is terminal and, in cross section, there is slight lobing.
vVhat appeared to be vestigial carpels were observed in many slaminate flowers. Centrad to the anther position, several parenchyma tollS
masses developed which appeared circular in cross sectioll. Their UIl
differentiated cells and conspicuous epidermis made lhcm easily distinguishable [rom the developing anthers, fig. 5.
The resting anther resumes its development in April. Protoplasmic changes are noticeable in the retention of dyes. Cell divisiom at fi.rst
7
BULLETIN 59~-FLORAL MORPHOLOGY OF WHITE OAK 5
Fig.5.-A cross section of the young ~\., stll11linatc flower showing the gall}O-
, \, phylJolis periallth. \, ,.
'/ \
Fig. 6-..\ (.:I'OSS scelicJI] of I he yOllllg
anther.
Fig. 7.,--.'\ cr()ss S(!ctillll of thl' lIlallll'C anthe!'.

6 PENNSYLVANIA AGRICULTURAL EXPERIMENT STATION
serve only to increase the size of the fundamental tissue of the anther but, by the middle of April, the archesporium can be distinguished.
Periclinal divisions of the archesporium and lhe subsequent formation of primary parietal and sporogenous tissue are completed by about the third week of April, fig. 0.
Approximately one month following the recognition of parietal and sporogenous tissues, anther dehiscence and pollination take place.
The innermost layer of parietal cells becomes conspicuons as n result of increase in cell size, noticeable cytoplasmic ind lIsions, and the binucleate condition. I t is obviollsly functioning as a tapetum. Dissolution of this nutritive layer takes place at about the time of spore tetrad formation in the anther.
Microspore formation is prominent during- the early pan of May, and it is accompanied by the digestion of the parenchymatous ~epta separating the contiguous locules of the anther.
Final separation of the microspore tetrads and the development of the two-celled microgametophyte take place [rom about the middle of lvlay until anther dehiscence. Both the tube and generative nuclei are discernible. With the maturation of the pollen, thickenings on the walls of the endothedal cells become evident. However, specialization of the outer layer of parietal cells shows considerable variation. In many instances, em additional middle layer is evident in the wall of the anther.
Anther dehiscence takes place along the stomium, delimited by H.
double row of lip cells, fig. 7.
PISTILLATE STRUCTURES
The pistillate flowers occur singly, or as small clusters of from two to five, to form a racemose inHorescence. They develop in the axil of a foliage leaf or bract. The female flower is surrounded by an involucre. The ovary is inferior, consisting of three carpels. Of the ovules produced, usually only one ripens as the seed.
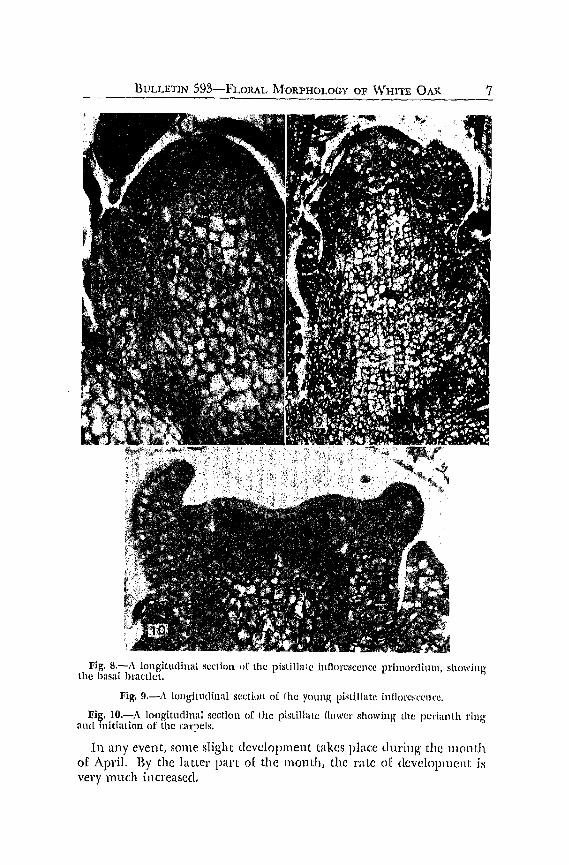
The earliest stages of the origin of the pistillate structure are llilHcult to recognize since their appearance coincides with that of the veg'etative bud. However, by early October, it is possible to distinguish the pistillate structure from the vegetative gTowth, largely through the failure of foliar primordia to develop and also because of the somewhat larger, naked appearance of the flowering primordiulll.
Very little, if any, development occurs from mid-Octoher ulltil February. In some cases, it was thought that slight elongation continued to take place during the winter. But variation in the size of the over-wintering pistillate structure might have accounted for this doubtful observation.
BVLLETIN 593~FLORAL MORPHOLOGY OF WHITE OAR 7
Fig. 8.-A longitudinal sectioll 1)[ the pistiJlal(~ illnClr~SCellC:C prilllot'(limn, showing' the basal bractlet.
Fig. 9.-:\ longitudinal scctillll of [he YOllng' pistillate inflorc!scencc.
Fig. 1O.-A ICHlgitudinal section of the pistilllile nowe!' showing lhe periallth ring and initiation of the Clll"peis.
In any event, some slight development takes place during' the month of April. By the latter pan of the lllonth, the rate of development i1i very much increased.

8 PENNSYLVANIA AGRICULTURAL EXPERIMENT STATION
By the time the mixed buds have begun to swell, during' the early part of May, a basal bractlet has developed 011 the side of the young axis, fig. 8. After the basal bractlet has made its appearance, continuous growth and elongation of the axis during the next week results in the appearance, in rapid succession, of a number of bl'actlets on the axis primordium, fig. 9.
A single Hower develops in the axil of tach of the lower bractlets. 1.1 mally, however, in addition to these fUllctional pistillate flowers, a number of bractleLs, with rudimentary flowers ill various stages of development, occur at the apex of the axis. Macroscopically, the stem tip, with its bractlets and rudimentary llowers, is seen as a slig'ht con· tinuation above the lateral functional Howers.
Immediately subsequent to this, and by the time the carpels have made their appearance, one or more bracteoles of the involucre already have appeared. Throughout the development of the flower and fruit, the cupule grows and increases in size, with more bracteo)cs being added at successively higher levels.
Although several hypotheses have been advanced concerning the morphological nature of the cupule in this family, there seems to he general agreement that the cupular involucre of the Hower and fruit of Quercus is a highly specialized structure. It is thought to he the product of fusion of the "secondary or tertiary branches or a llluch reduced dichasium" (2,5,/()).
The primordiulll of the pistillate flower arises as a hemispherical, or dome-shaped, structure. As it enlarges, about the middle of J'vIay, the apex of the primordium becomes flattened by more rapid growth of the peripheral area. Subsequently, there arise from this peripheral region usually six primordia. Their com1llon elongatioll gives rise to the gamophyllous perianth, fig'. Hl.
Following the origin of the periamh, the carpels arise frolll the flattened apex. Generally, they are three in number. Althoul4h two· carpellate ovaries were observed, the vascular anatomy indicat.es that the three carpellate condition is the common one, figs. 11,12. As a result of carpellary fusion and the adnation of the ovary anel the surrounding gamophyllous perianth, the flower is epigynous. Syncarpy does not involve the styles which are distinct. The tips and inner surfaces of these are stigmatic.
At lhe time of pollination, uSlUllly in the last week o[ I'l'1ay, the ovary rarely is mature and the ovules are just beginning to develop. This has been a common observation by those who have workeel with the Fagaceae and many other catkin-bearing plants.
At the time of pollination, rudimentary carpels have been ohserved in hazel, hornbeam, and alder. While pollination may take place in

BULLETIN 593-FLORAL :MORl'I-lOLOGY OF WI-UTE OAK 9
Fig. ll.-.\ ItJl1gitlidinal sectioll of the developing pistillate Hower.
Fig. I2.·-A cmss sl!t'lion of the pistillatc (tower al ttlc titne of poliillatioll.
Fig. IS.-.\. longitlldinal section o[ the pistillate lJowcr showing the ovules.
late i'day. syngamy docs not ordinarily occur until a month or more latcr.
In general, observations of (2. alba would indicate tbal pollination takes place between May 22 anc! 1\fay 28, with fertilization occulTing

10 PENNSYLVANIA AGRICULTURAL EXPERIMfNT STATION
during late June or early July. However, one well-developed elllhryo was noted from material collected June 9. This would necessitate allowing for some variation in the above statements.
Ovules develop at about the time of pollination. They become evident as protuberances from the axile placental folds of the inner carpellary walls. The six ovules are located at the same level, at the base of the locules. Two ovules occur in each of the three cells.
At first, growth is outward from the placental surface and slightly toward the base of the carpellary cavity. As the nucellns continues to elongate, it grows more rapidly on its lower and outer side, so that the ovule describes an arc of about 90 degTees. It finally assumes an erect, hemitropous position, fig. 13.
The two integuments appear simultaneollsly from the base of the nucellus in early June, about 10 clays after the nucellus is first rcco{!;nized. They elongate slowly and completely envelop the nucellus. They continue beyond the nucellus and both outer aml inner integ'uments grow into a long, much convoluted extension.
Although fertilization was not observed, ungerminated pollen grains were seen on the stigmatic surface of the style for several days subsequent to pollination. Some evidence of pollen tube growth was observed in the disrupted tissues of the inlegument. This suggested that the long attenuated integument might be serving, in some in" stances at least, as an integumentary obturator. The outer integument is relatively well supplied with vascular tissue. Although 110t observed, chalazogamy has been reported frequently by other workers in this and other genera of the Amentiferae.
At about the time of, or a little before, the appearance of the lWO integuments, changes in some of the hypodermal cells of the nuccllus indicate the location of the archesporium. Their cytoplasm lllay stain darker and the nuclei are more prominent with very noticeable nucleoli. The number of enlarged hypodermal cells varies from one to as many as six, arranged in a hypodermal plate.
Although division fig'ures were rarely seen, the subsequel1t anang-cment of the cells of the nucellus indicates that periclinal divisions of the archesporium give rise to primary parietal and primary sporo" genous cells. Indications are that the primary sporog'enolls cells need not act directly as megaspore mother cells but rather divide to produce a sporogenous mass, any cell of which might function as the primary megasporocyte, fig. 14.
In fact, i~ is impossible to be certain that each of these sp()ro,~'c110US cells. has ansen as the result of division of an archesporial cell. It is pOSSIble that some of the deep-seated cells have their origin in cells other than the hypodermal archesporium, fig. 15.

BULLETIN 5gB-FLORAL MORPHOLOGY OF WHITE OAK 11
Fig. 14.-A longitudinal view of the ovule showing divisioll of the archesporium to form primary ptn'idal lUld sporogcnous cells. Archesporial n~ll (a); primary parietal cell (pp); primary sporogenous cCII (ps).
Fig. lo.-The lluccllus with a JIlass o[ spol'ogenuus cells alld a two·layered parietal tissue.
Fig. lo.-Enlarging megaspore mothl~r cell.
Conrad describes a lIlulticellulat archcspOl'ial mass in (2. ·oelutinfl. It is not hypodermal, but extends far into the nucellar tissne. Its cells are, in reality, sporogenous, being potential megaspore mother cells.

12 PENNSYLVANIA AGR.ICULTURAL EXPERIMENT STATION ----About the middle of June, one or two of the sporogenous cells were
seen to enlarge, fig. 16. However, in none of the material examined did more than one of these megaspore mother cells complete sporogenes1s. The pattern followed is that of the Adoxa type.
Meiotic divisions of the functional megaspore mother cell occulTed during the last few days of June. As sporogenesis proceeded, distintegration of the other sporogenous tissue took place. Changes in the parietal and epidermal cells of the nucellus were soon obvious, indicating that they, too, were in the early stages of disintegration, fig. 17.
At first, the four megaspore nuclei are arranged in two groups of two each, with a pair at either end of the developing embryo sac. Most often, the two nuclei located at the micropylar end of the embryo sac, lie at right angles to the long axis of the nucellus, while the two nuclei toward the chalazal end are arranged parallel with it, fig". 18.
By early July, rather rapid disintegration of the nncellus at the micropylar region has taken place and the embryo sac is delimited hy the inner integument. The base of the nucellus remains and a caecum develops there, fig. 19.
This breakdown of most of the nucellus takes place before the final divisions of the megaspore nuclei. The nuclei of the mature gametophyte rearrange themselves so that the three antipodal nuclei are crowded into the tail of the caecum. The two polar nuclei occupy a more or less central position. The megagamete and two synergicls are arranged in typical fashion near the micropyle, fig. 20. Accompanying the final development of the gametophyte, disintegration of the inner cells of the inner integument takes place.
While mature megagametophytes were most often seen during the first week of .July, ovules ready for fertilization were noted in the last week of June. The material examined indicated that a variation of about two weeks is common in relation to any fixecl stage of development.
Indications are that a relatively low percentage of the pistillate flowers mature fruits. The formation of an abscission layer between the developing ovary and the involucre separates the young, undeveloped fruit from the tree. Whatever the physiological reason for the formation of this abscission layer, it forms early in many instances.
Some immature pistillate flowers, observed at the time of pollination late in May, already indicated the formation of the abscission layer. Developing fruits of various ages, examined as late as the end of July, showed many fruits continuing to form an abscission layer. These fruits dropped before maturity, fig. 21.
Apparently, wall sclerization leads to the death of certain of the cells in the area where the base of the developing ovary joins the

Fig. 17.-Virst divisioll of the megasporc mothcl' (cll. Fig. lB.-Second divisioll to fCll'lll four mcgaspore nuclei. Parit~tal tiHsue is disi11-
tcgratillg. Fig. 19.-Embryo sac Wilh [our megaspore nuclei. The llllCell\ls is disorganized at
the micropylar end. Caecum (c). Fig. 20.-Mature embryo sac. Antipodal llucleus (a); egg nucleus (c); polHl' 11\1-
deus (p); pl'Oc<lm bial stl'Hllcl (pc); synergic! (s).

1'1 PENNSYLVANIA AGRICULTURAL EXPERIMENT STATION
Fig. 21.-Early abscission layer forrriation at abollt the lime of pollination.
tissue of the involucre. No indication of cellular divisions was noted. But separation eventually takes place between the thick-walled cells and the relatively small, thin-walled cells adjacent to them.
LITERATURE CITED
(1) Benson, M. IB44. Contributions to the embryology o[ the Amclltiferae. 1'<\rt 1. Trans. Linn. Soc. Bot. II 3:409-422.
(2) Berridge, E. M. 1914. The structure of the flower of Fagaceae and its bearing on the affinities of the group. Ann. of Bot. 28:.~09-526.
(3) Conrad, A. H. 1900. A contribulion to the life history of QIt(~I'Ctis. !lol. Gaz. 29:408-418.
(1) Esau, Katherine. 1953. Plant Anatomy. John Wiley and Sons. Inc., New York. (5) Hjelmqvist, H. 1948. Studies 011 the floral morphology and phylogeny of the
Amentiferae. Botaniska Notiser 2:5-171. (6) James, W. O. and A. R. Clapham. 1935. The Biolog)) of Flmuet·s. Oxford llni-
versity Press, Oxford, Great Britain. .. (7) Langdon, L. M. 1939. Ontogenetic and analomicri:l studies of lhc !lowcr and
fruit of the Fagaceae and Juglandaceae. Bot. Gaz. 101:301-327. (8) Lendner, A. 1916. Sur la cupule des Fagacecs. Bul. Soc. Bot. GI'Ill~ve 8: 161-165. (9) Maheshwari, P. 1950. An Introduction to lhe Embryology of ArigiosjuJnlls.
McGraw-Hill Book Co., Inc., New York. . (10) Puri, V. 1951. The role of floral anatomy in the solution of morphological
problems. The Bot. Rev. 17:471-553. (11) Reece, P. L. 1938. The morphology of the flowers and inflorescences of lhe
Fagaceae. UnpUblished thesis, Cornell University.






























