Embed Size (px)
Citation preview

Plant Physiol. (1991) 95, 594-6030032-0889/91/95/0594/1 0/$01 .00/0
Received for publication July 2, 1990Accepted October 31, 1990
Flavonoid and Isoflavonoid Distribution in DevelopingSoybean Seedling Tissues and in Seed and Root Exudates'
Terrence L. GrahamDepartment of Plant Pathology, The Ohio State University, Columbus, Ohio 43210
ABSTRACT
The distribution of flavonoids, isoflavonoids, and their conju-gates in developing soybean (Glycine max L.) seedling organsand in root and seed exudates has been examined. Conjugatesof the isoflavones daidzein and genistein are major metabolitesin all embryonic organs within the dry seed and in seedling roots,hypocotyl, and cotyledon tissues at all times after germination.Primary leaf tissues undergo a programmed shift from isoflavo-noid to flavonoid metabolism 3 days after germination and be-come largely predominated by glycosides of the flavonols kamp-ferol, quercetin, and isorhamnetin by 5 days. Cotyledons containrelatively constant and very high levels of conjugates of bothdaidzein and genistein. Hypocotyl tissues contain a third uniden-tified compound, P19.3, also present in multiple conjugated forms.Conjugates of daidzein, genistein, and P19.3 are at their highestlevels in the hypocotyl hook and fall off progressively down thehypocotyl. These isoflavones also undergo a programmed anddramatic decrease between 2 and 4 days in the hypocotyl hook.All root sections are predominated by daidzein and its conjugates,particularly in the root tip, where they reach the highest levels inthe seedling. Light has a pronounced effect on the distribution ofthe isoflavones; in the dark, isoflavone levels in the root tips aregreatly reduced, while those in the cotyledons are higher. Finally,the conjugates of daidzein and genistein and several unidentifiedaromatic metabolites are selectively excreted into root and seedexudates. Analysis of seed exudates suggests that this is acontinuous, but saturable event.
Phenylpropanoid-derived metabolites are widely distrib-uted in the plant kingdom and have been proposed to playmany diverse roles (23). They have been implicated in theprotection of plants against both insects and pathogens andin the protection of plants against UV light damage. They aredeterminants offlower color and are involved in the attractionof pollinating insects. Because of the remarkable structuraldiversity shown by these metabolites and their derivatives andthe complexity of their distribution, it has long been proposedthat they may play additional, undefined roles in the plant,including a possible role in the regulation of growth anddevelopment (23, 28).
' Partial salary and research support were provided by state andfederal funds appropriated to the Ohio Agricultural and DevelopmentCenter (OARDC). This publication is OARDC manuscript 243-90.Partial research support was also provided by the U.S. Departmentof Agriculture under Cooperative Research Service Grant 89-37231-4493 to T. L. G.
Recent results from a number of laboratories have clearlyshown that the flavonoids and the less widely distributedisoflavonoids indeed possess additional biological activities.These activities include induction of nodulation genes invarious rhizobia species (21, 25), chemoattraction of Rhizo-bium leguminosarum biovar meliloti (10) and Bradyrhizo-bium japonicum (18), and possible regulation of auxintransport ( 17).
Research in our laboratory is focused on understanding theroles that flavonoid and isoflavonoid metabolites play in theresponses of soybeans (Glycine max L.) to biotic and abioticstress and the role(s) that they may play in signal exchangebetween soybeans and their microbial associates. Recently,we demonstrated the presence of previously unreported largeconstitutive pools of conjugates of the isoflavones daidzeinand genistein in soybean seedling tissues (14). These includethe simple 7-O-glucosyl- conjugates of daidzein and genistein(daidzin and genistin, respectively) and the more complexmalonylated conjugates (MGD2 and MGG, respectively). Evi-dence from our laboratory suggests that both the formationand hydrolysis of these isoflavone conjugates are subject toan intricate regulation program, and that they may play adirect role in the responses of soybean tissues to infection byPhytophthora megasperma f. sp. glycinea (PMG, 14) and toelicitor treatment (13).PMG infects all soybean seedling organs. However, race-
specific resistance, characterized by rapid and large accumu-lations of glyceollin, is expressed somewhat differently in eachindividual organ. Moreover, a series of important studies hasdemonstrated that the age and/or developmental state of thespecific organ infected by PMG and environmental condi-tions, particularly light, strongly influence race-specific accu-mulation of the glyceollins (4, 5, 20, 24, 26, 27). In addition,specific cell populations within a given organ may be involvedin glyceollin accumulation (13, 15, 29).
In research with the soybean symbiotic rhizobia, it has beenshown that daidzein is a potent inducer of the nodulationgenes in B. japonicum (19) and acts as a chemoattractant forB. japonicum (18). Like infection with PMG, infection ofroots by rhizobia species proceeds in distinct stages, each ofwhich is influenced by complex spatial and temporal inter-actions with specific host tissues at defined developmentalstages (for a recent overview, see ref. 21). Each phase of these
2Abbreviations: MGD, 6"-O-malonyl-7-O-f3-D-glucosyldaidzein;MGG, 6"-O-malonyl-7-O-3-o-glucosylgenistein; PMG, Phytophthoramegasperma Drechs. f. sp. glycinea (Hildeb.) Kuan & Erwin; ED50,the dose ofthe compound which is 50% effective in growth inhibition.
594 www.plantphysiol.orgon September 2, 2018 - Published by Downloaded from
Copyright © 1991 American Society of Plant Biologists. All rights reserved.

FLAVONOID DISTRIBUTION IN SOYBEAN
interactions may involve an exchange of specific signal mol-ecules between the host and symbiont (21, 25).To better understand the possible molecular interactions of
various soybean organs with their potential pathogens andsymbionts, we believed that it would be valuable to quantifythe distribution ofknown and unknown flavonoid and isofla-vonoid metabolites in various developing soybean seedlingtissues and in seed and root exudates. In addition to providingvaluable information for future experiments on the molecularmechanisms underlying the interactions of soybeans with itsmicrobial associates, such information could also be ofcriticalimportance in identifying additional determinants of hostresistance or compatibility and of host range. In an accom-panying paper (12), the development ofa rapid, sensitive, andhigh resolution HPLC metabolic profiling procedure is de-scribed which has allowed us for the first time to fingerprinta very wide range of soluble aromatic metabolites in crudeextracts of specific plant tissues or microbes. In this paper,this procedure is applied in an examination of the flavonoidsand isoflavonoids in discrete organs and tissues within thedeveloping soybean seedling and in seed and root exudates.The results demonstrate not only that the distribution patternis organ specific, but that the relative levels of specific flavon-oids and isoflavonoids are characteristic of each organ. More-over, within a tissue or organ, and within seed and rootexudates, the distribution of the flavonoids and isoflavonoidsvaries both with developmental stages, and in response tolight, in a manner which may have important implications inthe interactions of soybeans with its microbial associates.
MATERIALS AND METHODS
Growth of Soybean Plants
Williams, Williams 79, and Williams 82 soybean (Glycinemax L.) seedlings were grown from field-increased seed ob-tained from AF Schmitthenner (Department of Plant Pathol-ogy, The Ohio State University, Columbus, OH). Seedlingswere grown at 26C with 500 AuE/m2/s of light on a 14-hphotoperiod as reported previously (14). Etiolated soybeanseedlings were grown by germination and growth of surface-sterilized seeds (1% hypochlorite, 5 min) in total darknessbetween two 45.5 x 30.5 cm sheets of Anchor germinationpaper (Anchor Paper, Packaging Converters, Hudson, WI).After rolling the sheets into a cylinder, the cylinders were keptupright in sealed plastic bags until harvested.
Collection of Soybean Seed and Root Exudates
Surface-sterilized soybean seed were germinated and grownin specially designed chambers, described below, which al-lowed the seed to imbibe slowly and the roots to developsuspended in air at 100% RH. Black polystyrene plant-grow-ing trays, with wells measuring 1.7 cm diameter x 2.5 cmdeep (A. H. Hummert Seed Co., St. Louis, MO), were cut toform a grid containing five wells square. They were sterilizedby soaking in 70% ethanol prior to use. Individual soybeanseeds were loosely wrapped in moistened sterile germinationpaper (5 x 13 cm) and placed in the individual wells. Theseeds were oriented such that their radicle would extend outofthe germination paper and through the drainage hole. These
modified trays were then suspended in a sterilized closedpolypropylene freezer container (10 cm square x 12 cm deep)completely lined with absorbent tissues soaked in sterile water.The chambers were incubated at 26°C in complete darkness.Seeds germinated in this manner underwent very gradualhydration, with no apparent damage to the seed coat or toseed or seedling tissues. The resultant seedlings formed normallooking, succulent, white, and unblemished roots. Roots weregrown in this manner to avoid physical damage to surfacecells and nonspecific leakage of metabolites. Although growthof seedlings in aerated nutrient solutions was considered,preliminary results showed that changes in root metabolitesoccur under flooding conditions regardless of the care takenin aeration.
Seed exudate was collected by thoroughly washing thegermination paper for 10 replicate seeds with 10 mL HPLC-grade water. The resultant exudate was then passed slowlyunder vacuum through a C18 PrepSep column (Fisher Sci-entific, Springfield, NJ) previously washed with 100% meth-anol followed by HPLC grade water. Metabolites were theneluted in one step by washing slowly with 10 mL of HPLCgrade acetonitrile. The eluent was then taken to dryness,dissolved in 200 uL 80% ethanol, and used directly for HPLCanalysis.Root exudate was collected directly from roots suspended
in the chambers by placing short (3-5 mm) sections of pre-washed, sterile, water-soaked 2 mm diameter cotton string(wick) at the desired point along the root length. After 30min, the wicks from 20 replicate plants were harvested, andthe exudate was collected by centrifugation in a modifiedmicrofuge tube fit with plastic inserts which allowed collectionof the exudate in a lower chamber and retention of the wicksin an upper chamber. The volume of this exudate was meas-ured and the exudate was subjected directly to HPLC analysis.
Extraction and HPLC Analysis of Intact Soybean Tissues
Unblemished, healthy soybean tissues were harvested, ex-tracted, and analyzed as reported previously (12, 14). Allextractions were made in 80% ethanol (400 gL/0. 1 g tissue).HPLC separation and quantitation of the various metaboliteswere achieved as described previously (12, 14). Injections of20 ,uL were made on a 4.6 mm i.d. x 250 mm Hibar EcCartridge containing Merck Lichrosorb RP-18 10 ,m C 18reverse phase packing (Alltech Associates, Deerfield, IL). Aguard column containing the same packing was used toprotect the analytical column. Chromatography was achievedat 25°C with an Applied Biosystems Spectroflow model 430low pressure gradient former and an Applied BiosystemsSpectroflow 400 high pressure pump to form a linear gradientof 0 to 55% acetonitrile in pH 3 water for 25 min, followedby a step increase to 100% acetonitrile, which was held for 2min before a step return to pH 3 water. A flow rate of 1.5mL/min was used. HPLC grade water was adjusted to pH 3with phosphoric acid. The overall separation time, includinga 3-min water wash, was 30 min.
Spectrometric detection was at 236 nm. Calibration wasachieved with authentic standards and an Applied BiosystemsSpectroflow 783 Absorbance Detector coupled to a Spectra-Physics SP4290 Integrator. Standards of glyceollin and the
595
www.plantphysiol.orgon September 2, 2018 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.
Plant Physiol. Vol. 95, 1991
conjugates of daidzein and genistein were prepared based ontheir extinction coefficients as described previously (14). Allother standards were prepared by weight.
Criteria for the identity and purity ofindividual metabolitesincluded coinjection with authentic standards, ultravioletspectral analysis of individually collected peaks at severalpoints in the elution of the peak, and, in the case of all majormetabolites, fast atom bombardment mass spectral analysis.
RESULTS
Organ Specific Distribution of Flavonoids andIsoflavonoids
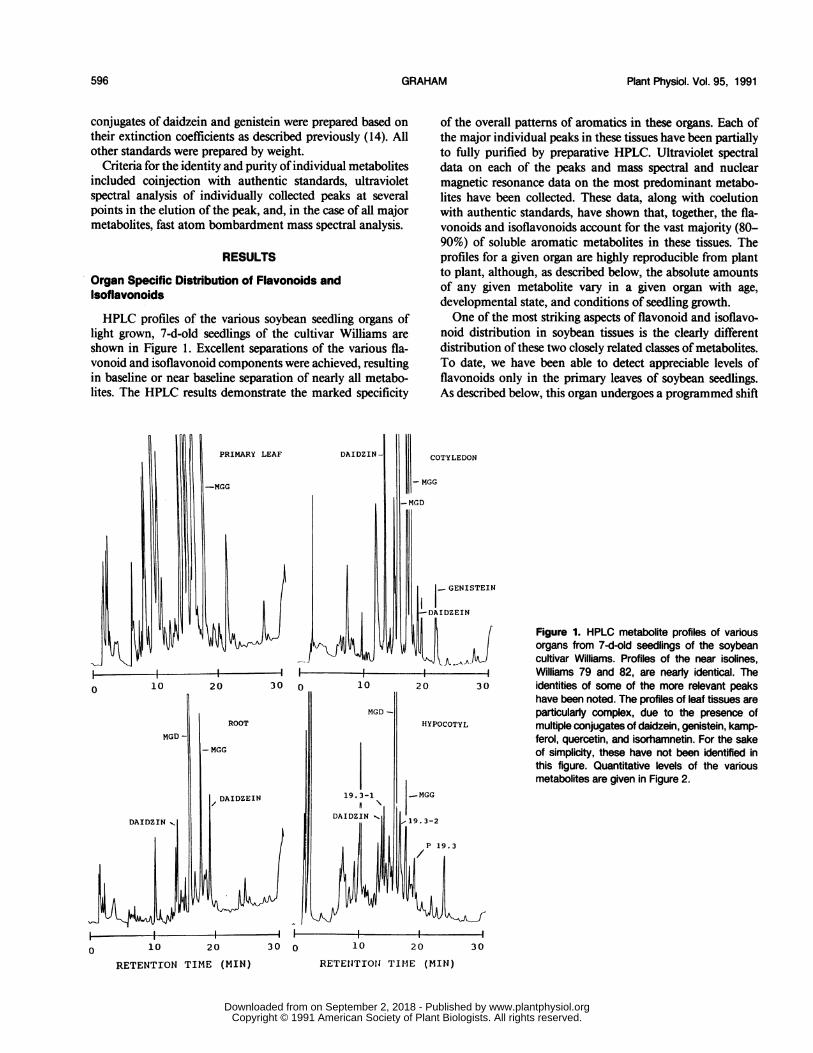
HPLC profiles of the various soybean seedling organs oflight grown, 7-d-old seedlings of the cultivar Williams areshown in Figure 1. Excellent separations of the various fla-vonoid and isoflavonoid components were achieved, resultingin baseline or near baseline separation of nearly all metabo-lites. The HPLC results demonstrate the marked specificity
0 10 20 30
MGD-
DAIDZIN
ROOT
MGG
DAIDZIN -
lk'---f
of the overall patterns of aromatics in these organs. Each ofthe major individual peaks in these tissues have been partiallyto fully purified by preparative HPLC. Ultraviolet spectraldata on each of the peaks and mass spectral and nuclearmagnetic resonance data on the most predominant metabo-lites have been collected. These data, along with coelutionwith authentic standards, have shown that, together, the fla-vonoids and isoflavonoids account for the vast majority (80-90%) of soluble aromatic metabolites in these tissues. Theprofiles for a given organ are highly reproducible from plantto plant, although, as described below, the absolute amountsof any given metabolite vary in a given organ with age,developmental state, and conditions of seedling growth.One of the most striking aspects of flavonoid and isoflavo-
noid distribution in soybean tissues is the clearly differentdistribution ofthese two closely related classes of metabolites.To date, we have been able to detect appreciable levels offlavonoids only in the primary leaves of soybean seedlings.As described below, this organ undergoes a programmed shift
- MGG
)TYLEDON
-MGD
STEINI_- GENIS
-DAIDZEIN
!4i -1Figure 1. HPLC metabolite profiles of variousorgans from 7-d-old seedlings of the soybeancultivar Williams. Profiles of the near isolines,Williams 79 and 82, are nearly identical. The
30 identities of some of the more relevant peakshave been noted. The profiles of leaf tissues areparticularly complex, due to the presence ofmultiple conjugates of daidzein, genistein, kamp-ferol, quercetin, and isorhamnetin. For the sakeof simplicity, these have not been identified inthis figure. Quantitative levels of the variousmetabolites are given in Figure 2.
30 0 10 20 30
RETENlTIONJ TIME (MIN)0 10 20
RETENTION TIME (MIN)
iL-i
-i i r
;I
596 GRAHAM
I
www.plantphysiol.orgon September 2, 2018 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.
FLAVONOID DISTRIBUTION IN SOYBEAN
from what is predominantly isoflavonoid metabolism in theplumule (d 0-3) to predominantly flavonoid metabolism inthe expanding primary leaf(after d 3). The profiles of all otherseedling organs are dominated, at all ages, by theisoflavonoids.
In addition to the multiple conjugates of the isoflavonesdaidzein and genistein previously identified in cotyledon tis-sues (14), we have identified several additional aromatic me-tabolites which have more specific distributions. Some ofthesewill be discussed further below, since they were more apparentin seed or root exudates. In whole tissue extracts, a metaboliteof particular interest has been identified in hypocotyl tissues(Fig. 1, P19.3). This aglycone has a retention time slightlylonger than daidzein, and, like daidzein, exists as multipleconjugates, which are denoted in Figure 1 as P19.3-1 andP19.3-2. We do not yet know the identity of P19.3, althoughit does not correspond to any of the known flavonoids orisoflavonoids for which we have standards.Although daidzein, genistein, and their conjugates are
found in all of the tissues, there are a few interesting andspecific aspects to their distribution. Free genistein and itsglucosyl conjugate, genistin, are at very low (often nondetect-able) levels in all seedling organs.
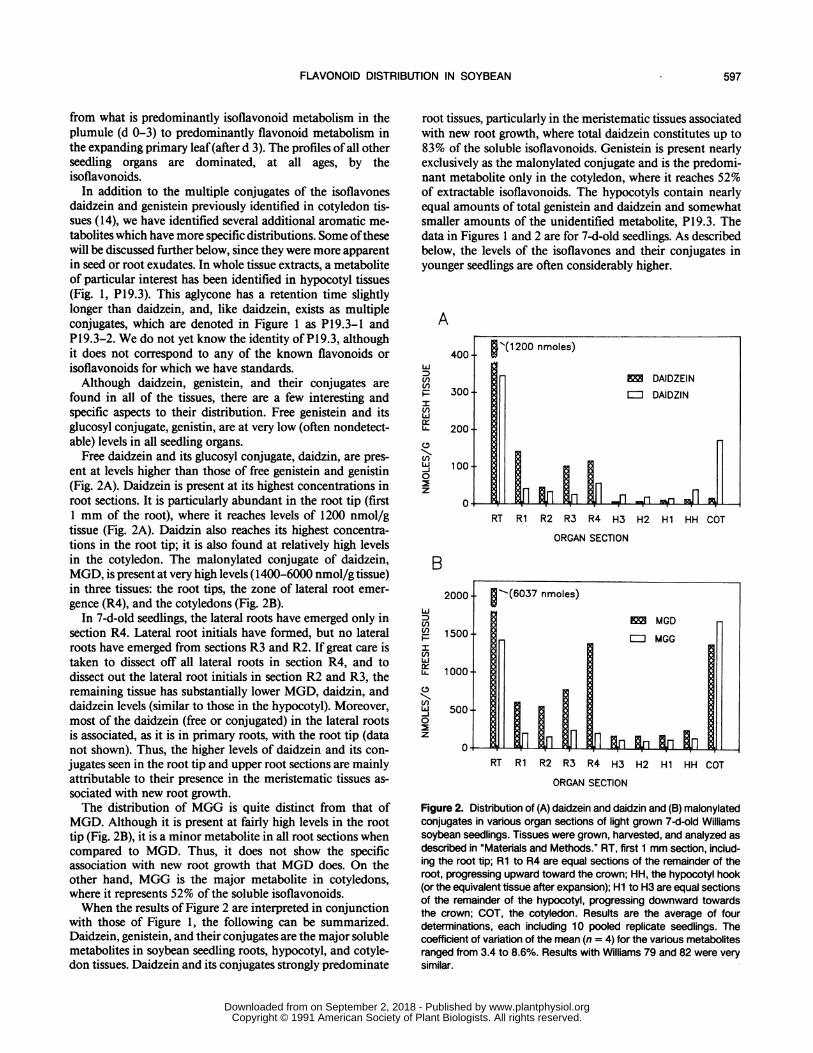
Free daidzein and its glucosyl conjugate, daidzin, are pres-ent at levels higher than those of free genistein and genistin(Fig. 2A). Daidzein is present at its highest concentrations inroot sections. It is particularly abundant in the root tip (first1 mm of the root), where it reaches levels of 1200 nmol/gtissue (Fig. 2A). Daidzin also reaches its highest concentra-tions in the root tip; it is also found at relatively high levelsin the cotyledon. The malonylated conjugate of daidzein,MGD, is present at very high levels (1400-6000 nmol/g tissue)in three tissues: the root tips, the zone of lateral root emer-gence (R4), and the cotyledons (Fig. 2B).
In 7-d-old seedlings, the lateral roots have emerged only insection R4. Lateral root initials have formed, but no lateralroots have emerged from sections R3 and R2. If great care istaken to dissect off all lateral roots in section R4, and todissect out the lateral root initials in section R2 and R3, theremaining tissue has substantially lower MGD, daidzin, anddaidzein levels (similar to those in the hypocotyl). Moreover,most of the daidzein (free or conjugated) in the lateral rootsis associated, as it is in primary roots, with the root tip (datanot shown). Thus, the higher levels of daidzein and its con-jugates seen in the root tip and upper root sections are mainlyattributable to their presence in the meristematic tissues as-sociated with new root growth.The distribution of MGG is quite distinct from that of
MGD. Although it is present at fairly high levels in the roottip (Fig. 2B), it is a minor metabolite in all root sections whencompared to MGD. Thus, it does not show the specificassociation with new root growth that MGD does. On theother hand, MGG is the major metabolite in cotyledons,where it represents 52% of the soluble isoflavonoids.When the results of Figure 2 are interpreted in conjunction
with those of Figure 1, the following can be summarized.Daidzein, genistein, and their conjugates are the major solublemetabolites in soybean seedling roots, hypocotyl, and cotyle-don tissues. Daidzein and its conjugates strongly predominate
root tissues, particularly in the meristematic tissues associatedwith new root growth, where total daidzein constitutes up to83% of the soluble isoflavonoids. Genistein is present nearlyexclusively as the malonylated conjugate and is the predomi-nant metabolite only in the cotyledon, where it reaches 52%of extractable isoflavonoids. The hypocotyls contain nearlyequal amounts of total genistein and daidzein and somewhatsmaller amounts of the unidentified metabolite, PI 9.3. Thedata in Figures 1 and 2 are for 7-d-old seedlings. As describedbelow, the levels of the isoflavones and their conjugates inyounger seedlings are often considerably higher.
A
400 R'(1 200 nmoles)
E) O DAIDZEIN() 300- EDDAIDZINLi 200
c,
0
00m f
RT Rl R2 R3 R4 H3 H2 Hi HH COT
ORGAN SECTION
B
wLcnU)Uf)wCLL
U)-J0
z
RT Rl R2 R3 R4 H3 H2 Hi HH COT
ORGAN SECTION
Figure 2. Distribution of (A) daidzein and daidzin and (B) malonylatedconjugates in various organ sections of light grown 7-dold Williamssoybean seedlings. Tissues were grown, harvested, and analyzed asdescribed in "Materials and Methods." RT, first 1 mm section, includ-ing the root tip; Ri to R4 are equal sections of the remainder of theroot, progressing upward toward the crown; HH, the hypocotyl hook(or the equivalent tissue after expansion); Hi to H3 are equal sectionsof the remainder of the hypocotyl, progressing downward towardsthe crown; COT, the cotyledon. Results are the average of fourdeterminations, each including 10 pooled replicate seedlings. Thecoefficient of variation of the mean (n = 4) for the various metabolitesranged from 3.4 to 8.6%. Results with Williams 79 and 82 were verysimilar.
597
www.plantphysiol.orgon September 2, 2018 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.
Plant Physiol. Vol. 95, 1991
A
400-
(nP 300
Zi:enJLL 200
EnSj 1000
z
0
B
2000
1500
1000'
500
RT Ri R2 R3 R4 H3 H2 Hi HH COT
ORGAN SECTION
RT Ri R2 R3 R4 H3 H2 Hi HH COT
ORGAN SECTION
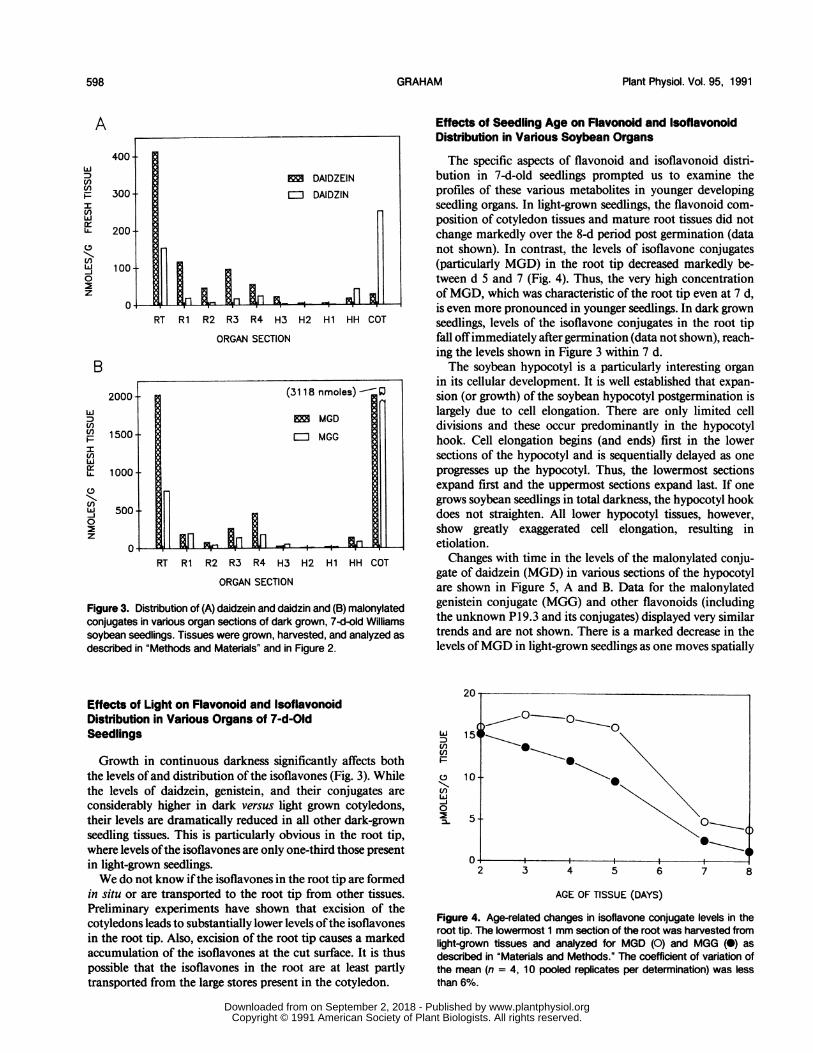
Figure 3. Distribution of (A) daidzein and daidzin and (B) malonylatedconjugates in various organ sections of dark grown, 7-d-old Williamssoybean seedlings. Tissues were grown, harvested, and analyzed asdescribed in "Methods and Materials" and in Figure 2.
Effects of Light on Flavonoid and IsoflavonoidDistribution in Various Organs of 7-d-OldSeedlings
Growth in continuous darkness significantly affects boththe levels ofand distribution of the isoflavones (Fig. 3). Whilethe levels of daidzein, genistein, and their conjugates are
considerably higher in dark versus light grown cotyledons,their levels are dramatically reduced in all other dark-grownseedling tissues. This is particularly obvious in the root tip,where levels ofthe isoflavones are only one-third those presentin light-grown seedlings.We do not know ifthe isoflavones in the root tip are formed
in situ or are transported to the root tip from other tissues.Preliminary experiments have shown that excision of thecotyledons leads to substantially lower levels ofthe isoflavonesin the root tip. Also, excision of the root tip causes a markedaccumulation of the isoflavones at the cut surface. It is thuspossible that the isoflavones in the root are at least partlytransported from the large stores present in the cotyledon.
Effects of Seedling Age on Flavonoid and IsoflavonoidDistribution in Various Soybean Organs
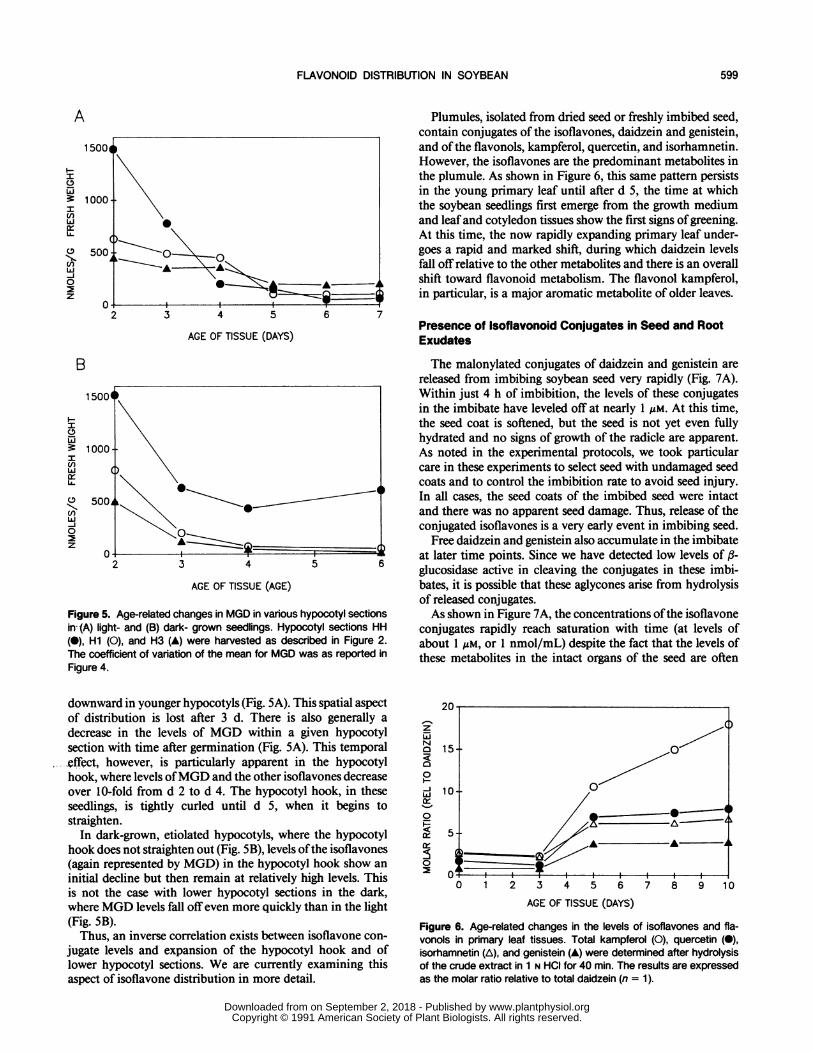
The specific aspects of flavonoid and isoflavonoid distri-bution in 7-d-old seedlings prompted us to examine theprofiles of these various metabolites in younger developingseedling organs. In light-grown seedlings, the flavonoid com-position of cotyledon tissues and mature root tissues did notchange markedly over the 8-d period post germination (datanot shown). In contrast, the levels of isoflavone conjugates(particularly MGD) in the root tip decreased markedly be-tween d 5 and 7 (Fig. 4). Thus, the very high concentrationofMGD, which was characteristic of the root tip even at 7 d,is even more pronounced in younger seedlings. In dark grownseedlings, levels of the isoflavone conjugates in the root tipfall offimmediately after germination (data not shown), reach-ing the levels shown in Figure 3 within 7 d.The soybean hypocotyl is a particularly interesting organ
in its cellular development. It is well established that expan-sion (or growth) of the soybean hypocotyl postgermination islargely due to cell elongation. There are only limited celldivisions and these occur predominantly in the hypocotylhook. Cell elongation begins (and ends) first in the lowersections of the hypocotyl and is sequentially delayed as oneprogresses up the hypocotyl. Thus, the lowermost sectionsexpand first and the uppermost sections expand last. If onegrows soybean seedlings in total darkness, the hypocotyl hookdoes not straighten. All lower hypocotyl tissues, however,show greatly exaggerated cell elongation, resulting inetiolation.Changes with time in the levels of the malonylated conju-
gate of daidzein (MGD) in various sections of the hypocotylare shown in Figure 5, A and B. Data for the malonylatedgenistein conjugate (MGG) and other flavonoids (includingthe unknown P19.3 and its conjugates) displayed very similartrends and are not shown. There is a marked decrease in thelevels ofMGD in light-grown seedlings as one moves spatially
20
ui15i
(110
w-J0
0
, , , ,
2 3 4 5 6 7 8
AGE OF TISSUE (DAYS)
Figure 4. Age-related changes in isoflavone conjugate levels in theroot tip. The lowermost 1 mm section of the root was harvested fromlight-grown tissues and analyzed for MGD (0) and MGG (0) asdescribed in "Materials and Methods." The coefficient of variation ofthe mean (n = 4, 10 pooled replicates per determination) was lessthan 6%.
w
LU
(f)t
wU-
0
z
GRAHAM598
www.plantphysiol.orgon September 2, 2018 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.
FLAVONOID DISTRIBUTION IN SOYBEAN
A
uij
w
U)
-J0
z
AGE OF TISSUE (DAYS)
B
1500
H
3 1000-U) i \w
U)C- 500f >;0~~~~~~0 ~~~0
2 3 4 5 6
AGE OF TISSUE (AGE)
Figure 5. Age-related changes in MGD in various hypocotyl sectionsin--(A) light- and (B) dark- grown seedlings. Hypocotyl sections HH(0), Hi (0), and H3 (A) were harvested as described in Figure 2.The coefficient of variation of the mean for MGD was as reported inFigure 4.
downward in younger hypocotyls (Fig. 5A). This spatial aspectof distribution is lost after 3 d. There is also generally adecrease in the levels of MGD within a given hypocotylsection with time after germination (Fig. 5A). This temporal.effect, however, is particularly apparent in the hypocotylhook, where levels ofMGD and the other isoflavones decreaseover 10-fold from d 2 to d 4. The hypocotyl hook, in theseseedlings, is tightly curled until d 5, when it begins tostraighten.
In dark-grown, etiolated hypocotyls, where the hypocotylhook does not straighten out (Fig. SB), levels ofthe isoflavones(again represented by MGD) in the hypocotyl hook show aninitial decline but then remain at relatively high levels. Thisis not the case with lower hypocotyl sections in the dark,where MGD levels fall offeven more quickly than in the light(Fig. SB).
Thus, an inverse correlation exists between isoflavone con-jugate levels and expansion of the hypocotyl hook and oflower hypocotyl sections. We are currently examining thisaspect of isoflavone distribution in more detail.
Plumules, isolated from dried seed or freshly imbibed seed,contain conjugates of the isoflavones, daidzein and genistein,and of the flavonols, kampferol, quercetin, and isorhamnetin.However, the isoflavones are the predominant metabolites inthe plumule. As shown in Figure 6, this same pattern persistsin the young primary leaf until after d 5, the time at whichthe soybean seedlings first emerge from the growth mediumand leafand cotyledon tissues show the first signs of greening.At this time, the now rapidly expanding primary leaf under-goes a rapid and marked shift, during which daidzein levelsfall off relative to the other metabolites and there is an overallshift toward flavonoid metabolism. The flavonol kampferol,in particular, is a major aromatic metabolite of older leaves.
Presence of Isoflavonoid Conjugates in Seed and RootExudates
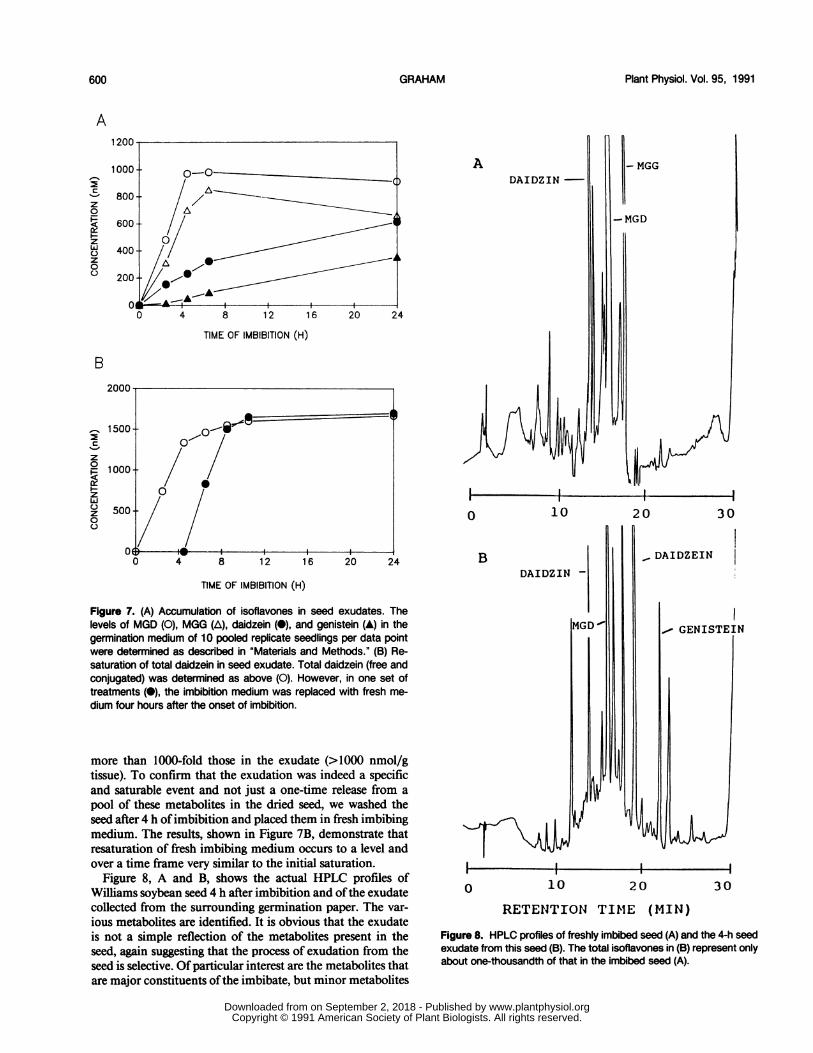
The malonylated conjugates of daidzein and genistein arereleased from imbibing soybean seed very rapidly (Fig. 7A).Within just 4 h of imbibition, the levels of these conjugatesin the imbibate have leveled off at nearly 1 ,M. At this time,the seed coat is softened, but the seed is not yet even fullyhydrated and no signs of growth of the radicle are apparent.As noted in the experimental protocols, we took particularcare in these experiments to select seed with undamaged seedcoats and to control the imbibition rate to avoid seed injury.In all cases, the seed coats of the imbibed seed were intactand there was no apparent seed damage. Thus, release of theconjugated isoflavones is a very early event in imbibing seed.
Free daidzein and genistein also accumulate in the imbibateat later time points. Since we have detected low levels of ,3-glucosidase active in cleaving the conjugates in these imbi-bates, it is possible that these aglycones arise from hydrolysisof released conjugates.As shown in Figure 7A, the concentrations ofthe isoflavone
conjugates rapidly reach saturation with time (at levels ofabout 1 ,uM, or 1 nmol/mL) despite the fact that the levels ofthese metabolites in the intact organs of the seed are often
20 -
zN015--00
0
H ~~~~~ A0 1 2 3 4 5 6 7 8 9 10
AGE OF TISSUE (DAYS)
Figure 6. Age-related changes in the levels of isoflavones and fla-vonols in primary leaf tissues. Total kampferol (0), quercetin (0),isorhamnetin (A), and genistein (A) were determined after hydrolysisof the crude extract in 1 N HCI for 40 min. The results are expressedas the molar ratio relative to total daidzein (n = 1).
599
www.plantphysiol.orgon September 2, 2018 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.
Plant Physiol. Vol. 95, 1991
A
z
z
01-
z00
TIME OF IMBIBITION (H)
B
c%-
z0
z
z0
TIME OF IMBIBITION (H)
Figure 7. (A) Accumulation of isoflavones in seed exudates. Thelevels of MGD (0), MGG (A), daidzein (0), and genistein (A) in thegermination medium of 10 pooled replicate seedlings per data pointwere determined as described in "Materials and Methods." (B) Re-saturation of total daidzein in seed exudate. Total daidzein (free andconjugated) was determined as above (0). However, in one set oftreatments (0), the imbibition medium was replaced with fresh me-
dium four hours after the onset of imbibition.
more than 1000-fold those in the exudate (>1000 nmol/gtissue). To confirm that the exudation was indeed a specificand saturable event and not just a one-time release from a
pool of these metabolites in the dried seed, we washed theseed after 4 h ofimbibition and placed them in fresh imbibingmedium. The results, shown in Figure 7B, demonstrate thatresaturation of fresh imbibing medium occurs to a level andover a time frame very similar to the initial saturation.
Figure 8, A and B, shows the actual HPLC profiles ofWilliams soybean seed 4 h after imbibition and ofthe exudatecollected from the surrounding germination paper. The var-
ious metabolites are identified. It is obvious that the exudateis not a simple reflection of the metabolites present in theseed, again suggesting that the process of exudation from theseed is selective. Of particular interest are the metabolites thatare major constituents ofthe imbibate, but minor metabolites
0
ADAIDZIN-
I
0 10
BDAIDZIN
10
- MGG
-MGD
IrS
2
i I
'0 30
_ DAIDZEIN
EN
4I3020
RETENTION TIM1E (MIN)
Figure 8. HPLC profiles of freshly imbibed seed (A) and the 4-h seedexudate from this seed (B). The total isoflavones in (B) represent onlyabout one-thousandth of that in the imbibed seed (A).
I i
GRAHAM600
I I
I I I
www.plantphysiol.orgon September 2, 2018 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.
FLAVONOID DISTRIBUTION IN SOYBEAN
in the seed per se. We are currently isolating these compoundsfor structural and biological studies.To determine if the conjugates also were secreted by the
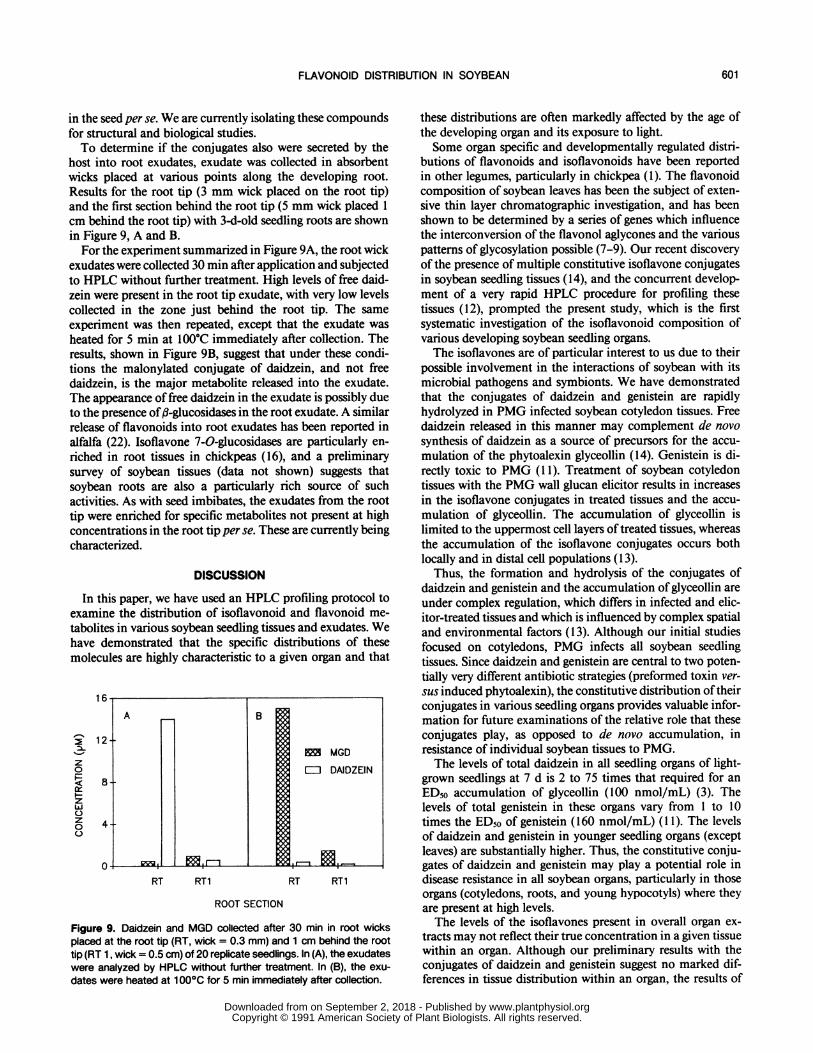
host into root exudates, exudate was collected in absorbentwicks placed at various points along the developing root.Results for the root tip (3 mm wick placed on the root tip)and the first section behind the root tip (5 mm wick placed 1cm behind the root tip) with 3-d-old seedling roots are shownin Figure 9, A and B.For the experiment summarized in Figure 9A, the root wick
exudates were collected 30 min after application and subjectedto HPLC without further treatment. High levels of free daid-zein were present in the root tip exudate, with very low levelscollected in the zone just behind the root tip. The sameexperiment was then repeated, except that the exudate washeated for 5 min at 100°C immediately after collection. Theresults, shown in Figure 9B, suggest that under these condi-tions the malonylated conjugate of daidzein, and not freedaidzein, is the major metabolite released into the exudate.The appearance of free daidzein in the exudate is possibly dueto the presence of,3-glucosidases in the root exudate. A similarrelease of flavonoids into root exudates has been reported inalfalfa (22). Isoflavone 7-0-glucosidases are particularly en-riched in root tissues in chickpeas (16), and a preliminarysurvey of soybean tissues (data not shown) suggests thatsoybean roots are also a particularly rich source of suchactivities. As with seed imbibates, the exudates from the roottip were enriched for specific metabolites not present at highconcentrations in the root tip per se. These are currently beingcharacterized.
DISCUSSION
In this paper, we have used an HPLC profiling protocol toexamine the distribution of isoflavonoid and flavonoid me-tabolites in various soybean seedling tissues and exudates. Wehave demonstrated that the specific distributions of thesemolecules are highly characteristic to a given organ and that
z
oa-
z
cL
z0
0
l6bA B
12IMMGD=J DAIDZEIN
8-
4-
RT RT1 RT RT1
ROOT SECTION
Figure 9. Daidzein and MGD collected after 30 min in root wicksplaced at the root tip (RT, wick = 0.3 mm) and 1 cm behind the roottip (RT 1, wick = 0.5 cm) of 20 replicate seedlings. In (A), the exudateswere analyzed by HPLC without further treatment. In (B), the exu-
dates were heated at 1 000C for 5 min immediately after collection.
these distributions are often markedly affected by the age ofthe developing organ and its exposure to light.Some organ specific and developmentally regulated distri-
butions of flavonoids and isoflavonoids have been reportedin other legumes, particularly in chickpea (1). The flavonoidcomposition of soybean leaves has been the subject of exten-sive thin layer chromatographic investigation, and has beenshown to be determined by a series of genes which influencethe interconversion of the flavonol aglycones and the variouspatterns of glycosylation possible (7-9). Our recent discoveryof the presence of multiple constitutive isoflavone conjugatesin soybean seedling tissues (14), and the concurrent develop-ment of a very rapid HPLC procedure for profiling thesetissues (12), prompted the present study, which is the firstsystematic investigation of the isoflavonoid composition ofvarious developing soybean seedling organs.The isoflavones are of particular interest to us due to their
possible involvement in the interactions of soybean with itsmicrobial pathogens and symbionts. We have demonstratedthat the conjugates of daidzein and genistein are rapidlyhydrolyzed in PMG infected soybean cotyledon tissues. Freedaidzein released in this manner may complement de novosynthesis of daidzein as a source of precursors for the accu-mulation of the phytoalexin glyceollin (14). Genistein is di-rectly toxic to PMG (1 1). Treatment of soybean cotyledontissues with the PMG wall glucan elicitor results in increasesin the isoflavone conjugates in treated tissues and the accu-mulation of glyceollin. The accumulation of glyceollin islimited to the uppermost cell layers of treated tissues, whereasthe accumulation of the isoflavone conjugates occurs bothlocally and in distal cell populations (13).
Thus, the formation and hydrolysis of the conjugates ofdaidzein and genistein and the accumulation of glyceollin areunder complex regulation, which differs in infected and elic-itor-treated tissues and which is influenced by complex spatialand environmental factors (13). Although our initial studiesfocused on cotyledons, PMG infects all soybean seedlingtissues. Since daidzein and genistein are central to two poten-tially very different antibiotic strategies (preformed toxin ver-sus induced phytoalexin), the constitutive distribution oftheirconjugates in various seedling organs provides valuable infor-mation for future examinations of the relative role that theseconjugates play, as opposed to de novo accumulation, inresistance of individual soybean tissues to PMG.The levels of total daidzein in all seedling organs of light-
grown seedlings at 7 d is 2 to 75 times that required for anED50 accumulation of glyceollin (100 nmol/mL) (3). Thelevels of total genistein in these organs vary from 1 to 10times the ED50 of genistein (160 nmol/mL) (11). The levelsof daidzein and genistein in younger seedling organs (exceptleaves) are substantially higher. Thus, the constitutive conju-gates of daidzein and genistein may play a potential role indisease resistance in all soybean organs, particularly in thoseorgans (cotyledons, roots, and young hypocotyls) where theyare present at high levels.The levels of the isoflavones present in overall organ ex-
tracts may not reflect their true concentration in a given tissuewithin an organ. Although our preliminary results with theconjugates of daidzein and genistein suggest no marked dif-ferences in tissue distribution within an organ, the results of
601
www.plantphysiol.orgon September 2, 2018 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 95, 1991
Barz and Hoesel (1) suggest that the analogous conjugates ofthe isoflavones formononetin and biochanin A may be local-ized in specific tissues within the root. We are currentlyinvestigating the turnover of the isoflavone conjugates andglyceollin in discrete cell populations in infected and elicitor-treated root, hypocotyl, and leaf tissues. Hopefully, thesestudies will allow us to better define the potential roles ofpreformed and induced isoflavones and glyceollin in theresistance of each of these tissues.We are particularly interested in the finding of an addi-
tional, unidentified compound and its conjugates in hypocotyltissues. This molecule is present nearly exclusively in hypo-cotyl tissues at levels equal to or greater than the daidzein andgenistein conjugates. It may represent an additional metabo-lite of particular importance to the hypocotyl.The dramatic differences in levels of and distribution of the
isoflavones and their conjugates with seedling developmentunder different light conditions will provide us with veryvaluable tools for future studies. Developmental and light-related effects on the infection of various soybean tissues havebeen well documented, particularly by Ward and co-workers(4, 5, 20, 26, 27) and Paxton and Chamberlain (24).The potential role of flavonoids and isoflavonoids in the
chemoattraction of Bradyrhizobium japonicum (18) and inthe induction of nodulation genes in B. japonicum (19)prompted the current examination of their distribution inseed and root exudates. B. japonicum infects soybean roots inthe zone of emerging root hairs (6). PMG infects roots afteraggregation and encystment of zoospores in the zone ofelongation (2). Thus, information on the exudation of thesecompounds from the seed and at specific points along thedeveloping root could provide valuable background for futureexperiments on soybean-PMG interactions as well.To obtain this information, seeds were germinated and
seedlings grown in a specially designed aeroponic chamber.Root exudates were collected over just a 30 min period byplacing short wicks at any specific point from the root tip tothe crown. Data from these experiments suggest that daidzeinand genistein conjugates are very rapidly released from im-bibing seed and roots at levels (1-10 uM) consistent with apotential role in chemotaxis or nodulation gene induction.Several lines of evidence suggest that the release of the isofla-vones may be a specific and regulated phenomenon. First ofall, the profiles of root and seed exudates are not a simplereflection of the profiles the intact tissues; in fact, we havedetected major and unique aromatic metabolites in seed androot exudates which are not present as major metabolites inintact root or seed tissues. The role of these metabolites assignal molecules remains to be established. Second, only avery small fraction of the total isoflavones in the seed arereleased, and the released isoflavones accumulate in a satu-rable manner. Finally, similar saturation kinetics of releaseare reestablished after the imbibing medium is replaced withfresh medium. Taken together, these results suggest the pos-sibility that soybean seed may possess a feedback mechanismto regulate the exudation of its very large stores of isoflavonesignal molecules.
ACKNOWLEDGMENTS
The author would like to acknowledge Drs. Fritz Schmitthenner,Arthur Olah, and Madge Graham for their helpful discussions andencouragement.
LITERATURE CITED
1. Barz W, Hoesel W (1979) Metabolism and degradation of phe-nolic compounds in plants. In T Swain, JB Harborne, CF VanSumere, eds, Biochemistry of Plant Phenolics, Recent Ad-vances in Phytochemistry, Vol 12. Plenum Press, New York,pp 339-369
2. Beagle-Ristaino JE, Rissler JF (1983) Histopathology of suscep-tible and resistant soybean roots inoculated with zoospores ofPhytophthora megasperma f. sp. glycinea. Phytopathology 73:590-595
3. Bhattacharyya MK, Ward EWB (1985) Differential sensitivityof Phytophthora megasperma f. sp. glycinea isolates to glyceol-lin isomers. Physiol Plant Pathol 27: 299-310
4. Bhattacharyya MK, Ward EWB (1986) Expression of gene spe-cific and age related resistance and the accumulation of gly-ceollin in soybean leaves infected with Phytophthora megas-perma f. sp. glycinea. Physiol Plant Pathol 29: 105-111
5. Bhattacharyya MK, Ward EWB (1986) Resistance, susceptibilityand accumulation of glyceollins I-IlI in soybean organs inoc-ulated with Phytophthora megasperma f. sp. glycinea. PhysiolMol Plant Pathol 29: 227-237
6. Bhuvaneswari TV, Bhagwat AA, Bauer WD (1981) Transientsusceptibility of root cells in four common legumes to nodu-lation by rhizobia. Plant Physiol 68: 1144-1149
7. Buttery BR, Buzzell RI (1975) Soybean flavonol glycosides:identification and biochemical genetics. Can J Bot 53: 219-224
8. Buzzell RI, Buttery BR (1973) Inheritance of flavonol glycosidesin soybeans. Can J Genet Cytol 15: 865-867
9. Buzzell RI, Buttery BR (1974) Flavonol glycoside genes in soy-bean. Can J Genet Cytol 16: 897-899
10. Caetano-Anolles G, Crist-Estes DK, Bauer WD (1988) Chemo-taxis of Rhizobium meliloti to the plant flavone luteolin re-quires functional nodulation genes. J Bacteriol 170: 3164-3169
11. Graham TL (1989) Constitutive conjugates of daidzein and gen-istein may play multiple roles in early race specific antibioticresistance in soybean. Phytopathology 79: 1199
12. Graham TL (1991) A rapid, high resolution high performanceliquid chromatography profiling procedure for plant and mi-crobial aromatic secondary metabolites. Plant Physiol 95: 584-593
13. Graham TL, Graham MY (1991) Glyceollin elicitors inducemajor but distinctly different shifts in isoflavonoid metabolismin proximal and distal soybean cell populations. Mol PlantMicrobe Interact (in press)
14. Graham TL, Kim JE, Graham MY (1990) Role of constitutiveisoflavone conjugates in the accumulation of glyceollin insoybean infected with Phytophthora megasperma. Mol PlantMicrobe Interact 3: 157-166
15. Hahn MG, Bonhoff A, Grisebach H (1985) Quantitative local-ization of the phytoalexin glyceollin I in relation to fungalhyphae in soybean roots infected with Phytophthora megas-perma f. sp. glycinea. Plant Physiol 77: 591-601
16. Hosel W, Barz W (1975) ,B-Glucosidase from Cicer arietinum L.Purification and properties ofisoflavone-7-O-glucoside-specific,B-glucosidases. Eur J Biochem 57: 607-616
17. Jacobs M, Rubery PH (1988) Naturally occurring auxin trans-port regulators. Science 241: 346-349
18. Khan MMA, Bauer WD (1988) Chemotaxis of Bradyrhizobiumjaponicum towards flavones and isoflavones from soybean(abstract No. 760). Plant Physiol 86: S-127
19. Kosslak RM, Bookland R, Barkei J, Paaren HE, Appelbaum ER(1987) Induction of Bradyrhizobium japonicum common nodgenes by isoflavones isolated from Glycine max. Proc NatlAcad Sci USA 84: 7428-7432
602 GRAHAM
www.plantphysiol.orgon September 2, 2018 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.

FLAVONOID DISTRIBUTION IN SOYBEAN
20. Lazarovits G, Stoessel P, Ward EWB (1981) Age related changesin specificity of glyceollin production in the hypocotyl reac-
tions of soybean to Phytophthora megasperma var. sojae.Phytopathology 71: 94-97
21. Long SR (1989) Rhizobium-legume nodulation: life together inthe underground. Cell 56: 203-214
22. Maxwell CA, Hartwig UA, Joseph CM, Phillips DA (1989) Achalcone and two related flavonoids released from alfalfa rootsinduce nod genes of Rhizobium meliloti. Plant Physiol 91:842-847
23. McClure JW (1975) Physiology and functions of flavonoids. InJB Harborne, IJ Mabry, H Mabry, eds, The Flavonoids. Aca-demic Press, New York, pp 970-1055
24. Paxton JD, Chamberlain DW (1969) Phytoalexin productionand disease resistance in soybeans as affected by age. Phyto-pathology 59: 755-777
25. Peters NK, Verma DPS (1990) Phenolic compounds as regulatorsof gene expression in plant-microbe interactions. Mol PlantMicrobe Interact 3: 4-8
26. Ward EWB, Buzzell RI (1983) Influence of light, temperatureand wounding on the expression ofsoybean genes for resistanceto Phytophthora megasperma f. sp. glycinea. Physiol Mol PlantPathol 23: 401-409
27. Ward EWB, Lazarovits G (1982) Temperature induced changesin specificity in the interaction of soybeans with Phytophthoramegasperma f. sp. glycinea. Phytopathology 72: 826-830
28. Wiermann R (1981) Secondary plant products and cell and tissuedifferentiation. In EE Conn, ed, The Biochemistry of Plants,Vol 7. Academic Press, New York, pp 85-116
29. Yoshikawa M, Yamamuchi K, Masago H (1978) Glyceollin: itsrole in restricting fungal growth in resistant soybean hypocotylsinfected with Phytophthora megasperma var. sojae. PhysiolPlant Pathol 12: 73-82
603
www.plantphysiol.orgon September 2, 2018 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.





























