Embed Size (px)
Citation preview

Fish Gill Morphology: Inside OutJONATHAN M. WILSON1* AND PIERRE LAURENT2,3
1Centro Interdisciplinar de Investigacao Marinha e Ambiental (CIIMAR),4150-180 Porto, Portugal2Centre d’Ecologie et de Physiologie Energetiques, CNRS, Strasbourg,67037 France3Department of Biology, McMaster University, Hamilton,Ontario Canada L8S 4K1
ABSTRACT In this short review of fish gill morphology we cover some basic gross anatomy aswell as in some more detail the microscopic anatomy of the branchial epithelia from representativesof the major extant groups of fishes (Agnathans, Elasmobranchs, and Teleosts). The agnathanhagfishes have primitive gill pouches, while the lampreys have arch-like gills similar to the higherfishes. In the lampreys and elasmobranchs, the gill filaments are supported by a completeinterbranchial septum and water exits via external branchial slits or pores. In contrast, the teleostinterbranchial septum is much reduced, leaving the ends of the filaments unattached, and themultiple gill openings are replaced by the single caudal opening of the operculum. The basicfunctional unit of the gill is the filament, which supports rows of plate-like lamellae. The lamellae aredesigned for gas exchange with a large surface area and a thin epithelium surrounding a well-vascularized core of pillar cell capillaries. The lamellae are positioned for the blood flow to becounter-current to the water flow over the gills. Despite marked differences in the gross anatomy ofthe gill among the various groups, the cellular constituents of the epithelium are remarkably similar.The lamellar gas-exchange surface is covered by squamous pavement cells, while large, mitochondria-rich, ionocytes and mucocytes are found in greatest frequency in the filament epithelium. Demandsfor ionoregulation can often upset this balance. There has been much study of the structure andfunction of the branchial mitochondria-rich cells. These cells are generally characterized by a highmitochondrial density and an amplification of the basolateral membrane through folding or thepresence of an intracellular tubular system. Morphological subtypes of MRCs as well as somemethods of MRC detection are discussed. J. Exp. Zool. 293:192–213, 2002. r 2002 Wiley-Liss, Inc.
The last comprehensive reviews of fish gillanatomy date from the 1980s (Hughes, ’84;Laurent, ’84, ’89). Rather than marking the endof morphological studies of the gills, they haveserved as a reference point from which work hascontinued. The fish’s gills anatomical complexityand functional importance and diversity havecontributed to its continuing study.This anatomical review will include description
of the basic gross anatomy of the gills followed bythe finer levels of organization, the filament andlamellae, and a detailed description of the celltypes in the branchial epithelia. We will limitourselves to a summary of the gross and micro-scopic morphology of representatives from theclasses Agnatha (jawless fishes), Elasmobranchio-morphi (cartilaginous fishes), and Osteichthyes(bony fishes). From the latter class we will onlycover the teleost fishes and refer the reader to theexcellent reviews of the gross and microscopicanatomy of the gill that have been made by
Hughes (’84) and Laurent (’84) for informationon the Chondrostei (e.g., Acipenser), Holostei (e.g.,Amia, Lepisosteus), and Dipnoi (e.g., Neocerato-dus, Lepidosiren, Protopterus). We will, however,make some cursory reference to representativesfrom these groups when interesting points arise.We refer to recent morphological studies whenever possible while also including citations ofimportant earlier works.
In addition to the reviews by Hughes (’84) andLaurent (’84, ’89), additional information can besought in a number of reviews made on particularaspects of fish gill morphology related to technique(TEM by Pisam and Rambourg, ’91; SEM by
*Correspondence to: Jonathan Wilson, CIIMAR, Centro Inter-disciplinar de Investigacao Marinha e Ambiental, Rua do CampoAlegre 823, 4150-180 Porto, Portugal. E-mail: [email protected]
Received 9 April 2002; Accepted 10 April 2002Published online in Wiley InterScience (www.interscience.wiley.
com). DOI: 10.1002/jez.10124
r 2002 WILEY-LISS, INC.
JOURNAL OF EXPERIMENTAL ZOOLOGY 293:192–213 (2002)

Olson, ’96) and function (Laurent and Perry, ’95;Jurss and Bastrop, ’96; Perry, ’97, ’98).
GROSS ANATOMY
The location and basic structures of the gills ofthe agnathan hagfishes and lampreys, elasmo-branchs, and teleosts are illustrated in Fig. 1. Inall fishes, the gills are bilaterally situated on eitherside of the pharynx and are composed of a series ofpouch-like or arch-like structures that provide thephysical support for the delicate gill filaments alsotermed primary lamellae. The gills of the lampreysand higher fishes share a similar arch-likearrangement with intervening branchial slits forthe water to pass laterally from the buccophar-
yngeal cavity through the gills and out. Thehagfish gill does not have recognizable arch-likestructures, and so its pouch gills will be dealt withseparately.
Lining the sides of the gill arches are rows ofregularly spaced filaments projecting posteriolat-erally. Each row or stack of filaments constitutes ahemibranch, while a set of hemibranchs, one oneach side of the arch, constitutes a holobranch.The gills of teleost fishes are composed of four suchholobranchs spaced between five branchial slits(chambers) (Fig. 1d) while the elasmobranchshave an extra hemibranch on the anterior side ofthe first branchial slit (Fig. 1c). The posterior sideof the final branchial chamber does not have ahemibranch. Not included in the above count of
Fig. 1. General schematics of (a) hagfish, (b) lamprey, (c) elasmobranch, and (d) teleost gills. More detailed illustrations ofthe (e) hagfish pouches and (f,g,h) branchial arches of the respective groups are in the lower panels. Arrows indicate thedirection of water flow.
FISH GILL MORPHOLOGY 193

hemibranchs is the pseudobranch, which is areduced hemibranch present in most teleosts andelasmobranchs in association with the spiracles(Fig. 1c; Laurent and Dunel, ’84). The spiraclesfunction as one-way valves for the entry ofrespiratory water. Within the elasmobranchs, thespiracle is reduced in fast-swimming pelagicspecies, while in bottom dwelling species thespiracle is enlarged. In the teleosts, the spiraclesare lost but the pseudobranch remains. Thepseudobranch does not function in gas exchangelike the other hemibranchs and in fact receivesoxygenated blood. Its function remains somewhatof a mystery (Laurent and Dunel, ’84; Bridgeset al., ’98). The lampreys have six holobranchs andseven branchial slits (pouches). The hagfish gillpouches (5–14 pairs) are either disk shaped orbiloped, and each pouch is composed of thehemibranchs of adjacent gill arches. The hagfishgill lacks distinct holobranchs.In the lampreys and elasmobranchs, the fila-
ments are supported for almost their entire lengthby an interbranchial septum containing a verticalsheet of connective tissue that runs from the archto the outer body wall (Fig. 1f,g). The afferent ortrailing edge1 of the filament is continuous withthe interbranchial septum, while the efferent orleading edge of the filament is unattached. In theteleosts, the interbranchial septum is reduced andthe majority of the distal length of the filamentsare unattached or free, giving them a filamentousappearance.In the teleosts and elasmobranchs, the indivi-
dual gill arches are supported by vertical elementsof the cartilaginous branchial skeleton, which runthrough the medial portion of each arch and arepartially calcified in the teleosts. In the elasmo-branchs, the interbranchial septum is supportedby skeletal branchial rays, which extend from themain skeletal support in each arch. In the teleosts,these branchial rays are present in the filamentsand in conjunction with abductor muscles areimportant in the positioning of the filament in thewater flow (see Hughes, ’84). The tips of filamentsfrom opposite hemibranchs in the branchial slitchambers come into close proximity to each otherto form a sieve like arrangement (Fig. 1g,h). Theagnathan visceral skeleton is totally unlike that ofthe jawed fishes as it forms a fenestrated frame-
work of continuous cartilage immediately underthe skin surrounding the pharynx. This branchialbasket is less developed in the hagfish than thelamprey.
The filaments are dorsoventrally flattened, andtheir surface area is greatly increased by second-ary folding to form lamellae (Fig. 2). The lamellaehave a thin plate-like appearance with a vascularcore covered by a thin epithelium on either side(Fig. 3c). The lamellar blood space is lined withpillar cells. The gill filament is considered thebasic functional unit or subdivision of the gill. Itsepithelium contains ionoregulatory cells while italso supports the lamellae, which are the basicrespiratory unit.
The lamellae of lampreys and higher fishes areproduced by cross-folding (perpendicular to longaxis of filament) of the upper and lower sides ofthe filament epithelium (Fig. 2). The lamellae arein evenly spaced rows along the length of thefilament (Fig. 2, 3f,g), although in the elasmo-branchs they are at an angle slightly oblique to thelong axis of the filament (efferent edge closer tothe pharynx than the afferent edge). The outermargins of the lamellae from adjacent filamentscome into close proximity to each other completing
Fig. 2. Scanning electron micrograph of two gill filamentsof the teleost tilapia (O. mossambicus). One filament is lyingflat while the other (left) has its leading edge facing up. Fromthese two views the orientation of the lamellae (L), which arepresent on both faces of the filament, is made obvious. Thearrow indicates the direction of water flow across thefilaments. The filament edge facing the water flow is theleading or efferent edge, referring to water and blood flows,respectively. The trailing or afferent edge of the filament isvery flat and typically contains MRCs. SEM morphometricstudies of the MRC typically make use of this area because it isflat (see Fig. 10c). Scale bar¼100mm (from J.M. Wilson,unpublished). Abbreviations: af., afferent; ef., efferent; L,Lamellae.
1In all fishes, the flow of water across the gill is counter-current tothe flow of blood. When referring to the flow of blood, the terms‘‘afferent’’ and ‘‘efferent’’ (‘‘to’’ and ‘‘from’’ the lamellae) are used.The terms ‘‘leading’’ and ‘‘trailing’’ edge are used in relation to thedirection of water flow across the filament and lamellae.
J.M. WILSON AND P. LAURENT194
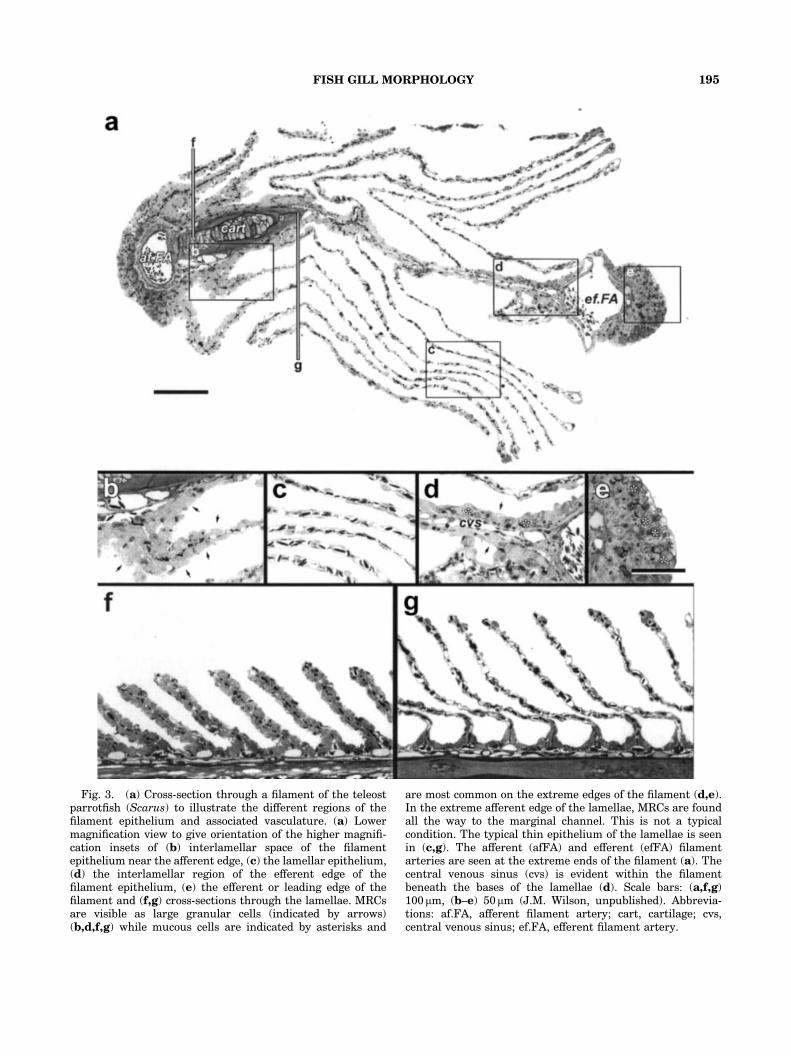
Fig. 3. (a) Cross-section through a filament of the teleostparrotfish (Scarus) to illustrate the different regions of thefilament epithelium and associated vasculature. (a) Lowermagnification view to give orientation of the higher magnifi-cation insets of (b) interlamellar space of the filamentepithelium near the afferent edge, (c) the lamellar epithelium,(d) the interlamellar region of the efferent edge of thefilament epithelium, (e) the efferent or leading edge of thefilament and (f,g) cross-sections through the lamellae. MRCsare visible as large granular cells (indicated by arrows)(b,d,f,g) while mucous cells are indicated by asterisks and
are most common on the extreme edges of the filament (d,e).In the extreme afferent edge of the lamellae, MRCs are foundall the way to the marginal channel. This is not a typicalcondition. The typical thin epithelium of the lamellae is seenin (c,g). The afferent (afFA) and efferent (efFA) filamentarteries are seen at the extreme ends of the filament (a). Thecentral venous sinus (cvs) is evident within the filamentbeneath the bases of the lamellae (d). Scale bars: (a,f,g)100 mm, (b–e) 50 mm (J.M. Wilson, unpublished). Abbrevia-tions: af.FA, afferent filament artery; cart, cartilage; cvs,central venous sinus; ef.FA, efferent filament artery.
FISH GILL MORPHOLOGY 195

a sieve-like arrangement for the respiratory waterto pass through. There is a space left between thetrailing edge of the lamellae and the lateral wall ofthe interbranchial septum so the water that haspassed between the lamellae can flow on outbehind the rows of lamellae. This water channelor canal facilitates the flow of water counter to thelamellar blood flow.The analogous structure to the gill filament in
the hagfish is the primary gill fold. The primarygill folds are arranged radially around the centralaxis of the pouch like the lines of longitude on aglobe (Fig. 1e). Some filaments extend to thecentre of the pouch while others have a low profile.The filament can be subdivided into afferent,respiratory, and efferent regions (Fig. 4). In therespiratory region, lamellae are present as sub-divisions and branchings of the filament throughthird to seventh order folding. The lamellae inhagfish are orientated in the same plane as thefilament, which is unlike the arrangement inlamprey and other fishes in which the lamellaeare perpendicular to the long axis of the filament.Water and blood flows are counter-current.Hughes (’84) has reviewed the relationship
between morphometric parameters of the gill(surface area, diffusion distances) in relation tohabit. A comparison of fast-swimming pelagic(e.g., tuna Thunnus albacares), sluggish benthic(e.g., toadfish Opsanus tua), and air-breathing (e.g.,tree-climbing perch Anabas testudineus) fishesillustrates the striking differences in filamentand lamellae size, shape, number, and spacing
that reflect the respiratory needs of the animals(total gill area relative to body mass¼a �Wb;W¼mass in g). Fast-swimming tunas have a largergas-exchange surface (3,151 �W0.875), while thesluggish toadfish (560.7 �W0.790) and air-breathingclimbing perch (556 �W0.615) fishes have reducedgas-exchange surfaces. The thickness of the bloodto water distance across the lamella is also muchsmaller in the tuna (0.533 mm) compared to theperch (10 mm) (cf. Hughes ’84). Air-breathingfishes generally make use of alternate gas-ex-change organs (modified opercular chamber andswim bladders) for gas exchange. The morpho-metrics of the gills of air-breathing fishes has beenreviewed recently by Graham (’97) and Roy andMunshi (’96).
In the elasmobranchs and agnathans the bran-chial slits open individually to the outside,although there are some exceptions. Outer foldsof skin create the vertical slit like opening to theoutside in the elasmobranchs and the branchio-pores of the agnathans (Fig. 1a,b,c). These open-ings are generally lateral, although in the skatesand rays they are ventral. The teleosts arecharacterized by the universal presence of a bonyoperculum. The operculum develops from a fold inthe hyoid arch and is a hard structure composed ofdermal bone, with a crescent-shaped caudal open-ing (Fig. 1d). The ventral edges are connected byan expandable membrane beneath the pharynx,and the space enclosed is the opercular cavity orspace. The presence of the operculum likely led tothe loss of the first hemibranch in the teleosts
Fig. 4. Section through the gill pouch of the Pacific hagfish(Eptatretus stouti) to illustrate the different subdivision of theprimary fold: afferent, respiratory, and efferent regions. Thissection is actually immunolabeled for Naþ,Kþ-ATPase similar
to Choe et al. (’99). However, in this case due to section pre-treatment the erythrocytes as well as the MRCs produce adetectable signal. Scale bars¼100mm (JM Wilson, unpub-lished).
J.M. WILSON AND P. LAURENT196

since water delivery to this area is poor (Romerand Parsons, ’86). The chimeras, which are alsomembers of the chondrichthyes, differ markedlyfrom the elasmobranchs in the gross morphologyof their gills and associated structures. Thenumerous gill slits are replaced by a singleopening covered by a large fleshy operculum thatextends caudally. The interbranchial septa arealso shorter, the last branchial chamber is closed,and the spiracles are lost. In these respects, theymore closely resemble the teleosts.In the hagfish, water enters the individual
pouches from the pharynx medially via internalbranchial ducts and passes out laterally via anexternal branchial duct (Fig. 1a,e). In the Bdellos-toma hagslime, the external branchial ducts leaddirectly to the outside via individual branchio-pores, while in the Myxine hagfish the ductsextend caudally to form a common external open-ing. At the end of the series of gill pouches on theleft side only is the pharyngocutaneous duct thatconnects the pharynx to the outside. This ductserves as an entry point for respiratory waterwhen the hagfish has its head buried into its prey(dead fish). Hagfish also have a nasal duct dorsalto the mouth that opens into the pharynx andserves as an important entry point for respiratorywater when the body of the hagfish is buried in themud.The water flow through the gills in lampreys is
modified in order to be compatible with theirspecialized suctorial feeding mechanism (Randall,’72). Lampreys are parasitic, and when theirdisc-shaped mouths are attached to a host fish orto the substrate, the water flow via this route tothe gills is blocked. The lampreys make use oftheir external branchiopores for both waterinhalation and exhalation (tidal breathing) (Fig.1f). They have also been observed to draw water inthrough their first few anterior pouches, which isthen exhaled through their posterior pouches(Nekvasil and Olson, unpublished observations).The lattice-like cartilaginous skeleton supportingthe pouches and a well-developed musculatureproduce the pumping action to circulate the waterin the gill pouches. The respiratory and feedingmechanisms are separated internally by a mod-ification of the pharynx. The pharynx in the adultlamprey is split into two ducts: one (dorsal)leading to the esophagus and a ventral blind-ended respiratory tube (branchial duct) thatadjoins the internal gill ducts. The flow of waterthrough the entrance of the respiratory tube iscontrolled by a value, the velum (Fig. 1b). During
feeding, the velum is closed and the body juicessucked from the host pass directly from the mouthto the digestive tract without affecting respirationand vice versa. This tidal mechanism also appearsto be the preferred route of water circulation infree-swimming lampreys and differs from theunidirectional flow through arrangement seen inall other fishes (with the exception of the sturgeon;Burggren and Randall, ’78).
EPITHELIA
The epithelia covering the filament and lamellaecan be differentiated, obviously by location, butalso by thickness, blood circulation (arterioarterialvs. arteriovenous, see Olson, 2002,this issue), and cell type composition (Fig. 3). Inthe hagfish, the thickness of the epitheliumdecreases gradually as the divisions of the surfaceare more complicated (up to sixth-order folding)compared to the lamellae of all other fishes, whichrepresent only first-order folding of the filament(Fig. 4).
In most other fishes, the epithelium coveringthe filament includes both the afferent andefferent edges as well as the spaces between thebases of the lamellae, which is commonly referredto as the filament interlamellar space (ILS) (Fig.3f,g). Within the filament, and bordering much ofthe filament epithelium, is the large centralvenous sinus (CVS) which is part of the arter-iovenous circulation (Olson, 2002, this issue). Thefilamental epithelium is thicker than the lamellarepithelium, typically being composed of three ormore cell layers. The majority of the surface iscovered by cuboidal and squamous pavement cells,while basal undifferentiated cells contact the basallamina and intermediate undifferentiated cells fillthe intervening space. Notably, also within thefilament epithelium are numerous, large mito-chondria-rich cells and mucous cells (see below).
In contrast, the lamellar epithelium overlays thearterioarterial circulation (Olson, 2002, this is-sue). The epithelium is typically one to three celllayers and composed of squamous pavement cellsand basal and intermediate non-differentiatedcells. Modified endothelial cells, called pillar cells,support and define the lamellar blood spaces. Theepithelium sits on a typical basal lamina composedof an electron-dense and a thinner electron lucentlayer. The thickness of the epithelium and basallamina is often less in fishes trying to minimizetheir blood to water diffusion distances in order tooptimize gas exchange (e.g., active pelagic teleost
FISH GILL MORPHOLOGY 197

T. albacares, 0.55 mm, versus sedentary benthicteleost Solea variegata, 2.80 mm, mean total blood–water distance; see Hughes, ’84). Mucous cells andMRCs are not as commonly encountered in thelamellar epithelium, although there are excep-tional cases.
CELL TYPES
Pillar cells (PC)
The pillar cell is a type of modified endothelialcell, which defines the blood spaces within thelamellae and is a cell type unique to the fish gill.They give the lamellae the appearance of a stringof beads when viewed in cross section (Fig. 3c,f,g).The pillar cell body spans the lamellar blood spaceand has a centrally located polymorphic nucleus(Fig. 5). The ends of the pillar cell flare out at thepoint of contact with the interstitial connectivetissue adjacent the basal lamina of the lamellarepithelia to form thin flanges that extend toneighboring pillar cells. The flanges of neighbor-ing pillar cells may form simple abutments orcomplex interdigitations. In the hagfish andlamprey, communicating gap junctions have beenfound between neighboring pillar cells [Bartelsand Decker (’85) and Bartels and Potter (’93),respectively]. The flanges are kept thin (0.02–1 mm; Hughes ’84) to minimize the blood to waterdiffusion distances and are generally devoid oforganelles except for microfilaments. Associatedwith the pillar cell body are columns of collagenbundles that entwine with collagen fibrils in theinterstitial connective tissue on the vascular sideof the basal lamina. These collagen bundles re-main extracellular but are enveloped by the pillarcell and sealed in by tight junctions (Newstead,’67). The hagfish is an exception, since thecollagen bundles are not deeply enveloped by thepillar cell body and tight junctions are absent.Instead, the collagen bundles are surrounded bygroups of pillar cells (Mallatt and Paulsen, ’86). Itis important to isolate the collagen from the bloodspace because it will cause blood clotting. Thesecollagen bundles presumably function to preventdistension and collapse of the blood space. Thepillar cells also have bundles of microfilaments inthe peripheral cytoplasm that run parallel to thecollagen bundles and have been found to becontractile although nervous control is not present(Laurent, ’84). Also crowded within the pillar cellbody are mitochondria, free ribosomes and variousmembrane bound organelles, although a Golgiapparatus, rough and smooth endoplasmic reticu-
lum (RER and SER, respectively) are not common.Unlike true endothelial cells, pillar cells lack densecore Weibel-Palade bodies (Newstead, ’67). Trueendothelial cells do, however, line the outermarginal channels of the lamellae (Newstead, ’67;Hughes and Weibel, ’72).
Pavement cells (PVC)
The most abundant cell type covering theepithelium (490% of the surface area) is thesquamous to cuboid-shaped cell commonly re-ferred to as the pavement or respiratory cell.The apical surface of PVCs is usually large andpolygonal and may have microridges (referred to
Fig. 5. Transmission electron micrograph (TEM) of across-section through the lamellae pillar cell of the rainbowtrout (O. mykiss). Note the thin flanges (large arrowhead) andcentrally located polymorphic nucleus of the pillar cell (pc).Smaller arrows indicate the basal lamina and the smallerarrowheads the extracellular collagen bundles spanning theblood space while embedded in the pillar cell. Electron-densered blood cells (rbc) are seen within the blood spaces. Scalebar¼5 mm (J.M. Wilson, unpublished).
J.M. WILSON AND P. LAURENT198

microplicae when viewed in cross-section) ormicrovilli (finger-like projections) (Laurent, ’84;Crespo, ’82; Olson, ’96; Wilson et al., 2002) (Figs.6, 7, 8a, 10b, c, and 11). When microridges arepresent, they typically form concentric rings. Adouble ridge marks the boarder of neighboringPVCs (Fig. 10c). The types of apical featurespresent show variation between species and alsowithin and between the different gill epithelia.Pavement cells typically have low mitochondrialdensities and unelaborated basolateral mem-branes. The nucleus in squamous pavement cellsis compressed while in cuboidal pavement cells itis rounded. Pavement cells have the typicalintracellular organelles, rough and smooth endo-plasmic reticulum, Golgi apparatus, lysosomes,and vesicles with contents of various electrondensities. In the non-teleost fishes, pavement cellsdisplay abundant mucous secretory granules inthe apical cytoplasm (Laurent, ’84; Bartels, ’85;Mallatt and Paulsen, ’86) (Fig. 6,11). Thesevesicles fuse with the plasma membrane and expeltheir contents. Pavement cells are joined bydesmosomes, and the peripheral cytoplasm con-tains microfilaments. The tight junctions asso-ciated with pavement cells have multiple strandsand are not characterized as leaky (Sardet et al.,’79; Kawahara et al., ’82; Bartels, ’88). In thehagfish and lamprey, communicating gap junc-tions have been found between neighboringpavement cells, and pavement cells and un-differentiated cells, however, not with MRCs
(Bartels ’88; Bartels and Potter ’93, respectively).The presence of communicating gap junctions issignificant because they allow the electrochemicalcoupling of the cells of the epithelium which isimportant in coordinating cell growth, differentia-tion, and function. No studies have been under-taken in elasmobranchs to specifically determinethe presence of gap junctions.
In teleost fishes there is conflicting evidence forthe presence of PVC gap junctions in the branchialepithelium. In freeze-fracture electron microscopystudies on five euryhaline species (Anguillaanguilla, Oncorhynchus mykiss, Fundulus hetero-clitus, Lebistes reticulatus, and Mugil capito),Sardet and co-workers (Sardet, ’77; Sardet et al.,’79) were unable find evidence for the presence ofgap junctions; however, Kawahara and co-workers(’82) using the same approach detected gapjunctions between gill PVCs of Oplegnethusfasciatus. Kawahara et al. (’82) were unable tofind gap junctions between PVCs and MRCs. In arecent study by Sandbacka et al. (’98), commu-nicating junctions between trout (O. mykiss) gillpavement cells in primary culture were demon-strated using a dye-coupling technique. They alsosuggested that the clustered intramembranousparticles of the P-face shown in Fig. 3 of Sardetet al. (’79) are gap junctions although they are notrecognized as such. Unfortunately, this figure
Fig. 6. TEM of the apical region of a pavement cell fromthe dogfish Squalus acanthias). Evident are the apicalsecretory granules (asterisks) and microfilament network, aswell as mitochondria (m), Golgi apparatus (g), and vesicles ofvarious shapes and sizes. Glycocalyx is also recognizableattached to the exterior surface. Scale bar¼0.5 mm (J.M.Wilson, unpublished).
Fig. 7. TEM of a mucocyte of the freshwater chum salmonlarvae (Oncorhynchus keta). Note the flattened basally locatednucleus, peripheral cytoplasm with abundant endoplasmicreticulum (er), and some mitochondria (m), and the largemucin granules dominating the space of the cell (asterisks).Scale bar¼5 mm (J.M. Wilson and E.P. Groot, unpublished).
FISH GILL MORPHOLOGY 199

Fig. 8. (a,b) TEMs of the MRC of the freshwater chumsalmon larvae (O. keta) using the potassium ferrocyanide-reduced osmium stain described by Pisam et al. (’87). Note thestrong staining of the tubular system (ts) and the plasmamembrane while other membrane systems remain weaklystained. (c) TEM showing the connections of the tubular
system with the basolateral membrane (indicated by arrows)in the neon tetra (Paracheirodon innesi) MRC using conven-tional heavy metal staining. Scale bars: (a) 1 mm; (b,c) 0.5 mm[(a,b) J.M. Wilson and E.P. Groot, unpublished; (c) J.M.Wilson, unpublished]. Abbreviations: MRC, mitochondria-richcell; PVC, pavement cell; tvs, tubulovesicular system.
J.M. WILSON AND P. LAURENT200

shows the fracture face of the apical membranes oftwo neighboring pavement cells and not thelateral membranes as would be necessary to makesuch a claim. It is possible that gap junctions inprimary cultures are an artefact of being underculture conditions as in situ attempts to demon-strate dye-coupling were unsuccessful due totechnique reasons (mucus layer). Thus, furtherwork seems justified to clarify this importantquestion.PVCs have been shown to be involved in the
covering and uncovering of MRCs under certainconditions (Goss et al., ’94; Bartels et al., ’96;Daborn et al., 2001), but the role of the PVC in theprocess is somewhat secondary to that of the MRC.Generally PVCs themselves have been shown to bemorphologically unresponsive to changes in en-vironmental conditions (Laurent, ’84). Part of theproblem stems from the fact that the MRCs aregenerally the focus of attention in morphologicalstudies and the changes in PVCs may be over-looked. It also might be difficult to recognizethese changes if they only occur in a subpopulationof PVCs since there are proportionally manymore PVCs. With that said there have beentwo studies that have shown that PVCs increaseapical surface area (densities of microvilli andmicroplicae) in response to hypercapnia andalkaline exposure [Goss et al. (’94) and Laurentet al. (2000), respectively]. Laurent et al. (’94)have also found studded subapical vesicles thatresemble the proton pump containing vesiclesfrom other animals (Brown et al., ’87). Thesemorphological characteristics of PVCs help tosupport the hypothesis that they are the sites ofproton pump-driven sodium uptake (Perry, ’97;Wilson et al., 2000a). Perhaps more attentionfocused on these cells will reveal more adapta-tional changes of this somewhat underrated celltype.
Nondifferentiated cells(basal and intermediate)
The basal and intermediate layers of theepithelium contain cells that are characterizedby a high nucleus to cytoplasm ratio. These cellsappear undifferentiated and serve as the progeni-tors for the other terminally differentiated epithe-lial cell types (PVCs and MRCs). In teleost fishes,at least, mitosis of undifferentiated cells is morecommon in the filament than lamellar epithelium(Chretien and Pisam, ’86; Laurent et al., ’94). Thenon-differentiated cells share some common fea-
tures with the PVCs (RER, lysosomes, vesicles,and vacuoles). However, relative to PVCs, non-differentiated cells have more abundant freeribosomes. In the lamellar epithelium, the nucleusof the basal undifferentiated cells often sits on thebasal lamina above the pillar cell body. In epitheliathat are thicker than two cell layers (filamentepithelium), intermediate non-differentiated cellsare found. Desmosomes are found between non-differentiated cells and pavement cells. Hemides-mosomes connect the basal undifferentiated cellsto the basal lamina anchoring the epithelium.Caveolae, non-clathrin-coated plasma membraneinvaginations, can also be found along this cellboarder as well.
Mucous cells (goblet cells)
Goblet type mucous cells are large ovoid cellsthat are composed mostly of large apical mucoussecretory granules (Figs. 3 and 7). The nucleusand cytoplasm are usually flattened and in a basalposition. Within the cytoplasm is the cellularmachinery for producing the mucin (ER, Golgi,mitochondria). The mucin granules are generallyelectron lucent or only moderately electron densewith some variability between granules within thesame cell. The mucous cells are commonly foundin the filament epithelium in the followingfrequency: efferent edges 4 afferent edge 4interlamellar space 4 base of lamellae outermargin of lamellae. However, their distribu-tion and numbers do show variability from thisgeneral plan. There tends to be a reduction inthe number of mucous cells in seawater versusfreshwater fish (Laurent and Hebibi, ’89).Goblet cells have been found in all fishes withthe exception of the hagfishes (Mallatt andPaulsen, ’86).
Mucous cells have also been characterized bythe types of mucin in their granules. The periodicacid-Schiff (PAS) method is used to detect neutralmucins, and the variations of the alcian bluemethod to detect acidic mucins (pH 2.5, carboxy-lated and sulfated mucins; pH 0.5, sulfatedmucins, and Quintarelli’s acid hydrolysis sialomu-cins) (e.g., Saboic-Moraes et al., ’96; Watrin andMayer-Gostan, ’96). Rojo et al. (’96) have also usedlectin histochemistry to describe different types ofmucous cells in the brown trout (Salmo trutta).The different lectins bind specifically to thedefined carbohydrate moieties present in themucin granules.
FISH GILL MORPHOLOGY 201

Neuroepithelial cells (NEC)
Neuroepithelial cells are found deep within thefilament epithelium along the full length of theefferent edge although concentrated near filamenttips (Dunel-Erb et al., ’94). Neuroepithelial cellshave been described in elasmobranch and teleostfishes. The fish neuroepithelial cells share com-mon characteristics with the neuroepithelial cellsdescribed within the wall of the respiratory tractof mammals (Laurent, ’84). These cells can bereadily identified by detecting the biogenic aminefluorescence resulting from formaldehyde treat-ment (Falck et al., ’62), which has been confirmedto include 5-HT by immunohistochemistry (Bailyet al., ’92). A defining feature of these cells is thepresence of dense-cored vesicles (80–100nm) andtheir location directly adjacent the basal lamina.The most significant characteristic of this cell is itsinnervation. Within the filament is a densesubepithelial nervous network composed of mostlyunmyelinated nerve fibers. Processes of unmyeli-nated nerve fibers cross the basal lamina, andmultiple contacts are made with the same NEC.These cells are believed to function as oxygensensors and to be involved in the regulation ofblood flow (see Sudin and Nilsson, 2002, thisissue).
Mitochondria-rich cells (MRCs)or chloride cells (CCs)
The descriptive morphological study of the gill isdominated by the mitochondria-rich chloride celland its subtypes (Pisam and Rambourg, ’91; Jurssand Bastrop, ’95; Perry, ’97; Evans et al., ’99).MRCs tend to be concentrated in the afferentregion of the filament epithelium and have anintimate association with the arteriovenous circu-lation, notably the central venous sinus, althoughin the interlamellar region MRCs are also asso-ciated with the basal channels of the lamellararterioarterial circulation (Laurent, ’84; Olson,2002, this issue). A nice example of the distribu-tion within the tilapia (Oreochromis mossambicus)gill filament can be found in the paper by Uchidaet al. (2000) in which laser scanning confocalmicroscopy was used to localize MRCs. Thedefining feature of this cell type is a highmitochondrial density. MRCs also generally havean amplification of their basolateral membraneeither through folding or the presence of a tubularsystem (Fig. 8), although the lamprey freshwater-type MRCs are an exception. Associated with thebasolateral membrane amplification is the impor-
tant ionoregulatory enzyme Naþ,Kþ-ATPase (seeMarshall, 2002, this issue). In the apical region ofMRCs is a collection of tubules and vesicles(tubulovesicular system) that is distinct from thetubular system (Fig. 9). The apical membrane canbe quite variable in appearance ranging fromconcave to convex, sometimes forming deep crypts.In addition, the surface topography has beenshown to be smooth, or having microvilli ofvarying densities and lengths as well as highlybranched, to give a sponge-like appearance. Theuse of morphological features has led to theclassification of different types of MRCs.
The term ‘‘chloride cell’’ relates to the functionof the MRC in Cl� elimination. In seawaterteleosts, the MRCs have quite convincingly beenshown to be sites of active Cl� elimination andhence the name is fitting (Marshall, 2002, thisissue). In the agnathans, elasmobranchs, andfreshwater teleosts, the evidence that MRCs areinvolved in Cl� fluxes is indirect or lacking alltogether. So in order to avoid any confusion on thematter, we will only use the term ‘‘chloride cell’’when referring to the seawater teleost MRC unlessotherwise noted. However, in general, numerousmitochondria in these cells are thought to supplythe ATP for ion-transport proteins to drive thevectorial transport of ions as part of ion and acid–base regulation (see Claiborne, 2002; Marshall,2002).
Teleost MRCs
The teleost MRC is characterized by its elabo-rate intracellular system of branching tubules thatare continuous with the basolateral membrane(tubular system; Fig. 8). This network of anasto-mosing tubules is closely associated with themitochondria and is only excluded from the areaof the Golgi apparatus and the band immediatelybeneath the apical membrane. The elements of thetubular system can be found along side theendoplasmic reticulum making identificationsomewhat problematic. Extracellular space mar-kers such as horseradish peroxidase and ruthe-nium red have been used to clearly establish thebasolateral continuity of the tubular system andits separation from the ER (Philpott, ’80). Pisamand co-workers (’87) have made effective use of thepotassium ferrocyanide reduced osmium staindeveloped by Karnovsky (’71) to unambiguouslydifferentiate the membrane elements of thetubular system from the ER without the needof extracellular markers. The selective heavy
J.M. WILSON AND P. LAURENT202

staining of the tubular system contrasted mark-edly with the poorly stained endoplasmic reticu-lum (Pisam, ’81) (Fig. 8a,b). Environment ionic
conditions have a marked effect on the morphol-ogy of the teleost MRCs, which of course relates tothe role of these cells in active ion transport
Fig. 9. TEMs of the apical regions of the MRCs of (a) Raja erinacei, (b) Acipenser beari, (c) Percafluviatilis, and (d) Protopterus annectens showing the tubulovesicular system. Scale bars¼1 mm (P.Laurent, unpublished). Abbreviation: em, external milieu.
FISH GILL MORPHOLOGY 203

(Perry, ’97; Evans et al., ’99). In sea water, the gillis involved in active ion elimination while infreshwater active ion-uptake takes place.The mitochondria-rich chloride cell (CC) and
accessory cell (AC) are two types of MRC thatare universally expressed in seawater teleosts(Fig. 10). These cells form multicellular complexesin the filament epithelium. The larger chloride cellmay be elongate, ovoid, or cuboidal in shape,depending upon the species. The tubular and thetubulovesicular systems are highly developed, andthe cell is packed with mitochondria. High packingof Naþ,Kþ-ATPase is associated with the CCtubular system (Sardet et al., ’79; Marshall,2002, this issue). The apical membrane of a CCis recessed from the surface of neighboring PVCsand may be further deepened through invagina-tion (Fig. 10c). The smaller accessory cell issuperficially located, semi-lunar or pear shaped,and generally has a less extensive tubular systemand a poorly developed tubulovesicular system.Notably, they have low levels of Naþ,Kþ-ATPaseunlike the CCs (Hootman and Philpott, ’80). TheAC sends cytoplasmic processes into the larger CC,which emerge at the apical surface to form acomplex mosaic. Notably, the tight junctions foundbetween CC and AC contain fewer strands than dothose formed with PVCs (Sardet et al., ’79;Kawakara et al., ’82). Therefore the AC inter-digitations within the CC apical membrane wouldgreatly increase the linear distance of leaky tightjunction, which is functionally important for theparacellular sodium efflux (see Marshall, 2002,this issue). There is also ample evidence that Cl�
efflux occurs transcellularly through the CC(Marshall, 2002, this issue).In freshwater teleost fishes there is less con-
sensus about clear MRC subtypes that mightrelate to the instability and variability of thefreshwater environment itself. Pisam and co-workers, however, have been able to describe aand b sub-types2 of MRCs with electron-lucent(light or pale) and electron-dense (dark) cyto-plasms, respectively, in a number of species,including the guppy (L.reticulatus), loach (Cobitistaenia), grudgeon (Gobio gobio), Atlantic salmon(Salmo salar), and Nile tilapia (Oreochromisniloticus) (Pisam et al., ’87, ’90, ’95). In additionto a pale cytoplasm, a cells were found at the edgesof the interlamellar space (ILS) in contact with the
basal lamina opposite the arterioarterial circula-tion. The b cells, on the other hand, were generallyobserved in the ILS in contact with the basallamina associated with the arteriovenous circula-tion. Also a manganese-lead (Mn-Pb) stain wasfound to selectively stain the contents of apopulation of variably sized membrane boundbodies in the apical cytoplasm of only the b-celltype. The material in the apical vesicles of theb-cell has been shown to include carbohydratematerial (Pisam et al., ’95) and recently Tsai andHwang (’98) have found binding of the lectin WGA(wheat germ agglutinin: sialic acid and N-acet-ylglucosaminyl residues) to the apical vesicles ofa subpopulation of MRCs in tilapia (presumablyb-cells). In a complementary study by Shikanoand Fujio (’98) on the guppy (L. reticulatus),it was shown that the a-cell has high levels ofNaþ,Kþ-ATPase immunoreactivity while the b-cellhas only weak labeling. In a study on rainbowtrout (O. mykiss), and an earlier study on Atlanticsalmon, differentiation of these two subtypes wasnot made (Pisam et al., ’88, ’89).
Transfer to sea water of euryhaline freshwaterfishes, results in the degeneration of the b-cellthrough apoptosis and hypertrophy of the a-cell(Pisam et al., ’87; Wendellar Bonga and van derMeij, ’89). The a-cell is transformed into what isrecognized as the seawater chloride cell (CC). Thetransformation consisted of an increase in the sizeof the a-cell, density of the cytoplasm, number ofmitochondria, the tubular system network with atighter polygonal mesh size, and the apicaltubulovesicular system. A mitochondria-poor celltype has been described by van der Heijden et al.(’99) that possesses a well-developed tubularsystem but has relatively few mitochondria. Thiscell increases in frequency following seawateracclimation and infrequently contacts the water.No function was suggested for this cell type.
Pisam et al. (’88, ’89, ’90) aided by their specificheavy metal stains have also characterized theaccessory cell type in some freshwater euryhalinespecies. These cells are associated with the a-celltype in tilapia and Atlantic salmon but b-cell typein brown trout (S. trutta) and are similar to the ACdescribed in the seawater fish gill, but they are lesscommon and do not send cytoplasmic processesinto the apical cytoplasm of the CC. Upon transferto sea water or in anticipation of transfer (smolt-ing), their numbers increase and interdigitationsare present. The AC are presumably present infishes living in fresh water in order to facilitate therapid switch to NaCl elimination. In the tilapia
2It should be noted that the a and b cells described by Pisam and co-workers are not meant to be analogous to the a and b cellsof the turtle bladder (e.g., Steinmetz and Andersen, ’82).
J.M. WILSON AND P. LAURENT204

Fig. 10. Transmission (TEM) and scanning electron(SEM) micrographs of the apical crypt of the seawatermitochondria-rich chloride cell (CC) and accessory cell (AC).(a) TEM showing CC and AC sharing an apical crypt. Arrowsindicate AC inclusions into the CC. Compare the shallow tightjunctions found between the AC and CC with those of thepavement cells PVCs (arrowheads). (b) High-magnificationSEM of the apical crypt showing AC inclusions into the CC
(arrows). (c) Lower-magnification SEM showing deep-holeappearance of MRCs and the large neighboring PVCs withtheir microridged surface arranged concentrically [(a) Moronelabrax, P. Laurent and S. Dunel-Erb, unpublished; (b) Soleasolea, modified from Laurent (’84) with permission fromAcademic Press; (c) O. mossambicus, J.M. Wilson, unpub-lished]. Scale bars: (a,b) 0.5 mm, (c) 10 mm.
FISH GILL MORPHOLOGY 205

(Alcolapia grahami) of extremely alkaline (pH 10)Lake Magadi in Kenya, Laurent et al. (’95) havefound active seawater ACs associated with largerCC type MRCs. The lake water has very highHCO3
� and CO32� concentrations of 40 and 265
mEq �L�1, respectively, with Naþ and Cl� at 342and 108 mEq �L�1, respectively. In the proposedmodel, the CC–AC complex facilitates paracellularNaþ efflux via the leaky tight junction while HCO3
�
is pumped out by the CC.In freshwater tilapia (O. mossambicus), Lee and
co-workers (’96) have defined three MRC typesaccording to the apical membrane appearance bySEM and categorized them as either wavy-convex,shallow-basin, or deep-hole. The ‘‘wavy-convex’’type has a convex apically exposed area withvariable ornamentation with microvilli and arelatively large two-dimensional area. The surfaceof the ‘‘shallow-basin’’ type MRCs is flat butrecessed below neighboring PVCs. The density ofthe ‘‘wavy-convex’’ type MRCs increases in lowNaCl and the ‘‘shallow-basin’’ type MRCs increasein low Ca2þ water, suggesting roles in NaþCl� andCa2þ uptake, respectively (Chang et al., 2001). The‘‘deep-hole’’ MRCs are also recessed betweenPVCs like the ‘‘shallow-basin’’ type, but they forma deeper apical crypt characteristic of seawaterCCs (Lee et al., 2000). In the fathead minnow(Pimephales promelas), the ‘‘deep-hole’’ MRCsappear in response to exposure to acid conditions,suggesting a role in acid excretion (Leino andMcCormick, ’84). However, in other species, likethe rainbow trout, this system of classification isnot applicable, and it remains to be determined ifthe subtypes of MRCs can be differentiated onapical appearance alone (Galvez et al., 2002).Although MRCs tend not to be as frequently
found in the lamellae as in the filament epitheliumin freshwater and especially seawater fishes,acclimation of trout to ion-poor (o0.01mM[Naþ]) conditions, elicits a massive proliferationof MRCs in the lamellar epithelium (Laurent et al.,’85; Perry and Laurent, ’89; Greco et al., ’96). Thenumber and area of exposed surface of these cellsboth increase dramatically. The effects of ion-poorwater on MRC proliferation can also be repro-duced using hormone treatments with cortisol andgrowth hormone (Laurent and Perry, ’90; Bindonet al., ’94). Since MRC are generally quite large,proliferation on the lamellae results in thethickening of the epithelium, and, consequently,the blood to water diffusion distance is increased(reviewed by Perry, ’98). This has negativeconsequences for gas exchange as blood PO2 and
hypoxia tolerance are reduced in fish with lamellarMRC proliferation. It should be noted, however,that ion-poor conditions are not always associatedwith the such massive numbers of lamellar MRCas is evident in the neon tetra (Paracheirodoninnesi), which is native to the ion poor waters ofthe Amazon (Wilson et al., ’96).
The estuarine, air-breathing mudskipper Peri-ophthalmodon schlosseri has quite exceptional gilllamellae because the basal layer is composedalmost exclusively of MRCs with a superficiallayer of very electron-dense pavement cells(Wilson et al., ’99). The MRCs communicate tothe outside via very deep apical crypts. Very fewACs could be found and MRC were isolated fromeach other by intervening filament-rich cells. Thelamellae also fuse together at various points, andthe interlamellar space is reduced to a narrowirregular channel that can no longer accommodatea ventilatory flow of water. It has been hypothe-sized that the numerous lamellar MRCs areinvolved in the active NH4
þ elimination observedin these animals (Randall et al., ’99; Wilson et al.,2000b). The apical marginal channels are stillquite thick, and the functional gas exchangesurface of these animals is found on the inside ofthe operculum where intraepithelial capillariesare found. Curiously, these intraepithelial capil-laries can also be found on the edges of thefilament making the gills of this fish quite atypical.
Elasmobranch MRCs
The elasmobranch MRCs are generally foundsingly in the filament epithelium and are cuboidalor ovoid in shape with a basally located nucleus(Laurent ’84; Fig. 11). In addition to their highmitochondrial densities, they have a basolateralmembrane amplified through heavy folding into abasal labyrinth and a densely packed sub-apicaltubulovesicular system. The tight junctions be-tween MRCs and neighboring PVCs consist ofmany strands and are not considered leaky. Theapical surfaces of elasmobranch MRCs range frombeing deeply invaginated to convexed and arecovered by microvilli of varying densities (Crespo,’82; Wilson et al., 2002).
Although the salt-secreting rectal gland replacesthe gill as the dominant salt secretory organ,branchial MRCs are still present in the gills insignificant numbers. Also the important ionoregu-latory enzyme Naþ,Kþ-ATPase is associated withthese cells (Piermarini and Evans, 2001; Wilsonet al., 2002); however, leaky tight junctions
J.M. WILSON AND P. LAURENT206

are absent, suggesting an alternative functionto NaCl elimination. Piermarini and Evans(2001) suggest roles in acid-base regulation (seeClaiborne, 2002, this issue).
Agnatha MRCs
The agnathans or jawless fishes represent themost primitive group of fishes and are representedby the osmoconforming marine hagfish (Myxine,Eptatretus) and hagslime (Bdellostoma) and theosmoregulating, anadromous lamprey (Lampretra,Petromyzon, Geotria). These differences in osmor-egulatory abilities are reflected in the differencesin MRC morphology in the agnathans (Morris andPickering, ’76; Youson and Freeman, ’76; Bartelset al., ’98). MRCs are found in both freshwater andseawater lampreys as well as in the marinehagfish. The morphological properties of theMRCs differ within these groups. Freshwaterlamprey have three different types of MRC(Bartels et al., ’98), one of which is associatedwith the preadaptation to sea water and is the onlytype found in the seawater-adapted lamprey. Theseawater type is commonly referred to as thechloride cell due to its morphological similaritywith the teleost chloride cell. In seaward-migrat-ing freshwater lamprey, the seawater-type (SW)MRCs are found in a continuous row along thelength of the interlamellar space of the filamentepithelium (Bartels et al., ’96). The swMRCs are
disc shaped and covered over by neighboring PVCswith the apically exposed surface containingmicrovilli limited to a diameter of 3–4 mm. Follow-ing acclimation to sea water, the overlying PVCsretract and the apical surface increases to extendthe full width of the interlamellar space. Thesurface bulges slightly and is relatively smoothwith few microvilli. The number of strands in thetight junction between neighboring seawaterMRCs or CC also decreases following seawatertransfer (Bartels and Potter, ’93). The shallowtight junction is an important morphologicalcharacteristic that gives a good indication of thecell’s function, namely, NaCl elimination.
In the lamprey, the freshwater MRC types aresometimes referred to as intercalated MRCs andone of the noticeable differences to the swMRC isthe lack of a tubular system (Bartels et al., ’98).One type is only present in the ammocoete and ischaracterized by electron-dense mitochondria,small secretory mucus granules, short microvilli,and/or microplicae and the presence of globularparticles in its apical membrane (as shown byfreeze-fracture). It is also found in groups in boththe lamellar and filament epithelia. The functionof this MRC type is not known, and it disappearsfollowing metamorphosis. The second type isgenerally found singly or in pairs in the inter-lamellar region and at the base of the lamellae,where it intercalates with neighboring pavementcells and/or other MRC types. It is characterizedby an apical membrane with elaborate micropli-cae, the absence of secretory mucous granules, thepresence of numerous small vesicles and membra-nous tubules, and the presence of rod-shapedparticles in either the apical or lateral membranes(as determined by freeze-fracture electron micro-scopy; Bartels et al., ’98). These rod-shapedparticles are similar to those described in themitochondria-rich intercalated cells of the mam-malian kidney collecting duct and toad and turtlebladders (Stetson and Steinmetz, ’85; Brown et al.,’87). The plasma membrane domains having denserod-shaped particle aggregates also show strongimmunoreactivity for the proton pump (vacuolartype Hþ-ATPase) (Brown et al., ’87). When theproton pumps are in the apical locations, thesecells function in acid excretion (A cells), while incells in which the particles have a basolaterallocation (B cells), the function is for base excre-tion. It would thus seem, on the basis of homologywith the amphibian and mammalian epithelia,that the hagfish and lamprey also have a protonpump in their MRCs. A direct demonstration
Fig. 11. TEM of the MRC from the dogfish (S. acanthias).Note the peripheral location of the mitochondria and theextensive basolateral folding (asterisks), and the densecollection of vesicles and tubules of the subapical tubulovesi-cular system (tvs). Arrows indicate the deep tight junctionsjoining the MRC to neighboring pavement cells. Scalebar¼5 mm (J.M. Wilson, unpublished data).
FISH GILL MORPHOLOGY 207

using immunohistochemical techniques has yet tobe made in the lamprey.In the hagfish, MRCs are restricted to the
lateral wall of gill pouches and the lateral half ofgill folds. All MRCs occur singly, being separatedby PVCs, and lack leaky tight junctions (Bartels,’88). The hagfish MRCs have abundant, largemitochondria and a tubular system continuouswith the basolateral membrane along with asubapical vesiculotubular system (Bartels, ’88).These cells also have perinuclear glycogen andhigh levels of carbonic anhydrase and Naþ,Kþ-ATPase (Conley and Mallatt, ’87; Choe et al., ’99).Communicating gap junctions are not foundbetween hagfish MRCs and its neighbors (PVCs,non-differentiated cells) (Bartels and Potter, ’90).Their apical membrane contains microvilli and isusually convex, and deep apical crypts are notcommon. Freeze-fracture electron microscopy hasshown the presence of rod-shaped particles in theapical membrane (Bartels, ’88), which may repre-sent proton pumps as in the lamprey. Sincehagfish are osmoconformers and therefore lackactive NaCl regulation, the presence of an apicalproton pump would suggest the involvement oftheir MRCs in acid–base regulation (see Claiborne,2002, this issue).
DETECTION OF MRCs
In addition to identifying MRC on the basis oftheir fine structure using electron microscopy, anumber of other techniques have been used forlocalization and quantification of this cell type.Keys and Willmer’s (’32) chloride-secreting cells
were identified on the basis of their greateraffinity for eosin, ovoid shape, and granularcytoplasm. Since then, some more specific histolo-gical stains have been used with success to identifyand enumerate MRCs. The acid Fuschin stain wasused by Shirai and Utida (’70) to identify MRCs ineel. The use of Champy-Maillet’s fixative (ZnI-OsO4) originally applied to fish MRCs by Garcia-Romeu and Masoni (’70) has been used by anumber of other investigators to quantify andlocalize MRCs (Avella et al., ’87; Madsen, ’90;Watrin and Mayer-Gostan, ’96; Wilson et al.,2002). This stain reacts specifically with mem-branes, and the reaction with the abundantmembrane systems of the MRC results in itsselective blackening (Fig. 12).Some techniques are directed at the detection of
the mitochondria. With higher numbers of mito-chondria associated with these cells, they give a
stronger signal than background. The most com-mon detection methods used are the mitochon-drial vital stains DASPMI (dimethylaminostyryl-methylpyridinium iodide, emission wavelength609 nm) and DASPEI (dimethylaminostyrylethyl-pyridinium iodide, emission wavelength 530 nm)(e.g., Karnaky et al. ’84; Li et al., ’95; Witters et al.,’96; van der Heijden et al., ’97; Rombough, ’99),which accumulate in active mitochondria. Thislimits their use to fresh, unfixed tissues. Thereare, however, newer products on the market thatare retained in the mitochondria after fixation andare being used on fish MRCs (Mitotracker,Molecular Probes; Galvez et al., 2002; Marshallet al., 2002). There have also been a few studiesusing immunohistochemical techniques to detectmitochondria using antibodies directed againstmitochondria specific proteins (HSP 60, Mickleet al., 2000; Ab-AMA, Sturla et al., 2001).
Another approach has been to target theenzyme Naþ,Kþ-ATPase, which has been shownto be expressed in high concentrations in MRCs.Ouabain specifically binds and inhibits theNaþ,Kþ-ATPase and both [3H]ouabain autoradio-graphy and anthroylouabain fluorescence havebeen found to localize to MRCs (Karnaky et al.,’76; McCormick, ’90). Histochemical methods have
Fig. 12. Localization of MRCs in the gills of a coho salmon(Oncorhynchus kisutch) using the Champy-Malliet solutionoriginally used in fish by Garcia-Romeu and Masoni (’70). Thetechnique preferentially stains the cells because of their ampleintracellular membrane systems (tubulovesicular and tubularsystems; see Figs. 8 and 9). Asterisks indicate some positivelylabeled cells in the filament and lamellae. Note the similarityin labelling with the Naþ,Kþ-ATPase immunohistochemicaltechnique (Fig. 13). Scale bar¼50 mm (J.M. Wilson, unpub-lished data).
J.M. WILSON AND P. LAURENT208

been used in the past to localize Naþ,Kþ-ATPaseto MRCs (de Renzis and Bornancin, ’84; Conleyand Mallatt, ’87). The use of antibodies directedagainst the Naþ,Kþ-ATPase has proven veryuseful, as is evident in the large number of studiespublished recently using this approach (Fig. 13).Practically all of these studies make use of eitherthe mouse monoclonal or rabbit polyclonal anti-bodies directed against conserved epitopes orregions of the Naþ,Kþ-ATPase a subunit originallyemployed by Witters et al. (’96) and Ura et al.(’96), respectively. In addition to the teleosts, themouse monoclonal has been used on a wide range
of fishes with positive results (Atlantic hagfishMyxine, Choe et al., ’99; lamprey, J.M. Wilson,unpublished; Atlantic sting ray, Piermarini andEvans, 2000, 2001; dogfish, Wilson et al., 2002).Lee et al. (’98) have also used other antibodiesagainst specific a-subunit isoforms that have beenfound to localize to MRCs in the tilapia (O.mossambicus) while Hwang et al. (’98) have alsodeveloped a specific tilapia Naþ,Kþ-ATPase a-subunit antibody.
Localization studies employing double-labelingtechniques have also been used that demonstratewhether the apical surface of the MRC is exposedas covered MRCs are presumably not active in iontransport. To this end Li et al. (’95), van derHeijden et al. (’97), and Lee et al. (2000) have usedfluorochrome-conjugated concanavalin A (Con-A),a lectin that binds specifically to a-mannopyrano-syl and a-glucopyranosyl glycoprotein residues, tolocalize the apical region of MRCs identified witheither a mitochondrial stain or Naþ,Kþ-ATPasemarker. PNA (peanut agglutinin binds specificallyto terminal b-galactose residues) and WGA (wheatgerm agglutinin binds specifically to N-acetylglu-cosamine and N-acetylneuraminic acid (sialic acid)residues) have been used by Galvez et al. (2002)and Tsai and Hwang (’98), respectively, to identifyspecific subpopulation of MRCs.
In hagfish, the PAS technique has also beenused to identify MRC through the reaction withthe abundant glycogen associated with the peri-nuclear region. EM studies have shown that thehagfish gill does not have goblet type mucous cellsprecluding the possibility that the PAS reaction isassociated with the neutral mucins found in somemucous cells (Mallatt and Paulsen, ’86).
OTHER EPITHELIAL CELL TYPES
In addition to the main cell types describedabove are lesser numbers of eosinophilic granulecells (EGC) and rodlet cells. The eosinophilicgranule cells may be homologous to mammalianmast cells (Holland and Rowley, ’98). The granulesare PAS negative and some show immunoreactiv-ity with monoclonal antibody 21G6, which hasbeen shown to be reactive with trout leukocytesand intestinal EGC. The rodlet cells, which have awell-defined capsule, and intracellular rodlets arelikely parasitic in origin (Barber et al., ’79). Themudskipper (P.schlosseri) possesses a uniquefilament rich cell (Wilson et al., ’99), which hasalso been found in the inner opercular epitheliumof the killifish (F. heteroclitus; Lacy, ’83). In the
Fig. 13. (a) Immunohistochemical localization of Naþ,Kþ-ATPase a-subunit in rainbow trout (O. mykiss) gill using themouse monoclonal a5 antibody in conjunction with a goat anti-mouse conjugated Alexa 488 (Molecular Probes) secondaryantibody. Arrows indicate some immunopositive cells in boththe filament and along the lamellae. Note the similarities inlabeling with the coho salmon (Fig. 12). (b) Correspondingphase-contrast image showing the tissue. Scale bar¼25 mm(J.M. Wilson, unpublished data).
FISH GILL MORPHOLOGY 209

mudskipper, these cells are laterally compressedbetween neighboring MRCs in the lamellarepithelium and appear to confer structural sup-port to the epithelium. The septum epithelium ofthe elasmobranch Scyliorhinus canicula containsthe usual complement of cells (PVCs, MRCs,undifferentiated cells, mucocytes) as well as largecells (10–30 mm in diameter) that contain a largeapical vacuole (8–27 mm in diameter) (Wright, ’73).The vacuoles are usually electron lucent whilesome show moderate electron density.
PROSPECTIVE STUDIES
The morphological approach will continue to bean integral part of the understanding and inter-pretation of the physiological functions of the gill(gas exchange, ionoregulation, acid–base regula-tion, detoxification). There will continue to beboth quantitative and qualitative morphologicaldata collected in studies of undescribed species aswell as changes during development and inresponse to endogenous and exogenous factors(i.e., hormones and changes in environment,respectively). Importantly, more direct indicatorsof branchial cell function will also be studied bylooking directly at the expression of the proteinsthat facilitate these various physiological pro-cesses. Detection of ion-transport proteins, usingimmunohistochemical techniques, has yieldedsome insight into the roles of both pavement cellsand mitochondria rich cells in ion regulation infreshwater fishes (see Marshall, 2002, this issue).Our understanding of the roles of these cells infreshwater ion regulation has been handicappedby the lack of a suitable surrogate model to applyelectrophysiological techniques which have pro-vided insightful in the study of the seawater fishgill (see Marshall, 2002). Most immunohistochem-ical studies to date have relied on nonhomologousantibodies and thus suffer from complications ofspecies cross reactive, but in the future fishspecific homologous antibodies will likely featuremore prominently. The cloning of fish ion trans-port proteins will facilitate the design and produc-tion of these antibodies. It is also now possible todetect low levels of mRNA expression in tissuesections using in situ PCR. Uncertainty surround-ing the functional role of MRCs in the agnathans,elasmobranchs, and freshwater fishes, the pre-sence of gap junctions in teleosts fishes, and therole of accessory MRCs will likely be addressedthrough the use of these techniques.
LITERATURE CITED
Avella M, Masoni A, Bornancin M, Mayer-Gostan N. 1987. Gillmorphology and sodium influx in the rainbow trout (Salmogairdneri) acclimated to artificial freshwater environments.J Exp Zool 241:159–169.
Bailly Y, Dunel-Erb S, Laurent P. 1992. The neuroepithelialcells of the fish gill filament indolamine immunocytochem-istry and innervation. Anat Rec 233:143–161.
Barber DL, Mills W, Jensen DN. 1979. New observations onthe rodlet cell (Rhabdospora thelohani) in the white suckerCatostomus commersoni (Lacepede): LM and EM studies.J Fish Biol 14:277–284.
Bartels H. 1985. Assemblies of linear arrays of particles in theapical plasma membrane of mitochondria-rich cells in thegill epithelium of the Atlantic hagfish (Myxine glutinosa).Cell Tissue Res 238:229–238.
Bartels H. 1988. Intercellular junctions in the gill epitheliumof the Atlantic hagfish, Myxine glutinosa. Cell Tissue Res254:573–583.
Bartels H, Decker B. 1985. Communicating junctions betweenpillar cells in the gills of the Atlantic hagfish, Myxineglutinosa. Experimentia 41:1039–1040.
Bartels H, Potter IC. 1990. Communicating (gap) junctionsbetween chloride cells in the gill epithelium of the lamprey,Geotria australis. Cell Tissue Res 259:393–395.
Bartels H, Potter IC. 1993. Intercellular junctions in thewater–blood barrier of the gill lamellae in the adult lamprey(Geotria australis, Lampetra fluviatilis). Cell Tissue Res274:521–532.
Bartels H, Moldenhauer A, Potter IC. 1996. Changesin the apical surface of chloride cells following accli-mation of lampreys to seawater. Am J Physiol 270:R125–R133.
Bartels H, Potter IC, Pirlich K, Mallatt J 1998. Categorizationof the mitochondria-rich cells in the gill epithelium of thefreshwater phases in the life cycle of lampreys. Cell TissueRes 291:337–349.
Bindon SD, Fenwick JC, Perry SF 1994. Branchial chloridecell proliferation in the rainbow trout, Oncorhynchusmykiss: implications for gas transfer. Can J Zool 72:1395–1402.
Bridges CR, Berenbrink M, Muller R, Waser W. 1998.Physiol-ogy and biochemistry of the pseudobranch: an unansweredquestion? Comp Biochem Physiol 119A:67–77.
Brown D, Gluck S, Hartwig J. 1987.Structure of the novelmembrane-coating material in proton-secreting epithelialcells and identification as an Hþ-ATPase. J Cell Biol105:1637–1648.
Burggren WW, Randall DJ. 1978. Oxygen uptake andtransport during hypoxic exposure in the sturgeon, Acipen-ser transmontanus. Resp Physiol 34:171–184.
Chang IC, Lee TH, Yang CH, Wei YY, Chou FI, Hwang P-P.2001. Morphology and function of gill mitochondria-richcells in fish acclimated to different environments. PhysBiochem Zool 74:111–119.
Choe KP, Edwards S, Morrison-Shetlar AI, Toop T, ClaiborneJB. 1999. Immunolocalization of Naþ/Kþ-ATPase in mito-chondria-rich cells of the Atlantic hagfish (Myxine glutinosa)gill. Comp Biochem Physiol 124A:161–168.
Chretien M, Pisam M. 1986. Cell renewal and differentiationin the gill epithelium of fresh- or salt-water-adaptedeuryhaline fish as revealed by [3H]thymidine radioautogra-phy. Biol Cell 56:137–150.
J.M. WILSON AND P. LAURENT210

Conley DM, Mallatt J. 1987. Histochemical localization ofNaþ-Kþ ATPase and carbonic anhydrase activity in gills of17 fish species. Can J Zool 66:2398–2405.
Crespo S, Soriano E, Sampera C, Balasch J. 1981. Zinc andcopper distribution in excretory organs of the dogfishScyliorhinus canicula and chloride cell response followingtreatment with zinc sulphate. Mar Biol 65:117–123.
Daborn K, Cozzi RRF, Marshall WS. 2001. Dynamics ofpavement cell–chloride cell interactions during abruptsalinity change in Fundulus heteroclitus. J Exp Biol204:1889–1899.
de Renzis G, Bornancin M. 1984. Ion transport and gillATPases. In: Hoar WS, Randall DJ, editors. Fish Physiology,Vol XB. New York: Academic Press. p 65–104.
Dunel-Erb S, Chevalier C, Laurent P. 1994. Distribution ofneuroepithelial cells and neurons in the trout gill filament:comparison in spring and winter. Can J Zool 72:1794–1799.
Evans DH, Piermarini PM, Potts WTW. 1999. Ionic transportin the fish gill epithelium. J Exp Zool 283:641–652.
Falck B, Hillarp NA, Thieme G, Torp A. 1962. Fluorescence ofcatecholamines and related compounds condensed withformaldehyde. J Histochem 10:348–354.
Galvez F, Reid SD, Hawkings G, Goss GG. 2002. Isolation andcharacterization of mitochondria-rich cell types from the gillof freshwater rainbow trout. Am J Physiol 282:R658–R668.
Garcia-Romeu F, Masoni A. 1970. Sur la mise en evidence descellules a chlorure de la branchie des poissons. Arch AnatMicrosc 59:289–294.
Goss GG, Laurent P, Perry SF. 1994. Gill morphology duringhypercapnia in brown bullhead (Ictalurus nebulosus): role ofchloride cells and pavement cells in acid–base regulation. JFish Biol 45:705–718.
Graham JB. 1997. Air-breathing fishes. San Diego: AcademicPress. p 299.
Greco AM, Fenwick JC, Perry SF. 1996. The effects of soft-water acclimation on gill structure in the rainbow troutOncorhynchus mykiss. Cell Tissue Res 285:75–82.
Holland JW, Rowley AF. 1998.Studies on the eosinophilicgranule cells in the gills of the rainbow trout, Oncorhynchysmykiss. Comp Biochem Physiol 120C:321–328.
Hootman SR, Philpott CW. 1980. Accessory cells in the teleostbranchial epithelium. Am J Physiol 238:R199–R206.
Hughes GM, Weibel ER. 1972. Similarity of supporting tissuein fish gills and the mammalian reticuloendothelium. JUltrastruct Res 39:106–114.
Hughes GM. 1984. General anatomy of the gills. In: Hoar WS,Randall DJ, editors. Fish physiology, Vol 10A. New York:Academic Press. p 1–72.
Hwang P-P, Fang MJ, Tsai J-C, Huang CJ, Chen ST. 1998.Expression of mRNA and protein of Naþ-Kþ-ATPase asubunit in gills of tilapia (Oreochromis mossambicus). FishPhysiol Biochem 18:363–373.
Jurss K, Bastrop R. 1995. The function of mitochondria-richcells (chloride cells) in teleost gills. Rev Fish Biol Fish 5:235–255.
Karnaky KJ, Degnan KJ, Garretson LT, Zadunaisky JA. 1984.Identification and quantification of mitochondria-rich cellsin transporting epithelia. Am J Physiol 246:R770–R775.
Karnaky KJ, Kinter LB, Kinter WB, Sterling CE. 1976.Teleost chloride cell. II. Autoradiographic localization ofgill Na,K-ATPase in killifish Fundulus heteroclitus adaptedto low and high salinity environments. J Cell Biol 70:157–177.
Karnovsky MJ. 1965. A formaldehyde–glutaraldehyde fixativeof high osmolality for use in electron microscopy. J Cell Biol27:137A–138A.
Kawahara T, Sasaki T, Higashi S. 1982. Intercellular junctionsin chloride and pavement cells of Oplegnethus fasciatus. JElectron Microsc 31:162–170.
Keys A, Willmer EN. 1932. ‘‘Chloride secreting cells’’ in thegills of fishes, with special reference to the common eel. JPhysiol 76:368–378.
Lacy ER. 1983. Histochemical and biochemical studies ofcarbonic anhydrase activity in the opercular epithelium ofthe euryhaline teleost, Fundulus heteroclitus. Am J Anat166:19–39.
Laurent P. 1984. Gill internal morphology. In: Hoar WS,Randall DJ, editors. Fish physiology, Vol Vol 10A. NewYork: Academic Press. p 73–183.
Laurent P. 1989. Gill structure and function. In: Wood SC,editor. Comparative pulmonary physiology. New York:Marcel Dekker. p 69–120.
Laurent P, Dunel-Erb S. 1984. The pseudobranch: morphol-ogy and function. In: Hoar WS, Randall DJ, editors. Fishphysiology, Vol 10B. New York: Academic Press. p 285–323.
Laurent P, Hebibi N. 1989. Gill morphology and fishosmoregulation. Can J Zool 67:3055–3063.
Laurent P, Perry SF. 1990.Effects of cortisol on gill chloridecell morphology and ionic uptake in the freshwater trout,Salmo gairdneri. Cell Tissue Res 259:429–442.
Laurent P, Perry SF. 1995. Morphological basis of acid–baseand ion regulation in fish. In: Heisler N, editor. Advances incomparative and environmental physiology. Mechanisms ofsystemic regulation: acid–base regulation, ion transfer andmetabolism. Heidelberg: Springer-Verlag. p 91–118.
Laurent P, Dunel-Erb S, Chevalier C, Lignon J. 1994a. Gillepithelial cells kinetics in a freshwater teleost, Oncor-hynchus mykiss during adaptation to ion-poor water andhormonal treatments. Fish Physiol Biochem 13:353–370.
Laurent P, Goss GG, Perry SF. 1994b. Proton pumps in fishgill pavement cells? Arch Int Physiol Biochim Biophys102:77–79.
Laurent P, Hobe H, Dunel-Erb S. 1985. The role ofenvironmental sodium chloride relative to calcium in gillmorphology of freshwater salmonid fish. Cell Tissue Res240:675–692.
Laurent P, Maina JN, Bergman HL, Narahara A, Walsh PJ,Wood CM. 1995. Gill structure of a fish from an alkalinelake: effect of short-term exposure to neutral conditions.Can J Zool 73:1170–1181.
Laurent P, Wilkie MP, Chevalier C, Wood CM. 2000. Theeffect of highly alkaline water (pH 9.5) on the morphologyand morphometry of chloride cells and pavement cells in thegills of the freshwater rainbow trout: relationship to ionictransport and ammonia excretion. Can J Zool 78:307–319.
Lee TH, Hwang PP, Lin HC, Huang FL. 1996. Mitochondria-rich cells in the branchial epithelium of the teleosts,Oreochromis mossambicus, acclimated to various hypotonicenvironments. Fish Physiol Biochem 15:513–523
Lee TH, Hwang P-P, Shieh YE, Lin CH. 2000. The relation-ship between ‘‘deep-hole’’ mitochondria-rich cells andsalinity adaptation in the euryhaline teleost, Oreochromismossambicus. Fish Physiol Biochem 23:133–140.
Lee TH, Tsai JC, Fang MJ, Yu MJ, Hwang PP. 1998. Isoformexpression of Naþ-Kþ-ATPase a-subunit in gills of theteleost Oreochromis mossambicus. Am J Physiol 275:R926–R932.
FISH GILL MORPHOLOGY 211

Leino RL, McCormick JH. 1984. Morphological and morpho-metrical changes in chloride cells of the gills of Pimephalespromelas after chronic exposure to acid water. Cell TissueRes 236:121–128.
Li J, Eygensteyn J, Lock RA, Verbost PM, van der HeijdenAJH, Wendelaar Bonga SE, Flik G. 1995. Branchial chloridecells in larvae and juveniles of freshwater tilapia Oreochro-mis mossambicus. J Exp Biol 198:2177–2184.
Madsen SS. 1990. Enhanced hypoosmoregulatory response togrowth hormone after cortisol treatment in immaturerainbow trout, Salmo gairdneri. Fish Physiol Biochem8:271–279.
Mallatt J, Paulsen C. 1986. Gill ultrastructure of the Pacifichagfish Eptatretus stouti. Am J Anat 177:243–269.
Marshall WS, Lynch EM, Cozzi RRF. 2002. Redistribution ofimmunofluorescence of CFTR anion channel and NKCCco-transporter in chloride cells during adaptation of thekillifish Fundulus heteroclitus to sea water. J Exp Biol205:1265–1273.
McCormick SD. 1990. Fluorescent labelling of Naþ,Kþ-ATPase in intact cells by use of a fluorescent derivative ofouabain: salinity and chloride cells. Cell Tissue Res 260:529–533.
Mickle J, Karnaky KJ, Jensen T, Miller DS, Terlouw S, GrossA, Corrigan B, Riordan JR, Cutting GR. 2000. Processingand localization of the cystic fibrosis transmembraneregulator in gill and operculum from Fundulus heteroclitus.Bull Mt Desert Isl Biol Lab 39:75–77.
Morris R, Pickering AD. 1976. Changes in the ultrastructureof the gills of the river lamprey, Lampetra fluviatilus (L.),during the anadromous spawning migration. Cell Tissue Res173:271–277.
Newstead JD. 1967. Fine structure of the respiratory lamellaeof teleostean gills. Z Zellforsch 79:396–428.
Olson KR. 1996. Scanning electron microscopy of the fish gill.In: Munshi JSD, Dutta HM, editors. Fish morphology.Horizon of new research. Rotterdam: AA Balkema.p 32–45.
Perry SF. 1997. The chloride cell: structure and function inthe gills of freshwater fishes. Annu Rev Physiol 59:325–347.
Perry SF. 1998. Relationships between branchial chloride cellsand gas transfer in freshwater fish. Comp Biochem Physiol119A:9–16.
Perry SF, Laurent P. 1989. Adaptational responses of rainbowtrout to lowered external NaCl concentration: contributionof the branchial chloride cell. J Exp Biol 147:147–168.
Philpott CW. 1980. Tubular system membranes of teleostchloride cells: osmotic response and transport sites. Am JPhysiol 238:R171–R184.
Piermarini PM, Evans DH. 2000. Effects of environmentalsalinity of Naþ/Kþ-ATPase in the gills and rectal gland of aeuryhaline elasmobranch (Dasyatis sabina). J Exp Biol203:2957–2966.
Piermarini PM, Evans DH. 2001. Immunochemical analysisof the vacuolar proton-ATPase b-subunit in the gillsof a eurhyaline stingray (Dasyatis sabina): effects ofsalinity and relation to Naþ/Kþ-ATPase. J Exp Biol204:3251–3259.
Pisam M. 1981. Membranous systems in the ‘‘chloride cell’’ ofteleostean fish gill; their modifications in response to thesalinity of the environment. Anat Rec 200:401–414.
Pisam M, Rambourg A. 1991. Mitochondria-rich cells in thegill epithelium of teleost fishes: an ultrastructural approach.Int Rev Cytol 130:191–232.
Pisam M, Boeuf G, Prunet P, Rambourg A. 1990. Ultra-structural features of mitochondria-rich cells in stenohalinefreshwater and seawater fishes. Am J Anat 187:21–31.
Pisam M, Caroff A, Rambourg A. 1987. Two types of chloridecells in the gill epithelium of a freshwater-adapted euryha-line fish, Lebistes reticulatus; their modifications duringadaptation to seawater. Am J Anat 179:40–50.
Pisam M, LeMoal C, Auperin B, Prunet P, Rambourg A. 1995.Apical structures of ‘‘mitochondria-rich’’ a and b cells ineuryhaline fish gill: their behaviour in various livingconditions. Anat Rec 241:13–24.
Pisam M, Prunet P, Boeuf G, Rambourg A. 1988. Ultra-structural features of chloride cells in the gill epithelium ofthe Atlantic salmon, Salmo salar, and their modificationsduring smoltification. Am J Anat 183:235–244.
Pisam M, Prunet P, Rambourg A. 1989. Accessory cells in thegill epithelium of freshwater rainbow trout Salmo gairdneri.Am J Anat 184:311–320.
Randall DJ, Wilson JM, Peng KW, Kok WK, Kuah SSL, ChewSF, Lam TJ, Ip YK. 1999. The mudskipper, Periophthalmo-don schloesseri, actively transports NH4
þ against a concen-tration gradient. Am J Physiol 277:R1562–R1567.
Randall DJ. 1972. Respiration. In: Hardistry MW, Potter IC,editors. The biology of lampreys, Vol II. London: AcademicPress. p 287–306.
Rojo MC, Blanquez MJ, Gonzalez ME. 1996. A histochemicalstudy of the distribution of lectin binding sites in thedeveloping branchial area of the trout Salmo trutta. J Anat189:609–621.
Rombough PJ. 1999. The gill of fish larvae. Is it primarily arespiratory or an ionoregulatory structure? J Fish Biol55:186–204.
Romer AS, Parsons TS. 1986. The vertebrate body. Philadel-phia: Saunders College Publishing. P 679.
Roy PK, Munshi JSD. 1996. Morphometrics of the respiratorysystem of air-breathing fishes of India. In: Munshi JSD,Dutta HM, editors. Fish morphology. Horizon of newresearch. Rotterdam: AA Balkema. p 203–234.
Saboia SMT, Hernandez-Blazquez FJ, Mota DL, BittencourtAM. 1996. Mucous cell types in the branchial epitheliumof the euryhaline fish Poecilia vivipara. J Fish Biol 49:545–548.
Sandbacka M, Lilius H, Enkvist MOK, Isomaa B. 1998.Rainbow trout gill epithelial cells in primary culturecommunicate through gap junctions as demonstrated bydye-coupling. Fish Physiol Biochem 19:287–292.
Sardet C, Pisam M, Maetz J. 1979. The surface epithelium ofteleostean fish gills. Cellular and junctional adaptations ofthe chloride cell in relation to salt adaptation. J Cell Biol80:96–117.
Sardet C. 1977. Ordered arrays of intramembrane particles onthe surface of fish gills. Cell Biol Int Rep 1:409–418.
Shikano T, Fujio Y. 1998. Immunolocalization of Naþ,Kþ-ATPase and morphological changes in two types of chloridecells in the gill epithelium during seawater and freshwateradaptation in a euryhaline teleost, Poecilia reticulata. J ExpZool 281:80–89.
Shirai N, Utida S. 1970. Development and degeneration of thechloride cell during seawater and freshwater adaptation ofthe Japanese eel Anguilla japonica. Z Zellforsch 103:247–264.
Stetson DL, Steinmetz PR. 1985. a and b types of carbonicanhydrase-rich cells in turtle bladder. Am J Physiol249:F553–F565.
J.M. WILSON AND P. LAURENT212

Sturla M, Masini MA, Prato P, Grattarola C, Uva B. 2001.Mitochondria-rich cells in gills and skin of an Africanlungfish, Protopterus annectens. Cell Tissue Res 303:351–358.
Tsai J-C, Hwang P-P. 1998. Effects of wheat germ agglutininand colchicine on microtubules of the mitochondria-richcells and Ca2þ uptake in tilapia (Oreochromis mossambicus)larvae. J Exp Biol 201:2263–2271.
Uchida K, Kaneko T, Miyazaki H, Hasegawa S, Hirano T.2000. Excellent salinity tolerance of Mozambique tilapia(Oreochromis mossambicus): elevated chloride cellactivity in the branchial and opercular epithelia ofthe fish adapted to concentrated seawater. Zool Sci 17:149–160.
Ura K, Soyano K, Omoto N, Adachi S, Yamauchi K. 1996.Localization of Na-K-ATPase in tissues of rabbit andteleosts using an antiserum directed against a partialsequence of the a-subunit. Zool Sci 13:219–227.
van der Heijden AJH, van der Meij CJM, Flik G, WendelaarBonga SE. 1999. Ultrastructure and distribution dynamicsof chloride cells in tilapia larvae in fresh water and seawater. Cell Tissue Res 297:119–130.
van der Heijden AJH, Verbost PM, Eygensteyn J, Li J,Wendelaar Bonga SE, Flik G. 1997. Mitochondria-rich cellsin gills of tilapia (Oreochromis mossambicus) adapted tofresh water or sea water: Quantification by confocal laserscanning microscopy. J Exp Biol 200:55–64.
Watrin A, Mayer-Gostan N. 1996. Simultaneous recognition ofionocytes and mucous cells in the gill epithelium of turbotand in the rat stomach. J Exp Zool 276:95–101.
Wendelaar Bonga SE, van der Meij CJM. 1989. Degenerationand death, by apoptosis and necrosis, of the pavement and
chloride cells in the gills of the teleost Oreochromismossambicus. Cell Tissue Res 255:235–243.
Wilson JM, Kok WK, Randall DJ, Vogl AW, Ip YK. 1999. Finestructure of the gill epithelium of the terrestrial mudskip-per, Periophthalmodon schlosseri. Cell Tissue Res 298:345–356.
Wilson JM, Laurent P, Tufts BL, Benos DJ, Donowitz M, VoglAW, Randall DJ. 2000. NaCl uptake by the branchialepithelium in freshwater teleost fish: an immunologicalapproach to ion-transport protein localization. J Exp Biol203:2279–2296.
Wilson JM, Randall DJ, Donowitz M, Vogl AW, Ip YK. 2000.Immunolocalization of ion-transport proteins to branchialepithelium mitochondria-rich cells in the mudskipper(Periophthalmodon schlosseri). J Exp Biol 203:2297–2310.
Wilson JM, Vogl AW, Randall DJ. 1996. Gill morphology of theneon tetra, Paracheirodon innesi. In: Val AL, Randall DJ,MacKinlay D, editors. Physiology of tropical fishes. Alphar-etta, GA: American Fisheries Society. p 123– 128.
Wilson JM, Vogl AW, Randall DJ. 2002. Branchial mitochon-dria-rich cells in the dogfish (Squalus acanthias). CompBiochem Physiol A 132:365–374.
Witters H, Berckmans P, Vangenechten C. 1996. Immunolo-calization of Naþ,Kþ-ATPase in the gill epithelium ofrainbow trout, Oncorhynchus mykiss. Cell Tissue Res283:461–468.
Wright DE. 1973. The structure of the gills of the elasmo-branch, Scyliorhinus canicula (L.). Z Zellforsch 144:489–509.
Youson JH, Freeman PA. 1976. Morphology of the gills oflarval and parasitic adult sea lamprey, Petromyzon marinusL. J Morphol 149:73–104.
FISH GILL MORPHOLOGY 213





























