Embed Size (px)
Citation preview

Harmful Algae 24 (2013) 10–19
First development and characterisation of polyclonal and monoclonal antibodiesto the emerging fresh water toxin cylindrospermopsin
Christopher T. Elliott a,*, Clare H. Redshaw b,c, Suja E. George a, Katrina Campbell a
a Institute for Global Food Security, School of Biological Sciences, Queen’s University, Belfast BT9 5AG, UKb European Centre for Environment and Human Health, University of Exeter Medical School, Royal Cornwall Hospital, Truro, Cornwall TR1 3HD, UKc School of Geography, Earth and Environmental Science, University of Plymouth, Drake Circus, Plymouth, Devon PL4 8AA, UK
A R T I C L E I N F O
Article history:
Received 10 July 2012
Received in revised form 30 December 2012
Accepted 30 December 2012
Keywords:
Cylindrospermopsin
Fresh water toxin
Antibody
Immunoassay
A B S T R A C T
As increasing incidences in the occurrence of cylindrospermopsin (CYN) appear, in addition to further
research on its toxicological nature, improved rapid methods to detect this toxin are required. Antibody
based assays are renowned for their ability to provide rapid, portable, simple to use tests. As yet however
there are no publications outlining how an antibody to CYN can be produced. A range of chemical
approaches was investigated to synthesise CYN immunogens for antibody production but failed to
generate a response. Finally, a modified Mannich reaction for immunogen synthesis was employed to
couple the toxin to two carrier proteins. Both protein conjugates were successfully used to raise both
polyclonal and monoclonal antibodies of high sensitivity to CYN. These antibodies were characterised
employing competitive indirect ELISA and an optical biosensor assay. By ELISA the sensitivity achieved
ranged from 27 to 131 pg/mL and by SPR 4.4 to 11.1 ng/mL thus demonstrating that the selection of
immunoassay platform is important for the detection level required by the end user for their application.
Low cross-reactivity to the much less toxic metabolite deoxyCYN was observed. This is the first reported
production of antibodies to this toxin.
� 2013 Elsevier B.V. All rights reserved.
Contents lists available at SciVerse ScienceDirect
Harmful Algae
jo u rn al h om epag e: ww w.els evier .c o m/lo cat e/ha l
1. Introduction
The appearance of a number of previously unreported marinebiotoxins, such as tetrodotoxin, palytoxin and cyclic imines, withinEuropean waters, has led to concerns regarding risks to humanhealth (Paredes et al., 2011), and is evidenced by the formation of anumber of European Food Safety Authority opinion reports onthese emerging toxins (EFSA, 2008, 2009, 2010). Similar emergenceof cyanobacterial produced toxins has also been observed intemperate freshwater systems with the appearance of anatoxin,and their analogues (Cadel-Six et al., 2007; Codd et al., 2005; Rawnet al., 2007). This has resulted in concerns over both human healthand environmental consequences due to contamination ofdrinking water, food stuffs and bioaccumulation in exposedaquatic organisms (Kinnear, 2010). The presence of these emergingtoxins and the increased occurrence of blooms of establishedcyanobacteria (such as the microcystin producer Microcystis spp.)is largely attributed to increased water temperature due to climatechange and the degradation of water quality; particularlyeutrophication due to high agricultural fertiliser usage (Hudnellet al., 2008; Paerl and Huisman, 2008).
* Corresponding author.
E-mail address: [email protected] (C.T. Elliott).
1568-9883/$ – see front matter � 2013 Elsevier B.V. All rights reserved.
http://dx.doi.org/10.1016/j.hal.2012.12.005
The best known cyanobacteria responsible for producingcylindrospermopsin (CYN) is Cylindrospermopsis raciborskii, al-though in recent years a number of other genera includingAphanizomenon, Anabaena, Raphidiopsis, Umezakia and Lyngbia
have emerged as CYN producers in a number of geographicallocations across Europe, the Americas, E and SE Asia, West Africa,the Middle East and Australasia (Banker et al., 2000; Bernard et al.,2003; Berry and Lind, 2010; Blahova et al., 2009; Brient et al., 2009;Carson, 2000; Cheng et al., 2009; Li et al., 2001; Messineo et al.,2009; Preussel et al., 2009).
The structure and relative stereochemistry of CYN(C15H21N5O7S; molecular weight 415.43 g/mole; Fig. 1) was firstelucidated by Ohtani and Moore (1992) as a highly water-solublealkaloid with a zwitterionic tricyclic guanidine moiety andsulphate ester functionality, bridged with a 5-substituted-2,4-dioxypyrimidine (uracil) moiety. 7-epi-CYN (Fig. 1), a diasteriomerof CYN, has been identified by Banker et al. (2000) and has a similartoxic potency in mouse bioassays (Banker et al., 2000, 2001;Griffiths and Saker, 2003). The toxicity of the slightly less polardeoxy-CYN (C15H21N5O6S; molecular weight 399; Fig. 1), which isproduced as a major analogue by some cyanobacterial spp., isunder debate as it is non-toxic by mouse bioassay, but is a potentprotein synthesis inhibitor and can inhibit glutathione synthesis(Griffiths and Saker, 2003; Li et al., 2001; Looper et al., 2005; Norriset al., 1999).

A
Cyli ndrospermopsin (CYN), favor ed enol tautomer Cyli ndrospermopsin (CYN), keto tautomer
A
Deoxy-Cyli ndrospermopsin (deoxy-CYN)7-epi-Cyli ndrospermopsin (7-epi-CYN)
A B
C
D
B
B
C
C
D
D
A B
C
D
Fig. 1. Cylindrospermopsin (CYN) and its natural cyanobacterial produced analogues 7-epi-CYN and deoxy-CYN. The positive charge is shared between the 3 nitrogen atoms,
and is not upon the carbon atom as illustrated.
C.T. Elliott et al. / Harmful Algae 24 (2013) 10–19 11
The first documented large scale human CYN poisoning eventoccurred in 1979 in Australia, although the aetiology of this eventwas not elucidated until several years later, thereby earning CYNpoisoning the colloquial name of ‘‘Palm Island Mystery Disease’’(Griffiths and Saker, 2003). High environmental CYN concentra-tions have been reported in farm dams and highly eutrophic waters(up to 800 mg/L; Griffiths and Saker, 2003). Exposure to thesepotent hepatotoxins from submerged summer blooms can haveserious health effects ranging from gastroenteritis to pathologicalimpacts upon the intestine, spleen, thymus, kidneys, liver andheart (Carson, 2000; Falconer et al., 1999; Griffiths and Saker,2003). The primary mechanism of toxicity is by no means fullyunderstood, however evidence suggests inhibition of proteinsynthesis (Carson, 2000; Froscio et al., 2001, 2003; Griffiths andSaker, 2003), glutathione synthesis inhibition (Froscio et al., 2003;Griffiths and Saker, 2003; Norris et al., 2002), involvement ofcytochrome P450 enzymes in CYN activation/metabolite formationand hepatocytotoxicity, (Froscio et al., 2003; Norris et al., 2001,2002), inhibition of pyrimidine nucleotide synthesis (Beyer et al.,2009), DNA strand breakage (Griffiths and Saker, 2003) andalteration of ratios of free cholesterol and phospholipids in redblood cell membranes (Reisner et al., 2004), all have a role to pay inthe toxic effects observed. Due to the toxic effects the tolerabledaily intake for CYN was initially calculated to be 0.02 g/kg bodyweight/day based on a no observable effect level in mice. It wasestimated that guideline values for adults, children, and infantswere 0.48, 0.16, and 0.11 g/L, respectively, based on drinking waterconsumption of 2 L for a 60-kg adult, 1 L for a 10-kg child, and0.75 L for a 5-kg infant (Duy et al., 2000). The guideline values forthe number of cyanobacterial cells (based on 0.026 pg CYN/cell)were 4231 in infants, 6154 in children, and 18,461 in adults (Duyet al., 2000). Concerns over the emergence of this potent toxin haveled a number of countries to introduce drinking water guidelinesfor CYN, which range from 0.3 mg/L (France) to 1 mg/L (Brazil,Australia and New Zealand). Nevertheless, CYN poisoning has beenreported in wild and domestic animals more than any otherorganisms, due to the higher probability of exposure directly fromtoxic cyanobacterial blooms in lakes and ponds. In reports of toxinexposure in cattle the incidence of mortality and morbidity washigher in calves than in adult cattle (Thomas et al., 1998). Thisimplies that toxicity of CYN in livestock may be greater in young
animals but may be a consideration for livestock (e.g. cattle, goats)whereby it could present in milk. CYN accumulation in plants(Kittler et al., 2012), snails (Berry and Lind, 2010) and finfish (Galloet al., 2009) has been reported. With the emergence of cyanotoxins,such as CYN, it is imperative that both qualitative and quantitativeanalytical methodologies are developed for the detection of thesetoxins in a range of matrices, including water and food, to ensurehuman and animal wellbeing.
A number of advanced analytical techniques, e.g. HPLC–MS/MShave been developed for these purposes (Blahova et al., 2009;Eaglesham et al., 1999; Gallo et al., 2009; Nybom et al., 2008;Oehrle et al., 2010; Rawn et al., 2007), however there is a paucity ofrapid, low-cost, high-throughput techniques available, which areessential for monitoring these emerging cyanotoxins. To meet thisneed, monoclonal and polyclonal antibodies were raised againstCYN, which could be utilised in various immunological assayssuch as enzyme-linked immunoassays (ELISAs) and the opticalbiosensing technique of surface-plasmon resonance (SPR). Theaim of this study was to assess different protein conjugationapproaches for the synthesis of immunogens for antibodyproduction and to characterise both antibody types by comparingboth traditional ELISA and advanced surface plasmon resonance(SPR) methodologies.
2. Materials and methods
2.1. Chemicals and reagents
Bovine thyroglobulin (BTG), ovalbumin (OVA), 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide hydrochloride (EDC), N-hydroxysuccinimide (NHS), sodium hypochlorite, succinic anhy-dride, sodium borohydride, butandiol-diglycidyl-ether, methylmorpholine, isobutylchloroformate, glacial acetic acid, aceticanhydride, perchloric acid, potassium carbonate, N-(t-Boc)-2-bromoethylamine (IUPAC: tert-butyl N-(2-bromoethyl)carbamate,ethylenediamine, 2,2-(ethylenedioxy)-bis(ethylamine) (Jeffa-mine), formaldehyde and all other organic solvents used in thestudy (Analar grade) were obtained from Sigma–Aldrich ChemicalCompany (Dorset, UK). The Nunc Maxisorp plates were purchasedfrom Fisher Scientific UK (Leicestershire, UK) and the horseradishperoxidase-linked anti-mouse immunoglobulin from DAKO

C.T. Elliott et al. / Harmful Algae 24 (2013) 10–1912
(Cambridge, UK). TMB/E solution was obtained from MilliporeLimited (Watford, UK). Slide-A-Lyzer dialysis cassettes (10K MWCO,0.5–3 mL Capacity) and Imject Mariculture KLH (keyhole limpethemocyanin) were supplied by Medical Supply Company Ltd.(Dublin, Republic of Ireland). PD10 columns were obtained from GEHealthcare, UK and the immunoassay stabiliser was purchased fromSource BioScience Life Sciences (Nottingham, UK). Freund’s adjuvant(complete and incomplete) was obtained from Sigma–Aldrich(Bornem, Belgium), Quil-ATM was obtained from Brenntag BiosectorA/S (Frederikssund, Denmark) and the adjuvant Pam3Cys-Ser-(Lys)4
(PCSL) was purchased from EMC microcollections (Tubingen,Germany). Horseradish peroxidise (HRP) and mouse monoclonalantibody isotyping strips were obtained from Roche DiagnosticsLimited (West Sussex, UK) and (CYN) from n’Tox (France).
Ethanolamine, HBS–EP buffer (pH 7.4, 0.01 M HEPES, 0.15 MNaCl, 3 mM EDTA, 0.005% polysorbate), and an amine coupling kitwere provided by GE Healthcare Biacore AB (Uppsala, Sweden).
2.2. Preparation of protein conjugates for immunogens
2.2.1. Immunogen 1: CYN–acid–BTG prepared via carbodiimide
reaction
CYN was derivatised to CYN–acid through the chlorinationreaction in a similar manner as described by Banker et al. (2001). Inbrief, CYN (1.5 mg) was dissolved in 100 mL of deionised water.Sodium hypochlorite (1.1 mL of 1 mg/mL) was added and themixture was left stirring in the dark for seven days at roomtemperature. The final mixture was dried and analysed by UPLC–MS (Quattropremier XE) for the presence of the CYN–acid product(see details later).
The crude CYN–acid product was then coupled to BTG via thecarbodiimide reaction with minor modifications to that reportedfor dinitrocarbanilide mimics (Connolly et al., 2002). Briefly, theCYN–acid (�0.75 mg) was added to deionised water and activatedby the addition of 1-ethyl-3-(3-dimethylaminopropyl) carbodii-mide (EDC) (1 mg) and N-hydroxysuccinimide (NHS) (0.5 mg) in 2-(N-morpholino)ethanesulfonic acid (MES) buffer (0.05 M MES,0.5 M NaCl, pH 4.7). The activated CYN–acid was added to BTG(2.5 mg) dissolved in phosphate buffered saline (pH 7.2), and themixture was incubated overnight at room temperature. The BTGconjugate was purified by dialysis against 0.15 M saline solution.
2.2.2. Immunogen 2: CYN–acid to BTG prepared via acid anhydride
reaction
The CYN–acid derivative, prepared as described in Section 2.2.1,was coupled to BTG via an acid anhydride intermediate asdescribed for steroids (Erlanger et al., 1957), monensin (Crookset al., 1997) and chloramphenicol succinate (Fodey et al., 2007).CYN–acid (�0.44 mg) was dissolved in 0.5 mL anhydrous dimethylformamide (DMF). Methyl morpholine (20 mL) was added to themixture which was immediately cooled at �20 8C for 10 min.Isobutylchloroformate (20 mL) was added and the mixture wasstirred for 15 min at room temperature. The activated CYN–acidwas added slowly to BTG (2.8 mg) dissolved in 1 mM sodiumacetate solution and allowed to react for 1 h at 4 8C. The conjugatewas purified by dialysis against 0.15 M saline solution.
2.2.3. Immunogen 3: CYN–succinate–BTG via carbodiimide reaction
CYN was derivatised to CYN succinate in an analogous approachas presented for 30-azido-30-deoxythymidine (Tadayoni et al.,1993). CYN (1 mg) with succinic anhydride (1 mg) dissolved inanhydrous pyridine (1 mL) at room temperature for 48 h stirring inthe dark. After the incubation period, pyridine was removed fromthe final mixture by solvent evaporation. This derivative wasconjugated to BTG (2.5 mg) using the carbodiimide method asdescribed in Section 2.2.1.
2.2.4. Immunogen 4: CYN–BTG via an epoxide linker
CYN–BTG was prepared using the approach of coupling throughan epoxide linker (Hermanson, 1996). Sodium borohydride(10 mg) was dissolved in 5 mL of deionised water that wasadjusted to pH 11 with 0.1 M sodium hydroxide. To this 20 mL ofbutanediol-diglycidyl-ether was added and the epoxide solutionwas degassed with nitrogen. BTG (3 mg) was dissolved in 500 mL ofdeionised water and 180 mL of the epoxide solution was added.The mixture was incubated for 40 h under nitrogen at roomtemperature before purification by PD-10 gel filtration to removeexcess epoxide linker. CYN was dissolved in degassed 0.1 Msodium hydroxide (0.5 mL). The hapten solution was added to theepoxide modified BTG under nitrogen and left to incubate in thedark at room temperature for 40 h. Finally, the BTG conjugate waspurified by dialysis against saline solution.
2.2.5. Immunogens 5 and 6: CYN–KLH and CYN–OVA via modified
Mannich reaction
CYN was coupled to KLH and OVA using a modification of theMannich reaction as described for saxitoxin (Campbell et al., 2007).CYN was added to KLH or OVA (3 mg) dissolved in 500 mL ofphosphate buffer. Formaldehyde (13.5 mL) was added and themixture was stirred in the dark at room temperature. Theconjugate was purified by dialysis in 0.15 M saline solution
2.3. Preparation of CYN–HRP conjugates for competitive direct ELISA
CYN–HRP conjugates were prepared as described for immuno-gens 1–6 as outlined in Section 2.2 to be used for the competitivedirect ELISA format.
2.4. Coating antigen in competitive indirect ELISA
Both CYN–KLH and CYN–OVA protein conjugates (immunogens7 and 8) were also employed for ELISA coating antigens in thecompetitive indirect ELISA format.
2.5. Preparation of CYN biosensor chip surfaces
The carboxymethylated surface of a CM5 certified gradebiosensor chip (Biacore AB) was equilibrated to room temperatureand activated using an amine coupling kit. Briefly, EDC and NHSfrom the kit were mixed (1:1, v/v) and applied to the chip surfacefor 30 min. Excess solution was removed, and Jeffamine (0.1 M) inborate buffer was added to the chip surface and allowed to react for1 h. The chip surface was deactivated by exposure to ethanolamine(1 M) for 30 min followed by immobilisation of using threedifferent surface chemistries.
Chip 1: The carboxylic acid group of CYN-acid was activatedusing EDC/NHS and coupled to the free amine on the Jeffaminelinker.Chip 2: CYN was also activated using the epoxide chemistry inthe same manner as the conjugation to BTG and coupled to anethylenediamine linker used instead of Jeffamine.Chip 3: CYN was activated using formaldehyde chemistry andcoupled to the amine of the Jeffamine linker
Each sensor chip surface was then washed with deionized water,dried using a stream of nitrogen gas, and stored desiccated at 4 8Cwhen not in use.
2.6. Immunisation procedures for antibody production
2.6.1. Polyclonal antibody production
Two rabbits were immunised with each of the CYN immuno-gens (1–6) prepared. Prior to these immunisations serum was

C.T. Elliott et al. / Harmful Algae 24 (2013) 10–19 13
collected from each rabbit to use as a baseline control for titrecomparisons with blood samples taken post immunisations. Foreach rabbit 200 mg of the immunogen preparation (in sterilesaline) was added slowly to an equal volume of Freund’s adjuvantwith constant vortexing to produce an emulsion. For the initialinjection Freund’s complete adjuvant was used while the incom-plete form was used for all subsequent immunisations. Theemulsion was delivered subcutaneously and divided equallybetween four injection sites on the animal. An interval of twoweeks was used between the first three immunisations, whileremaining injections were delivered on a monthly basis. A bloodsample (�5 mL) was collected ten days after each immunisationfor monitoring for the presence of antibodies to CYN.
2.6.2. Monoclonal antibody production
Prior to immunisation sera from a blood sample was collectedfrom each mouse to use as a baseline control for titre comparisonswith blood samples taken post immunisation. For the first twoimmunisations of mice, 40 mg of immunogen was mixed with50 mg of the adjuvant Quil-A in a total volume of 200 mL of sterilesaline. This was administered subcutaneously, equally dividedbetween two injection sites, with an interval of two weeksbetween immunisations. Subsequent immunisations were deliv-ered by the same route on a monthly basis using the same amountof immunogen (40 mg) and 50 mg of the adjuvant Pam3Cys-Ser-(Lys)4 (PCSL). A blood sample (�0.1 mL) was collected ten daysafter each immunisation to determine the presence of antibodiesto CYN. When a sufficiently high antibody titre was observed a finalimmunisation was performed intraperitoneally, without adjuvant.Three days later the mouse was sacrificed and the spleen removedfor fusion with an SP2 murine myeloma cell line to produceantibody producing hybridomas (Galfre and Milstein, 1981); ablood sample from the heart was also obtained to use as a positivecontrol in the protocol for screening the clones for specificantibodies to CYN. All protocols carried out using mice wereperformed in accordance with a licence issued by the Departmentof Health, Social Services and Public Safety in the UK under Theanimals (scientific procedures) Act 1986.
2.7. Screening procedures for determination of antibody titre
Sera samples from all the immunised animals were assessed fortheir antibody content by performing an ELISA chequerboardtitration using both competitive direct and indirect assay formats.Each rabbit serum was tested by direct ELISA using all CYN–HRPconjugates produced. Test-bleed sera was also analysed by SPRusing the biosensor chips produced. In theory this allowed for theassessment of the different chemistries employed with the varyingorientations of the immobilised CYN on the protein conjugatebecause of the group reacted.
During the monoclonal fusion the hybridomas formed wereassessed for specific antibody production by both competitiveindirect ELISA and a SPR biosensor (Biacore Q, GE Healthcare)screening assay.
2.7.1. Competitive direct ELISA protocol for antibody screening
Serial dilutions (1/1K to 1/8K) of antibody (100 mL/well) in1 mM sodium acetate buffer were coated onto Falcon plates andincubated overnight at room temperature. All sera from immuno-gen approaches 1–5 were tested with each CYN–HRP fromapproaches 1–5. The antibody was removed and the HRP seriallydiluted in 1% BSA solution were added (50 mL/well) to appropriatewells in a chequerboard design. Negative and positive (1000 ng/mL) controls (50 mL/well) were incorporated into the assay todetermine binding and inhibition. The CYN–HRP and standardswere discarded and the plates washed with wash buffer. TMB/E
(100 mL/well) was added to each well and the reaction stoppedafter 12 min using 2.5 mM H2SO4 (25 mL/well). The plates wereread at 450 nm using a TECAN Safire2.
2.7.2. Competitive indirect ELISA protocol for antibody screening
Nunc Maxisorp plates were coated with either CYN–OVA orCYN–KLH conjugate (100 mL/well) in PBS (0.01 M, pH 7.4) andincubated at 4 8C overnight in the dark. The plates were blockedusing a commercial immunoassay stabiliser solution (100 mL/well)and incubated in an incubator/shaker for 1 h at 37 8C. The blockingbuffer was discarded and antisera or hybridoma supernatant(diluted 1:5 in PB buffer, pH 7.2) was added to the plate (100 mL/well). For hybridoma screening only CYN–OVA was used as thecoating antigen, the positive control was the sera obtained fromthe final heart bleed of the mouse and the negative control was cellculture medium diluted in PB buffer (pH 7.2). The plates wereincubated at 4 8C overnight in the dark, the solution discarded andthe plate washed four times using wash buffer. Secondary antibody(horseradish peroxidase-linked anti-species immunoglobulin) at1:2000 dilution was added to each well (100 mL/well) andincubated in the incubator shaker for 1 h at 37 8C in the dark.The secondary antibody was discarded and the plates washed fourtimes with wash buffer. TMB/E (100 mL/well) was added to eachwell and the reaction stopped after a few minutes using 2.5 mMH2SO4. The plates were read at 450 nm using a TECAN Safire2.
2.7.3. Biosensor protocol for antibody screening
The screening of the test sera for CYN antibodies was assessedon each chip surface by injecting dilutions (1 in 100) of the serumover the chip surface at a flow rate of 12 mL/min for 2 min. Reportpoints were recorded before (5 s) and after each injection (30 s),and the relative response units were determined. The chip surfacewas regenerated with a 25 mL injection of hydrochloric acid(10 mM) with 10% SDS at a flow rate of 25 mL/min. A typicalscreening cycle was 6–8 min.
2.8. Assessment of antibody sensitivity and specificity
2.8.1. Polyclonal antibodies
The sensitivity and specificity of the final polyclonal antiseraproduced in rabbits was assessed using a competitive indirectELISA and the Biacore Q biosensor assay with a CYN surfaceproduced using formaldehyde chemistry. Various parameters wereinitially investigated when developing both the ELISA andbiosensor assays to achieve optimal conditions. The conditionsapplied for each antibody characterised fully are summarised inTable 1.
Stock solutions of CYN and deoxyCYN standard were used toprepare working standards. For ELISA a set of working standards,ranging in concentration from 0 to 100 ng/mL in pH 7.2 PB and forbiosensor 0–100 ng/mL in pH 7.4 HBS–EP buffer, were prepared toproduce calibration curves for each toxin based on dose–responseby each method. The response units relative to the toxinconcentration curves were evaluated using a non linear regressioncurve fit using Graphpad Prism for Windows (version 5) in additionto determining the % binding at each toxin concentration, themidpoint (IC50) and the dynamic range (IC20–IC80) of each standardcurve and the specificity of the assay to other compounds. Thespecificity or % cross-reactivity for deoxyCYN relative to CYN wascalculated using a % ratio of mid-points for each of the standardcurves.
2.8.2. Monoclonal antibodies
On selection of the best performing monoclonal antibody, thecell line was grown and antibody was concentrated in cell culturemedium using Viva spin centrifuge tubes (30,000 MWCO) followed

Table 1ELISA and SPR assay parameters for each antibody.
Antibody Raised to ELISA SPR biosensor
Coating antigen Antibody
titre
Antibody
titre
Flow
rate (mL/min)
Contact
time (min)
Regeneration solution(s) Contact
time (s)
Polyclonal 1 CYN–KLH CYN–OVA (0.5 mg/mL) 1 in 8K 1 in 100 12 2 10 mM HCl/1% SDS 60
Polyclonal 2 CYN–KLH CYN–OVA (1.0 mg/mL) 1 in 16K 1 in 100 12 2 250 mM NaOH
10 mM HCl/1% SDS
60
30
Polyclonal 3 CYN–OVA CYN–KLH (5.0 mg/mL) 1 in 8K 1 in 100 12 2 250 mM NaOH/20% acetonitrile
250 mM NaOH/20% acetonitrile
40
40
Polyclonal 4 CYN–OVA CYN–KLH (5.0 mg/mL) 1 in 4K 1 in 100 12 2 250 mM NaOH
10 mM HCl/1% SDS
60
30
Monoclonal 1 CYN–KLH CYN–OVA (0.5 mg/mL) 1 in 5K 1 in 250 12 2 75 mM NaOH 30
Monoclonal 2 CYN–KLH CYN–OVA (0.5 mg/mL) 1 in 5K 1 in 250 12 2 75 mM NaOH 30
C.T. Elliott et al. / Harmful Algae 24 (2013) 10–1914
by purification via affinity chromatography using a protein G-sepharose gel column (MAbTrap Kit). Dialysis of the antibody over24 h in 0.15 M saline (3� 4 L) was performed. The proteinconcentration and isotyping of each antibody was determined atA280 nm and a mouse monoclonal antibody isotyping kit,respectively. The monoclonal antibodies were then assessed byELISA and SPR in the same way as the polyclonal antibodies usingthe parameters as outlined in Table 1.
3. Results and discussion
3.1. Preparation of CYN immunogens
Six different immunogens were prepared comparing directlyfive different chemical approaches and two different carrierproteins. The rationale behind assessing different chemicalapproaches was to link the carrier protein to different sites ofthe CYN molecule or its derivatised analogue, as illustrated inFig. 2, to determine the best orientation of the CYN molecule toinduce an immune response.
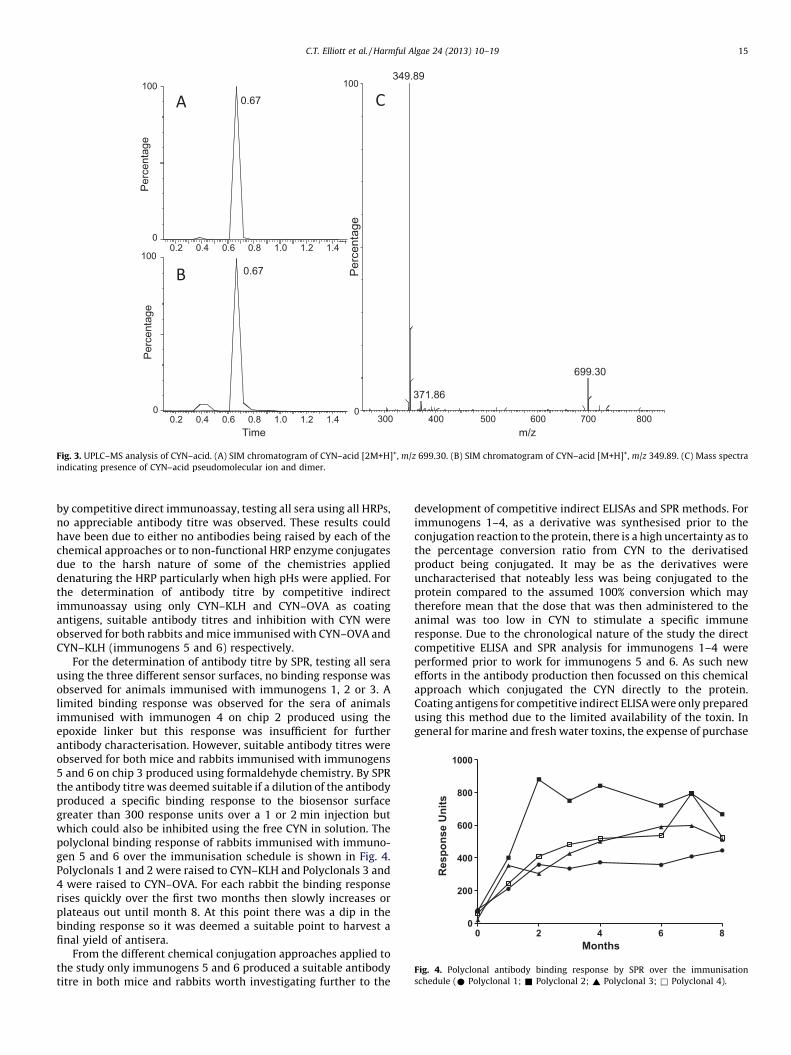
For immunogens 1 and 2 the production of CYN–acid (molecularweight: 349 g/mole) was confirmed by UPLC–mass spectrometry.Aliquots (20 mL) of the NaOCl–CYN reaction mixture were mixedwith 80 mL of eluent (98:2% (v/v) water:ACN modified with 0.1%TFA) and analysed every 24 h by isocratic UPLC–MS with a C18column (0.35 mL/min flow rate; 5 mL injection volume; 50 8Ccolumn temperature). Selective ion monitoring (SIM) in positivemode for pseudomolecular ions, sodium adducts and in the case of
Fig. 2. Conjugation sites on the CYN m
CYN–acid a dimer, was performed so that the relative proportions ofstarting material (CYN; [M+H]+, m/z 416; [M+Na]+, m/z 438),intermediate product (5-chloro-CYN; [M+H]+, m/z 450; [M+Na]+, m/z 472) and final product (CYN–acid; [M+H]+, m/z 350; [M+Na]+, m/z372; [2M+H]+, m/z 699) could be evaluated. The reaction mixtureanalysed on day 7 contained no detectable levels of 5-chloro-CYN orCYN and was deemed ready for use in conjugation (Fig. 3). Althoughit can be confirmed that the CYN–acid was formed, the quantity orpercentage conversion could not be determined due to the lack ofavailable analytical standards. For immunogens 3 and 4 spacermolecules were incorporated into the conjugation process. Succinicanhydride was reacted with the hydroxyl reaction site (Fig. 2C) of theCYN molecule to form a hemisuccinate derivative which creates aterminal carboxylic acid functional group that is then conjugated tothe amine of the protein via the carbodiimide reaction. Similarly, thebis-epoxide homobifunctional compound 1,4-butanediol diglycidylether was employed to cross-link the amine of the protein with thehydroxyl reaction site of the CYN molecule (Fig. 2C).
For immunogens 5 and 6 two different proteins were bothconjugated via a modified Mannich reaction which most probablyreacted through the indole group of the guanidinium moiety of theCYN (Fig. 2C) as was previously reported for cyclopiazonic acid(Huang and Chu, 1993).
3.2. Antibody titres by ELISA and SPR
The three screening methods were utilised to assess antibodytitre for the sera produced. For the determination of antibody titre
olecule and derivatised analogues.
349.89
699.30
371.86
C
B
A 0.67
0.67
m/z
300 40 0 50 0 60 0 70 0 80 00
100
Time
0.2 0.4 0.6 0.8 1.0 1.2 1.40
1000.2 0.4 0.6 0.8 1.0 1.2 1.4
0
100
Percentage
Percentage
Percentage
Fig. 3. UPLC–MS analysis of CYN–acid. (A) SIM chromatogram of CYN–acid [2M+H]+, m/z 699.30. (B) SIM chromatogram of CYN–acid [M+H]+, m/z 349.89. (C) Mass spectra
indicating presence of CYN–acid pseudomolecular ion and dimer.
0 2 4 6 80
200
400
600
800
1000
Months
Re
sp
on
se
Un
its
Fig. 4. Polyclonal antibody binding response by SPR over the immunisation
schedule (* Polyclonal 1; & Polyclonal 2; ~ Polyclonal 3; & Polyclonal 4).
C.T. Elliott et al. / Harmful Algae 24 (2013) 10–19 15
by competitive direct immunoassay, testing all sera using all HRPs,no appreciable antibody titre was observed. These results couldhave been due to either no antibodies being raised by each of thechemical approaches or to non-functional HRP enzyme conjugatesdue to the harsh nature of some of the chemistries applieddenaturing the HRP particularly when high pHs were applied. Forthe determination of antibody titre by competitive indirectimmunoassay using only CYN–KLH and CYN–OVA as coatingantigens, suitable antibody titres and inhibition with CYN wereobserved for both rabbits and mice immunised with CYN–OVA andCYN–KLH (immunogens 5 and 6) respectively.
For the determination of antibody titre by SPR, testing all serausing the three different sensor surfaces, no binding response wasobserved for animals immunised with immunogens 1, 2 or 3. Alimited binding response was observed for the sera of animalsimmunised with immunogen 4 on chip 2 produced using theepoxide linker but this response was insufficient for furtherantibody characterisation. However, suitable antibody titres wereobserved for both mice and rabbits immunised with immunogens5 and 6 on chip 3 produced using formaldehyde chemistry. By SPRthe antibody titre was deemed suitable if a dilution of the antibodyproduced a specific binding response to the biosensor surfacegreater than 300 response units over a 1 or 2 min injection butwhich could also be inhibited using the free CYN in solution. Thepolyclonal binding response of rabbits immunised with immuno-gen 5 and 6 over the immunisation schedule is shown in Fig. 4.Polyclonals 1 and 2 were raised to CYN–KLH and Polyclonals 3 and4 were raised to CYN–OVA. For each rabbit the binding responserises quickly over the first two months then slowly increases orplateaus out until month 8. At this point there was a dip in thebinding response so it was deemed a suitable point to harvest afinal yield of antisera.
From the different chemical conjugation approaches applied tothe study only immunogens 5 and 6 produced a suitable antibodytitre in both mice and rabbits worth investigating further to the
development of competitive indirect ELISAs and SPR methods. Forimmunogens 1–4, as a derivative was synthesised prior to theconjugation reaction to the protein, there is a high uncertainty as tothe percentage conversion ratio from CYN to the derivatisedproduct being conjugated. It may be as the derivatives wereuncharacterised that noteably less was being conjugated to theprotein compared to the assumed 100% conversion which maytherefore mean that the dose that was then administered to theanimal was too low in CYN to stimulate a specific immuneresponse. Due to the chronological nature of the study the directcompetitive ELISA and SPR analysis for immunogens 1–4 wereperformed prior to work for immunogens 5 and 6. As such newefforts in the antibody production then focussed on this chemicalapproach which conjugated the CYN directly to the protein.Coating antigens for competitive indirect ELISA were only preparedusing this method due to the limited availability of the toxin. Ingeneral for marine and fresh water toxins, the expense of purchase

0.00 1 0.01 0.1 1 10 10 0 100 00
20
40
60
80
100
120
CYN Concentration (ng/mL)
No
rmalised
op
tical d
en
sit
y
0.00 1 0.01 0.1 1 10 10 0 100 00
20
40
60
80
100
120
DeoxyCYN Concentration (ng/mL)
No
rmalised
op
tical d
en
sit
y
0.00 1 0.01 0.1 1 10 10 0 100 00
20
40
60
80
100
CYN Concentration (ng/mL)
No
rmalised
resp
on
se
0.00 1 0.01 0.1 1 10 10 0 100 00
20
40
60
80
100
DeoxyCYN Concentration (ng/mL)
No
rma
lis
ed
re
sp
on
se
(A) (B)
(C) (D)
Fig. 5. Calibration curves for polyclonal antibodies determined by ELISA for (A) CYN and (B) deoxyCYN and by SPR for (C) CYN and (D) deoxyCYN (* Polyclonal 1; & Polyclonal
2; ~ Polyclonal 3; ! Polyclonal 4).
C.T. Elliott et al. / Harmful Algae 24 (2013) 10–1916
and limited availability makes the challenge of finding a suitablechemistry to produce effective immunogens a difficult task.
3.3. Polyclonal antibody characterisation
For polyclonal antibody production, animals demonstrating asuitable titre received a total of nine immunisations before theirsacrifice whereby a final bleed was taken. The assay parametersused for both indirect competitive ELISA and SPR for the finalbleeds of the four polyclonal sera (Fig. 5) from immunogens 5 and 6are summarised in Table 1.
All four polyclonal antisera demonstrated high sensitivity,determined by the IC50, for CYN by ELISA ranging from 27 to131 pg/mL (Table 2). The dynamic range (IC20–IC80) deemed to bethe theoretical lower and upper detection limits for the assay wassimilar for all four polyclonal antibodies with Polyclonal 1 showingthe lowest theoretical detection limit of 10 pg/mL. For allantibodies the dynamic range was in the pg/mL range. CurrentlydeoxyCYN is not considered to be toxic compared to CYN thereforethe low percentage cross-reactivity demonstrated for all 4antibodies of up to 6.2% of this analogue of CYN in the SPR assayis beneficial so that an assay developed will not detect thedeoxyCYN to the same extent the much more potent CYN. Thisfinding strongly suggests that the hydroxyl molecule present inCYN but absent in deoxyCYN plays an important role in the abilityof the antibody to bind to the toxin. These polyclonal antibodiesshowed no cross-reactivity towards microcystins but this was notunexpected due to the much greater differences in molecularstructure.
By SPR all four polyclonal antibodies showed a similar titre withthose raised to KLH as the carrier protein displaying marginally
better sensitivity to CYN compared to those raised to OVA. Thesensitivity of the polyclonal antibodies by SPR ranged between 4.4and 9.6 ng/mL (Table 2). The dynamic range (IC20–IC80) was similarfor all four polyclonal antibodies with Polyclonal 1 showing thelowest theoretical detection limit of 1.9 ng/mL. The SPR assaydemonstrated slightly higher theoretical detection limits yetsimilar low % cross-reactivity for all 4 antibodies towardsdeoxyCYN of between 9.6 and 12.5% (Table 2).
3.4. Monoclonal antibody characterisation
For the purposes of monoclonal antibody production the micereceived a total of five immunisations and the best performingmouse in terms of titre detected was identified as belonging to amouse immunised with CYN–KLH. This animal was sacrificed, thespleen harvested and a fusion performed. From the fusion over atwo-week period 714 hybridomas were screened for antibodiesbinding to CYN conjugate. For those screened 10% (71) demon-strated antibody binding greater than the final heart bleedpolyclonal control and 26% (18) of those tested positive forinhibition when challenged with CYN. The best 4 performinghybridomas in terms of intensity of reaction observed were clonedand following the cloning procedure two clones which demon-strated the best inhibition were grown, purified and characterised.The final protein concentration of the purified monoclonalantibodies Monoclonal 1 and Monoclonal 2 was 0.948 and1.115 mg/mL determined by UV A280. Isotyping of the monoclonalantibodies demonstrated that both were IgG1 heavy chain with klight chains.
By ELISA and SPR the sensitivity of the monoclonal antibodies,Monoclonal 1 and Monoclonal 2, was on average slightly higher

Ta
ble
2S
en
siti
vit
ya
nd
spe
cifi
city
of
ea
cha
nti
bo
dy
by
ELI
SA
an
dS
PR
.
An
tib
od
yR
ais
ed
toE
LIS
AB
iose
nso
r
CY
NIC
50
(ng
/mL)
CY
NIC
20–
IC8
0
(ng
/mL)
De
ox
yC
YN
IC5
0(n
g/m
L)
De
ox
yC
YN
IC2
0–
IC8
0(n
g/m
L)
%C
Rto
de
ox
yC
YN
CY
NIC
50
(ng
/mL)
CY
NIC
20–
IC8
0
(ng
/mL)
De
ox
yC
YN
IC5
0(n
g/m
L)
De
ox
yC
YN
IC2
0–
IC8
0(n
g/m
L)
%C
Rto
de
ox
yC
YN
Po
lycl
on
al
1C
YN
–K
LH0
.02
70
.01
–0
.08
1.3
10
.25
–6
.82
.14
.41
.9–
10
.04
5.8
13
.4–
13
49
.6
Po
lycl
on
al
2C
YN
–K
LH0
.04
70
.01
–0
.19
1.1
10
.16
–9
.44
.27
.64
.0–
14
.77
7.2
29
.3–
20
4.5
9.8
Po
lycl
on
al
3C
YN
–O
VA
0.0
97
0.0
6–
0.1
71
.55
0.6
1–
6.1
6.2
8.9
3.9
–1
9.6
71
.32
5.5
–1
86
.81
2.5
Po
lycl
on
al
4C
YN
–O
VA
0.1
31
0.0
7–
0.2
22
.14
0.7
1–
4.9
6.1
9.6
4.5
–1
9.9
80
.93
9.2
–1
61
.81
1.9
Mo
no
clo
na
l1
CY
N–
KLH
0.1
21
0.0
4–
0.3
63
.81
.6–
8.7
3.2
9.7
4.4
–2
0.6
92
.03
8.7
–2
05
.31
0.5
Mo
no
clo
na
l2
CY
N–
KLH
0.1
08
0.0
6–
0.1
82
.10
.56
–6
.45
.11
1.1
5.7
–2
1.2
98
.94
0.1
–2
28
.11
1.2
0.00 1 0.01 0.1 1 10 10 00
20
40
60
80
100
120
Concentration (ng/mL)
No
rmalised
op
tical d
en
sit
y0.00 1 0.01 0.1 1 10 10 0 100 00
20
40
60
80
100
Concentration (ng/ml)
No
rmali
sed
resp
on
se
(A)
(B)
Fig. 6. Calibration curves for monoclonal antibodies determined by (A) ELISA and (B)
SPR for CYN and deoxyCYN (* CYN monoclonal 1; ^ deoxyCYN monoclonal 1; *
CYN monoclonal 2; ^ deoxyCYN monoclonal 2.
C.T. Elliott et al. / Harmful Algae 24 (2013) 10–19 17
than that of the polyclonal antibodies (Fig. 6). By ELISA thesensitivities were 108 and 121 pg/mL with % cross-reactivities todeoxyCYN of 3.2% and 5.1% and by SPR the sensitivities were 9.8and 11.1 ng/mL with % cross-reactivity for deoxyCYN at 10.5–11.2% (Table 2). The benefit of monoclonal antibodies over theirpolyclonal counterparts is well documented such as theirhomogeneous immortal reproducible characteristics comparedto polyclonal sera. There is no variability in specificity and affinityof monoclonal antibodies compared to polyclonal sera though theyare more laborious to produce and have a limited stability inchanges in pH or salt concentration.
3.5. Comparison of methods
This level of sensitivity and specificity for all antibodies wouldallow for the development of either fit for purpose ELISA or SPRassays to be constructed for food and environmental samples. TheELISA assay employed demonstrated a range in factor of 60–360times more sensitivity compared to the corresponding SPR assay.However, the SPR assay employed was a real time assay wherebythe sample being analysed could be monitored for CYN during theanalysis which would present a rapid result. For SPR one importantparameter to note is that the regeneration solutions employed hadto be optimised for each animal with Polyclonal 3 proving to bindstrongly to the chip surface thereby requiring harsher conditions tobe removed for the automated regeneration of the next sample. It isimportant that these regeneration conditions are consideredwhenever any automated sensor system is being employed toensure that the chip is fully conditioned to its original state for thenext analysis to avoid any issues with carryover in the system.When food or environmental samples are being analysed or theantibodies are being employed in different sensor technologies

C.T. Elliott et al. / Harmful Algae 24 (2013) 10–1918
these regeneration conditions may need re-optimised to theanalysis conditions at that time.
For the polyclonal sera a higher level of non-specific back-ground response was observed for the antibodies raised to OVA asthe carrier protein compared to those raised to KLH whenevaluated by ELISA. This is most likely due to non-specific bindingof the antibody to KLH as the coating antigen. This effect is reducedwhen free CYN is immobilised onto sensor surfaces which theneliminates non-specific protein–protein interactions.
For both the polyclonal and the monoclonal antibodiesgenerated in this study the selection of an appropriate platformfor the assay development would be more dependent of therequirements of the end user. The ELISA based tests would offergreater sensitivity whereas the SPR based assay would allow forrapid analysis of a sample at that time. The sample preparation ofthe environmental or food sample would also need to beconsidered and the ELISA based test would offer more scope inthe dilution of the sample to reduce interfering components basedon its improved sensitivity. For rapid portable testing theantibodies could be incorporated or employed using otherimmunoassay-based platforms such as lateral flow devices, flowcytometry or electrochemical sensors.
4. Conclusion
Antibodies of high sensitivity and specificity to CYN weresuccessfully produced for a range of future applications in rapid,immunobased detection platforms. This is the first report of thesuccessful production of antibodies to this toxin. One of thelimitations which hampers all researchers in the field of naturaltoxin analysis is either the availability or expense of the toxin topurchase. Since no work has been published on the development ofantibodies to this toxin it was important to find a suitablechemistry to use as an effective immunogen and to discuss all theapproaches taken to achieve the success reported. Using a modifiedMannich reaction for immunogen synthesis both polyclonal andmonoclonal antibodies of high sensitivity were produced. It wasdemonstrated that these antibodies could be used in two assayformats of ELISA and SPR. Future work will be employed in theapplication of these antibodies on different immunoassay basedplatforms for food and environmental sample analysis to deter-mine CYN presence.
Acknowledgements
This research was funded by The Department of Employmentand Learning All Island Research programme (ASSET), the InterregProgramme through the project ‘‘Atlantox: Advanced Tests aboutNew Toxins appeared in the Atlantic Area’’ and the EU FP7 project‘‘mAQUA: Universal microarrays for the evaluation of fresh-waterquality based on detection of pathogens and their toxins’’ of Grantagreement no: 265409. The authors would like to acknowledgeboth Brett Greer and Eliza Barszczewska-Lyner for their technicalsupport.[SS]
References
Banker, R., Carmeli, S., Werman, M., Teltsch, B., Porat, R., Sukenik, A., 2001. Uracilmoiety is required for toxicity of the cyanobacterial hepatotoxin. Journal ofToxicology and Environmental Health. Part A 62, 281–288.
Banker, R., Teltsch, B., Sukenik, A., Carmeli, S., 2000. 7-Epi, a toxic minor metaboliteof the cyanobacterium Aphanizomenon ovalisporum from Lake Kinneret, Israel.Journal of Natural Products 63, 387–389.
Bernard, C., Harvey, M., Briand, J.F., Bire, R., Krys, S., Fontaine, J.J., 2003. Toxicologicalcomparison of diverse Cylindrospermopsis raciborskii strains: evidence of liverdamage caused by a French C. raciborskii strain. Environmental Toxicology 18,176–186.
Berry, J.P., Lind, O., 2010. First evidence of paralytic ‘‘ralytic shellfish toxins’’ and in aMexican freshwater system, LagoCatemaco, and apparent bioaccumulation of
the toxins in ‘‘tegogolo’’ snails (Pomacea patula catemacensis). Toxicon 55,930–938.
Beyer, D., Suranyi, G., Vasas, G., Roszik, J., Erdodi, F., M-Hamvas, M., Bacsi, I., Batori,R., Serfozo, Z., Szigeti, Z.M., Vereb, G., Demeter, Z., Gonda, S., Mathe, C., 2009.Cylindrospermopsin induces alterations of root histology and microtubuleorganization in common reed (Phragmites australis) plantlets cultured in vitro.Toxicon 54 (4), 440–449.
Blahova, L., Oravec, M., Marsalek, B., Sejnohova, L., Simek, Z., Blaha, L., 2009. The firstoccurrence of the cyanobacterial alkaloid toxin in the Czech Republic asdetermined by immunochemical and LC/MS methods. Toxicon 53 (5), 519–524.
Brient, L., Lengronne, M., Bormans, M., Fastner, J., 2009. First occurrence of Cylin-drospermopsin in freshwater in France. Environmental Toxicology 24 (4),415–420.
Cadel-Six, S., Peyraud-Thomas, C., Brient, L., de Marsac, N.T., Rippka, R., Mejean, A.,2007. Different genotypes of anatoxin-producing cyanobacteria coexist in theTarn river, France. Applied and Environmental Microbiology 73 (23), 7605–7614.
Campbell, K., Stewart, L.D., Fodey, T.L., Haughey, S.A., Doucette, G.J., Kawatsu, K.,Elliott, C.T., 2007. An assessment of specific binding proteins suitable for thedetection of paralytic shellfish poisons (PSP) using optical biosensor technolo-gy. Analytical Chemistry 79 (15), 5906–5914.
Carson, B., 2000. Cylindrospermopsin: Review of Toxicological Literature. Integrat-ed Laboratory Systems, North Carolina.
Cheng, X.L., Shi, H.L., Adams, C.D., Timmons, T., Ma, Y.F., 2009. Effects of oxidativeand physical treatments on inactivation of Cylindrospermopsis raciborskii andremoval of cylindrospermopsin. Water Science and Technology 60 (3), 689–697.
Codd, G., Lindsay, J., Young, F., Morrison, L., Metcalf, J., 2005. In: Huisman, J.,Matthijs, H., Visser, P. (Eds.), Harmful Cyanobacteria. Springer,Netherlands, pp. 1–23.
Crooks, S.R.H., Traynor, I.M., Elliott, C.T., McCaughey, W.J., 1997. Detection ofmonensin residues in poultry liver using an enzyme immunoassay. Analyst122, 161–163.
Duy, T.N., Lam, P.K.S., Shaw, G.R., Connell, D.W., 2000. Toxicology and risk assess-ment of freshwater cyanobacterial (blue-green algal) toxins in water. Reviewsof Environment Contamination and Toxicology 163, 113–186.
Eaglesham, G.K., Norris, R.L., Shaw, G.R., Smith, M.J., Chiswell, R.K., Davis, B.C.,Neville, G.R., Seawright, A.A., Moore, M.R., 1999. Use of HPLC–MS/MS to moni-tor, a blue-green algal toxin, for public health purposes. Environmental Toxi-cology 14, 151–154.
EFSA, 2008. Opinion of the scientific panel on contaminants in the food chain on arequest from the European Commission on marine biotoxins in shellfish –azaspiracids. The EFSA Journal 723, 1–52.
EFSA, 2009. Scientific opinion of the panel on contaminants in the food chain on arequest from the European Commission on marine biotoxins in shellfish –influence of processing in the levels of lipophilic marine biotoxins in bivalvemolluscs. The EFSA Journal 1016, 1–10.
EFSA, 2010. Scientific opinion on marine biotoxins in shellfish – cyclic imines(spirolides, gymnodimines, pinnatoxins and pteriatoxins). The EFSA Journal1628, 1–39.
Erlanger, B.F., Borek, F., Beiser, S.M., Lieberman, S., 1957. Steroid-protein conjugates.I. Preparation and characterization of conjugates of bovine serum albumin withtestosterone and with cortisone. Journal of Biological Chemistry 228, 713–727.
Falconer, I.R., Hardy, S.J., Humpage, A.R., Froscio, S.M., Tozer, G.J., Hawkins, P.R.,1999. Hepatic and renal toxicity of the blue-green alga (cyanobacterium)Cylindrospermopsis raciborskii in male Swiss albino mice. Environmental Toxi-cology 14, 143–150.
Fodey, T., Murilla, G., Cannavan, A., Elliott, C., 2007. Characterisation of antibodies tochloramphenicol, produced in different species by enzyme-linked immunosor-bent assay and biosensor technologies. Analytica Chimica Acta 592, 51–57.
Froscio, S.M., Humpage, A.R., Burcham, P.C., Falconer, I.R., 2001. Cell-free proteinsynthesis inhibition assay for the cyanobacterial toxin. Environmental Toxicol-ogy 16, 408–412.
Froscio, S.M., Humpage, A.R., Burcham, P.C., Falconer, I.R., 2003. Cylindrospermop-sin-induced protein synthesis inhibition and its dissociation from acute toxicityin mouse hepatocytes. Environmental Toxicology 18, 243–251.
Galfre, G., Milstein, C., 1981. Preparation of monoclonal antibodies: strategies andprocedures. Methods in Enzymology 72, 3–46.
Gallo, P., Fabbrocino, S., Cerulo, M.G., Ferranti, P., Bruno, M., Serpe, L., 2009.Determination of Cylindrospermopsin in freshwaters and fish tissue by liquidchromatography coupled to electrospray ion trap mass spectrometry. RapidCommunications in Mass Spectrometry 23 (20), 3279–3284.
Griffiths, D.J., Saker, M.L., 2003. The Palm island mystery disease 20 years on: areview of research on the cyanotoxin. Environmental Toxicology 18 (2), 78–93.
Hermanson, G.T. (Ed.), 1996. Homobifunctional cross-linkers: bioconjugate tech-niques. Academic Press, London, pp. p.220–p.221.
Huang, X., Chu, F.S., 1993. Production and characterization of monoclonal andpolyclonal antibodies against the mycotoxin cyclopiazonic acid. Journal ofAgricultural and Food Chemistry 41, 329–333.
Hudnell, H.K., Dortch, Q., Zenick, H., 2008. An overview of the interagency. Interna-tional Symposium on Cyanobacterial Harmful Algal Blooms (ISOC-HAB): Ad-vancing the Scientific Understanding of Freshwater Harmful Algal Blooms. In:Hudnell, H.K. (Ed.), Cyanobacterial Harmful Algal Blooms: State of the Scienceand Research Needs. Springer, New York, pp. 1–16.
Kinnear, S., 2010. A decade of progress on bioaccumulation research. Marine Drugs8 (3), 542–564.

C.T. Elliott et al. / Harmful Algae 24 (2013) 10–19 19
Kittler, K., Schreiner, M., Krumbein, A., Manzei, S., Koch, M., Rohn, S., Maul, R., 2012.Uptake of the cyanobacterial toxin in Brassica vegetables. Food Chemistry,http://dx.doi.org/10.1016/j.foodchem.2012.01.107.
Li, R., Carmichael, W.W., Brittain, S., Eaglesham, G.K., Shaw, G.R., Liu, Y., Watanabe,M.M., 2001. First report of the cyanotoxins Cylindrospermopsin and deoxycylindrospermopsin from Raphidiopsis curvata (cyanobacteria). Journal of Phy-cology 37, 1121–1126.
Looper, R.G., Runnegar, M.T.C., Williams, R.M., 2005. Synthesis of the putativestructure of 7-deoxy: C7 oxygenation is not required for the inhibition ofprotein synthesis. Angewandte Chemie International Edition 44, 3879–3881.
Messineo, V., Bogialli, S., Melchiorre, S., Sechi, N., Luglie, A., Casiddu, P., Marlani,M.A., Padedda, B.M., Di Corcia, A., Mazza, R., Carloni, E., Bruno, M., 2009.Cyanobacterial toxins in Italian freshwaters. Limnologica 39 (2), 95–106.
Norris, R.L., Eaglesham, G.K., Pierens, G., Shaw, G.R., Smith, M.J., Chiswell, R.K.,Seawright, A.A., Moore, M.R., 1999. Deoxy, cylindrospermopsin an analog ofcylindrospermopsin from Cylindrospermopsis raciborskii. Environmental Toxi-cology 14, 163–165.
Norris, R.L.G., Seawright, A.A., Shaw, G.R., Senogles, P., Eaglesham, G.K., Smith, M.J.,Chiswell, R.K., Moore, M.R., 2002. Hepatic xenobiotic metabolism of cylindros-permopsin in vivo in the mouse. Toxicon 40, 471–476.
Norris, R.L.G., Seawright, A.A., Shaw, G.R., Smith, M.J., Chiswell, R.K., Moore, M.R.,2001. Distribution of 14C cylindrospermopsin in vivo in the mouse. Environ-mental Toxicology 16, 498–505.
Nybom, S.M.K., Salminen, S.J., Meriluoto, J.A.O., 2008. Specific strains of probioticbacteria are efficient in removal of several different cyanobacterial toxins fromsolution. Toxicon 52 (2), 214–220.
Oehrle, S.A., Southwell, B., Westrick, J., 2010. Detection of various freshwatercyanobacterial toxins using ultra-performance liquid chromatography tandemmass spectrometry. Toxicon 55, 965–972.
Ohtani, I., Moore, R.E., 1992. Cylindrospermopsin: A potent hepatotoxin from theblue-green alga Cylindrospermopsis raciborskii. Journal of the American Chemi-cal Society 114, 7941–7942.
Paerl, H.W., Huisman, J., 2008. Blooms like it hot. Science 320 (5872), 57–58.Paredes, I., Rietjens, I.M.C.M., Vieites, J.M., Cabado, A.G., 2011. Update of risk
assessments of main marine biotoxins in the European Union. Toxicon 58(4), 336–354.
Preussel, K., Wessel, G., Fastner, J., Chorus, I., 2009. Response of cylindrospermopsinproduction and release in Aphanizomenon flos-aquae (Cyanobacteria) to varyinglight and temperature conditions. Harmful Algae 8 (5), 645–650.
Rawn, D.F.K., Niedzwiadek, B., Lau, B.P.Y., Saker, M., 2007. Anatoxin-a and itsmetabolites in blue-green algae food supplements from Canada and Portugal.Journal of Food Protection 70 (3), 776–779.
Reisner, M., Carmeli, S., Werman, M., Sukenik, A., 2004. The cyanobacterial toxincylindrospermopsin inhibits pyrimidine nucleotide synthesis and alters cho-lesterol distribution in mice. Toxicological Sciences 82 (2), 620–627.
Tadayoni, B.M., Friden, P.M., Walus, L.R., Musso, G.F., 1993. Synthesis in vitrokinetics, and in vivo studies on protein conjugates of AZT: evaluation as atransport system to increase brain delivery. Bioconjugate Chemistry 4,139–145.
Thomas, A.D., Saker, M.L., Norton, J.H., Olsen, R.D., 1998. Cyanobacterium Cylin-drospermopsis raciborskii as a probable cause of death in cattle in Queensland.Australian Veterinary Journal 76, 592–594.





























