Embed Size (px)
Citation preview

Identification of binding sites of TEFM with mitochondrial
RNA polymerase
James Graber
Rowan University Summer Undergraduate Research Experience 2015
Department of Cell Biology
Dr. Dmitry Temiakov

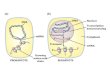
Mitochondria has an unique genetic system
• “powerhouse of the cell”• ATP production
• Maintains its own independent genome• Similarities to bacterial genetics (circular DNA, multigenic transcripts, etc)• T7 phage-like RNA polymerase
• Mitochondrial genetics play an important role in mitochondrial diseases, neurological disorders, senescence, and cell development

TEFM behaves as a molecular switch between replication and transcription• Transcription Elongation Factor of
Mitochondria (TEFM)• Transcription terminates
prematurely at CSBII (G6AG8), creating replication primer• In presence of TEFM, RNA
polymerase effectively transcribes through CSBII

The molecular mechanism of TEFM’s anti-termination activity is not known
While the structure of RNA polymerase is known, TEFM’s structure is not. Identification of binding sites will allow complete structural mapping of TEFM and RNA polymerase interaction. With this information, the molecular mechanism of TEFM’s anti-termination activity can be better understood.

Identification of binding sites in Elongation Complex by photo cross-linking

• Site-specific substitution with the amber codon (TAG)• Sites encoding charged amino acid residues along
the N-terminal domain• Amber Mutations
• K55• K63• S72• R89• Q110• K122
The tertiary structure of TEFM’s N-terminal domain is unknown
Secondary structure:

Three forms of chromatography to purify TEFM: Affinity, Ion Exchange, and Size Exclusion
lysate Ni-NTA Eluate
Heparin Eluate Gel Filtration fractions

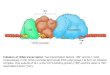
Figure 1: Photo Cross-linking Figure 2: Transcription Assay
TEFM-R89Bpa mutant does not cross-link or prevents transcript elongation

Conclusions• TEFM-R89Bpa does not crosslink with RNA polymerase or scaffold (Figure 1)
• Amino acid residue R89 of TEFM is not a binding site in the elongation complex
• Substitution of R89 with Bpa in TEFM does not have detrimental effects on TEFM’s anti-termination activity (Figure 2)• R89 has no significant function in preventing termination at CSBII
• Juxtaposition of data suggests that amino acid residue R89 does not play a role in TEFM’s binding to the elongation complex or TEFM’s anti-termination activity
• Further in vitro analysis of N-terminal residues using unnatural amino acid mutagenesis of photocrosslinker Bpa should be explored so that the molecular mechanism of TEFM’s anti-termination activity can be better understood

References1. K. Agaronyan, Y. I. Morozov, M. Anikin, and D. Temiakov, Replication-transcription switch in
human mitochondria, Science. 30 January 2015: 347 (6221), 548-551. doi:10.1126/science.aaa0986.
2. T. S. Young, I. Ahmad, J. A. Yin, and P. G. Schultz, An Enhanced System for Unnatural Amino Acid Mutagenesis in E. coli, Journal of Molecular Biology, Volume 395, Issue 2, 15 January 2010, Pages 361-374, ISSN 0022-2836, http://dx.doi.org/10.1016/j.jmb.2009.10.030. (http://www.sciencedirect.com/science/article/pii/S0022283609012704)
3. Y. I. Morozov, K. Agaronyan, A. C. M. Cheung, M. Anikin, P. Cramer, and D. Temiakov, A novel intermediate in transcription initiation by human mitochondrial RNA polymerase, Nucleic Acids Research. 2014; 42 (6), 3884-3893. doi:10.1093/nar/gkt1356.
4. “Mitochondria.” University of California, Davis. n.d. Web. 30 July 2015. <https://smartsite.ucdavis.edu/access/content/user/00002950/bis10v/week2/13mitochondria.html>.
5. “QuikChange II Site-Directed Mutagenesis Kits - Details & Specifications.” Agilent Technologies. 2015. Web. 30 July 2015. <http://www.genomics.agilent.com/article.jsp?pageId=384&_requestid=602465>.

Questions?