Embed Size (px)
Citation preview

Microbes and Infection 12 (2010) 635e642www.elsevier.com/locate/micinf
Original article
Filaria-induced IL-10 suppresses murine cerebral malaria
Sabine Specht a,1,*, Daniel Fernandez Ruiz a,1, Bettina Dubben a, Susanne Deininger a,Achim Hoerauf a
a Institute of Medical Microbiology, Immunology and Parasitology, University Hospital Bonn, Sigmund Freud Strasse 25, 53105 Bonn, Germany
Received 5 January 2010; accepted 15 April 2010
Available online 24 April 2010
Abstract
Filarial nematodes achieve long survival in their hosts due to their capacity to modulate immune responses. Therefore, immunomodulation byfilarial nematodes may alter responses to concomitant infections such as malaria. Cerebral malaria (CM), a severe complication of Plasmodiumfalciparum infections, is triggered as a consequence of the immune response developed against malaria parasites. The question arises whetherprior infection with helminth parasites is beneficial against CM. In the present work a murine model for subsequent has been used to assess thishypothesis. C57BL/6 mice were infected with the rodent filarial parasite Litomosoides sigmodontis and the murine model parasite for CM,Plasmodium berghei ANKA. Previously filaria-infected C57BL/6 mice showed significantly reduced CM rates. CD8þ T cell recruitment to thebrain, a hallmark for CM development, was reduced in protected mice. Furthermore, in contrast to P. berghei single-infected animals, filaria-infected mice had significantly higher levels of circulating IL-10. The requirement for IL-10 in CM protection was demonstrated by the lack ofprotection in IL-10 KO mice. This suggests that the anti-inflammatory IL-10 elicited by filarial nematodes is able to suppress the overwhelminginflammatory reaction otherwise triggered against malaria parasites in C57BL/6 mice, preventing full progress to CM.� 2010 Elsevier Masson SAS. All rights reserved.
Keywords: Cerebral malaria; Filariasis; IL-10; Litomosoides sigmodontis; Plasmodium berghei
1. Introduction
Filarial nematodes and malaria parasites are wide spreadand co-endemic in the tropics and in some regions even sharethe same transmitting vector species [1,2]. Hence, the possi-bility of being concomitantly infected is high for individualsliving in co-endemic regions. As a result of infection withfilarial and plasmodial parasites, be it simultaneous or subse-quent, the development of pathology or ability to combat theinfections may be profoundly altered.
Filarial nematodes use multiple immunoregulatory mech-anisms that allow them to survive for prolonged periods oftime inside the host by evading immune attack. An importantway to achieve this is by the induction of immunoregulatory
* Corresponding author. Tel.: þ49 228 287 11453; fax: þ49 228 287 19573.
E-mail address: [email protected] (S. Specht).1 These authors contributed equally.
1286-4579/$ - see front matter � 2010 Elsevier Masson SAS. All rights reserved.
doi:10.1016/j.micinf.2010.04.006
cytokines such as IL-10. As an anti-inflammatory cytokine,IL-10 impairs the production of pro-inflammatory cytokinesand the expression of the class II major histocompatibilitycomplex (MHC) [3]. In response to filarial antigen macro-phages produce IL-10, which in turn down-regulates theirexpression of the co-stimulatory molecules CD80 and CD86[4]. In contrast, IFN-g production is increased after in vitroneutralisation of IL-10 in PMBCs from filaria-infected patients[5]. The murine model with Litomosoides sigmodontis repro-duces many of the features of human filariasis, includingimmunosuppression through IL-10 [6,7]. Beyond, Th2 cyto-kines IL-4 and IL-5 are produced in response to a L. sigmo-dontis infection causing rapid expansion of T cells, B cells,macrophages, neutrophils and eosinophils in the thoraciccavity leading to containment of the worms [8,9]. Hostimmune responses against plasmodial parasites result inparasite elimination but also in excessive inflammation whichcan develop into life threatening cerebral malaria (CM).Central steps in the progress to CM are the release of

636 S. Specht et al. / Microbes and Infection 12 (2010) 635e642
pro-inflammatory cytokines and the subsequent sequestrationof infected erythrocytes and immune cells in the capillaries ofthe brain [10]. Relevant features of the pathophysiology ofhuman CM can be reproduced in the malaria model P. bergheiANKA in C57BL/6 mice [11]. The sequestration of predom-inantly cytotoxic CD8þ T has been shown to be a key inducerof CM pathogenesis [12].
Co-infections can alter the immune responses elicitedagainst single pathogens [13,14]. Filarial-derived immuno-suppression might alter the pro-inflammatory aetiology of CMand could potentially be beneficial for the host. In order toassess this hypothesis, CM development was analysed in thepresent work in a murine model of consecutive infection withL. sigmodontis and P. berghei ANKA in C57BL/6 mice.Filaria-infected C57BL/6 mice showed a decrease in thesequestration of CD8þ T cells in the brain and reduced malariasymptoms in an IL-10 dependent manner after P. bergheiinfection. Our results reflect that filarial infections mayconstitute a natural means of protection from CM pathology.
2. Materials and methods
2.1. Mice and infections
Wild type and IL-10 KO C57BL/6 mice were maintainedand bred at the animal facility of the University HospitalBonn. L. sigmodontis was maintained by cyclic passagebetween the cotton rat (Sigmodon hispidus) and mites (Orni-thonyssus bacoti), as described before [15]. Stocks of P. ber-ghei infected red blood cells (iRBC) were stored in liquidnitrogen and injected in a C57BL/6 mouse in order to generateiRBC for experimental P. berghei infections. For the experi-ments, six to eight weeks old female C57BL/6 mice werenaturally infected with the filarial nematode L. sigmodontis. 60days later, the same animals were inoculated with 50,000 P.berghei ANKA iRBC. When lower concentrations (10,000iRBC and lower) were used, a generation of CM pathology inall of the control mice was not guaranteed. Each infectionexperiment was performed with 4e8 mice per group unlessotherwise noted.
2.2. P. berghei ANKA parasitaemia and spleen index
Blood samples were collected daily from the tail vein.Giemsa-stained (Merck, Darmstadt, Germany) thin bloodsmears were prepared and parasitaemia was quantified as thepercentage of iRBCs per 500 RBCs per slide. The spleen indexwas calculated as the ratio of spleen wet weight (g) versusbody weight (g) � 100.
2.3. Cell extraction from spleen and brain
Mice were perfused before removal of the organs by PBS toflush out blood cells. Spleens were aseptically isolated,weighed and then homogenized through a sieve. Erythrocyteswere lysed by incubating the cell pellet in cold 0.1 M Trisammonium chloride solution for 10 min and then cells were
spun down (4 �C, 5 min, 400 g). The remaining leukocyteswere washed with PBS, suspended in RPMI medium (PAAlaboratories, Pasching, Austria) and filtered through a nylonnet. Cells were stored at 4 �C until further use. Brains wereresected and homogenized to single-cell suspensions througha sieve. After 35% Percoll separation cell pellets were washedwith PBS, filtered through nylon net and kept at 4 �C in RPMImedium for FACS analysis.
2.4. Pleural wash
The thoracic cavity was flushed with one ml PBS. Theobtained thoracic cavity fluid was centrifuged (4 �C, 5 min,400 g) and the supernatant was stored at �20 �C for cytokinemeasurements.
2.5. Flow cytometry
Cells from spleen and brain were stained following stan-dard protocols after Fc receptor blockage with rat IgG anti-bodies (SigmaeAldrich, Steinheim, Germany). Combinationsof the following antibodies were used: anti-CD4-FITC (cloneRM4-4, ebioscience, San Diego, CA), anti-CD3e-PerCP-Cy5.5 (clone 145-2C11), anti-CD8a-PE or anti-CD8aePE-Cy7 (clone 53-6.7), anti-CD25-APC (clone PC61), NK1.1-PE(clone PK136), gd-TCR-PE (clone GL3), CD11b-FITC (Mac-1a chain, clone M1/70) and B220-PE (CD45R, clone RA3-6B2, BD Pharmingen, Heidelberg, Germany). Unbound anti-bodies were removed by washing twice with PBS. The cellswere fixed in 2% paraformaldehyde and analysed using a BDFACS Canto (BD Biosciences, Heidelberg, Germany).
2.6. Enzyme-linked immunosorbent assay
Concentrations of IL-10, IL-4, IL-5, TGF-b and IFN-g weredetermined by sandwich-ELISA using standard protocols. Theantibody pairs for capture and detection (biotinylated) werepurchased from BD Pharmingen. Recombinant cytokines (BDPharmingen) were used as standards. All ELISAs were devel-oped after incubation with streptavidineperoxidase complex(1:10,000; RocheeBoehringer, Mannheim, Germany), using3,30, 5,50-tetramethylbenzidine (Roth, Karlsruhe, Germany) at6 mg/ml in dimethyl sulfoxide as the substrate. For TGF-b measurements, a prior activation of this cytokine was per-formed by incubating samples in 0.02 N HCl for 15 min andsubsequent bufferingwith 0.02NNaOH.The sampleswere readon a spectrophotometer at 450 nm.
2.7. Statistics
Statistical analyses were performed using GraphPad Prismsoftware. Normality of data distributions was assessed by theKolmogoroveSmirnov normality test. Parametrically distrib-uted data were analysed by unpaired T test or one-way ANOVAwith Tukey’s multiple comparisons post-hoc test to checkdifferences between single groups and represented asmean� SD. Non-parametrically distributed data were analysed
637S. Specht et al. / Microbes and Infection 12 (2010) 635e642
using ManneWhitney test or KruskaleWallis test plus Dunn’smultiple comparisons post-hoc test to check differencesbetween single groups and represented as median and range.
3. Results
3.1. Reduced incidence of CM in mice after filarialinfection
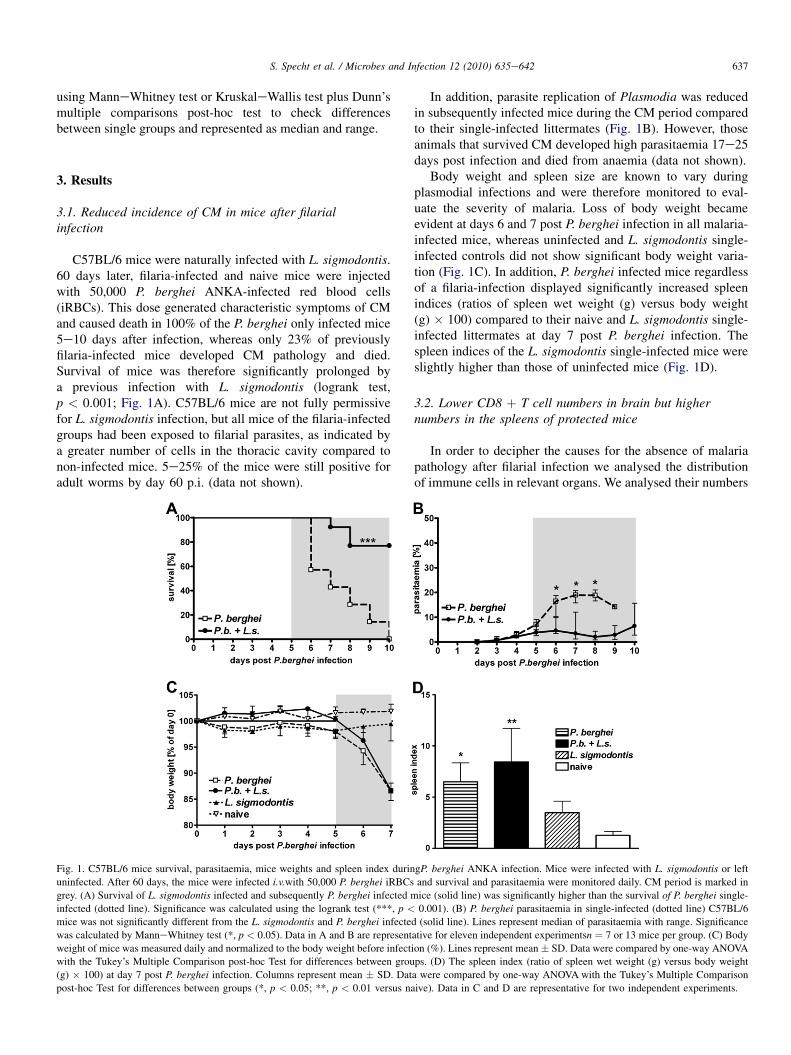
C57BL/6 mice were naturally infected with L. sigmodontis.60 days later, filaria-infected and naive mice were injectedwith 50,000 P. berghei ANKA-infected red blood cells(iRBCs). This dose generated characteristic symptoms of CMand caused death in 100% of the P. berghei only infected mice5e10 days after infection, whereas only 23% of previouslyfilaria-infected mice developed CM pathology and died.Survival of mice was therefore significantly prolonged bya previous infection with L. sigmodontis (logrank test,p < 0.001; Fig. 1A). C57BL/6 mice are not fully permissivefor L. sigmodontis infection, but all mice of the filaria-infectedgroups had been exposed to filarial parasites, as indicated bya greater number of cells in the thoracic cavity compared tonon-infected mice. 5e25% of the mice were still positive foradult worms by day 60 p.i. (data not shown).
Fig. 1. C57BL/6 mice survival, parasitaemia, mice weights and spleen index durin
uninfected. After 60 days, the mice were infected i.v.with 50,000 P. berghei iRBCs
grey. (A) Survival of L. sigmodontis infected and subsequently P. berghei infected m
infected (dotted line). Significance was calculated using the logrank test (***, p <
mice was not significantly different from the L. sigmodontis and P. berghei infecte
was calculated by ManneWhitney test (*, p < 0.05). Data in A and B are represent
weight of mice was measured daily and normalized to the body weight before infect
with the Tukey’s Multiple Comparison post-hoc Test for differences between grou
(g) � 100) at day 7 post P. berghei infection. Columns represent mean � SD. Dat
post-hoc Test for differences between groups (*, p < 0.05; **, p < 0.01 versus na
In addition, parasite replication of Plasmodia was reducedin subsequently infected mice during the CM period comparedto their single-infected littermates (Fig. 1B). However, thoseanimals that survived CM developed high parasitaemia 17e25days post infection and died from anaemia (data not shown).
Body weight and spleen size are known to vary duringplasmodial infections and were therefore monitored to eval-uate the severity of malaria. Loss of body weight becameevident at days 6 and 7 post P. berghei infection in all malaria-infected mice, whereas uninfected and L. sigmodontis single-infected controls did not show significant body weight varia-tion (Fig. 1C). In addition, P. berghei infected mice regardlessof a filaria-infection displayed significantly increased spleenindices (ratios of spleen wet weight (g) versus body weight(g) � 100) compared to their naive and L. sigmodontis single-infected littermates at day 7 post P. berghei infection. Thespleen indices of the L. sigmodontis single-infected mice wereslightly higher than those of uninfected mice (Fig. 1D).
3.2. Lower CD8 þ T cell numbers in brain but highernumbers in the spleens of protected mice
In order to decipher the causes for the absence of malariapathology after filarial infection we analysed the distributionof immune cells in relevant organs. We analysed their numbers
gP. berghei ANKA infection. Mice were infected with L. sigmodontis or left
and survival and parasitaemia were monitored daily. CM period is marked in
ice (solid line) was significantly higher than the survival of P. berghei single-
0.001). (B) P. berghei parasitaemia in single-infected (dotted line) C57BL/6
d (solid line). Lines represent median of parasitaemia with range. Significance
ative for eleven independent experimentsn ¼ 7 or 13 mice per group. (C) Body
ion (%). Lines represent mean � SD. Data were compared by one-way ANOVA
ps. (D) The spleen index (ratio of spleen wet weight (g) versus body weight
a were compared by one-way ANOVA with the Tukey’s Multiple Comparison
ive). Data in C and D are representative for two independent experiments.
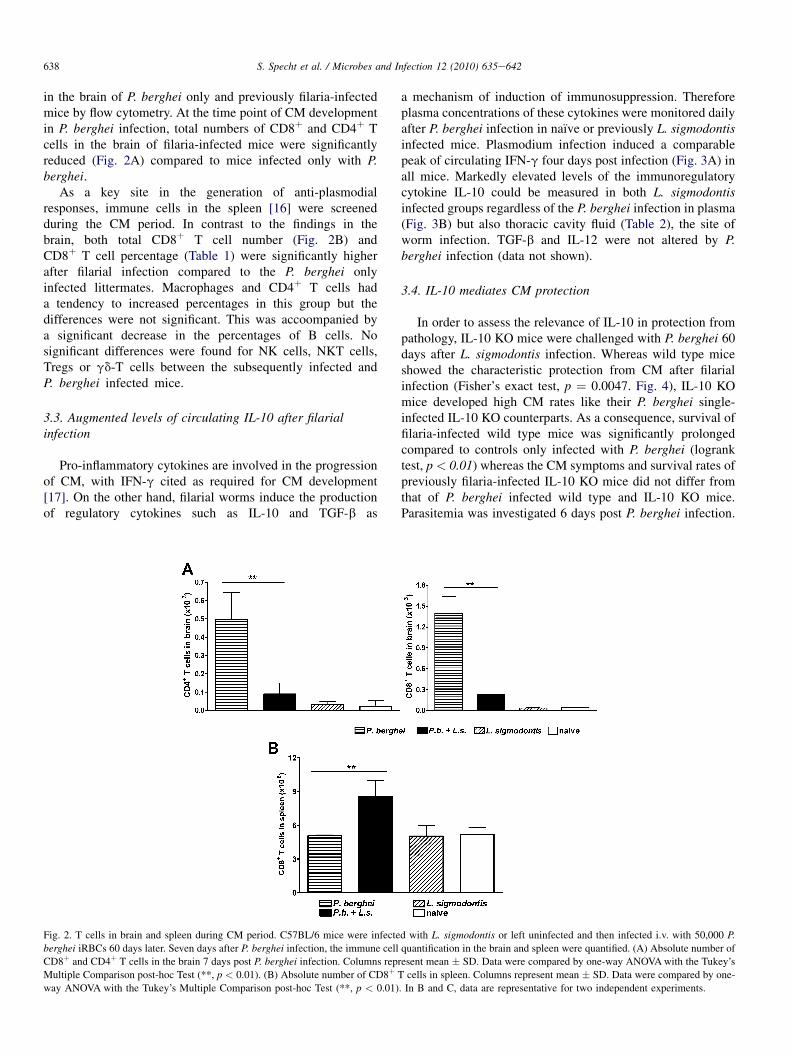
638 S. Specht et al. / Microbes and Infection 12 (2010) 635e642
in the brain of P. berghei only and previously filaria-infectedmice by flow cytometry. At the time point of CM developmentin P. berghei infection, total numbers of CD8þ and CD4þ Tcells in the brain of filaria-infected mice were significantlyreduced (Fig. 2A) compared to mice infected only with P.berghei.
As a key site in the generation of anti-plasmodialresponses, immune cells in the spleen [16] were screenedduring the CM period. In contrast to the findings in thebrain, both total CD8þ T cell number (Fig. 2B) andCD8þ T cell percentage (Table 1) were significantly higherafter filarial infection compared to the P. berghei onlyinfected littermates. Macrophages and CD4þ T cells hada tendency to increased percentages in this group but thedifferences were not significant. This was accoompanied bya significant decrease in the percentages of B cells. Nosignificant differences were found for NK cells, NKT cells,Tregs or gd-T cells between the subsequently infected andP. berghei infected mice.
3.3. Augmented levels of circulating IL-10 after filarialinfection
Pro-inflammatory cytokines are involved in the progressionof CM, with IFN-g cited as required for CM development[17]. On the other hand, filarial worms induce the productionof regulatory cytokines such as IL-10 and TGF-b as
Fig. 2. T cells in brain and spleen during CM period. C57BL/6 mice were infecte
berghei iRBCs 60 days later. Seven days after P. berghei infection, the immune cell
CD8þ and CD4þ T cells in the brain 7 days post P. berghei infection. Columns rep
Multiple Comparison post-hoc Test (**, p < 0.01). (B) Absolute number of CD8þ T
way ANOVA with the Tukey’s Multiple Comparison post-hoc Test (**, p < 0.01)
a mechanism of induction of immunosuppression. Thereforeplasma concentrations of these cytokines were monitored dailyafter P. berghei infection in naıve or previously L. sigmodontisinfected mice. Plasmodium infection induced a comparablepeak of circulating IFN-g four days post infection (Fig. 3A) inall mice. Markedly elevated levels of the immunoregulatorycytokine IL-10 could be measured in both L. sigmodontisinfected groups regardless of the P. berghei infection in plasma(Fig. 3B) but also thoracic cavity fluid (Table 2), the site ofworm infection. TGF-b and IL-12 were not altered by P.berghei infection (data not shown).
3.4. IL-10 mediates CM protection
In order to assess the relevance of IL-10 in protection frompathology, IL-10 KO mice were challenged with P. berghei 60days after L. sigmodontis infection. Whereas wild type miceshowed the characteristic protection from CM after filarialinfection (Fisher’s exact test, p ¼ 0.0047. Fig. 4), IL-10 KOmice developed high CM rates like their P. berghei single-infected IL-10 KO counterparts. As a consequence, survival offilaria-infected wild type mice was significantly prolongedcompared to controls only infected with P. berghei (logranktest, p < 0.01) whereas the CM symptoms and survival rates ofpreviously filaria-infected IL-10 KO mice did not differ fromthat of P. berghei infected wild type and IL-10 KO mice.Parasitemia was investigated 6 days post P. berghei infection.
d with L. sigmodontis or left uninfected and then infected i.v. with 50,000 P.
quantification in the brain and spleen were quantified. (A) Absolute number of
resent mean � SD. Data were compared by one-way ANOVAwith the Tukey’s
cells in spleen. Columns represent mean � SD. Data were compared by one-
. In B and C, data are representative for two independent experiments.
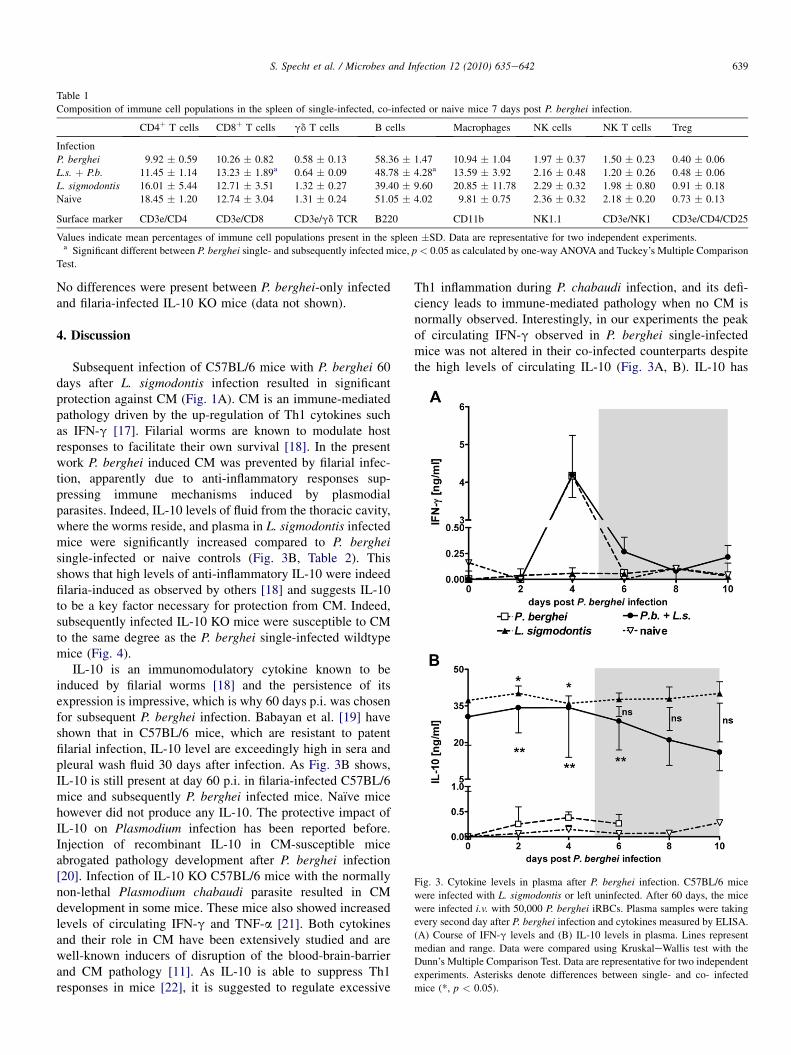
Table 1
Composition of immune cell populations in the spleen of single-infected, co-infected or naive mice 7 days post P. berghei infection.
CD4þ T cells CD8þ T cells gd T cells B cells Macrophages NK cells NK T cells Treg
Infection
P. berghei 9.92 � 0.59 10.26 � 0.82 0.58 � 0.13 58.36 � 1.47 10.94 � 1.04 1.97 � 0.37 1.50 � 0.23 0.40 � 0.06
L.s. þ P.b. 11.45 � 1.14 13.23 � 1.89a 0.64 � 0.09 48.78 � 4.28a 13.59 � 3.92 2.16 � 0.48 1.20 � 0.26 0.48 � 0.06
L. sigmodontis 16.01 � 5.44 12.71 � 3.51 1.32 � 0.27 39.40 � 9.60 20.85 � 11.78 2.29 � 0.32 1.98 � 0.80 0.91 � 0.18
Naive 18.45 � 1.20 12.74 � 3.04 1.31 � 0.24 51.05 � 4.02 9.81 � 0.75 2.36 � 0.32 2.18 � 0.20 0.73 � 0.13
Surface marker CD3e/CD4 CD3e/CD8 CD3e/gd TCR B220 CD11b NK1.1 CD3e/NK1 CD3e/CD4/CD25
Values indicate mean percentages of immune cell populations present in the spleen �SD. Data are representative for two independent experiments.a Significant different between P. berghei single- and subsequently infected mice, p < 0.05 as calculated by one-way ANOVA and Tuckey’s Multiple Comparison
Test.
Fig. 3. Cytokine levels in plasma after P. berghei infection. C57BL/6 mice
were infected with L. sigmodontis or left uninfected. After 60 days, the mice
were infected i.v. with 50,000 P. berghei iRBCs. Plasma samples were taking
every second day after P. berghei infection and cytokines measured by ELISA.
(A) Course of IFN-g levels and (B) IL-10 levels in plasma. Lines represent
median and range. Data were compared using KruskaleWallis test with the
Dunn’s Multiple Comparison Test. Data are representative for two independent
experiments. Asterisks denote differences between single- and co- infected
mice (*, p < 0.05).
639S. Specht et al. / Microbes and Infection 12 (2010) 635e642
No differences were present between P. berghei-only infectedand filaria-infected IL-10 KO mice (data not shown).
4. Discussion
Subsequent infection of C57BL/6 mice with P. berghei 60days after L. sigmodontis infection resulted in significantprotection against CM (Fig. 1A). CM is an immune-mediatedpathology driven by the up-regulation of Th1 cytokines suchas IFN-g [17]. Filarial worms are known to modulate hostresponses to facilitate their own survival [18]. In the presentwork P. berghei induced CM was prevented by filarial infec-tion, apparently due to anti-inflammatory responses sup-pressing immune mechanisms induced by plasmodialparasites. Indeed, IL-10 levels of fluid from the thoracic cavity,where the worms reside, and plasma in L. sigmodontis infectedmice were significantly increased compared to P. bergheisingle-infected or naive controls (Fig. 3B, Table 2). Thisshows that high levels of anti-inflammatory IL-10 were indeedfilaria-induced as observed by others [18] and suggests IL-10to be a key factor necessary for protection from CM. Indeed,subsequently infected IL-10 KO mice were susceptible to CMto the same degree as the P. berghei single-infected wildtypemice (Fig. 4).
IL-10 is an immunomodulatory cytokine known to beinduced by filarial worms [18] and the persistence of itsexpression is impressive, which is why 60 days p.i. was chosenfor subsequent P. berghei infection. Babayan et al. [19] haveshown that in C57BL/6 mice, which are resistant to patentfilarial infection, IL-10 level are exceedingly high in sera andpleural wash fluid 30 days after infection. As Fig. 3B shows,IL-10 is still present at day 60 p.i. in filaria-infected C57BL/6mice and subsequently P. berghei infected mice. Naıve micehowever did not produce any IL-10. The protective impact ofIL-10 on Plasmodium infection has been reported before.Injection of recombinant IL-10 in CM-susceptible miceabrogated pathology development after P. berghei infection[20]. Infection of IL-10 KO C57BL/6 mice with the normallynon-lethal Plasmodium chabaudi parasite resulted in CMdevelopment in some mice. These mice also showed increasedlevels of circulating IFN-g and TNF-a [21]. Both cytokinesand their role in CM have been extensively studied and arewell-known inducers of disruption of the blood-brain-barrierand CM pathology [11]. As IL-10 is able to suppress Th1responses in mice [22], it is suggested to regulate excessive
Th1 inflammation during P. chabaudi infection, and its defi-ciency leads to immune-mediated pathology when no CM isnormally observed. Interestingly, in our experiments the peakof circulating IFN-g observed in P. berghei single-infectedmice was not altered in their co-infected counterparts despitethe high levels of circulating IL-10 (Fig. 3A, B). IL-10 has
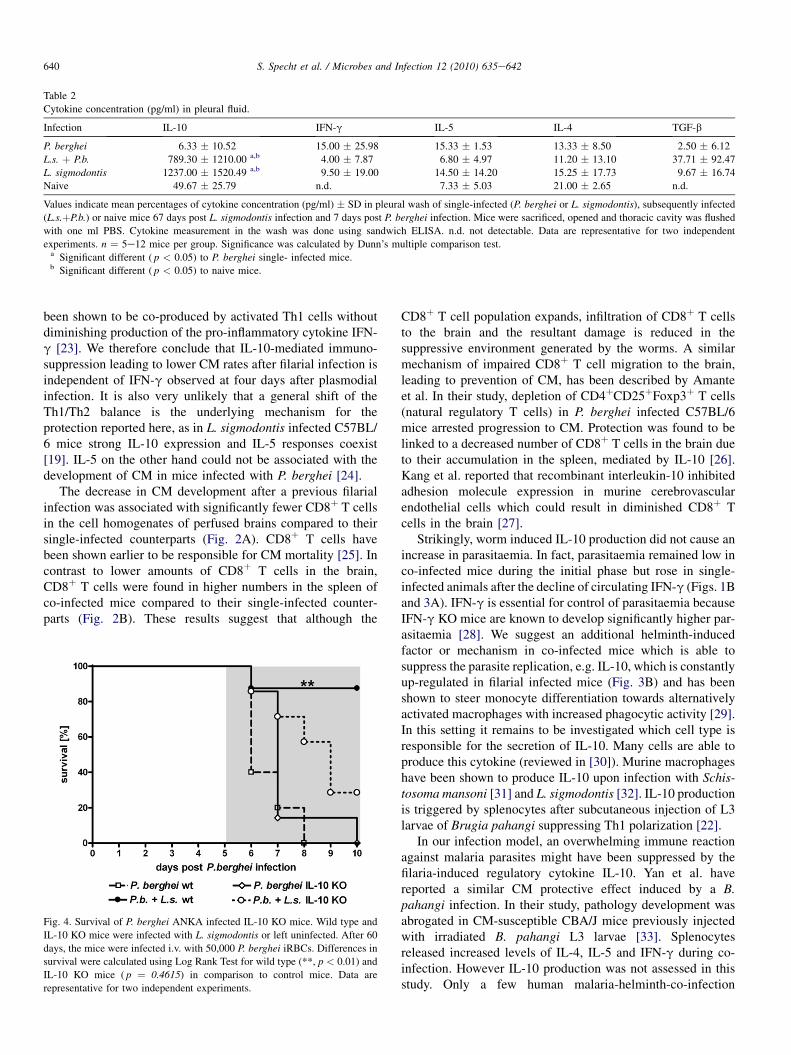
Table 2
Cytokine concentration (pg/ml) in pleural fluid.
Infection IL-10 IFN-g IL-5 IL-4 TGF-b
P. berghei 6.33 � 10.52 15.00 � 25.98 15.33 � 1.53 13.33 � 8.50 2.50 � 6.12
L.s. þ P.b. 789.30 � 1210.00 a,b 4.00 � 7.87 6.80 � 4.97 11.20 � 13.10 37.71 � 92.47
L. sigmodontis 1237.00 � 1520.49 a,b 9.50 � 19.00 14.50 � 14.20 15.25 � 17.73 9.67 � 16.74
Naive 49.67 � 25.79 n.d. 7.33 � 5.03 21.00 � 2.65 n.d.
Values indicate mean percentages of cytokine concentration (pg/ml) � SD in pleural wash of single-infected (P. berghei or L. sigmodontis), subsequently infected
(L.s.þP.b.) or naive mice 67 days post L. sigmodontis infection and 7 days post P. berghei infection. Mice were sacrificed, opened and thoracic cavity was flushed
with one ml PBS. Cytokine measurement in the wash was done using sandwich ELISA. n.d. not detectable. Data are representative for two independent
experiments. n ¼ 5e12 mice per group. Significance was calculated by Dunn’s multiple comparison test.a Significant different ( p < 0.05) to P. berghei single- infected mice.b Significant different ( p < 0.05) to naive mice.
640 S. Specht et al. / Microbes and Infection 12 (2010) 635e642
been shown to be co-produced by activated Th1 cells withoutdiminishing production of the pro-inflammatory cytokine IFN-g [23]. We therefore conclude that IL-10-mediated immuno-suppression leading to lower CM rates after filarial infection isindependent of IFN-g observed at four days after plasmodialinfection. It is also very unlikely that a general shift of theTh1/Th2 balance is the underlying mechanism for theprotection reported here, as in L. sigmodontis infected C57BL/6 mice strong IL-10 expression and IL-5 responses coexist[19]. IL-5 on the other hand could not be associated with thedevelopment of CM in mice infected with P. berghei [24].
The decrease in CM development after a previous filarialinfection was associated with significantly fewer CD8þ T cellsin the cell homogenates of perfused brains compared to theirsingle-infected counterparts (Fig. 2A). CD8þ T cells havebeen shown earlier to be responsible for CM mortality [25]. Incontrast to lower amounts of CD8þ T cells in the brain,CD8þ T cells were found in higher numbers in the spleen ofco-infected mice compared to their single-infected counter-parts (Fig. 2B). These results suggest that although the
Fig. 4. Survival of P. berghei ANKA infected IL-10 KO mice. Wild type and
IL-10 KO mice were infected with L. sigmodontis or left uninfected. After 60
days, the mice were infected i.v. with 50,000 P. berghei iRBCs. Differences in
survival were calculated using Log Rank Test for wild type (**, p < 0.01) and
IL-10 KO mice ( p ¼ 0.4615) in comparison to control mice. Data are
representative for two independent experiments.
CD8þ T cell population expands, infiltration of CD8þ T cellsto the brain and the resultant damage is reduced in thesuppressive environment generated by the worms. A similarmechanism of impaired CD8þ T cell migration to the brain,leading to prevention of CM, has been described by Amanteet al. In their study, depletion of CD4þCD25þFoxp3þ T cells(natural regulatory T cells) in P. berghei infected C57BL/6mice arrested progression to CM. Protection was found to belinked to a decreased number of CD8þ T cells in the brain dueto their accumulation in the spleen, mediated by IL-10 [26].Kang et al. reported that recombinant interleukin-10 inhibitedadhesion molecule expression in murine cerebrovascularendothelial cells which could result in diminished CD8þ Tcells in the brain [27].
Strikingly, worm induced IL-10 production did not cause anincrease in parasitaemia. In fact, parasitaemia remained low inco-infected mice during the initial phase but rose in single-infected animals after the decline of circulating IFN-g (Figs. 1Band 3A). IFN-g is essential for control of parasitaemia becauseIFN-g KO mice are known to develop significantly higher par-asitaemia [28]. We suggest an additional helminth-inducedfactor or mechanism in co-infected mice which is able tosuppress the parasite replication, e.g. IL-10, which is constantlyup-regulated in filarial infected mice (Fig. 3B) and has beenshown to steer monocyte differentiation towards alternativelyactivated macrophages with increased phagocytic activity [29].In this setting it remains to be investigated which cell type isresponsible for the secretion of IL-10. Many cells are able toproduce this cytokine (reviewed in [30]). Murine macrophageshave been shown to produce IL-10 upon infection with Schis-tosoma mansoni [31] and L. sigmodontis [32]. IL-10 productionis triggered by splenocytes after subcutaneous injection of L3larvae of Brugia pahangi suppressing Th1 polarization [22].
In our infection model, an overwhelming immune reactionagainst malaria parasites might have been suppressed by thefilaria-induced regulatory cytokine IL-10. Yan et al. havereported a similar CM protective effect induced by a B.pahangi infection. In their study, pathology development wasabrogated in CM-susceptible CBA/J mice previously injectedwith irradiated B. pahangi L3 larvae [33]. Splenocytesreleased increased levels of IL-4, IL-5 and IFN-g during co-infection. However IL-10 production was not assessed in thisstudy. Only a few human malaria-helminth-co-infection

641S. Specht et al. / Microbes and Infection 12 (2010) 635e642
studies considered IL-10 plasma levels in patients, with con-flicting results. In vitro data [34] revealed that whole blood ofhelminth-infected children in Ghana released more IL-10 thanthose of uninfected children in response to iRBC, whereas theINF-g levels were unaltered.
We have recently published that one third of filarial infectedBALB/cmice is completely protected from challengeP. bergheisporozoites. This sterilitywas dependent on IL-10 [35]. BALB/cmice develop a fully patent filarial infection including micro-filaremia, whereas they do not develop CM and succumb to P.berghei infection due to anemia. In C57 BL/6 mice filarialparasites do not develop to fecundity, but they do develop CM.Notably, protectivemechanisms in patent and non-patent modelsystems were dependent on IL-10, however acting on differentmalaria parasite and disease stages. This most likely reflects themultitude of mechanisms that can change the outcome ofa malaria infection and might explain the differences observedin humans.
In summary, a delicate equilibrium must be achieved toavoid CM: a Th1 reaction sufficient to restrain the parasites isnecessary, but an exaggerated response will drive CMpathology. Our data suggest that in murine P. berghei infec-tion, previous infection with L. sigmodontis is able to shift thehost immune response to establish such equilibrium. First,filaria-infected mice had significantly higher levels of circu-lating IL-10 that is able to prevent pathology, potentially byimpeding CD4þ and CD8þ T cell migration into the brain.Second, despite the high IL-10 levels, the production of IFN-gnecessary to kill the parasites was not reduced. It was recentlypublished in a study of Gambian children suffering fromPlasmodium falciparum malaria that those with uncomplicatedmalaria showed higher levels of a T cell population in bloodcoproducing IL-10 and IFN-g in contrast to children withsevere malaria [36]. This benefit of IL-10 and IFN-gproduction on malaria outcome in our mouse model seems toreflect the situation in humans. Thus, therapies that emulatethe immunosuppression induced by filarial nematode infec-tions could provide a tool to reduce the risk of malarial CM.
References
[1] Who, World Malaria Report 2008. World Health Organisation, Geneva,
2008.
[2] Who, Global Programme to Eliminate Lymphatic Filariasis. World
Health Organization, Geneva, 2009, pp. 437e444.[3] R. de Waal Malefyt, J. Abrams, B. Bennett, C.G. Figdor, J.E. de Vries,
Interleukin 10(IL-10) inhibits cytokine synthesis by human monocytes:
an autoregulatory role of IL-10 produced by monocytes. J Exp Med 174
(1991) 1209e1220.
[4] N.W. Brattig, U. Rathjens, M. Ernst, F. Geisinger, A. Renz, F.W.
Tischendorf, Lipopolysaccharide-like molecules derived from Wolbachia
endobacteria of the filaria Onchocerca volvulus are candidate mediators
in the sequence of inflammatory and antiinflammatory responses of
human monocytes. Microbes Infect 2 (2000) 1147e1157.
[5] S. Mahanty, M. Ravichandran, U. Raman, K. Jayaraman, V. Kumar-
aswami, T.B. Nutman, Regulation of parasite antigen-driven immune
responses by interleukin-10 (IL-10) and IL-12 in lymphatic filariasis.
Infect Immun 65 (1997) 1742e1747.
[6] S. Specht, L. Volkmann, T. Wynn, A. Hoerauf, Interleukin-10 (IL-10)
counter-regulates IL-4-dependent effector mechanisms in murine filari-
asis. Infect Immun 72 (2004) 6287e6293.
[7] W.H. Hoffmann, A.W. Pfaff, H. Schulz-Key, P.T. Soboslay, Determinants
for resistance and susceptibility to microfilaraemia in Litomosoides sig-
modontis filariasis. Parasitology 122 (2001) 641e649.
[8] S. Babayan, T. Attout, S. Specht, A. Hoerauf, G. Snounou, L. Renia, M.
Korenaga, O. Bain, C. Martin, Increased early local immune responses
and altered worm development in high-dose infections of mice suscep-
tible to the filaria Litomosoides sigmodontis. Med Microbiol Immunol
194 (2005) 151e162.
[9] L. Volkmann, O. Bain, M. Saeftel, S. Specht, K. Fischer, F. Brombacher,
K.I. Matthaei, A. Hoerauf, Murine filariasis: interleukin 4 and interleukin
5 lead to containment of different worm developmental stages. Med
Microbiol Immunol 192 (2003) 23e31.
[10] L. Schofield, G.E. Grau, Immunological processes in malaria patho-
genesis. Nat Rev Immunol 5 (2005) 722e735.
[11] N.H. Hunt, G.E. Grau, Cytokines: accelerators and brakes in the patho-
genesis of cerebral malaria. Trends Immunol 24 (2003) 491e499.[12] J. Nitcheu, O. Bonduelle, C. Combadiere, M. Tefit, D. Seilhean, D.
Mazier, B. Combadiere, Perforin-dependent brain-infiltrating cytotoxic
CD8 þ T lymphocytes mediate experimental cerebral malaria patho-
genesis. J Immunol 170 (2003) 2221e2228.[13] F.C. Hartgers, M. Yazdanbakhsh, Co-infection of helminths and malaria:
modulation of the immune responses to malaria. Parasite Immunol 28
(2006) 497e506.
[14] S. Specht, A. Hoerauf, Does helminth elimination promote or prevent
malaria? Lancet 369 (2007) 446e447.
[15] K.M. Al-Qaoud, A. Taubert, H. Zahner, B. Fleischer, A. Hoerauf,
Infection of BALB/c mice with the filarial nematode Litomosoides sig-
modontis: role of CD4 þ T cells in controlling larval development. Infect
Immun 65 (1997) 2457e2461.
[16] C.R. Engwerda, L. Beattie, F.H. Amante, The importance of the spleen in
malaria. Trends Parasitol 21 (2005) 75e80.[17] W. Rudin, N. Favre, G. Bordmann, B. Ryffel, Interferon-gamma is essential
for the development of cerebralmalaria. Eur J Immunol 27 (1997) 810e815.
[18] A. Hoerauf, J. Satoguina, M. Saeftel, S. Specht, Immunomodulation by
filarial nematodes. Parasite Immunol 27 (2005) 417e429.[19] S. Babayan, M.N. Ungeheuer, C. Martin, T. Attout, E. Belnoue, G.
Snounou, L. Renia, M. Korenaga, O. Bain, Resistance and susceptibility
to filarial infection with litomosoides sigmodontis are associated with
early differences in parasite development and in localized immune
reactions. Infect Immun 71 (2003) 6820e6829.
[20] S. Kossodo, C. Monso, P. Juillard, T. Velu, M. Goldman, G.E. Grau,
Interleukin-10 modulates susceptibility in experimental cerebral malaria.
Immunology 91 (1997) 536e540.
[21] C. Li, I. Corraliza, J. Langhorne, A defect in interleukin-10 leads to
enhanced malarial disease in Plasmodium chabaudi chabaudi infection in
mice. Infect Immun 67 (1999) 4435e4442.[22] J. Osborne, E. Devaney, Interleukin-10 and antigen-presenting cells
actively suppress Th1 cells in BALB/c mice infected with the filarial
parasite Brugia pahangi. Infect Immun 67 (1999) 1599e1605.
[23] S. Rutz, M. Janke, N. Kassner, T. Hohnstein, M. Krueger, A. Scheffold,
Notch regulates IL-10 production by T helper 1 cells. Proc Natl Acad Sci
U S A 105 (2008) 3497e3502.
[24] S. de Kossodo, G.E. Grau, Profiles of cytokine production in relation with
susceptibility to cerebral malaria. J Immunol 151 (1993) 4811e4820.
[25] E. Belnoue, M. Kayibanda, A.M. Vigario, J.C. Deschemin, N. van
Rooijen, M. Viguier, G. Snounou, L. Renia, On the pathogenic role of
brain-sequestered alphabeta CD8 þ T cells in experimental cerebral
malaria. J Immunol 169 (2002) 6369e6375.
[26] F.H. Amante, A.C. Stanley, L.M. Randall, Y. Zhou, A. Haque, K.
McSweeney, A.P. Waters, C.J. Janse, M.F. Good, G.R. Hill, C.R. Eng-
werda, A role for natural regulatory T cells in the pathogenesis of
experimental cerebral malaria. Am J Pathol 171 (2007) 548e559.
[27] H. Kang, P.Y. Yang, Y.C. Rui, Adenovirus viral interleukin-10 inhibits
adhesion molecule expressions induced by hypoxia/reoxygenation in
cerebrovascular endothelial cells. Acta Pharmacol Sin 29 (2008) 50e56.

642 S. Specht et al. / Microbes and Infection 12 (2010) 635e642
[28] Z. Su, M.M. Stevenson, Central role of endogenous gamma interferon in
protective immunity against blood-stage Plasmodium chabaudi AS
infection. Infect Immun 68 (2000) 4399e4406.
[29] M.M. Tiemessen, A.L. Jagger, H.G. Evans, M.J. van Herwijnen, S. John,
L.S. Taams, CD4þCD25þFoxp3þ regulatory T cells induce alternative
activation of human monocytes/macrophages. Proc Natl Acad Sci U S A
104 (2007) 19446e19451.
[30] A. O’Garra, P. Vieira, T(H)1 cells control themselves by producing
interleukin-10. Nat Rev Immunol 7 (2007) 425e428.
[31] P.O. Flores Villanueva, H. Reiser, M.J. Stadecker, Regulation of T helper
cell responses in experimental murine schistosomiasis by IL-10. Effect
on expression of B7 and B7-2 costimulatory molecules by macrophages.
J Immunol 153 (1994) 5190e5199.
[32] S. Specht, M. Saeftel, M. Arndt, E. Endl, B. Dubben, N.A. Lee, J.J. Lee,
A. Hoerauf, Lack of eosinophil peroxidase or major basic protein impairs
defense against murine filarial infection. Infect Immun 74 (2006)
5236e5243.
[33] Y. Yan, G. Inuo, N. Akao, S. Tsukidate, K. Fujita, Down-regulation of
murine susceptibility to cerebral malaria by inoculation with third-stage
larvae of the filarial nematode Brugia pahangi. Parasitology 114 (4)
(1997) 333e338.
[34] F.C. Hartgers, B.B. Obeng, Y.C. Kruize, A. Dijkhuis, M. McCall, R.W.
Sauerwein, A.J. Luty, D.A. Boakye, M. Yazdanbakhsh, Responses to
malarial antigens are altered in helminth-infected children. J Infect Dis
199 (2009) 1528e1535.[35] D. Fernandez Ruiz, B. Dubben, M. Saeftel, E. Endl, S. Deininger, A.
Hoerauf, S. Specht, Filarial infection induces protection against P. ber-
ghei liver stages in mice. Microbes Infect 11 (2009) 172e180.
[36] M. Walther, D. Jeffries, O.C. Finney, M. Njie, A. Ebonyi, S. Deininger,
E. Lawrence, A. Ngwa-Amambua, S. Jayasooriya, I.H. Cheeseman, N.
Gomez-Escobar, J. Okebe, D.J. Conway, E.M. Riley, Distinct roles for
FOXP3 and FOXP3 CD4 T cells in regulating cellular immunity to
uncomplicated and severe Plasmodium falciparum malaria. PLoS Pathog
5 (2009) e1000364.





























