Embed Size (px)
Citation preview

F A C U L T Y O F S C I E N C E
U N I V E R S I T Y O F C O P E N H A G E N
PhD thesis
Therése Oskarsson
Filamentous growth in Eremothecium fungi
Molecular characterization of the Ashbya gossypii ARF3 module
Academic advisors: Steen Holmberg, Department of Biology, University of Copenhagen
Co- supervisor: Jürgen Wendland, Carlsberg laboratory
Submitted: 13/03/2014

Institutnavn: Natur- og Biovidenskabelige Fakultet Name of department: Department of Biology Author: Therése Oskarsson Titel og evt. undertitel: Filamentous growth in Eremothecium fungi Title / Subtitle: Molecular characterization of the Ashbya gossypii ARF3 module Academic advisor: Professor Steen Holmberg Submitted: 2014-03-13 Grade:

”The world is a thing of utter inordinate complexity
and richness and strangeness that is absolutely awesome.”
Douglas Adams, 1952-2001

Preface
i
Preface
This thesis encompasses the results of three years of PhD studies at Carlsberg laboratory and the
Univerity of Copenhagen. My research was a part of the Ariadne project, funded by the Marie
Curie Programme and was mainly carried out at Carlsberg laboratory, Copenhagen, Denmark,
under the supervision of Prof. Jürgen Wendland and Prof. Steen Holmberg. Five weeks was also
spent working with the development of Eremothecium pathogenicity assays in the group of Prof.
Antonio di Pietro, University of Córdoba, Córdoba, Spain.
This thesis begins with a general introduction of fungal biology and the topics presented in this
thesis and the metods used during my research. Results and discussion of the three topics which
has been the focus my research are presented in three separate chapters, followed by a general
summary and future topics that could be adressed given more time. nnnnnnnnnnnnnnnnnnnnnnn

Contents
ii
Contents
PREFACE ........................................................................................................................ I
CONTENTS ..................................................................................................................... II
SUMMARY ..................................................................................................................... IV
RESUMÉ ......................................................................................................................... V
ACKNOWLEDGEMENTS .............................................................................................. VI
ABBREVIATIONS ........................................................................................................ VIII
CHAPTER 1: INTRODUCTION ..................................................................................... 10
1.1 Fungi as model organisms ................................................................................................................................... 10 1.1.1 Ashbya gossypii .............................................................................................................................................. 10 1.1.2 Eremothecium cymbalariae ............................................................................................................................ 13
1.2 Factors for polarized hyphal growth .................................................................................................................. 13 1.2.1 Dynamics of the apical Spitzenkörper ............................................................................................................ 13 1.2.2 Phosphoinositides ........................................................................................................................................... 15 1.2.3 The Actin cytoskeleton ................................................................................................................................... 15 1.2.4 Clathrin mediated endocytosis ....................................................................................................................... 17
1.3 The Ras superfamily of small GTPases .............................................................................................................. 21 1.3.1 The mechanism of GTP/GDP switching ........................................................................................................ 22 1.3.2 The Arf3 small GTPase and its regulators ...................................................................................................... 24
1.4 Focus and aim ....................................................................................................................................................... 26
CHAPTER 2: METHODS .............................................................................................. 28
2.1 Strains, media and growth conditions ................................................................................................................ 28 2.1.1 Plate assays ..................................................................................................................................................... 29 2.1.2 Pathogenicity assays ....................................................................................................................................... 29
2.2 Isolation of genomic DNA .................................................................................................................................... 30
2.3 PCR and cloning ................................................................................................................................................... 31 2.3.1 Generation of deletion cassettes ..................................................................................................................... 31 2.3.2 Vector generation by homologous recombination .......................................................................................... 32 2.3.3 Cloning and functional expression of truncated AgGts1 ................................................................................ 32 2.3.4 Construction of E. cymbalariae centromere plasmids .................................................................................... 33
2.4 Transformation .................................................................................................................................................... 33
2.5 Cytological staining and microscopy .................................................................................................................. 34

Contents
iii
CHAPTER 3: FUNCTIONAL ANALYSIS OF THE A. GOSSYPII ARF3-YEL1-GTS1 MODULE ....................................................................................................................... 35
3.1 Results ................................................................................................................................................................... 35 3.1.1 Generation of A. gossypii arf3 and yel1 mutants ............................................................................................ 35 3.1.2 Growth phenotypes and mycelial morphology ............................................................................................... 37 3.1.3 Actin and chitin localization ........................................................................................................................... 39 3.1.4 Endosome visualization using FM4-64 .......................................................................................................... 41 3.1.5 Truncation of AgGts1 ..................................................................................................................................... 44 3.1.6 AgGts1-GFP localization ................................................................................................................................ 48
3.2 Discussion .............................................................................................................................................................. 50 3.2.1 The A. gossypii Arf3-Yel1-Arf3 module ........................................................................................................ 50 3.2.2 AgGTS1-GFP localizes at sites of growth and septation ................................................................................ 52 3.2.3 Unregulated AgArf3 causes impaired endocytosis and morphological abnormalities .................................... 52 3.2.4 AgARF3-dependent and independent apical actin localization ....................................................................... 54
CHAPTER 4: EREMOTHECIUM PATHOGENICITY ASSAYS ..................................... 56
4.1 Results ................................................................................................................................................................... 56 4.1.1 Cellophane penetration assay ......................................................................................................................... 56 4.1.2 Fruit pathogenicity assay ................................................................................................................................ 57 4.2.3 Galleria mellonella killing assay .................................................................................................................... 58
4.2 Discussion .............................................................................................................................................................. 59 4.2.1 Characteristics of filamentous growth in Eremothecium fungi ...................................................................... 59 4.2.2 Eremothecium spores causes G. mellonella immune reaction ........................................................................ 60
CHAPTER 5: ESTABLISHING MOLECULAR TOOLS IN E. CYMBALARIAE ............ 61
5.1 Results ................................................................................................................................................................... 61 5.1.1 Establishing E. cymbalariae PCR based gene targeting ................................................................................. 61 5.1.2 Characterization of Ectec1 and ecym5230 ...................................................................................................... 64 5.1.3 Assessment of CEN and ARS function in E. cymbalariae ............................................................................. 66
5.2 Discussion .............................................................................................................................................................. 68 5.2.1 Establishing PCR-based gene targeting in E. cymbalariae ............................................................................ 68 5.2.2 Wt-like phenotypes of E. cymbalariae TEC1 and ECYM5230 ....................................................................... 69 5.2.3 A. gossypii centromeres are stable in E. cymbalariae..................................................................................... 70
CHAPTER 6: SUMMARY AND FUTURE PROSPECTS .............................................. 72
CHAPTER 7: REFERENCES ........................................................................................ 73
APPENDIX I: STRAINS ................................................................................................ 84
APPENDIX II: PRIMERS ............................................................................................... 85
APPENDIX III: PLASMIDS ............................................................................................ 87

Summary
iv
Summary
The fungal kingdom encompasses a diverse group of organisms some of which have a great
impact on human lives, either as domesticated benefactors or as human and crop pathogens.
Using the filamentous fungus Ashbya gossypii and its close relative Eremothecium cymbalariae
as model organisms, this thesis deals with some of the aspects of hyphal growth, which is an
important virulence factor for pathogenic fungi infecting both humans and plants.
Hyphal establishment through continuous polar growth is a complex process, requiring the
careful coordination of a large subset of proteins involved in polarity establishment and
maintenance, cytoskeleton dynamics and intracellular transport. The first part of this thesis
addresses the A. gossypii Arf3 small GTPase and its GEF- and GAP regulators; Yel1 and Gts1,
which has been implicated in polar growth in a wide range of organisms. We could demonstrate
that manipulations of the regulatory AgGts1 rendered A. gossypii strains with severe actin
localization, endocytic and morphological phenotypes, presumably due to unregulated AgArf3
activity. As a homolog of the extensively more studied mammalian ARF6, we hypothesize that
the continuous AgArf3 activity causes an abnormal accumulation of PI(4,5)P2 in the plasma
membrane, which in turn can be linked to the slow endocytic uptake of FM4-64, the sub-apical
actin localization and subsequently the atypical mycelial morphology observed in Aggts1
mutants. Furthermore, we demonstrated that in addition to the AgArf3 GAP-regulatory activity of
AgGts1, the protein could have additional actin organizing properties.
In the second and third part, this thesis addresses the use of A. gossypii and its relative E.
cymbalariae as model organisms for filamentous growth. A series of assays analyzed the
capability of Eremothecium genus fungi to invade and colonize both plant- and insect hosts. We
found that neither A. gossypii nor E. cymbalariae are able to penetrate any host tissue, and
although A. gossypii is classified as a plant pathogen it is strictly dependent on its insect vectors
for infection. In addition, we optimized a series of molecular tools for E. cymbalariae to enable a
faster and more efficient approach for genetic comparisons between Eremothecium genus fungi.

Resumé
v
Resumé
Svampe er en stor gruppe af organismer med stor mangfoldighed. Nogle arter af svampe har
påvirket mäniskors liv, enten som velgørere eller ved at forårsage sygdom på personer eller
afgrøder. Denne afhandling avvänder trådsvampen Ashbya gossypii og dens nære slægtning
Eremothecium cymbalariae som modelorganismer at analysere nogle aspekter involveret i trådet
vækst, en væsentlig faktor der bidrager til evnen af svampe til at forårsage sygdom hos både
personer eller planter.
Hyfer i trådsvampe dannes ved kontinuerlig polar vækst, en kompleks proces, der kræver
omhyggelig koordinering af et stort antal proteiner, der bidrager til etablering og vedligeholdelse
af polaritet. Den første del af denne afhandling omhandler ARF3 i A. gossypii og dens GFF og
GAP regulatorer, YEL1 og GTS1. Disse gener koder for proteiner der tidligere er identificeret i
polar vækst i forskellige organismer. Gennem manipulation af den regulatoriske AgGTS1 kan vi
påvise at ureguleret AgArf3 aktivitet forårsager store defekter i lokalisering af actin cytoskelet,
cellemorfologi og evnen til endocytose. Vores hypotese er, at hvis AgArf3 er en homolog af
pattedyrsproteiner Arf6, kan de have lignende funktioner i cellen. Baseret på funktion hos Arf6,
forårsager ureguleret AgArf3 aktivitet en unormal akkumulering af plasma membranbundet
PI(4,5)P2. Denne ændring i plasmamembraet kan igen knyttes til de fænotyper observerede i
Aggts1; langsom endocytose, subapikal lokalisering af aktin og unormal mycelmorfologi. Vi kan
også vise på ekstra funtioner af AgGts1 udover at regulere AgArf3 hvis proteinet påvirker
organiseringen af actin uafhængig af AgArf3.
Den anden og tredje del er afsat til brugen af A. gossypii og E. cymbalariae som
modelorganismer for trådformede vækst. I en række forsøg blev analyseret svampe evne til at
invadere og kolonisere forskellige vævstyper fra både plante-og dyreriget. Vi fandt, at ingen af de
testede svampe selvstændigt kan trænge ind i vævet i en værtsorganisme, og skønt A. gossypii er
klassificeret som et plantpatogen er afhængighed af et insekt vektor at inficere sin vært. For
yderligere at lette brugen af E. cymbalariae som en model organisme, har vi udviklet en serie af
molekylære værktøjer til hurtigere og mere effektive sammenligninger mellem svampe fra
slægten Eremothecium.

Acknowledgements
vi
Acknowledgements
I would like to express my deepest and sincerest thanks to all the people who have supported me
and in any way contributed to the completion of this thesis.
First of all, I would like to thank Professor Jürgen Wendland for giving me the opportunity to
join his lab at Carlsberg laboratory as a PhD student. It has been three interesting and challenging
years that I will never forget.
Thanks to Professor Steen Holmberg for being my academic advisor and for the support during
the completion of this thesis.
Klaus Lengeler, thank you for all the comments, discussions and support during the completion
of this manuscript. Also, thank you for always taking the time to answer any questions that has
arisen during these three years, no matter how time-consuming or trivial they might have been.
Andrea Walther, thank you for all the support during endless microscopy sessions for always
being available for comments and discussion during the progression of my project, and for
always being interested in new results and developments.
Thank you Lisa for sharing projects, office space and travelling time, working with you has
always been inspirational. And thank you for always being a good friend when I have needed
one.
In addition to the above mentioned persons, I would like to mention the other members of the
Carlsberg yeast group: Ana, Claudia, Davide, Jevgenia and Natalia. Thank you for creating the
culturally colorful, friendly environment in which we all shared a piece of life for a while. I will
miss you.n
To all the PIs, post docs and researchers involved in the Ariande project, thank you for
organizing such an interesting and exciting project and for all the effort put in to courses,
discussions and excursions. Special thanks to Professor Antonio di Pietro who invited me into
his lab in Cordóba, and to all the members of his lab who made me feel welcome during my stay
there.

Acknowledgements
vii
To my fellow PhD students: Clara, Elisabetta “Betta”, Elzbieta “Ellie”, Filomena, Katja,
Lisa, Mennat, Miriam, Pankaj, Sonia and Vikram. We started out as associates in a common
project, but after three years of sharing research, fortunes, troubles and humor you all have
become great friends and I wish you all the best!
In addition to the gratitude I owe all of those directly involved i this project, there are some
persons whom deserve my depest thanks on a more personal level: Anna, Josefin and Linda,
thank you for being the amazing friends that you are and for helping me to stay focused during
these challenging years.
Most importantly, I would like to thank my wonderful family, my father Jan-Erik, mother
Margareta, sisters Emelie and Frida and my sweet little nephew Thor. Whithout your love and
support this thesis would not have been possible.
Therése Oskarsson

Abbreviations
viii
Abbreviations
a a amino acid
ADP adenosine diphosphate
AFM Ashbya full media
Ag Ashbya gossypii
ARF ADP ribosylation factors (protein family)
ARF3 ADP ribosylation factor (gene)
ARS autonomously replicating sequence
bp base pairs
CDE centromeric DNA element
CEN centromere
CFW calcofluor white
CSM complete supplement mixture (media)
DIC differential interference contrast
Ec Eremothecium cymbalariae
EDTA ethylenediaminetetraacetic acid
ER endoplasmic reticulum
GAP GTPase activating protein
GDP guanosine diphosphate
GEF guanine nucleotide exchange factor
GFP green fluorescent protein
GLN glutamine-rich (protein domain)
GTP guanosine triphosphate
GTS1 glycine threonine serine repeat 1 (gene)
kb kilo (x1000) base-pairs
NPF nucleation-promoting factor
ORF open reading frame
PBS phosphate buffered saline
PCR polymerase chain reaction
PDB potato dextrose broth (media)

Abbreviations
ix
PEG poly-ethylene glycol
pFA plasmid for Functional Analysis
PH pleckstrin homology
PI phosphoinositide
PI(4,5)P2 phosphatidylinositol-4,5-bisphosphate
PRO proline-rich (protein domain)
RAS (protein superfamily)
Sc Saccharomyces cerevisiae
SDS sodium doedylsulphate
STE NaCl-Tris-EDTA
STM saccharose-TRIS-MgCl2
TE Tris-EDTA (buffer)
TEC1 transposon enhancement control 1 (gene)
TES 2-((1,3-dihydroxy-2-(hydroxymethyl)propanyl)amino)ethanesulfonic
acid
Tris tris(hydroxymethyl)aminomethane
UBA ubiquitinin associated (protein domain)
YEL1 yeast EFA6-like (gene)
WASP Wilscott-Aldrich syndrome protein
wt wild-type
X-Gal 5-bromo-4-chloro-3-indolyl- β-D-galactopyranoside
YPD yeast peptone dextrose (media)
YT yeast extract and tryptone (media)

Chapter 1: Introduction
10
Chapter 1: Introduction
1.1 Fungi as model organisms
The fungal kingdom encompasses a large and diverse group of organisms, spanning from single
cell yeasts to multicellular, filamentous molds and mushrooms. Approximately 800000 fungal
species are described today, some of which influence human lives in several ways (Hedges,
2002). Some fungi are beneficial for us; e.g. the yeast Saccharomyces cerevisiae, which was one
of the first organisms domesticated by humans. Today, S. cerevisiae is perpetually used in baking
and alcohol fermentation, and with the development of the fields of molecular biology and
biotechnology new functions for the yeast has emerged, both as a model organism and as a tool
for processing and developing biological compounds (Botstein and Fink, 2011).
Other fungi are pathogenic and affect us either directly by causing disease in primarily
immunocompromised individuals (Pfaller and Diekema, 2007), or indirectly by infecting cereal
and crops, significantly reducing the crop yield worldwide each year (Dean et al., 2012). Due to
the impact on human lives by pathogenic fungi, a significant research effort today is concentrated
pathogenic fungi, including this thesis. Mapping and characterizing conserved virulence factors
might not only result in increased understanding of the fungal infection process, but could also
render new targets for antifungal drug development (Botstein and Fink, 2011, Hedges, 2002,
Mustacchi et al., 2006).
1.1.1 Ashbya gossypii
The ascomycete Ashbya gossypii, also known as Eremothecium gossypii, is a haploid, strictly
filamentous fungus of the Eremothecium genus discovered by Ashby and Novell in 1926. Using
insects of the Pyrrhocoridae family, i.e. “cotton-stainers”, as vectors, A. gossypii acts as a plant
pathogen, causing stigmatomycosis in cotton and dry rot in citrus fruit (Ashby and Novell, 1926,
Dietrich et al., 2013). While the fungus had a significant agricultural impact at the time of
discovery, the development of modern day insecticides has more or less eliminated fungal
infection by insect vectors. Today, A. gossypii is primarily used in biotechnology due to its
Riboflavin overproducing properties (Kato and Park, 2012, Wendland and Walther, 2005a).

Chapter 1: Introduction
11
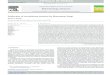
The A. gossypii life-cycle (Figure 1) is initiated by the germination of the needle-shaped
ascospore, which after germination forms multinucleated hyphae with compartments separated by
chitin-rich septa. Germination starts with isotropic growth, coupled with an increase of nuclei
through meiosis. After switching to polarized growth, juvenile mycelia is formed which is
characterized by lateral branching. As the hyphae mature, they switch to dichotomous hyphal
branching and increases growth speed from 6-10 µm/h to 200 µm/h (Wendland and Walther,
2005b, Knechtle et al., 2003). No sexual cycle has been identified in A. gossypii (Wasserstrom et
al., 2013), but spores are formed in sporangia of older hyphae, a process which is linked to the
overproduction of Riboflavin, also known as vitamin B2 (Kato and Park, 2012, Stahmann et al.,
2001).
Figure 1. The A. gossypii life cycle. The A. gossypii life cycle is initiated with a short period of isotropic growth (A), followed by the appearance of a primary germ tube along the gem bubble equatorial line. After the establishment of a second germ tube (B), the juvenile mycelia is characterized by lateral branching (C) and the growing hypha is divided into multinucleated compartments by chitin-rich septa. In the mature mycelia, growth speed increases 20-fold, and the hyphae switches from lateral- to dichotomous tip branching (D-E). Sporulation occurs in older hyphal compartments, a process linked to the overproduction of Riboflavin (F).

Chapter 1: Introduction
12
As a member of the Saccharomyces complex clade 12 (Figure 2), A. gossypii is a close relative of
S. cerevisiae (Kurtzman and Robnett, 2003). However, while the S. cerevisiae ancestor went
through a whole genome duplication event, the A. gossypii linage evolved a compact genome.
The completion of the A. gossypii genome sequencing in 2004 revealed a small 9.2 million bp
genome, with roughly 5000 genes located on seven chromosomes. In addition, the genome shows
only few duplications and lacks transposons and sub-telomeric repeats altogether. Despite the
differences in both phenotype and genome evolution between A. gossypii and S. cerevisiae, they
remain close relatives with a genetic functional homology of 95 %, and a high degree of synteny
(Dietrich et al., 2004, Prillinger et al., 1998). The close relationship to S. cerevisiae, together with
the development of efficient tools for molecular manipulations (Steiner et al., 1995, Wright and
Phillipsen, 1991, Wendland et al., 2000, Wendland and Walther, 2005a), A. gossypii has emerged
as an interesting model organism for studying evolution and filamentous growth in fungi.
Figure 2. Fungi of the Saccharomyces complex. Clade 1 and Clade 12, the Saccharomyces and Eremothecium clades, are highlighted in the phylogenetic tree. Modified from from Kurtzman and Robnett, 2003.

Chapter 1: Introduction
13
1.1.2 Eremothecium cymbalariae
Eremothecium cymbalariae, isolated by Borzi in 1888, is a filamentous ascomycete of the same
genus as A. gossypii. The E. cymbalariae genome was published in 2011, and reveals large
similarities to the A. gossypii genome. The 9.7 million bp encodes 4712 genes, of which 97 %
have an A. gossypii homolog and 95 % have a homolog in S. cerevisiae (Wendland and Walther,
2011, Kurtzman and Robnett, 2003).
E. cymbalariae grows as multinucleated hyphae with dichotomous hyphal tip branching, however
lacking the chitin-rich septa found in A. gossypii. As A. gossypii, no sexual cycle has been
identified in E. cymbalariae, but needle-shaped ascospores are produced in aerial sporangia:
However, it produces fewer spores than its relative, and also lacks the characteristic riboflavin
overproduction displayed in A. gossypii (Wendland and Walther, 2011).
1.2 Factors for polarized hyphal growth
Polar growth can be seen in a varying extent in all eukaryotic organisms. Unicellular organisms
perform short growth periods, e.g during cell division, while multicellular organisms use
continous polar growth for the development of specialized cell structures like roots, neurons and
the fungal hyphae. As the mechanism for polarity establishment is highly conserved, and hyphal
formation by continuous polar growth is a key feature for many pathogenic fungi, studying the
molecular processes that govern hyphal development might offer new targets for the development
of antifungal drugs. Discussed below are some of the most important factors for polarized
growth, and their impact on the development of the fungal hyphae.
1.2.1 Dynamics of the apical Spitzenkörper
Elongated cell structures like the fungal hyphae require a long-term maintenance of polarity,
requiring careful coordination of continuous delivery of membrane and cell wall material, up-
regulation of the polarisome complex and remodeling of the cytoskeleton (Figure 3). In the
fungal hyphae, a specialized structure called the Spitzenkörper organizes the addition of new
plasma membrane- and cell wall compounds by coordinating actin polarization and vesicle
trafficking at the hyphal apex (Grove and Bracker, 1970, Girbardt, 1957, Sudbery, 2011).
Regulated by small GTPases of the Ras superfamily (Park and Bi, 2007), secretory vesicles are

Chapter 1: Introduction
14
transported along the cytoskeletal microtubules from the Golgi apparatus towards the hyphal
apex (Howard and Aist, 1980, Fischer et al., 2008). At the Spitzenkörper, the vesicles switch
from microtubule tracks to actin filaments leading directly to the plasma membrane. The rapid
and dynamic polymerization of actin at the Spitzenkörper tip is critical for vesicle delivery and
maintenance of hyphal polarity and growth. Thus, an essential feature for the Spitzenkörper is the
polarisome protein complex, which mediates actin polymerization and forms a link between the
plasma membrane and the growing actin filament (Fischer et al., 2008, Uphadyay and Shaw,
2008).
Just below the hyphal apex and the Spitzenkörper is an area characterized by a high amount of
endocytic vesicle formation, detectable by an accumulation of actin patches (Taheri-Talesh et al.,
2008). Just as exocytosis is essential for delivering new material to the hyphal apex, the endocytic
machinery is thought to recycle and redistribute polarity-associated proteins from areas no longer
at the hyphal apex (Atkinson et al., 2002, Shaw et al., 2011).
Figure 3. Cytoskeleton- and organelle organization of the growing hyphae. Hyphal growth is mediated by a specialized structure, the Spitzenkörper, which organizes actin cable polymerization by the polarisome protein complex as well as transport of secretory vesicles to the plasma membrane. Polarity-associated membrane bound proteins are recycled by endocytosis, and endocytic vesicles are formed at an area close to the Spitzenkörper.

Chapter 1: Introduction
15
1.2.2 Phosphoinositides
Although a minor constituent of the plasma membrane, phosphoinositides (PIs) are a group of
phosphorylated lipids which has been found to have a wide range of cellular functions, including
acting as secondary messengers in signal transduction cascades, membrane anchors for various
proteins, organelle organization and vesicular trafficking. The regulatory role of PIs such as the
plasma membrane-associated PI(4,5)P2, is assigned to the irregular distribution throughout the
membranes. While PIs are synthesized from a at the associated membrane by specific inositide
kinases, and the kinase activity is in turn regulated by proteins of the Ras superfamily of small
GTPases (Mayinger, 2012, Oude Weernink et al., 2007, Balla, 2005, Brown et al., 2001)
As PI(4,5)P2 is located at the plasma membrane it is the PI most associated with polar growth. In
S. cerevisiae PI(4,5)P2, generated by the kinase Mss4, has been shown to mediate both the
assembly of clathrin coated endocytic vesicles and interactions between the plasma membrane
and the actin cytoskeleton (Desrivières et al., 1998, Gaidarov and Keen, 1999, Sun et al., 2007)
1.2.3 The Actin cytoskeleton
The microtubule and actin cytoskeletons are essential for many diverse features of an eukaryotic
cell, including vesicular transport and polar growth. Although the function of the microtubule
cytoskeletion is mostly assinged to nuclear- and organelle migration, it is also involved in polar
growth as the microtubule cytoskeleton functions as tracks for vesicle transport, linking the ER
and Golgi apparatus to sites of growth (Barnes et al., 1990, Lichius et al., 2011). In contrast, the
dynamic actin cytoskeleton is associated with actin cables and patches at the growing hyphal tip,
structures linked to short-distance vesicle transport and endocytosis. In addition, actin rings is
associate with the formation of septa throughout the hyphae (Evangelista et al., 2001, Wendland
and Walther, 2005b).
The assembly of the actin cytoskeleton is mediated by a large array of proteins, many of which
are regulated by membrane-bound PIs (Hilpelä et al., 2004). The most commonly involved in
actin nucleation are proteins of the formin family and the Arp2/3 complex (Figure 4). In S.
cerevisiae, formins mediate polymerization of unbranched actin cables used in vesicle transport
(Aghamohammadzadeh and Ayscough, 2010). As key components of the polarisome protein

Chapter 1: Introduction
16
complex, formins act by binding actin monomers and incorporation them to the growing filament
(Evangelista et al., 2001, Rida and Surana, 2005, Pollard, 2007). This formin-dependent actin
polymerization is regulated by small GTPases of the Ras superfamily, which activate formin
activity by blocking an auto-inhibitory binding between to formins (Li and Higgs, 2003, Wang et
al., 2009, Evangelista et al., 1997). In S. cerevisiae, the bestt studied formin regulator is the
conserved small GTPase Cdc42. In its active, GTP-bound state, Cdc42 associates with the plasma
membrane and recruits Spitzenkörper polarisome components, including the formin Bni1. Thus,
in addition to activation of the formin activity, Cdc42 acts as a linker between the polarisome and
the plasma membrane which enables direct targeting of the actin filament to the plasma
membrane (Evangelista et al., 1997, Chen et al., 2012).
Figure 4. Actin nucleation by formins and the Arp2/3 complex. (A) In the hyphal apex, polarisome associated formins mediate nucleation of unbranched actin filaments. (B) A branched actin network is assembled by the Arp2/3 complex assisted by associated NPFs such as the WASP-homolog Las17. Once activated, Arp2/3 anchors actin filaments to the preexisting actin network.
Cortical actin patches consists of clusters of branched actin filaments, primarily associated with
endocytosis (Pollard, 2007, Aghamohammadzadeh and Ayscough, 2010). The clustering of actin
filaments depend on the Arp2/3 complex, a conserved, seven subunit protein complex which
anchors new actin branches to a preexisting actin network. As the Arp2/3 complex has a low
intrinsic actin nucleation ability, actin assembly is activated by nucleation promoting factors,
NPFs (Machesky et al., 1994, Winter et al., 1999). Several NPFs have been identified in yeast so

Chapter 1: Introduction
17
far, including the strong WASP-like homolog Las17 and the weaker Abp1 and Pan1 (Lee et al.,
2000, Walther and Wendland, 2004b, Boettner et al., 2012, Machesky and Insall, 1998). Like
formins, NPFs are regulated by small GTPases of the Ras superfamily (Goley and Welch, 2006,
Bompard and Caron, 2004).
1.2.4 Clathrin mediated endocytosis
In eukaryotic organisms, vesicle trassport is not only a key festure for filamentous growth but
also essential for the biological functions of the cell. Intracellular transport is necessary for
transporting newly synthesized proteins between the ER and the Golgi apparatus for various post-
translational modifications before further distribution to the correct subcellular localization.
Exocytosis enables correct positioning of membrane bound proteins as well as the excretion of
soluble proteins into the extracellular fluid, necessary for cell-cell signaling. In turn, endocytosis
enables internalization of proteins and lipids from the cell surface, and is used to recycle plasma
membrane-bound proteins and to regulate their expression and localization (Figure 5) (Takai et
al., 2001, Kaksonen et al., 2003).
Figure 5. Intracellular protein traffic. Newly synthesized proteins are transported between the endoplasmic reticulum, Golgi apparatus and lysosomes for post-translational modifications and further to the correct subcellular localization. Endocytosis is used to recycle and control the localization of protein components of the plasma membrane.

Chapter 1: Introduction
18
Several different kinds of endocytosis have been identified, but the most studied and best
characterized is clathrin-mediated endocytosis which exists in all eukaryotic organisms. Still, the
clathrin-mediated endocytic machinery is very complex, requiring the timing and coordination of
a large number of proteins required for cargo sorting, membrane invagination and vesicle scission
(Figure 6) (McMahon and Boucrot, 2011, Kaksonen et al., 2003). Most of the key genes
encoding S. cerevisiae endocytic proteins have homologues in A. gossypii, (Table 1) and for this
reason the endocytic machinery is thought to function in a similar manner for both species.
Figure 6. Organization of the components of clathrin mediated endocytosis. Early endocytic factors Syp1 and Ede1, and clathrin subunits assemble at the site of endocytosis. While Syp1 causes formation of a membrane curvature, Ede1 recruits other proteins of the endocytic complex. Coat proteins like the Sla1-End3-Pan1 complex mediate the assembly of the clathrin coat, and form a link between the forming vesicle and the actin cytoskeleton through the NFP Pan1. Assembly of the actin filament network by the Arf2/3 complex mediated further membrane invagination and the formin vesicle is released from the plasma membrane by accumulation of amphiphysins. Once the scission event has released the vesicle, the clathrin coat is disassembled, an event regulated by several proteins including small GTPases of the Ras superfamily
In S. cerevisiae, nucleation is initiated by the early arriving BAR-protein Syp1 and the EH-
domain protein Ede1, which binds to the phosphatidylinositol-containing plasma membrane and
cause a membrane curvature (Heath and Insall, 2008, Antonescu et al., 2011) and enables
interaction with other proteins in the endocytic pathway (Confalonieria and Di Fiorea, 2002,
McMahon and Boucrot, 2011). Also arriving at this time point is the clathrin heavy- and light
chain. The subunits interact to form clathrin penta- and hexameres which serve as building blocks

Chapter 1: Introduction
19
for the clathrin coat (Figure 7) (Ungewickell and Branton, 1981, Crowther and Pearse, 1981,
Higgins and McMahon, 2002).
Table 1. S. cerevisiae clathrin-mediated endocytic proteins and their A. gossypii homologues. The S.
cerevisiae actin-clathrin linker Sla2 does not have a homologue in A. gossypii. S. cerevisiae proteins A. gossypii homologues Function(s) in endocytosis
Early nucleation
Ede1 ABR149W Protein-protein interactions
Syp1 AEL147W Membrane curvature
Chc1 AER359W Clathrin heavy chain
Clc1 AGR309C Clathrin light chain
Mid- to late nucleation and coat assembly
Sla2 - Actin-Clathrin linker
Sla1 AGR170C Actin assembly, Sla-End3-Pan1 complex
End3 AER416C Actin assembly, Sla-End3-Pan1 complex
Pan1 ADR018C Actin assembly, Sla-End3-Pan1 complex
Yap1801 AEL209W Clathrin adaptor, Pan1 interaction
Yap1802 AEL209W Clathrin adaptor, Pan1 interaction
Ent1 ACL157C Clathrin adaptor, Actin assembly
Ent2 ACL157C Clathrin adaptor, Actin assembly
Las17 AGR285W Nucleation-promoting factor
Myo3 AEL306C Nucleation-promoting factor
Myo5 AEL306C Nucleation-promoting factor
Abp1 AGL237C Nucleation-promoting factor
Arp2 ADR316W Actin filamentation
Arp3 AFR419C Actin filamentation
Scission
Rvs161 AER193W Actin filamentation
Rvs167 AFR140C Actin filamentation
Vps1 ABL001W GTPase, Actin organization
Dnm1 AAL174C GTPase
Slj2 (Inp52) AFL228W Polyphosphatidylinositol phosphatase
Uncoating
Ark1 ADL217W Kinase
Prk1 ADL217W Kinase
Arf3 ACL078W GTPase
Gts1 ACL055W ArfGAP
Lsb5 AFR709C Actin filamentation, Pan1 disassembly

Chapter 1: Introduction
20
Figure 7. Structure and assembly of the clathrin coat. Clathrin trisklerions (A) are composed of three heavy chains and three light chains. The trisklerions polymerize to form penta- and hexamere units (B) which build the clathrin coat in the forming vesicle (C).
In mammals, the AP2 complex plays a central role in clathrin coat formation and cargo selection.
In contrast, although the AP2 complex is part of yeast endocytosis, it is not essential (Schmid,
1997, Yeung et al., 1999). Instead, continuous clathrin recruitment to the forming vesicle is
dependent on the clathrin adaptors Yap1801/2 and Ent1/2 (Wendland et al., 1999a, Stahelin et al.,
2003). As the coat protein Sla2 arrives at the nucleation site, it functions as a linker between the
plasma membrane, the clathrin coat and the NPF Pan1 of the Sla1-End3-Pan1 complex (McCann
and Craig, 1997, Tang et al., 2000). Further membrane invagination at the site of nucleation is
driven by accumulation of actin filaments assembled by the Arp2/3 complex, which is activated
by several NPFs present at the endocytic site (Boettner et al., 2012, Aghamohammadzadeh and
Ayscough, 2010).
Scission of the newly formed vesicle from the plasma membrane is achieved by the
amphiphysins Rvs161 and Rvs167 (Youn et al., 2010), together with Vps1 and Dmn1, related to
mammalian dynamin (Smaczynska-de Rooij et al., 2010, Boettner et al., 2012, Gammie et al.,
1995, McMahon and Boucrot, 2011). Once the scission event releases the vesicle from the cell
membrane, the clathrin coat is disassembled. Several of the coat proteins, including the Pan1-
Sal1-End3 complex and the Ent1/2 clathrin adaptors, are regulated by phosphorylation. Thus,
several proteins implicated in endocytic vesicle uncoating are kinases, including Ark1 and Prk1
or phosphatases, like synaptojanins and small GTPases (Toret et al., 2008, Costa and Ayscough,
2005). Actin filaments protruding from the newly formed vesicle forms a link to the actin
cytoskeleton, which is used as a track for transportation throughout the cell (Toshima et al.,
2006, Qualmann and Kessels, 2002)

Chapter 1: Introduction
21
1.3 The Ras superfamily of small GTPases
In the 1980s, the first small GTPases of the Ras superfamily, Ha-Ras and Ki-Ras, were identified
as human oncogenes (Takai et al., 2001). Since then, a large number of small GTPases, varying
between 20-40 kDa, has been identified in various eukaryotic species spanning mammals, insects,
plants and fungi. The characterization of the small GTPases implements them in diverse cellular
processes such as vesicular traffic, cell signaling, cytoskeleton organization and gene expression.
Since their discovery, the over 150 members of the Ras superfamily have been divided into four
sub-branches depending on sequence and biological function: Rho, Rab, Ras and Arf (Table 2)
(Takai et al., 2001, Wennerberg et al., 2005).
Table 2. Proteins of the Ras superfamily
Ras sub-family Protein function(s) No. genes
Yeast Mammalia
Arf Intracellular traffic 7 16
Rab Intracellular traffic 11 42
Ran Nucleocytoplasmic transport
Cytoskeleton organization 2 1
Ras Gene expression 4 19
Rho Gene expression
Cytoskeleton organization 6 19
Small GTPases function as regulators of cellular processes by switching between an active and
inactive state in a GTP-dependent manner. A common feature for all small GTPases is the
GTP/GDP binding consensus sequence, the G-domain, which renders a high affinity for binding
GTP and GDP (Bourne et al., 1991). The GTPase cycling between the active GTP-bound state
and the inactive GDP-bound state is a naturally slow process, but the cycling is enhanced by two
groups of regulatory proteins: Guanine nucleotide exchange factors (GEFs) and GTPase
activating proteins (GAPs). The inactive small GTPase is activated by disassociation of the
bound GDP, catalyzed by the regulatory GEF (Rensland et al., 1991, Shapiro et al., 1993). The
subsequent binding of GTP causes a conformational change of the protein, allowing interaction
between the small GTPase and the downstream effector. In turn, the small GTPase is inactivated
by a rate-limiting hydrolysis of GTP. This reaction is regulated by GAPs, which increase the
intrinsic GTPase, promoting the transition into the inactive, GDP bound form (Figure 8) (Trahey
and McCormick, 1987, Takai et al., 2001).

Chapter 1: Introduction
22
Figure 8. Illustration of small GTPase switching between active and inactive states. An upstream signal triggers GEF aided GDP-GTP exchange, causing a conformational change which activates the GTPase. In turn, the small GTPase is inactivated by GAP-assisted hydrolysis of GTP to GDP and Pi.
1.3.1 The mechanism of GTP/GDP switching
The 166 residue, 20 kDa G-domain of guanine nucleotide binding proteins like the small
GTPases of the Ras superfamily, is highly conserved and the mechanism of GTP/GDP binding is
similar for all proteins. The G-domain structure consists of five α-helices organized around a six
stranded β-sheet. The guanine nucleotide binding properties arise from five conserved sequence
motifs (Figure 9) (Bourne et al., 1991). While the N/TKXD and GXXXXGKS/T motifs bind any
nucleotide phosphate (Saraste et al., 1990), the aspartic acid side chain of the DXXG motif and
the alanine of the S/CAK/L/T motif render the guanine nucleotide specificity (Zhong et al.,
1995). Two areas in the G-domain are particularly prone to conformational changes when
interacting with GTP/GDP. The conformation of these areas, named Switch I and Switch II, are
very similar in all guanine nucleotide binding proteins when bound to GTP, whereas the structure
varies to some extent in the GDP bound state (Corbett and Alber, 2001, Vetter and Wittinghofer,
2001). In the activated G-domain, the GTP is fixated by hydrogen bonds between GTP-oxygen
and NH-groups in threonine and glycine residues in Switch I and II, respectively (Vetter and
Wittinghofer, 2001).

Chapter 1: Introduction
23
Figure 9. Conserved sequence elements for the guanine nucleotide binding properties of the G-domain. Sequence elements I and IV enable a general affinity for nucleotide phosphates, while elements III and V renders guanine nucleotide specificity.
The slow GDP release from the G-domain is catalyzed by GEFs (Figure 10A). Different families
of GEFs may be structurally unrelated, but they all catalyze the release of GTP in a similar
manner (Cherfils and Chardin, 1999). By forcing a structural change of the G-domain through
Switch I and II interaction, the bonds between the G-domain P-loop, the GTP and the magnesium
ion is inhibited and the GDP is released (Kawashima et al., 1996, Vetter and Wittinghofer, 2001,
Cherfils and Chardin, 1999). A new guanine nucleotide, together with a Mg2+
ion, rapidly
replaces the GDP once the GEF has dissociated from the G-domain. (Zurita et al., 2010).
The process of GTP-to-GDP conversion varies between protein families. While G-domains of the
G-protein family appear to be catalytically active due to an intrinsic arginine residue (Mittal et
al., 1996), G-domain hydrolysis of GTP in the RAS superfamily is an approximately hundred-
fold slower process (Rensland et al., 1991, Shapiro et al., 1993, Bourne et al., 1991). In order to
rapidly cancel G-domain protein signaling, the speed of hydrolysis of GTP to GDP is enhanced
up to 105-fold by GAP regulators (Gideon et al., 1992). The GAP-dependent switch from the
active, GTP–bound state to the inactive GDP-bound state is a coordinated event which requires
an Mg2+
ion to interact with the oxygen atoms of two water molecules, two phosphoryl groups of
the bound GTP, an α-helix serine residue and a Switch I threonine residue (Figure 10C) (Zurita et
al., 2010, Vetter and Wittinghofer, 2001). As the GAP regulator interact with Switch II, a Switch
II glycine residue is repositioned and a GAP arginine finger is placed at the GTPase active site.
The positively charged arginine triggers the displacement of one water molecule from the active
site, creating an electrostatic field which catalyzes the hydrolysis of GTP to GDP and Pi
(Scheffzek et al., 1996, Mittal et al., 1996, te Heesen et al., 2007).

Chapter 1: Introduction
24
Figure 10. Mechanism of G-domain GTP/GDP switching. (A) Small GTPases are activated by its corresponding GEF. The GEF forces a conformational change in the G-domain Switch regions and P-loop, which breaks the binding of the GDP and magnesium ion. (B) Free cytosolic GTP associates with the small GTPase through interactions between the GTP, Switch I and II, the G-domain P-loop and one magnesium ion. (C) Hydrolysis of bound GTP to GDP is mediated by the regulatory GAP protein, which inserts a catalytically active arginine finger in the G-domain active site. The charged arginine triggers a magnesium-dependent hydrolysis reaction resulting in GDP and Pi. Modified from Vetter and Wittinghofer, 2001.
1.3.2 The Arf3 small GTPase and its regulators
ADP-ribosylation factors, Arfs, are a group of proteins within the Ras superfamily of small
GTPases. Many Arfs are key regulators during different stages of intracellular membrane
trafficking and they have been implicated in regulation of both clathrin and COPI-coated vesicle
formation and trafficking between the ER, the Golgi apparatus and the plasma membrane (Spang
et al., 2010, D’Souza-Schorey and Chavrier, 2006, East and Kahn, 2011). One of the best studied
Arfs is the mammalian Arf6, which has been demonstrated to regulate endocytic vesicle
formation and actin cytoskeleton organization (Toret et al., 2008). The regulatory properties of
Arf6 are thought to be partly mediated by activation of enzymes responsible for increased
membrane content of PI(4,5)P2 (Honda et al., 1999, D’Souza-Schorey and Chavrier, 2006). In
turn, PI(4,5)P2 is known to effect clathrin-mediated endocytosis, capping of actin filaments and
several actin binding proteins (Wenk and De Camilli, 2004, Hilpelä et al., 2004, Antonescu et al.,
2011). Other possible effectors of Arf6 are the clathrin adaptor AP2, kinases implemented in
vesicle scission and the Rac1 GTPase (D’Souza-Schorey and Chavrier, 2006).
S. cerevisiae Arf3, homologous to mammalian Arf6 is not as extensively studied as its
mammalian counterpart, and the precise role of Arf3 is unclear (Lee et al., 1994). Initial
characterization has identified Arf3 as a non-essential protein in polarity establishment rather

Chapter 1: Introduction
25
than endocytosis (Huang et al., 2003). However, later studies have shown that Arf3 might be
involved in endocytosis by increasing the plasma membrane content of PI(4,5)P2 (Smaczynska-
de Rooij et al., 2008) and through recruitment of the coat protein Lsb5 (Costa and Ayscough,
2005, Toret et al., 2008). There is also indications that Arf3, like mammalian Arf6, might have a
role in actin assembly (Lambert et al., 2007). A characteristic feature for the S. cerevisiae Arf3, is
an N-terminal glycine myristoylation (Huang et al., 2003), which enables Arf3-membrane
interaction. In the GDP-bound Arf3, the N-terminal helix is folded into the G-domain, while
conformational changes during GDP/GTP switching causes a delocalization of the myristoylated
helix enabling helix-membrane interaction (Itzen and Goody, 2011, Antonny et al., 1997).
Like all small GTPases, Arf3 cycles between an active, GTP-bound and an inactive, GDP- bound
state, aided by specific GEF and GAP regulators. Yel1 has been identified as the sole Arf3-GEF
(Figure 11), and displays a significant C-terminal homology to Efa6, a GEF of mammalian Arf6.
(Gillingham and Munro, 2007a, Franco et al., 1999). The Sec7-domain of Ye1l is a common
feature for all ArfGEFs, and catalyzes nucleotide exchange through interactions with the Arf
Switch I and Switch II regions. The structural change of the Arf G-domain enables the GEF
Sec7-domain glutamic finger to compete electrostatically with the bound GDP, thereby
efficiently ejecting it from the G-domain altogether (Casanova, 2007, Renault et al., 2003). In
addition to the Sec7 domain, Yel1 contains a C-terminal PI(4,5)P2–interacting PH domain, which
is thought to mediate Yel1 localization at the plasma membrane (Casamayor and Snyder, 2002,
Gillingham and Munro, 2007a).
Figure 11. Domain organization of S. cerevisiae Arf3, Yel1 and Gts1.

Chapter 1: Introduction
26
Arf hydrolysis of GTP to GDP is catalyzed by specific GAPs, defined by the presence of the
ArfGAP domain: a four cysteine zinc-finger motif, terminating with a catalytic arginine residue
(CX2CX16CX2CX4R) (Cukierman et al., 1995). There is a general idea that ArfGAPs may not
only function as regulators of Arf activity, but also downstream effectors of Arf (Spang et al.,
2010). Indeed, Gts1, the only GAP identified for S. cerevisiae Arf3, was initially characterized as
a clock gene, and has since then been implicated in several cellular processes including
oscillation of energy metabolism, clathrin mediated endocytosis and flocculation (Yaguchi et al.,
2007, Bossier et al., 1997, Xu and Tsurugi, 2007). Most GAPs have additional domains, and the
domain organization between different GAPs can vary significantly. The S. cerevisiae Gts1 has
two domains downstream of the catalytic ArfGAP; the central ubiquitinin-associated, UBA,
domain and the C-terminal glutamine-rich, GLN, domain. The functions of the UBA and GLN
domains in Gts1 has not previously been described, although UBA-motifs are usually
implemented in either binding of ubiquitinin directly, or binding of ubiquitinated proteins
(Hofmann and Bucher, 1996).
1.4 Focus and aim
This thesis is part of a project within the Marie Curie Initial Training Network, Ariadne, aimed at
identifying and characterizing conserved fungal virulence factors which might pose targets for
the development of new antifungal drugs. While research in the consortium spans human- and
plant pathogens, this thesis is mainly focused on the use of Ashbya gossypii as a model organism
for filamentous fungi due to the molecular tools readily available for manipulation of the A.
gossypii genome.
Hyphal growth is a key virulence factor for many pathogenic fungi, and several steps of hyphal
formation and growth are regulated by members of the conserved Ras superfamily of small
GTPases. For this purpose, this thesis aims to characterize and analyze the interactions of the A.
gossypii homolog of the S. cerevisiae small GTPase ARF3, and its regulators, YEL1 and GTS1.
Previous studies in S. cerevisiae have implemented Arf3 in establishment of polarity, actin
assembly and endocytosis, all important factors for filamentous growth. However, the precise
function and mode of action of Arf3 is not clear, and the use of a filamentous fungus might give
additional clues to how Arf3 regulates polarity and hyphal growth.

Chapter 1: Introduction
27
To enable a more correct comparison between A. gossypii and the fungal pathogens targeted by
the Ariadne project, another aim of this thesis has been the development of pathogenicity assays
to establish if, and how Eremothecium fungi might invade host tissue. To enable further
investigation of the Eremothecium genus, the final part of this thesis addresses the development
of molecular tools for E. cymbalariae, a close relative to A. gossypii.nnnnnnnnnnnnnnnnnnnnnnn

Chapter 2: Methods
28
Chapter 2: Methods
2.1 Strains, media and growth conditions
All strains, primers and plasmids used and generated in these experiments are listed in Appendix
I, Appendix II and Appendix III, respectively.
A. gossypii leu2 was used as background strain for gene deletion, and served as wt control during
phenotyping of mutant strains. All A. gossypii strains were grown at 30 °C in AFM media,
consisting of 2 % glucose, 1 % yeast extract, 1 % casein peptone and when necessary 2 % agar
for solidification. For antibiotic selection, 200 µg/ml G418/Geneticin and/or 50 µg/ml ClonNat
was used. A. gossypii was sporulated in CSM minimal medium supplemented with 0.1 % myo-
inositol consisting of 2 % glucose, 0.17 % yeast nitrogen base w/o amino acids and w/o
ammonium sulphate, 0.069 % CSM with 0.1 % asparagine, and when necessary 2 % agar for
solidification and antibiotic selection as described above. After 5-7 days, the mycelia were
degraded by incubation at 37 °C in 1x TE buffer (10 mM Tris, 1 mM EDTA) with 1 mg/ml
Zymolyase, and the spores were harvested by centrifugation and resuspension in 0.03 % Triton
X-100 spore buffer.
DBVPG7215 was used for optimization of gene deletion, plasmid stability- and pathogenicity
assays in Eremothecium cymbalariae. All E. cymbalariae strains were cultivated in the same
media and under the same conditions as A. gossypii. E. cymbalariae was sporulated on AFM agar
plates. After 5-7 days at 30 °C, the spores were harvested by mechanical disruption of the
sporangia. 10 ml 0.03 % Triton X100 spore buffer was added to the over-grown agar plate and
the sporangia were disrupted using a sterile spatula, releasing the spores into the spore buffer.
The spore buffer and the spores were collected and the spores were harvested by centrifugation
before immediate use.
Saccharomyces cerevisiae BY4741 was used for pAG marker exchange and GFP integration by
homologous recombination. In all cases, S. cerevisiae was grown on YPD media, consisting of 2
% glucose, 1 % yeast extract, 2 % casein peptone and when necessary 2 % agar for solidification.
For antibiotic selection, 200 µg/ml G418/Geneticin and/or 50 µg/ml ClonNat was used.

Chapter 2: Methods
29
Fusarium oxysporum f. sp lycopersici strain 4287 (FGSC 9935) was used as a positive control in
Eremothecium genus pathogenicity assays. F. oxysporum microconidia was obtained from
cultures grown in PDB as described elsewhere (di Pietro and Roncero, 1998). The spores were
obtained by sterile filtration and used immediately after isolation.
E. coli DH5α was used for cloning and propagation of plasmids used in this study. E. coli was
grown in 2YT media containing 1.6 % tryptone peptone, 1% yeast extract, 0.5 % NaCl and when
necessary 2 % agar for solidification. For antibiotic selection, 100 µg/ml Ampicillin or 50 µg/ml
Kanamycin was used. Propagated plasmids were isolated using the PureYieldTM
Plasmid
Midiprep System (Promega) (Ish-Horowicz and Burke, 1981).
2.1.1 Plate assays
Phenotypic analysis of colony growth and temperature stress was performed by spotting fresh
mycelia samples at the marked center of AFM agar plates with appropriate selection. Plates were
incubated at temperatures ranging from 20 °C to 37 °C for 7 days, after which the colony
diameter was measured. For each assay, were combined and summarized as average growth for
each genotype and temperature. For each sample growth condition, the standard deviation was
calculated according to the formula: - 2/(n-1).
2.1.2 Pathogenicity assays
The ability of Eremothecium fungi to penetrate physical barriers and colonize various tissues was
assayed using artificial media as well as fruit- and animal tissue. The ability to penetrate a
physical barrier was assayed during optimal conditions in artificial AFM- or CSM agar media.
Circular cellophane sheets, approximately the size of a petri dish, were prepared by autoclaving
in distilled water. Under sterile conditions, a single cellophane sheet was transferred to solidified
agar plates, after which 5 µl spore suspension was inoculated at the center of the cellophane
sheets and inoculated plates were incubated at 30 °C for 2-6 days. After removal of the
cellophane sheet, the plate was incubated for an additional 2-3 at 30 °C in order to evaluate
mycelial regrowth.

Chapter 2: Methods
30
Fruit pathogenicity assays were performed on fresh, undamaged apples and oranges. The fruit
were sterilized in 70% ethanol for 5 min, and under sterile conditions the fruit was cut into 0.5
mm slices and placed in petri dishes. For the apple assay, the exposed fruit tissue was inoculated
with 5µl spore suspensions from A. gossypii, E. cymbalariae and F. oxysporum. For the orange
assay, 5 µl spore suspension from the three species was inoculated separately in the exocarp,
mesocarp and endocarp to assess penetration of different parts of the orange tissue. The samples
were incubated at 30 °C and 100 % humidity. After 7 days, the colony growth on each fruit slice
was assessed. Tissue from the downward facing surface of the fruit slice was examined by
microscopy for the presence of mycelia.
The larvae of the greater wax moth, Galleria mellonella, were used for assaying the ability of
Eremothecium fungi to colonize animal tissue. A. gossypii, E. cymbalariae and F. oxysporum
spores were injected into Galleria mellonella larvae according to a protocol described elsewhere
(Navarro-Velasco et al., 2011). Before injection the spores were suspended in PBS to a density
which could pass the syringe. Heat-killed F. oxysporum microcornidia and A. gossypii spores,
generated by 1 h incubation at 65 °C, were used as negative control. The G. mellonella larvae
were incubated at 37 °C and scored daily as healthy (no melanization), sick (moderate
melanization) or dead (intense melanization and non-responsive to physical stimuli).
2.2 Isolation of genomic DNA
Genomic DNA was obtained by alkaline extraction and isopropanol precipitation (Birnboim and
Doly, 1979). The cell wall was digested by incubation at 37 °C in STE buffer (1 M Sorbitol, 50
mM Tris, 100 mM EDTA) with 1 mg/ml Zymolyase and 50 µg/ml RNAse A. Proteins were
denatured by addition of SDS to a final concentration of 1 % followed by incubation at 65 °C for
30 min. Potassium acetate to a final concentration of 0.64 M, followed by 30 min incubation on
ice. Cell debris was removed from the supernatant by centrifugation, and isopropanol was used
to precipitate the DNA. For recovery of plasmids from S. cerevisiae and Eremothecium fungi, the
genomic DNA was subsequently transformed into E. coli DH5α by electroporation.
For isolation of larger amounts of A. gossypii and E. cymbalariae DNA, vacuum filtered mycelia
were frozen using liquid nitrogen and pulverized using a mortar. The pulverized mycelia was

Chapter 2: Methods
31
dissolved in TES buffer (100 mM Tris pH 8, 10 mM EDTA, 1 % SDS) and incubated at 60 °C
for 1 h. After centrifugation, 1 volume of 5 M ammonium acetate was added to the supernatant,
followed by incubation on ice for another 1 h. After removal of cell debris by centrifugation, the
DNA was obtained by isopropanol precipitation, dissolved in TE buffer (10 mM TRIS pH 8, 1
mM EDTA) with 50 µg/ml RNAse A and incubated at 37 °C for 1 h (Birnboim and Doly, 1979).
Highly purified DNA was obtained by including a phenol extraction step prior to isopropanol
precipitation (Kirby, 1956).
2.3 PCR and cloning
2.3.1 Generation of deletion cassettes
For PCR-based gene targeting, primers were designed to amplify a dominant resistance marker,
adding target ORF specific flanks enabelling genomic integration of the cassette through
homologous recombination. To enable correct integration of the resistance markers, 40- and 100
bp homology regions were used for A. gopssypii and E. cymbalariae respectively. The deletion
cassettes were amplified by standard PCR conditions (Sambrook and Russel, 2001): 95 °C initial
denaturation for 2 min, followed by 35 cycles of 1 min denaturation at 95 °C, 1 min annealing at
52 °C, 2 min elongation at 72 °C and 5 min final elongation at 72°C.
Gene disruption cassettes were generated in a two-step process (Noble and Johnson, 2005). The
target ORF was amplified from genomic DNA using PCR conditions as described above. For
EcTEC1, an amplicon-native XbaI restriction site and a XhoI restriction site added by the
antisense primer, was used to clone the amplicon into the basic vector pBluescript SK+. The
KANMX resistance marker (Steiner and Philippsen, 1994) was released from pFA-KANMX6 by
EcoRV/PvuII restriction enzyme digest, and cloned to pSK-EcTEC1 through an EcoRV
restriction site (Sambrook and Russel, 2001). Clonal insertion of the KANMX marker disrupted
the EcTEC1 ORF into a 0.93 kb upstream fragment and a 1.36 kb downstream fragment. Before
transformation of E. cymbalariae, the TEC1::KANMX disruption construct was released by XbaI
and XhoI restriction enzyme digest.
In homokaryotic mutants, correct integration of the marker was verified by PCR at both up- and
downstream flanks of the resistancee marker. In addition, complete loss of the target ORF was

Chapter 2: Methods
32
verified by PCR amplification of an internal, ORF-specific fragment. This fragment could be
amplified in wt and heterokaryotic strains, but not in homokaryotic strains.
2.3.2 Vector generation by homologous recombination
S. cerevisiae homologous recombination was used to generate a C-teminal tagged AgGTS1-GFP
construct and for pAG URA3- to NATMX marker exchange. Integration cassettes were amplified
by PCR, using primers adding flanking 40 bp homology regions for plasmid integration. Using
pFA-MoGFP-NAT1 and pFA-AgNATMX4 as templates, the cassettes were ampliofied standard
PCR conditions: 95 °C initial denaturation for 5 min, followed by 35 cycles of 1 min denaturation
at 95°C, 1 min annealing at 52-55 °C, 2 min elongation at 72 °C and 5 min final elongation at 72
°C (Sambrook and Russel, 2001). For the AgGTS1-GFP construct, the GFP-NAT1 cassette was
integrated into pAG19275(GTS1) and NATMX marker exchange was performed on
pAG19275(GTS1) and pAG17522(ARF3). Correct integration of the GFP-NAT cassette was
verified by sequencing performed by LCG Genomics, Germany.
2.3.3 Cloning and functional expression of truncated AgGts1
A. gossypii Gts1 domains were identified by comparing the amino acid sequence (Gattiker et al.,
2007) to conserved domain motifs using PROSITE (Sigrist et al., 2012). Predicted domains from
the PROSITE analysis served as template for two AgGTS1 truncations, named after the domains
in the truncated protein: AgGts1(ArfGAP), containing the N-terminal ArfGTPase activating
protein (ArfGAP) domain, and AgGts1(UPG), containing the C-terminal Ubiquitin associated-
(UBA), Proline-rich (PRO) and Glutamine-rich (GLU) domains.
The AgGTS1(ArfGAP) and AgGTS1(UPG) sequences were amplified from A. gossypii genomic
DNA by standard PCR conditions (Sambrook and Russel, 2001): 95 °C initial denaturation for 2
min, followed by 35 cycles of 30 sec denaturation at 95 °C, 30 sec annealing at 52 °C, 2 min
elongation at 72 °C and 5 min final elongation at 72 °C. Primers were designed to add 5’XhoI and
3’XbaI sites to the amplicons. While the AgGTS1(ArfGAP) sequence contained the native
AgGTS1 start codon, an ATG start codon was added directly in front of the AgGTS1(UPG)
sequence. The AgGTS1 truncations were cloned to pRS418-AgTEFp-LacZ through XbaI/XhoI

Chapter 2: Methods
33
restriction sites, enabling the use of the AgTEF promoter for expression regulation (Sambrook
and Russel, 2001). Correct insert was verified by sequencing of the resultant plasmids. All
sequencing reactions were performed by LCG Genomics, Germany.
2.3.4 Construction of E. cymbalariae centromere plasmids
pFA-KANMX6 was used as vector backbone for construction of a low copy-number plasmids
with E. cymbalariae CEN/ARS. Centromeres from chromosome I and V, EcCEN1 and EcCEN5,
were amplified by standard condition PCR: 94 °C initial denaturation for 2 min, 35 cycles of 1
min denaturation at 93 °C, 1 min annealing at 52 °C, 2 min elongation at 72 °C and a final 5 min
elongation at 72 °C. Using SpeI and SacI restriction sites added to the amplicons by the forward
and reverse primers, the centromere sequences were cloned into pFA-KANMX6 (Sambrook and
Russel, 2001). The resultant plasmid was sequenced by LCG Genomics, Germany.
2.4 Transformation
Transformation of A. gossypii and E. cymbalariae was performed by electroporation according to
the PCR-based gene targeting protocol (Wendland et al., 2000). After transformation the samples
were incubated on AFM agar plates for 6 hours at 30 °C after which the plates were covered by 7
ml 0.5 % agarose with G418/Geneticin for a final antibiotic concentration of 200 mg/ml.
Recombinant heterokaryotic colonies were isolated after 2-3 days incubation at 30 °C. IN order
to isolate single homokaryotic mycelia, spores obtained from heterokaryotic mycelia were
germinated overnight under selective conditions at room temperature. The next day, growing
germlings were micomainpulated using a MSM Micromanipulator (Singer Instruments). Two
separate isolates were obtained for each gene deletion.
Transformation of S. cerevisiae BY4741 was performed according to the LiAc/ss carrier DNA
method (Gietz and Woods, 2002). The samples were plated on YPD agar plates with 50µg/ml
ClonNat, and incubated at 30°C for 3-4 days.
Transformation of E. coli was achieved by electroporation (Dower et al., 1988). After 1 h
incubation at 37 °C, the transformants were plated on 2YT agar plates with appropriate antibiotic

Chapter 2: Methods
34
selection. For blue-white screening of lacZ disruption by clonal insert (Vieira and Meesing,
1982), X-gal to a final concentration of 50 µg/ml was added to the growth medium.
Transformants were selected after overnight incubation at 37 °C.
2.5 Cytological staining and microscopy
A. gossypii and E. cymbalariae mycelial morphology was analyzed using confocal microscopy.
For differential interface contrast (DIC) and GFP imaging, a small mycelial inoculum from an
overnight liquid culture was grown in 10 ml AFM with appropriate selection at 30 °C for 4 h.
Microscopy was performed using an Axio Imager M1 microscope (Zeiss, Germany) and mycelial
images were taken using a MicroMax1024 CCD camera (Princeton Instruments, USA).
Microphotographs were edited using Metamorph 7 software (Molecular Devices LLC, USA).
Fluorescence microscopy was performed using the appropriate filter settings for GFP
fluorescence (excitation 470±20 nm, emission 525 ±25 nm), DAPI (excitation 350 ±25 nm,
emission 460 ±25 nm) and RED (excitation 545 ±15 nm, emission 620 ±30 nm).
A. gossypii septal sites were stained using CFW (Sigma Aldrich) (Pringle et al., 1989). The stain
was added to A. gossypii liquid cultures to a final concentration of 40µg/ml. After incubation at
room temperature for 10 min, the sample was observed with the microscope using the DAPI filter
set. Actin was stained using Rhodamin-Phalloidin (Molecular Probes) on A. gossypii mycelia
fixed in PBS with 3.7 % formaldehyde (Oberholzer et al., 2002). The samples were observed
with microscope using the GFP filter set. To visualize endosomes and vacuoles in A. gossypii, 3-
4 h old mycelial cultures were harvested, diluted in 200 µl AFM with 2 µM FM4-64 (Molecular
Probes) and incubated at room temperature for 15-120 min. Internalization of the FM4-64 dye
was observed by microscopy using the RED filter set (Fischer-Parton et al., 2000).
nnnnnnnnnnnnnnnnn

Chapter 3: Functional analysis of the A.gossypii Arf3-Yel1-Gts1 module
35
Chapter 3: Functional analysis of the A. gossypii Arf3-
Yel1-Gts1 module
3.1 Results
3.1.1 Generation of A. gossypii arf3 and yel1 mutants
S. cerevisiae GTS1 was initially characterized as a clock-gene, since deletion mutants have few
phenotypes except an oscillation phenotype regarding timing of budding (Mitsui et al., 1994). As
GTS1 has previously been deleted in A. gossypii we could compare S. cerevisiae and A. gossypii
gts1 deletion strains and in contrast to S. cerevisiae, a strong growth phenotype was immediately
obvious in A. gossypii gts1 strains. As Gts1 is an Arf3-GAP regulatory protein, we wanted to
generate deletion strains of the AgArf3 small GTPase and the Arf3-GEF AgYel1, which would
enable characterization of the whole A. gossypii Arf3 small GTPase module.
Agarf3 and Agyel1 deletion mutants were generated using PCR-based gene targeting, during
which the target ORF was replaced with a G418/Geneticin resistance marker, GEN3 (Wendland
et al., 2000). Primer pairs #6195-#6196 and #6201-#6202, corresponding to A. gossypii genes
ACL078W (ARF3) and ABR218C (YEL1) were designed to amplify the GEN3 cassette (Figure
12) (Sambrook and Russel, 2001), and homocaryotic Agarf3 and Agyel1 mutants were verified by
PCR (Figure 13).
Figure 12. A. gossypii gene deletion by PCR-based gene targeting. Deletion-cassettes containing a resistance marker and flanking homology region to the target ORF is amplified using S1-21 primers. The deletion cassettes integrate in the genome by homologous recombination, replacing the target ORF with the resistance marker. Correct integration of the deletion cassette is verified by PCR up- and downstream of resistance marker using G1-G2 and G3-G4 primers. Complete deletion of the target gene is verified by amplification of a target OFR specific sequence, using I1-I2 primers.

Chapter 3: Functional analysis of the A.gossypii Arf3-Yel1-Gts1 module
36
Figure 13. Agarose gel electrophoresis of PCR verification fragments. AgARF3 (upper panel) and AgYEL1 (lower panel) deletion were verified by amplifying the GEN3 flanking regions using G1-G2 and G3-G4 primers (see methods). Loss of the target ORF was verified by amplification of an ORF-specific sequence using I1-I2 primers.
Figure 14. Agarose gel electrophoresis of pAG17522(ARF3) (upper panel) and pAG19275(GTS1) (lower panel) internal fragments. Complementation of A. gossypii arf3 and gts1 phenotypes was verified by PCR using ORF-specific internal primers.

Chapter 3: Functional analysis of the A.gossypii Arf3-Yel1-Gts1 module
37
No phenotypes could be observed in Agyel1 but phenotypes observed in the Aggts1 and Agarf3
strains (see section 3.1.2) could be complemented by re-introduction of the deleted ORF using
pAG plasmids pAG19275(GTS1) and pAG17522(ARF3). In addition to the AgGTS1 ORF,
pAG19275 contains the partial sequences of AgMND1 and AgATG1. pAG17522 contains the full
length AgRKI1 ORF as well as the partial sequences of AgRPS7A/B and ACL079C. Re-
introduction of the ORFs was verified by PCR amplification of the ORF-specific internal
sequence (Figure 14).
3.1.2 Growth phenotypes and mycelial morphology
As the obvious growth retardation of the Aggts1 mutants was one of the reasons for our interest in
the Arf3 small GTPase complex, the full scale of Aggts1 growth retardation was carefully
documented. Along with the Agarf3 and Agyel1 mutants, Aggts1 growth were investigated by
growth- and temperature assays, where a small mycelial inoculum was allowed to grow on AFM
agar plates for seven days at 20, 30 and 37 °C. While the Agarf3 and Agyel1 mutants show wt-
like growth during all tested conditions, the Aggts1 strains display severe growth retardation with
successively decread growth at 20, 30 and 37 °C (Figure 15). The decreased growth phenotype of
the Aggts1 mutant was completely rescued by introduction of the AgGTS1 ORF in
pAG19275(GTS1) (see section 3.1.5, Figure 24).
Figure 15. Colony growth of A. gossypii arf3, yel1 and gts1. Colony growth after 7 days on AFM media,
20-37°C, represented as median growth and percentage of wt growth. Standard deviation error bars are
calculated for values with 5 or more replicates.

Chapter 3: Functional analysis of the A.gossypii Arf3-Yel1-Gts1 module
38
By measuring Aggts1 radial growth continuously over 7 days, it was concluded that the retarded
growth phenotype of Aggts1 is most likely due to a generally slower growth and not because of a
log-phase at any time during growth (Figure. 16) The slow growth rate of Aggts1 mutants has not
been seen in S. cerevisiae, which maintain wt-like growth at 30 °C (Mitsui et al., 1994).
Figure 16. A. gossypii wt and gts1 colony growth on full media, 30 °C, 3-7 days after inoculation.
DIC imaging of the Aggts1 mutant mycelia show an abnormal hyphal morphology (Figure 17).
While wt A. gossypii has smooth hyphae with dichotomous branching in mature mycelia, Aggts1
mutants are irregularly shaped with indications of sub-apical hyphal swelling. Lateral branching
is observed in an irregular pattern and although dichotomous tip branching is most prevalent; tip
branching into 3 or 4 new hyphae is readily observed. Aggts1 also exhibits a sporulation
deficiency which were complemented by re-introduction of the AgGTS1 ORF. Standard condition
sporulation in liquid media generated no detectable spores, and spores from solid media were
extremely scarce and of an abnormal morphology (Figure 17).

Chapter 3: Functional analysis of the A.gossypii Arf3-Yel1-Gts1 module
39
FIgure 17. DIC imaging of wt and Aggts1 hyphal- and spore morphology. Arrows indicates Aggts1 sub-apical hyphal swelling and multiple hyphal tip branching. Scale bar for hyphal images is 25 µm and for spore images 10 µm.
3.1.3 Actin and chitin localization
The actin cytoskeleton is tightly coupled to the maintenance of polar growth and endocytosis. In
growing hyphae the actin filaments accumulate at the site of growth, the Spitzenkörper.
Additionally, actin patches occur at a high frequency at an area just below the site of growth, an
area associated with endocytic recycling of proteins associated with polar growth (Shaw et al.,
2011, Sudbery, 2011). ScArf3 has previously been implicated to be in is involved in development
of polarity (Huang et al., 2003) and the Agwal1 and Agsac6 mutants, which have similar growth
retardation phenotypes as Aggts1, have sub-apical actin localization phenotypes (Walther and
Wendland, 2004a, Jorde et al., 2011). Thus, we analyzed the actin localization in the deletion
mutants of the A. gossypii Arf3 small GTPase complex by Rhodamin-Phalloidin actin staining. In
wt A. gossypii as well as the Agarf3 and Agyel1 deletion strains, actin filaments and patches

Chapter 3: Functional analysis of the A.gossypii Arf3-Yel1-Gts1 module
40
cluster at the growing hyphal tip. Actin patches can also be seen evenly distributed throughout
the hyphae. The Aggts1 actin cytoskeleton is not polarized at the hyphal tips, but like Agwal1 and
Agsac6 (Walther and Wendland, 2004a, Jorde et al., 2011), at an area in the sub-apical hyphae.
There is also a clear decrease of actin patches distributed in hyphae of the Aggts1 deletion mutnt
(Figure 18).
Figure 18. Actin localization in A. gossypii mutant hyphae. Agyel1 and Agarf3 have wt-like actin localization at the hyphal tip. The dot-like structures in the hyphae represent actin patches, i.e. former sites of endocytosis. Aggts1 have a sub-apical actin localization phenotype (hyphal tip is marked by (*) and a decreased amount of actin patches in the hyphae. Scale bar is 10 μm
Deletion of AgWAL1 causes a septation defect in A. gossypii (Walther and Wendland, 2004a).
Due to the accumulation of chitin in mature A. gossypii septa, any septation phenotypes can be
easily visualized by chitin staining using the fluorescent dye CFW. Due to the similar growth-,
sporulation and actin localization phenotypes between Agwal1 and Aggts1, we investigated the
ability of Agarf3, Agyel1 and Aggts1 to form mature chitin-rich septa. While chitin accumulates
at septal sites in all three mutants, there is a difference in the septal distribution throughout the
hyphae between the A. gossypii wt and Aggts1. Compared to wt septation, Aggts1 show an

Chapter 3: Functional analysis of the A.gossypii Arf3-Yel1-Gts1 module
41
irregular pattern of septation, and furthermore, an increased chitin content in the cell wall at sites
of sub-apical swelling (Figure 19).
Figure 19. Accumulation of chitin at septal sites in A. gossypii mutant hyphae. While Aggts1 has chitin-rich septa, the septal sites are irregularly dispersed throughout the hyphae. Increased flourescence of the hyphal cell wall visualizes the sub-apical hyphal swellings observed in Aggts1. Scale bar is 10 μm.
3.1.4 Endosome visualization using FM4-64
While mammalian Arf6 has a clear role in uncoating of clathrin-coated endocytic vesicles (Toret
et al., 2008), the role or the S. cerevisiae homologue Arf3 and its GAP and GEF regulators are
less clear. Although not essential, ScArf3 is implicated in endocytosis through a yet unknown
mechanism (Huang et al., 2003). In order to study whether deletion of the AgArf3 small GTPase
module has any effects on endocytosis, the deletion strains were stained with the lipophilic stain
FM4-64, and endocytic uptake of the dye enabled tracing of both early endosomes and vacuoles
at time points 15, 60 and 120 min after addition of the dye (Figure 20).
In A. gossypii wt, endosomes are rapidly visualized by FM4-64. Stained endosomes could also be
detected in Agyel1, and to a lesser extent Agarf3 within 15 min after addition of the dye.
Endosome progression from endocytosis and early endosome to vacuolar fusion occurs similar to
wt in Agyel1. Although the FM4-64 uptake in Agarf3 is slower than wt, the progression of the

Chapter 3: Functional analysis of the A.gossypii Arf3-Yel1-Gts1 module
42
Figure 20. Endocytic uptake of the lipophilic dye FM4-64 in Agarf3, Agyel1 and Aggts1. Hyphae were grown in AFM and pictures were acquired using fluorescence microscopy at time points 15, 60 and 120 min after addition of the dye. Uptake of the dye is delayed in various degrees in the Agarf3 and Aggts1, furthermore Agarf3 appear to have an endosome-vacuolar fusion phenotype. Scale bar is 10 µm.

Chapter 3: Functional analysis of the A.gossypii Arf3-Yel1-Gts1 module
43
endosome through the hyphae appears to be wt-like until the point of vacuolar fusion. 60 to 120
min after addition of FM4-64, the dye stains the vacuoles of the wt hyphae. However, Agarf3
appear to have a endosome-vacuolar fusion phenotype, the endosomes cluster together in the
older hyphae, but does not appear to fuse in order to form vacuoles. The endosome fusion
phenotype is rescued by re-introduction of the AgARF3 ORF in pAG17522(ARF3) (Figure 21).
Figure 21. Complementation of the Agarf3 endosome fusion phenotype by pAG17522(ARF3). The dye can be detected in the vacuoles of A. gossypii wt and pAG17522(ARF3) complementation strain 60 min after addition of the FM4-64 dye. In Agarf3, endosomes cluster without fusing to form a vacuole.
FM4-64 staining of Aggts1 shows a delay in internalization of the dye at the hyphal tip compared
to the A. gossypii wt, and early endosomes are not detectable until 60 min after addition of the
dye. A slower rate of dye internalization is also indicated by the rate of which the FM4-64 dye
reached the older hyphae. Vacuoles of the Aggts1 mutant are visualized after 120 min, while the
dye is detectable in the older wt hyphae already after 15 min, and is clearly visible after 60 min.

Chapter 3: Functional analysis of the A.gossypii Arf3-Yel1-Gts1 module
44
3.1.5 Truncation of AgGts1
Due to the many, and severe phenotypes of Aggts1, most of which could not be observed in the
corresponding S. cerevisiae deletion strains, we wanted to investigate whether AgGts1 might
have any functions additional to the Arf3-GAP activity. Using PROSITE (Sigrist et al., 2012),
four AgGts1 domains were identified and used for two truncated versions of AgGts1 (Figure 22).
Primers #6338-#6339 and #6340-#6341 were designed to amplify the truncations, which were
cloned to shuttle vector pRS418-AgTEFp-LacZ. Expression of the AgGTS1(ArfGAP) truncation
in the Aggts1 background was verified by XbaI/XhoI restriction enzyme digest of the rescued
plasmid. The AgGTS1(UPG) truncation was verified by PCR amplification of an internal
fragment (Figure 23).
Figure 22. The domains of A. gossypii Gts1 as identified by PROSITE: the Arf GTPase activating protein (ArfGAP) domain, the ubiquitinin associated (UBA) domain, the proline-rich (PRO) domain and the glutamine-rich (GLN) domain. Two truncated versions of the Gts1 protein were created, ArfGAP (a a 1-174) and UPG (a a 166-471).
As stated before, colonies of the Aggts1 mutants show a significantly slower growth than A.
gossypii wt, with a correlation between growth retardation and increasing temperature. The
growth retardation observed in Aggts1 is complemented by introduction of the full length
AgGTS1 sequence. AgTEFp overexpression of the AgGTS1(ArfGAP) sequence complements the
Aggts1 mutant phenotype up to 80 % at low- to moderate temperatures (20-30 °C), but only to 35
% at 37 °C. The AgTEFp regulated AgGTS1(UPG) truncation is insufficient for complementation
of the Aggts1 growth retardation and only rescues 35% of the wt growth speed (Figure 24).

Chapter 3: Functional analysis of the A.gossypii Arf3-Yel1-Gts1 module
45
Figure 23. Verification of AgGTS1(ArfGAP) and AgGTS1(UPG) in the Aggts1 background. AgGTS1(UPG) is verified by PCR and AgGTS1(ArfGAP) is verified by XbaI/XhoI restriction enzyme digest. To the left are the general plasmid map used for transformation of Aggts1.
Figure 24. Colony growth of A. gossypii gts1 deletion and complementation strains. Aggts1 is transformed using plasmids containing the full length AgGTS1 ORF, and two AgGTS1 truncations: AgGTS1(ArfGAP) and AgGTS1(UBA). Colony growth is measured after 7 days on AFM media, 20-37°C, and represented as median growth and percentage of wt growth.Standard deviation error bars are calculated for values with 5 or more replicates.

Chapter 3: Functional analysis of the A.gossypii Arf3-Yel1-Gts1 module
46
A clear phenotype of Aggts1 is the sub-apical actin localization similar to those of Agwal1 and
Agsac6 (Walther and Wendland, 2004a, Jorde et al., 2011). Fluorescence microscopy of
Rhodamin-Phalloidin stained Aggts1 complemented with the full length AgGTS1 ORF,
AgGTS1(ArfGAP) and AgGTS1(UBA) indicate that the actin miss-localization phenotype is
complemented by all three versions of the Gts1 protein (Figure 25).
Figure 25. Rhodamin-Phalloidin staining of AgGTS1 deletion, truncation and complementation strains. The hyphal tip of gts1 is marked by (*). Scale bar is 10 µm.
In S. cerevisiae, the Arf3GAP activity of Gts1 depends on the highly conserved, N-terminal
ArfGAP domain (Cukierman et al., 1995). In order to investigate whether the ArfGAP domain is
essential for the Arf3-GAP function of AgGts1, we analyzed the ability of the truncated
AgGTS1(ArfGAP) to complement the Aggts1 slow endocytosis phenotype. Endosome staining
using FM4-64 indicate that the slow endocytic rate of Aggts1 is restored to wt-like when
complemented with a plasmid expressing the AgGTS1(ArfGAP) sequence (Figure 26). Wt-like
endocytic rate was also restored by introduction of the complete AgGTS1 ORF. In contrast, the
AgGTS1(UBA) truncation, which lacks the catalytic ArfGAP domain responsible for regulation
of AgArf3 activity, is unable to restore the Aggts1 endocytic rate.

Chapter 3: Functional analysis of the A.gossypii Arf3-Yel1-Gts1 module
47
Figure 26. Visualization of endocytosis by FM4-64. Hyphal tip endosomes are visible after 15 min in Aggts1 strains complemented by the full length AgGTS1 ORF and the truncated AgGTS1(ArfGAP). AgGTS1(UBA) does not complement the Aggts1 slow endocytosis phenotype. Scale bar is 10 µm.

Chapter 3: Functional analysis of the A.gossypii Arf3-Yel1-Gts1 module
48
3.1.6 AgGts1-GFP localization
Proteins of the Ras superfamily are generally conserved, and many of their GEF and GAP
regulators show a moderate substrate unspecificity in vitro. Even so, in vivo, every small GTPase
have specific regulators, thus, it is generally thought that GEF and GAP specificity depends on
subcellular co-localization with the target substrate (Takai et al., 2001). In order to study the
localization of AgGts1, we constructed an AgGts1-GFP fusion protein by integrating a MoGFP-
NAT1 cassette downstream of the AgGTS1 locus in pAG19275(AgGTS1) through homologous
recombination in S. cerevisiae.
The MoGFP-NAT1 cassette was amplified by primers #6180-#6181, designed to add 40 bp
homology regions to pAG19275(AgGTS1). Integration of the GFP cassette into the pAG plasmid
enables expression of the GTS1-GFP be regulated by the endogenous GTS1 promoter (Figure
27). The GFP-fusion construct was introduced in the Aggts1 background, and in addition to
compementing the Aggts1 growth phenotype, the mutant was verified by restriction enzyme
digest of the rescued plasmid (Figure 28). Florescence microscopy of A. gossypii hyphae
expressing the AgGts1-GFP construct show pattern of GFP signaling along the cell membrane,
with distinct increase in florescence at actin patches, hyphal tips and septal sites (Figure 29).
Figure 27. Generation of pAG19275(AgGTS1-GFP) by homologous recombination of MoGFP-NAT1 to pAG19275(AgGTS1), using short flanking regions homologous to the pAG plasmid. The final construct, pAG19275(AgGTS1-GFP), enables expression of the AgGTS1-GFP fusion protein under control of the endogenous AgGTS1 promoter.

Chapter 3: Functional analysis of the A.gossypii Arf3-Yel1-Gts1 module
49
Figure 28. Verification of Aggts1 pAG19275(AgGTS1-GFP) by NcoI restriction enzyme digest of rescued plasmids.
Figure 29. AgGts1 localization in A. gossypii hyphae. Localization of C-terminal AgGts1-GFP in a growing hypha with a well-defined septal site. Photographs show DIC images of growing hyphae, the AgGts1-GFP signal (green), and DIC/GFP as well as bright field (Red)/GFP overlay. Bright field image is not shown. Scale bar is 10 µm.

Chapter 3: Functional analysis of the A.gossypii Arf3-Yel1-Gts1 module
50
3.2 Discussion
Conserved small GTPases of the Ras superfamily have been shown to be key regulators of many
stages during hyphal growth, an important virulence factor of many pathogenic fungi. One of the
more extensively studied small GTPases is the mammalian Arf6, which regulates different stages
of vesicle trafficking and actin cytoskeleton organization at the plasma membrane, and is
associated with polar cells such as neurons, phagocytic cells and tumor cells. S. cerevisiae Arf3
has previously been established as the homolog of the extensively studied mammalian Arf6, and
is also involved in endocytosis and actin cytoskeleton rearrangements. However, unlike
mammalian Arf6, a link between ScArf3 and polarity has not been established.
In this series of experiments we studied the A. gossypii homologs of the S. cerevisiae Arf3
module and the mammalian Arf6, in an effort to evaluate the impact of the Arf3 module on the
endocytosis and hyphal polarity. As A. gossypii is a close relative to S. cerevisiae, the Arf3
module is assumed to function in an equivalent manner, while the strictly filamentous nature of
A. gossypii enables characterization of the Arf3 module on polar cells.
3.2.1 The A. gossypii Arf3-Yel1-Arf3 module
The S. cerevisiae Arf3, like its mammalian homolog, is thought to modulate the local plasma
membrane content of PI(4,5)P2 through activation of phosphoinositide kinases, e.g. S. cerevisiae
Mss4 (Smaczynska-de Rooij et al., 2008, Desrivières et al., 1998, Honda et al., 1999). As there is
an accumulation of evidence suggesting that increasing plasma membrane concentrations of the
lipid PI(4,5)P2 have a regulatory role in both endocytic vesicle generation and uncoating, changes
in plasma membrane composition might render an explanation to the phenotypes observed in A.
gossypii arf3 and gts1mutants.
Although the effect of A. gossypii Arf3 on PI(4,5)P2 pools were not investigated in this project
due to time limitations, the equivalent S. cerevisiae model might explain the phenotypes observed
in A. gossypii arf3. In S. cerevisiae, Arf3-mediated increase of the membrane PI(4,5)P2 content
facilitates endocytosis as PI(4,5)P2 acts as an anchor for many endocytic proteins through their
PI(4,5)P2-binding domains (Smaczynska-de Rooij et al., 2008). Supporting the link between
Arf3, PI(4,5)P2 and endocytosis proposed for S. cerevisiae, we can demonstrate that while A.

Chapter 3: Functional analysis of the A.gossypii Arf3-Yel1-Gts1 module
51
gossypii arf3 deletion strains are viable, FM4-64 uptake at the hyphal tip occurs at a lower extent
compared to A. gossypii wt hyphae (Figure 20-21), indicating a slower vesicle formation. In
addition, the effect of Arf3 on PI(4,5)P2 concentrations can also be linked to the observed Agarf3
inability of endosome-vacuole fusion. Vesicle coat disassembly is thought to be crucial for
membrane fusion, and endosome fusion phenotypes similar to Agarf3 has previously been
observed in mutant strains deficient in uncoating (Pishvaee et al., 2000, Harris et al., 2000).
While several mechanisms cooperate in the regulation of vesicle uncoating, one important factor
is the dephosphorylation of PI(4,5)P2 to phosphoinositole by synaptojanin, which stimulating
endosome coat disassembly (Sun et al., 2007, Toret et al., 2008, Chung et al., 1997). Assuming
that Agarf3, like the Scarf3 mutants, lacks the increased PI(4,5)P2 concentration associated with
vesicle formation, synaptojanin-mediated PI(4,5)P2 dephosphporylation is drastically reduced.
While other mechanisms triggering coat disassembly might still exist, this could reduce the rate
of uncoating and give rise to the observed endosome fusion phenotype (Toret et al., 2008, Brown
et al., 2001).
While 15 GEFs has been identified for the mammalian Arf6, YEL1 encodes the only identified
Arf3 GEF in both S. cerevisiae and A. gossypii. Through the catalytic Sec7 domain, Yel1
promotes Arf3 activation by accelerating GDP- to GTP-exchange (Casanova, 2007, Gillingham
and Munro, 2007a, Gillingham and Munro, 2007b), and deletion of YEL1 severely delays Arf3
re-activation after GTP hydrolysis (Gillingham and Munro, 2007a). A complete abolishment of
AgArf3 activity in Agyel1 deletion strains would render phenotypes similar to the ones observed
in Agarf3, however, we could not identify any non-wt Agyel1 phenotypes suggesting an AgYel1-
independent AgArf3 activity. This could be due to one or more yet unidentified AgArf3 GEFs
which may possess overlapping functions with AgYel1, or it may be caused by the initial activity
of newly synthesized AgArf3 proteins, which could associate with GTP and remain active until
inactivated by AgGts1 GAP activity (Zurita et al., 2010). Previous studies of both mammalian
and fungal Arf3 has shown that while inactive Arf3 is cytosolic, active Arf3 interacts with the
plasma membrae through a myrisoylated N-terminal α-helix (D’Souza-Schorey and Chavrier,
2006). Thus, any AgArf3 activity in Agyel1 deletion strains might be determined by examining
the localization of an AgArf3-GFP construct.

Chapter 3: Functional analysis of the A.gossypii Arf3-Yel1-Gts1 module
52
In contrast to Agyel1 and Agarf3 deletion strains, deletion of AgGTS1 renders an overactive
AgArf3. As AgGts1 is the only known AgArf3 GAP based on homology to S. cerevisiae, deletion
strains is severely crippled in GTP hydrolysis, which efficiently renders the AgArf3 small
GTPase unable to “shut off” (Gideon et al., 1992, Cukierman et al., 1995, Yaguchi et al., 2007).
Deletion of AgGTS1 causes a drastic phenotype in hyphal growth and morphology (Figure 15-
17), actin assembly (Figure 18) and endocytosis (Figure 20). However, due to the nature of
AgGts1 as an AgArf3 GAP regulator, the severe phenotypic effects observed in Aggts1 deletion
strains could be caused either directly by the loss of AgGTS1, or indirectly by the overactive
AgArf3. In an effort to establish the causative factor for the Aggts1 phenotypes, the possible roles
for AgGts1 are discussed further below.
3.2.2 AgGTS1-GFP localizes at sites of growth and septation
GFP-tagged proteins are commonly used for studying cellular processes and in situ protein
localization in various species (Shimomura et al., 1962). The localization pattern of the AgGTS1-
GFP fusion protein (Figure 29) is consistent with the phenotypes observed in Aggts1 deletion
strains. Localization at actin patches is normally associated with proteins of the endocytic
complex and coat proteins, suggesting that AgGts1, like ScGts1 might be directly involved in the
formation of the endocytic vesicle (Yaguchi et al., 2007). Co-localization with the Spitzenkörper
is usually assigned to proteins involved in polarity establishment and polar growth, including
organization of the actin cytoskeleton (Araujo-Palomares et al., 2009, Delgado-Alvarez et al.,
2010). The link between AgGts1 and actin is further supported by the association of AgGts1 to
sites of septation, an event that is tightly linked to the organization of the actin cytoskeleton in
constricting actomyosin rings (Walther and Wendland, 2003, Chang et al., 1996).
3.2.3 Unregulated AgArf3 causes impaired endocytosis and morphological abnormalities
As discussed above, the phenotypic effects observed in Aggts1 deletion strains could be caused
by the inability of AgArf3 to transition from the active GTP-bound state to the inactive GDP-
bound state. Similar morphological-, polarity- and endocytic phenotypes as observed in Aggts1
have been described for a constitutively active mammalian Arf6 in several independent studies

Chapter 3: Functional analysis of the A.gossypii Arf3-Yel1-Gts1 module
53
(Albertinazzi et al., 2003, Brown et al., 2001, D’Souza-Schorey and Chavrier, 2006, Paleotti et
al., 2005), and a role for S. cerevisiae Arf3 in polarity and endocytosis has been suggested as well
(Huang et al., 2003, Tsai et al., 2008, Smaczynska-de Rooij et al., 2008). The truncated
AgGTS1(ArfGAP), rendering an artificial AgGts1 protein, lacking the proposed UBA, PRO and
GLN domains were used to analyze whether the phenotypes observed in Aggts1 were caused by
the unregulated AgArf3. Since the known function of the remaining ArfGAP domain is to act as a
catalyst of GTP hydrolysis (Scheffzek et al., 1996), it enables proper AgArf3 regulation. The
ability of AgGTS1(ArfGAP) to complement several of the Aggts1 phenotypes show the
importance of the ArfGAP domain in the function of AgGts1, further indicating that the observed
phenotypes could be due to inability of AgArf3 regulation, rather than a direct consequence of the
Aggts1 deletion.
The morphological and endocytic effects of unregulated AgArf3 activity give clues to the protein
mechanism of action. While many functions has been suggested for the mammalian and fungal
homologs of AgArf3 (Costa et al., 2005, Huang et al., 2003, Lambert et al., 2007, Brown et al.,
2001, Radhakrishna et al., 1999), substantial evidence for Arf3-mediated accumulation of plasma
membrane PI(4,5)P2 has been shown in both mammals and other fungal species (Honda et al.,
1999, Brown et al., 2001, Smaczynska-de Rooij et al., 2008). Mammalian Arf6 overexpression
studies have shown that unregulated Arf6 activity causes the formation of vacuoles containing
plasma membrane components, efficiently removing endocytic factors like PI(4,5)P2 from the
plasma membrane (Aikawa and Martin, 2005, Brown et al., 2001, Aikawa and Martin, 2003). As
PI(4,5)P2 is an essential component for endocytic vesicle formation, the unregulated Arf6 activity
causes a secondary inhibitory effect on endocytosis (Smaczynska-de Rooij et al., 2008, Brown et
al., 2001), as observed in the delayed internalization of FM4-64 by Aggts1.
Inhibition of endocytosis could render a possible explanation to the abnormal hyphal morphology
and growth rate observed in Aggts1. Hyphal growth is mediated at the apical Spitzenkörper
structure, through the careful coordination of exo- and endocytosis (Grove and Bracker, 1970,
Girbardt, 1957). While exocytosis functions to maintain hyphal polarity and growth by addition
of membrane compounds at the hyphal apex, endocytosis regulates the distribution of the same
compounds from the area below the Spitzenkörper. Defective endocytosis caused by the
overactive AgArf3 could potentially redistribute polarizing proteins normally concentrated at the

Chapter 3: Functional analysis of the A.gossypii Arf3-Yel1-Gts1 module
54
apex to the sub-apical hyphae, which would give rise to the morphological abnormalities and
reduced growth rate seen in Aggts1 (Taheri-Talesh et al., 2008, Atkinson et al., 2002, Shaw et al.,
2011).
3.2.4 AgARF3-dependent and independent apical actin localization
A clear phenotype of Aggts1 is the localization of the actin cytoskeleton not at the hyphal apex
but at an area in the sub-apical hyphae (Figure 18). The finding that both the AgArf3-regulating
AgGTS1(ArfGAP)- and the AgGTS1(UPG), which lacks the ArfGAP domain responsible for
AgArf3 regulation, rescues the actin localization phenotype gives rise to several possible roles for
AgGts1 in regulation of the actin cytoskeleton dynamics.
As discussed previously, the notion that PI(4,5)P2 regulates several aspects in the actin
cytoskeleton dynamics in S. cerevisiae suggests a causative role for the unregulated AgArf3 in the
Aggts1 sub-apical actin localization. In mammals, high plasma membrane content of PI(4,5)P2
stimulates actin polymerization by activating WASP family NPFs and the Arp2/3 complex
(Rohatgi et al., 1999, Miki et al., 1996). PI(4,5)P2 further effects actin cytoskeleton dynamics by
stimulating uncapping of actin filaments, enabling further actin polymerization (Kim et al.,
2007), and inhibiting the activity of actin-depolymerizing proteins (Janmey et al., 1987).
Unregulated AgArf3 could mediate a constant PI(4,5)P2-dependent actin polarization at the area
of increased endocytosis below the apical Spitzenkörper. An additional consequence of the
increased sub-apical actin accumulation would then be the depletion of free actin monomers from
the cytoplasm, efficiently rendering actin polymerization at the hyphal apex impossible.
However, the finding that the sub-apical actin localization is rescued by introduction of the
AgGTS1(UPG) construct, lacking the AgArf3-regulatory ArfGAP domain, suggest that AgGts1
has an additional role in the dynamics of the actin cytoskeleton. The mechanism of which AgGts1
might aid in the actin assembly is largely unknown, although a link between Gts1, Arf3 and the
endocytic coat protein Lsb5 has been identified in S. cerevisiae (Toret et al., 2008, Costa and
Ayscough, 2005). To further suggest a role for AgGts1 in actin organization, the sub-apical actin
localization in Aggts1 is similar to those observed in Agsac6 and Agwal1 deletion strains (Jorde et
al., 2011, Walther and Wendland, 2004a). As SAC6 encodes the only actin-bundling fimbrin

Chapter 3: Functional analysis of the A.gossypii Arf3-Yel1-Gts1 module
55
identified in A. gossypii (Adams et al., 1989), and AgWAL1 encodes a NPF homologous to the
mammalian WASP (Walther and Wendland, 2004a), they are both directly implicated in the
organization of the actin cytoskeleton, and like Aggts1, they both have endocytic deficiencies and
show signs of abnormal polar establishment (Jorde et al., 2011, Walther and Wendland, 2004a).
Although the relationship between AgGts1, AgSac6 and AgWal1 is unclear, it does highlight the
link between endocytosis, polarity establishment and the dynamics of the actin cytoskeleton and
shows the complexity of continuous filamentous growth.

Chapter 4: Eremothecium pathogenicity assays
56
Chapter 4: Eremothecium pathogenicity assays
4.1 Results
Although A. gossypii is known for causing stigmatomycosis in cotton and dry rot in citrus fruit,
the exact mechanism of infection is not known (Wendland and Walther, 2005a). A. gossypii has
already been used as a model organism for filamentous growth, and development of
pathogenicity assays for the Eremothecium genus might increase the applicability of A. gossypii
to other pathogenic fungi. Here we use pathogenicity assays developed for the pathogenic fungi
F. oxysporum to analyze the pattern of pathogenicity in the Eremothecium genus, were A.
gossypii and E. cymbalariae are considered representative species.
4.1.1 Cellophane penetration assay
Physical barriers are the main protection against pathogens in both plant and animal tissue and
the penetration of a physical barrier is often a key event during infection. To explore the ability of
A. gossypii and E. cymbalariae to penetrate a physical barrier by sheer force, the cellophane
penetration assay is set up to form a barrier between the fungus and the nutrient-rich media.
While the positive control, F. oxysporum, penetrates the cellophane within 2 days of growth, and
regardless of media condition, neither A. gossypii or E. cymbalariae hyphae penetrates the
mycelia during the 6 days trial (Figure 30).
Figure 30. Cellophane penetration by A. gossypii, E. cymbalariae and F. oxysporum when grown on full media. Upper row: Photos of fungal colonies before removal of the cellophane. Bottom row: mycelial regrowth after removal of the cellophane sheet. F. oxysporum was incubated 4 days while A. gossypii and E. cymbalariae 6 days before removal of the cellophane sheet.

Chapter 4: Eremothecium pathogenicity assays
57
4.1.2 Fruit pathogenicity assay
The fruit pathogenicity assay was constructed to evaluate the ability of A. gossypii to grow on the
surface of-, and invade fruit tissue. After 7 days at 30 °C and 100 % humidity, the positive
control F. oxysporum has invaded and caused rotting of the apple tissue as well as the orange
exo-, meso- and endocarp (Figure 31). Microscopic analysis of the fruit tissue confirms that the
mycelia have completely invaded the fruit tissue and mycelia can be identified in tissue samples
opposite of the site of infection. Microscopic analysis shows that even though E. cymbalariae
spores were present in both the apple and fruit assays, the spores were unable to germinate in the
fruit tissue and growing mycelia could not be identified in any assay. While the A. gossypii wt
and leu2 spores were unable to germinate in the apple assay, hyphal filaments could be found in
the orange endocarp assay. However, microscopic analysis of the orange tissue suggests that the
colonies are superficial at the sites of infection, and the mycelia do not penetrate the orange
tissue.
Figure 31. Invasive growth of apple and orange tissue. A. gossypii wt, Agleu2 and F. oxysporum wt spore suspensions were inoculated at the exposed tissue surface. The pictures are taken after 7 days incubation at 30 °C, 100 % humidity.

Chapter 4: Eremothecium pathogenicity assays
58
4.2.3 Galleria mellonella killing assay
G. mellonella larvae were chosen as an animal model for Eremothecium fungal infections, and
sickness could easily be scored by daily control of larval melanization. As melanization of the
normally pale yellow larvae is a clear indicator of activation of the innate immune system,
melanized larvae were scored as sick or, if non-responsive to physical stimuli -dead (Burgwyn
Fuchs et al., 2010). Injection of A. gossypii and E. cymbalariae spores into G. mellonella larvae
induced a rapid melanization reaction of the hemocoel. Within 15 min, 40 % of the larvae
injected with A. gossypii spores, and all 100 % of the larvae injected with E. cymbalariae spores
showed a clear darkening of the hemocoel. The rapid, intense melanization response also occurs
in all 100 % of the heat killed A. gossypii spores used as a negative control. In contrast, the
control group with larvae injected with F. oxysporum microcornidia shows no signs of illness at
this point.
Daily scoring of the fungal infection progress revealed that the F. oxysporum control kills 100 %
of the larvae within 4 days of injection. Spores from A. gossypii and E. cymbalariae kill more
than 50 % of the larvae in each assay over the course of 7 days (Figure 32). However, larvae
injected with heat-killed A. gossypii spores have a 100 % mortality rate in just 2 days after
injection. Furthermore, microscopical analysis of the larval hemocoel does not reveal any
germinated pores of any Eremothecium fungi, which suggests that the G. mellonella larvae are
not killed by an infection, but rather a physical effect of the spores.
Figure 32. Virulence of A. gossypii, E. cymbalariae and F. oxysporum on G. mellonella. Kaplan-Meier plots of G. mellonella survival after injection of spores to the larval hemocoel.

Chapter 4: Eremothecium pathogenicity assays
59
4.2 Discussion
Fungal pathogens have great impact on human society, both through human infections and
agricultural and economic loss. In order to fight fungal infections, and to find new targets for
antifungal chemicals it is necessary to increase the understanding of the mechanisms of fungal
growth and pathogenesis. Knowledge of A. gossypii mechanism of infection could greatly
enhance the use of the fungus as a model organism for filamentous growth, thus, we aimed at
developing assays for evaluating the ability of Eremothecium fungi to infect and cause disease in
plant and animal tissue. While not taking into account any molecular mechanism for infection,
we investigated whether Eremothecium fungi could penetrate a physical barrier, a key event
during host infection, and proliferate and cause disease in fruit and animal tissue.
4.2.1 Characteristics of filamentous growth in Eremothecium fungi
In order to infect host tissue, many pathogenic fungi have evolved mechanisms to penetrate host
defense barriers. Either by degrading enzymes or sheer force, the fungal hyphae invades the host
tissue where it can proliferate in a nutrient-rich environment (Dean et al., 2012). By exploring the
capability of invasive growth in fungi of the Eremothecium genus, we hoped to gain insight in the
mechanism of pathogenicity for A. gossypii.
Previous reports have shown that phytopathogenic fungi such as F. oxysporum are able to
penetrate an artificial barrier such as a cellophane sheet and invade the underlying media, and
deletion of genes encoding essential pathogenicity factors often reduces the ability of pathogenic
fungi to invade cellophane covered agar (Pérez-Nadales and Di Pietro, 2011, Rispail and Di
Pietro, 2009). In order to analyze the invasive growth of Eremothecium species, we
systematically compared cellophane invasion of A. gossypii, E. cymbalariae and F. oxysporum.
As shown previously, F. oxysporum was able to penetrate the cellophane and invade the
underlying media within days of vegetative growth. In contrast, while the vegetative growth of A.
gossypii and E. cymbalariae was wt-like, they were both unable to penetrate the cellophane.
A. gossypii has previously been reported as a cotton- and citrus fruit pathogen spread by insects
of the Pyrrhocoridae family, although the mechanism of infection is not clear (Ashby and
Novell, 1926, Dietrich et al., 2013). We can demonstrate that E. cymbalariae is non-pathogenic,
and unable to germinate in neither apple- nor orange tissue. While not being able to germinate in

Chapter 4: Eremothecium pathogenicity assays
60
apple tissue, A. gossypii spores geminate in the nutrient rich orange endocarp, but do not invade
the tissue. Additionally, despite the fact of being the cause of dry rot, and irregardless of growing
in the endocarp, A. gossypii did not germinate- or grow on the surface of the citrus fruit. Due to
the apparent inability of A. gossypii to invade host tissue, we hypothesize that the fungus is
dependent on insects of the Pyrrhocoridae family not only as a vector for spreading, but also for
the mechanical force necessary for gaining access to the sites of growth. The use of insect vectors
for fungal plant infection has been noted previously (Medrano et al., 2009), and as Pyrrhocoridae
insects are known pests feeding on both cotton seeds and various fruit, the A. gossypii spores
might be delivered to the infection site through the insect feeding on the plant host (Kimura et al.,
2008).
4.2.2 Eremothecium spores causes G. mellonella immune reaction
Designing animal models for fungal pathogenicity there are ethical and logistical reasons for
using invertebrates for initial infection models (Arvanitis et al., 2013). While several invertebrate
animal models have been developed, we chose the larvae of the greater wax moth, G. mellonella
for assaying Eremothecium animal infection. The relatively large size of G. mellonella larvae
enabled injection of spores to the larval hemocoel without causing significant trauma to the
larvae, although injection of Eremothecium spores cause a rapid melanization of the larvae. As
the G. mellonella immune system is based on humoral phagocytic cells, which after ingestion of
the pathogen forms melanin-rich aggregates in the hemocoel (Ratcliffe, 1985, Lionakis, 2011),
the rapid melanization strongly suggest an intense immune reaction to the Eremothecium spores.
While the G. mellonella immune response to Eremothecium spores can be followed by scoring
melanization and dead larvae (Burgwyn Fuchs et al., 2010, Navarro-Velasco et al., 2011), it is
unlikely that the Eremothecium fungi are able to establish an infection. The observation that heat-
killed A. gossypii spores also induces rapid melanization, and the fact that no germinated spores
could be identified in the larval hemocoel, suggests that the larval melanization is caused by the
humoral phagocytes reacting to the presence of spores and does not correspond to the progress of
fungal infection. Thus, it is likely that under the tested conditions, A. gossypii and E. cymbalariae
are non-virulent in the sense that they do not invade invertebrate host tissue.

Chapter 5: Establishing molecular tools in E. cymbalariae
61
Chapter 5: Establishing molecular tools in E.
cymbalariae
5.1 Results
5.1.1 Establishing E. cymbalariae PCR based gene targeting
In an effort to establish PCR-based gene targeting in E. cymbalariae, a close relative to A.
gossypii, we sought to analyze whether 100 bp homology regions would suffice for correct
resistance marker integration into the E. cymbalariae genome. As target genes, we chose
ECYM7434 and ECYM5230. While ECYM5230 encodes a hypothetical protein with no
identifiable homologs in A. gossypii or S. cerevisiae, the 2292 bp ECYM7435 ORF encoding a
homolog to A. gossypii transcriptional regulator Tec1.
For PCR-based gene targeting, deletion cassettes with 100 bp homology regions were amplified
using primers #6333-#6334 (EcTEC1) and #6329-#6330 (ECYM5230) and ~100ng of DNA were
transformed into E. cymbalariae germlings in order to obtain two independent strains for each
target gene. The number of transformant colonies were compared those generated using
EcTEC1::KANMX disruption cassettes with a 10-fold increase in homologt region size (Table 3).
No difference in correct KANMX integration could be observed when comparing gene deletion
using 100 bp homology regions and ~1000 bp homology regions. Two independent
homokaryotic strains were isolated for ecym5230, Ectec1 and EcTEC1::KANMX (Figure 34).
Table 3. Transformation of gene disruption constructs and gene deletion cassettes in E.
cymbalariae. pRS417-AgTEFp-GFP was used as positive control.
Sample Deletion method Homology region Transformants obtaied/analyzed/verified
ECYM5230 deletion cassette 100 bp 5/5/4
EcTEC1 deletion cassette 100 bp 9/5/5
EcTEC1 TEC1::KanMX disruption cassette
0.93-1,36 kb 6/5/5
Positive control
- - >100
Negative control
- - 1

Chapter 5: Establishing molecular tools in E. cymbalariae
62
PCR amplification of the target ORF up- and downstream regions verified that the PCR-based
deletion cassettes integrated into the genome and replaced the ORF with the KANMX marker.
Since E. cymbalariae mycelia consist of multinucleated compartments, primary heterokaryotic
transformants contain both wt- and mutant nuclei. Sporulation and selective germination of
uninuclear spores enabled isolation of homokaryotic mutant mycelia, in which loss of the target
ORF could be verified by PCR amplification of an internal, ORF-specific fragment. In
TEC1::KANMX disruption mutants, the internal fragment is not missing, but integration of the
KANMX marker should lead to an 1.,4 kb increase of the amplicon size (Figure 33).
Figure 33 Construction of a disruption cassette for gene deletion of TEC1 in E. cymbalariae. The amplified EcTEC1 ORF is cloned into the basic plasmid pBluescript SK+. ORF disruption is achieved by clonal insertion of the KANMX resistance marker, containing the E. coli Kan
R gene flanked by the A. gossypii
TEF promoter and terminator sequences. The disrupted ORF creates two flanks, which were used for homologous recombination.

Chapter 5: Establishing molecular tools in E. cymbalariae
63
Figure 34. Agarose gel electrophoresis of PCR verification fragments. EcTEC1::KANMX gene disruption (upper panel), EcTEC1 (middle panel) and ECYM5230 (bottom panel) deletions were verified by amplifying the KANMX flanking regions using G1-G2 and G3-G4 primers. Loss of the target ORF was verified by amplification of an ORF-specific sequence using I1-I2 primers.

Chapter 5: Establishing molecular tools in E. cymbalariae
64
5.1.2 Characterization of Ectec1 and ecym5230
When optimizing gene deletion in E. cymbalariae, we chose to target two genes which pose
special interest areas that might improve the use of E. cymbalariae as a model organism beside A.
gossypii: the unknown, no-homolog gene ECYM5230 and EcTEC1, encoding a
transcriptionregulator involved in filamentous and invasive growth as well as sporulation
(Grünler et al., 2010).
The homokaryotic E. cymbalariae tec1 and ecym5230 deletion strains were used to investigate
possible roles for the corresponding EcTec1 and Ecym5230 proteins. Growth assays of the E.
cymbalariae mutants on full media at 30 °C, indicate wt-like phenotypes of both ecym5230
strains and Ectec1 deletion strains, independent of method for integration of the KANMX marker
(Figure 35 and Figure 36). DIC imaging of hyphal morphology is consistent with the wt-like
phenotypes of the ecym5230, Ectec1 and EcTEC1::KANMX mutants (Figure 37). Furthermore,
the increased sporangia or spore formation observed in A. gossypii could not be observed
microscopically in Ectec1 deletion mutants (data not shown).
Figure 35. Colony growth of E. cymbalariae ecym5230, Ectec1 and EcTEC1::KANMX mutants. Colony appearance after 7 and 14 days of growth on full media, 30 °C.

Chapter 5: Establishing molecular tools in E. cymbalariae
65
Figure 36. Colony growth of E. cymbalariae ecym5230, tec1 and TEC1::KANMX mutants. Median colony growth (mm) after 7-14 days on AFM media at 30 °C. Standard deviation error bars are calculated for strains with 5 or more replicates. Figure 37. Differential interference contrast (DIC) imaging of E. cymbalariae wt, Ectec1, EcTEC1::KANMX and ecym5230. Scale bar is 50 µm.

Chapter 5: Establishing molecular tools in E. cymbalariae
66
5.1.3 Assessment of CEN and ARS function in E. cymbalariae
The ARS sequence motif enables plasmid propagation in eukaryotes, and are essential for
plasmid replication. In contrast, centromeres are not essential for eukaryotic vectors, but they do
increase the mitotic and meiotic stability of the plasmid (Clarke and Carbon, 1980). While S.
cerevisiae ARS, but not CENs, are stable in A. gossypii (Wright and Phillipsen, 1991), it is
unknown whether S. cerevisiae and A. gossypii CEN/ARS elements are stable in E. cymbalariae.
Alignments of the A. gossypii and E. cymbalariae centromeric DNA elements, CDEs, together
with the S. cerevisiae CDE consensus sequences, show that the CDEs, specially the cysteine
residues, are conserved between the species (Figure 38). Thus, we analyzed the stability of
plasmids containing ARS/CEN sequences of S. cerevisiae and A. gossypii in E. cymbalariae, and
compare those to the stability of the native ARS/CEN sequences.
Figure 38: Interspecies alignment of centromeric DNA elements CDEI, CDEII and CDEIII. Nucleotide sequences from A. gossypii and E. cymbalariae are compared to the corresponding consensus sequences of S. cerevisiae. The less conserved CDEII sequences are represented by sequence length and percentage of A-T content. In the S. cerevisiae consensus sequences, U represents purine (A or G), R represents A or T, and N represents any base (A, T, C or G).

Chapter 5: Establishing molecular tools in E. cymbalariae
67
The plasmid pFA-KANMX6 served as backbone for construction of two E. cymbalariae
centromeric vectors rendering G418/Geneticin antibiotic resistance. E. cymbalariae genomic
sequences containing ARS elements and CEN1 or CEN5 were amplified using primers #4800-
#4801 and #5030-#4817 respectively, and cloned to pFA-KANMX6 through SpeI/SacI restriction
sites added during PCR amplification (Figure 39).
Figure 39. E. cymbalariae CEN/ARS vectors. (A) Construction of E. cymbalariae CEN1 and CEN5 plasmids by cloning the centromeric sequence to pFA-KanMX6 through SpeI/SacI restriction sites. (B) The structures of CEN1 and CEN5, showing the number and relative positions of the autonomously replicating sequences (ARS), as well as the centromere DNA element (CDE) I, II and III.
E. cymbalariae germlings were transformed with the pFA-KANMX6-EcCEN1 and pFA-
KANMX6-EcCEN5 as well as plasmids containing A. gossypii and S. cerevisiae CEN/ARS
(Figure 40). Initial transformants were obtained for all plasmids during selective growth on 200
µg/ml G418/Geneticin. However, pRS-plasmids with ScCEN/ARS sequences appear unstable,
and antibiotic resistance is lost within a few days of growth in liquid selective full media (Data
not shown). Even so, ScCENs are more stable on solid media and the plasmids are maintained
during selective growth (Figure 40 C-D). The vector with an AgCEN/ARS is mitotically stable
with 72.2% plasmid retention after 10 days incubation without selective pressure. In comparison,
a sole A. gossypii ARS renders 100% plasmid loss in non-selective conditions (Figure 40 A-B).
Both EcCEN plasmids have an observed antibiotic retention of 100 % after 10 days of non-
selective growth (Figure 40 E-F). However, in both cases introduction of EcCEN plasmids also

Chapter 5: Establishing molecular tools in E. cymbalariae
68
cause a slow growth phenotype which raise the question of genomic integration. Furthermore, we
were unable to recover the plasmids by re-transformation of E. cymbalariae DNA into E. coli.
Plasmid Origin Resistance
marker CEN ARS
A pLC-Shuttle-A A. gossypii A. gossypii KanMX, NatMX B pHC-Shuttle-B - A. gossypii KanMX C pRS417-ScTEFp1-LacZ S. cerevisiae S. cerevisiae GEN3 D pRS415-KanMX S. cerevisiae S. cerevisiae KanMX
E pFA-EcCEN1 E. cymbalariae E. cymbalariae KanMX F pFA-EcCEN5 E. cymbalariae E. cymbalariae KanMX
Figure 42. Loss of G418/Geneticin resistance in E. cymbalariae plasmid transformants. Upper row:
regrowth on selective full media after 10 days of selective growth. Lower row: regrowth on selective full
media after 10 days of non-selective growth. CEN and ARS sequences are obtained from A. gossypii (A-
B), S. cerevisiae (C-D) and E. cymbalariae (E-F). Table: CEN and ARS sequences tested originate from
A. gossypii, E. cymbalariae and S. cerevisiae
5.2 Discussion
5.2.1 Establishing PCR-based gene targeting in E. cymbalariae
Homologous recombination has been established as a tool for engineering DNA mutations in
many fungal species, and under optimal conditions it allows for a single mutation without any
unspecific genomic alterations (Klinner and Schäfer, 2004). However, in many species
homologous recombination is unspecific, and the transformation efficiency often requires time-

Chapter 5: Establishing molecular tools in E. cymbalariae
69
consuming screening for the correct mutation. In an effort to reduce non-specific integration,
sequences for homologous recombination are usually several hundred bp, and construction of the
cassettes is often a time-consuming multi-step process (Krawchuk and Wahls, 1999, Tatebayashi
et al., 1994).
Gene deletion in the filamentous fungus A. gossypii is uncommonly efficient, and a mere 40 bp
homology region is sufficient for a correct integration of a resistance marker cassette into a
specific locus in the A. gossypii genome. The efficient homologous recombination of A. gossypii
has enabled the development of PCR-based gene targeting, which uses a one-step PCR reaction
for deletion cassette amplification, and is therefore time-saving compared to the traditional use of
gene disruption cassettes (Wendland et al., 2000). In an effort to establish PCR-based gene
targeting in E. cymbalariae, a close relative to A. gossypii (Wendland and Walther, 2011), we
used deletion cassettes with 100 bp homology regions for transformation of E. cymbalariae
mycelia. Our results show that although the initial transformation frequency of E. cymbalariae is
relatively low, correct integration of the deletion cassette could be verified in the vast majority of
the tested transformants (Table 3). Furthermore, there was no obvious difference in
transformation frequency using PCR-based deletion cassettes with 100 bp homology regions and
the disruption cassette with ~1000 bp homology regions. PCR verification of the correct
integration confirms that the PCR-based gene targeting in E. cymbalariae is accurate, and correct
integration of the marker occurs with high fidelity.
5.2.2 Wt-like phenotypes of E. cymbalariae TEC1 and ECYM5230
The growth phenotypes and hyphal morphology of Ectec1 and ecym5230 are both equivalent to
E. cymbalariae wt. The lack of ECYM5230 orthologs were the main cause of our interest in the
gene, and the reason for creating an ecym5230 deletion strain. However, while we can show that
ECYM5230 is non-essential, the lack of a deletion phenotype severely limits the understanding of
the gene function. Additional clues to the Ecym5230 protein function could be gained by further
genetic manipulations, e.g. Ecym5230-GFP localization and expression studies, but due to time
limits, neither was done in this study.

Chapter 5: Establishing molecular tools in E. cymbalariae
70
Unlike ECYM5230, EcTec1 have known orthologs in both S. cerevisiae and A. gossypii and in
both species TEC1 is a transcription regulator implicated in mating, filamentous growth and
adhesion. Be that as it may, the S. cerevisiae and A. gossypii deletion phenotypes differ in some
aspects which caused our interest in the E. cymbalariae TEC1. While S. cerevisiae tec1 deletion
strains are incapable of pseudohyphal- and invasive growth (Köhler et al., 2002), Agtec1 strains
are more similar to the Ectec1 strains and maintain filamentous growth at a slightly reduced rate
compared to A. gossypii wt. Furthermore, A. gossypii TEC1 deletion strains do not abolish
invasive growth, and also displays a clear over-sporulation phenotype (Grünler et al., 2010,
Grünler, 2010). Although no clear oversporulation phenotype was observed in E. cymbalariae
tec1 deletion strains, both phenotypes have to be further evaluated in E. cymbalariae at a later
occasion.
5.2.3 A. gossypii centromeres are stable in E. cymbalariae
The development of interspecies shuttle vectors has greatly facilitated genetic engineering and
DNA manipulations in several species (Clarke and Carbon, 1985). In order to establish a system
of stable shuttle vectors for the filamentous fungus E. cymbalariae, we have compared the
stability of replication origins and centromeres originating from S. cerevisiae, A. gossypii and E.
cymbalariae during selective and non-selective growth.
Plasmid pHC-Shuttle-B, containing only an A. gossypii self-replicating sequence, AgARS, is
rapidly lost during non-selective growth. There is a general consensus that the instability of an
ARS-only plasmid is caused by an inability of the plasmid to segregate during mitotic division
(Stinchcomb et al., 1979, Clarke and Carbon, 1985). Even though the hyphae of Eremothecium
fungi are multinucleated, the nuclei in growing hyphae move towards the hyphal apex, and non-
segregating plasmids will have a strong bias towards remaining in the nuclei of the older hyphae
(Gibeaux et al., 2012). Thus, even though ARS-only plasmids might be present in older mycelia,
there is little to no chance of migration of an ARS-only plasmid to the hyphal apex at the edge of
a colony. Introduction of a functional centromere into the plasmid backbone enables segregation
of the plasmid throughout the hyphae (Murray and Szostak, 1983). As G418/Geneticin resistance
is maintained in hyphal samples transformed with an AgCEN-plasmid, pLC-Shuttle-A, we
conclude that A. gossypii centromeric sequences mediate mitotic segregation in E. cymbalariae.

Chapter 5: Establishing molecular tools in E. cymbalariae
71
While G418/Geneticin resistance is maintained using native E. cymbalariae centromeres,
transformed mycelia also show a slow grow phenotype. Failed efforts to recover the EcCEN
plasmids by transformation of E. coli with E. cymbalariae genomic DNA further strengthens the
hypothesis that using native E. cymbalariae centromeres enables genomic integration of the
plasmid.
Plasmids containing S. cerevisiae CEN6 and ARSH4 sequences are not stable in E. cymbalariae.
As the G418/Geneticin resistance is maintained during selective growth on solid media, the
ScARS is functional, however the resistance marker loss in liquid cultures suggest that the ARS
activity might be weak. In addition to analyzing the ScCEN/ARS activity, the use of two
G418/Geneticin resistance marker modules, KANMX and GEN3, enabled us to assess the function
of A. gossypii and S. cerevisiae TEF promoter and terminator activity in E. cymbalariae. Both
modules contain the E. coli KanR gene: in the KANMX module, expression is regulated by the
AgTEF promoter and terminator (Steiner and Philippsen, 1994), and in GEN3 it is regulated by
the ScTEF promoter and terminator (Wendland et al., 2000). As G418/Geneticin resistant
transformants are generated using either KANMX or GEN3 plasmids, expression regulation in E.
cymbalariae can be achieved using either S. cerevisiae or A. gossypii regulatory sequences.

Chapter 6: Summary and future prospects
72
Chapter 6: Summary and future prospects
The conserved nature of small GTPases makes them interesting targets for comparative studies.
This study focus on the A. gossypii ARF3 module due to the impact of excess of Arf3 activity on
filamentous growth, an important factor for fungal pathogenicity. Comparing the A. gossypii
ARF3 small GTPase with its yeast and mammalian homologs, it is likely that Arf3 acts by
modulating the plasma membrane content of the lipid PI(4,5)P2 by directing the activity of
phosphatidylinositide kinases. While the unregulated activity of Arf3 causes endocytic, actin
localization- and morphological phenotypes which can all be linked to an excess of membrane
PI(4,5)P2 content, the direct effect of Arf3 on membrane composition and PI(4,5)P2
concentrations has not been addressed in this study. In addition to addressing the correlation
between Arf3 and PI(4,5)P2, further linkage between the excess of Arf3 activity in the gts1
deletion strains could be demonstrated through either overexpression of the YEL1 GEF, or
construction of a constitutively active ARF3.
In order to compare A. gossypii to other fungi, we established a set of pathogenicity assays to
determine the limits of filamentous growth in species of the Eremothecium genus. In addition,
this study also establishes molecular tools for E. cymbalariae, a close relative to A. gossypii. Due
to the fact that the yeast S. cerevisiae only does short rounds of polar growth during budding, it is
valuable to compare the impact of the modified Arf3 module in other filamentous fungi, such as
E. cymbalariae. As we established that neither A. gossypii or E. cymbalariae are able to penetrate
host tissue, and thus are limited in the development of an infection structure, it would be
interesting to evaluate the Arf3 module in a pathogenic fungus. This could possibly address the
importance of filamentous growth during fungal infections and would enlighten whether the Arf3
module poses potential targets for antifungal drugs. In addition, the conserved nature of small
GTPases, further investigations of the A. gossypii ARF3 module could be valuable when
compared to mammalian Arf6, a known oncogene. As Arf6 is regulated by several different
GEFs and GAPs, research in the regulation of Arf6 activity is particularly difficult. The fact that
A. gossypii ARF3 only has one GAP and one GEF, GTS1 and YEL1 respectively, facilitates
deletion of regulatory proteins and enables further understanding of the Arf3-, and consequently
the mammalian Arf6 function.

Chapter 7: References
73
Chapter 7: References
ADAMS, A. E., BOTSTEIN, D. & DRUBIN, D. G. 1989. A yeast actin-binding proteinis
encoded by SAC6, a gene found by suppression of an actin mutation. Science, 243, 231-
233.
AGHAMOHAMMADZADEH, S. & AYSCOUGH, K. R. 2010. The yeast actin cytoskeleton and
its function in endocytosis. Fungal Biology Reviews, 24, 37-46.
AIKAWA, Y. & MARTIN, T. F. J. 2003. ARF6 regulates a plasma membrane pool of
phosphatidylinositol (4,5)bisphosphate required for regulated exocytosis. Journal of Cell
Biology, 162, 647-659.
AIKAWA, Y. & MARTIN, T. F. J. 2005. ADP-rbosylation factor 6 regulation of
phosphatidylinositol 4,5-bisphosphate synthesis, endocytosis and exocytosis. methods in
Enzymology, 404.
ALBERTINAZZI, C., ZA, L., PARIS, S. & DE CURTIS, I. 2003. ADP-ribosylation factor 6 and
functional PIX/p95-APP1 complex are required for RAc1B-meidated neurite outgrowth.
Molecular Biology of the Cell, 14, 1295-1307.
ANTONESCU, C. N., AGUET, F., DANUSER, G. & SCHMID, S. L. 2011.
Phosphatidylinositol-(4,5)-bisphosphate regulates clathrin-coated pit initiation,
stabilization and size. molecular Biology of the Cell, 22, 2588-2600.
ANTONNY, B., BERAUD-DUFOUR, S., CHARDIN, P. & CHABRE, M. 1997. N-terminal
hydrophobic residues of the G-protein ADP-ribosylation factor-1 insert into membrane
phosholipids upon GDP to GTP exchange. Biochemistry, 36, 4675-4684.
ARAUJO-PALOMARES, C. L., RIQUELME, M. & CASTRO-LONGORIA, E. 2009. The
polarisome component SPA-2 localizes at the apex of Neurospora crassa and partially
colocalizes with the Spitzenkörper. Fungal Genetics and Biology, 46, 551-563.
ARVANITIS, M., GLAVIS-BLOOM, J. & MYLONAKIS, E. 2013. Invertebrate models of
fungal infection. Biochemica et Biophysica Acta, 1839, 1378-1383.
ASHBY, S. F. & NOVELL, W. 1926. The fungi of stigmatomycosis. Ann. Bot. , 40, 69-84.
ATKINSON, H. A., DANIELS, A. & READ, N. D. 2002. Live-cell imaging of endocytosisi
during conidial germination in the rice blast fungus, magnaporthe grisea. Fungal Genetics
and Biology, 37, 233-244.
BALLA, T. 2005. Inositol-lipid binding motifs: signal integrators through protein-lipid and
protein-protein interactions. Journal of Cell Science, 118, 2093-2104.
BARNES, G., DRUBIN, D. G. & STEARNS, T. 1990. The cytoskeleton if Saccharomyces
cerevisiae. Current opinion in cell biology, 2.
BIRNBOIM, H. C. & DOLY, J. 1979. A rapid alkaline extraction procedure for screening
recombinant plasmid DNA. Nucleic Acid Research, 24, 1513-1523.
BOETTNER, D. R., CHI, R. J. & LEMMON, S. K. 2012. Lessons form yeast from clathrin-
mediated endocytosis. Nature Cell Biology, 14, 2-10.
BOMPARD, G. & CARON, E. 2004. Regulation of WASP/WAVE proteins -making a long story
short. Journal of Cell Biology, 166, 957-962.
BOSSIER, P., GOETHALS, P. & RODRIGUES-POUSADA, C. 1997. Constitutive flocculation
in Saccharomyces cerevisiae through overexpression of the GTS1 gene , coding for a Glo-
type Zn-finger-containing protein. Yeast, 13, 717-725.

Chapter 7: References
74
BOTSTEIN, D. & FINK, G. R. 2011. Yeast: An experimental organism for 21st century biology.
Genetics, 189, 695-704.
BOURNE, H. R., SANDERS, D. A. & MCCORMICK, F. 1991. The GTPase superfamily:
conserved structure and molecular mechanism. Nature, 349, 117-127.
BROWN, F. D., ROZELLE, A. L., YIN, H. L., BALLA, T. & DONALDSON, J. G. 2001.
Phosphatidylinositol 4,5-bisphosphate and Arf6-regulated membrane traffic. Journal of
Cell Biology, 154, 1007-1018.
BURGWYN FUCHS, B., O'BRIEN, E., EL KHOURY, J. B. & MYLONAKIS, E. 2010.
Methods for using Galleria mellonella as a model host to study fungal infections.
Virulence, 1, 475-482.
CASAMAYOR, A. & SNYDER, M. 2002. Bud-site selection and cell polarity in budding yeast.
Current Opinion in Microbiology, 5, 179-186.
CASANOVA, J. E. 2007. Regulation of Arf activation: the Sec7 family of Guanine Nucleotide
exchange factors. Traffic.
CHANG, F., WOOLLARD, A. & NURSE, P. 1996. Isolation and characterization of fission
yeast mutants defective in the assembly and placement of the contractile actin ring.
Journal of Cell Science, 109, 131-142.
CHEN, H., KUO, C.-C., KANG, H., HOWELL, A. S., ZYLA, T. R., JIN, M. & LEW, D. J.
2012. Cdc42p regulation of the yeast formin Bni1p mediated by the effector Gic2p.
molecular Biology of the Cell, 23, 3814-3826.
CHERFILS, J. & CHARDIN, P. 1999. GEFs: structural basis for their activation of small GTP-
binding proteins. TRENDS in Biochemical Sciences, 24, 306-311.
CHUNG, J.-K., SEKIYA, F., KANG, H.-S., LEE, C., HAN, J.-S., KIM, S. R., BAE, Y. S.,
MORRIS, A. J. & RHEE, S. G. 1997. Synaptojanin inhibition of Phospholipase D activity
by hydrolysis of phosphatidylinositole 4,5-bisphosphate. The Journal of Biological
Chemistry, 272, 15980-15985.
CLARKE, L. & CARBON, J. 1980. Isolation of a yeast centromere and construction of
functional small circular chromosomes. Nature 287, 504-509.
CLARKE, L. & CARBON, J. 1985. The structure and function of yeast centromeres. Annual
Review of Genetics, 19, 29-56.
CONFALONIERIA, S. & DI FIOREA, P. P. 2002. The Eps15 homology (EH) domain. FEBS
letters, 513, 24-29.
CORBETT, K. D. & ALBER, T. 2001. The many faces of Ras: reconition of small GTP-bining
proteins. TRENDS in Biochemical Sciences, 26, 710-716.
COSTA, R. & AYSCOUGH, K. R. 2005. Interactions between Sla1p, Lsb5p and Arf3p in yeast
endocytosis. Biochemical Society Transactions, 33, 1273-1275.
COSTA, R., WARREN, D. T. & AYSCOUGH, K. R. 2005. Lsb5p interacts with actin regulators
Sla1p and Las17p, ubiquitinin and Arf3p to couple actin dynamics to membrane trafficing
processes. Biochemical Journal, 387, 649-658.
CROWTHER, R. A. & PEARSE, M. F. 1981. Assembly and packaging of clathrin into coats.
Journal of Cell Biology, 91, 790-797.
CUKIERMAN, E., HUBER, I., ROTMAN, M. & CASSEL, D. 1995. The ARF1 GTPase-
activating protein: zinc finger motif and Golgi complex localization. Science, 270, 1999-
2002.
D’SOUZA-SCHOREY, C. & CHAVRIER, P. 2006. ARF proteins: roles in membrane traffic and
beyond. Nature reviews-Molecular Cell Biology, 7, 347.

Chapter 7: References
75
DEAN, R., VAN KAN, J. A. L., PRETORIUS, Z. A., HAMMOND-KOSACK, K. E., DI
PIETRO, A., SPANU, P. D., RUDD, J. J., DICKMAN, M., KAHMANN, R., ELLIS, J. &
FOSTER, G. D. 2012. The top 10 fungal pathogens in molecular plant pathology.
Molecular Plant Pathology, 13, 414-430.
DELGADO-ALVAREZ, D. L., CALLEJAS-NEGRETE, O. A., GÓMEZ, N., FREITAG, M.,
ROBERSON, R. W., SMITH, L. G. & MOURIÑO-PÉREZ, R. R. 2010. Visualization of
F-actin localization and dynamics with live cell markers in Neurospora crassa. Fungal
Genetics and Biology, 47, 573-586.
DESRIVIÈRES, S., COOKE, F. T., PARKER, P. J. & HALL, M. N. 1998. MSS4, a
Phosphatidylinositol-4-5-phosphate 5-kinase required for organization of the actin
cytoskeleton. The Journal of Biological Chemistry, 273, 15787-15793.
DI PIETRO, A. & RONCERO, M. I. G. 1998. Cloning, expression and role in pathogenicity of
pg1 encoding the major extracellular endopolygalacturonase of the vascular wilt pathogen
Fusarium oxysporum. Molecular Plant-Microbe Interactions, 11, 91-98.
DIETRICH, F. S., VOEGELI, S., BRACHAT, S., LERCH, A., GATES, K., STEINER, S.,
MOHR, C., PÖHLMANN, R., LUEDI, P., CHOI, S., WING, R. A., FLAVIER, A.,
GAFFNEY, T. D. & PHILLIPSEN, P. 2004. The Ashbya gossypii genome as a tool for
mapping the ancient Saccharomyces cereviciae genome. Science, 304, 304-307.
DIETRICH, F. S., VOEGELI, S., KUO, S. & PHILLIPSEN, P. 2013. Genomes of ashbya fungi
isolated from insects reveal four mating-type loci, numerous translocations, lack of
transposons, and distinct gene duplications. G3, 3, 1225-1239.
DOWER, W. J., MILLER, J. F. & RAGSDALE, C. W. 1988. High efficiency transformation of
E. coli by high voltage electroporation. Nucleic Acid Research, 16, 6127-6145.
DÜNKLER, A. & WENDLAND, J. 2007. Use of MET3 promoters for regulated gene expression
in Ashbya gossypii. Current Genetics, 52, 1-10.
EAST, M. P. & KAHN, R. A. 2011. Models for the functions of Arf GAPs. Seminars in Cell &
Developmental Biology 22 (2011) 3–9, 22, 3-9.
EVANGELISTA, M., BLUNDELL, K., LONGTINE, M. S., CHOW, C. J., ADAMES, N.,
PRINGLE, J. R., PETER, M. & BOONE, C. 1997. Bni1p, a yeast formin linking Cdc42p
and the actin cytoskeleton during polarized morphogenesis. Science, 276, 118-122.
EVANGELISTA, M., PRUYNE, D., AMBERG, D. C., BOONE, C. & BRETSCHER, A. 2001.
Formins direct Arp2/3-independent actin filament assembly to polarize cell growth in
yeast. Nature Cell Biology, 4, 260-269.
FISCHER-PARTON, S., PARTON, R. M., HICKEY, P. C., DIJKSTERHUIS, J., ATKINSON,
H. A. & READ, N. D. 2000. Confocal microscopy of FM4-64 as a tool for analysing
endocytosis and vesicle trafficking in living fungal hyphae. Journal of Microscopy, 198,
246-259.
FISCHER, R., ZEKERT, N. & TAKESHITA, N. 2008. Polarized growth in fungi -interplay
between the cytoskeleton, positional markers and membrane domains. Molecular
Microbiology, 68, 813-826.
FRANCO, M., PETERS, P. J., BORETTO, J., VAN DONSELAAR, E., NERI, A., D'SOUZA-
SCHOREY, C. & CHAVRIER, P. 1999. EFA6, a sec7 domain -containing exchange
factor for ARF6, coordinates membranerecycling and actin cytoskeleton organization. The
EMBO Journal, 18, 1480-1491.
GAIDAROV, I. & KEEN, J. H. 1999. Phosphoimositide-Ap-2 interactions required for targeting
plasma membrane chlathrin-coated pits. Journal of Cell Biology, 148, 755-764.

Chapter 7: References
76
GAMMIE, A. E., KURIHARA, L. J., VALLEE, R. B. & ROSE, M. D. 1995. DNM1, a
Dynamin-related gene, participates in endosomal trafficing in yeast. The Journal of Cell
Biology, 130, 553-566.
GASTMANN, S., DÜNKLER , A., WALTHER, A., KLEIN, K. & WENDLAND, J. 2007. A
molecular toolbox for manipulating Eremothecium coryli. Microbiological Research, 162,
299-307.
GATTIKER, A., RISCHATSCH, R., DEMOUGIN, P., VOEGELI, S., DIETRICH, F. S.,
PHILLIPSEN, P. & PRIMIG, M. 2007. Ashbya genome database 3.0: a cross-species
genome and transcriptome browser for yeast biologists. BMC Genomics, 8.
GIBEAUX, R., LANG, C., POLITI, A. Z., JASPERSEN, S. L., PHILIPPSEN, P. & ANTHONY,
C. 2012. Electron tomography of the microtubule cytoskeleton in the multinucleated
hyphae of Ashbya gossypii. Journal of Cell Science, 125, 5830-5839.
GIDEON, P., JOHN, J., FRECH, M., LAUTWEIN, A., CLARK, R., SCHEFFLER, J. E. &
WITTINGHOFER, A. 1992. Mutational and kinetic analyses of the GTPase-activating
protein (GAP)-p21 interaction: The C-terminal domain of GAP is not sufficient for full
activity. Molecular and Cellular Biology, 12, 2050-2056.
GIETZ, D. & WOODS, R. A. 2002. Transformation of yeast by lithium acetate/single-stranded
carrier DNA/polyethylene glycol method. Methods in Enzymology, 350, 87-96.
GILLINGHAM, A. K. & MUNRO, S. 2007a. Identification of a Guanine Nucleotide Exchange
Factor for Arf3, the Yeast Orthologue of Mammalian Arf6. PLoS one, 2, 1-9.
GILLINGHAM, A. K. & MUNRO, S. 2007b. The small G proteins of the Arf family and their
regulators. Annual Review of Cell Developmental Biology, 23, 579-611.
GIRBARDT, M. 1957. Der Spitzenkörper von Polystictus versicolor. Planta, 50, 47-50.
GOLEY, E. D. & WELCH, M. D. 2006. The ARP2/3 complex: an actin nucleator comes of age.
Nature Reviews - Molecular Cell Biology, 7, 713-726.
GROVE, S. N. & BRACKER, C. E. 1970. Protoplasmic organization of hyphal tips among
fungi: vesicles and Spitzenkörper. Journal of Bacteriology, 104, 989-1009.
GRÜNLER, A. 2010. Molecular biological analysis of cell adhesion in Ashbya gossypii. Phd
Thesis, Friedrich-Schiller-Universität Jena Germany.
GRÜNLER, A., WALTHER, A., LÄMMEL, J. & WENDLAND, J. 2010. Analysis of flocculins
in Ashbya gossypii reveals FIG2 regulation by TEC1. Fungal Genetics and Biology, 47,
619-628.
HARRIS, T. W., HARTWIEG, E., HOROVITZ, H. R. & JORGENSEN, E. M. 2000. Mutations
in Synaptojanin disrupt synaptic vesicle recycling. Journal of Cell Biology, 150, 589-600.
HEATH , R. J. W. & INSALL, R. H. 2008. F-BAR domains: multifunctional regulators of
membrane curvature. Journal of Cell Science, 121, 1951-1954.
HEDGES, S. B. 2002. The origin and evolution of model orgamisms. Nature, 3, 838-849.
HIGGINS, M. K. & MCMAHON, H. T. 2002. Snap-shots of clathrin-mediated endocytosis.
TRENDS in Biochemical Sciences, 27, 257-263.
HILPELÄ, P., VARTIAINEN, M. K. & LAPPALAINEN, P. 2004. Regulation of the actin
cytoskeleton by PI(4,5)P2 and PI(4,5)P3. Current Topics in Microbiological Immunology,
282, 117-163.
HOFMANN, K. & BUCHER, P. 1996. The UBA domain: a sequence motif prescent in multiple
enzyme classes of the ubiquitination pathway. TRENDS in Biochemical Sciences, 21, 172-
173.
HONDA, A., NOGAMI, M., YOKOZEKI, T., YAMAZAKI, M., NAKAMURA, H.,
WATANABE, H., KAWAMOTO, K., NAKAYAMA, K., MORRIS, A. J., FROHMAN,

Chapter 7: References
77
M. A. & KANAHO, Y. 1999. Phosphatidylinositol 4-phosphate 5-kinase alpha is a
downstream effector of the small G protein ARF6 in membrane ruffle formation. Cell, 99,
521-532.
HOWARD, R. J. & AIST, J. R. 1980. Cytoplasmic microtubules and fungal morphogenesis:
ultrastructural effects of methyl benzimidazole-2-ylcarbamate determines by freeze-
substitution of hyphal tip cells. Journal of Cell Biology, 87, 55-64.
HUANG, C.-F., LIU, Y.-W., TUNG, L., LIN, C.-H. & LEE, F.-J., S 2003. Role for Arf3p in
Development of Polarity, but Not Endocytosis, in Saccharomyces cerevisiae. Molecular
Biology of the Cell, 14, 3834-3847.
ISH-HOROWICZ, D. & BURKE, J. F. 1981. Rapid and efficient cosmid cloning. Nucleic Acid
Research, 9, 2989-2998.
ITZEN, A. & GOODY, R. S. 2011. GTPases involved in vesicular trafficking: Structures and
mechanisms. Seminars in Cell & Developmental Biology, 22, 48-56.
JANMEY, P. A., IIDA, K., YIN, H. L. & STOSSEL, T. P. 1987. Phosphatidylinositise micelles
and phosphoinositide-containing vesicles dissociate endogenous gesolin-acting complexes
and promote actin assembly from the fast-growing end of actin filaments blocked by
gesolin. Journal of biological chemistry, 262, 12228-12236.
JORDE, S., WALTHER, A. & WENDLAND, J. 2011. The Ashbya gossypii fimbrin SAC6 is
requiredfor fast polarizedhyphaltipgrowthandendocytosis. Microbiological Research.
KAKSONEN, M., SUN, Y. & DRUBIN, D. G. 2003. A Pathway for Association of Receptors,
Adaptors, and Actin during Endocytic Internalization. Cell, 115, 475-487.
KATO, T. & PARK, E. Y. 2012. Riboflavin production by Ashbya gossypii. Biotechology letters,
34, 611-618.
KAWASHIMA, T., BERTHET-COLOMINAS, C., WULFF, M., CUSACK, S. & LEBERMAN,
R. 1996. The structure of the Escherichia coli EF-Tu.EF-Ts complex at 2.5A resolution.
Nature, 379, 511-518.
KIM, K., MCCULLY, M. E., BHATTACHARYA, N., BUTLER, B., SEPT, D. & COOPER, J.
A. 2007. Structure/function analysis of the interaction of phosphatidylinositol 4,5-
bisphosphae with actin-capping protein: implicatons forhow capping proteins binf the
actin filament. Journal of biological chemistry, 282, 5871-5879.
KIMURA, S., TOKUMARU, S. & KUGE, K. 2008. Eremothecium ashbyo causes soybean
yeast-spot and is assocuated with stink.bug Riptorus clavatus. Journal of General Plant
Pathology, 74, 275-280.
KIRBY, K. S. 1956. A new method for the isolation of ribonucleic acids from mammalian
tissues. Biochemical Journal, 64, 405-408.
KLINNER, U. & SCHÄFER, B. 2004. Genetic aspects of targeted insertion mutagenesis in
yeasts. FEMS Microbiology Reviews, 28, 201-223.
KNECHTLE, P., DIETRICH, F. S. & PHILLIPSEN, P. 2003. Maximal polar growth potential
depends on the polarisome component AgSpa2 in the filamnetous fungus Ashbya
gossypii. Molecular Biology of the Cell, 14, 4140-4154.
KRAWCHUK, M. & WAHLS, W. P. 1999. High-efficiency gene targeting in
Schizosaccharomyces pombe using a modlular, PCR-based approach with long tracts of
flanking homology. Yeast, 15, 1419-1427.
KURTZMAN, C. P. & ROBNETT, C. J. 2003. Phylogenetic relationships among yeasts of the
'Saccharomyces complex' determined from multigene sequence analyses. FEMS Yeast
Research, 3, 417-432.

Chapter 7: References
78
KÖHLER, T., WESCHE, S., TAHERI, N., BRAUS, G. H. & MÖSCH, H. U. 2002. Dual role of
the Saccharomyces cerevisiae TEA/ATTS family transcription factor Tec1p in regulation
of gene expression and cellular development. Eukaryotic cell, 1, 673-686.
LAMBERT, A. A., PERRON, M. P., LAVOIE, E. & PALLOTTA, D. 2007. The Saccharomyces
cerevisiae Arf3 protein is involved in actin cable and cortical patch formation. FEMS
Yeast Research, 7, 782–795.
LEE, F.-J. S., STEVENS, L. A., KAO, Y. L., MOSS, J. & VAUGHAN, M. 1994.
Characterization of a Glucose-repressible ADP-ribosylation Factor (ARF3) from
Saccharomyces cerevisiae. The Journal of Biological Chemistry, 269, 20931-20937.
LEE, W. L., BEZANILLA, M. & POLLARD, T. D. 2000. Fission yeast myosin-1, Myo1p,
stimulates actin assembly by Arp2/3 complex and shares function with WASp. Journal of
Cell Biology, 15, 789-800.
LI, F. & HIGGS, H., N 2003. The mouse formin mDia1 is a potent actin nucleation factor
regulated by autoinhibition. Current Biology, 13, 1335-1340.
LICHIUS, A., BEREPIKI, A. & READ, N. D. 2011. Form follows function -The versatile fungal
cytoskeleton. Fungal Biology, 115, 518-540.
LIONAKIS, M. S. 2011. Drosophila and Galleria insect model hosts. Virulence, 2, 521-527.
MACHESKY, L. M., ATKINSON, S. J., AMPE, C., VANDEKERCKHOVE, J. & POLLARD,
T. D. 1994. Purification of a cortical complex containing two unconventionaö actins from
Achanthamoeba by affinity chromatography on profilin-agarose. Journal of Cell Biology,
127, 107-115.
MACHESKY, L. M. & INSALL, R. H. 1998. Scar1 and the related Wiscott-Aldrich syndrome
protein WASP, regulate the actin cytoskeleton through the Arf2/3 complex. Current
Biology, 8, 1347-1356.
MAYINGER, P. 2012. Phosphoinositides and vesicular membrane traffic. Biochemica et
Biophysica Acta, 1821, 1104-1113.
MCCANN, R. O. & CRAIG, S. W. 1997. TheI/LWEQ module: a conserved aequence that
sigifies F-actin binding in functionally diverse proteins from yeast to mammals. Proc Natl
Acad Sci U S A, 94, 5679-5684.
MCMAHON, H. T. & BOUCROT, E. 2011. Molecular mechanism and physiological functions
of clathrin-mediated endocytosis. Nature reviews-Molecular Cell Biology, 12, 517-533.
MEDRANO, E. G., ESQUIVEL, J., BELL, A., GREENE, J., ROBERTS, P., BACHELER, J.,
MAROIS, J., WERIGHT, D., NICHOLS, R. & LOPEZ, J. 2009. Potential for Nezara
viridula (Hemiptera: Pentatomidae) to transmit bacterial and funagl pathogens in cotton.
Current Microbiology, 59, 405-412.
MIKI, H., MIURA, K. & TAKENAWA, T. 1996. N-WASP, a novel actin-depolymerizing
protein, regulates the cortical cytoskeletal rearrangement in a PIP2-dependent manner
downstream of tyrosine kinases. EMBO J, 15, 5326-5335.
MITSUI, K., YAGUCHI, S.-I. & TSURUGI, K. 1994. The GTS1 Gene, Which Contains a Gly-
Thr Repeat, Affects the Timing of Budding and Cell Size of the Yeast Saccharomyces
cerevisiae. Molecular and Cellular Biology, 14, 5569-5578.
MITTAL, R., AHMADIAN, M. R., GOODY, R. S. & WITTINGHOFER, A. 1996. Formation of
a transition-state analog of the Ras GTPase reacyion by Ras-GDP, Tetrafluoroaluminate
and GTPase-activating proteins. Science, 273, 115-117.
MURRAY, A. W. & SZOSTAK, J. W. 1983. Pedigree analysis of plasmid segregation in yeast.
Cell, 34, 961-970.

Chapter 7: References
79
MUSTACCHI, R., HOHMANN, S. & NIELSEN, J. 2006. Yeast systems biology to unravel the
network of life. Yeast, 23, 227-238.
NAVARRO-VELASCO, G. Y., PRADOS-ROSALES, R. C., ORTIS-URQUIZA, A.,
QUESADA-MORAGA, E. & DI PIETRO, A. 2011. Galleria mellonella as model host for
the trans-kingdom pathogen Fusarium oxysporum. Fungal Genetics and Biology, 48,
1124-1129.
NOBLE, S. M. & JOHNSON, A. D. 2005. Strains and strategies for large-scale gene deletion
studies of the diploid human pathogen Candida albicans. Eukaryotic cell, 4, 298-309.
OBERHOLZER, U., MARCIL, A., LEBERER, E., THOMAS, D. Y. & WHITEWAY, M. 2002.
Myosin I is required for hypha formationin Candida albicans. Eukaryotic cell, 1, 213-228.
OUDE WEERNINK, P. A., LÓPEZ DE JESÚS, M. & SCHMIDT, M. 2007. Phospholipase D
signalliing: orchestration by PIP2 and small GTPases. Naunyn Schmiedebergs Arch
Pharmacol., 374, 399-411.
PALEOTTI, O., MACIA, E., LUTON, F., KLEIN, S., PARTISANI, M., CHARDIN, P.,
KIRCHHAUSEN, T. & FRANCO, M. 2005. The small G-protein Arf6GTP recruits the
AP-2 adaptor complex to membranes. Journal of biological chemistry, 208, 21661-21666.
PARK, H.-O. & BI, E. 2007. Central roles of small GTPases in the development of cell polarity
in yeast and beyond. Microbiology and Molecular Biology Reviews, 71, 48-96.
PÉREZ-NADALES, E. & DI PIETRO, A. 2011. The membrane mucin Msb2 regulates invasive
growth and plant infection in Fusarium oxysporum. The Plant Cell, 23, 1171-1185.
PFALLER, M. A. & DIEKEMA, D. J. 2007. Epidemiology of invasive candidiasis: a persisitent
public health problem. Clinical Microbiological Reviews, 20, 133-163.
PISHVAEE, B., COSTAGUTA, G., YEUNG, B. G., RYAZANTSEV, S., GREENER, T., L.E,
G., EISENBERG, E., MCCAFFERY, J. M. & PAYNE, G. S. 2000. A yeast DNA J
protein required for uncoating o clathrin coated vesicles in vivo. Nature Cell Biology, 2,
958-963.
POLLARD, T. D. 2007. Regulation of filament assembly by Arp2/3 complex and formins.
Annual Review of Biophysics and Biomolecular Structure, 36, 451-477.
PRILLINGER, H., SCHWEIGKOFLER, W., BREITENBACH, M., BRIZA, P.,
STAUDACHER, E., LOPANDIC, K., MOLNÁR, O., WEIGANG, F., IBL, M. &
ELLINGER, A. 1998. Phytopathogenic filamentous (Ashbya, Eremotecium) and
dimorphic fungi (Holleya, Nematospora) with needle-shaped ascospores as new members
within the Saccharomycetaceae. Yeast, 13, 945-960.
PRINGLE, J. R., PRESTON, R. A., ADAMS, A. E., STEARNS, T., DRUBIN, D. G., HAARER,
B. K. & JONES, E. W. 1989. Flourescence microscopy methods for yeast. Methods in
Cell Biology, 31, 357-435.
QUALMANN, B. & KESSELS, M. M. 2002. Endocytosis and the cytoskeleton. International
Review of Cytology, 220, 93-144.
RADHAKRISHNA, H., AL-AWAR, O., KHACHIKIAN, Z. & DONALDSON, J. G. 1999.
ARF6 requirement for Rac ruffling suggests a role form membrane trafficking in cortical
actin rearrangements. Journal of Cell Biology, 112, 855-866.
RATCLIFFE, N. A. 1985. Invertebarte immunity -a primer for the non-specialist. Immunology
Letters, 10, 235-270.
RENAULT, L., GUIBERT, B. & CHERFILS, J. 2003. Structural snapshots of the mechanism
and inhibition if a guanine nucleotide exchange factor. Nature, 426, 525-530.
RENSLAND, H., LAUTWEIN, A., WITTINGHOFER, A. & GOODY, R. S. 1991. Is there a
rate-limiting step before GTP cleavage by H-ras p-21. Biochemistry, 30, 11181-11185.

Chapter 7: References
80
RIDA, P. C. & SURANA, U. 2005. Cdc42-dependent localization of polarisome component
Spa2 to the incipient bud site is independent of the GDP/GTP exchange factor Cdc24.
European Journal of Cell Biology, 84, 939-949.
RISPAIL, N. & DI PIETRO, A. 2009. Fusarium oxysporum Ste12 controls invasive growth and
virulence downstream of the Fmk MAPK cascade. Molecular Plant-Microbe Interactions,
22, 830-839.
ROHATGI, R., MA, L., MIKI, H., LOPEZ, M., KIRCHHAUSEN, T., TAKENAWA, T. &
KIRSCHNER, M. W. 1999. The interaction between N-WASP and the Arp2/3 complex
links Cdc42-dependent signals to actin assembly. Cell, 97, 221-231.
SAMBROOK, J. & RUSSEL, D. W. 2001. Molecular cloning: a laboratory manual. Cold Spring
Harbor Laboratory Press, Cold Spring Harbor, NY.
SARASTE, M., SIBBALD, P. R. & WITTINGHOFER, A. 1990. The P-loop -a common motif in
ATP- and GTP-binding proteins. TRENDS in Biochemical Sciences, 15, 430-434.
SCHADE, D., WALTHER, A. & WENDLAND, J. 2003. The development of a transformation
system for the dimorphic plant pathogen Holleya sinecuada based on Ashbya gossypii
DNA elements. Fungal Genetics and Biology, 40, 65-71.
SCHEFFZEK, K., LAUTWEIN, A., KABSCH, W., AHMADIAN, M. R. & WITTINGHOFER,
A. 1996. Crystal structure of the GTPase-activating domain of human p120GAP and
implications for the interaction with Ras. Nature, 238, 591-596.
SCHMID, S. L. 1997. Clathrin-coated vesicle formation and protein sorting. Annual Review of
Biochemistry, 66, 511-548.
SHAPIRO, A. D., RIEDERER, M. A. & PFEFFER, S. R. 1993. Biochemical analysis of rab9, a
ras-like GTPase involved in protein transport from late endosomes to the tans Golgi
network. Journal of biological chemistry, 268, 6925-6931.
SHAW, B. D., CHUNG, D.-W., WANG, C.-L., QUINTANILLA, L. A. & UPADHYAY, S.
2011. A role for endocytic recycling in hyphal growth. Fungal Biology, 115, 541-546.
SHIMOMURA, O., JOHNSON, F. H. & SAIGA, Y. 1962. Extraction , purification and
properties of aequorin, a bioluminiscent protein from the luminous hydromedusan,
Aequorea. Journal of Cellular and Comparative Physiology, 59, 223-239.
SHORT, J. M., FERNANDEZ, J. M., SORGE, J. A. & HUSE, W. D. 1988. Lambda ZAP: a
bactereophage lamda expression vector with in vivo excision properties. Nucleic Acid
Research, 16, 7583-7600.
SIGRIST, C. J. A., DE CASTRO, E., CERUTTI, L., CUCHE, B. A., HULO, N., BRIDGE, A.,
BOUGUELERET, L. & XENARIOS, I. 2012. New and continuing developments at
PROSITE. Nucleic Acids Research.
SMACZYNSKA-DE ROOIJ, I. I., ALLWOOD, E. G., AGHAMOHAMMEDZADEH, S.,
HETTEMA, E. H., GOLDBERG, M. W. & AYSCOUGH, K. R. 2010. A role for the
dynaim-like protein Vps1 during endocytosis in yeast. Journal of Cell Science, 123, 3496-
3506.
SMACZYNSKA-DE ROOIJ, I. I., COSTA, R. & AYSCOUGH, K. R. 2008. Yeast Arf3p
modulates plasma membrane PtdIns(4,5)P2 levels to facilitate endocytosis. Traffic, 9,
559.
SPANG, A., SHIBA, Y. & RANDAZZO, P. A. 2010. Arf GAPs: Gatekeepers of vesicle
generation. FEBS letters, 584, 2646-2651.
STAHELIN, R. V., LONG, F., PETER, B. J., MURRAY, D., P., D. C., MCMAHON, H. T. &
CHO, W. 2003. Contrasting membrane interaction mechanisms of AP108 N-terminal

Chapter 7: References
81
homology (ANTH) and epsin N-erminal homology (ENTH) domains. The Journal of
Biological Chemistry, 278, 28993-28999.
STAHMANN, K. P., ARST, H. N. J., ALTHÖFER, H., REVUELTA, J. L., MONSCHAU, N.,
SCHLÜPEN, C., GÄTGENS, C., WIESENBURG, A. & SCHLÖSSER, T. 2001.
Riboflavin, overproduced during sporulation of Ashbya gossypii, protects its hyaline
spores against utraviolet light. Environmental Microbiology, 3, 545-550.
STEINER, S. & PHILIPPSEN, P. 1994. Sequence and promoter analysis of the highly expressed
TEF gene of the filamentous fungus Ashbya gossypii. Molecular and General Genetics,
242, 263-271.
STEINER, S., WENDLAND, J., WRIGHT, M. C. & PHILLIPSEN, P. 1995. Homologous
recombination as the main mechanism for DNA integration and cause of rearrangements
in the filamentous ascomycete Ashbya gossypii. Genetics, 140, 973-987.
STINCHCOMB, D. T., STRUHL, K. & DAVIS, R. W. 1979. Isolation and characterization of a
yeast chromosomal replicator. Nature, 282, 39-43.
SUDBERY, P. 2011. Flourescent proteins illustrate the structure and function of the hyphal tip
apparatus. Fungal Genetics and Biology, 48, 849-857.
SUN, Y., CARROLL, S., KAKSONEN, M., TOSHIMA, J. Y. & DRUBIN, D. G. 2007.
PtdIns(4,5)P2 turnover is required for multiple stages during clathrin-mediated and actin-
dependent endocytic internalization. Journal of Cell Biology, 177, 355-367.
TAHERI-TALESH, N., HORIO, T., ARAUJO-BAZÁN, L., DOU, X., ESPESO, E. A.,
PEÑALVA, M. A., OSMANI, S. A. & OAKLEY, B. R. 2008. The tip growth apparatus
of Aspergillus nidulans. Molecular Biology of the Cell, 19, 1439-1449.
TAKAI, Y., SASAKI, T. & MATOZAKI, T. 2001. Small GTP-binding proteins. Physiological
Rewiews, 81, 153-208.
TANG, H.-Y., XU, J. & CAI, M. 2000. Pan1p, End3p and Sla1p, three yeast proteins required for
normal cortcal actin cytoskeleton organization, associate with each other and play
essential roles in cell wall morphogenesis. Molecular and Cellular Biology, 20, 12-25.
TATEBAYASHI, K., KATO, J. & OIKEDA, H. 1994. Structural analyses of DNA fragments
integrated by illegitime recombination in Schizosaccharomyces pombe. molecular and
General Genetics, 244, 11-19.
TE HEESEN, H., GERWERT, K. & SCHLITTER, J. 2007. Role of the arginine finger in RAS-
RasGAP revealed by Qm/MM calculations. FEBS letters, 581, 5677-5684.
TORET, C. P., LEE, L., SEKIYA-KAWASAKI, M. & DRUBIN, D. G. 2008. Multiple Pathways
Regulate Endocytic Coat Disassembly in Saccharomyces cerevisiae for Optimal
Downstream Trafficking. Traffic, 9, 848-859.
TOSHIMA, J. Y., TOSHIMA, J., KAKSONEN, M., MARTIN, A. C., KING, D.S & DRUBIN,
D. G. 2006. Spatial dynamics of receptor-mediated endocytic trafficking in budding yeast
revealed by using flourescent alpha-factor derivatives. Proc Natl Acad Sci U S A, 103,
5793-5798.
TRAHEY, M. & MCCORMICK, F. 1987. A cytoplasmic protein stimulates normal N-ras p21
GTPase, but doea not effect oncogene mutants. Science, 238, 542-545.
TSAI, P.-C., LEE, S.-W., LIU, Y.-W., CHU, C.-W., CHEN, K.-Y., HO, J.-C. & LEE, F.-J. S.
2008. Afi1p Functions as an Arf3p Polarization-specific Docking Factor for Development
of Polarity. The Journal of Biological Chemistry, 283, 16915–16927.
UNGEWICKELL, E. & BRANTON, D. 1981. Assembly units of clathrin coats. Nature, 289.
UPHADYAY, S. & SHAW, B. D. 2008. The role of actin, fimbrin and endocytosis in growyj of
hyphae in Aspergillus nidulans. Molecular Microbiology, 68, 690-705.

Chapter 7: References
82
WACH, A., BRACHAT, A., PÖHLMANN, R. & PHILIPPSEN, P. 1994. New heterologous
modules for classical or PCR-based gene disruption in Saccharomyces cerevisiae. Yeast,
10.
WALTHER, A. & WENDLAND, J. 2003. Septation and cytokinesis in fungi. Fungal Genetics
and Biology, 40, 187-196.
WALTHER, A. & WENDLAND, J. 2004a. Apical localization of actin patches and vacuolar
dynamics in Ashbya gossypii depend on the WASP homolog Wal1p. Journal of Cell
Science, 117.
WALTHER, A. & WENDLAND, J. 2004b. Polarized hyphal growth in Candida albicans
requires the Wilscott-Aldricg syndrome protein homolog Wal1p. Eukaryotic cell, 3, .471–
482.
WANG, J., NEO, S. P. & CAI, M. 2009. Regulation of the yeast formin Bni1p by the actin-
regulating kinase Prk1p. Traffic, 10, 528-535.
WASSERSTROM, L., LENGELER, K. B., WALTHER, A. & WENDLAND, J. 2013. Molecular
determinants of sporulation in Ashbya gossypii. Genetics, 135, 87-99.
WENDLAND, B., STEECE, K. E. & EMR, S. D. 1999a. Yeast epsins contain an essential N-
terminal ENTH domain bind clathrin and are required for endocytosis. The EMBO
Journal, 18, 4383-4393.
WENDLAND, J., AYAD-DURIEUX, Y., KNECHTLE, P., C, R. & PHILLIPSEN, P. 2000.
PCR-based gene targeting in the filamentous fungus Ashbya gossypii. Gene, 242, 381-
391.
WENDLAND, J., PÖHLMANN, R., DIETRICH, F., STEINER, S., MOHR, C. & PHILIPPSEN,
P. 1999b. Compact ofganization if rRNA genes in the filamentous fungus Ashbya
gossypii. Current Genetics, 35, 618-625.
WENDLAND, J. & WALTHER, A. 2005a. Ashbya gossypii: a model for funga developmental
biology. Nature Reviews -Microbiology, 3, 421-429.
WENDLAND, J. & WALTHER, A. 2005b. Tip growth and endocytosis in fungi. Plant Cell
Monographs.
WENDLAND, J. & WALTHER, A. 2011. Genome evolution in the Eremotecium clade of the
Saccharomyces complex revealed by comparative genomics. Genes Genomes Genetics, 1,
539-548.
WENK, M. R. & DE CAMILLI, P. 2004. Protein.lipis interactions and phosphoinositide
metabolism in membrane traffick: insights from vesicle recycling in nerve terminals. Proc
Natl Acad Sci U S A, 101, 8262-8269.
WENNERBERG, K., ROSSMAN, K. L. & CHANNING, J. D. 2005. The Ras superfamily at a
glace. Journal of Cell Science, 118, 843-846.
VETTER, I. R. & WITTINGHOFER, A. 2001. The guanine nucleotide-binding switch in three
dimentsions. Science, 294, 1299-1304.
VIEIRA, J. & MEESING, J. 1982. The pUC plasmids, an M13mp7-derived system for insertion
mutagenesis and sequencing with synthetic universal primers. Gene, 19, 259-268.
WINTER, D., LECHLER, T. & LI, R. 1999. Activation of the yeast Arp2/3 complex by BEE1p,
a WASP-family protein. Current Biology, 9, 501-504.
WRIGHT, M. C. & PHILLIPSEN, P. 1991. Replicative transformation of the filamentous fungus
Ashbya gossypii with plasmids containing Saccharomyces cerevisiae ARS elements.
Gene, 109, 99-105.
XU, Z. & TSURUGI, K. 2007. Role of Gts1p in regulation of energy-metabolism oscillation in
continuous cultures of the yeast Saccharomyces cerevisiae. Yeast, 24, 161-170.

Chapter 7: References
83
YAGUCHI, S.-I., SHEN, H. & TSURUGI, K. 2007. Localization of Gts1p in cortical actin
patches of yeast and its possible role in endocytosis. European Journal of Cell Biology,
86, 275-285.
YEUNG, B. G., PHAN, H. L. & G.S., P. 1999. Adaptor complex-independent Clathrin function
in yeast. Molecular Biology of the Cell, 10, 3643-3659.
YOUN, J.-Y., FRIESEN, H., KISHIMOTO, T., W.M., H., KURAT, C. F., YE, W.,
CECCARELLI, D. F., SICHERI, F., KOHLWEIN, S. D., MCMAHON, H. T. &
ANDREWS, B. J. 2010. Dissecting BAR domain function in the yeast amphiphysins
Rvs161 and Rvs167 during endocytosis. Molecular Biology of the Cell, 21, 3054-3069.
ZHONG, M. J., CHEN-HWANG, M. C. & HWANG, Y. W. 1995. Switching nucleotide
specificity of Ha-Ras p21 by a single amino acid substitution at aspartate 119. Journal of
biological chemistry, 270, 10002-10117.
ZURITA, A., ZHANG , Y., DARDEN, T. & BIRNBAUMER, L. 2010. Obligatory role in GTP
hydrolysis for the amide carbonyl oxygen of the Mg(2+)-coordinating Thr of regulatory
GTPases. Proc Natl Acad Sci U S A, 107, 9596-9601.

Appendix I: Strains
84
Appendix I: Strains Strain Genotype Source Derivate of
Ashbya gossypii ATCC10895 wt Ashby and Novell, 1926 - Agleu2 wt (leu2) Mohr and Philippsen ATCC10895 ALK17-1 gts1::GEN3, leu2 Lengeler, this study Agleu2 ALK17-2 gts1::GEN3, leu2 Lengeler, this study Agleu2 ATO010 arf3::GEN3, leu2 This study Agleu2 ATO012 arf3::GEN3, leu2 This study Agleu2 ATO002 yel1::GEN3, leu2 This study Agleu2 ATO004 yel1::GEN3, leu2 This study Agleu2 ATO005 gts1::GEN3/GTS1, leu2 This study ALK17-1 ATO006 gts1::GEN3/GTS1, leu2 This study ALK17-2 ATO007 gts1::GEN3/GTS1-GFP, leu2 This study ALK17-1 ATO013 gts1::GEN3/GTS1(ArfGAP),
leu2 This study ALK17-1
ATO014 gts1::GEN3/GTS1(ArfGAP), leu2
This study ALK17-2
ATO017 gts1::GEN3/GTS1(UBA), leu2 This study ALK17-1 ATO018 gts1::GEN3/GTS1(UBA), leu2 This study ALK17-2 ATO015 arf3::GEN3/ARF3, leu2 This study ATO010 ATO016 arf3::GEN3/ARF3, leu2 This study ATO012 AWE37 wal1::KanMX6, leu2 Walther and Wendland
2004 Agleu2
ASJ22 sac6::KanMX6, leu2 Jorde et al., 2011 Agleu2
Eremothecium cymbalariae DBVPG7215 wt Kurtzman - ETO12 tec1::GEN3 This study DBVPG7215 ETO13 tec1::GEN3 This study DBVPG7215 ETO14 TEC1::KanMX This study DBVPG7215 ETO15 TEC1::KanMX This study DBVPG7215 ETO16 ecym5230::GEN3 This study DBVPG7215 ETO17 ecym5230::GEN3 This study DBVPG7215 ETO18 pHC-NatMX-KanMX This study DBVPG7215 ETO19 pLC-KanMX-LacZ This study DBVPG7215 ETO20 pFA-EcCEN1 This study DBVPG7215 ETO21 pFA-EcCEN5 This study DBVPG7215 Saccharomyces cerevisiae BY4749 MATa, his3, leu2, met15, ura3 Euroscarf CEN.PK2 MATa/α, his3, leu2, trp1, ura3 Hoepfner et al, 2000 F234 Carlsberg brewery strain Carlsberg YTO001 pRS418-MoGFP-Nat1 This study BY4749 YTO002 pRS418-AgTEFp-GTS1-GFP This study BY4749 YTO003 pRS418-AgTEFp-GTS1-GFP This study CEN.PK2 YTO004 pRS418-AgTEFp-GTS1-GFP This study F234 Fusarium oxysporum f. sp. Lycopersici FGSC 9935 wt O'Donnell et al. 2004 FGSC 9935

Appendix II: Primers
85
Appendix II: Primers Primer No.
Primer name Primer sequence 5’-3’
392 5’-GFP cataaccttcgggcatggcatc 511 AgTEF-prom aggatttgccactgaggttcttc 529 pFAup ttgtgcgttagaacgcggctac 530 pFAdo tgcaggttaacctggcttatc 1112 G2new gccagtttagtctgaccatc 1113 G3new tcgcagaccgataccagg 1202 N2new gcgtttccctgctcgcaggtc 1214 ScTEFp-up1 gggtaatttgtcgcggtctggg 1215 ScTEFt-down1 gcccatcagattgatgtcctcc 4321 G1-ACL055W cgatcacgtgacaatgcaac 4322 G4-ACL055W gtcctactactaggtggcg
4323 S1-ACL055W CAGCAAGAGGTAAAGCCAGAGAACGGTGGGCTTCGACGGACA
GGCgaagcttcgtacgctgcaggtc
4324 S2-ACL055W TCATGGCTCTAACTGAGCCCCTGATGAGATGTGCTTGTGGCGT
CActgatatcatcgatgaattcgag 4325 I1-ACL055W ctcgctgaatagcgccag 4326 I2-ACL055W cctgctgtaaagagccatg 4800 3'-Ecym_CEN1 CTTCTTactagtcgctaactctcgtgccattgg 4801 5'-Ecym_CEN1 AAACTAgagctcctgtccctgaccttgacg 4817 3'-Ecym_CEN5 CTTCTTactagtctattccataaactttctataacgg 4832 G1-EcymTEC1 cattgcagcagcagcagcaag 4833 G4-EcymTEC1 ggtaacggttcagtacagtg 4834 I1-EcymTEC1 cttcgggcacggatcattcgg 4835 I2-EcymTEC1 ggcgtactacgctcgctac 5030 CEN5-1 AAACTAgagctcccgtctgggcagttgatatgag
6062 pAGlib_swap1
TATGCGGCATCAGAGCAGATTGTACTGAGAGTGCACCATACCACgaagcttcgtacgctgcaggtc
6063 pAGlib_swap2
AATTTGTGAGTTTAGTATACATGCATTTACTTATAATACAGTtttttaggggcagggcatgctcatg
6180 AgGTS1-GFP-S1
CCAACGGCCTCGCAGGGCGCTTATCCTGGCTACAACTACCAGATgggtgctggcgcaggtgcttc
6181 AgGTS1-GFP-S2
GTTACGTTTGTTTCACGTGGTTTTTAATTACGTACTGATACAGAAtctgatatcatcgatgaattc
6195 S1-ACL078W
AGACTACCGAAGTCTAGCGGCGTGGGCAAGATCACAGACGGCGCAgaagcttcgtacgctgcaggtc
6196 S2-ACL078W
CACGGCCGCGCCGGCCCCGCTCTACTGTTACAAGCCCGCGCAGGCctgatatcatcgatgaattcgag
6197 G1-ACL078W ttggacgatctacggtgtacg 6198 G4-ACL078W accgacaacaacaacctgctc 6199 I1-ACL078W agttcaacatgtgggacgttgg 6200 I2-ACL078W agctggttgttcaggttgtcg 6201 S1-ABR218C GAGCTACTGTTTCCTGGGGAGACAGAGTAAGGCGCACGCCGA
TATgaagcttcgtacgctgcaggtc 6202 S2-ABR218C CTTTGGTGGTACATAGGAATGATACATAATGGAAACTCAGGTT
CGctgatatcatcgatgaattcgag 6203 G1-ABR218C cattctcgtggaagacacacc 6204 G4-ABR218C cggtcagcatgtgtaacatgc 6205 I1-ABR218C acgaacgacaacagccatagc 6206 I2-ABR218C caagttgtctggctgttgctc

Appendix II: Primers
86
Primer No.
Primer name Primer sequence 5’-3’
6327 G1-Ecym5230 ctgtctatatataccaagag 6328 G4-Ecym5230 ctgctctctgtgtaactgcg
6329 S1-Ecym5230
AGACACAACCAACACTCTATATATATATATACACTCTCACCCATATACGACGCCGCCGCACACTGCATCGACTCTGACATCTAAAAACACAAATCAAACAgaagcttcgtacgctgcaggtc
6330 S2-Ecym5230
GGCCACCCGGCAACGCCACTACTACTATGTTCTAATATATGCTTCGTTCGCCATGTATTGTGTGTACGATACACCAACATGCAACACCGGGGCTACTTATctgatatcatcgatgaattcgag
6331 I1-Ecym5230 gttgcgctgcattgcgctg 6332 I2-Ecym5230 cacccggcaacgccactac 6333 S1-EcTEC1 ATGATGAATGAAGAAGGAGCAGGGGCTAGGTTTGAGAATCTGT
TTAATTCGCAGCTGTATTACCAGCAGCCAGGAGGAGGGGGGGGTGGGGATGGTCAGGgaagcttcgtacgctgcaggtc
6334 S2-EcTEC1 TTATCCTGACCGAGAGGCGTCATCAGAATATTCTTGAGCTTCAAGAGAAACAACGGTCTGCTGCTGTGGGAGCGGTATGCTCTGTTGGGGGGACACATGCctgatatcatcgatgaattcgag
6336 3'-EcTEC1-XbaI ctgtcatctagacatattccaacggtgttgtagg
6338 5'-GTS1fwd-XhoI ctgtcactcgaggatactaaagcaccatgcgc 6339 5'-GTS1rev-XbaI ctgtcatctagactagctattcagcgagctgtag 6340 3'-GTS1fwd-XhoI ctgtcactcgagatgtacagctcgctgaatagcg 6341 3'-GTS1rev-XbaI ctgtcatctagactacatctggtagttgtagc
Lower case letters represent homology region to PCR target sequence. Upper case letters represent bases for homologous recombination. Italics/underlined represent restriction sites. Bold letters represent added codons.

Appendix III: Plasmids
87
Appendix III: Plasmids Plasmid No.
Plasmid Name Marker(s) Source
120 pSK+Bluescript Amp (Short et al., 1988) 121 pFA-KanMX6 Amp, Kan (Wach et al., 1994) 150 pRS415-KanMX Amp, Kan (Schade et al., 2003) 651 pRS417-AgTEFp-LacZ Amp, GEN3 (Dünkler and Wendland, 2007) 690 pRS417-AgTEFp-GFP Amp, GEN3 (Dünkler and Wendland, 2007) 706 pHC-Shuttle-AgTEFp-StlacZ Amp, Kan (Gastmann et al., 2007) 738 pLC-Shuttle-KanMX-NATMX3 Amp, Nat, Kan (Gastmann et al., 2007) 840 pFA-AgNATMX4 Amp, Nat Lab collection C169 pRS417-ScTEFp-LacZ Amp, GEN3 Lab collection C455 pRS418-AgTEFp-LacZ Amp, Nat Lab collection C470 pFA-MoGFP-Nat1 Amp, Nat Lab collection C682 pFA-EcCEN1 Amp, Kan This study C686 pFA-EcCEN5 Amp, Kan This study C781 pAG19275(GTS1)-NatMX Amp, Nat This study C783 pAG17522(ARF3)-URA3 Amp, URA3 (Wendland et al., 1999b) C787 pAG19275(GTS1-GFP) Amp, Nat This study C832 pRS418-AgTEFp-GTS1(ArfGAP) Amp, Nat This study C834 pSK+EcymTEC1 Amp This study C841 pSK+EcymTEC1::KanMX(fwd) Amp, Kan This study C842 pSK+EcymTEC1::KanMX(rev) Amp, Kan This study C871 pRS417-AgTEFp-(GTS1-GFP) Amp, Kan This study C936 pAG19275(GTS1)-URA3 Amp, URA3 (Wendland et al., 1999b) C938 pRS418-AgTEFp-GTS1(UBA) Amp, Nat This study C943 pAG17522(ARF3)-NatMX Amp, Nat This study