Embed Size (px)
Citation preview

Fetal Neurobehavioral Development: A Tale of Two Cities
Janet A. DiPietro and Laura E. CaulfieldJohns Hopkins University
Kathleen A. CostiganJohns Hopkins Medical Institutions
Mario Merialdi and Ruby H. N. NguyenJohns Hopkins University
Nelly ZavaletaInstituto de Investigacion Nutricional
Edith D. GurewitschJohns Hopkins Medical Institutions
Longitudinal neurobehavioral development was examined in 237 fetuses of low-risk pregnancies from 2distinct populations—Baltimore, Maryland, and Lima, Peru—at 20, 24, 28, 32, 36, and 38 weeksgestation. Data were based on digitized Doppler-based fetal heart rate (FHR) and fetal movement (FM).In both groups, FHR declined while variability, episodic accelerations, and FM–FHR coupling increased,with discontinuities evident between 28 and 32 weeks gestation. Fetuses in Lima had higher FHR andlower variability, accelerations, and FM–FHR coupling. Declines in trajectories were typically observed1 month sooner in Lima, which magnified these disparities. Motor activity differences were lessconsistent. No sex differences in fetal neurobehaviors were detected. It is concluded that populationfactors can influence the developmental niche of the fetus.
Most of the vast amount of study on the physical and mental devel-opment of children has begun with birth, more or less completelyneglecting the possible influence of environment during the period ofintra-uterine development. (Sontag & Wallace, 1934, p. 1050)
Consideration of the environmental context provided by thepregnant woman to her developing fetus remains as vanguard aconcept today as it was in the 1930s for the investigators in the
Fels Longitudinal Study. Well-established models of child devel-opment after birth include both proximal and distal influences ondevelopment, ranging from biological factors to parental caregiv-ing to the social and political environment (Pachter & Harwood,1996). The developmental niche of a given child reflects theparticular setting for development within these multiple levels ofinfluence (Super & Harkness, 1986). Cross-cultural research hasbeen used to distinguish biological, maturative processes fromthose that are guided by environmental influences. Such studiestypically compare cultures that broadly differ on many dimensions,including ethnicity, socioeconomic status, and general living con-ditions (Freedman & DeBoer, 1979). Cross-cultural differences inneonatal neurobehaviors have been observed (Choi & Hamilton,1986; Coll, Spekoski, & Lester, 1981; Eishima, 1992; Keefer,Tronick, Dixon, & Brazelton, 1982)—as have differences in therate and proficiency of motor development in the first few years oflife—between, for example, infants in the United States and Brazil(Santos, Gabbard, & Goncalves, 2001), Paraguay (Kaplan & Dove,1987), and the Yucatan (Solomons & Solomons, 1975). Althoughsome variation in motor development has been attributed to vari-ations in aspects of the physical or social environment, in general,the sources of these differences are poorly understood.
The uterus is the developmental niche of the fetus. In contrast toother periods of development, there has been little coherent con-ceptualization of the environmental factors that may influence fetalneurobehavioral development as it unfolds. Despite the acknowl-edgement that both environmental and biological processes serveas sources of variation in postnatal development, there is a ten-dency to regard differences that are apparent at birth, or before, asreflecting genetic influences alone. However, there is increasingrecognition of the influence of the maternal womb environment on
Janet A. DiPietro, Department of Population and Family Health Sci-ences, Johns Hopkins University; Laura E. Caulfield and Mario Merialdi,Department of International Health, Johns Hopkins University; KathleenA. Costigan and Edith D. Gurewitsch, Division of Maternal–Fetal Medi-cine, Johns Hopkins Medical Institutions; Ruby H. N. Nguyen, Departmentof Epidemiology, Johns Hopkins University; Nelly Zavaleta, Instituto deInvestigacion Nutricional, Lima, Peru.
The U.S.-based study was supported by National Institute of ChildHealth and Human Development (NICHD) Grant R01 HD27592, awardedto Janet A. DiPietro. The Peru study was conducted through a grant fromthe Nestle Research Foundation, and analysis was supported by NICHDGrant R01 HD42675, both awarded to Laura E. Caulfield. The ConsiglioNazionale delle Richerche of Italy provided partial support for MarioMerialdi.
We thank the Division of Maternal–Fetal Medicine of the Johns Hop-kins Medical Institutions and the staff of Hospital San Jose and the regionalhealth authorities of Southern Lima. We greatly appreciate the statisticalsupport provided by Sterling Hilton. Most of all, we are grateful for thediligent and generous participation of our study families, without whomthis research would not have been possible.
Correspondence concerning this article should be addressed to Janet A.DiPietro, Department of Population and Family Health Sciences, JohnsHopkins Bloomberg School of Public Health, 624 North Broadway, Bal-timore, MD 21205. E-mail: [email protected]
Developmental Psychology Copyright 2004 by the American Psychological Association2004, Vol. 40, No. 3, 445–456 0012-1649/04/$12.00 DOI: 10.1037/0012-1649.40.3.445
445

constitutional factors (Devlin, Daniels, & Roeder, 1997). Theperspective that the sequence and timing of prenatal developmentare universal phenomena is reflected in the small sample sizes (i.e.,often 20 or fewer participants) of most studies of fetal neurobe-havioral development. Few studies consider the sociodemographiccharacteristics of participants despite evidence for social classeffects on fetal neurobehavioral development in at-risk (M. John-son et al., 1992) and normal (DiPietro, Costigan, Shupe, Pressman,& Johnson, 1998) pregnancies in the United States. Knowledgeabout fetal development has been generated predominantly fromsamples of pregnant middle-class women from developed coun-tries with the assumption that these findings reflect universalantenatal processes.
Interest in antenatal development has burgeoned with theknowledge that processes beginning before birth contribute to laterhealth in general (Barker, 1995; Gillman & Rich-Edwards, 2000;Phillips, 2001) and to neurological disorders such as cerebral palsyin particular (Lamb & Lang, 1992; Nelson & Ellenberg, 1986).The view that fetal neurobehaviors reflect the developing nervoussystem within normal populations has received support from manysources (DiPietro, Irizarry, Hawkins, Costigan, & Pressman, 2001;Hepper, 1995; I. Nijhuis & ten Hof, 1999; J. G. Nijhuis, 1986;Sandman, Wadhwa, Hetrick, Porto, & Peeke, 1997). In addition,neurobehavioral development is affected in fetuses with congenitalanomalies (Hepper & Shahidullah, 1992; Horimoto et al., 1993;Romanini & Rizzo, 1995; Vindla, Sahota, Coppens, & James,1997) and in fetuses exposed to other deleterious antenatal condi-tions including growth restriction (Bekedam, Visser, de Vries, &Prechtl, 1985; I. Nijhuis et al., 2000; Vindla, James, & Sahota,1999), maternal diabetes (Kainer, Prechtl, Engele, & Einspieler,1997; Mulder, 1993), and substance exposure (Gingras &O’Donnell, 1998; Mulder, Morssink, van der Schee, & Visser,1998; Szeto, 1983).
Understanding of the developmental trajectories of normativefetal neurobehavior has emerged over time through synthesis offindings obtained with the more common cross-sectional approachand findings from longitudinal studies. The general pattern ofdecline in fetal heart rate (FHR) accompanied by an increase intime-dependent (G. S. Dawes, Moulden, Sheil, & Redman, 1992;Fleisher, DiPietro, Johnson, & Pincus, 1997) or time-independent(G. S. Dawes, Houghton, Redman, & Visser, 1982; van Leeuwen,Lange, Bettermann, Gronemeyer, & Hatzmann, 1999) variabilityis well known. Accelerations, defined as episodic excursions inFHR, form the cornerstone of antepartum clinical monitoring(Ware & Devoe, 1994). Reports of developmental trends in fetalmotor activity are less consistent. Some report that the fetusbecomes less active as term approaches (DiPietro, Hodgson, Costi-gan, Hilton, & Johnson, 1996b; Roodenburg, Wladimiroff, van Es,& Prechtl, 1991; ten Hof et al., 2002), but others fail to showchanges during the third trimester (Manning, Platt, & Sipos, 1979;Patrick, Campbell, Carmichael, & Probert, 1982). Differences inhow fetal movement (FM) is defined across studies make compar-isons difficult (ten Hof et al., 1999), and this inconsistency maypartially explain discrepancies. For example, fetuses may makefewer individual movements over time without a correspondingchange in the proportion of the time they spend moving (DiPietroet al., 1998; Roberts, Griffin, Mooney, Cooper, & Campbell,
1980). Therefore, inclusion of both types of measures provides amore comprehensive characterization of movement.
A third feature of fetal neurobehavioral development that hasreceived attention is the relationship between heart rate and motoractivity. As gestation advances, fetal motor activity becomes in-creasingly associated with transient accelerations of FHR. Thestrength of this relationship indicates general fetal well-being(Baser, Johnson, & Paine, 1992), and its developmental nature hasbeen interpreted as a sign of progressive integration betweensympathetic and parasympathetic innervation of the autonomicnervous system (DiPietro et al., 1996b; T. R. B. Johnson, Besinger,Thomas, Strobino, & Niebyl, 1992; Timor-Tritsch, Dierker, Zador,Hertz, & Rosen, 1978; Vintzileos, Campbell, & Nochinson, 1986).The linkage between somatic and cardiac functioning is also at thebasis of the emergence of fetal behavioral states as gestationprogresses (Groome & Watson, 1992; I. J. M. Nijhuis et al., 1999;J. G. Nijhuis, Prechtl, Martin, & Bots, 1982; van Vliet, Martin,Nijhuis, & Prechtl, 1985).
Consistent with the long-standing interest and well-developedliterature on sex differences in infants and children in general, themost frequently investigated moderator of development beforebirth has been fetal sex. Despite persistent clinical conviction,antenatal differences in mean heart rate between male and femalefetuses have not been documented (G. S. Dawes et al., 1982;DiPietro et al., 1996b; Druzin, Hutson, & Edersheim, 1986). Heartrate during labor was found to be significantly higher in femalefetuses in one study (N. Dawes, Dawes, Moulden, & Redman,1999) but not in another (Petrie & Segalowitz, 1980) that explicitlytested for sex differences. Similarly, variability in FHR was re-ported as greater in male fetuses in one study (DiPietro et al.,1998) but not in another (I. Nijhuis et al., 2000). The mostconsistently documented postnatal sex difference is that boys aremore physically active than girls. Male fetuses are reported tomake more frequent leg movements (Almli, Ball, & Wheeler,2001); female fetuses, more frequent mouthing movements (Hep-per, Shannon, & Dorman, 1998). However, a meta-analysis of sixstudies concluded that there are no antenatal sex differences inactivity level (Eaton & Enns, 1986). Conflicting results have beengenerated within our own work, with male fetuses being active formore of the observation time in one sample (DiPietro et al., 1996b)but not in another (DiPietro et al., 1998).
Two investigative groups, our own (DiPietro et al., 1996a,1996b) and a team in the Netherlands (I. Nijhuis et al., 1998; tenHof et al., 2002), have modeled normative fetal neurobehavioraldevelopment using statistical techniques that are becoming morecommonly applied in developmental science. Results generated bythese projects, which measured antenatal neurobehavior at eithermonthly or bimonthly intervals, have provided perhaps the mostcomprehensive information regarding development during the sec-ond half of gestation. However, both are limited in sample size(Ns � 31 and 29, respectively) and are based on relatively homo-geneous groups of women in the United States and England. Nostudy to date has either explicitly compared development beforebirth cross-culturally or comprehensively evaluated fetal neurobe-havioral development in a developing country.
We had two primary goals for this project. The first was toexamine the levels and trajectories of development in two differentcultures: one marked by the degree of affluence typical in the
446 DIPIETRO ET AL.

developed world; the other, by the pervasive disadvantages en-countered in developing countries. To this end, we implementedparallel longitudinal data collection at two sites: Baltimore, Mary-land, in the United States and Lima, Peru. By recruiting onlylow-risk women with normally progressing pregnancies who gavebirth to healthy offspring, we were able to focus on normativedevelopment in both locations. If antenatal development is drivenprimarily by intrinsic maturative processes, one would not expectfetal development to differ in these two locales. If, however, theenvironmental milieu of the pregnancy affects antenatal develop-ment, significant variation both in the level and trajectories of fetalneurobehavior would be expected between Baltimore and Lima.Our second goal was to confirm preliminary findings regarding thetrajectory of fetal neurobehavioral development from our earlier,small sample. In particular, we focused on those findings sugges-tive of a period of discontinuity at the beginning of the thirdtrimester, which has implications for the understanding of theneuromaturational processes of preterm infants delivered at thistime. In previous reports, discontinuities were detected in five ofeight aspects of fetal neurobehavioral development, clusteringaround the 28th to 32nd weeks of gestation (DiPietro et al., 1996a,1996b). In each instance, the slope of the trajectory of neurobe-havioral development slowed after this gestational period. Thus,we sought to replicate this finding with a larger U.S. sample and toestablish whether it is a universal phenomenon by applying thesame statistical analysis to a group of fetuses drawn from anentirely different population. A secondary goal, afforded by thelarger sample sizes, was to reconcile the discrepancies in theliterature regarding whether sex differences in fetal neurodevelop-ment exist.
Method
Participants
Eligibility for enrollment in Baltimore and Lima was restricted tononsmoking women with uncomplicated pregnancies carrying singletonfetuses. Accurate dating of the pregnancy, based on early first trimesterpregnancy testing or examination and/or confirmation by ultrasound, wasrequired. Gestational age was ultimately established by using the bestclinical estimate that was based on all available dating information(DiPietro & Allen, 1991). Fifty percent of each sample consisted ofprimiparous women.
A total of 185 self-referred pregnant women were enrolled in Baltimoreat Johns Hopkins Hospital. Women learned of the project through adver-tisements placed in local university publications and by word of mouth.Women resided either in Baltimore or the surrounding suburbs and did notnecessarily receive prenatal care or deliver their infants at the recruitinghospital. Forty-eight participants were either prospectively or retrospec-tively excluded for the following reasons: preterm labor, preterm delivery,or both (21; 11%); gestational diabetes (6; 3%); congenital malformation(2; 1%); fetal death in utero or nonviable delivery (2; 1%); growth retar-dation or other condition of antepartum origin detected in the newborn (6;3%); and lack of continued participation because of scheduling difficulties,moving, etc. (12; 6%).
Participants in Lima were part of a randomized controlled trial investi-gating the role of prenatal zinc supplementation in fetal growth anddevelopment. In order to approximate comparison with the populationnorms in Lima, the current analysis was based on women who made up thecontrol arm of the larger study and thus did not receive zinc supplemen-
tation. However, as is consistent with the recommendation of the PeruvianMinistry of Health, women in this group received daily supplementation ofiron and folic acid. There were no sociodemographic differences betweenwomen in the control and the experimental arms of the study. Recruitmentproceeded at the Hospital Materno Infantil San Jose in Villa El Salvador,an impoverished district at the periphery of the city. Women who areserved by this hospital are considered to be at low risk for poor pregnancyoutcomes, and those in the study enrolled in prenatal care prior to 16 weeksgestation. Lima, like Baltimore, is at sea level.
The same exclusionary criteria for developing pregnancy complicationsand infant conditions as in Baltimore were applied. Of the 117 women whobegan fetal testing, 16 were excluded for the following reasons: pretermdelivery (3; 2.6%); congenital malformation (3; 2.5%), fetal death in uteroor nonviable delivery (2; 5%); condition of antepartum origin detected inthe newborn (1; 1%); and lack of continued participation because ofscheduling difficulties, moving, etc. (7; 7%).
The final samples comprised 137 Baltimore and 101 Lima participants.Although a range of socioeconomic levels was represented in the U.S.sample, as a group they represented a well-nourished, middle- to upper-class population receiving a high standard of prenatal care. The Peruviansample represented a much less affluent population that was disadvantagedin terms of income, status, and all that these entail. However, the healthcare system provides a relatively high level of prenatal care, including earlydetection, monthly prenatal visits, screening, and referral services.
Design and Procedure
In both locations, fetal monitoring commenced at 20 weeks gestation andwas repeated at 24, 28, 32, 36, and 38 weeks gestation. In Baltimore,women were assessed at the same time of day during each visit (either 1:00p.m. or 3:00 p.m.), but testing conditions in Lima made this degree ofcontrol impossible. Testing in Lima took place between 9 a.m. and 6 p.m.However, no systematic diurnal effects for FHR or FM parameters havebeen found when testing is done within daytime hours (I. Nijhuis et al.,2000; ten Hof et al., 2002). All women were instructed to eat 11⁄2 hr priorto testing but not thereafter. A member of the Baltimore investigative teamsupervised implementation of the study in Lima to ensure fidelity in datacollection procedures. A brief real-time ultrasound scan was conducted todetermine fetal position and provide photographs to parents. Fetal moni-toring proceeded for 50 min, with the mother resting comfortably in asemirecumbent, left-lateral position. This duration was consistent with thatchosen in our prior studies and exceeds the 30 to 40 min of recording timeestablished for intrafetal stability (I. Nijhuis et al., 1998; Ribbert, Fidler, &Visser, 1991). The standards and methods of data collection and analysis atboth sites were identical with one exception: Data collection at 32 weeksin Baltimore was shortened to 30 min to facilitate an experimental manip-ulation following the undisturbed recording.
Fetal data were collected using a Toitu MT320 fetal actocardiograph(Toitu Co., Ltd., Tokyo, Japan). This monitor detects FM and FHR throughthe use of a single, wide array transabdominal Doppler transducer andprocesses this signal through a series of filters. The actograph detects FMby preserving the remaining signal after bandpassing frequency compo-nents of the Doppler signal that are associated with FHR and maternalsomatic activity. Reliability studies comparing actograph-based versusultrasound-visualized FM have found the performance of this monitor to behighly accurate in detecting both fetal motor activity and quiescence(Besinger & Johnson, 1989; DiPietro, Costigan, & Pressman, 1999; Maeda,Tatsumura, & Utsu, 1999).
Fetal data in both sites were collected from the output port of the monitorand digitized at 1000 Hz through an internal A/D (analog/digital) boardusing streaming software. Data were analyzed offline with software devel-oped for this project. Digitized FHR data underwent error rejection pro-cedures based on moving averages of acceptable values as needed. Fetal
447FETAL NEUROBEHAVIORAL DEVELOPMENT
variables included three cardiac measures: FHR, variability (the standarddeviation of each 1-min epoch aggregated over time), and accelerations.Based on standard clinical definitions, accelerations were defined as oc-curring when FHR values attained 10 beats per minute (bpm) abovebaseline for 15 s or longer. FM measures were based on the actographsignal, which ranges from 0 to 100 in arbitrary units. A movement bout wasconsidered to begin when the first spike of the actograph attained ampli-tude of 15 units and to end when there was a cessation of 15 unit signalsfor at least 10 s. The number of movements was counted, and the durationof each movement was measured. Total motor activity was computed as thenumber of movement bouts multiplied by the mean movement duration (inseconds) divided by 3,000 (the number of seconds per 50 min of record-ing). This variable represents the proportion of time the fetus spent movingduring the observation period. FHR–FM coupling was calculated as theproportion of fetal movements associated with excursions in FHR � 5 bpmover baseline within 5 s before the start of a movement or within 15 s afterthe start of a movement, consistent with previously developed criteria(Baser et al., 1992; DiPietro et al., 1996a).
Data Analysis
All fetal and maternal measures were examined for normality; two FMoutliers were found, one in Baltimore and one in Lima. Because twovariables (accelerations and movement bouts) at 32 weeks required weight-ing relative to the shortened recording length, care was taken to detectinstances in which the 32-week data did not conform to expectations basedon patterns of earlier and later data. Weighted least squares analysis wasused to model the developmental trends of the FHR, FM, and FHR–FMcoupling measures over time separately for each sample. This methodestimates the correlation structure generated by the repeated measurementson the same fetus and uses the estimate to weight the observations in theregression analysis. The robustness of the estimated unstructured correla-tion matrix was assessed using generalized estimating equations method-ology (GEE; Zeger & Liang, 1986). This technique produces appropriateestimates of regression parameters and their variances (Diggle, Liang, &Zeger, 1994). Moreover, unlike repeated measures analysis of varianceprocedures, GEE does not exclude subjects with missing data from the finalmodel. There were few instances of missing data that were due to eithernoncompliance or technical problems at any visit prior to the final one (2cases at 28 weeks in Baltimore; 1 to 2 cases at each gestational age inLima). However, a substantial number of women who delivered at term(i.e., 37 to 41 weeks) delivered before their scheduled 38-week visit (33%in Baltimore; 9% in Lima), usually because they delivered earlier duringthat week.
To model nonlinear trends, we included knotted splines in each model at24, 28, 32, and 36 weeks gestation, and thus there were at least twocontiguous data points before a deviation was established. Knotted splinesallow nonlinear trends to be modeled by permitting the slope to change atthe “knot”; consideration of significance reflects the degree to which theslopes prior to and following this point diverge. In the event that changesin slope were significant at more than one point, the placement of the splineknot was determined by the magnitude of the deviance from the saturatedmodel. Terms for fetal sex were added to each linear model. Comparisonsbetween the general developmental trends in Baltimore and Lima wereconducted by modeling slopes by location.
Results
Maternal and Infant Characteristics
Table 1 presents maternal and infant characteristics. There wereclear differences in maternal sociodemographic characteristics be-tween the Baltimore and Lima samples. The level of maternal
education diverged greatly, with some degree of graduate training(43%) being the modal value in the U.S. sample compared with ahigh school education (49.5%) in Peru. Almost a third of the Limasample did not have a high school diploma. Explicit comparisonsof income and occupation were not possible because of differencesin monetary value and the low levels of maternal employmentduring childbearing in Lima. Peruvian women were of Mestizodescent. The U.S. sample was 85% White, 12% African American,and 3% Hispanic or Asian. The Baltimore women were about 8years older than the Lima women.
Anthropometric measures can provide indicators of nutritionalstatus in mothers and fetal growth in infants. Peruvian womenwere shorter and lighter, but the difference in body mass index(BMI) between groups was only marginally significant. Similarly,although Peruvian infants were smaller at birth, the ponderalindex, which reflects the relationship between weight and length,was not lower, nor was head circumference. Despite the Peruvianinfants’ lower birth weight, the average gestational age at deliverywas 3 days later in Lima than in Baltimore.
Developmental Trends
Table 2 presents the results of the linear and spline-based GEEanalyses for the Baltimore and Lima samples over gestation.Results include the coefficients (estimates), the standard errors,and the Z scores for significance testing. Each estimate indicatesthe magnitude of weekly change for each variable in its unit ofmeasurement. For example, from 20 weeks to 38 weeks, FHRdeclined at a rate of 0.41 bpm per week. Mean raw data values areplotted in Figures 1 through 7. Highly significant positive linear
Table 1Maternal and Infant Characteristics
CharacteristicBaltimore(n � 137)
Lima(n � 101) t or �2 df
Maternal
Age (years) 31.3 23.4 13.29* 236Education (%) 167.10* 3
Less than high school 0 30.7High school 5.1 49.51–3 years postsecondary 16.0 19.8Baccalaureate degree 35.8 0Graduate training 43.1 0
Primiparous (%) 55.7 53.5 0.04 1Prepregnancy weight (kg) 68.2 54.8 8.62* 236Height (cm) 166.2 152.3 17.66* 236Body mass index (kg/m2) 24.7 23.6 1.91† 236
Infant
Male (%) 50 50 0.01 1Gestational age at birth
(weeks) 39.4 40.0 �3.61* 236Birth weight (g) 3,520 3,214 5.55* 228Birth length (cm) 51.7 49.8 6.76* 227Head circumference (cm) 34.7 34.7 0.01 195Ponderal index (g/cm3) 2.56 2.60 �1.34 227
† p � .10. * p � .01.
448 DIPIETRO ET AL.
trends over gestation were detected for all three FHR measures(mean, variability, and accelerations) at both sites, with decliningmean FHR but increasing variability and accelerations. Couplingbetween FHR and FM increased significantly over time at bothsites. The number of fetal movements declined over time in bothsites, but the total amount of activity and mean FM durationdeclined significantly in Peru only.
Values listed in Table 2 under “Discontinuity (spline)” indicatethe gestational period (if any) at which a significant change inslope was detected. Discontinuities in development were evidentfor FHR variability, accelerations, and coupling with FM in bothBaltimore and Lima, clustering between 28 and 32 weeks gesta-tion. FHR showed a change in trajectory in Lima only, at 28weeks. Examination of the data plotted in the figures reveals thatthe slope of the trajectory following each spline is reduced relativeto the trajectory to that point. Inspection of Figure 1 indicates anapparent change in slope for FHR in Baltimore at 32 weeks.Examination of the data revealed substantial interfetal variabilityin the 36-week FHR, which thus suppressed detection of a changein slope (Z � �1.48, p � .14). Exclusion of a single case ofunusually high FHR (170 bpm) resulted in a marginally significantdiscontinuity at 32 weeks (estimate � �.223, SE � .125, Z ��1.78, p � .07). There were two significant spline terms withsimilar deviance values for FHR variability (at 24 and 28 weeks)in the Baltimore sample. After visual inspection of the plots, the28-week point was selected as the more conservative estimatebecause it included three, instead of two, data points in the initialtrajectory.
Trajectories for FM were less consistent. No significant splines
were detected in Baltimore for the number of movements, whichwas the only FM measure to display a gestational trend. In Peru,the number of movements and total motor activity declined mostprecipitously after 28 and 36 weeks, respectively.
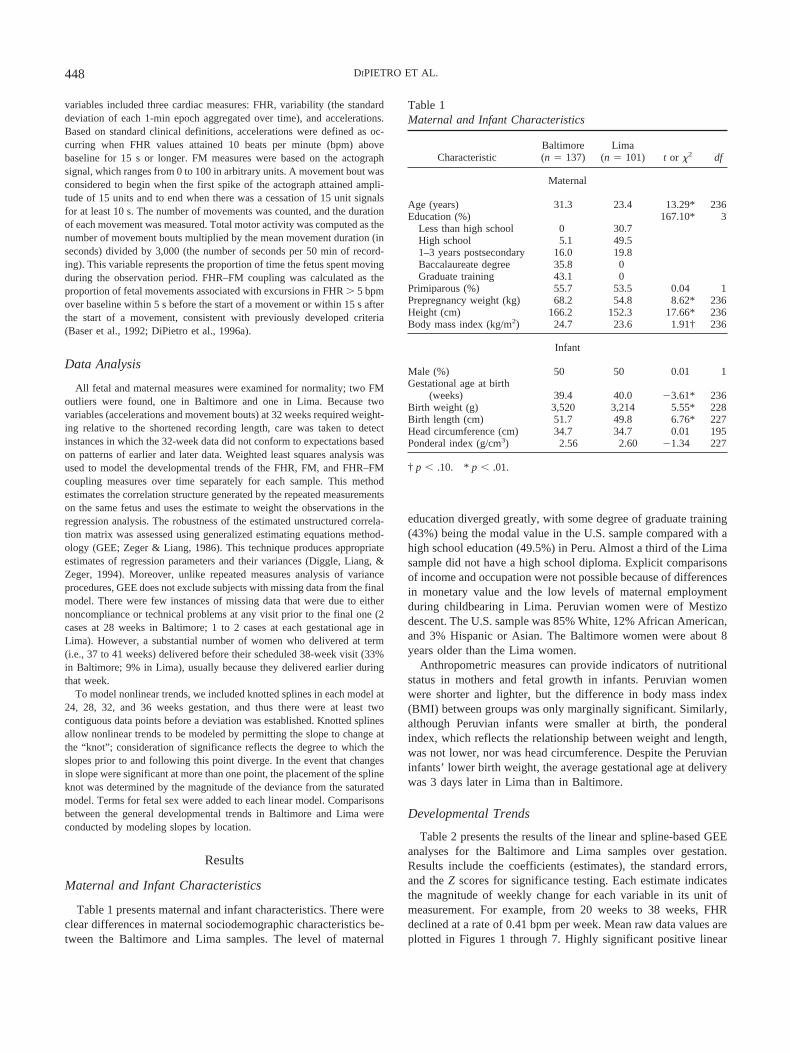
Figure 1. There was an initial decline in fetal heart rate in both localesfollowed by a significant leveling of slope in Lima at 28 weeks and a trendtoward a decline at 28 weeks in Baltimore (see Table 2). Mean fetal heartrate during gestation was significantly higher in Lima than in Baltimore(i.e., Z � 3.23, *p � .01). The coefficient (�) indicates that magnitude; onaverage, there was a difference of 2.17 beats per minute (bpm), althoughthis was not constant over gestation.
Table 2Fetal Neurobehavioral Development: Results of Generalized Estimating Equations AnalysisShowing Trends in Development Over Gestation
Fetal measure
Gestation Discontinuity (spline)
Estimate SE Z GA Estimate SE Z
Baltimore
Heart rate �.410 .031 �13.00* Nonea
Heart rate variability .203 .011 18.48* 28a .116 .031 3.79*Accelerations .357 .021 17.02* 32 .292 .063 4.67*FM–FHR coupling .010 .001 16.82* 32 .010 .002 4.64*Number of movements �.625 .083 �7.50* NoneMovement duration �.413 .361 �1.14 NoneTotal motor activity �.002 .001 �1.59 None
Lima
Heart rate �.298 .040 �7.41* 28 �.417 .109 �3.84*Heart rate variability .113 .011 10.14* 24a .389 .060 6.49*Accelerations .173 .015 11.14* 32 .218 .052 4.15*FM–FHR coupling .009 .001 13.38* 28 .008 .002 4.13*Number of movements �.574 .092 �6.26* 28 1.373 .296 4.62*Movement duration �.457 .130 �3.51* NoneTotal motor activity �.011 .001 �7.69* 36 .032 .012 2.56*
Note. GA � gestational age at which a significant change in slope was detected; FM � fetal movement;FHR � fetal heart rate.a See text for additional description of discontinuity.* p � .01.
449FETAL NEUROBEHAVIORAL DEVELOPMENT
Baltimore–Lima Comparisons
Results of the comparisons in overall levels of fetal neurobe-haviors between Baltimore and Lima are provided in the uppercorners of Figures 1 through 7. The coefficients reflect the adjustedmean difference over gestation; the Z scores are used to determinesignificance levels of the comparison between groups across ges-tation. There were large, significant mean differences in the threeFHR measures and FM–FHR coupling. Fetuses in Baltimore hadslower FHR, greater FHR variability, more accelerations, andhigher levels of FM–FHR coupling. Information concerning thenature of these differences as they relate to developmental trajec-tories is provided by comparisons of the onsets of the discontinui-ties. For three of these four measures (FHR, FHR variability, andFM–FHR coupling), the change in slope signifying the onset of thedeceleration in development occurred 1 month earlier in Peru thanin the United States. Moreover, after the initial decline in the rateof increase in FHR variability, FHR variability plateaued with aslight decline with advancing gestation (see Figure 2). This processbegan at 32 weeks in Lima (estimate � .247, SE � .042, Z � 5.87,p � .01) but at 36 weeks in Baltimore (estimate � .301, SE �.095, Z � 3.19, p � .01). With respect to accelerations, a discon-tinuity was detected in both groups at 32 weeks, but inspection ofFigure 3 reveals a very different pattern of development beyondthis point. In Baltimore, accelerations continued to increase but ata slower rate; fetuses in Lima showed little continued developmenton this measure and then declined at 36 weeks (Z � 3.97, p � .01).Trajectories for FM–FHR coupling were similar between groups(see Figure 4), but levels of coupling were consistently lower inLima than in Baltimore.
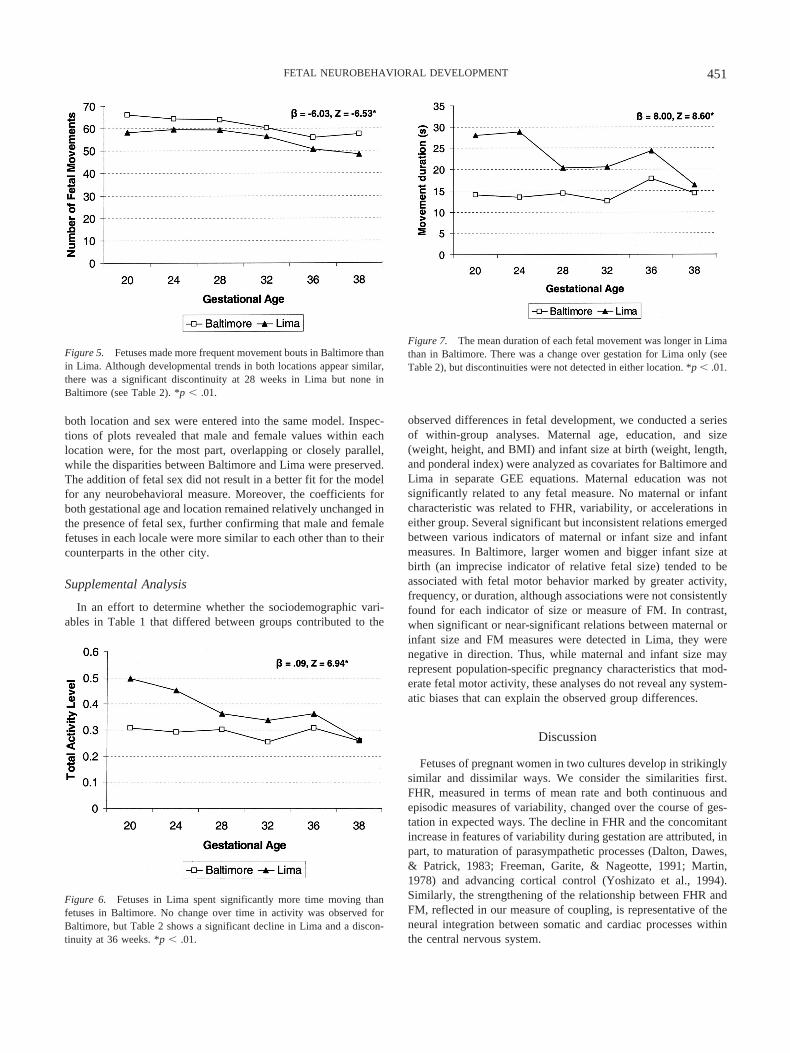
The plotted values and trajectories for the number of fetalmovements (see Figure 5) are remarkably similar between groups,although the Baltimore sample made significantly more move-ments. Across gestation, Baltimore fetuses made between 3.6 and9.1 more movements per 50-min recording period than did fetusesin Lima. In contrast, fetuses in Lima spent significantly more of
the observation period in motor activity (see Figure 6) driven bythe longer mean duration of each movement (see Figure 7). Thisdifference was more pronounced earlier in gestation than as termneared. Discontinuities had relatively little impact on differencesbetween fetal motor activity between study locations.
Fetal Sex
No sex differences in fetal neurobehaviors in either the Balti-more or Lima samples were detected, and there were no interac-tions between fetal sex and gestational age. To determine whetherpotential fetal sex effects might be masked by within-locationanalyses, we conducted an additional series of analyses in which
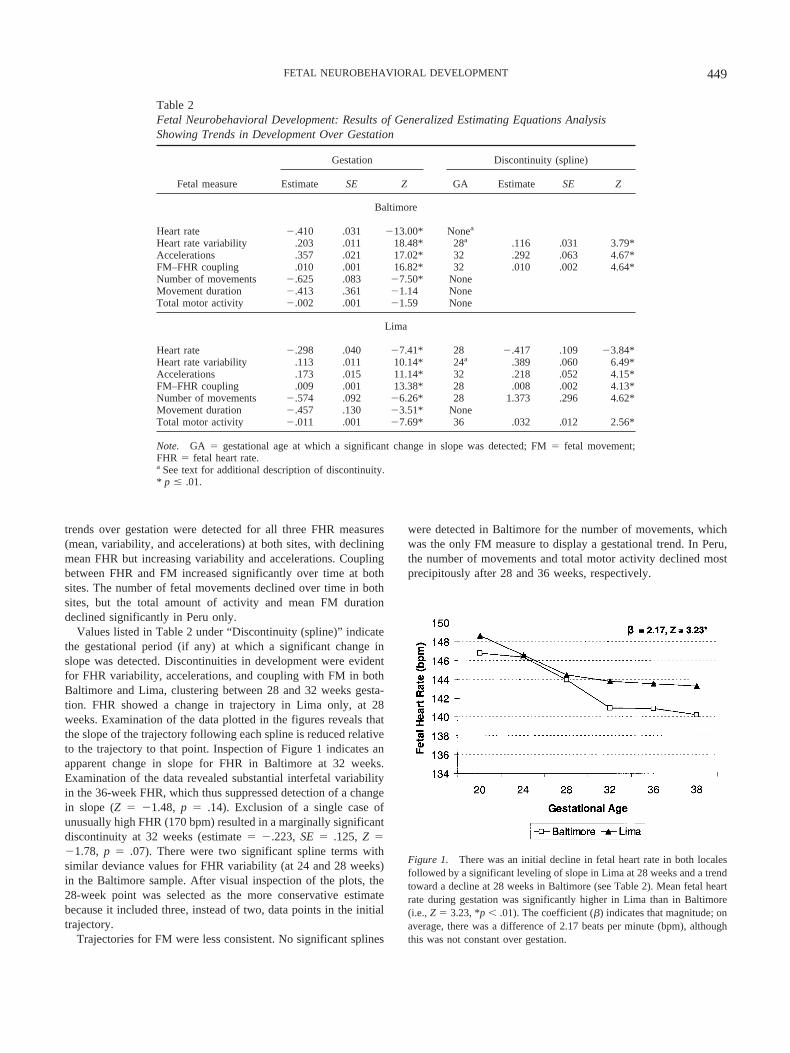
Figure 3. Episodic accelerations in fetal heart rate per 50-min recordingwere more frequent in Baltimore than in Lima. Significant discontinuityexisted at 32 weeks in both locations (see Table 2) followed by a continu-ing increase in incidence in Baltimore but a decline in Lima. *p � .01.
Figure 4. Fetal movement (FM) – fetal heart rate (FHR) coupling washigher in Baltimore than in Lima. From Table 2, note the increase overgestation with deceleration at 32 weeks in Baltimore and at 28 weeks inLima. *p � .01.
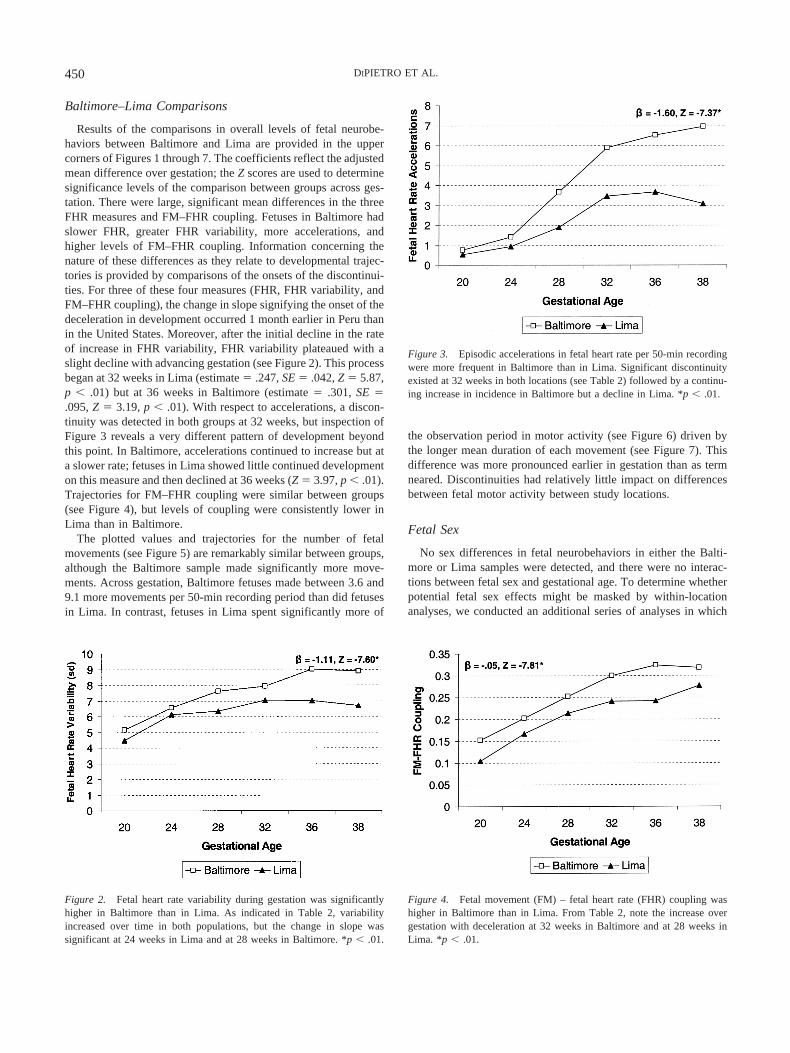
Figure 2. Fetal heart rate variability during gestation was significantlyhigher in Baltimore than in Lima. As indicated in Table 2, variabilityincreased over time in both populations, but the change in slope wassignificant at 24 weeks in Lima and at 28 weeks in Baltimore. *p � .01.
450 DIPIETRO ET AL.
both location and sex were entered into the same model. Inspec-tions of plots revealed that male and female values within eachlocation were, for the most part, overlapping or closely parallel,while the disparities between Baltimore and Lima were preserved.The addition of fetal sex did not result in a better fit for the modelfor any neurobehavioral measure. Moreover, the coefficients forboth gestational age and location remained relatively unchanged inthe presence of fetal sex, further confirming that male and femalefetuses in each locale were more similar to each other than to theircounterparts in the other city.
Supplemental Analysis
In an effort to determine whether the sociodemographic vari-ables in Table 1 that differed between groups contributed to the
observed differences in fetal development, we conducted a seriesof within-group analyses. Maternal age, education, and size(weight, height, and BMI) and infant size at birth (weight, length,and ponderal index) were analyzed as covariates for Baltimore andLima in separate GEE equations. Maternal education was notsignificantly related to any fetal measure. No maternal or infantcharacteristic was related to FHR, variability, or accelerations ineither group. Several significant but inconsistent relations emergedbetween various indicators of maternal or infant size and infantmeasures. In Baltimore, larger women and bigger infant size atbirth (an imprecise indicator of relative fetal size) tended to beassociated with fetal motor behavior marked by greater activity,frequency, or duration, although associations were not consistentlyfound for each indicator of size or measure of FM. In contrast,when significant or near-significant relations between maternal orinfant size and FM measures were detected in Lima, they werenegative in direction. Thus, while maternal and infant size mayrepresent population-specific pregnancy characteristics that mod-erate fetal motor activity, these analyses do not reveal any system-atic biases that can explain the observed group differences.
Discussion
Fetuses of pregnant women in two cultures develop in strikinglysimilar and dissimilar ways. We consider the similarities first.FHR, measured in terms of mean rate and both continuous andepisodic measures of variability, changed over the course of ges-tation in expected ways. The decline in FHR and the concomitantincrease in features of variability during gestation are attributed, inpart, to maturation of parasympathetic processes (Dalton, Dawes,& Patrick, 1983; Freeman, Garite, & Nageotte, 1991; Martin,1978) and advancing cortical control (Yoshizato et al., 1994).Similarly, the strengthening of the relationship between FHR andFM, reflected in our measure of coupling, is representative of theneural integration between somatic and cardiac processes withinthe central nervous system.
Figure 5. Fetuses made more frequent movement bouts in Baltimore thanin Lima. Although developmental trends in both locations appear similar,there was a significant discontinuity at 28 weeks in Lima but none inBaltimore (see Table 2). *p � .01.
Figure 6. Fetuses in Lima spent significantly more time moving thanfetuses in Baltimore. No change over time in activity was observed forBaltimore, but Table 2 shows a significant decline in Lima and a discon-tinuity at 36 weeks. *p � .01.
Figure 7. The mean duration of each fetal movement was longer in Limathan in Baltimore. There was a change over gestation for Lima only (seeTable 2), but discontinuities were not detected in either location. *p � .01.
451FETAL NEUROBEHAVIORAL DEVELOPMENT

With respect to FM, given the potential range in the number ofmovements a fetus could make in 50 min, the mean values andtrajectories for this measure were surprisingly consistent betweengroups. The average number of fetal movements across gestationand study site ranged from 48 to 66, or between 0.96 and 1.32 perminute. Observations that the fetus moves, on average, once perminute have been reported from studies that rely on variousmethods of identifying FM (DiPietro et al., 1998; Manning et al.,1979; Nasello-Paterson, Natale, & Connors, 1988; Roberts et al.,1980; Roodenburg et al., 1991). There was convergence in fetalactivity level at term such that both groups of fetuses spentapproximately 25% of the observation time moving. The trajecto-ries of development for mean movement duration were also quitesimilar, again showing convergence at term. Investigations ofperiodicity in motility in animal and human fetuses provide strongevidence for endogenously generated cyclic motor activity (Rob-ertson, 1985), and the similarity observed between samples sup-ports this phenomenon.
Results from the current study confirm earlier observations thata developmental transition occurs between the 28th and 32ndgestational weeks (DiPietro et al., 1996a, 1996b). Fetuses in bothLima and Baltimore exhibited developmental discontinuities in allmeasures that involved FHR, including its relation to FM, withinthis period. Fetal motor measures were much less consistent in thisregard, but when discontinuities were detected, they clusteredwithin this period. Others have also reported transitions in fetalfunctions during this period. The inspiratory component of fetalbreathing movements peaks between 28 and 32 weeks (Kozuma,Nemoto, Okai, & Mizuno, 1991), and fetal breathing rates plateauduring this time (Pillai & James, 1990; Roodenburg et al., 1991).The fetus attains mature levels and patterns of responsiveness tovibroacoustic stimuli between 29 and 32 weeks (Kisilevsky, Muir,& Low, 1992; Kuhlman, Burns, Depp, & Sabbagha, 1988), andthere is a concomitant increase in habituation (Groome, Gotlieb,Neely, & Waters, 1993). The onset of these neurobehavioralchanges coincides with a period of rapid increase in neural devel-opment and myelination, including cortical and vagal processes(Kinney, Karthigason, Borenshteyn, Flax, & Kirschner, 1994;Sachis, Armstrong, Becker, & Bryan, 1982). In a study of thedevelopment of neural control of the heart, measures of FHRvariability were compared between anencephalic and normal fe-tuses at varying gestational ages (Yoshizato et al., 1994). A criticaltransition period during which higher cortical control was exertedwas documented between 27 and 30 weeks gestation.
On the basis of the existing literature and the current findings,we propose that the deceleration in neurobehavioral maturationafter 32 weeks suggests that antenatal neural development throughterm is somewhat overdetermined. Ancillary support for this po-sition is provided by the ultimate developmental and cognitivesuccess of preterm infants who are born after this gestationalperiod, despite immaturity in other organ systems. This is not toimply that development ceases after this period or that certainaspects of fetal neurobehavior do not continue to develop in alinear fashion. For example, there is a linear increase in periods ofwakefulness as fetuses progress through postterm pregnancies(Junge, 1979; van de Pas, Nijhuis, & Jongsma, 1994). At the veryleast, the consistently decelerative trajectories observed in these
two samples of fetuses indicate that the rate of development beginsto taper off well before term.
Despite the overall similarities in trends between the Baltimoreand Lima samples, the disparities are equally revealing. Theseresults make clear that there is population-based variation both inthe level and the trajectories of fetal neurobehavior. The Limasample failed to attain the same level of decline in FHR and wasconsistently lower in both FHR variability and FM–FHR couplingthroughout gestation. Moreover, after 24 weeks gestation, thisgroup displayed approximately half the number of accelerations inFHR, the primary clinical indicator of fetal well-being. Differencesin level were exacerbated by variation in developmental trajecto-ries. For three of these measures, the deceleration in developmentthat is denoted by the significant spline term occurred at least 4weeks before the corresponding change was evident in the Balti-more sample. For FHR and FM–FHR coupling, this was expressedas lack of further significant change beginning as early as 28weeks gestation, compared with 32 weeks in Baltimore. Withrespect to FHR variability, not only did the decline in slope begin1 month earlier in Peru (at 24 vs. 28 weeks), but the decline inlevel expressed in both samples as gestation progressed was initi-ated a month earlier in Peru (at 32 vs. 36 weeks). The most strikingcontrast in trajectory was displayed for accelerations. The discon-tinuity observed in both samples at 32 weeks preceded a continu-ing increase at a reduced rate in Baltimore but a collapse ofsubsequent development in Peru. Thus, on the measures mostclearly affiliated with neural development, the Peruvian fetuseslagged significantly behind their Baltimore counterparts.
What is the source of these disparities? Women at each sitereflected widely different populations of affluence and disadvan-tage, yet all had good prenatal care, uneventful pregnancies, andhealthy newborns. Recruitment techniques were somewhat differ-ent for the two populations, but participation was ultimately de-pendent on volunteerism in both groups. Post hoc analyses failedto show that the differences in measured sociodemographic char-acteristics of the sample contributed to FHR measures within eachgroup, making it difficult to conclude that they generated between-groups differences. Although fetal motor activity showed somerelations with maternal and infant size, the directions of theseassociations varied within each group and did not parallel thebetween-groups differences. More surprisingly, maternal educa-tion, an important proxy that confers a wide sphere of influence ona woman’s life, failed to differentiate fetal neurobehavior withineither sample.
The potentially important genetic influence of ethnicity cannotbe ascertained by this study, nor could a targeted study be easilymounted because of corresponding social class differences withinPeru. However, we have been unable to identify published reportsthat show an effect of maternal ethnicity per se on any aspect offetal development. Racial differences in fetal heart rate patternshave been observed (Ogueh & Steer, 1998) but are typicallyconfounded with socioeconomic status (M. Johnson et al., 1992).Similarly, an effort to match on sociodemographic characteristicsbetween locales would have created a new set of potential con-founds. Finally, numerous unmeasured characteristics of thematernal–fetal intrauterine milieu exist that may have contributedto these results, including hemoglobin levels, uteroplacental suf-ficiency, by-products of maternal psychosocial stress, and nutri-
452 DIPIETRO ET AL.

tional status. There is reason to suspect that the two groups differedon the latter: Although the lack of group differences in BMI forwomen and in the ponderal index for infants indicates that thePeruvian sample was not malnourished and the fetuses were notgrowth restricted, women in Lima tend to be deficient in micro-nutrients (Sacco, Caulfield, Zavaleta, & Retamozo, 2003). Thesefindings also do not rule out the possibility that disparities of asimilar nature or magnitude might have been detected had wecompared different Peruvian or U.S. samples with each other.Results based on a low-income group in Baltimore are consistentin direction, but not magnitude, with some of the Baltimore–Limadisparities detected here (DiPietro et al., 1998).
We were unsuccessful in documenting sex differences in anyaspect of fetal neurobehavioral development despite previous re-ports to the contrary by others and ourselves. Our conclusion isthat male and female fetuses develop similarly in terms of undis-turbed, baseline measures of function and that earlier positivefindings reflect Type I errors as a result of the vagaries of smallsamples. However, these conclusions are tempered by the quanti-tative nature of the FM data collected through the actograph, andthere may be qualitative differences in motor behavior between thesexes that require ultrasound visualization to detect.
Several other issues relevant to fetal research in general areraised by these results. In this study as in others, conclusions aboutdisparities in motor development are dependent on the manner inwhich motor activity is defined. Data on FM during gestationprovided a much less consistent portrait both within and acrossgroups. For example, Baltimore fetuses exhibited a greater numberof movements, but Lima fetuses spent more of the observationperiod moving. This confirms the importance of definitional cri-teria for FM in comparing across studies. These data also under-score the need for repeated measures designs in studies of thefetus. Our conclusions about potential group differences in activitylevel, for example, would have been quite different had data beencollected only at 38 weeks. Similarly, a single assessment at 24weeks would have obscured the progressive divergence in accel-erations as gestation advanced. Just as a single week in the first 2years of life is not representative of all of infancy, neither does asingle gestational age reflect the fetal period.
Among developmentalists, there is long-standing acceptance ofthe notion that continuity exists among processes that span theprenatal and postnatal periods (Als, 1982; Prechtl, 1984). Morerecently, Barker (1995) and colleagues have reinvigorated interestin this construct with their proposal that fetal programming pro-vides a substrate for subsequent health and well-being in a varietyof domains throughout life. An adverse prenatal environment hasbeen linked with hypertension and cardiovascular disease (Barker,2002; Szitanyi, Janda, & Poledne, 2003), insulin resistance anddiabetes (Hales & Ozanne, 2003; Phillips & Barker, 1997), and anumber of other undesirable health outcomes, although detectionof these associations has not been universal across studies (Laurenet al., 2003). The failure of this construct to similarly ignite thedevelopmental community may be a result of the existing accep-tance of the importance of the fetal period but also may be due tothe fact that the original literature focused almost exclusively onweight at birth as an indicator of the prenatal environment. Morerecent discourse in this arena has acknowledged that birth weightis an imperfect proxy for mechanisms through which antenatal
factors challenge growth and development, and attention has beenreoriented to the intrauterine processes that may link fetal growthto outcomes. These processes, which include aberrations infetal nutrient or oxygen delivery (Godfrey, 2002; Harding, 2001)and metabolic or endocrinological consequences, such ashypothalamic–pituitary–adrenal (Betram & Hanson, 2002; Mat-thews, 2002) and sympathetic–adrenal (Young, 2002) activation,likely have direct application to fetal development as well asgrowth and have been implicated in an array of postnatal devel-opmental processes.
The results from the current study indicate that fetal neurobe-havioral development represents a confluence of epigenetic, envi-ronmental, and maturative processes. The processes brought tobear on shaping fetal neurobehavioral development within groupsand individuals are probably not unlike those that influence devel-opment after birth but are mediated through the direct biologicaland physiological constraints imposed by the maternal uterineenvironment. Although there are many consistencies in the natureand trajectories of fetal neurobehavioral development in the twosamples studied, there were unattributable but unquestionablecross-cultural discrepancies. These results suggest that differentdevelopmental trajectories expressed in the fetal period underlieexisting observations of cross-cultural differences on neonatalneurobehavioral assessments (Choi & Hamilton, 1986; Coll et al.,1981; Eishima, 1992; Keefer et al., 1982). The task of identifyingpotential mechanisms while adequately controlling for other rele-vant factors will be a complex one but one that serves as animpetus for future research. Efforts to determine whether fetalneurobehaviors will predict either different developmental trajec-tories or variations in temperament between cultures in the post-natal period are under way in both cities. Despite the currentambiguity in understanding the mechanisms through which theintrauterine milieu engages the developing fetal nervous system,these findings indicate that the broader context of the antenatalenvironment must be considered in the putative role of the fetus asthe precursor to the child.
References
Almli, C. R., Ball, R. H., & Wheeler, M. E. (2001). Human fetal andneonatal movement patterns: Gender differences and fetal-to-neonatalcontinuity. Developmental Psychobiology, 38, 252–273.
Als, H. (1982). Toward a synactive theory of development: Promise for theassessment and support of infant individuality. Infant Mental HealthJournal, 3, 229–243.
Barker, D. J. (1995). The fetal and infant origins of disease. EuropeanJournal of Clinical Investigation, 25, 457–463.
Barker, D. (2002). Fetal programming of coronary heart disease. Trends inEndocrinology and Metabolism, 13, 364–368.
Baser, I., Johnson, T. R. B., & Paine, L. L. (1992). Coupling of fetalmovement and fetal heart rate accelerations as an indicator of fetalhealth. Obstetrics and Gynecology, 80, 62–66.
Bekedam, D., Visser, G., de Vries, J., & Prechtl, H. (1985). Motor behav-iour in the growth retarded fetus. Early Human Development, 12, 155–165.
Besinger, R. E., & Johnson, T. R. B. (1989). Doppler recordings of fetalmovement: Clinical correlation with real-time ultrasound. Obstetricsand Gynecology, 74, 277–280.
Betram, C., & Hanson, M. (2002). Prenatal programming of postnatalendocrine responses by glucocorticoids. Reproduction, 124, 459–467.
453FETAL NEUROBEHAVIORAL DEVELOPMENT

Choi, E., & Hamilton, R. (1986). The effects of culture on mother–infantinteraction. Journal of Obstetric, Gynecologic, and Neonatal Nursing,15, 256–261.
Coll, C., Spekoski, C., & Lester, B. (1981). Cultural and biomedicalcorrelates of neonatal behavior. Developmental Psychobiology, 14, 147–154.
Dalton, K., Dawes, G. S., & Patrick, J. E. (1983). The autonomic nervoussystem and fetal heart rate variability. American Journal of Obstetricsand Gynecology, 146, 456–462.
Dawes, G. S., Houghton, C. R. S., Redman, C. W. G., & Visser, G. H. A.(1982). Pattern of the normal human fetal heart rate. British Journal ofObstetrics and Gynecology, 89, 276–284.
Dawes, G. S., Moulden, M., Sheil, O., & Redman, C. W. G. (1992).Approximate entropy, a statistic of regularity, applied to fetal heart ratedata before and during labor. Obstetrics and Gynecology, 80, 763–768.
Dawes, N., Dawes, G., Moulden, M., & Redman, C. (1999). Fetal heart ratepatterns in labor vary with sex, gestational age, epidural analgesia, andfetal weight. American Journal of Obstetrics and Gynecology, 180,181–187.
Devlin, B., Daniels, M., & Roeder, K. (1997). The heritability of IQ.Nature, 388, 468–471.
Diggle, P., Liang, K., & Zeger, S. (1994). Analysis of longitudinal data.New York: Oxford University Press.
DiPietro, J. A., & Allen, M. C. (1991). Estimation of gestational age:Implications for developmental research. Child Development, 62, 1184–1199.
DiPietro, J. A., Costigan, K. A., & Pressman, E. K. (1999). Fetal movementdetection: Comparison of the Toitu actograph with ultrasound from 20weeks gestation. Journal of Maternal-Fetal Medicine, 8, 237–242.
DiPietro, J. A., Costigan, K. A., Shupe, A. K., Pressman, E. K., & Johnson,T. R. B. (1998). Fetal neurobehavioral development: Associations withsocioeconomic class and fetal sex. Developmental Psychobiology, 33,79–91.
DiPietro, J. A., Hodgson, D. M., Costigan, K. A., Hilton, S. C., & Johnson,T. R. B. (1996a). Development of fetal movement – fetal heart ratecoupling from 20 weeks through term. Early Human Development, 44,139–151.
DiPietro, J. A., Hodgson, D. M., Costigan, K. A., Hilton, S. C., & Johnson,T. R. B. (1996b). Fetal neurobehavioral development. Child Develop-ment, 67, 2553–2567.
DiPietro, J. A., Irizarry, R., Hawkins, M., Costigan, K., & Pressman, E.(2001). Cross-correlation of fetal cardiac and somatic activity as anindicator of antenatal neural development. American Journal of Obstet-rics and Gynecology, 185, 1421–1428.
Druzin, M., Hutson, J., & Edersheim, T. (1986). Relationship of baselinefetal heart rate to gestational age and fetal sex. American Journal ofObstetrics and Gynecology, 154, 1102–1103.
Eaton, W., & Enns, L. (1986). Sex differences in human motor activitylevel. Psychological Bulletin, 100, 19–28.
Eishima, K. (1992). A study on neonatal behavior comparing between twogroups from different cultural backgrounds. Early Human Development,28, 265–277.
Fleisher, L. A., DiPietro, J. A., Johnson, T. R. B., & Pincus, S. (1997).Complementary and non-coincident increases in heart rate variabilityand irregularity during fetal development. Clinical Science, 92, 345–349.
Freedman, D., & DeBoer, M. (1979). Biological and cultural differences inearly child development. Annual Review of Anthropology, 8, 579–600.
Freeman, R. K., Garite, T. J., & Nageotte, M. P. (1991). Physiologic basisof fetal monitoring. In R. K. Freeman, T. J. Garite, & M. P. Nageotte(Eds.), Fetal heart rate monitoring (2nd ed., pp. 7–20). Baltimore, MD:Williams & Wilkins.
Gillman, M., & Rich-Edwards, J. (2000). The fetal origins of adult disease:
From sceptic to convert. Paediatric and Perinatal Epidemiology, 14,192–193.
Gingras, J. L., & O’Donnell, K. J. (1998). State control in the substance-exposed fetus: I. The fetal neurobehavioral profile: An assessment offetal state, arousal, and regulation competency. Annals of the New YorkAcademy of Sciences, 846, 262–276.
Godfrey, K. (2002). The role of the placenta in fetal programming: Areview. Placenta, 16, S20-S27.
Groome, L. J., Gotlieb, S. J., Neely, C. L., & Waters, M. D. (1993).Developmental trends in fetal habituation to vibroacoustic stimulation.American Journal of Perinatology, 10, 46–49.
Groome, L. J., & Watson, J. E. (1992). Assessment of in utero neurobe-havioral development: I. Fetal behavioral states. Journal of Maternal-Fetal Investigation, 2, 183–194.
Hales, C., & Ozanne, S. (2003). The dangerous road of catch-up growth.Journal of Physiology, 547, 5–10.
Harding, J. (2001). The nutritional basis of the fetal origins of adultdisease. International Journal of Epidemiology, 30, 15–23.
Hepper, P. G. (1995). Fetal behavior and neural functioning. In J. P.Lecanuet, W. P. Fifer, N. A. Krasnegor, & W. P. Smotherman (Eds.),Fetal development: A psychobiological perspective (pp. 405–417).Hillsdale, NJ: Erlbaum.
Hepper, P. G., & Shahidullah, S. (1992). Habituation in normal andDown’s syndrome fetuses. Quarterly Journal of Experimental Psychol-ogy: Human Experimental Psychology, 44, 305–317.
Hepper, P., Shannon, E., & Dorman, J. (1998). Sex differences in fetalmouth movements. Lancet, 350, 1820.
Horimoto, N., Koyangi, T., Maeda, H., Satoh, S., Takashima, T., Minami,T., et al. (1993). Can brain impairment be detected by in utero behav-ioural patterns? Archives of Disease in Childhood, 69, 3–8.
Johnson, M., Paine, L., Mulder, H., Cezar, C., Gregor, C., & Johnson, T.(1992). Population differences of fetal biophysical and behavioral char-acteristics. American Journal of Obstetrics and Gynecology, 166, 138–142.
Johnson, T. R. B., Besinger, R. E., Thomas, R. L., Strobino, D. M., &Niebyl, J. R. (1992). Quantitative and qualitative relationships betweenfetal heart rate accelerations and fetal movement. Journal of Maternal-Fetal Medicine, 1, 251–253.
Junge, H. D. (1979). Behavioral states and state related heart rate andmotor activity patterns in the newborn infant and the fetus antepartum:A comparative study. Journal of Perinatal Medicine, 7, 85–107.
Kainer, F., Prechtl, H., Engele, H., & Einspieler, C. (1997). Assessment ofthe quality of general movements in fetuses and infants of women withtype-I diabetes mellitus. Early Human Development, 50, 13–25.
Kaplan, H., & Dove, H. (1987). Infant development among the Ache ofeastern Paraguay. Developmental Psychology, 23, 190–198.
Keefer, C., Tronick, E., Dixon, S., & Brazelton, T. (1982). Specificdifferences in motor performance between Gusii and American new-borns and a modification of the Neonatal Behavioral Assessment Scale.Child Development, 53, 754–759.
Kinney, H. C., Karthigason, J., Borenshteyn, N., Flax, J., & Kirschner, D.(1994). Myelination in the developing brain: Biochemical correlates.Neurochemical Research, 19, 983–996.
Kisilevsky, B. S., Muir, D. W., & Low, J. A. (1992). Maturation of humanfetal responses to vibroacoustic stimulation. Child Development, 63,1497–1508.
Kozuma, S., Nemoto, A., Okai, T., & Mizuno, M. (1991). Maturationalsequence of fetal breathing movements. Biology of the Neonate, 60,36–40.
Kuhlman, K. A., Burns, K. A., Depp, R., & Sabbagha, R. (1988). Ultra-sonic imaging of normal fetal response to external vibratory acousticstimulation. American Journal of Obstetrics and Gynecology, 258, 47–51.
454 DIPIETRO ET AL.

Lamb, B., & Lang, R. (1992). Aetiology of cerebral palsy. British Journalof Obstetrics and Gynecology, 99, 176–178.
Lauren, L., Jarvelin, M., Elliott, P., Sovio, U., Spellman, A., McCarthy, M.,et al. (2003). Relationship between birthweight and blood lipid concen-trations in later life: Evidence from the existing literature. InternationalJournal of Epidemiology, 32, 862–876.
Maeda, K., Tatsumura, M., & Utsu, M. (1999). Analysis of fetal move-ments by Doppler actocardiogram and fetal B-mode imaging. Clinics inPerinatology, 26, 829–851.
Manning, F. A., Platt, L. D., & Sipos, L. (1979). Fetal movements inhuman pregnancies in the third trimester. Obstetrics and Gynecology,54, 699–702.
Martin, C. (1978). Regulation of the fetal heart rate and genesis of FHRpatterns. Seminars in Perinatology, 2, 131–146.
Matthews, S. (2002). Early programming of the hypothalamo-pituitary-adrenal axis. Trends in Endocrinology and Metabolism, 13, 373–380.
Mulder, E. J. H. (1993). Diabetes in pregnancy as a model for testingbehavioral teratogenicity in man. Developmental Brain Dysfunction, 6,210–228.
Mulder, E. J., Morssink, L. P., van der Schee, T., & Visser, G. H. (1998).Acute maternal alcohol consumption disrupts behavioral state organiza-tion in the near term fetus. Pediatric Research, 44, 774–779.
Nasello-Paterson, C., Natale, R., & Connors, G. (1988). Ultrasonic eval-uation of fetal body movements over twenty-four hours in the humanfetus at twenty-four to twenty-eight weeks’ gestation. American Journalof Obstetrics and Gynecology, 158, 312–316.
Nelson, K. B., & Ellenberg, J. H. (1986). Antecedents of cerebral palsy:Multivariate analysis of risk. New England Journal of Medicine, 315,61–86.
Nijhuis, I., & ten Hof, J. (1999). Development of fetal heart rate andbehavior: Indirect measures to assess the fetal nervous system. EuropeanJournal of Obstetrics & Gynecology and Reproductive Biology, 87, 1–2.
Nijhuis, I., ten Hof, J., Mulder, E., Nijhuis, J., Narayan, H., Taylor, D., etal. (1998). Numerical fetal heart rate analysis: Nomograms, minimalduration of recording, and intrafetal consistency. Prenatal and NeonatalMedicine, 3, 314–322.
Nijhuis, I., ten Hof, J., Mulder, E., Nijhuis, J., Narayan, H., Taylor, D., etal. (2000). Fetal heart rate in relation to its variation in normal andgrowth retarded fetuses. European Journal of Obstetrics & Gynecologyand Reproductive Biology, 89, 27–33.
Nijhuis, I. J. M., ten Hof, J., Nijhuis, J. G., Mulder, E. J. H., Narayan, H.,Taylor, D. J., et al. (1999). Temporal organisation of fetal behavior from24 weeks gestation onwards in normal and complicated pregnancies.Developmental Psychobiology, 34, 257–268.
Nijhuis, J. G. (1986). Behavioural states: Concomitants, clinical implica-tions, and the assessment of the condition of the nervous system.European Journal of Obstetrics & Gynecology and Reproductive Biol-ogy, 21, 301–308.
Nijhuis, J. G., Prechtl, H. F. R., Martin, C. B., & Bots, R. S. G. (1982). Arethere behavioural states in the human fetus? Early Human Development,6, 47–65.
Ogueh, O., & Steer, P. (1998). Ethnicity and fetal heart rate variation.Obstetrics and Gynecology, 91, 324–328.
Pachter, L., & Harwood, R. (1996). Culture and child behavior and psy-chosocial development. Developmental and Behavioral Pediatrics, 17,191–198.
Patrick, J., Campbell, K., Carmichael, L., & Probert, C. (1982). Influenceof maternal heart rate and gross fetal body movements on the dailypattern of fetal heart rate near term. American Journal of Obstetrics andGynecology, 144, 533–538.
Petrie, B., & Segalowitz, S. (1980). Use of fetal heart rate, other perinataland maternal factors as predictors of sex. Perceptual and Motor Skills,50, 871–874.
Phillips, D. (2001). Programming of adrenocortical function and the fetalorigins of adult disease. Journal of Endocrinological Investigation, 24,742–746.
Phillips, D., & Barker, D. (1997). Association between low birthweight andhigh resting pulse in adult life: Is the sympathetic nervous systeminvolved in programming the insulin resistance syndrome? DiabeticMedicine, 14, 673–677.
Pillai, M., & James, D. (1990). Hiccups and breathing in human fetuses.Archives of Disease in Childhood, 65, 1072–1075.
Prechtl, H. F. R. (1984). Continuity and change in early neural develop-ment. In H. Prechtl (Ed.), Continuity in neural functions from prenatalto postnatal life (pp. 1–15). Philadelphia: J. B. Lippincott.
Ribbert, L., Fidler, V., & Visser, G. (1991). Computer-assisted analysis ofnormal second trimester fetal heart rate patterns. Journal of PerinatalMedicine, 19, 53–59.
Roberts, A. B., Griffin, D., Mooney, R., Cooper, D. J., & Campbell, S.(1980). Fetal activity in 100 normal third trimester pregnancies. BritishJournal of Obstetrics and Gynaecology, 87, 480–484.
Robertson, S. S. (1985). Cyclic motor activity in the human fetus aftermidgestation. Developmental Psychobiology, 18, 411–419.
Romanini, C., & Rizzo, G. (1995). Fetal behaviour in normal and com-promised fetuses: An overview. Early Human Development, 43, 117–131.
Roodenburg, P. J., Wladimiroff, J. W., van Es, A., & Prechtl, H. F. R.(1991). Classification and quantitative aspects of fetal movements dur-ing the second half of normal pregnancy. Early Human Development,25, 19–35.
Sacco, M., Caulfield, L., Zavaleta, N., & Retamozo, L. (2003). Dietarypattern and usual nutrient intakes of Peruvian women during pregnancy.European Journal of Clinical Nutrition, 57, 1492–1497.
Sachis, P. N., Armstrong, D., Becker, L., & Bryan, A. (1982). Myelinationof the human vagus nerve from 24 weeks postconceptional age toadolescence. Journal of Neuropathology and Experimental Neurology,41, 466–472.
Sandman, C. A., Wadhwa, P., Hetrick, W., Porto, M., & Peeke, H. (1997).Human fetal heart rate dishabituation between thirty and thirty-twoweeks gestation. Child Development, 68, 1031–1040.
Santos, D., Gabbard, C., & Goncalves, V. (2001). Motor developmentduring the first year: A comparative study. Journal of Genetic Psychol-ogy, 162, 143–153.
Solomons, G., & Solomons, H. (1975). Motor development in Yucatecaninfants. Developmental Medicine and Child Neurology, 17, 41–46.
Sontag, L. W., & Wallace, R. F. (1934). Preliminary report of the FelsFund. American Journal of Diseases of Children, 48, 1050–1057.
Super, C., & Harkness, S. (1986). The developmental niche: A conceptu-alization at the interface of child and culture. International Journal ofBehavioral Development, 9, 545–569.
Szeto, H. (1983). Effects of narcotic drugs on fetal behavioral activity:Acute methadone exposure. American Journal of Obstetrics and Gyne-cology, 146, 211–216.
Szitanyi, P., Janda, J., & Poledne, R. (2003). Intrauterine undernutritionand programming as a new risk of cardiovascular disease later in life.Physiological Research, 52, 389–395.
ten Hof, J., Nijhuis, I. J. M., Mulder, E. J. H., Nijhuis, J. G., Narayan, H.,Taylor, D. J., et al. (1999). Quantitative analysis of fetal generalizedmovements: Methodological considerations. Early Human Develop-ment, 56, 57–73.
ten Hof, J., Nijhuis, I. J. M., Mulder, E. J. H., Nijhuis, J. G., Narayan, H.,Taylor, D. J., et al. (2002). Longitudinal study of fetal body movements:Nomograms, intrafetal consistency and relationship with episodes ofheart rate patterns A and B. Pediatric Research, 52, 568–575.
Timor-Tritsch, I. E., Dierker, L. J., Zador, I., Hertz, R. H., & Rosen, M. G.(1978). Fetal movements associated with fetal heart rate accelerations
455FETAL NEUROBEHAVIORAL DEVELOPMENT

and decelerations. American Journal of Obstetrics and Gynecology, 131,276–280.
van de Pas, M., Nijhuis, J. G., & Jongsma, H. W. (1994). Fetal behaviourin uncomplicated pregnancies after 41 weeks of gestation. Early HumanDevelopment, 40, 29–38.
van Leeuwen, P., Lange, S., Bettermann, H., Gronemeyer, D., & Hatz-mann, W. (1999). Fetal heart rate variability and complexity in thecourse of pregnancy. Early Human Development, 54, 259–269.
van Vliet, M. A., Martin, C. B., Nijhuis, J. G., & Prechtl, H. F. R. (1985).Behavioural states in the fetuses of nulliparous women. Early HumanDevelopment, 12, 121–135.
Vindla, S., James, D., & Sahota, D. (1999). Computerised analysis ofunstimulated and stimulated behaviour in fetuses with intrauterinegrowth restriction. European Journal of Obstetrics & Gynecology andReproductive Biology, 83, 37–45.
Vindla, S., Sahota, D., Coppens, M., & James, D. (1997). Computerizedanalysis of behavior in fetuses with congenital abnormalities. Ultra-sound in Obstetrics and Gynecology, 9, 302–309.
Vintzileos, A. M., Campbell, W. A., & Nochinson, D. (1986). Relationbetween fetal heart rate accelerations, fetal movements, and fetal breath-ing movements. American Journal of Perinatology, 3, 16–18.
Ware, D. J., & Devoe, L. D. (1994). The nonstress test: Reassessment ofthe “gold standard”. Clinics in Perinatology, 21, 779–796.
Yoshizato, T., Koyanagi, T., Takashima, T., Satoh, S., Akazawa, K., &Nakano, H. (1994). The relationship between age-related heart ratechanges and developing brain function: A model of anencephalic humanfetuses in utero. Early Human Development, 36, 101–112.
Young, J. (2002). Programming of sympathoadrenal function. Trends inEndocrinology and Metabolism, 13, 381–385.
Zeger, S., & Liang, K. (1986). Longitudinal data analysis for discrete andcontinuous outcomes. Biometrics, 42, 121–130.
Received October 13, 2003Revision received December 22, 2003
Accepted December 31, 2003 �
456 DIPIETRO ET AL.