Embed Size (px)
Citation preview

INFECON AND IMMUNMTY, Sept. 1976, p. 660-666Copyright © 1976 American Society for Microbiology
Vol. 14, No. 3Printed in U.S.A.
Fetal and Adult Bovine Interferon Production During BovineViral Diarrhea Virus Infection
C. R. RINALDO, JR.,'* D. W. ISACKSON, J. C. OVERALL, JR., L. A. GLASGOW,T. T. BROWN,2 S. I. BISTNER, J. H. GILLESPIE, AND F. W. SCOTT
Departments ofPediatrics and Microbiology, University of Utah College of Medicine, Salt Lake City, Utah84132*; and Departments ofMicrobiology, Pathology, and Large Animal Medicine, Obstetrics and Surgery,
New York State College of Veterinary Medicine, Cornell University, Ithaca, New York 14853
Received for publication 15 April 1976
Levels of interferon in adult bovine serum and in fetal bovine serum andtissues were examined during the course oftransplacental bovine viral diarrheavirus infection. The cows produced circulating interferon between 2 and 9 daysafter viral inoculation, with mean peak levels in the serum on day 4. Interferoncould be routinely detected in fetal tissues (e.g., thymus, spleen, and kidney)between days 4 and 21 after viral inoculation of the cows at 149 to 150 days ofgestation (mid-second trimester) and in fetal serum from day 13 through day 21.Interferon was also detectable in the serum and tissues of fetuses from damsinfected at day 95 of gestation (the beginning of the second trimester). Ingeneral, no differences were found between the ability of the adult and fetus toproduce interferon. Fetal lamb kidney cells were more sensitive to the antiviraleffects of bovine interferon than were fetal bovine kidney cells. The antiviralsubstance from the fetal and adult animals was characterized as interferon bystandard criteria.
There are many examples both in man andexperimental animals of enhanced susceptibil-ity of the fetus to viral infection (3, 9, 30, 34).Congenital rubella virus and cytomegalovirusinfections are representative of such diseases ofthe human fetus. It has been well documentedthat infection of the fetus with rubella virus orcytomegalovirus during pregnancy can resultin severe congenital defects of the central nerv-ous system (CNS), eye, ear, heart, and skeleton(14, 18). In contrast, infection of children oradults with either ofthese viruses usually leadsto a relatively mild or asymptomatic illness(25).Bluetongue virus (21), hog cholera virus (15),
and bovine viral diarrhea (BVD) virus (8, 23,49) are examples of agents which may causesevere disease in the fetuses of domestic ani-mals. In the cow, which has a 9-month gesta-tion period, infection with BVD virus duringthe first trimester of pregnancy can result infetal mummification, stillbirth, or abortion (4,8, 24). Cerebellar and ocular lesions may de-velop in the fetuses of cows infected with BVDvirus during the second trimester of gestation
' Present address: Harvard Medical School, Departmentof Medicine, Massachusetts General Hospital, Boston, MA02114.
2 Present address: Department of Pathology, College ofVeterinary Medicine, Oklahoma State University, Stillwa-ter, OK 74074.
(4-6, 41). The BVD virus has little or no effecton the fetuses of cows infected during the lasttrimester of pregnancy (8, 24).The precise mechanisms involved in the age-
related resistance of the fetus to viral infectionhave not been well defined. Several investiga-tions have suggested that the inability of thefetal and newborn animal to control viral infec-tion may be related to an inadequate interferonresponse (1, 2, 13, 19, 22, 27, 32, 40). In contrastto these results, other studies have shown thatthe capacity to produce interferon is not re-duced in the immature host (10, 11, 31, 36, 44,48). Studies from this laboratory have shownthat the fetal lamb is as competent as the adultsheep to produce interferon in response to viralinfection either after direct inoculation in utero(33, 35) or in cell cultures derived from fetusesof various gestational ages (38).To more fully define the role of interferon in
age-related host resistance, the present studieswere undertaken to evaluate interferon produc-tion in the pregnant heifer and in the bovinefetus after transplacental infection with BVDvirus between 65 and 150 days of gestation.
MATERIALS AND METHODSCells and media. Fetal bovine kidney (FBK) cells
were prepared from kidneys removed asepticallyfrom bovine fetuses obtained from the MidvalePacking Co., Midvale, Utah. The renal capsule was
660
on June 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

INTERFERON IN BVD VIRUS INFECTION 661
removed, and the cortical and medullary regionswere minced and washed with phosphate-bufferedsaline. Cells were extracted in a 0.25% (wt/vol)trypsin sodium citrate-potassium chloride solutionfor 1 h, centrifuged, and resuspended in Eagle mini-mum essential medium consisting of Earle bal-anced salt solution supplemented with minimum es-sential medium vitamins and amino acids (GrandIsland Biological Co., Grand Island, N.Y.), 1% (volVvol) of a 300 ,ug/liter solution of glutamine, 100 U ofpenicillin per ml, 100 ,ug ofstreptomycin per ml, 10%(vol/vol) fetal calf serum (Grand Island Biological),and NaHCO3 as a buffer. The FBK cells were main-tained in 32-oz glass prescription bottles (BrockwayGlass Co., Brockway, Pa.) and used in the second tofifth passage. Procedures have previously been de-scribed (37) for the culturing of fetal lamb kidney(FLK) and mouse L cells.
Viruses. The cytopathogenic Holmes strain ofBVD virus used in these experiments was originallyisolated from the maternal and fetal tissues of apregnant cow that died ofnatural infection (42). Thestock virus was prepared and assayed in fetal bovinespleen cells, and the median tissue culture infectivedose was calculated by the Karber method (28). Theviral pools used in these experiments had a titer ofapproximately 104 median tissue culture infectivedoses per 0.1 ml. A detailed description has beenreported (37) for the preparation and assay ofvesicu-lar stomatitis virus (VSV) and Sindbis virus pools.
Experimental animals and procedures. The studywas divided into three phases. Pregnant Holstein-Friesian heifers with known breeding dates werehoused in an isolated barn in Ithaca, N.Y. Therewere 11 heifers used in phase I of the experiment, 10heifers in phase II, and 6 heifers in phase III. All ofthe heifers, with the exception of one heifer in phaseII, did not have serum-neutralizing antibody titersagainst BVD virus. The animal with neutralizingantibody was used as a control. The heifers wereserologically surveyed each month to ensure thatthe animals remained susceptible to the virus. Be-fore inoculation with virus, the pregnant heiferswere moved into environmentally controlled indi-vidual isolation units. Heifers in phases I and II ofthe study were intravenously inoculated with250,000 median tissue culture infective doses of theHolmes strain ofBVD virus at either 149 or 150 daysof gestation. Heifers in phase III of the study wereinoculated similarly with the same amount of thevirus at either 65, 75, 85, 95, or 115 days ofgestation.At sequential times after inoculation of the
heifers with BVD virus, the fetuses were surgicallyremoved by cesarean section from their dams. Im-mediately thereafter they were exsanguinated, andfetal serum and tissues were stored at -70°C forinterferon assay.
Interferon assay. The fetal and adult bovine sam-ples were transferred in dry ice by air freight to theUniversity of Utah College of Medicine. The sam-ples were assayed by the 50% plaque reduction tech-nique in FBK and FLK cells using VSV as thechallenge virus (37). An internal laboratory refer-ence bovine interferon was included with each as-say; titers varied from two- to threefold betweendifferent assays on FLK cells.
RESULTSInterferon production in the adult bovine.
The levels of circulating interferon in nine sus-ceptible pregnant heifers during the course ofBVD virus infection (phase II studies) are pre-sented in Fig. 1. As assayed in FBK cells, inter-feron was detectable from day 3 through day 6postviral inoculation and reached mean peaktiters of 36 U/ml on day 4. Interferon titersranged from undetectable levels to 100 U/ml inthe separate heifers. The serum interferon re-sponse of the heifers used in phase I and IIIexperiments was equivalent to that shown forphase II.During the course of the phase II studies, it
was discovered that FLK cells were much moresensitive to the antiviral effects of bovine inter-feron than were FBK cells. Therefore, phase IIand III samples were assayed for interferonactivity in both bovine (FBK) and sheep (FLK)cells. The titers of interferon in the serum ofthenine phase II BVD virus-infected heifers wereconsistently several-fold higher in the sheepthan in the bovine cells (Fig. 1). The mean peaktiter in FLK cells was 590 U/ml, approximately16-fold higher than in the FBK cells. The peaktiters reached levels greater than 1,000 U/ml inseveral animals. Although the kinetics of the
10,000A INTERFERON
ASSAYED ONFLK CELLS
0 INTERFERONASSAYED ONFBK CELLS
1,000
wcr-
¢, T'X
O 1 2 3 4 5 6 7 8 9 10DAYS POST- INOCULATION
FIG. 1. Mean levels and range of interferon titersinduced in the sera of nine dams inoculated intrave-nously with BVD virus and assayed in FLK and inFBK cells.
VOL. 14, 1976
on June 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

662 RINALDO ET AL.
interferon response followed a similar patternwith both assay systems, the enhanced sensi-tivity of the FLK cells allowed detection of se-rum interferon from day 2 through day 9.
In comparing the sensitivity of the sheep andbovine cells, an interferon standard kindly do- t
nated by J. D. Todd (Jensen-Salsbery Labs, -Kansas City, Mo.) was assayed in both cell tlines. The standard consisted of nasal secre-tions from calves which had been inoculated >with infectious bovine rhinotracheitis virus and >,was known to contain antiviral activity which 2had been characterized as interferon (46). Thebovine interferon sample titered 200 U/ml in -6FBK cells and 715 U/ml in FLK cells, i.e., tnearly a fourfold increase in the sheep cells. >
Interferon production in the bovine fetus:phase I studies. Data on the production of in-terferon in the bovine fetus after inoculation of Qtheir dams with BVD virus during the phase I >studies are presented in Table 1. Interferon waspresent in fetal serum from 13 to 21 days after 2inoculation of the cows with virus and ranged mfrom 4 to 70 U/ml as assayed in FBK cells. Low ,levels of interfering activity were also noted in ithe serum of fetuses taken at 27 and 56 days .postviral inoculation. Interferon was detectedin fetal tissues between 4 and 21 days after viral .Einfection of the dams with highest levels con-sistently present in the liver, thymus, and kid- aney. Interferon was found irregularly and atlower levels in spleen, lung, mesenteric lymph 2node, and cotyledon.One possible explanation for the low or unde- *
tectable levels of interferon in fetuses removed C
from 21 to 70 days was that they were not >infected with BVD virus. However, these fe- 'tuses were shown to be infected as evidenced by
.
the presence of elevated levels of serum immu- onoglobulins M and G1 (T. T. Brown et al., un- -published observations), and cerebellar and eocular lesions typical ofBVD virus infection (5, t6).BVD virus is known to cause severe lesions >
in the CNS of fetuses infected at 150 days of 2gestation (5, 6, 7, 41). However, interferon was ;detectable in the CNS tissue of only one fetus 0during the phase I studies (Table 1). Low levels 0
of interfering activity (4 U/ml) were found in ,cerebrospinal fluid of fetuses removed at 17, 21, .and 27 days postviral inoculation. Since thetiters were low, the interfering activity found <in the one CNS tissue sample and in the three E-spinal fluid samples could not be characterizedas interferon. However, a recent report hasshown exogenous interferon to be capable ofcrossing from blood to spinal fluid in adult mon-keys (17).
3-,.
00
0
E
,s .X.
n.0
0*
00
00
INFECT. IMMUN.
le v v10 v v- v Iv vvvvvvvvvvvvvvv
0v v 0 00v 0
v vIttttVvvV vVvvvcq
v v v vv v
V ~ V VIV VVVV
CD: CD QCD CDt= CD CDt L
0 000V V V V
0100CD10100"0 lw'
V-, V4V VV VV V
> = UD v w = 1" Ild v v v I"3-404 VV V V
CNO-
S. 0
4 Q
;0%
10
U
-Cd04
I.~
.5 *~ 0)
, m
S-0-4
0 0000000000tt NV V -V VVVVV
"' 10 w00 I" v v v v vVN~-400V V V V V VvV -vvvvv
vvVV-I VVVVV
#' t- O0 t' O 'CtCt 100
CD~C4 CD3 C~1 CDI~'1
00000000'I'0d,
V V-- V V ,-4V
> _C_ CD CDCS CDU=
tO La LO LO LO LO in LO LO
r- r-.cs4eDIuz4 r4oo4c--_
I- Cq 10O
00 t-LO10 l 10 l' l' 10010
$42
euE -t4)k I 4140 04 204E
-4-a cd 4.1,
"-I0 0.:) z .:)01a .0 ;d
on June 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

INTERFERON IN BVD VIRUS INFECTION 663
The data suggest that the majority of fetalinterferon production in response to intrauter-ine BVD virus infection occurs from day 4through day 21 postinoculation. Furthermore,levels of interferon in the sera of bovine fetuses(Table 1) were comparable to those of adult seraas assayed in FBK cells (Fig. 1).
Interferon production in the bovine fetus:phase II studies. Based on the results of thephase I experiments, the phase II studies con-
centrated on interferon production in fetusesremoved from dams between 12 and 21 dayspostinoculation of BVD virus. Interferon, asassayed in FLK cells, was present in the major-ity of fetal serum, kidney, spleen, and thymussamples but in only a few of the cotyledon,lung, and liver samples. The titers of inter-feron, however, were similar in all tissues. In-terferon was not demonstrable in CNS tissue.Titers of interferon in fetal serum and tissuesas assayed in FLK cells (Table 2) were gener-ally higher than the interferon levels detectedin fetal samples from the phase I studies whichused the FBK cell assay system (Table 1).
Fetus no. 06 from a control heifer with spe-cific antiserum activity against BVD virus didnot contain interferon in serum or tissues (Ta-ble 2). This fetus also failed to produce serum-neutralizing antibody against BVD virus(Brown et al., unpublished observations). Themother of this fetus did not produce circulatinginterferon in response to inoculation with BVDvirus. It is therefore presumed that the anti-BVD antibody present in the serum protectedthe cow, and hence the fetus, against infectionwith BVD virus.
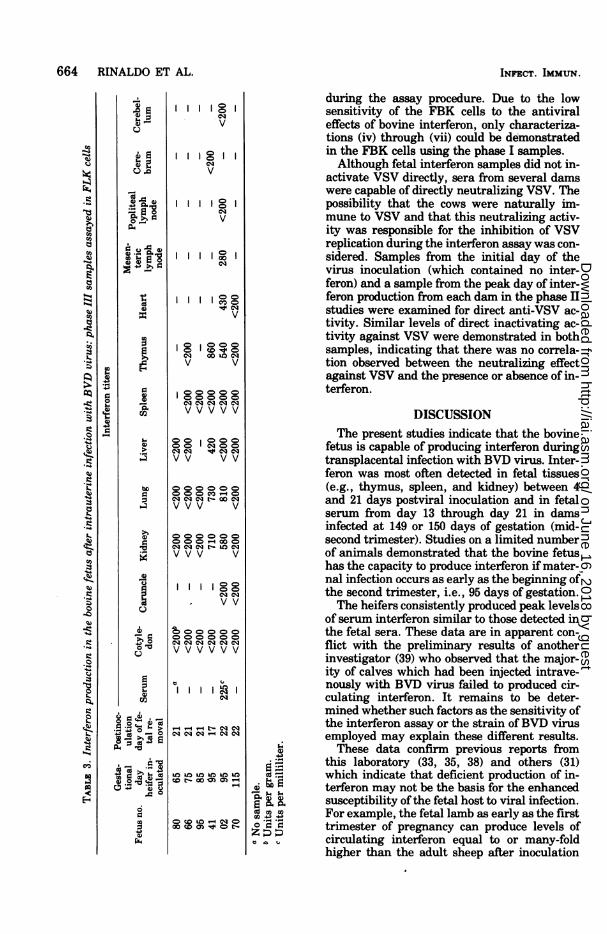
Interferon production in the bovine fetus:phase III studies. In phase III of these investi-gations, six pregnant heifers were inoculatedwith BVD virus during the late first and earlysecond trimesters of gestation (65 to 115 days).Fetuses were removed from these animals be-tween 17 and 22 days postinoculation, and thetissues were examined for evidence of inter-feron production. Table 3 contains data whichdemonstrate that fetuses from dams inoculatedwith BVD virus at 95 days of gestation (no. 41and 02) produced interferon levels similar tothose found in the phase II studies. Tissuesfrom the other four fetuses of the phase IIIstudies did not contain detectable interferon.Fetuses no. 41 and 02 were demonstrated to beinfected with BVD virus by the occurrence ofimmunoglobulin M in the serum and cerebellarand ocular lesions (6; Brown et al., unpublishedobservations). Ofthe four fetuses which did notproduce interferon, no. 70 was dead at the timeofcesarotomy and serum immunoglobulins andcerebellar or ocular degeneration could not bedemonstrated in fetuses no. 80, 66, and 95.Characterization of the fetal and adult bo-
vine interferon. The antiviral substance in-duced in the fetal and adult bovine by BVDvirus was characterized as interferon by thefollowing criteria: (i) inactivation by trypsintreatment; (ii) activity against both VSV andSindbis virus in FLK cells; (iii) lack of activityin FLK cells treated with actinomycin C; (iv)resistance to pH 2 for 24 h at 4°C; (v) lack ofdetectable activity in mouse L cells; (vi) stabil-ity at 560C for 30 min; (vii) failure of the inter-fering activity to be washed off the FLK cells
TABLE 2. Interferon production in the bovine fetus after intrauterine infection with BVD virus: phase IIsamples assayed in FLK cells
Gesta- Postin- Interferon titerstional ocula-
Fetus day tion day Mesen-no. heifer offtlSrmCotyle- KinyLn ie pen
Thy- teric Cere- Cere-inc-of fetal Serum Kidney Lung Liver Spleen ms lmhbuihlulinocu removal don mus lymph brum bellum
lated rmvlnode87 149 12 500a 460b -C 720 200 760 880 - - -01 149 14 500 <400 850 - - <400 400 - <400 -63 150 15 480 <200 610 670 770 1000 360 - <200 <20062 150 16 370 <200 - <400 <400 - - - - -78 150 17 250 - 1200 <400 - <200 <200 - - -94 150 18 425 <200 450 850 <200 950 250 - <200 -48 150 19 300 <200 500 <200 <200 600 200 - <200 -
8,974 150 20 245 <200 <200 <200 <200 <200 930 - <200 <20056 150 21 450 - 600 - - - - 750 - -06Y 150 21 <20 <200 <200 <200 - <200 <200 - <200 <200
a Units per milliliter.b Units per gram.c No sample.d Control heifer immune to BVD-MD virus.
VOL. 14, 1976
on June 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
664 RINALDO ET AL.
18eqv
Igv
Igeqv
II Ogv
C1 00 ell clv vCD CDCD Q
cq cq esq cq cv v v v v
I-C=
eqeq =eq>X. VV VV
bDa 0V0CV V-4053 Cq " "t--4 VV V
O OO-4 0
eq eq Ce t- La e
VVV V
IIgeq eqv
CQ
,VV
t- eq eq4eq eq cq - cq eq
et- Cs QO -O H so eq 0s
Cs c' 0s t e-
CD m00W Q t
INFECT. IMMUN.
co'
bCoCoCoCo -4. 4.)
S-
during the assay procedure. Due to the lowsensitivity of the FBK cells to the antiviraleffects of bovine interferon, only characteriza-tions (iv) through (vii) could be demonstratedin the FBK cells using the phase I samples.Although fetal interferon samples did not in-
activate VSV directly, sera from several damswere capable of directly neutralizing VSV. Thepossibility that the cows were naturally im-mune to VSV and that this neutralizing activ-ity was responsible for the inhibition of VSVreplication during the interferon assay was con-sidered. Samples from the initial day of thevirus inoculation (which contained no inter-feron) and a sample from the peak day of inter-feron production from each dam in the phase IIstudies were examined for direct anti-VSV ac-tivity. Similar levels of direct inactivating ac-tivity against VSV were demonstrated in bothsamples, indicating that there was no correla-tion observed between the neutralizing effectagainst VSV and the presence or absence of in-terferon.
DISCUSSIONThe present studies indicate that the bovine
fetus is capable of producing interferon duringtransplacental infection with BVD virus. Inter-feron was most often detected in fetal tissues(e.g., thymus, spleen, and kidney) between 4and 21 days postviral inoculation and in fetalserum from day 13 through day 21 in damsinfected at 149 or 150 days of gestation (mid-second trimester). Studies on a limited numberof animals demonstrated that the bovine fetushas the capacity to produce interferon if mater-nal infection occurs as early as the beginning ofthe second trimester, i.e., 95 days of gestation.The heifers consistently produced peak levels
of serum interferon similar to those detected inthe fetal sera. These data are in apparent con-flict with the preliminary results of anotherinvestigator (39) who observed that the major-ity of calves which had been injected intrave-nously with BVD virus failed to produced cir-culating interferon. It remains to be deter-mined whether such factors as the sensitivity ofthe interferon assay or the strain ofBVD virusemployed may explain these different results.These data confirm previous reports from
this laboratory (33, 35, 38) and others (31)which indicate that deficient production of in-terferon may not be the basis for the enhancedsusceptibility ofthe fetal host to viral infection.For example, the fetal lamb as early as the firsttrimester of pregnancy can produce levels ofcirculating interferon equal to or many-foldhigher than the adult sheep after inoculation
Io0)
U CoCo )CoCoh
Co0 rCo~
Cs
056-
It
S)
V.)
co
co
Co
3o
Co5..4
V.)
Co
5..c)
2Q.
EC
C)
O0
d
C),
on June 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

INTERFERON IN BVD VIRUS INFECTION 665
with Chikungunya virus (33, 35). Further stud-ies are necessary to determine whether an inef-ficient interferon response may be of signifi-cance in the enhanced susceptibility of the firsttrimester bovine fetus to BVD virus infection.The fetuses used in phase I and II studies
have been shown to be transplacentally in-fected with BVD virus as evidenced by grossand microscopic lesions in the brain and eyes(5, 6) and by the production ofelevated levels ofserum immunoglobulins M and G1 (Brown etal., unpublished observations). Although sev-eral attempts were made to isolate BVD virusfrom fetal tissues, recovery was successful inonly a few instances. Unfortunately the strainof BVD virus used in these studies caused in-consistent cytopathic effect in tissue culture,and attempts to assay virus using viral inter-ference techniques were also unsuccessful. Theearliest time of postmaternal inoculation thatBVD was isolated from fetal tissues was 12 days(fetus no. 87, Table 2). Histopathologic lesionsand serologic evidence of infection were notedat 12 days and later. However, our data oninterferon production suggest that the bovinefetus may be infected as early as 4 to 7 days.Other investigators, using the NADL strain ofBVD and direct fetal inoculation, have re-covered virus from brain, kidney, lung, spleen,lymph node, buffy coat, and placenta (4, 8). Thehistopathologic changes we observed in theCNS would suggest that BVD virus replicatedwithin CNS tissue in this model infection. It isof interest that we failed to demonstrate inter-feron in any of the CNS tissue samples exam-ined. It is at present unknown whether fetalCNS tissue infected with virus is incapable ofproducing interferon or whether interferon pro-duced by the other fetal tissues fails to enter theCNS.
It is possible that the presence of interferonin fetuses prior to 14 days after infection of thedams may represent passively transferred ma-ternal interferon. Transplacental transfer of in-terferon has been observed during the course ofnonpathogenic viral infections in rodents (20,26, 31). In contrast, data from this laboratory(33) have shown that interferon induced by anonreplicating togavirus in pregnant sheep orin fetal lambs fails to pass transplacentally ineither direction. The basic similarities betweenthe bovine and sheep placental structureswould suggest that interferon is unlikely to bepassively transferred in the bovine. Furtherevidence against the transplacental transfer ofmaternal interferon in this study is the obser-vation that interferon was not detectable in theserum of two fetuses (no. 44 and 47, Table 1) at
a time when it was present in their maternalserum.An interesting observation in the present
studies is that the antiviral activity of bovineinterferon is greatly enhanced in sheep as com-pared with bovine cells. Although interferonsgenerally display a characteristic species spe-cifity, it is known that certain interferons areactive in cells of different animal species (29).For example, other investigators have shownthat bovine interferon exhibits antiviral activ-ity in porcine (R. Ahl and A. Rump, personalcommunication), human (45), and certain mon-key (43) cells but is inactive in chicken chorioal-lantoic membrane (47) and other types of mon-key cells (45). Human interferon has been re-ported to have greatly enhanced antiviral activ-ity in rabbit kidney cells (12) and various typesof bovine and porcine cells (16), comparable tothe pattern ofincreased activity observed in ourstudies of bovine interferon in sheep cells. Thepresent results suggest, therefore, that sheepcells may provide a highly sensitive cell systemfor the assay of bovine interferon.
ACKNOWLEDGMENTSThis investigation was supported by a subcontract from
the New York State College of Veterinary Medicine atCornell University through Public Health Service grantHDAI-05757 from the National Institute of Child Healthand Human Development and Public Health Service grantAI10217 from the National Institute of Allergy and Infec-tious Diseases. J. C. Overall is an investigator of the How-ard Hughes Medical Institute.
LITERATURE CITED1. Barbosa, L. H., W. T. London, R. Hamilton, and C.
Buckler. 1974. Interferon response of the fetal rhesusmonkey after viral infection. Proc. Soc. Exp. Biol.Med. 146:398-400.
2. Baron, S., and A. Isaacs. 1961. Mechanism of recoveryfrom viral infection in the chick embryo. Nature(London) 191:97-98.
3. Blattner, R. J., A. P. Williamson, and F. M. Heys.1973. Role of viruses in the etiology of congenitalmalformations. Prog. Med. Virol. 15:1-41.
4. Braun, R. K., B. I. Osburn, and J. W. Kendrick. 1973.Immunologic response of bovine fetus to bovine viraldiarrhea virus. Am. J. Vet. Res. 34:1127-1132.
5. Brown, T. T., A. deLahunta, S. I. Bistner, F. W. Scott,and K. McEntee. 1974. Pathogenetic studies of infec-tion of the bovine fetus with bovine viral diarrheavirus. I. Cerebellar atrophy. Vet. Pathol. 11:486-505.
6. Brown, T. T., A. deLahunta, S. I. Bistner, F. W. Scott,and K. McEntee. 1975. Pathogenetic studies of infec-tion of the bovine fetus with bovine viral diarrheavirus. II. Ocular lesions. Vet. Pathol. 12:394-404.
7. Brown, T. T., A. deLahunta, F. W. Scott, R. F. Kahrs,K. McEntee, and J. H. Gillespie. 1973. Virus inducedcongenital anomalies of the bovine fetus. II. Histopa-thology of cerebellar degeneration (hypoplasia) in-duced by the virus of bovine viral diarrhea-mucosaldisease. Cornell Vet. 63:561-578.
8. Casaro, A. P. E., J. W. Kendrick, and P. C. Kennedy.1971. Response of the bovine fetus to bovine viraldiarrhea-mucosal disease virus. Am. J. Vet. Res.32:1543-1562.
VOL. 14, 1976
on June 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

666 RINALDO ET AL.
9. Catalano, L. W., Jr., and J. L. Sever. 1971. The role ofviruses as causes of congenital defects. Annu. Rev.Microbiol. 25:255-282.
10. Cole, G. A., and C. L. Wisseman, Jr. 1969. Pathogen-esis of type 1 dengue virus infection in suckling,wealing and adult mice. I. The relation of virus repli-cation to interferon and antibody formation. Am. J.Epidemiol. 89:669-680.
11. Craighead, J. E. 1966. Growth of parainfluenza type 3virus and interferon production in infant and adultmice. Br. J. Exp. Pathol. 47:235-241.
12. Desmyter, J., W. E. Rawls, and J. L. Melnick. 1968. Ahuman interferon that crosses the species line. Proc.Natl. Acad. Sci. U.S.A. 59:69-76.
13. Desmyter, J., W. E. Rawls, J. L. Melnick, M. D. Yow,and F. F. Barrett. 1967. Interferon in congenital ru-bella: response to live attenuated measles vaccine. J.Immunol. 99:771-777.
14. Dudgeon, J. A. 1969. Congenital rubella. Pathogenesisand immunology. Am. J. Dis. Child. 118:35-44.
15. Dunne, H. W., and C. D. Clark. 1968. Embryonic death,fetal mummification, stillbirth, and neonatal deathin pigs of gilts vaccinated with attenuated live-virushog cholera vaccine. Am. J. Vet. Res. 29.787-796.
16. Gresser, I., M-T. Bandu, D. Brouty-Boye, and M.Tovey. 1974. Pronounced antiviral activity of humaninterferon on bovine and porcine cells. Nature (Lon-don) 251:543-545.
17. Habif, D. V., R. Lipton, and K. Cantell. 1975. Inter-feron crosses blood-cerebrospinal fluid barrier inmonkeys. Proc. Soc. Exp. Biol. Med. 149:287-289.
18. Hanshaw, J. B. 1971. Congenital cytomegalovirus in-fection: a fifteen year perspective. J. Infect. Dis.123:555-561.
19. Heineberg, H., E. Gold, and F. C. Robbins. 1964. Dif-ferences in interferon content in tissues of mice ofvarious ages infected with Coxsackie Bi virus. Proc.Soc. Exp. Biol. Med. 115:947-953.
20. Ho, M., B. Postic, and Y. H. Ke. 1968. The systemicinduction of interferon, p. 19-38. In G. E. W. Wolsten-holme and M. O'Conner (ed.), Interferon. J. & A.Churchill, London.
21. Howell, P. G., and D. W. Verwoerd. 1971. Bluetonguevirus. Virol. Monogr. no. 9:35-74.
22. Isaacs, A., and S. Baron. 1960. Antiviral action ofinter-feron in embryonic cells. Lancet 2:946-947.
23. Kahrs, R. F. 1973. Effects of bovine viral diarrhea onthe developing fetus. J. Am. Vet. Med. Assoc.163:877-878.
24. Kendrick, J. W. 1971. Bovine viral diarrhea-mucosaldisease virus infection in pregnant cows. Am. J. Vet.Res. 32:533-544.
25. Kibrick, S., and R. M. Loria. 1974. Rubella and cyto-megalovirus. Current concepts of congenital and ac-quired infection. Pediatr. Clin. North Am. 21:513-526.
26. Korsantiya, B. M., and A. A. Smorodintsev. 1971.Transplacental transmission of endogenous inter-feron in pregnant mice inoculated with influenza orNewcastle disease viruses. Nature (London) 232:560-561.
27. Lavelle, G. C., and T. J. Starr. 1968. Interferon re-sponse and age-related resistance of germfree mice tomouse hepatitis virus. J. Reticuloendothel. Soc.5:422-435.
28. Lennette, E. H., and N. J. Schmidt (ed.). 1969. Diag-nostic procedures for viral and rickettsial infections,4th ed., p. 49. Am. Public Health Assoc., Inc., NewYork.
29. Lockart, R. Z., Jr. 1973. Criteria for acceptance of aviral inhibitor as an interferon and a general descrip-tion of the biological properties ofknown interferons,p. 11-27. In N. B. Finter (ed.), Interferons and inter-feron inducers, 2nd ed. American Elsevier Publishing
Co., Inc., New York.30. Mims, C. A. 1968. Pathogenesis of viral infections of the
fetus. Prog. Med. Virol. 10:194-237.31. Mendelson, J., R. Kapusta, and V. Dick. 1970. Mater-
nal and fetal interferon production in the rat. Am. J.Obstet. Gynecol. 107:902-907.
32. Morahan, P. S., and S. E. Grossberg. 1970. Age-relatedcellular resistance of the chicken embryo to viralinfections. I. Interferon and natural resistance tomyxoviruses and vesicular stomatitis virus. J. Infect.Dis. 121:615-623.
33. Overall, J. C., Jr., and L. A. Glasgow. 1970. Fetalresponse to viral infection: interferon production insheep. Science 167:1139-1141.
34. Overall, J. C., Jr., and L. A. Glasgow. 1970. Virusinfections of the fetus and newborn infant. J. Pediatr.77:315-333.
35. Overall, J. C., Jr., and L. A. Glasgow. 1971. Host resist-ance to virus infection in the fetus. I. Interferon (IF)production. Pediatr. Res. 5:403.
36. Reinarz, A. B. G., M. G. Broome, and B. P. Sagik.1971. Age-dependent resistance of mice to Sindbisvirus infection: viral replication as a function of hostage. Infect. Immun. 3:268-273.
37. Rinaldo, C. R., Jr., J. C. Overall, Jr., B. C. Cole, and L.A. Glasgow. 1973. Mycoplasma-associated inductionof interferon in ovine leukocytes. Infect. Immun.8:796-803.
38. Rinaldo, C. R., Jr., J. C. Overall, Jr., and L. A. Glas-gow. 1975. Viral replication and interferon produc-tion in fetal and adult ovine leukocytes and spleencells. Infect. Immun. 12:1070-1077.
39. Rosenquist, B. D. 1973. Induction and biological signifi-cance of interferon, with particular reference to cat-tle. J. Am. Vet. Med. Assoc. 163:821-824.
40. Sawicki, L. 1961. Influence of age of mice on the recov-ery from experimental Sendai virus infection. Nature(London) 192:1258-1259.
41. Scott, F. W., R. F. Kahrs, A. deLahunta, T. T. Brown,K. McEntee, and J. H. Gillespie. 1973. Virus inducedcongenital anomalies of the bovine fetus. I. Cerebel-lar degeneration (hypoplasia), ocular lesions and fe-tal mummification following experimental infectionwith bovine viral diarrhea-mucosal disease virus.Cornell Vet. 63:536-560.
42. Scott, F. W., R. F. Kahrs, and I. M. Parsonson. 1972. Acytopathogenic strain of bovine viral diarrhea-mu-cosal disease virus isolated from a bovine fetus. Cor-nell Vet. 62:74-84.
43. Sellers, R. F., and M. Fitzpatrick. 1962. An assay ofinterferon produced in rhesus monkey and calf kid-ney tissue cultures using bovine enterovirus M6 aschallenge. Br. J. Exp. Pathol. 43:674-683.
44. Subrahmanyan, T. P. 1968. A study of the possiblebasis of age-dependent resistance of mice to poxvirusdiseases. Aust. J. Exp. Biol. Med. Sci. 46:251-265.
45. Sutton, R. N. P., and D. A. J. Tyrrell. 1961. Someobservations on interferon prepared in tissue cul-tures. Br. J. Exp. Pathol. 42:99-105.
46. Todd, J. D., F. J. Volenec, and I. M. Paton. 1972.Interferon in nasal secretions and sera of calves afterintranasal administration of avirulent infectious bo-vine rhinotracheitis virus: association of interferon innasal secretions with early resistance to challengewith virulent virus. Infect. Immun. 5:699-706.
47. Tyrrell, D. A. J. 1959. Interferon produced by cultures ofcalf kidney cells. Nature (London) 184:452-453.
48. Vilcek, J. 1964. Production of interferon by newbornand adult mice infected with Sindbis virus. Virology22:651-652.
49. Ward, G. M., S. J. Roberts, K. McEntee, and J. H.Gillespie. 1969. A study of experimentally inducedbovine viral diarrhea-mucosal disease in pregnantcows and their progeny. Cornell Vet. 59:525-538.
INFECT. IMMUN.
on June 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from







![Leukocyte Dysfunction in the Bovine Homologue of Chediak ...iai.asm.org/content/10/4/928.full.pdf · RENSHAWETAL. mined with [1-_4C]glucose and [6-"4C]glucose by a modification ofthe](https://img.pdfslide.us/doc/110x75/5b7cf5227f8b9ada6d8bb486/leukocyte-dysfunction-in-the-bovine-homologue-of-chediak-iaiasmorgcontent104928fullpdf.jpg)





















