Embed Size (px)
Citation preview

Fertilization Stimulates AnaerobicFuel Degradation of Antarctic Soilsby Denitrifying MicroorganismsS H A N E M . P O W E L L , * , †
S U S A N H . F E R G U S O N , ‡ I A N S N A P E , ‡ A N DS T E V E N D . S I C I L I A N O §
School of Agricultural Science, University of Tasmania,Hobart 7001, Australia, Department of Heritage andEnvironment, Australian Antarctic Division,Kingston 7050, Australia, and University of Saskatchewan,Saskatoon, Canada S7N 5A8
Human activities in the Antarctic have resulted inhydrocarbon contamination of these fragile polar soils.Bioremediation is one of the options for remediation of thesesites. However, little is known about anaerobic hydrocarbondegradation in polar soils and the influence of bioreme-diation practices on these processes. Using a field trial atOld Casey Station, Antarctica, we assessed the influenceof fertilization on the anaerobic degradation of a 20-year oldfuel spill. Fertilization increased hydrocarbon degradationin both anaerobic and aerobic soils when compared tocontrols, but was of most benefit for anaerobic soils whereevaporation was negligible. This increased biodegradationin the anaerobic soils corresponded with a shift in thedenitrifier community composition and an increasedabundance of denitrifiers and benzoyl-CoA reductase. Amicrocosm study using toluene and hexadecane confirmedthe degradative capacity within these soils underanaerobic conditions. It was observed that fertilizedanaerobic soil degraded more of this hydrocarbon spikewhen incubated anaerobically than when incubatedaerobically. We conclude that denitrifiers are activelyinvolved in hydrocarbon degradation in Antarctic soils andthat fertilization is an effective means of stimulating theiractivity. Further, when communities stimulated to degradehydrocarbons under anaerobic conditions are exposedto oxygen, hydrocarbon degradation is suppressed. Thecommonly accepted belief that remediation of polar soilsrequires aeration needs to be reevaluated in light of this newdata.
IntroductionPolar soils have been contaminated by petroleum hydro-carbons as the result of accidental spillage during fueltransport and transfer, and from leaking vehicles and storagetanks (1). There are no reliable estimates of the amount ofcontaminated soil in the Antarctic, but it is possibly in theorder of ∼1 million m3 (2). The most common managementresponse to this problem has been to rely on naturalattenuation or, occasionally, to excavate and ship the soil to
a temperate region for remediation (2). Excavation isexpensive and can have severe environmental impacts. Thiscombination of cost and environmental risk has promotedresearch into in situ alternatives such as bioremediation.
In laboratory studies, Antarctic soil organisms degradehydrocarbons such as n-alkanes, monoaromatics, and naph-thalenes under aerobic conditions (3, 4), and field resultsindicate that a range of catabolic genes exist in Antarcticsoils (5). Unfortunately, degradation rates are limited due tonutrient deficiencies, low temperatures, poor water holdingcapacities, and frozen water (6-8), but these limitations canbe overcome by techniques such as fertilization, temperaturecontrol, and irrigation (7, 8).
It is commonly assumed that there is sufficient aerationof Antarctic soils to prevent oxygen limitation (1). Thisassumption is based on gravimetric water contents between3% and 11% and bulk densities between 1 and 1.2 g cm-3 (2)with a corresponding air volume fraction between 40% and50%. The relatively low oxygen demand (∼600 µg O2 g-1 soilday-1) characteristic of many Antarctic soils suggests thatthere will be sufficient oxygen to prevent hypoxia (9).However, some Antarctic soils display gleying, an indicationof oxygen limitation (10-12), and even in aerobic soilsanaerobic conditions exist in anoxic microsites (13). Underintensive bioremediation where there is a carbon source(hydrocarbons) and nutrients (fertilizer) present in excess,it is probable that certain soil regions will become hypoxicor anoxic.
Degradation of hydrocarbons by anaerobic organisms isdifferent from aerobic degradation, as they do not use theoxygenases used by aerobic organisms (14, 15). Anerobicalkane degradation is poorly understood, but appears to beinitiated by a fumarate addition via a radical reaction,eventually forming an alkyl-CoA compound later oxidizedby â-oxidation (16-18). Degradation of alkyl-substitutedaromatic hydrocarbons commences with the addition offumarate to alkyl groups that is then oxidized to producebenzoyl coenzyme A (CoA). Benzoyl-CoA is reduced bybenzoyl-CoA reductase and used for energy and carbon bythe cell (19). Recently, two research groups developedconserved primers to amplify portions of the genes respon-sible for benzoyl-CoA reductase (14, 20). These primers areuseful but are limited to the detection of the genes for alkyl-substituted aromatic degradation by some anaerobic organ-isms.
Denitrifiers are common anaerobic soil organisms (21),and several isolates can metabolize hydrocarbons (16, 22,23) using benzoyl-CoA reductase and the fumarate additionpathway. Denitrification is a respiratory process; however,denitrifying bacteria can either be obligately or facultativelyanaerobic. Denitrification, the sequential reduction of nitrateto di-nitrogen gas, involves the genes nar/nap, nir, nor, andnos encoding for nitrate reductase, nitrite reductase, nitricoxide reductase, and nitrous oxide reductase, respectively(21). The nir and nosZ genes are considered indicative ofdenitrifying bacteria (24). There are two distinct forms ofnitrite reductase, one with a cytochrome center (nirS) andone with a copper center (nirK); thus a quantitative assess-ment of denitrifiers in soil is more easily accomplished usingprimers specific for nosZ (25). However, in situ confirmationof denitrification activity is technically challenging. As nitratereduction will not occur until oxygen levels have becomedepleted and iron reduction will not occur until nitrate hasbeen consumed, the sequential use of electron acceptors isan accepted method of determining the dominant bio-geochemical process (26).
* Corresponding author phone: +61 03 6226 2776; fax: +61 036226 2642; e-mail: [email protected].
† University of Tasmania.‡ Australian Antarctic Division.§ University of Saskatchewan.
Environ. Sci. Technol. 2006, 40, 2011-2017
10.1021/es051818t CCC: $33.50 2006 American Chemical Society VOL. 40, NO. 6, 2006 / ENVIRONMENTAL SCIENCE & TECHNOLOGY 9 2011Published on Web 02/14/2006

Denitrification is thought to be a minor process in polarenvironments because of nitrate limitations and oxygensurpluses. However, in the case of bioremediation, substantialnitrate is usually added and metabolic activity is stimulated.Thus, the nitrate limitation is removed and partial oxygenpressures reduced. In Antarctica, little is known aboutdenitrifiers in soil with all previous work investigatingdenitrifiers in aquatic systems (27-29) and marine sediments(30). These investigators all found that denitrification doesoccur in Antarctica but to a limited extent. In contrast,denitrification is an important biogeochemical process inthe Arctic (31, 32). Under nitrate-reducing conditions,hydrocarbon degradation by Arctic soil isolates was notlimited by low temperatures with little difference seen inhydrocarbon degradation at 20 °C as compared to that at 7°C (6).
We hypothesized that, in fertilized soils, denitrifiers wouldcontribute significantly to hydrocarbon degradation. If so, itmay not be necessary to aerate contaminated soil sites. It iseven possible that denitrification-based hydrocarbon bio-degradation may prove to be more active at lower temper-atures than alternative treatments involving expensive soilheating. This would allow low cost in situ treatment toproceed at a sufficient rate to achieve remediation goals ina reasonable time, thus reducing operational costs and theenvironmental risk. To investigate these possibilities, weexamined the differences in hydrocarbon biodegradation andthe structure of denitrifying communities in nutrient-treated,hydrocarbon contaminated soils in Antarctica.
Materials and MethodsField Site and Experiment. The study site is located in EastAntarctica near the Old Casey Research Station and has beendescribed elsewhere (2, 33). Approximately 1000 m3 of soilis contaminated from historical fuel spills that occurred over20 years ago. Total petroleum hydrocarbons (TPH) concen-trations at the site range between 10 000 and 47 000 mg kg-1.The main contaminant is Special Antarctic Blend (SAB) diesel,which contains predominantly n-alkanes. A landfarming trialwas initiated in 1998 and has been previously described (2).Briefly, contaminated soil was sieved (<5.0 mm) andhomogenized before being added to hexane-cleaned metalcontainers (240 (w) × 250 (l) × 150 (d) mm) with perforatedbottoms. The soil added to the containers had approximately23 000 mg fuel kg-1 soil (unpublished data). There weresubstantial evaporative losses in the first 3 weeks with thefuel concentration measured as 14 300 mg fuel kg-1 soil after3 weeks. The containers were variously amended and reburiedadjacent to the contaminated site so that environmentalconditions were similar to the main plume. Soil temperaturesat the site have been measured as between -4 and 23 °C (7).Two treatments are considered here: a fertilization trial inwhich controlled release fertilizer pellets were added to thesoil to yield soil concentrations of 696 mg NO3
- kg-1, 883 mgNH4 kg-1, 366 mg PO4
-2 kg-1, 43 mg SO4-2 kg-1, and 35 mg
Ca2+ kg-1 on a dry mass basis; and a control treatment thatreceived no additional nutrients. The containers weresampled at the top (0-35 mm), middle (35-70 mm), andbottom (70-100 mm) after 60 months of incubation, andsamples were stored at -18 °C in glass containers.
Sub-samples of soil (10 g) were spiked with an internalstandard (1000 mg L-1 cyclo-octane and 1-bromoeicosaneand 100 mg L-1 d8-naphthalene and p-terphenyl). Water (10mL) and hexane (10 mL) were added, and the samples weretumbled overnight. Analysis of the hexane layer was under-taken on an Agilent 6890 GC fitted with a split/splitlessinjector (280 °C) and a flame ionization detector (FID) (300°C). Separation was performed on a 50 m × 0.22 mm i.d. ×0.25 µm film BP-1 column with helium as the carrier gas. TheGC oven program was 40 °C for 5 min, and then ramped to
310 °C (at 10 °C min-1) and held for 15 min. The instrumentwas cross calibrated with an in-house SAB standard. Thequantity of fuel was approximated by the sum of resolvedpeaks and the undefined complex mixture between C9 andC28 and normalized to the amount of internal standardrecovered.
DNA Extraction. Total microbial DNA was extracted usinga modified mini-beadbeater method (34, 35). Briefly, 0.5 gof soil, 0.5 g of zirconia/silica beads (0.1 mm diameter), 0.5mL of CTAB buffer (hexadecyltrimethylammonium bromide5wt %/vol, 0.35 M NaCl, 120 mM potassium phosphate buffer,pH 8.0), and 0.5 mL of a phenol-chloroform-isoamyl alcoholmixture (25:24:1) were added to a beadbeater tube. This washomogenized twice in a Bio101 beadbeater (three rounds of30 s each at 5000 rpm), and the supernatants were pooled.The pooled supernatant was extracted with an equal volumeof chloroform-isoamyl alcohol (24:1). The nucleic acids wereprecipitated with 2 volumes of 30% (wt/vol) poly(ethyleneglycol) 6000 in 1.6 M NaCl for 2 h at room temperature. Thetubes were centrifuged at 18 000g at 4 °C for 10 min. Thepellet was washed in cold 70% (vol/vol) ethanol and driedunder vacuum prior to resuspension in RNase free water.
Quantitative PCR. Gene abundance was quantified usingreal-time PCR with the primer pairs nosZF and nosZR (25),nirS1F and nirS1R (36), and bzAQ4F and bzAQ4R (14). ThenosZ thermal cycling program consisted of an initial hotstartat 97 °C for 15 min, followed by 45 cycles of 94 °C for 20 s,annealing for 40 s, 72 °C for 40 s, read at 77 °C for 45 s. Theannealing temperature was 54 °C for quantification of thenosZ gene, 53 °C for nirS, and 50 °C for bzdQ. Dissociationcurves were performed every run to confirm reactionspecificity. Quantitative PCR was carried out in an ABI-3000real-time PCR machine using the QuantiTect SYBR GreenPCR MasterMix (Qiagen; Hilden, Germany). The 25 µLreactions contained 2 µL of template DNA and a final primerconcentration of 0.5 µM each.
Standard curves were generated using 10-fold serialdilutions of genomic DNA obtained from the reference strainP. stutzeri for nosZ and nirS. Standards for bzdQ were purifiedPCR products obtained from soil extracts amplified withbzAQ4F and bzAQ4R. DNA concentrations were measuredspectrophotometrically at 260 nm using an Ultraspec 2000UV/visible spectrophotometer (Pharmacia Biotech, Cam-bridge, UK). Copy numbers were calculated on the basis ofthe size of the bzdQ product or assuming that a P. stutzerigenome is 6 Mbp (37) and that each genome contained onecopy of both the nosZ and the nirS genes. The standard curveswere linear over 5 orders of magnitude, and the r2 value wasalways over 0.9. Amplification efficiency for different soilswas calculated by adding increasing concentrations ofstandards to a known background. The amplification ef-ficiency averaged 70%, and percent deviation of duplicates(based on CT values) was 2.1% (n ) 32). Results are expressedas copy number per microgram of DNA extracted per gramof soil.
Denaturing Gradient Gel Electrophoresis Analysis. Forcommunity composition assessments, the nosZF primer hada GC-clamp added (nosZFC): GGCGGCGCGCCGCCCGC-CCCGCCCCCGTCGCCCCG(C/T)TGTTC(A/C)TCGACAGC-CAG. PCR conditions were 95 °C for 5 min, followed by 30cycles of 94 °C for 40 s, 55 °C for 40 s, 72 °C for 1 min, followedby 72 °C for 10 min. Primers were used at 0.5 µM concentra-tion with the Taq MasterMix kit used as suggested by themanufacturer (Qiagen). Denaturing gradient gel electro-phoresis (DGGE) based on the protocol described by Muyzeret al. (38) was performed using the Bio-Rad DCode system(Bio-Rad Laboratories, Hercules). PCR products (5 µL) wereloaded onto 8% (w/v) polyacrylamide gradient gels (37:1acrylamide/bis-acrylamide; 1.5 mm thick) with a 30-70%denaturing gradient (100% denaturing acrylamide contains
2012 9 ENVIRONMENTAL SCIENCE & TECHNOLOGY / VOL. 40, NO. 6, 2006
7 M urea and 40% (v/v) formamide). A 3 mL stacking gel (0%denaturant) was added to the top of the gel. Gels were runfor 15 min at 80 V prior to loading samples and then for 16h at 80 V in 1× TAE buffer (40 mM Tris, 20 mM acetic acid,and 1 mM EDTA) at 60 °C. Gels were stained with SYBRGreenI (10 000 × solution in DMSO; Sigma-Aldrich Chemie GmbH,Steinheim, Germany) diluted in 1 × TAE and destained indeionized water prior to ultraviolet (UV) transillumination.Gels were photographed with a Nikon CoolPix 4500 cameraequipped with a SYBRGreen filter.
Laboratory Microcosm Experiment. A microcosm ex-periment was set up to demonstrate that hydrocarbondegradation under anaerobic conditions was promoted byfertilization. Four different soils (control top, control bottom,fertilized top, and fertilized bottom) from the field trial wereused to establish the microcosms. Toluene (58 µL) andhexadecane (65 µL) were added to 5 g of sterile sand (OttawaSand Standard S2303, Fisher Scientific) and thoroughly mixed.Three grams of this was mixed with 27 g of sterile sand tocreate a spiked sand containing approximately 12 mg kg-1
of toluene and 300 mg kg-1 hexadecane. Spiked sand (4 g)was added to 36 g of control or fertilized bottom soil, and 5g of spiked sand was added to 45 g of control or fertilized topsoils. Next, 5 g of these soils was added to each of eight 50mL tubes for each soil type. This amount of soil gave aheadspace:soil ratio of approximately 10:1. Remaining spikedsand was frozen at -20 °C prior to hydrocarbon analysis.Four tubes of each of the four soil types were sealed andincubated aerobically. Another four tubes of each soil typewere flushed with a mixture of H2 (5%), CO2 (5%), and N2
(90%) prior to sealing. All were incubated at 10 °C for 30days.
After 30 days incubation, hydrocarbons were extractedfrom the microcosms following the Canadian CouncilMinisters of the Environment guidelines (39). Briefly, the F1fraction, containing the toluene, was extracted by adding a2:1 ratio of methanol to soil and shaking for 60 min. Thetubes were centrifuged, and the methanol extract was directlyinjected on-column. The F2 and F3 fractions, containing thehexadecane, were extracted by accelerated solvent extraction(Soxtex Avanti model 2050, Hoganas, Sweden) using a 1:1mixture of hexane and acetone. Extracts were analyzed witha Varian model CP-3800 GC equipped with a FID and a WCOTfused silica capillary column (CP-Sil 5 CB; 100% dimethylpolysiloxane; 15 m × 0.53 mm × 0.15 µm; Varian Canada;Mississauga, ON). This chromatographic data were processedusing Varian’s Star Workstation software (ver. 6.2).
Isolation of Hydrocarbon Degrading Denitrifiers. Soil(1 g) from the bottom samples was added to tubes containing5 g of 5 mm sterile glass beads and 10 mL of sterile phosphatebuffered saline and vortexed at high speed for 60 s. Aliquotswere spread plated onto YTSN plates (per liter: 0.25 g ofyeast extract, 0.25 g of tryptone, 0.25 g of starch, 0.5 g ofNaNO3, and 15 g of agar) (40). The plates were incubated at10 °C in anaerobic GasPaks (BBL). After 3 weeks, colonieswere picked from the plates and sub-cultured until pure.Isolates were tested for their ability to utilize either hexa-decane or toluene as a sole carbon source. Cultures (20 µL)grown in YTSN were inoculated into 180 µL of mineralmedium (per liter: 1 g of K2HPO4, 0.2 g of MgSO4‚7H2O, 0.01g of FeSO4‚7H2O, 0.02 g of CaCl2, 0.02 g of MnCl2‚4H2O, 0.5g of NaNO3) in triplicate in 96-well titer trays. Controlsconsisting of inoculated wells with no added carbon sourceand uninoculated medium were included. Five microlitersof either hexadecane or toluene was added, and the plateswere incubated at 10 °C. Growth was measured at 650 nm.
Statistical Analysis. All data were tested for normalityusing the Anderson-Darling normality test and for homo-geneity of variances by Bartlett and Levene’s test. Geneabundance results were log transformed and assessed by
analysis of variance, general linear model (ANOVA-GLM).The field experiment (n ) 4) was analyzed using the treatment(control or fertilized) and the depth (top, middle, bottom) asfactors. The microcosm experiment (n ) 4) was analyzedusing treatment (control or fertilized), depth (top or bottom),and incubation (aerobic or anaerobic) as factors. For non-normal data with unequal variances, the Kruskal Wallis testwas used to assess treatment effects when there were onlytwo treatments, and the Mood’s median test was used withmultiple treatments (41). DGGE patterns were processed andclustered using Bionumerics software (Applied Maths, Ko-rtrijk, Belgium). For principal component analysis (PCA), allbands were divided into common band classes. PCA scoreswere analyzed by ANOVA-GLM to test for treatment effects(42). PCA scores were non-normally distributed but hadhomogeneous variances between treatments.
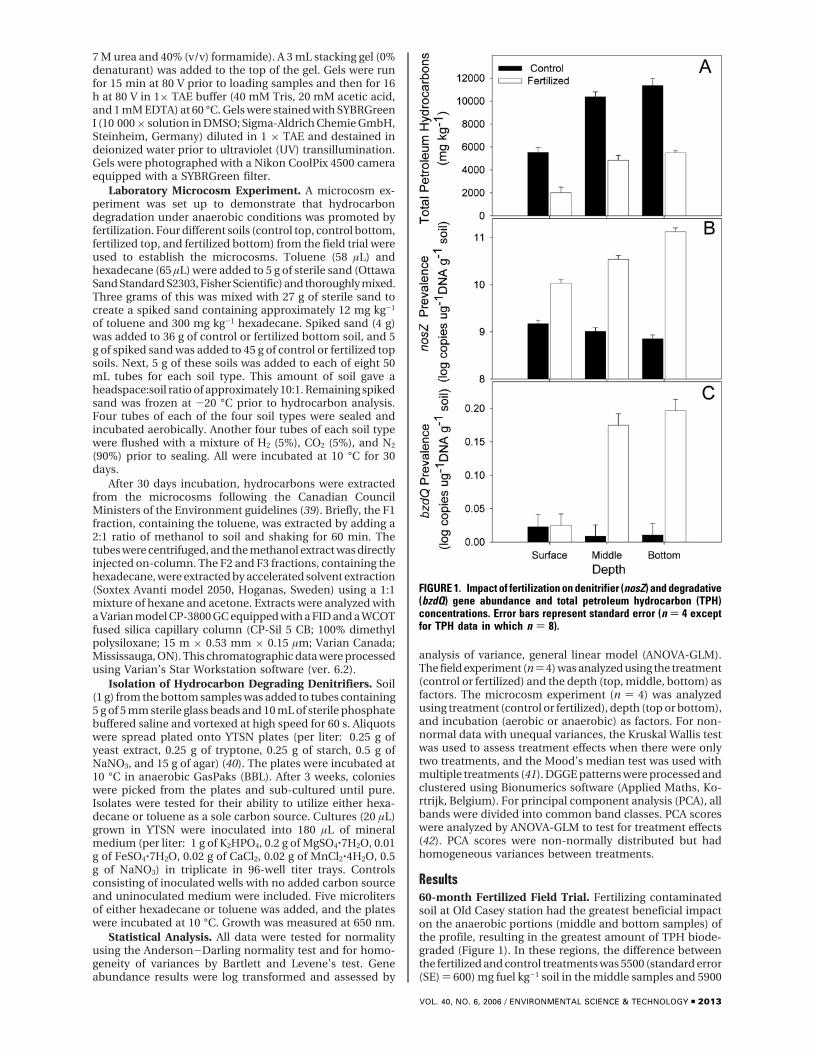
Results60-month Fertilized Field Trial. Fertilizing contaminatedsoil at Old Casey station had the greatest beneficial impacton the anaerobic portions (middle and bottom samples) ofthe profile, resulting in the greatest amount of TPH biode-graded (Figure 1). In these regions, the difference betweenthe fertilized and control treatments was 5500 (standard error(SE) ) 600) mg fuel kg-1 soil in the middle samples and 5900
FIGURE 1. Impact of fertilization on denitrifier (nosZ) and degradative(bzdQ) gene abundance and total petroleum hydrocarbon (TPH)concentrations. Error bars represent standard error (n ) 4 exceptfor TPH data in which n ) 8).
VOL. 40, NO. 6, 2006 / ENVIRONMENTAL SCIENCE & TECHNOLOGY 9 2013
(SE ) 670) mg fuel kg-1 soil in the bottom samples. In contrast,this difference was only 3500 (SE ) 630) mg fuel kg-1 soil forthe top samples. A direct comparison of biodegradation ratesbetween top and middle or bottom depths is confounded byevaporation, which is limited to the top (43).
Fertilization substantially increased nosZ copy numberin the anaerobic portions of the soil profile (Figure 1).Fertilization increased (P < 0.001; ANOVA-GLM) nosZ copynumbers by 10-fold in the top samples (from 1.2 × 109 to 2.3× 1010) and by 1000-fold in the bottom samples (from 9.8 ×108 to 1.6 × 1011). This increase was also observed for benzoyl-CoA gene copy numbers, fertilization again having thegreatest impact for anaerobic sections of the soil (P < 0.002;ANOVA-GLM). Fertilization increased bzdQ copy numbers10-fold in the bottom samples but actually decreased bzdQabundance in the top samples (Figure 1).
The changes in degradation induced by fertilizationcorresponded to significant changes in denitrifier communitycomposition. The ratio of the number of copies of nirS µg-1
DNA g-1 to the number of copies of nosZ µg-1 DNA g-1
decreased substantially (P < 0.001 ANOVA-GLM) underfertilization from 0.32 to 0.08. There was, however, no depth-specific effect (P < 0.697) with an average ratio of 0.07 in thebottom samples as compared to a ratio of 0.1 in the topsamples. Although the number of copies of nirS µg-1 DNAg-1 was greater in the fertilized soils, the ratio decreased,which indicates that there is a change in the structure of thedenitrifying community, perhaps due to the preferentialstimulation of the nirK containing organisms.
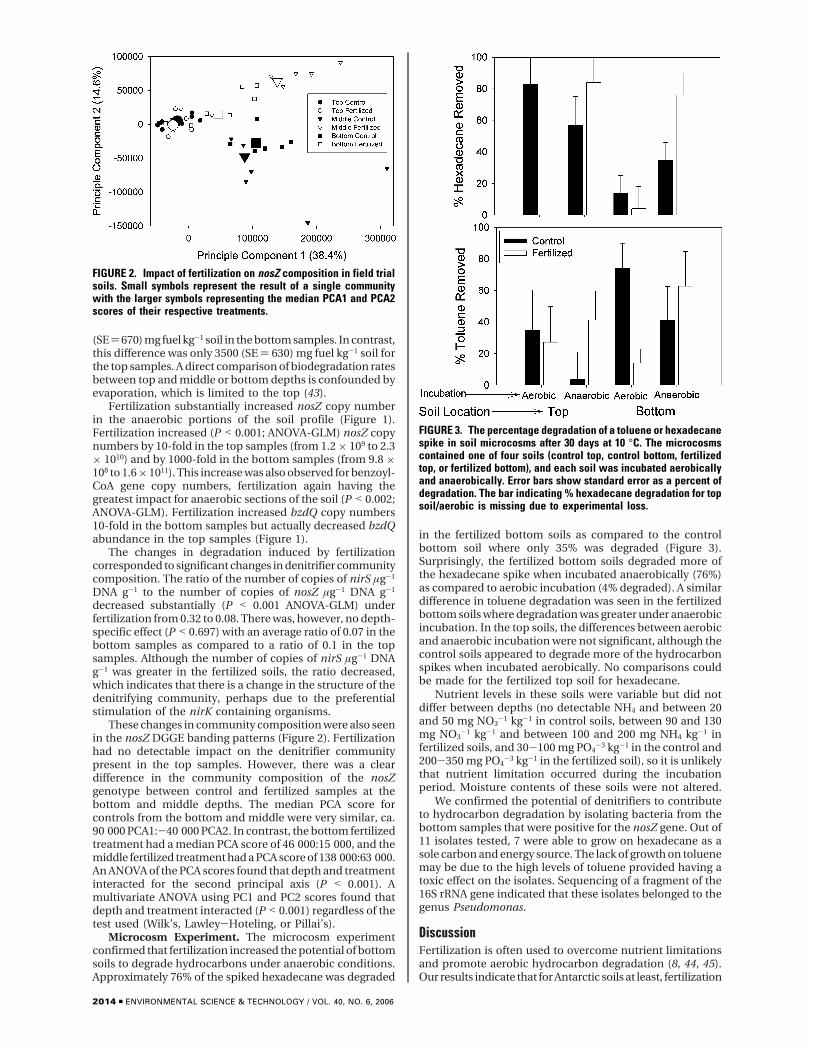
These changes in community composition were also seenin the nosZ DGGE banding patterns (Figure 2). Fertilizationhad no detectable impact on the denitrifier communitypresent in the top samples. However, there was a cleardifference in the community composition of the nosZgenotype between control and fertilized samples at thebottom and middle depths. The median PCA score forcontrols from the bottom and middle were very similar, ca.90 000 PCA1:-40 000 PCA2. In contrast, the bottom fertilizedtreatment had a median PCA score of 46 000:15 000, and themiddle fertilized treatment had a PCA score of 138 000:63 000.An ANOVA of the PCA scores found that depth and treatmentinteracted for the second principal axis (P < 0.001). Amultivariate ANOVA using PC1 and PC2 scores found thatdepth and treatment interacted (P < 0.001) regardless of thetest used (Wilk’s, Lawley-Hoteling, or Pillai’s).
Microcosm Experiment. The microcosm experimentconfirmed that fertilization increased the potential of bottomsoils to degrade hydrocarbons under anaerobic conditions.Approximately 76% of the spiked hexadecane was degraded
in the fertilized bottom soils as compared to the controlbottom soil where only 35% was degraded (Figure 3).Surprisingly, the fertilized bottom soils degraded more ofthe hexadecane spike when incubated anaerobically (76%)as compared to aerobic incubation (4% degraded). A similardifference in toluene degradation was seen in the fertilizedbottom soils where degradation was greater under anaerobicincubation. In the top soils, the differences between aerobicand anaerobic incubation were not significant, although thecontrol soils appeared to degrade more of the hydrocarbonspikes when incubated aerobically. No comparisons couldbe made for the fertilized top soil for hexadecane.
Nutrient levels in these soils were variable but did notdiffer between depths (no detectable NH4 and between 20and 50 mg NO3
-1 kg-1 in control soils, between 90 and 130mg NO3
-1 kg-1 and between 100 and 200 mg NH4 kg-1 infertilized soils, and 30-100 mg PO4
-3 kg-1 in the control and200-350 mg PO4
-3 kg-1 in the fertilized soil), so it is unlikelythat nutrient limitation occurred during the incubationperiod. Moisture contents of these soils were not altered.
We confirmed the potential of denitrifiers to contributeto hydrocarbon degradation by isolating bacteria from thebottom samples that were positive for the nosZ gene. Out of11 isolates tested, 7 were able to grow on hexadecane as asole carbon and energy source. The lack of growth on toluenemay be due to the high levels of toluene provided having atoxic effect on the isolates. Sequencing of a fragment of the16S rRNA gene indicated that these isolates belonged to thegenus Pseudomonas.
DiscussionFertilization is often used to overcome nutrient limitationsand promote aerobic hydrocarbon degradation (8, 44, 45).Our results indicate that for Antarctic soils at least, fertilization
FIGURE 2. Impact of fertilization on nosZ composition in field trialsoils. Small symbols represent the result of a single communitywith the larger symbols representing the median PCA1 and PCA2scores of their respective treatments.
FIGURE 3. The percentage degradation of a toluene or hexadecanespike in soil microcosms after 30 days at 10 °C. The microcosmscontained one of four soils (control top, control bottom, fertilizedtop, or fertilized bottom), and each soil was incubated aerobicallyand anaerobically. Error bars show standard error as a percent ofdegradation. The bar indicating % hexadecane degradation for topsoil/aerobic is missing due to experimental loss.
2014 9 ENVIRONMENTAL SCIENCE & TECHNOLOGY / VOL. 40, NO. 6, 2006

also stimulates denitrifying bacteria to degrade hydrocarbonsin the anaerobic portion of the soil profile. Further, whenthese communities are subsequently exposed to oxygen,hydrocarbon degradation is inhibited. This stimulatory effectwas not observed in aerobic sections of the soil profile. Similarresults were found in temperate soils where fertilizationincreased denitrification activity in aerobic soils (46), althoughthis activity was not directly linked to hydrocarbon degrada-tion. Although denitrifying bacteria that are capable ofhydrocarbon degradation have been isolated (20, 47, 48), therole of denitrifiers in hydrocarbon degradation in Antarcticahas not been explored previously, largely because Antarcticsoils are assumed to be oxic.
We estimated oxygen concentrations at different depthsin the soil by assuming a steady state of oxygen in the soilecosystem with the only O2 input being from gas diffusionand the only sink being microbial respiration. We assumedno sources or outputs of O2 in the soil ecosystem. Microbialrespiration rates of 350 µg O2 g-1 h-1 were taken from Bolter(9). Measured bulk density (1.224 g cm-3), moisture contents(7.4% w/w top, 14% middle, and 18% bottom) and theMillington-Quirk equation were used to convert the bulkair O2 diffusion coefficient of 0.178 cm2 s-1 (49) to the soilair O2 diffusion coefficient.
where Dsoil is the bulk soil diffusion coefficient, Dair is the airdiffusion coefficient of 0.178 cm2 s-1, fair is the air filledporosity, and ftotal is the total porosity. We estimated oxygeninput due to diffusion (Ndiffusion) to the soil system using eq2.
where ∆O2 is the difference between atmospheric, assumedto be 300 mg L-1 (49), and soil O2 concentrations, which weassumed was always at a maximum. We used depths of 17.5,52.5, and 85 mm corresponding to top, middle, and bottomsamples. The soil sink was calculated by converting microbialrespiration rates to a volumetric basis and multiplying theserates by the depth of the soil profile. Where soil oxygenconcentrations were negative, we assumed the soil haddepleted all available oxygen (49).
Despite water-filled pore spaces of 8%, 16%, and 22% forthe top, middle, and bottom, after 60 months O2 concentra-tions were calculated to be less than zero in the bottomsamples. These calculations indicate that the soils at Caseystation are very likely micro-oxic or anaerobic with depth.The air diffusion coefficients as calculated by the Millington-Quirk formula were 0.0668, 0.0519, and 0.0446 cm2 s-1 forthe top, middle, and bottom, respectively. Some recent worksuggests that the Marshall equation (Dsoil ) fair
2.5/ftotalDair)may be more appropriate for soil that has been sieved andrepacked (12). This equation results in slightly differentdiffusion coefficients (0.0626, 0.0519, and 0.046 cm2 s-1 forthe top, middle, and bottom). However, given the uncertaintyassociated with the microbial respiration rates, this is notsignificant.
While our data cannot conclusively demonstrate in situdenitrification, several lines of evidence suggest that thebottom of the containers were actively denitrifying. First,oxygen diffusion and consumption calculations indicate thatthese soils were anoxic or, at best, micro-oxic. Second, therewas readily available nitrate in the bottom samples. Third,there were significant increases in nosZ prevalence in thebottom samples, and finally we isolated several strains ofbacteria able to utilize hexadecane. We do not have directredox measurements of the soil so our results do not exclude
the possibility that iron-reducing bacteria are also active.However, the high concentrations of nitrate (109 mg kg-1)would provide a selective advantage for denitrifiers. Similarlines of evidence have been used by other investigators (50)to demonstrate which microbial groups are involved incontaminant degradation.
Increased hydrocarbon degradation observed in fertilizedbottom samples as compared to control bottom sampleswas mirrored by an increase in the prevalence of nosZ andbzdQ genes (Figure 1). The bzdQ primers can amplify othergenes, and thus it is possible that the bzdQ gene copiesnumbers are overestimates. At the very least, the differencesin the numbers of bzdQ detected represent a change in themicrobial community structure. Unfortunately, gene targetsdo not currently exist for alkane degradation by denitrifiers,so we were unable to link increased alkane degradation toan increase in an alkane catabolic gene specific for denitrifiers.As expected, the composition of the denitrifier communityalso changed with a greater difference observed betweenfertilized and control samples from the bottom samples thanbetween fertilized and control samples from the top soils(Figure 2). This implies that fertilization had the greatestimpact on the denitrifier community in the anaerobic regionof the soil. This depth-specific effect was expected becauseof the depletion of the dominant electron acceptor (oxygen),in the soil profile. Supporting our argument that denitrifiersare responsible for fuel degradation, the microcosm experi-ment demonstrated that fertilized soils from the anoxic(bottom) region of the soils lost more of a hexadecane andtoluene spike when incubated anaerobically than whenincubated aerobically (Figure 3). Presumably, the microbialcommunity had adapted to hydrocarbon degradation underanaerobic conditions and was unable to change to aerobicdegradation in 30 days. The implication for managementstrategies is that aerating anaerobic soil may reduce hydro-carbon biodegradation rather than increase it, at least in theshort-term. Further work with 14C-labeled hydrocarbons isrequired to determine the ultimate fate of hydrocarbons inthe soil under anaerobic conditions.
The results presented here demonstrate that fertilizationhas an unexpected benefit in Antarctic soil bioremediationsystems, the disproportional stimulation of the anaerobicportion of the soil profile. For denitrifiers, fertilization is notonly providing nutrients but also electron acceptors. How-ever, as noted by Roy and Greer (46), denitrifiers may bestimulated in the aerobic portion of a soil profile and notdegrade hydrocarbons but compete with aerobic hydrocar-bon degraders for scarce nitrogen. Our work does not addresswhether denitrifiers are a net benefit in a mixed aerobic/anaerobic system. Promisingly, recent work suggests thatoxygen, not temperature, limits hydrocarbon degradation inpolar soils (49) and that hydrocarbon degradation at lowtemperatures limits residual toxicity (51). Our data suggestthat management plans need to incorporate the role ofdenitrifiers in hydrocarbon degradation to optimize biore-mediation strategies.
This is the only report of denitrifiers in Antarctic soils andone of the few reports of the role of denitrifiers in hydrocarbondegradation in polar soils. The results presented here alsoindicate that aerating anaerobic soil would work at cross-purposes to fertilization. The implications that denitrifiersare responsible for hydrocarbon degradation are very positivefor polar remediation technologies. Other investigators,working on Arctic soils, found that under nitrate-reducingconditions hydrocarbon degradation was not as sensitive todecreases in temperature as aerobic hydrocarbon degrada-tion (52). This implies that management techniques thatpromote denitrifier-based hydrocarbon degradation may notrequire heating to be effective. For these reasons, the role ofdenitrifying bacteria in the biodegradation of hydrocarbons
Dsoil ) Dair × (fair/ftotal)2 × fair
4/3 (1)
Ndiffusion ) (soil air O2 diffusion coefficient × cm2)/(depth × ∆O2) (2)
VOL. 40, NO. 6, 2006 / ENVIRONMENTAL SCIENCE & TECHNOLOGY 9 2015

in cold terrestrial environments shows great promise as aneffective remediation tool and warrants further investigation.
AcknowledgmentsWe wish to thank P. McA Harvey (AAD) for carrying out thehydrocarbon analysis of the field trial. This work wassupported by Australian Antarctic Science grant 1163.
Literature Cited(1) Aislabie, J. M.; Balks, M. R.; Foght, J. M.; Waterhouse, E. J.
Hydrocarbon spills on Antarctic soils: Effects and management.Environ. Sci. Technol. 2004, 38, 1265-1274.
(2) Ferguson, S. H.; Woinarski, A. Z.; Snape, I.; Morris, C. E.; Revill,A. T. A field trial of in situ chemical oxidation to remediatelong-terin diesel contaminated Antarctic soil. Cold Reg. Sci.Technol. 2004, 40, 47-60.
(3) Aislabie, J.; Foght, J.; Saul, D. Aromatic hydrocarbon-degradingbacteria from soil near Scott Base, Antarctica. Polar Biol. 2000,23, 183-188.
(4) Eckford, R.; Cook, F. D.; Saul, D.; Aislabie, J.; Foght, J. Free-living heterotrophic nitrogen-fixing bacteria isolated from fuel-contaminated antarctic soils. Appl. Environ. Microbiol. 2002,68, 5181-5185.
(5) Luz, A. P.; Pellizari, V. H.; Whyte, L. G.; Greer, C. W. A surveyof indigenous microbial hydrocarbon degradation genes in soilsfrom Antarctica and Brazil. Can. J. Microbiol. 2004, 50, 323-333.
(6) Eriksson, M.; Ka, J. O.; Mohn, W. W. Effects of low temperatureand freeze-thaw cycles on hydrocarbon biodegradation in Arctictundra soil. Appl. Environ. Microbiol. 2001, 67, 5107-5112.
(7) Ferguson, S. H.; Franzmann, P. D.; Snape, I.; Revill, A. T.; Trefry,M. G.; Zappia, L. R. Effects of temperature on mineralisation ofpetroleum in contaminated Antarctic terrestrial sediments.Chemosphere 2003, 52, 975-987.
(8) Ferguson, S. H.; Franzmann, P. D.; Revill, A. T.; Snape, I.; Rayner,J. L. The effects of nitrogen and water on mineralisation ofhydrocarbons in diesel-contaminated terrestrial Antarctic. ColdReg. Sci. Technol. 2003, 37, 197-212.
(9) Bolter, M. Environmental conditions and microbiologicalproperties from soils and lichens from Antarctica (Wilkes Land).Polar Biol. 1992, 11, 591-599.
(10) Beyer, L.; Bolter, M. Formation, ecology, and geography ofcryosols of an ice-free oasis in coastal East Antarctica near CaseyStation (Wilkes Land). Aust. J. Soil Res. 1998, 37, 209-244.
(11) Beyer, L.; Bockheim, J. G.; Campbell, I. B.; Claridge, G. G. C.Genesis, properties and sensitivity of Antarctic Gelisols. Antarct.Sci. 1999, 11, 387-398.
(12) Moldrup, P.; Olesen, T.; Gamst, J.; Schjonning, P.; Yamaguchi,T.; Rolston, D. E. Predicting the gas diffusion coefficient inrepacked soil: Water-induced linear reduction model. Soil Sci.Soc. Am. J. 2000, 64, 1588-1594.
(13) Hoejberg, O.; Revsbech, N. P.; Tiedje, J. Denitrification in soilaggregates analyzed with microsensors for nitrous oxide andoxygen. Soil Sci. Soc. Am. J. 1994, 58, 1691-1698.
(14) Song, B.; Ward, B. B. Genetic diversity of benzoyl coenzyme Areductase genes detected in denitrifying isolates and estuarinesediment communities. Appl. Environ. Microbiol. 2005, 71,2036-2045.
(15) Heider, J.; Spormann, A. M.; Beller, H. R.; Widdel, F. Anaerobicbacterial metabolism of hydrocarbons. FEMS Microbiol. Rev.1999, 22, 459-473.
(16) Ehrenreich, P.; Behrends, A.; Harder, J.; Widdel, F. Anaerobicoxidation of alkanes by newly isolated denitrifying bacteria.Arch. Microbiol. 2000, 173, 58-64.
(17) Rabus, R.; Wilkes, H.; Behrends, A.; Armstroff, A.; Fischer, T.;Pierik, A. J.; Widdel, F. Anaerobic initial reaction of n-alkanesin a denitrifying bacterium: Evidence for (1-methylpentyl)-succinate as initial product and for involvement of an organicradical in n-hexane metabolism. J. Bacteriol. 2001, 183, 1707-1715.
(18) Spormann, A. M.; Widdel, F. Metabolism of alkylbenzenes,alkanes, and other hydrocarbons in anaerobic bacteria. Bio-degradation 2000, 11, 85-105.
(19) Harwood, C. S.; Burchhardt, G.; Herrmann, H.; Fuchs, G.Anaerobic metabolism of aromatic compounds via the benzoyl-CoA pathway. FEMS Microbiol. Rev. 1999, 22, 439-458.
(20) Hosoda, A.; Kasai, Y.; Hamamura, N.; Takahata, Y.; Watanabe,K. Development of a PCR method for the detection andquantification of benzoyl-CoA reductase genes and its applica-
tion to monitored natural attenuation. Biodegradation 2005,16, 591-601.
(21) Zumft, W. G. Cell biology and molecular basis of denitrification.Microbiol. Mol. Biol. Rev. 1997, 61, 533-616.
(22) Song, B.; Palleroni, N. J.; Haggblom, M. M. Isolation andcharacterization of diverse halobenzoate-degrading denitrifyingbacteria from soils and sediments. Appl. Environ. Microbiol.2000, 66, 3446-3453.
(23) Heider, J.; Spormann, A. M.; Beller, H. R.; Widdel, F. Anaerobicbacterial metabolism of hydrocarbons. FEMS Microbiol. Rev.1998, 22, 459-473.
(24) Mesa, S.; Alche, J. D.; Bedmar, E. J.; Delgado, M. J. Expressionof nir, nor and nos denitrification genes from Bradyrhizobiumjaponicum in soybean root nodules. Physiol. Plant. 2004, 120,205-211.
(25) Kloos, K.; Mergel, A.; Rosch, C.; Bothe, H. Denitrification withinthe genus Azospirillum and other associative bacteria. Aust. J.Plant Physiol. 2001, 28, 991-998.
(26) Kirk, G. The Biogeochemistry of Submerged Soils; John Wiley &Sons, Ltd.: Hoboken, NJ, 2004.
(27) Gooseff, M. N.; McKnight, D. M.; Runkel, R. L.; Duff, J. H.Denitrification and hydrologic transient storage in a glacialmeltwater stream, McMurdo Dry Valleys, Antarctica. Limnol.Oceanogr. 2004, 49, 1884-1895.
(28) Lee, P. A.; Mikucki, J. A.; Foreman, C. M.; Priscu, J. C.; DiTullio,G. R.; Riseman, S. F.; de Mora, S. J.; Wolf, C. F.; Kester, L.Thermodynamic constraints on microbially mediated processesin lakes of the McMurdo Dry Valleys, Antarctica. Geomicrobiol.J. 2004, 21, 221-237.
(29) Ward, B. B.; Priscu, J. C. Detection and characterization ofdenitrifying bacteria from a permanently ice-covered Antarcticlake. Hydrobiologia 1997, 347, 57-68.
(30) Mountfort, D. O.; Kaspar, H. F.; Downes, M.; Asher, R. A.Partitioning effects during terminal carbon and electron flowin sediments of a low-salinity meltwater pond near BratinaIsland, McMurdo Ice Shelf, Antarctica. Appl. Environ. Microbiol.1999, 65, 5493-5499.
(31) Chapin, D. M. Nitrogen mineralization, nitrification, anddenitrification in a high arctic lowland ecosystem, Devon Island,NWT, Canada. Arct. Alp. Res. 1996, 28, 85-92.
(32) Christensen, T. R. Potential and actual trace gas fluxes in Arcticterrestrial ecosystems. Polar Res. 1999, 18, 199-206.
(33) Snape, I.; Riddle, M. J.; Stark, J. S.; Cole, C. M.; King, C. K.;Duquesne, S.; Gore, D. B. Management and remediation ofcontaminated sites at Casey Station, Antarctica. Polar Rec. 2001,37, 199-214.
(34) Kowalchuk, G. A.; Bodelier, P. L. E.; Heilig, G. H. J.; Stephen, J.R.; Laanbroek, H. J. Comunity analysis of ammonia-oxidizingbacteria, in relation to oxygen availability in soils and root-oxygenated sediments, using PCR, DGGE and oligonucleotideprobe hybridization. FEMS Microbiol. Ecol. 1998, 27, 339-350.
(35) Burgmann, H.; Pesaro, M.; Widmer, F.; Zeyer, J. A strategy foroptimizing quality and quantity of DNA extracted from soil. J.Microbiol. Methods 2001, 45, 7-20.
(36) Braker, G.; Fesefeldt, A.; Witzel, K. P. Development of PCR primersystems for amplification of nitrite reductase genes (nirK andnirS) to detect denitrifying bacteria in environmental samples.Appl. Environ. Microbiol. 1998, 64, 3769-3775.
(37) Fogel, G. B.; Collins, C. R.; Li, J.; Brunk, C. F. Prokaryotic genomesize and SSU rDNA copy number: estimation of microbialrelative abundance from a mixed population. Microb. Ecol. 1999,38, 93-113.
(38) Muyzer, G.; de Waal, E. C.; Utterlinden, A. G. Profiling of complexmicrobial population by denaturing gradient gel electrophoresisanalysis and polymerase chain reaction-amplified gene codingfor 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695-700.
(39) Lorca, C. University of Saskatchewan, 2005.(40) Siciliano, S. D.; Fortin, N.; Mihoc, A.; Wisse, G.; Labelle, S.;
Beaumier, D.; Ouellette, D.; Roy, R.; Whyte, L. G.; Banks, M. K.;Schwab, P.; Lee, K.; Greer, C. W. Selection of specific endophyticbacterial genotypes by plants in response to soil contamination.Appl. Environ. Microbiol. 2001, 67, 2469-2475.
(41) Sokal, R. R.; Rohlf, F. J. Biometry: The Principles and Practiceof Statistics in Biological Research, 3rd ed.; W. H. Freeman andCo.: New York, 1995.
(42) Glimm, E.; Holger, H.; Engelen, B.; Smalla, K.; Backhaus, H.Statistical comparisons of community catabolic profiles. J.Microbiol. Methods 1997, 30, 71-80.
(43) Kang, S. H.; Oulman, C. S. Evaporation of petroleum productsfrom contaminated soils. J. Environ. Eng. (N.Y.) 1996, 122, 384-387.
2016 9 ENVIRONMENTAL SCIENCE & TECHNOLOGY / VOL. 40, NO. 6, 2006

(44) Cavanagh, J. E.; Nichols, P. D.; Franzmann, P. D.; McMeekin,T. A. Hydrocarbon degradation by Antarctic coastal bacteria.Antarct. Sci. 1998, 10, 386-397.
(45) Aislabie, J.; McLeod, M.; Fraser, R. Potential for biodegradationof hydrocarbons in soil from the Ross Dependency, Antarctica.Appl. Microbiol. Biotechnol. 1998, 49, 210-214.
(46) Roy, R.; Greer, C. W. Hexadecane mineralization and denitri-fication in two diesel-fuel contaminated soils. FEMS Microbiol.Ecol. 2000, 32, 17-23.
(47) Bonin, P. C.; Michotey, V. D.; Mouzdahir, A.; Rontani, J. F.Anaerobic biodegradation of squalene: Using DGGE to monitorthe isolation of denitrifying Bacteria taken from enrichmentcultures. FEMS Microbiol. Ecol. 2002, 42, 37-49.
(48) Gilewicz, M.; Monpert, G.; Acquaviva, M.; Mille, G.; Bertrand,J. C. Anaerobic oxidation of 1-N-heptadecene by a marinedenitrifying bacterium. Appl. Microbiol. Biotechnol. 1991, 36,252-256.
(49) Rike, A. G.; Haugen, K. B.; Engene, B. In situ biodegradation ofhydrocarbons in arctic soil at sub-zero temperatures - fieldmonitoring and theoretical simulation of the microbial activa-
tion temperature at a Spitsbergen contaminated site. Cold Reg.Sci. Technol. 2005, 41, 189-209.
(50) Braddock, J. F.; Mccarthy, K. A. Hydrologic and microbiologicalfactors affecting persistence and migration of petroleumhydrocarbons spilled in a continuous-permafrost region.Environ. Sci. Technol. 1996, 30, 2626-2633.
(51) Coulon, F.; Pelletier, E.; Gourhant, L.; Delille, D. Effects of nutrientand temperature on degradation of petroleum hydrocarbonsin contaminated sub-Antarctic soil. Chemosphere 2005, 58,1439-1448.
(52) Eriksson, M.; Sodersten, E.; Yu, Z.; Dalhammar, G.; Mohn, W.W. Degradation of polycyclic aromatic hydrocarbons at lowtemperature under aerobic and nitrate-reducing conditions inenrichment cultures from northern soils. Appl. Environ.Microbiol. 2003, 69, 275-284.
Received for review September 14, 2005. Revised manuscriptreceived January 20, 2006. Accepted January 23, 2006.
ES051818T
VOL. 40, NO. 6, 2006 / ENVIRONMENTAL SCIENCE & TECHNOLOGY 9 2017





























