Embed Size (px)
Citation preview

www.elsevier.com/locate/yviro
Virology 330 (20
Feline immunodeficiency virus envelope glycoprotein mediates apoptosis
in activated PBMC by a mechanism dependent on gp41 function
Himanshu Garg1, Anjali Joshi, Wayne A. Tompkins*
Immunology Program, College of Veterinary Medicine, North Carolina State University, Raleigh, NC 27606, USA
Received 27 August 2004; returned to author for revision 17 September 2004; accepted 5 October 2004
Abstract
Feline Immunodeficiency Virus (FIV) is a lentivirus that causes immunodeficiency in cats, which parallels HIV-1-induced
immunodeficiency in humans. It has been established that HIV envelope (Env) glycoprotein mediates T cell loss via a mechanism that
requires CXCR4 binding. The Env glycoprotein of FIV, similar to HIV, requires CXCR4 binding for viral entry, as well as inducing
membrane fusion leading to syncytia formation. However, the role of FIV Env in T cell loss and the molecular mechanisms governing this
process have not been elucidated. We studied the role of Env glycoprotein in FIV-mediated T cell apoptosis in an in vitro model. Our studies
demonstrate that membrane-expressed FIV Env induces apoptosis in activated feline peripheral blood mononuclear cells (PBMC) by a
mechanism that requires CXCR4 binding, as the process was inhibited by CXCR4 antagonist AMD3100 in a dose-dependent manner.
Interestingly, studies regarding the role of CD134, the recently identified primary receptor of FIV, suggest that binding to CD134 may not be
important for induction of apoptosis in PBMC. However, inhibiting Env-mediated fusion post CXCR4 binding by FIV gp41-specific fusion
inhibitor also inhibited apoptosis. Under similar conditions, a fusion-defective gp41 mutant was unable to induce apoptosis in activated
PBMC. Our findings are the first report suggesting the potential of FIV Env to mediate apoptosis in bystander cells by a process that is
dependent on gp41 function.
D 2004 Elsevier Inc. All rights reserved.
Keywords: FIV; Envelope; Apoptosis; Fusion; Syncytia; CD134; CXCR4
Introduction
Feline Immunodeficiency Virus (FIV) infection, similar
to HIV-1, is characterized by a progressive and irrever-
sible depletion of CD4+ T lymphocytes leading to
immunodeficiency in cats. Various hypotheses have been
proposed for HIV-mediated T cell loss, including direct
cytopathic effect due to infection (Lenardo et al., 2002),
syncytia formation (Ferri et al., 2000), and autoimmune
killing of T cells (Golding et al., 1989). Several inves-
0042-6822/$ - see front matter D 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.virol.2004.10.007
* Corresponding author. Immunology Program, College of Veterinary
Medicine, North Carolina State University, 4700 Hillsborough Street,
Raleigh, NC 27606. Fax: +1 919 515 4237.
E-mail address: [email protected] (W.A. Tompkins).1 Current address: National Cancer Institute, Frederick, MD 21702,
United States.
tigators have suggested that the loss of T cells in HIV
infection is partly due to apoptosis of uninfected bystander
cells (Alimonti et al., 2003; Famularo et al., 1997;
Gougeon and Montagnier, 1993; Meyaar et al., 1992;
Muro-Cacho et al., 1995). In this regard, Finkel et al.
(1995) showed that apoptosis of lymph node T cells in
HIV-infected individuals primarily occurs in uninfected
bystander cells that lie in close proximity to productively
infected cells. The mechanism(s) behind FIV-mediated T
cell depletion are not clear, although observations mostly
parallel findings in HIV-1.
Several viral proteins including HIV Env (Laurent-
Crawford et al., 1995), Vpr (Desai et al., 2002), and Tat
(Purvis et al., 1995) have been shown to induce apoptosis.
However, determinants of HIV pathogenicity most often
map to the Env glycoprotein (Etemad-Moghadam et al.,
2001), suggesting that it may be a key regulator of CD4+ T
04) 424–436
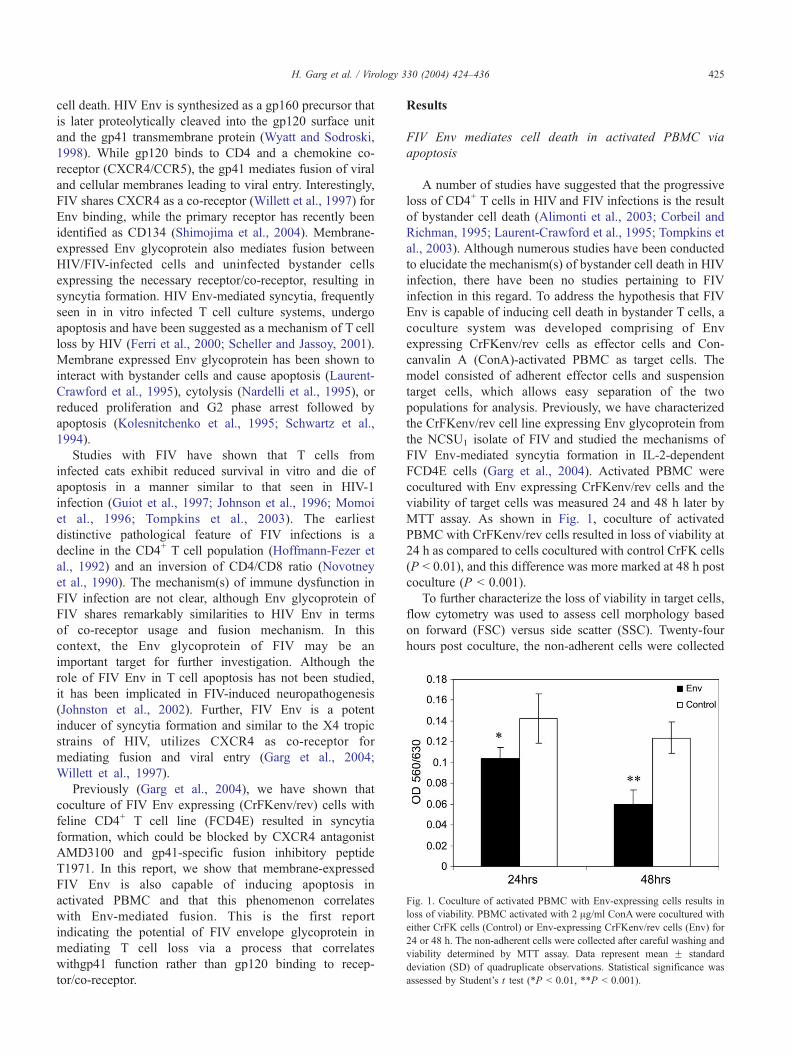
Fig. 1. Coculture of activated PBMC with Env-expressing cells results in
loss of viability. PBMC activated with 2 Ag/ml ConAwere cocultured with
either CrFK cells (Control) or Env-expressing CrFKenv/rev cells (Env) for
24 or 48 h. The non-adherent cells were collected after careful washing and
viability determined by MTT assay. Data represent mean F standard
deviation (SD) of quadruplicate observations. Statistical significance was
assessed by Student’s t test (*P b 0.01, **P b 0.001).
H. Garg et al. / Virology 330 (2004) 424–436 425
cell death. HIV Env is synthesized as a gp160 precursor that
is later proteolytically cleaved into the gp120 surface unit
and the gp41 transmembrane protein (Wyatt and Sodroski,
1998). While gp120 binds to CD4 and a chemokine co-
receptor (CXCR4/CCR5), the gp41 mediates fusion of viral
and cellular membranes leading to viral entry. Interestingly,
FIV shares CXCR4 as a co-receptor (Willett et al., 1997) for
Env binding, while the primary receptor has recently been
identified as CD134 (Shimojima et al., 2004). Membrane-
expressed Env glycoprotein also mediates fusion between
HIV/FIV-infected cells and uninfected bystander cells
expressing the necessary receptor/co-receptor, resulting in
syncytia formation. HIV Env-mediated syncytia, frequently
seen in in vitro infected T cell culture systems, undergo
apoptosis and have been suggested as a mechanism of T cell
loss by HIV (Ferri et al., 2000; Scheller and Jassoy, 2001).
Membrane expressed Env glycoprotein has been shown to
interact with bystander cells and cause apoptosis (Laurent-
Crawford et al., 1995), cytolysis (Nardelli et al., 1995), or
reduced proliferation and G2 phase arrest followed by
apoptosis (Kolesnitchenko et al., 1995; Schwartz et al.,
1994).
Studies with FIV have shown that T cells from
infected cats exhibit reduced survival in vitro and die of
apoptosis in a manner similar to that seen in HIV-1
infection (Guiot et al., 1997; Johnson et al., 1996; Momoi
et al., 1996; Tompkins et al., 2003). The earliest
distinctive pathological feature of FIV infections is a
decline in the CD4+ T cell population (Hoffmann-Fezer et
al., 1992) and an inversion of CD4/CD8 ratio (Novotney
et al., 1990). The mechanism(s) of immune dysfunction in
FIV infection are not clear, although Env glycoprotein of
FIV shares remarkably similarities to HIV Env in terms
of co-receptor usage and fusion mechanism. In this
context, the Env glycoprotein of FIV may be an
important target for further investigation. Although the
role of FIV Env in T cell apoptosis has not been studied,
it has been implicated in FIV-induced neuropathogenesis
(Johnston et al., 2002). Further, FIV Env is a potent
inducer of syncytia formation and similar to the X4 tropic
strains of HIV, utilizes CXCR4 as co-receptor for
mediating fusion and viral entry (Garg et al., 2004;
Willett et al., 1997).
Previously (Garg et al., 2004), we have shown that
coculture of FIV Env expressing (CrFKenv/rev) cells with
feline CD4+ T cell line (FCD4E) resulted in syncytia
formation, which could be blocked by CXCR4 antagonist
AMD3100 and gp41-specific fusion inhibitory peptide
T1971. In this report, we show that membrane-expressed
FIV Env is also capable of inducing apoptosis in
activated PBMC and that this phenomenon correlates
with Env-mediated fusion. This is the first report
indicating the potential of FIV envelope glycoprotein in
mediating T cell loss via a process that correlates
withgp41 function rather than gp120 binding to recep-
tor/co-receptor.
Results
FIV Env mediates cell death in activated PBMC via
apoptosis
A number of studies have suggested that the progressive
loss of CD4+ T cells in HIV and FIV infections is the result
of bystander cell death (Alimonti et al., 2003; Corbeil and
Richman, 1995; Laurent-Crawford et al., 1995; Tompkins et
al., 2003). Although numerous studies have been conducted
to elucidate the mechanism(s) of bystander cell death in HIV
infection, there have been no studies pertaining to FIV
infection in this regard. To address the hypothesis that FIV
Env is capable of inducing cell death in bystander T cells, a
coculture system was developed comprising of Env
expressing CrFKenv/rev cells as effector cells and Con-
canvalin A (ConA)-activated PBMC as target cells. The
model consisted of adherent effector cells and suspension
target cells, which allows easy separation of the two
populations for analysis. Previously, we have characterized
the CrFKenv/rev cell line expressing Env glycoprotein from
the NCSU1 isolate of FIV and studied the mechanisms of
FIV Env-mediated syncytia formation in IL-2-dependent
FCD4E cells (Garg et al., 2004). Activated PBMC were
cocultured with Env expressing CrFKenv/rev cells and the
viability of target cells was measured 24 and 48 h later by
MTT assay. As shown in Fig. 1, coculture of activated
PBMC with CrFKenv/rev cells resulted in loss of viability at
24 h as compared to cells cocultured with control CrFK cells
(P b 0.01), and this difference was more marked at 48 h post
coculture (P b 0.001).
To further characterize the loss of viability in target cells,
flow cytometry was used to assess cell morphology based
on forward (FSC) versus side scatter (SSC). Twenty-four
hours post coculture, the non-adherent cells were collected
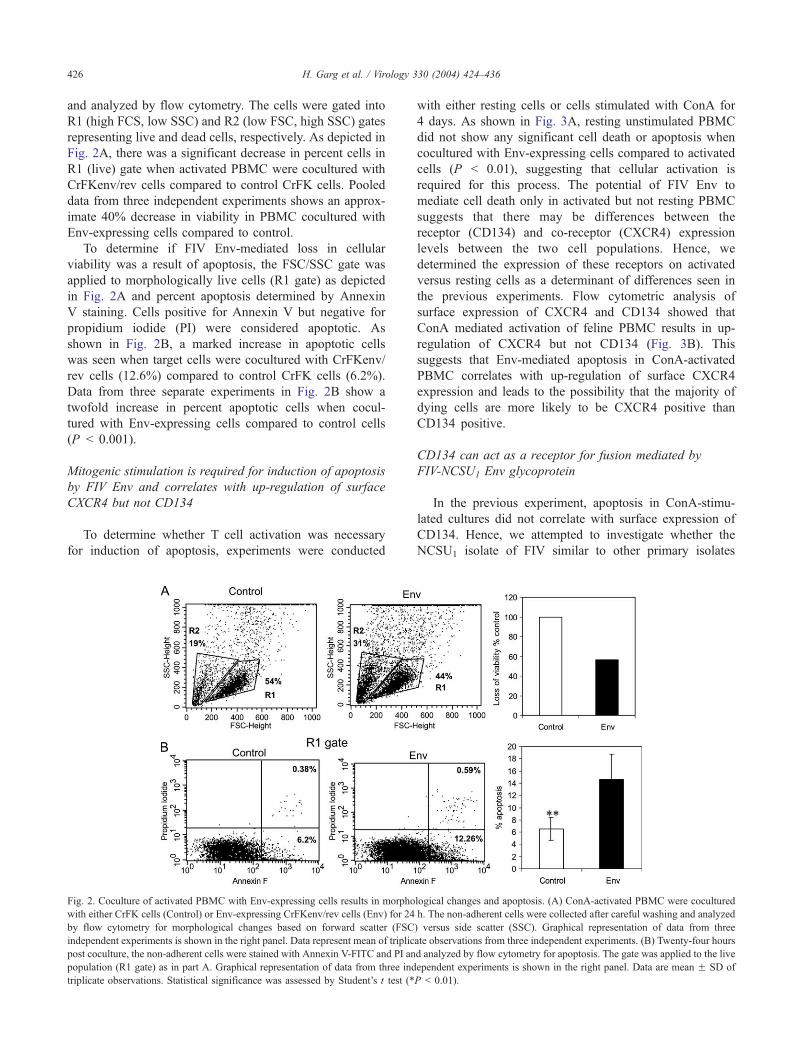
H. Garg et al. / Virology 330 (2004) 424–436426
and analyzed by flow cytometry. The cells were gated into
R1 (high FCS, low SSC) and R2 (low FSC, high SSC) gates
representing live and dead cells, respectively. As depicted in
Fig. 2A, there was a significant decrease in percent cells in
R1 (live) gate when activated PBMC were cocultured with
CrFKenv/rev cells compared to control CrFK cells. Pooled
data from three independent experiments shows an approx-
imate 40% decrease in viability in PBMC cocultured with
Env-expressing cells compared to control.
To determine if FIV Env-mediated loss in cellular
viability was a result of apoptosis, the FSC/SSC gate was
applied to morphologically live cells (R1 gate) as depicted
in Fig. 2A and percent apoptosis determined by Annexin
V staining. Cells positive for Annexin V but negative for
propidium iodide (PI) were considered apoptotic. As
shown in Fig. 2B, a marked increase in apoptotic cells
was seen when target cells were cocultured with CrFKenv/
rev cells (12.6%) compared to control CrFK cells (6.2%).
Data from three separate experiments in Fig. 2B show a
twofold increase in percent apoptotic cells when cocul-
tured with Env-expressing cells compared to control cells
(P b 0.001).
Mitogenic stimulation is required for induction of apoptosis
by FIV Env and correlates with up-regulation of surface
CXCR4 but not CD134
To determine whether T cell activation was necessary
for induction of apoptosis, experiments were conducted
Fig. 2. Coculture of activated PBMC with Env-expressing cells results in morpho
with either CrFK cells (Control) or Env-expressing CrFKenv/rev cells (Env) for 24
by flow cytometry for morphological changes based on forward scatter (FSC
independent experiments is shown in the right panel. Data represent mean of triplic
post coculture, the non-adherent cells were stained with Annexin V-FITC and PI an
population (R1 gate) as in part A. Graphical representation of data from three ind
triplicate observations. Statistical significance was assessed by Student’s t test (*
with either resting cells or cells stimulated with ConA for
4 days. As shown in Fig. 3A, resting unstimulated PBMC
did not show any significant cell death or apoptosis when
cocultured with Env-expressing cells compared to activated
cells (P b 0.01), suggesting that cellular activation is
required for this process. The potential of FIV Env to
mediate cell death only in activated but not resting PBMC
suggests that there may be differences between the
receptor (CD134) and co-receptor (CXCR4) expression
levels between the two cell populations. Hence, we
determined the expression of these receptors on activated
versus resting cells as a determinant of differences seen in
the previous experiments. Flow cytometric analysis of
surface expression of CXCR4 and CD134 showed that
ConA mediated activation of feline PBMC results in up-
regulation of CXCR4 but not CD134 (Fig. 3B). This
suggests that Env-mediated apoptosis in ConA-activated
PBMC correlates with up-regulation of surface CXCR4
expression and leads to the possibility that the majority of
dying cells are more likely to be CXCR4 positive than
CD134 positive.
CD134 can act as a receptor for fusion mediated by
FIV-NCSU1 Env glycoprotein
In the previous experiment, apoptosis in ConA-stimu-
lated cultures did not correlate with surface expression of
CD134. Hence, we attempted to investigate whether the
NCSU1 isolate of FIV similar to other primary isolates
logical changes and apoptosis. (A) ConA-activated PBMC were cocultured
h. The non-adherent cells were collected after careful washing and analyzed
) versus side scatter (SSC). Graphical representation of data from three
ate observations from three independent experiments. (B) Twenty-four hours
d analyzed by flow cytometry for apoptosis. The gate was applied to the live
ependent experiments is shown in the right panel. Data are mean F SD of
P b 0.01).
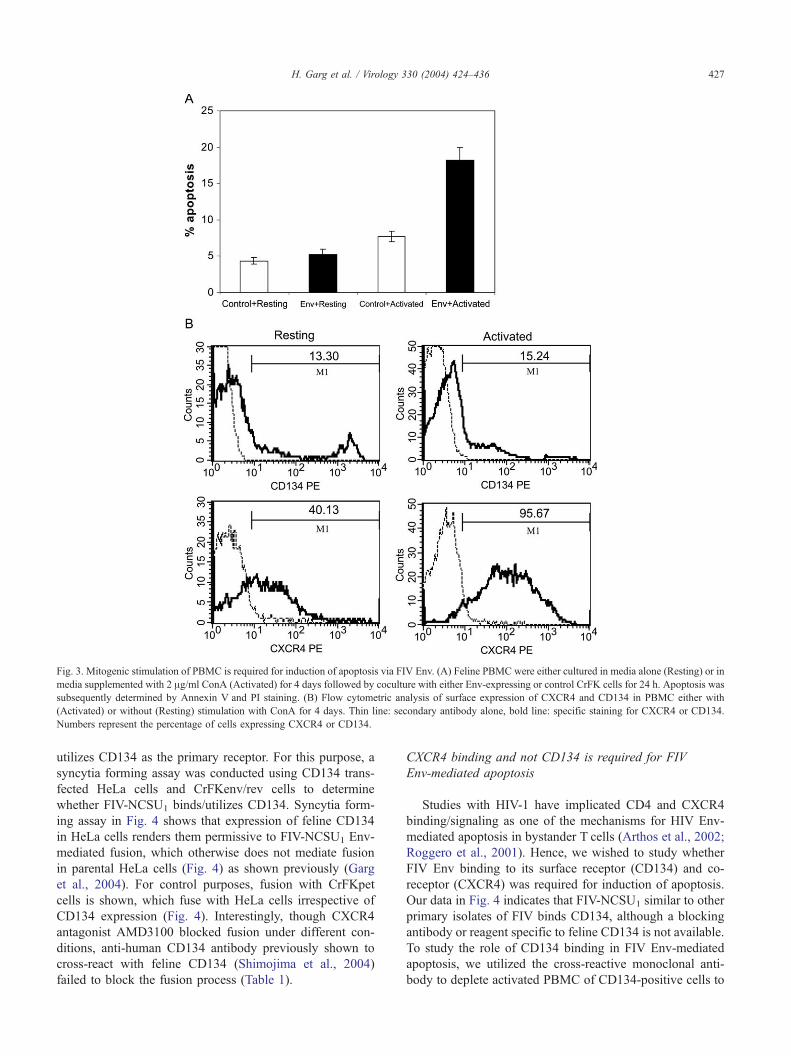
Fig. 3. Mitogenic stimulation of PBMC is required for induction of apoptosis via FIV Env. (A) Feline PBMC were either cultured in media alone (Resting) or in
media supplemented with 2 Ag/ml ConA (Activated) for 4 days followed by coculture with either Env-expressing or control CrFK cells for 24 h. Apoptosis was
subsequently determined by Annexin V and PI staining. (B) Flow cytometric analysis of surface expression of CXCR4 and CD134 in PBMC either with
(Activated) or without (Resting) stimulation with ConA for 4 days. Thin line: secondary antibody alone, bold line: specific staining for CXCR4 or CD134.
Numbers represent the percentage of cells expressing CXCR4 or CD134.
H. Garg et al. / Virology 330 (2004) 424–436 427
utilizes CD134 as the primary receptor. For this purpose, a
syncytia forming assay was conducted using CD134 trans-
fected HeLa cells and CrFKenv/rev cells to determine
whether FIV-NCSU1 binds/utilizes CD134. Syncytia form-
ing assay in Fig. 4 shows that expression of feline CD134
in HeLa cells renders them permissive to FIV-NCSU1 Env-
mediated fusion, which otherwise does not mediate fusion
in parental HeLa cells (Fig. 4) as shown previously (Garg
et al., 2004). For control purposes, fusion with CrFKpet
cells is shown, which fuse with HeLa cells irrespective of
CD134 expression (Fig. 4). Interestingly, though CXCR4
antagonist AMD3100 blocked fusion under different con-
ditions, anti-human CD134 antibody previously shown to
cross-react with feline CD134 (Shimojima et al., 2004)
failed to block the fusion process (Table 1).
CXCR4 binding and not CD134 is required for FIV
Env-mediated apoptosis
Studies with HIV-1 have implicated CD4 and CXCR4
binding/signaling as one of the mechanisms for HIV Env-
mediated apoptosis in bystander T cells (Arthos et al., 2002;
Roggero et al., 2001). Hence, we wished to study whether
FIV Env binding to its surface receptor (CD134) and co-
receptor (CXCR4) was required for induction of apoptosis.
Our data in Fig. 4 indicates that FIV-NCSU1 similar to other
primary isolates of FIV binds CD134, although a blocking
antibody or reagent specific to feline CD134 is not available.
To study the role of CD134 binding in FIV Env-mediated
apoptosis, we utilized the cross-reactive monoclonal anti-
body to deplete activated PBMC of CD134-positive cells to
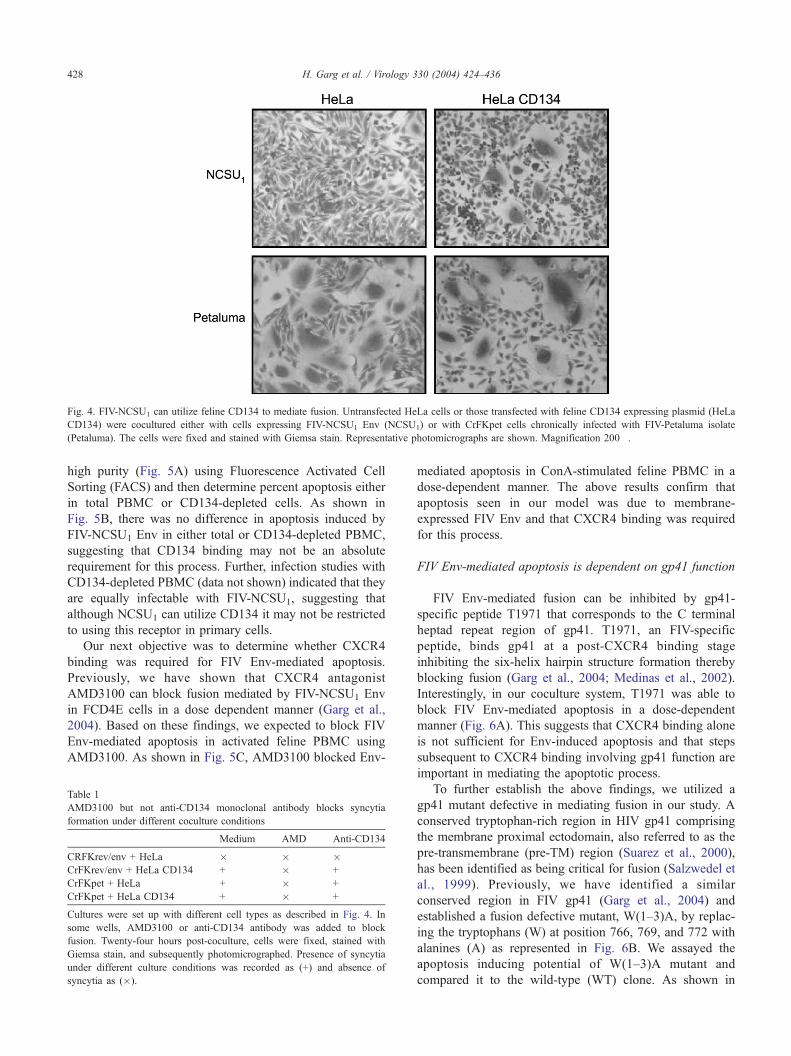
Fig. 4. FIV-NCSU1 can utilize feline CD134 to mediate fusion. Untransfected HeLa cells or those transfected with feline CD134 expressing plasmid (HeLa
CD134) were cocultured either with cells expressing FIV-NCSU1 Env (NCSU1) or with CrFKpet cells chronically infected with FIV-Petaluma isolate
(Petaluma). The cells were fixed and stained with Giemsa stain. Representative photomicrographs are shown. Magnification 200�.
H. Garg et al. / Virology 330 (2004) 424–436428
high purity (Fig. 5A) using Fluorescence Activated Cell
Sorting (FACS) and then determine percent apoptosis either
in total PBMC or CD134-depleted cells. As shown in
Fig. 5B, there was no difference in apoptosis induced by
FIV-NCSU1 Env in either total or CD134-depleted PBMC,
suggesting that CD134 binding may not be an absolute
requirement for this process. Further, infection studies with
CD134-depleted PBMC (data not shown) indicated that they
are equally infectable with FIV-NCSU1, suggesting that
although NCSU1 can utilize CD134 it may not be restricted
to using this receptor in primary cells.
Our next objective was to determine whether CXCR4
binding was required for FIV Env-mediated apoptosis.
Previously, we have shown that CXCR4 antagonist
AMD3100 can block fusion mediated by FIV-NCSU1 Env
in FCD4E cells in a dose dependent manner (Garg et al.,
2004). Based on these findings, we expected to block FIV
Env-mediated apoptosis in activated feline PBMC using
AMD3100. As shown in Fig. 5C, AMD3100 blocked Env-
Table 1
AMD3100 but not anti-CD134 monoclonal antibody blocks syncytia
formation under different coculture conditions
Medium AMD Anti-CD134
CRFKrev/env + HeLa � � �CrFKrev/env + HeLa CD134 + � +
CrFKpet + HeLa + � +
CrFKpet + HeLa CD134 + � +
Cultures were set up with different cell types as described in Fig. 4. In
some wells, AMD3100 or anti-CD134 antibody was added to block
fusion. Twenty-four hours post-coculture, cells were fixed, stained with
Giemsa stain, and subsequently photomicrographed. Presence of syncytia
under different culture conditions was recorded as (+) and absence of
syncytia as (�).
mediated apoptosis in ConA-stimulated feline PBMC in a
dose-dependent manner. The above results confirm that
apoptosis seen in our model was due to membrane-
expressed FIV Env and that CXCR4 binding was required
for this process.
FIV Env-mediated apoptosis is dependent on gp41 function
FIV Env-mediated fusion can be inhibited by gp41-
specific peptide T1971 that corresponds to the C terminal
heptad repeat region of gp41. T1971, an FIV-specific
peptide, binds gp41 at a post-CXCR4 binding stage
inhibiting the six-helix hairpin structure formation thereby
blocking fusion (Garg et al., 2004; Medinas et al., 2002).
Interestingly, in our coculture system, T1971 was able to
block FIV Env-mediated apoptosis in a dose-dependent
manner (Fig. 6A). This suggests that CXCR4 binding alone
is not sufficient for Env-induced apoptosis and that steps
subsequent to CXCR4 binding involving gp41 function are
important in mediating the apoptotic process.
To further establish the above findings, we utilized a
gp41 mutant defective in mediating fusion in our study. A
conserved tryptophan-rich region in HIV gp41 comprising
the membrane proximal ectodomain, also referred to as the
pre-transmembrane (pre-TM) region (Suarez et al., 2000),
has been identified as being critical for fusion (Salzwedel et
al., 1999). Previously, we have identified a similar
conserved region in FIV gp41 (Garg et al., 2004) and
established a fusion defective mutant, W(1–3)A, by replac-
ing the tryptophans (W) at position 766, 769, and 772 with
alanines (A) as represented in Fig. 6B. We assayed the
apoptosis inducing potential of W(1–3)A mutant and
compared it to the wild-type (WT) clone. As shown in
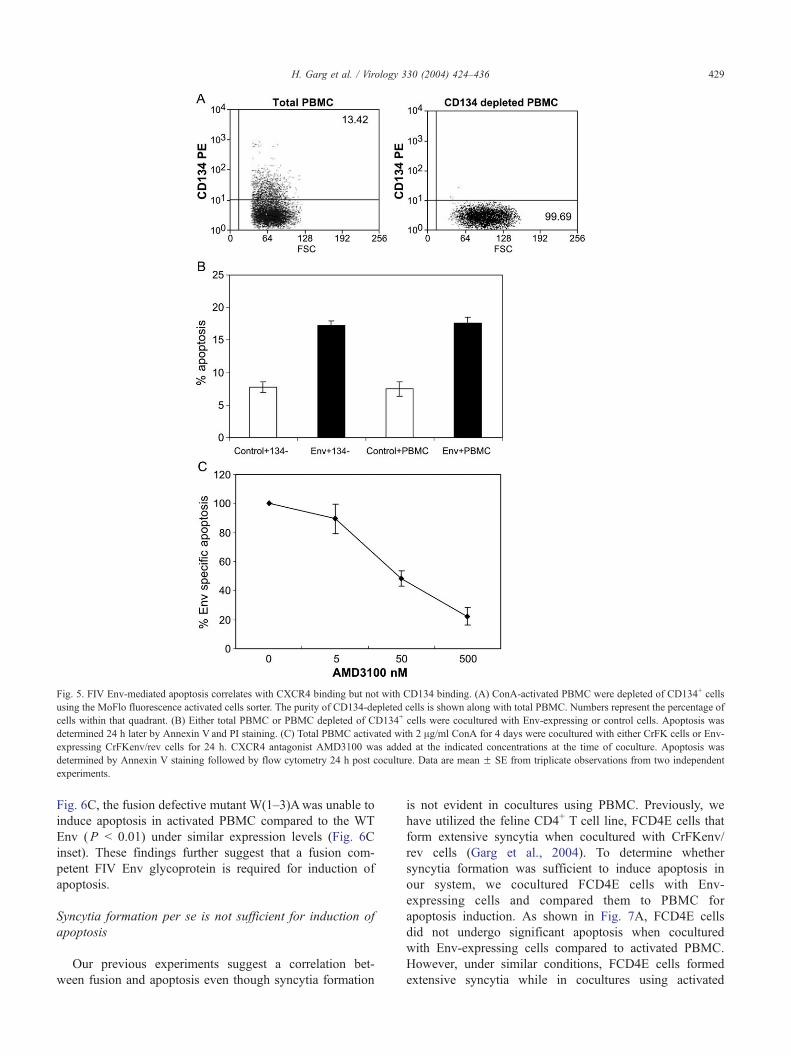
Fig. 5. FIV Env-mediated apoptosis correlates with CXCR4 binding but not with CD134 binding. (A) ConA-activated PBMC were depleted of CD134+ cells
using the MoFlo fluorescence activated cells sorter. The purity of CD134-depleted cells is shown along with total PBMC. Numbers represent the percentage of
cells within that quadrant. (B) Either total PBMC or PBMC depleted of CD134+ cells were cocultured with Env-expressing or control cells. Apoptosis was
determined 24 h later by Annexin V and PI staining. (C) Total PBMC activated with 2 Ag/ml ConA for 4 days were cocultured with either CrFK cells or Env-
expressing CrFKenv/rev cells for 24 h. CXCR4 antagonist AMD3100 was added at the indicated concentrations at the time of coculture. Apoptosis was
determined by Annexin V staining followed by flow cytometry 24 h post coculture. Data are mean F SE from triplicate observations from two independent
experiments.
H. Garg et al. / Virology 330 (2004) 424–436 429
Fig. 6C, the fusion defective mutant W(1–3)Awas unable to
induce apoptosis in activated PBMC compared to the WT
Env (P b 0.01) under similar expression levels (Fig. 6C
inset). These findings further suggest that a fusion com-
petent FIV Env glycoprotein is required for induction of
apoptosis.
Syncytia formation per se is not sufficient for induction of
apoptosis
Our previous experiments suggest a correlation bet-
ween fusion and apoptosis even though syncytia formation
is not evident in cocultures using PBMC. Previously, we
have utilized the feline CD4+ T cell line, FCD4E cells that
form extensive syncytia when cocultured with CrFKenv/
rev cells (Garg et al., 2004). To determine whether
syncytia formation was sufficient to induce apoptosis in
our system, we cocultured FCD4E cells with Env-
expressing cells and compared them to PBMC for
apoptosis induction. As shown in Fig. 7A, FCD4E cells
did not undergo significant apoptosis when cocultured
with Env-expressing cells compared to activated PBMC.
However, under similar conditions, FCD4E cells formed
extensive syncytia while in cocultures using activated
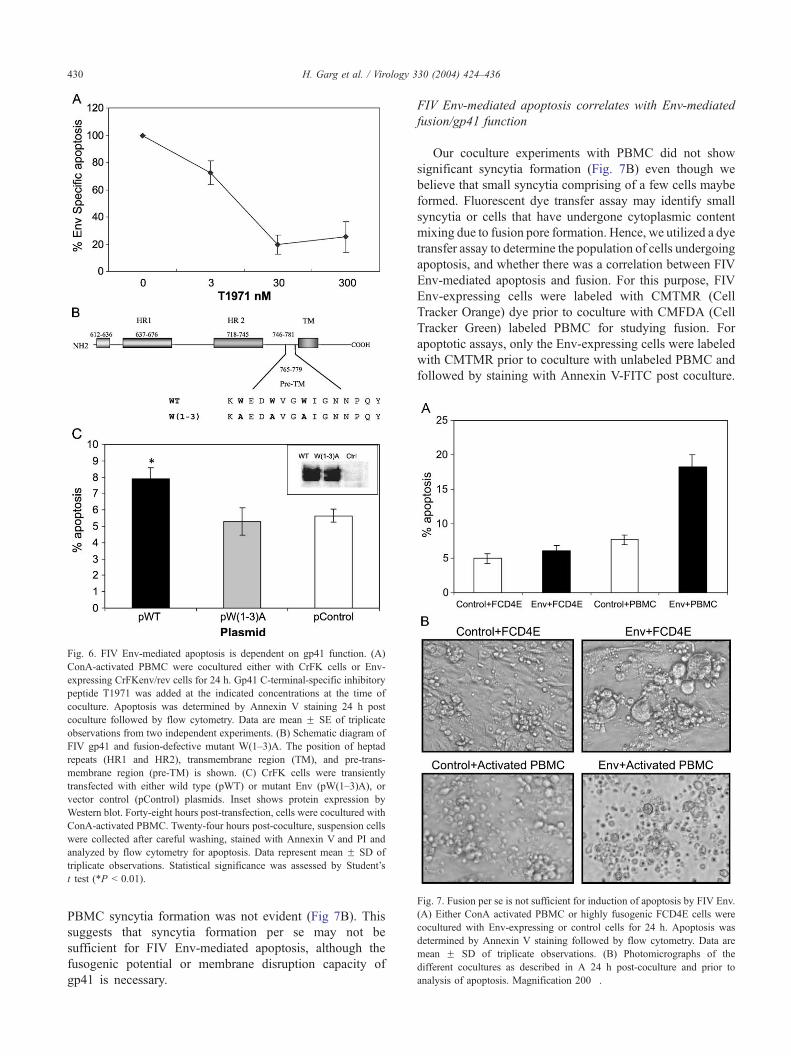
Fig. 6. FIV Env-mediated apoptosis is dependent on gp41 function. (A)
ConA-activated PBMC were cocultured either with CrFK cells or Env-
expressing CrFKenv/rev cells for 24 h. Gp41 C-terminal-specific inhibitory
peptide T1971 was added at the indicated concentrations at the time of
coculture. Apoptosis was determined by Annexin V staining 24 h post
coculture followed by flow cytometry. Data are mean F SE of triplicate
observations from two independent experiments. (B) Schematic diagram of
FIV gp41 and fusion-defective mutant W(1–3)A. The position of heptad
repeats (HR1 and HR2), transmembrane region (TM), and pre-trans-
membrane region (pre-TM) is shown. (C) CrFK cells were transiently
transfected with either wild type (pWT) or mutant Env (pW(1–3)A), or
vector control (pControl) plasmids. Inset shows protein expression by
Western blot. Forty-eight hours post-transfection, cells were cocultured with
ConA-activated PBMC. Twenty-four hours post-coculture, suspension cells
were collected after careful washing, stained with Annexin V and PI and
analyzed by flow cytometry for apoptosis. Data represent mean F SD of
triplicate observations. Statistical significance was assessed by Student’s
t test (*P b 0.01).
Fig. 7. Fusion per se is not sufficient for induction of apoptosis by FIV Env.
(A) Either ConA activated PBMC or highly fusogenic FCD4E cells were
cocultured with Env-expressing or control cells for 24 h. Apoptosis was
determined by Annexin V staining followed by flow cytometry. Data are
mean F SD of triplicate observations. (B) Photomicrographs of the
different cocultures as described in A 24 h post-coculture and prior to
analysis of apoptosis. Magnification 200�.
H. Garg et al. / Virology 330 (2004) 424–436430
PBMC syncytia formation was not evident (Fig 7B). This
suggests that syncytia formation per se may not be
sufficient for FIV Env-mediated apoptosis, although the
fusogenic potential or membrane disruption capacity of
gp41 is necessary.
FIV Env-mediated apoptosis correlates with Env-mediated
fusion/gp41 function
Our coculture experiments with PBMC did not show
significant syncytia formation (Fig. 7B) even though we
believe that small syncytia comprising of a few cells maybe
formed. Fluorescent dye transfer assay may identify small
syncytia or cells that have undergone cytoplasmic content
mixing due to fusion pore formation. Hence, we utilized a dye
transfer assay to determine the population of cells undergoing
apoptosis, and whether there was a correlation between FIV
Env-mediated apoptosis and fusion. For this purpose, FIV
Env-expressing cells were labeled with CMTMR (Cell
Tracker Orange) dye prior to coculture with CMFDA (Cell
Tracker Green) labeled PBMC for studying fusion. For
apoptotic assays, only the Env-expressing cells were labeled
with CMTMR prior to coculture with unlabeled PBMC and
followed by staining with Annexin V-FITC post coculture.
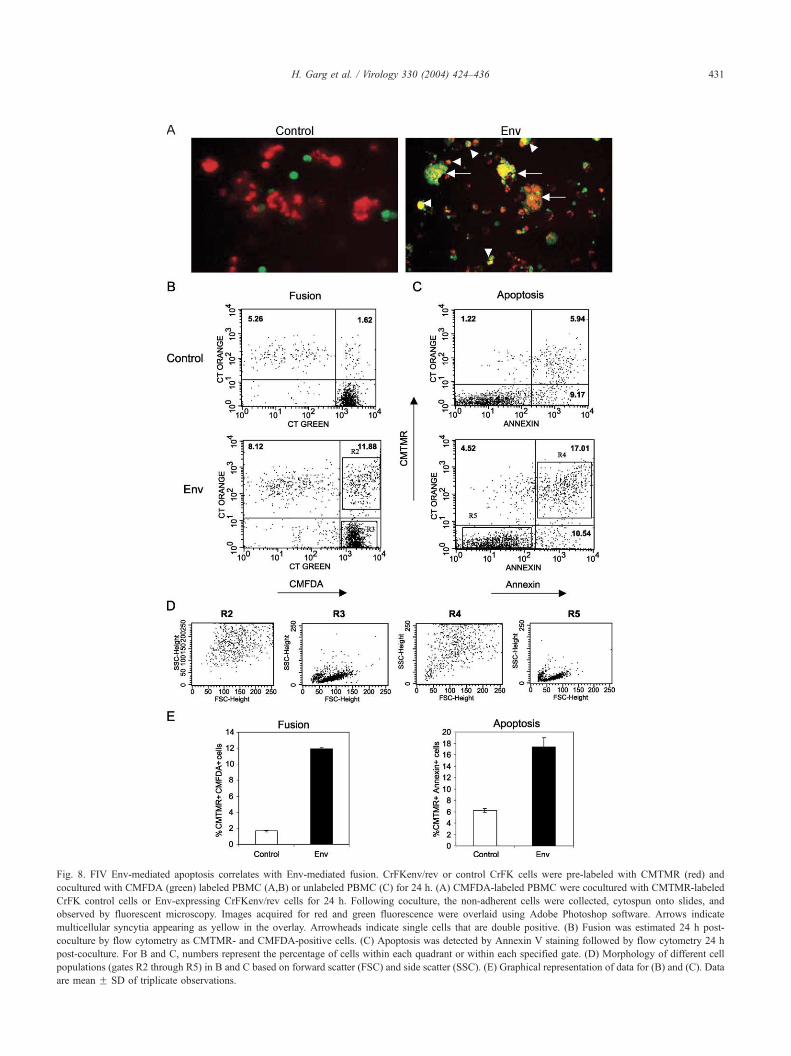
Fig. 8. FIV Env-mediated apoptosis correlates with Env-mediated fusion. CrFKenv/rev or control CrFK cells were pre-labeled with CMTMR (red) and
cocultured with CMFDA (green) labeled PBMC (A,B) or unlabeled PBMC (C) for 24 h. (A) CMFDA-labeled PBMC were cocultured with CMTMR-labeled
CrFK control cells or Env-expressing CrFKenv/rev cells for 24 h. Following coculture, the non-adherent cells were collected, cytospun onto slides, and
observed by fluorescent microscopy. Images acquired for red and green fluorescence were overlaid using Adobe Photoshop software. Arrows indicate
multicellular syncytia appearing as yellow in the overlay. Arrowheads indicate single cells that are double positive. (B) Fusion was estimated 24 h post-
coculture by flow cytometry as CMTMR- and CMFDA-positive cells. (C) Apoptosis was detected by Annexin V staining followed by flow cytometry 24 h
post-coculture. For B and C, numbers represent the percentage of cells within each quadrant or within each specified gate. (D) Morphology of different cell
populations (gates R2 through R5) in B and C based on forward scatter (FSC) and side scatter (SSC). (E) Graphical representation of data for (B) and (C). Data
are mean F SD of triplicate observations.
H. Garg et al. / Virology 330 (2004) 424–436 431

H. Garg et al. / Virology 330 (2004) 424–436432
Twenty-four hours post coculture, fusion was quantified by
flow cytometry as CMTMR+ CMFDA+ double-positive cells
and apoptosis was determined by staining with FITC labeled
Annexin V (CMTMR+/�Annexin+/�). As seen in Fig. 8A,
fused cells (appearing as yellow in overlay) were distinctly
seen by fluorescent microscopy when activated PBMC
were cocultured with Env-expressing CrFKenv/rev cells
compared to control cells. Fusion here did not necessarily
represent syncytia (arrows) as single cells that were double
positive were also seen (arrowheads) most likely due to
formation of fusion pore without progressing to complete
fusion. A quantification of fusion/dye transfer by flow
cytometry revealed that approximately 10% of the cells
were double positive in Env cocultures (11.88%) when
compared to background control cocultures (1.62%). We
then determined percent apoptosis in cocultured target cells
by Annexin V staining under identical coculture conditions
(except that the target PBMC were unlabeled). Interest-
ingly, a similar increase of 10% (5.94–17.01%) was seen
in CMTMR+Annexin V+ cells in Env cocultures compared
to control cocultures. Further, there was no significant
increase in apoptosis of CMTMR� cells (9.17–10.54%,
Fig. 8C). Since the previously unlabelled target cells
(activated PBMC) had taken up the CMTMR dye, this
population most likely represents cells that have formed
fusion pores with Env-expressing cells. To further validate
the above findings, we determined whether the fused and
apoptotic populations were morphologically similar based
on the FSC/SSC characteristics. As seen in Fig. 8D, the
fused CMTMR+CMFDA+ (R2 gate) cells were almost
identical to the apoptotic CMTMR+Annexin V+ (R4 gate)
cells based on forward and side scatter characteristics.
Moreover, they were morphologically distinct from
CMTMR negative (R3 gate) and non-apoptotic (R5 gate)
cells. These results suggest that apoptosis in feline PBMC
was primarily occurring post fusion pore formation as a
result of coculture with Env-expressing cells, indicating a
strong correlation between FIV Env-mediated fusion and
apoptosis.
Discussion
Feline immunodeficiency virus infection is a relevant and
well-characterized animal model for human AIDS (English
et al., 1993; Mizuno et al., 2001). Based on the similarities
between FIV and HIV pathogenesis, it is speculated that
both viruses share common mechanism(s) for inducing
immunodeficiency. Even though it has been known for
sometime that FIV causes T cell depletion in a manner
similar to HIV-1 (Guiot et al., 1997; Johnson et al., 1996;
Tompkins et al., 2003), the molecular mechanisms involved
in FIV-induced T cell loss have not been established. The
Env glycoprotein of HIV has been implicated in apoptotic
killing of bystander T cells leading to immunodeficiency
(Etemad-Moghadam et al., 2001). Conflicting reports
suggest the role of gp120 binding to CD4 and chemokine
receptors versus fusion mediated by Env glycoprotein in
initiating apoptotic signaling. The objective of the present
study was to determine (1) whether FIV Env glycoprotein
was capable of inducing apoptosis in bystander cells and (2)
if this process was dependent on receptor (CD134), co-
receptor (CXCR4) binding, or Env-mediated fusion.
The fact that FIV Env utilizes CD134 and not CD4 as its
primary receptor (Hosie et al., 1993; Shimojima et al., 2004)
but can still initiate apoptosis in bystander cells indicates
that CD4 signaling may not be necessary for lentiviral
Env-mediated apoptosis. We show here that membrane-
expressed FIV Env can induce apoptosis in activated PBMC
but not resting cells, and this correlates with increased
surface expression of the CXCR4 but not CD134. While
CXCR4 is a well-known activation marker (Bleul et al.,
1997), our observations with regard to lack of CD134 up-
regulation following ConA stimulation are consistent with
findings in other species where ConA-mediated activation
failed to up-regulate surface CD134 expression (Barten et
al., 2001). Interestingly, similar to HIV-1, FIV Env binding
to CXCR4 is required for apoptosis, suggesting that it is a
common requirement for both HIV and FIV Env-induced T
cell death. However, inhibition of FIV Env-mediated
apoptosis at steps post CXCR4 binding using either gp41-
specific inhibitory peptides or a mutation in gp41 pre-
transmembrane region suggests that CXCR4 itself may not
be sufficient for activating the apoptotic signaling pathway.
Although our studies regarding the primary receptor CD134
were not conclusive in the absence of suitable blocking
antibodies, inhibition of the fusion process at steps post
receptor and co-receptor binding abrogates apoptosis,
suggesting that the binding of CD134 and/or CXCR4 is
not sufficient for induction of FIV-NCSU1 Env-mediated
apoptosis. The observation that fusion-inhibiting agents/
mutations inhibit T cell apoptosis indicates a strong
correlation between the two processes. Our findings are
contrary to the hypothesis that CXCR4 binding and signal-
ing is sufficient for lentivirus Env-mediated apoptosis
(Arthos et al., 2002; Penn et al., 1999), but are in agreement
with recent reports showing that HIV Env-mediated
apoptosis can be blocked by T20, an HIV gp41-specific
fusion inhibitor (Barretina et al., 2003; Blanco et al., 2003;
Scheller and Jassoy, 2001).
The correlation between HIV Env-mediated fusion and
apoptosis remains controversial. While some reports show
the potential of soluble Env glycoprotein to induce
apoptosis in T cells (Corbeil and Richman, 1995), others
have demonstrated the necessity of membrane-bound Env
glycoprotein for induction of apoptosis (Laurent-Crawford
et al., 1995). Though it has been established that HIV Env
can signal via chemokine receptors (CXCR4/CCR5)
(Davis et al., 1997), and some authors suggest that HIV
Env mediates apoptotic signaling via engaging CD4 and/or
CXCR4 (Arthos et al., 2002; Vlahakis et al., 2001), others
have shown that signaling via either of these receptors is

H. Garg et al. / Virology 330 (2004) 424–436 433
not required for HIV Env-mediated apoptosis (Biard-
Piechaczyk et al., 2000; Blanco et al., 1999). While
pathogenicity of HIV is often related to the fusogenic
potential of the Env glycoprotein (Alimonti et al., 2003;
Gougeon and Montagnier, 1993), the requirement of fusion
between infected and uninfected bystander cells for
apoptosis remains controversial.
HIV infections are characterized highly cytopathic
syncytia-inducing (SI) and relatively less pathogenic non-
syncytia-inducing (NSI) strains based on the in vitro
phenotype of the viruses. The likelihood of finding SI
phenotype in a patient is associated with poor immuno-
logical parameters, rapid CD4+ T cell loss and disease
progression (Koot et al., 1993; Richman and Bozzette,
1994). The mechanism by which SI strains tend to be more
pathogenic than NSI strains is not established. SI strains
utilize CXCR4 as co-receptor, which is expressed practi-
cally on all CD4+ T cells while NSI strains generally utilize
CCR5, which is expressed only on a limited number of
CD4+ T cells especially in the gut-associated lymphoid
tissue (Anton et al., 2000). Based on co-receptor expression
by T cells, X4 viruses tend to be more infectious, which may
be a cause of their increased pathogenicity (Fouchier et al.,
1996). On the other hand, cells infected with SI strains
would be more effective at causing apoptosis of CXCR4
expressing bystander T cells via binding by Env glycopro-
tein and mediating fusion. Even though NSI strains are
considered less cytopathic, they have been shown to be
capable of CD4+ T cell killing by a mechanism that is not
directly related to the rate of viral replication (Yu et al.,
1994). In a recent report, LaBonte et al. (2003) have shown
that CCR5 utilizing viruses can cause cytolysis of CCR5+
target cells by a process that is again dependent on
membrane fusion.
Our findings utilizing the highly fusogenic FCD4E cells
suggests that although syncytia formation itself may not be
the determining factor in inducing apoptosis, the fusogenic
potential and membrane disruption properties of FIV Env
are required for this process. Further analysis of this
phenomenon is necessary utilizing membrane specific
lipophilic dyes and nondiffusible cytoplasmic labeling. We
believe that the discrepancy between FCD4E cells and
PBMC might be due to differences in membrane structure
and organization as well as signaling factors. This correlates
with the observation that although pathogenicity of HIV
correlates with the fusogenic potential of Env glycoprotein,
formation of syncytia in vivo are rare.
Based on the above evidence, it is clear that interaction of
lentiviral Env glycoprotein with target cell membranes may
result in membrane destabilization subsequently leading to
apoptosis of target cells. Our findings are in agreement with
others (Roumier et al., 2003), further strengthening the
hypothesis that gp41-mediated membrane perturbation may
be important for FIV as well as HIV-induced apoptosis. This
is the first report showing the potential of membrane-
expressed FIV Env to induce apoptosis in bystander cells.
Also, by blocking Env-mediated apoptosis at the level of
gp41, we show a strong correlation between fusion and
apoptosis mediated by FIV Env. Our findings not only
strengthen the hypothesis that lentiviral Env glycoproteins
may be involved in T cell apoptosis but also indicate strong
similarities between FIV and HIV pathogenesis.
Materials and methods
Cells and reagents
Crandell Feline Kidney (CrFK), CrFK cells expressing
FIV-NCSU1 Env glycoprotein (CrFKenv/rev) (Garg et al.,
2004), and CrFKpet (CrFK cells chronically infected with
the Petaluma isolate of FIV) were maintained in Dulbecco’s
Modified Eagle’s Medium (DMEM) (Mediatech) supple-
mented with 10% fetal bovine serum (FBS), penicillin (100
U/ml), and streptomycin (100 Ag/ml). Medium for
CrFKenv/rev cells was supplemented with G-418 (Gibco-
BRL, Gaithesberg, MD) at 700 Ag/ml. HeLa cells were
maintained in DMEM medium supplemented with 10%
FBS, penicillin (100 U/ml) and streptomycin (100 Ag/ml).
FCD4E, a CD4+ T cell line highly permissive for FIV-
NCSU1 infection (English et al., 1993) was cultured in
RPMI medium supplemented as described below. Periph-
eral blood lymphocytes were obtained from FIV negative
cats by Percoll density gradient centrifugation as described
previously (English et al., 1993). The donor cats were
housed at the Laboratory Animal Resource facility of the
College of Veterinary Medicine, North Carolina State
University, as per the federal guidelines and institutional
policies. Purified PBMC were cultured in RPMI 1640
medium (Mediatech) supplemented with 10% FBS, strep-
tomycin (100 Ag/ml), penicillin (100 IU/ml), l-glutamine
(2 mM), HEPES (15 mM), sodium pyruvate (2 mM) and h-mercaptoethanol (2.5 � 10�5 M). PBMC were activated
with 2 Ag/ml ConA (Sigma) and cultured for 4 days prior
to use in the coculture experiments. Plasmid expressing
feline CD134 was a kind gift from Dr. Brian Willett
(University of Glasgow). CXCR4 antagonist AMD3100
was a kind gift from Dr. Edward Hoover (Colorado State
University). FIV gp41-specific fusion inhibitor T1971,
shown to inhibit FIV Env-mediated fusion, was a kind gift
from Robyn Medinas (Trimeris Inc. Durham, NC).
Flow cytometry and cell sorting
Cell surface CXCR4 expression was measured using the
cross-reacting anti-CXCR4 monoclonal antibody (44717,
BD Biosciences) followed by staining with Phycoerythrin
(PE) conjugated goat anti-mouse IgG (Jackson ImmunoR-
esearch Laboratories, PA). PE-conjugated anti CD134 anti-
body BerACT35 (Ancell Corporation, Bayport, MN)
previously shown to cross-react with the feline CD134
homolog (Shimojima et al., 2004) was used to detect surface

H. Garg et al. / Virology 330 (2004) 424–436434
CD134 expression. Cells were acquired using the FACSCa-
libur flow cytometer (BD Biosciences) and analyzed using
the CellQuest software. For blocking experiments, the
nonconjugated anti-CD134 antibody was used. PBMC were
sorted into purified CD134+ Gaithesberg and CD134�
populations using a high-speed MoFlo fluorescence acti-
vated cell sorter (DakoCytomation) at the flow cytometry
facility at North Carolina State University.
Coculture experiment
CrFK or CrFKenv/rev cells were seeded in 24-well plates
at 2 � 105 cells/well and allowed to adhere overnight. After
24 h, medium was aspirated from the wells and 4 � 105
PBMC activated with 2 Ag/ml ConA for 4 days were added
to the wells. Inhibitors AMD3100 or T1971 were added at
the time of coculture. The cells were cocultured for 24 or
48 h. Subsequently, the suspension cells were collected by
careful washing and used for viability or apoptotic assays.
Transfection of cells
HeLa cells were transfected using Effectene transfection
reagent (Qiagen, CA) as per the manufacturer’s instructions.
Twenty-four hours post transfection, the cells were seeded
in 96-well plates and used for syncytia forming assay.
Syncytia-forming assay
HeLa cells either untransfected or transfected with feline
CD134 were seeded in 96-well plates at 104 cells per well
and allowed to adhere overnight. Subsequently, CrFK cells
expressing FIV-NCSU1 Env (CrFKenv/rev) or chronically
infected with FIV-Petaluma (CrFKpet) were added at 1000
cells per well. In some wells, either AMD3100 or anti-
CD134 antibody was added for blocking. The cells were
cocultured for 24 h after which the plates were fixed and
stained with Giemsa stain as described previously (Garg
et al., 2004).
Measurement of viability
Viability of PBMC was measured by the MTT [3-(4,5-
dimethylthiozol-2-yl)-2,5-diphenyl tetrazolium bromide]
(Sigma) assay as described previously (Mosmann, 1983)
with slight modifications. Briefly, ConA-activated PBMC
were cocultured with either CrFK or CrFKenv/rev cells for
24 or 48 h following which the non-adherent cells were
collected and re-plated in 96-well plates at 100 Al/well.Thirty microlitres of MTT reagent (5 mg/ml in PBS) was
added to each well and incubated for 4 h. Subsequently, the
plates were centrifuged, supernatant discarded and 100 Al of0.1 N HCl in isopropanol added to the wells. The plates
were incubated for an additional 1 h and then 100 Al ofdistilled water added per well. Finally, the plates were read
at 530/650 nm in an ELISA plate reader (Tecan).
Detection of apoptosis
Morphological change in suspension cells was detected
24 h post coculture by measuring the forward scatter (FSC)
versus side scatter (SSC) by flow cytometry on a FACSCa-
libur flow cytometer (BD Biosciences). At least 15000
events were collected and analyzed using the CellQuest
software. Apoptosis was detected by staining suspension
cells with FITC-conjugated Annexin V, a 35–36 kDa protein
that binds to phosphatidylserine exposed on the surface of
apoptotic cells. Twenty-four hours post coculture, the
suspension cells were stained with Annexin V and PI
(Roche Diagnostics, IN) as per the manufacturer’s instruc-
tions and analyzed by flow cytometry. Cells positive for
Annexin V but negative for PI were considered apoptotic,
while PI-positive cells were excluded as dead cells.
Inhibitors AMD3100 or T1971 were added at the time of
coculture for some experiments.
Detection of fusion and apoptosis using fluorescent labeling
of cells
To determine FIV Env-mediated fusion in activated
PBMC, a two-color dye redistribution assay was developed.
Briefly, CrFK or CrFKenv/rev cells were labeled with
cytoplasmic dye 5- and 6-{[(4-choloromethyl) benzoyl]
amino} tetramethyl rhodamine (CMTMR/Cell Tracker
Orange, Molecular Probes) (5AM in PBS for 30 min),
washed and seeded in 24-well plates at 2 � 105 cells per
well. Next day, activated PBMC labeled with cytoplasmic
dye 5-chloromethylfluorescein diacetate (CMFDA/Cell
Tracker Green, Molecular Probes) (0.5 AM in PBS for 30
min) were added to the wells at 4 � 105 cells per well. The
cells were cocultured for 24 h, following which suspension
cells were analyzed either by fluorescent microscopy or by
flow cytometry. A similar assay with slight modification
was used to detect apoptosis in activated PBMC after
coculture with Env-expressing cells. CrFKenv/rev or non-
transfected CrFK cells labeled with CMTMR dye were
cocultured with unlabeled ConA-activated PBMC. Apopto-
sis was detected the subsequent day by staining suspensions
cells with Annexin V. Cells positive for Annexin V but
negative for CMTMR were single unfused PBMC that were
apoptotic. Cells positive for Annexin V as well as CMTMR
were Env-expressing cells undergoing apoptosis, most
likely post fusion.
Acknowledgments
The authors would like to thank Dr. Edward Hoover
(Colorado State University) for AMD3100, Robyn Medinas
(Trimeris Inc. Durham, NC) for T1971, and Dr. Brian
Willett (University of Glasgow) for the feline CD134
expression plasmid. The authors are also thankful to Dr.
Frederick Fuller (N.C. State University) for critical review

H. Garg et al. / Virology 330 (2004) 424–436 435
of the manuscript. This work was supported in part by
National Institute of Health grant AI43858.
References
Alimonti, J.B., Ball, T.B., Fowke, K.R., 2003. Mechanisms of CD4+ T
lymphocyte cell death in human immunodeficiency virus infection and
AIDS. J. Gen. Virol. 84, 1649–1661.
Anton, P.A., Elliott, J., Poles, M.A., McGowan, I.M., Matud, J., Hultin,
L.E., Grovit-Ferbas, K., Mackay, C.R., Chen, I.S.Y., Giorgi, J.V.,
2000. Enhanced levels of functional HIV-1 co-receptors on human
mucosal T cells demonstrated using intestinal biopsy tissue. AIDS 14,
1761–1765.
Arthos, J., Cicala, C., Selig, S.M., White, A.A., Ravindranath, H.M., Van
Ryk, D., Steenbeke, T.D., Machado, E., Khazanie, P., Hanback, M.S.,
Hanback, D.B., Rabin, R.L., Fauci, A.S., 2002. The role of the CD4
receptor versus HIV co-receptors in envelope-mediated apoptosis in
peripheral blood mononuclear cells. Virology 292, 98–106.
Barretina, J., Blanco, J., Armand-Ugon, M., Gutierrez, A., Clotet, B., Este,
J.A., 2003. Anti-HIV-1 activity of enfuvirtide (T-20) by inhibition of
bystander cell death. Antiviral Ther. 8, 155–161.
Barten, M.J, Gummert, J.F, van Gelder, T., Shorthouse, R., Morris,
R.E., 2001. Flow cytometric quantitation of calcium-dependent and
-independent mitogen-stimulation of T cell functions in whole
blood: inhibition by immunosuppressive drugs in vitro. J. Immunol.
Methods 253, 95–112.
Biard-Piechaczyk, M., Robert-Hebmann, V., Richard, V., Roland, J.,
Hipskind, R.A., Devaux, C., 2000. Caspase-dependent apoptosis of
cells expressing the chemokine receptor CXCR4 is induced by cell
membrane-associated human immunodeficiency virus type 1 Envelope
glycoprotein (gp120). Virology 268, 329–344.
Blanco, J., Jacotot, E., Cabrera, C., Cardona, A., Clotet, B., De Clercq, E.,
Este, J.A., 1999. The implication of the chemokine receptor CXCR4 in
HIV-1 Envelope protein-induced apoptosis is independent of the G
protein-mediated signaling. AIDS 13, 909–917.
Blanco, J., Barretina, J., Ferri, K.F., Jacotot, E., Gutierrez, A., Armand-
Ugon, M., Cabrera, C., Kroemer, G., Clotet, B., Este, J.A., 2003. Cell-
surface-expressed HIV-1 envelope induces the death of CD4 T cells
during GP41-mediated hemifusion-like events. Virology 305, 318–329.
Bleul, C., Wu, C.L., Hoxie, J.A., Springer, T.A., Mackay, C.R., 1997. The
HIV co-receptors CXCR4 and CCR5 are differentially expressed and
regulated on human T lymphocytes. Proc. Natl. Acad. Sci. U.S.A. 94,
1925–1930.
Corbeil, J., Richman, D.D., 1995. Productive infection and subsequent
interaction of CD4-gp120 at the cellular membrane is required for HIV-
induced apoptosis of CD4+ T cells. J. Gen. Virol. 76, 681–690.
Davis, C.B., Dikic, I., Unutmaz, D., Hill, C.M., Arthos, J., Siani, M.A.,
Thompson, D.A., Schlessinger, J., Littman, D.R., 1997. Signal trans-
duction due to HIV-1 Envelope interactions with chemokine receptors
CXCR4 or CCR5. J. Exp. Med. 86, 1793–1798.
Desai, B.M., Zhang, D., Dayes, N., Green, D.R., Weiner Muthumani, K.,
Hwang, D.S., 2002. HIV-1 Vpr induces apoptosis through caspase 9 in
T cells and peripheral blood mononuclear cells. J. Biol. Chem. 277,
37820–37831.
English, R.V., Johnson, C.M., Gebhard, D.H., Tompkins, M.B., 1993. In
vivo lymphocyte tropism of feline immunodeficiency virus. J. Virol. 67,
5175–5186.
Etemad-Moghadam, B., Rhone, D., Steenbeke, T., Sun, Y., Manola, J.,
Gelman, R., Fanton, J.W., Racz, P., Tenner-Racz, K., Axthelm, M.K.,
Letvin, N.L., Sodroski, J., 2001. Membrane-fusing capacity of the
human immunodeficiency virus envelope proteins determines the
efficiency of CD+ T-cell depletion in macaques infected by a simian-
human immunodeficiency virus. J. Virol. 75, 5646–5655.
Famularo, G., De Simone, C., Marcellini, S., 1997. Apoptosis: mechanisms
and relation to AIDS. Med. Hypotheses 48, 423–429.
Ferri, K.F., Jacotot, E., Geuskens, M., Kroemer, G., 2000. Apoptosis and
karyogamy in syncytia induced by the HIV-1-Envelope glycoprotein
complex. Cell Death Differ. 7, 1137–1139.
Finkel, T.H., Tudor-Williams, G., Banda, N.K., Cotton, M.F., Curiel, T.,
Monks, C., Baba, T.W., Ruprecht, R.M., Kupfer, A., 1995. Apoptosis
occurs predominantly in bystander cells and not in productively
infected cells of HIV- and SIV-infected lymph nodes. Nat. Med. 1,
129–134.
Fouchier, R.A., Meyaard, L., Brouwer, M., Hovenkamp, E., Schuitemaker,
H., 1996. Broader tropism and higher cytopathicity for CD4+ T cells of
a syncytium-inducing compared to a non-syncytium-inducing HIV-1
isolate as a mechanism for accelerated CD4+ T cell decline in vivo.
Virology 219, 87–95.
Garg, H., Fuller, F., Tompkins, W., 2004. Mechanism of feline immuno-
deficiency virus envelope glycoprotein mediated fusion. Virology 321,
274–286.
Golding, H., Shearer, G.M., Hillman, K., Lucas, P., Manischewitz, J., Zajac,
R.A.,Clerici,M.,Gress,R.E.,Boswell,R.N.,Golding,B., 1989.Common
epitope in human immunodeficiencyvirus (HIV) I-GP41andHLAclass II
elicits immunosuppressive autoantibodies capable of contributing to
immune dysfunction in HIV I-infected individuals. J. Clin. Invest. 83,
1430–1435.
Gougeon, M.L., Montagnier, L., 1993. Apoptosis in AIDS. Science 260,
1269–1270.
Guiot, A.L., Rigal, D., Chappuis, G., 1997. Spontaneous programmed cell
death (PCD) process of lymphocytes of FIV-infected cats: pharmaco-
logical modulation in vitro. Int. J. Immunopharmacol. 19, 167–179.
Hoffmann-Fezer, G., Thum, J., Ackley, C., Herbold, M., Mysliwietz, J.,
Thefeld, S., Hartmann, K., Kraft, W., 1992. Decline in CD4+ cell
numbers in cats with naturally acquired feline immunodeficiency virus
infection. J. Virol. 66, 1484–1488.
Hosie, M.J., Willett, B.J., Dunsford, T.H., Jarrett, O., Neil, J.C., 1993. A
monoclonal antibody which blocks infection with feline immunodefi-
ciency virus identifies a possible non-CD4 receptor. J. Virol. 67,
1667–1671.
Johnson, C.M., Benson, N.A., Papadi, G.P., 1996. Apoptosis and CD4+
lymphocyte depletion following feline immunodeficiency virus infec-
tion of a T-lymphocyte cell line. Vet. Pathol. 33, 195–203.
Johnston, J.B., Silva, C., Power, C., 2002. Envelope gene-mediated
neurovirulence in feline immunodeficiency virus infection: induction
of matrix metalloproteinases and neuronal injury. Virology 76,
2622–2633.
Kolesnitchenko, V., Wahl, L.M., Tian, H., 1995. Human immunodeficiency
virus 1 Envelope-initiated G2-phase programmed cell death. Proc. Natl.
Acad. Sci. U.S.A. 92, 11889–11893.
Koot, M., Keet, I.P., Vos, A.H., de Goede, R.E., Roos, M.T., Coutinho,
R.A., Miedema, F., Schellekens, P.T., Tersmette, M., 1993. Prognostic
value of HIV-1 syncytium-inducing phenotype for rate of CD4+ cell
depletion and progression to AIDS. Ann. Intern. Med. 118, 681–688.
LaBonte, J.A., Madani, N., Sodroski, J., 2003. Cytolysis by CCR5-using
human immunodeficiency virus type 1 envelope glycoproteins is
dependent on membrane fusion and can be inhibited by high levels
of CD4 expression. J. Virol. 77, 6645–6659.
Laurent-Crawford, A.G., Coccia, E., Krust, B., Hovanessian, A.G., 1995.
Membrane-expressed HIV Envelope glycoprotein heterodimer is a
powerful inducer of cell death in uninfected CD4+ target cells. Res.
Virol. 146, 5–17.
Lenardo, M.J., Angleman, S.B., Bounkeua, V., Bolton, D.L., 2002.
Cytopathic killing of peripheral blood CD4(+) T lymphocytes by
human immunodeficiency virus type 1 appears necrotic rather than
apoptotic and does not require Env. J. Virol. 76, 5082–5093.
Medinas, R.J., Lambert, D.M., Tompkins, W.A., 2002. C-Terminal gp40
peptide analogs inhibit feline immunodeficiency virus: cell fusion and
virus spread. J. Virol. 76, 9079–9086.
Meyaar, L., Otto, S.A., Jonker, R.R., Mijnster, M.J., Keet, R.P., Miedema,
F., 1992. Programmed death of T cells in HIV-1 infection. Science 257,
217–219.

H. Garg et al. / Virology 330 (2004) 424–436436
Mizuno, T., Goto, Y., Baba, K., Masuda, K., Ohno, K., Tsujimoto, H., 2001.
TNF-alpha-induced cell death in feline immunodeficiency virus-
infected cells is mediated by the caspase cascade. Virology 287,
446–455.
Momoi, Y., Mizuno, T., Nishimura, Y., Endo, Y., Ohno, K., Watari, T.,
Goitsuka, R., Tsujimoto, H., Hasegawa, A., 1996. Detection of
apoptosis induced in peripheral blood lymphocytes from cats infected
with feline immunodeficiency virus. Arch. Virol. 141, 1651–1659.
Mosmann, T., 1983. Rapid colorimetric assay for cellular growth and
survival: application to proliferation and cytotoxicity assays. J. Immunol.
Methods 65, 55–63.
Muro-Cacho, C.A., Pantaleo, G., Fauci, A.S., 1995. Analysis of apoptosis
in lymph nodes of HIV-infected persons. Intensity of apoptosis
correlates with the general state of activation of the lymphoid tissue
and not with stage of disease or viral burden. J. Immunol. 154,
5555–5566.
Nardelli, B., Gonzalez, C.J., Schechter, M., Valentine, F.T., 1995. CD4+
blood lymphocytes are rapidly killed in vitro by contact with autologous
human immunodeficiency virus-infected cells. Proc. Natl. Acad. Sci.
U.S.A. 92, 7312–7316.
Novotney, C., English, R.V., Housman, J., Davidson, M.G., Nasisse, M.P.,
Jeng, C.R., Davis, W.C., Tompkins, M.B., 1990. Lymphocyte pop-
ulation changes in cats naturally infected with feline immunodeficiency
virus. AIDS 4, 1213–1218.
Penn, M.L., Grivel, J.C., Schramm, B., Goldsmith, M.A., Margolis, L.,
1999. CXCR4 utilization is sufficient to trigger CD4+ T cell depletion
in HIV-1-infected human lymphoid tissue. Proc. Natl. Acad. Sci. U.S.A.
96, 663–668.
Purvis, S.F., Jacobberger, J.W., Sramkoski, R.M., Patki, A.H., Lederman,
M.M., 1995. HIV type 1 Tat protein induces apoptosis and death in
Jurkat cells. AIDS Res. Hum. Retroviruses 11, 443–450.
Richman, D.D., Bozzette, S.A., 1994. The impact of the syncytium-
inducing phenotype of human immunodeficiency virus on disease
progression. J. Infect. Dis. 169, 968–974.
Roggero, R., Robert-Hebmann, V., Harrington, S., Roland, J., Vergne, L.,
Jaleco, S., Devaux, C., Biard-Piechaczyk, M., 2001. Binding of human
immunodeficiency virus type 1 gp120 to CXCR4 induces mitochondrial
transmembrane depolarization and cytochrome c-mediated apoptosis
independently of Fas signaling. J. Virol. 75, 7637–7650.
Roumier, T., Castedo, M., Perfettini, J.L., Andreau, K., Metivier, D.,
Zamzami, N., Kroemer, G., 2003. Mitochondrion-dependent cas-
pase activation by the HIV-1 envelope. Biochem. Pharmacol. 66,
1321–1329.
Salzwedel, K., West, J.T., Hunter, E., 1999. A conserved tryptophan-rich
motif in the membrane-proximal region of the human immunodefi-
ciency virus type 1 gp41 ectodomain is important for Env-mediated
fusion and virus infectivity. J. Virol. 73, 2469–2480.
Scheller, C., Jassoy, C., 2001. Syncytium formation amplifies apoptotic
signals: a new view on apoptosis in HIV infection in vitro. Virology
282, 48–55.
Schwartz, O., Alizon, M., Heard, J.M., Danos, O., 1994. Impairment of T
cell receptor-dependent stimulation in CD4+ lymphocytes after contact
with membrane-bound HIV-1 Envelope glycoprotein. Virology 198,
360–365.
Shimojima, M., Miyazawa, T., Ikeda, Y., McMonagle, E.L., Haining, H.,
Akashi, H., Takeuchi, Y., Hosie, M.J., Willett, B.J., 2004. Use of
CD134 as a primary receptor by the feline immunodeficiency virus.
Science 303, 1192–1195.
Suarez, T., Nir, S., Goni, F.M., Saez-Cirion, A., Nieva, J.L., 2000. The pre-
transmembrane region of the human immunodeficiency virus type-1
glycoprotein: a novel fusogenic sequence. FEBS Lett. 477, 145–149.
Tompkins, M.B., Bull, M.E., Dow, J.L., Ball, J.M., Collisson, E.W.,
Winslow, B.J., Phadke, A.P., Vahlenkamp, T.W., Tompkins, W.A.,
2003. Feline immunodeficiency virus infection is characterized by B7 +
CTLA4 + T cell apoptosis. J. Infect. Dis. 185, 1077–1093.
Vlahakis, S.R., Algeciras-Schimnich, A., Bou, G., Heppelmann, C.J.,
Villasis-Keever, A., Collman, R.C., Paya, C.V., 2001. Chemokine-
receptor activation by Env determines the mechanism of death in HIV-
infected and uninfected T lymphocytes. J. Clin. Invest. 107, 207–215.
Willett, B.J., Picard, L., Hosie, M.J., Turner, J.D., Adema, K., Clapham,
P.R., 1997. Shared usage of the chemokine receptor CXCR4 by the
feline and human immunodeficiency viruses. J. Virol. 71, 6407–6415.
Wyatt, R., Sodroski, J., 1998. The HIV-1 Envelope glycoproteins: fusogens,
antigens, and immunogens. Science 280, 1884–1888.
Yu, X., McLane, M.F., Ratner, L., O’Brien, W., Collman, R., Essex, M.,
Lee, T.H., 1994. Killing of primary CD4+ T cells by non-syncytium-
inducing macrophage-tropic human immunodeficiency virus type 1.
Proc. Natl. Acad. Sci. U.S.A. 91, 10237–10241.





























