Embed Size (px)
Citation preview

Vol. 4. .153-358. June 1995 Cancer Epidemiology, Biomarkers & Prevention 353
3 The abbreviation used is: 0-Dma, 0-demethylangolensin.
Fecal Lignan and Isoflavonoid Excretion in Premenopausal Women
Consuming Flaxseed Powder1
Mindy S. Kurzer,2 Johanna W. Lampe,Margaret C. Martini, and Herman Adlercreutz
Department of Food Science and Nutrition, University of Minnesota, St. Paul,
Minnesota 55108 [M. S. K., J. W. L.. M. C. M.], and Department of Clinical
Chemistry, University of Helsinki, Meilahti Hospital, Helsinki, Finland [H. A.]
Abstract
Lignans and isoflavonoids are diphenolic compoundsfound in plant foods, particularly whole grains andlegumes. They have been shown to have anticarcinogenicproperties in animal and cell studies, and have beenassociated with reduced cancer risk in epidemiobogicalstudies. In order to perform further epidemiobogical andmetabolic studies on these compounds, it is necessary to
be able to monitor concentrations in biological samples.In this study, we examined the effects of consumption of
flaxseed, a concentrated source of lignans, on fecal lignanexcretion and evaluated the effect of high lignanconsumption on fecal excretion of isoflavonoids. Thirteenwomen were studied for two diet periods of threemenstrual cycles each in a cross-over design. During thecontrol period, they consumed their usual diets; duringthe treatment period they consumed their usual dietssupplemented with 10 g/day ground flaxseed. Feces werecollected on days 7-1 1 of the last menstrual cycle in eachdiet period. Five-day fecal composites were analyzed forlignans and isoflavonoids by isotope dilution gas
chromatography-mass spectrometry. Fecal excretion ofthe lignans enterodiol, enterolactone, and matairesinobincreased significantly with flax consumption, from80.0 ± 80.0 (SD) to 2560 ± 3100; 640 ± 480 to 10,300± 7580; and 7.33 ± 10.0 to 11.9 ± 8.06 nmol/day,respectively. There were no differences in fecal excretionof the isoflavonoids, daidzein, equol, genistein, andO-demethylangolensin.
Introduction
Numerous components of plant foods have been associatedwith decreased cancer risk. Among these are lignans and isofla-vones, two groups of diphenolic compounds most concentratedin whole grains and legumes (Fig. 1). Epidemiobogical studieshave shown decreased cancer risk with increased excretion ofthese compounds, while animal, cellular, and metabolic studies
Received 9/2/94: revised 12/21/94: accepted 12/21/94.
I This research was supported by NCI Grant N01-CN-05288-O1 and MinnesotaAgricultural Experiment Station Project 18-34. J. W. L. was supported by
National (‘ancer Institute Grant 5T32-CA-09607. Development of the method
was supported by NIH Grant I ROI CA 56289-01.
2 To whom requests for reprints should be addressed, at Department of Food
Science and Nutrition, University of Minnesota, 1334 Eckles Avenue, St. Paul,MN 55108.
have shown important biological effects that contribute to their
potential as chemopreventive agents.Lignans have been shown to inhibit skin, breast, colon,
and lung cancer cell growth (1, 2). Consumption of flaxsecd, aconcentrated source of lignans (3), has been shown to inhibit
mammary tumor promotion (4) and development of early mark-ens of risk of mammary cancinogenesis (5). Proposed mecha-nisms by which lignans may inhibit carcinogenesis includeantivinal (6) and antioxidant (7) activities. In addition, thesimilarities in structure among lignans, estradiol, and the syn-thetic antiestrogen tamoxifen (Fig. 1) suggest that lignans mayalso exert their anticarcinogenic effects in part as a result ofantiestrogenic effects. In fact, lignans have been shown toinhibit placental (8) and adipocyte (9) estrogen synthesis; toinhibit cstnadiol-induced proliferation of MCF-7 human breast
carcinoma cells (10); and to stimulate sex hormone-bindingglobulin synthesis, with subsequent decrease in free estradiol(11). Flaxseed consumption has been shown to alter the men-strual cycle in premenopausal women in a potentially cancer-
preventive direction (12).Isoflavonoids have been reported to inhibit cancer cell
growth and development in lung (13), stomach (14), lcukocyte(15, 16), and breast (17, 18) carcinoma cells. Proposed mech-
anisms include in vitro inhibition of bacterial growth (19, 20),as well as in vitro inhibition of placental (8) and adipocyte (9,21) estrogen synthesis. Isoflavonoids have been shown in vivato have antispasmodic effects in guinea pig ileum (22), antihy-
pertensive effects in rats (23), and hypolipemic effects inrabbits (24).
Despite great interest in the physiological effects of big-nans and isoflavonoids, little is known about human metabo-
lism, absorption, and actual exposure. Mammalian lignans aresynthesized by cobonic microflona from plant precursors foundnaturally in many plant foods, particularly oilseeds and grains(3, 25). Flaxsecd is an oilseed known to be a concentratedsource of lignan precursors (3). Consumption of flaxsced ne-suits in large increases in urinary excretion of entenodiol and
enterolactone, the primary mammalian lignans derived frombacterial metabolism of their plant precursors, secoisolanicires-mob and matainesinol (26). Entenolactone also originates frombacterial oxidation of enterodiol.
Genistein and daidzein arc the two primary dietary isofla-vones found in high concentrations in legumes. Genistein and
daidzein can also originate from bacterial metabolism of theirrespective glycosides, as well as metabolism of two otherisoflavones, biochanin A and formononetin. Daidzein can be
metabolized further by intestinal microfbora to the isofla-vonoids 0-Dma3 and equol (25, 27).
Human studies of the physiological effects of dietary big-nans and isoflavones require accurate measurement of these
on April 14, 2019. © 1995 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from

HO
�cIciOL �
Daidzein Genistein
OcH�CHaNtcH3)2
H3 “‘ H
Tamoxifen
HO,
Enterodiol Enterolactone
Fig. 1. Structures of selected isoflavones (daidzein and genistein), estrogens
(tamoxifen and estradiol). and lignans (enterodiol and enterolactone).
354 Fecal Lignan and Isoflavonoid Excretion
compounds in biological fluids, yet few studies have reportedsuch data. Adlercreutz et a!. (1 1, 28-30) have reported urinaryand serum (31, 32) concentrations of entenolactone, enterodiol,and matairesinol, as well as genistein, daidzein, equol, and0-Dma, in cross-sections of omnivorous and vegetarian sub-jects in Boston, Helsinki, and Japan. Xu ci a!. (33) have
published the only human study of isoflavone bioavaibability,reporting plasma, urinary, and fecal daidzein and genisteinconcentrations in adult women. Although they report fecalisoflavone concentrations after consumption of isoflavone-nichsoymilk, Xu et a!. (33) include no baseline data. Recently,Adlercreutz et a!. (34) described a method for simultaneousmeasurement of fecal lignans and isoflavonoids, reporting pre-liminary data in omnivorous and vegetarian women. There havebeen no further reports of fecal lignan concentrations.
Fecal concentrations of isoflavonoids and lignans are ofinterest for several reasons: (a) fecal concentrations may reflectthe direct exposure of colon cells to anticarcinogenic lignansand isoflavonoids; (b) the known antiviral and antifungal ad-tions of lignans and isoflavonoids may exert beneficial intralu-minal effects. Their antioxidant actions also may be importantvia prevention of the activation of cocancinogens to carcino-gens; (c) since both lignans and isoflavones are metabolized byintestinal microflora, increased lignan consumption may alter
lignan or isoflavone metabolism and excretion, perhaps byaltering microfloral numbers or enzymatic activities; and (d)knowledge of fecal concentrations and the metabolic intenac-tions among these compounds will complete the metabolicpicture necessary to perform further epidemiological studies.
This study is the first metabolic experiment to reportlignan concentrations in human feces before and after consump-tion of a concentrated source of plant lignans. We also report
typical fecal isoflavonoid concentrations in female subjectsconsuming a low isoflavone diet, as well as the effect of high
lignan consumption on isoflavonoid excretion. Entenobactone,
enterodiol, matairesinob, genistein, daidzcin, equol, and 0-Dmawere measured in 5-day fecab composites collected from 13healthy young women after 3 menstrual cycles on their habitual
diets and after 3 menstrual cycles in which their usual dietswere supplemented with 10 g/day ground flaxseed. In addition
to determining the fecal concentrations of individual com-pounds, correlations were performed to evaluate the associa-tions among individual isoflavonoids and bignans.
Materials and Methods
Subjects. This study was a substudy of a larger study of theeffects of flaxseed consumption on lignan excretion and themenstrual cycle (12, 26). For the large study, 18 healthy pre-menopausal women were studied. Health status of the subjectswas verified by health history, routine blood and urine screen-ing, and physical exam. Selected subjects were sedentary, om-nivorous nonsmokers, who typically consumed diets containing<10 g dietary fiber/day. All subjects reported regular menstrual
cycles and stable body weights for at least I year, had not usedoral contraceptives for at beast 3 months, had not been onantibiotics for the previous month, and were not on any regularmedication.
For this study, 13 women selected from the original 18subjects were willing to collect feces. The prestudy averages for
age, height, weight, and body mass index of these 13 subjectswere 27.8 ± 4.3 year, 1.64 ± 0.05 m, 60.6 ± 8.5 kg, and22.5 ± 2.2 kg/m2, respectively (Table 1).
Study Design and Diet. The protocol for this study was ap-proved by the University of Minnesota Institutional ReviewBoard Human Subjects Committee. The study design and diethave been described in detail previously (26). The study was
performed as a balanced, randomized, cross-oven design fob-bowing the individual menstrual cycles of each subject. After an
acclimation period of I menstrual cycle, each subject wasstudied for 6 menstrual cycles. For the first 3 menstrual cycles,one-half of the subjects were on their habitual diets (control
diet) and one-half were on their habitual diets supplementedwith 10 g/day ground flaxseed (Enneco Corp., Manitowoc, WI;
treatment). For the last 3 menstrual cycles, the groups switched
diets.Subjects were free living during the entire study. Although
they consumed their habitual diets, they were given instructionsto minimize isoflavone and lignan consumption by avoiding alllegumes (due to the high isoflavone content) and flaxsced-
containing products (certain prepared bread and cereal items).Food intake was monitored by 3-day diet records kept twice permenstrual cycle. Diet records were analyzed for nutrient con-
tent with the use of a computerized nutrient analysis program(Nutritionist III; N-Squared Computing, Silvcnton, OR).
For the treatment period, each daily dose of 10 g/day
ground flaxseed was packaged individually and stored frozen.Subjects received weekly supplies of flaxseed, with instructionsto keep frozen until consumption. They were also instructed toconsume the entire contents of each individual package everyday, splitting it into 2 or 3 servings and adding it raw to
prepared foods. Daily flaxseed intake was recorded by eachsubject, and uneaten portions were returned and weighed.
Sample Collection and Analysis. The 13 subjects collectedfeces on days 7-1 1 of the last menstrual cycle of each dietperiod. The methods used for fecal collection and preparationhave been described previously (35). Each fecab sample was
collected in a plastic bag. Immediately after defecation, 10 mlof 10% (w/v) ascorbic acid were poured over the feces, air was
on April 14, 2019. © 1995 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from

Cancer Epidemiology, Biomarkers & Prevention 355
Table I Subject characteristics and dietary intake”
Control diet Flax supplemented
Age (yr)
Height (m)
Weight (kg)
BMI” (kg/m2)
Energy intake (kcal/day)
Protein intake (g/day)
Fat intake (g/day)
Carbohydrate intake (g/day)
Dietary fiber (g/day)
27.80 ± 4.30 (23.00-34.00)
1.64 ± 0.05 (1.55-1.75)
61.00 ± 9.00 (53.00-85.00)
22.50 ± 2.20 (20.10-27.70)
1834.00 ± 1 10.00 (1 182.00-2564.00)
79.00 ± 5.00(51.00-111.00)
75.00 ± 5.00 (42.00-103.00)
213.00 ± 17.00 (124.00-312.00)
6.00 ± 1.00 (1.00-11.00)
27.80 ± 4.30 (23.0O-34.0())
1.64 ± 0.00 (1.55-1.75)
61.()0 ± 9.00(51.00-85.00)
22.50 ± 2.20(20.10-27.70)
2015.00 ± 191.00 (1085.00-3208.00)
82.00 ± 9.00(39.00-141.00)
84.00 ± 8.00 (41(10-137(X))
249.00 ± 28.00(140.00-405(X))
11(X) ± 1.00(7.00-17.00)
“ Arithmetic X ± SD (range): n 13.
I’ BMI, body mass index.
forced from the bag, and the fecal sample was then sealed andstored at -20#{176}C. A 5-day fecal composite was created byweighing all individual fecal samples for each subject to de-(ermine total wet weight, adding 0.1% (w/v) sodium azide, and
homogenizing the feces. Aliquots of the composite were frozenat -20#{176}C for later bignan and isoflavonoid determination.Duplicate abiquots were freeze dried for determination of dry
weight.
Fecal lignans and isoflavonoids were analyzed with theuse of a modification of the urinary method developed byAdlencneutz et a!. (29). Fecal lignans and isoflavonoids wereextracted and separated by ion exchange chromatography andquantified by capillary gas chromatography-mass spectrometnyin the selective ion-monitoring mode. The complete fecabmethod has been published separately (34). Briefly, deuterated
bignan and isoflavonoid standards were added to 0.3-0.6-gfecab samples. After extraction and filtration in a 1 :9 acetone:ethanol solution, samples were left overnight in 70% ethanol toprecipitate proteins. Samples were then run through a series of
Sep-Pak C1,, (Waters Associates, Inc., Milford, MA), DEAE-Sephadex-acetate (Pharmacia Fine Chemicals, Uppsaba, Swe-den), and QAE-Sephadex-canbonate (Pharmacia Fine Chemi-
cabs) columns. Following denivatization, analyses were carriedout with the use of a Hewlett-Packard 5995 quadrapole gaschromatography-mass spectrometry instrument. Samples wereanalyzed in triplicate, and all samples from a given subject were
analyzed in the same run. Triplicate quality control sampleswere also analyzed in each run.
Intra-assay coefficients of variation for the fecab methodwere found to be 6.4, 10.1, 5.1, 6.2, 6.2, 9.8, and 7.4% for0-Dma, equol, enterodiol, enterolactone, daidzein, genistein,and matainesinol, respectively. Interassay coefficients of van-ation were 8.0, 12.1, 5.5, 8.6, 9.3, 24.7, and 6.8% for 0-Dma,equob, enterodiob, entenobactonc, daidzein, genistein, and ma-tairesinol, respectively. The recoveries of added phytoestrogens
were between 80.0 and 103.6%.
Statistics. Although bignans and isoflavonoids have been hy-pothesized to have many physiologically significant effects, it
is unknown whether it is the concentration or total quantityexcreted that is the important parameter to be followed. As aresult, all data were analyzed on a pen g dry weight, per g wetweight, and pen day basis. Statistical analyses were performedwith the use of the Statistical Analysis System (36). The effectof diet on fecal bignan and isoflavonoid excretion was evaluatedwith the use of pained t tests. Pearson correlations were per-formed within each diet period to evaluate associations amongthe individual lignans and isoflavonoids. Since some of thevariables were not normally distributed, t tests and correlationswere performed on log-transformed as well as nontransformed
data. Results were considered significant at levels of P < 0.05.Data are presented as both nontransformed and geometricmeans. For correlation analyses of log-transformed data, 0.1was entered in place of zero values.
Results
Diet and Body Weight. Diet and body weight results for the18 subjects of the large study have been published previously
( 26). Mean consumption of energy and macronutnients for the13 subjects in this substudy are shown in Table 1. These datainclude the contribution from the 10 g/day flaxseed, which
included 184 Id (44 kcal), 2 g protein, 4 g fat, and 4 g dietaryfiber/day. Flax consumption resulted in no significant differ-ences in consumption of energy, carbohydrate, protein, or fat
expressed as g/day on percentage energy. Fiber intake increasedsignificantly (P < 0.0001), from 6 to 1 1 g/day, with flaxsupplementation.
There were no significant differences in body weight orbody mass index between the control and flaxsecd treatment
periods.
Fecal Lignans and Isoflavonoids. Fecal enterolactone, en-terodiob, and matairesinol excretion increased significantly with
flaxseed consumption, when expressed pen g wet weight, per gdry weight, or per day (Table 2). Although the entenodiob:enterolactone ratio appeared to increase with flaxseed, the in-crease was not significant. There were no significant effects of
flaxseed consumption on fecal isoflavonoid excretion (Table 3).Within the control diet, when results were expressed pen g
dry weight, per g wet weight, on per day, all three isoflavonoids(0-Dma, daidzein, and genistein) were correlated positivelywith each other (Tables 4 and 5). The lignan entenodiol was
correlated positively with each of the isoflavonoids, as well aswith the lignan precursor matairesinol.
When correlations were performed within the flax treat-ment period, 0-Dma was no longer associated significantly
with daidzein and genistein, although daidzein and genistemnwere still correlated positively with each other. The correlations
between enterodiol and the isoflavonoids 0-Dma, daidzein, andgenistein were lost, although entenodiol was still associatedpositively with matairesinol. When results were expressed asper g wet weight, enterolactone was associated negatively withits precursor, matairesinol. When expressed as pen day, equolwas associated positively with matairesinol and enterobactone.
Discussion
This is the first report of a metabolic study in which lignanconcentrations were measured in human feces before and after
on April 14, 2019. © 1995 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from
Control diet
Enterodiol
(nmol/g dry wt)
(nmol/g wet wt)
(nmollday)
Enterolactone
(nmol/g dry wt)
(nmol/g wet wt)
(nmol/day)
“ Arithmetic X ± SD (geometric mean); n = 13.
5 Significantly different from control diet using paired t tests; P < 0.0001.
‘ Significantly different from control diet using paired t tests; P < 0.01.
‘I Significantly different from control diet using paired t tests; P < 0.001.
� Significantly different from control diet using paired t tests; P < 0.05.
356 Fecal Lignan and Isoflavonoid Excretion
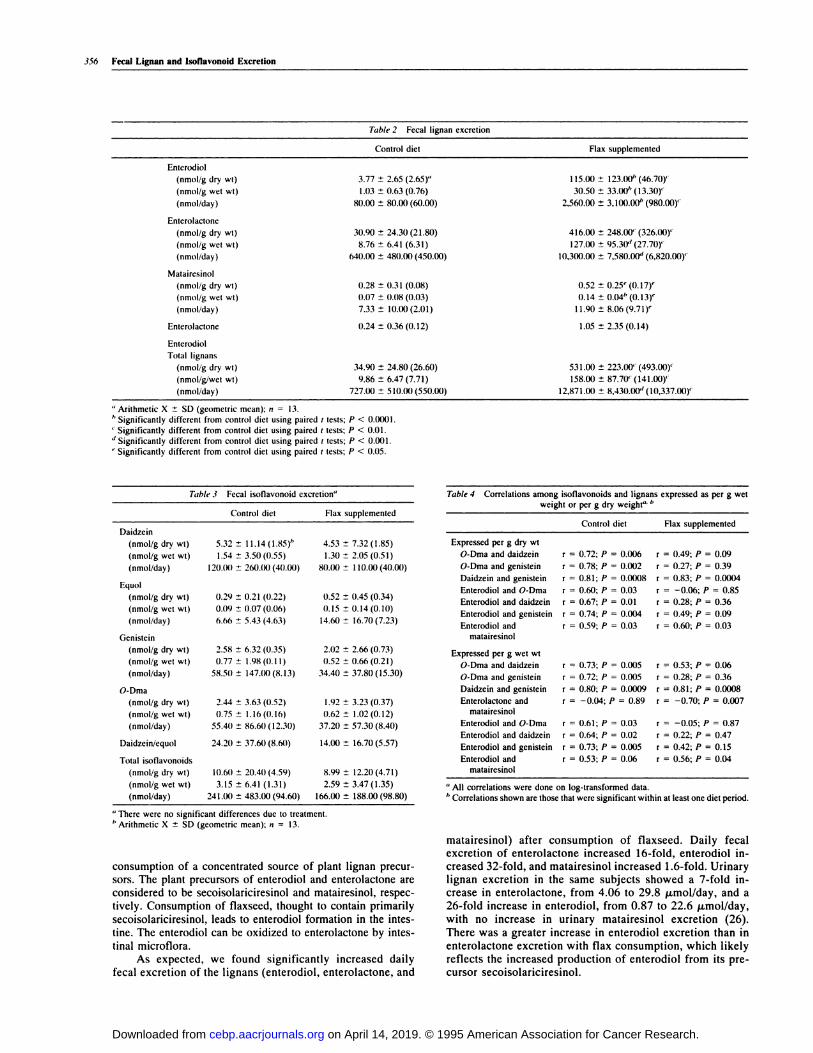
Table 2 Fecal lignan excretion
Matairesinol
(nmol/g dry wt)
(nmol/g wet wt)
(nmol/day)
Enterolactone
Enterodiol
Total lignans
(nmol/g dry wt)
(nmol/gjwet wt)
(nmol/day)
3.77 ± 2.65 (2.65)”
1 .03 ± 0.63 (0.76)
80.00 ± 80.00(60.00)
30.90 ± 24.30(21.80)
8.76 ± 6.41 (6.31)
640.00 ± 480.00(450.00)
0.28 ± 0.31 (0.08)
0.07 ± 0.08 (0.03)
7.33 ± 10.()0 (2.01)
0.24 ± 0.36 (0.12)
34.90 ± 24.80 (26.60)
9.86 ± 6.47 (7.71)
727.00 ± 510.00(550.00)
Flax supplemented
1 15.00 ± 123.00” (46.70)’
30.50 ± 33.00” (13.30)’
2,560.00 ± 3,100.00’� (980.00)’
416.00 ± 248.00’ (326.00)’
127.00 ± 95.30” (27.70)’
10,300.00 ± 7,580.00’� (6,820.00)’
0.52 ± 0.2Y (0.17)�
0.14 ± 0.04” (0.13)�
11.90 ± 8.06 (9.71)�
1.05 ± 2.35 (0.14)
531.00 ± 223.00’ (493.00)’
158.00 ± 87.70’ (141.00)’
I 2,87 1 .00 ± 8,430.00” (10,337.00)’
Ta ble 3 Fecal isoflavonoid excretion”
Control diet Flax supplemented
Daidzein
(nmol/g dry wt)
(nmol/g wet wt)
(nmollday)
5.32 ± 1 1.14 (1.85)”
1.54 ± 3.50 (0.55)
120(X) ± 260(X) (40.00)
4.53 ± 7.32 (1.85)
1.30 ± 2.05 (0.51)
80.00 ± 1 10.00 (4().00)
Equol
(nmollg dry wt)
(nmol/g wet wt)
(nmol/day)
0.29 t 0.21 (0.22)
0.09 ± 0.07 (0.06)
6.66 ± 5.43 (4.63)
0.52 � 0.45 (0.34)
0.15 ± 0.14 (0.10)
14.60 ± 16.70 (7.23)
Genistein
(nmol/g dry wt)
(nmol/g wet wt)
(nmollday)
2.58 � 6.32 (0.35)
0.77 ± 1.98(0.11)
58.50 ± 147(10 (8.13)
2.02 ± 2.66 (0.73)
0.52 ± 0.66 (0.21)
34.40 ± 37.80 (15.30)
0-Dma
(nmollg dry wt)
(nmol/g wet wt)
(nmollday)
2.44 ± 3.63 (0.52)
0.75 ± 1.16 (0.16)
55.40 ± 86.60 (12.30)
1.92 ± 3.23 (0.37)
0.62 ± 1.02 (0.12)
37.20 ± 57.30 (8.40)
Daidzeinlequol 24.20 ± 37.60 (8.6()) 14.00 ± 16.70 (5.57)
Total isoflavonoids
(nmol/g dry wt)
(nmol/g wet wt)
(nmol/day)
10.60 ± 20.40 (4.59)
3.15 ± 6.41 (1.31)
241.00 ± 483.00 (94.60)
8.99 ± 12.20 (4.71)
2.59 ± 3.47 (1.35)
166.00 ± 188.00 (98.80)
“ There were no significant differences due to treatment.
I’ Arithmetic X ± SD (geometric mean); n = 13.
consumption of a concentrated source of plant lignan precur-sons. The plant precursors of enterodiol and cntcnolactone areconsidered to be secoisolanicinesinol and matainesinol, respec-
tively. Consumption of flaxseed, thought to contain primarilysecoisobaniciresinol, leads to enterodiob formation in the intes-tine. The enterodiol can be oxidized to entenolactone by intes-
tinal microflora.As expected, we found significantly increased daily
fecal excretion of the bignans (entenodiol, enterolactone, and
Table 4 Correlations among isoflavonoids and lignans expressed as per g wet
weight or per g dry weight” b
Control diet Flax supplemented
Expressed per g dry wt
0-Dma and daidzein r = 0.72; P = 0.006 r = 0.49; P = 0.09
0-Dma and genistein r = 0.78; P = 0.002 r = 0.27; P = 0.39
Daidzein and genistein r = 0.81; P = 0.0008 r = 0.83; P = 0.0004
Enterodiol and 0-Dma r 0.60; P 0.03 r -0.06; P 0.85
Enterodiol and daidzein r = 0.67; P = 0.01 r = 0.28; P = 0.36
Enterodiol and genistein r = 0.74; P = 0.004 r 0.49; P 0.09
Enterodiol and r = 0.59; P = 0.03 r = 0.60; P = 0.03
matairesinol
Expressed per g wet wt
0-Dma and daidzein r = 0.73; P = 0.005 r = 0.53; P = 0.06
0-Dma and genistein r = 0.72; P = 0.005 r = 0.28; P = 0.36
Daidzein and genistein r = 0.80; P = 0.0009 r = 0.81; P 0.0008
Enterolactone and r = -0.04; P = 0.89 r = -0.70; P = 0.007
matairesinol
Enterodiol and 0-Dma r = 0.61; P = 0.03 r = -0.05; P 0.87
Enterodiol and daidzein r = 0.64; P = 0.02 r = 0.22; P = 0.47
Enterodiol and genistein r = 0.73; P = 0.005 r = 0.42; P = 0.15
Enterodiol and r = 0.53; P = 0.06 r 0.56; P 0.04
matairesinol
“ All correlations were done on log-transformed data.
I, Correlations shown are those that were significant within at least one diet period.
matairesinol) after consumption of flaxseed. Daily fecalexcretion of enterobactone increased 16-fold, entenodiob in-
creased 32-fold, and matairesinol increased I .6-fold. Urinarybignan excretion in the same subjects showed a 7-fold in-crease in enterolactone, from 4.06 to 29.8 �mol/day, and a26-fold increase in enterodiol, from 0.87 to 22.6 �tmol/day,with no increase in urinary matairesinol excretion (26).
There was a greater increase in enterodiol excretion than in
entenolactone excretion with flax consumption, which likelyreflects the increased production of enterodiol from its pre-cursor secoisolaniciresinol.
on April 14, 2019. © 1995 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from

Cancer Epidemiology, Biomarkers & Prevention 357
Table 5 Correlations among isoflavonoids and lignans expressed
as per day” �
Control diet Flax supplemented
0-Dma and daidzein r 0.75; P = 0.003 r 0.47; P 0.11
0-Dma and genistein r = 0.71; P = 0.006 r 0.21; P 0.49
Daidzein and genistein r = 0.83; P = 0.0(104 r 0.81; P 0.0009
Equol and matairesinol r = 0.39; P = 0.19 r 0.62; P 0.01
Equal and enterolactone r 0.45; P = 0.13 r 0.60; P 0.03
Enterodiol and 0-Dma r = 0.62; P = 0.02 r 0.08; P 0.79
Enterodiol and daidzein r - 0.72; P = 0.006 r 0.26; P 0.39
Enterodioi and genistein r = 0.74: P = 0.004 r = 0.46; P = 0.11
Enterodiol and matairesinol r = 0.63; P = 0.02 r 0.56; P 0.04
“ All correlations were done on log-transformed data.I,Correlations shown are those that were significant within at least one diet period.
The fecal cnterodiol:entenobactonc ratio in our 13 subjectsincreased from 0.24 to 1.05 with flax consumption. Using an in
vitro fermentation system to evaluate lignan production bynumerous foods incubated with human fecab bacteria, Thomp-
son et a!. (3) reported the enterodiol:entcrobactone ratio pro-duced by flaxseed flour to be 3.4. Although it is difficult tocompare these very different methodologies, both studies showgreaten entenodiol than entenobactone production, likely reflect-ing the higher concentration of secoisobaniciresinol than
matairesinol in flaxseed.In our study, low matairesinol excretion after flaxseed
consumption suggests that there is very little matairesinolpresent in the flaxseed. At the same time, correlation analysesshowed fecab matairesinol excretion to be associated positivelywith fecal enterodiol excretion. This suggests that althoughmatairesinob levels were low, flax may have been the primarysource of matainesinol just as it is known to be the primary
source of the enterodiob precursor secoisolariciresinob. On theother hand, bow levels of matainesinob may compete with 5cc-oisolanicinesinol for the same demethylating and dehydroxylat-ing enzymes, leading to decreased conversion of matairesinol to
enterolactone in the presence of high secoisolaniciresinob levels.
This is the first report of basal levels of fecal isoflavonoidsin subjects consuming a typical Western diet, which is expect-edly low in isoflavones. Although flaxseed is a concentratedsource of plant lignans, it is a minor source of isoflavones. Ourfinding that fecal isoflavonoids did not change with flaxseed
consumption was therefore unsurprising. Although consump-tion of high bevels of bignans did not significantly influenceisoflavonoid excretion, it did appear to influence isoflavonoid
metabolism by intestinal microflora. The significant associationbetween fecal 0-Dma and its isoflavone precursor daidzein,seen during the control diet, was not observed during the flaxtreatment period. This suggests that high bignan consumptionmay redirect daidzein metabolic pathways away from 0-Dmaformation and toward equob formation. In fact, with flax con-sumption, 0-Dma excretion decreased and equob excretion in-
creased, although neither change was statistically significant.On the other hand, we did not find a significant correlationbetween fecab daidzein and equol excretion. This is not surpris-
ing since it is well known that some people do not synthesizeequol significantly after consumption of a concentrated daid-zein food source such as soy (37). Since we did not separate outequol producers from nonproducers, it is expected that anycorrelations involving equol would be difficult to show.
The associations between entenodiol and the isoflavonoids
daidzein, genistein, and 0-Dma observed during the control
diet were lost during the flax treatment period. This would be
expected if the entenodiol precursors were increased signifi-
cantly with the addition of flaxseed, while the isoflavonoids,found mainly in the habitual diets, remained unchanged. Theassociations between enterodiol and the isoflavonoids observed
during the control period likely reflect common sources in thehabitual diets.
In conclusion, this is the first report of a metabolic study
in which lignan concentrations were measured in human fecesbefore and after consumption of a concentrated source of plantlignan precursors. It is also the first report of fecal isoflavonoid
concentrations in subjects consuming a typical Western diet,which is low in isoflavones. Consumption of 10 g/day groundflaxseed resulted in significantly increased fecal concentrationsof the lignans entenobactone, enterodiol, and matairesinol, whileno changes were observed in fecal concentrations of the isofla-vonoids daidzein, equol, genistein, on 0-Dma. To more fully
understand the exposure of differing populations to these com-pounds, as well as the interactions among them, it will benecessary to perform further dietary studies in which bignan and
isoflavone intakes are controlled and levels of the compounds
and their precursors are measured in urine, blood, and feces.
Improved understanding of the concentrations of lignans andisoflavones in biological fluids will enable us to perform furtherepidemiological studies in which bignans and isoflavonoidsmay be used as biomarkens of cancer risk.
Acknowledgments
We thank Susan Fredstrom for assisting with study coordination; Anja Koskela,
Sirkka Adlercreutz, and Inga Wiik for technical assistance; and the 13 subjects for
their cooperation.
References
1. Kardono, L. B., Tsauri, S., Padmawinata, K., Pezzuto, J. M., and Kinghorn,
A. D. Cytotoxic constituents of the bark of Plumeria rubra collected in Indonesia.
J. Nat. Prod., 53: 1447-1455, 1990.
2. Hirano, T., Fukuoka, K., Oka, K., Naito, T., Hosaka, K., Mitsuhashi, H., and
Matsumoto, Y. Antiproliferative activity of mammalian lignan derivatives against
the human breast carcinoma cell line, ZR-75-1. Cancer Invest., 8: 595-602, 1990.
3. Thompson, L. U., Robb, P., Serraino, S., and Cheung, F. Mammalian lignan
production from various foods. Nutr. Cancer, 16: 43-52, 1991.
4. Serraino, M., and Thompson, L. U. The effect of flaxseed consumption of theinitiation and promotional stages of mammary carcinogenesis. Nutr. Cancer. I 7:
153-159, 1992.
5. Serraino, M., and Thompson, L. U. The effect of flaxseed supplementation onearly risk markers for mammary carcinogenesis. Cancer Lett., 60: 135-142, 1991.
6. Schroder, H. C., Merz, H., Steffen, R., Muller, W. E., Sam, P. 5., Trumm, S.,
Schulz, J., and Eich, E. Differential in s’itro anti-HIV activity of natural lignans.Z. Naturforsch., 45: 1215-1221, 1990.
7. Yamashita, K., Nohara, Y., Katayama, K., and Namiki, M. Sesame seed
lignans and ‘y-tocopherol act synergistically to produce vitamin E activity in rats.
J. Nutr., 122: 2440-2446, 1992.
8. Adlercreutz, H., Bannwart, C., W#{228}h#{228}lS,K., M#{228}kelS,T., Brunow, G., Hase, T.,Arosema, P. J., Kellis, J. T., Jr., and Vickery, L. E. Inhibition of human aromata.se
by mammalian lignans and isoflavonoid phytoestrogens. J. Steroid Biochem. Mol.
Biol., 44: 147-153, 1993.
9. Wang, C., MSkelS, T., Hase, T., Adlercreutz, H., and Kurzer, M. S. Lignans
and flavonoids inhibit aromatase enzyme in human preadipocytes. J. Steroid
Biochem. Mol. Biol., 50: 205-212, 1994.
10. Mousavi, Y., and Adlercreutz, H. Enterolactone and estradiol inhibit each
other’s proliferative effect on MCF-7 breast cancer cells in culture. J. Steroid
Biochem. Mol. Biol., 4!: 615-619, 1992.
1 1. Adlercreutz, H., Mousavi, Y., Clark, J., HOckerstedt, K., H#{228}mSlainen, E.,
WShSlS, K., Makel#{228},T., and Hase, T. Dietary phytoestrogens and cancer: in euro
and in vivo studies. J. Steroid Biochem. Mol. Biol., 41: 331-337, 1992.
12. Phipps, W. R., Martini, M. C., Lampe, J. W., Slavin, J. L., and Kurzer, M. S.
Effect of flax seed ingestion on the menstrual cycle. J. Clin. Endocrinol. Metab.,
77: 1215-1219, 1993.
on April 14, 2019. © 1995 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from

358 Fecal Lignan and Isoflavonold Excretion
13. Lee, Y. S., Seo, J. S., Chung, H. T., and Jang, J. J. Inhibitory effects of
biochanin A on mouse lung tumor induced by benzo(a)pyrene. J. Korean Med.
Sci., 6: 325-328, 1991.
14. Matsukawa, Y., Marui, N., Sakai, T., Satomi, Y., Yoshida, M., Matsumoto,
K.. Nishino, H., and Aoike, A. Genistein arrests cell cyle progression at G2-M.
Cancer Res., 53: 1328-1331, 1993.
15. Traganos, F., Ardelt. B., Halko, N., Bruno, S., and Darzynkiewicz, Z. Effectsof genistein on the growth and cell cycle progression of normal human lympho-
cytes and human leukemic MOLT-4 and HL-60 cell. Cancer Res., 52:
62(X)-6208, 1992.
16. Hirano. T., Gotoh, M., and Oka, K. Natural flavonoids and lignans are potent
cytostatic agents against human leukemic HL-60 cells. Life Sci., 55: 1061-1069,
1994.
17. Peterson, G., and Barnes, S. Genistein inhibition of the growth of humanbreast cancer cells: independence from estrogen receptors and the multi-drug
resistance gene. Biochem. Biophys. Res. Commun., 179: 661-667, 1991.
18. Monti, E., and Sinha, B. K. Antiproliferative effect of genistein and adria-mycin against estrogen-dependent and -independent human breast carcinoma cell
lines. Anticancer Res., 14: 1221-1226, 1994.
19. Osawa, K., Yasuda, H., Maruyama, T., Morita, H., Takeya, K., and Itokawa,
H. Isoflavonones from the heartwood of Swartzia polvphvlla and their
antibacterial activity against cariogenic bacteria. Chem. Pharmacol. Bull.
(Tokyo). 40: 2970-2974, 1992.
20. linuma, M., Tanaka, T., Mizuno. M., Yamamoto, H., Kobayashi, Y.,and Yonemori, S. Phenolic constituents in Erv:hrina x bidwilli and their activity
against oral microbial organisms. Chem. Pharmacol. Bull. (Tokyo), 40: 2749-
2752, 1992.
21. Campbell. D. R., and Kurzer, M. S. Flavonoid inhibition of aromatase
enzyme activity in human preadipocytes. J. Steroid Biochem. Mol. Biol., 46:
381-388. 1993.
22. Herrera, M. D., Marhuenterodiola, E., and Gibson, A. Effects of genistein, an
isoflavone isolated from Genusta tridentaza, on isolated guinea-pig ileum and
guinea-pig ileai myenteric plexus. Planta. Med., 58: 314-316, 1992.
23. Wu, E. S., Loch, J. T. 3rd, Toder, B. H., Borrelli, A. R., Gawlak, D., Radov,
L. A., and Gensmantel, N. P. Flavones. III. Synthesis, biological activities, and
conformational analysis of isoflavone derivatives and related compounds. J. Med.
Chem., 35: 3519-3525, 1992.
24. Gopalan, R., Gracias, D., and Madhavan, M. Serum lipid and lipoprotein
fractions in bengal gram and biochanin A induced alterations in atherosclerosis.
Indian Heart J., 43: 185-189, 1991.
25. Borriello, S. P., Setcheil, K. D. R., Axelson, M., and Lawson, A. M.Production and metabolism of lignans by the human faecal flora. J. AppI.
Bacteriol., 58: 37-43, 1985.
26. Lampe. J. W., Martini, M. C., Kurzer, M. S., Adiercreutz, H., and Slavin, J.L. Urinary lignan and isoflavonoid excretion in premenopausal women consum-
ing flaxseed powder. Am. J. Clin. Nutr., 60: 122-128, 1994.
27. Setchell, K. D. R., and Adlercreutz, H. Mammalian lignans and phyto-
estrogens: recent studies on their formation, metabolism and biological role in
health and disease. in: I. R. Rowland (ed), Role of the Gut Flora in Toxicity and
Cancer, pp. 315-345, London: Academic Press, 1988.
28. Adlercreutz, H., Fotsis, T., Bannwart, C., WSh#{228}lS,K., M#{228}kel#{228},T., Brunow,
G., and Hase, T. Determination of urinary lignans and phytoestrogen metabolites,potential antiestrogens and anticarcinogens, in urine of women on various habit-
ual diets. J. Steroid Biochem., 25: 791-797, 1986.
29. Adlercreutz, H., Fotsis, T., Bannwart, C., WShSlS, K., Brunow, G., and Hase,
T. Isotope dilution gas chromatographic-mass spectrometric method for thedetermination of lignans and isoflavonoids in human urine, including identifica-
tion of genistein. Clin. Chim. Acta., 199: 263-278, 1991.
30. Adlercreutz, H., Honjo, H., Higashi, A., Fotsis, T., HSm#{228}lainen, E., Hase-
gawa, T., and Okada, H. Urinary excretion of lignans and isoflavonoid phy-
toestrogens in Japanese men and women consuming a traditional Japanese diet.
Am. J. Clin. Nutr., 54: 1093-1100, 1991.
31. Adlercreutz, H., Fotsis, T., Lampe, J., WShSlS, K., M#{228}kel#{228},T., Brunow, G.,
and Hase, T. Quantitative determination of lignans and isoflavonoids in plasma of
omnivorous and vegetarian women by isotope dilution gas chromatography-mass
spectrometry. Scand. J. Clin. Lab. Invest. Suppl., 215: 5-18, 1993.
32. Adlercreutz, H., Markkanen, H., and Watanabe, S. Plasma concentrations of
phyto-estrogens in Japanese men. Lancet, 342: 1209-1210, 1993.
33. Xu, X., Wang, H-J., Murphy, P. A., Cook, L., and Hendrich, S. Daidzein is
a more bioavailable soymilk isoflavone than is genistein in adult women. J. Nutr.,
124: 825-832, 1994.
34. Adlercreutz, H., Fotsis, T., Kurzer, M. S., W#{228}h#{228}l#{228},K., MSkelS, T., and Hase,T. Isotope dilution gas chromatographic-mass spectrometric method for thedetermination of unconjugated lignans and isoflavonoids in human feces, with
preliminary results in omnivorous and vegetarian women. Anal. Biochem. 225:
101-108, 1995
35. Adlercreutz, H. Diet and sex hormone metabolism. in: I. R. Rowland (ed),
Nutrition, Toxicity and Cancer, pp. 137-195. Boston: CRC Press, 1991.
36. SAS Institute, Inc. Statistical Analysis System, Ver. 6.08 (CMS). Cary, NC:
SAS Institute, Inc., 1993.
37. Setchell, K. D. R., Borriello, S. P., Hulme, P., Kirk, D. N., and Mclean, M.Nonsteroidal estrogens of dietary origin: possible roles in hormone-dependent
disease. Am. J. Clin. Nutr., 40: 569-578, 1984.
on April 14, 2019. © 1995 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from

1995;4:353-358. Cancer Epidemiol Biomarkers Prev M S Kurzer, J W Lampe, M C Martini, et al. women consuming flaxseed powder.Fecal lignan and isoflavonoid excretion in premenopausal
Updated version
http://cebp.aacrjournals.org/content/4/4/353
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cebp.aacrjournals.org/content/4/4/353To request permission to re-use all or part of this article, use this link
on April 14, 2019. © 1995 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from













![Excretion [2015]](https://img.pdfslide.us/doc/110x75/55d39c87bb61eb05278b46dd/excretion-2015-55d47f0693bf7.jpg)















