Embed Size (px)
Citation preview

Fast Activation of Dihydropyridine-sensitive Calcium Channels of
Skeletal Muscle
Multiple Pathways of Channel Gating
JIANJIE MA,* ADOM GONZ.i.LEZ,+* a n d R u t CHEN*
Front the *Department of Physiology and Biophysics, Case Western Reserve University School of Medicine, Cleveland, Ohio 44106; and ,*Department of Molecular Biophysics and Physiology~ Rush Medical College, Chicago, Illinois 60612
ABSTRACT Dihydropyridine (DHP) receptors of the transverse tubule membrane play two roles in excitation- contraction coupling in skeletal muscle: (a) they function as the voltage sensor which undergoes fast transition to control release of calcium from sarcoplasmic reticulum, and (b)they provide the conduct ing unit of a slowly acti- vating L-type calcium channel. To unders tand this dual function of the DHP receptor, we studied the effect of de- polarizing condit ioning pulse on the activation kinetics of the skeletal muscle DHP-sensitive calcium channels re- constituted into lipid bilayer membranes. Activation of the incorporated calcium channel was imposed by depo- larizing test pulses f rom a holding potential of - 8 0 mV. The gating kinetics of the channel was studied with ensemble averages of repeated episodes. Based on a first latency analysis, two distinct classes of channel openings occurred after depolarization: most had delayed latencies, distributed with a mode at 70 ms (slow gating); a small number of openings had short first latencies, <12 ms (fast gating). A depolarizing condit ioning pulse to +20 mV placed 200 ms before the test pulse ( - 1 0 mV), led to a significant increase in the activation rate of the ensemble averaged-current; the time constant o f activation went from "fin = 110 ms (reference) to T,,~ = 45 ms after condi- tioning. This enhanced activation by the condit ioning pulse was due to the increase in frequency of fast open events, which was a steep function of the intermediate voltage and the interval between the condit ioning pulse and the test pulse. Additional analysis demonstrated that fast gating is the property of the same individual chan- nels that normally gate slowly and that the channels adopt this property after a sojourn in the open state. The rapid secondary activation seen after depolarizing prepulses is not compatible with a linear activation model for the calcium channel, but is highly consistent with a cyclical model. A six-state cyclical model is proposed for the DHP-sensitive Ca channel, which pictures the normal pathway of activation of the calcium channel as two voltage- dependen t steps in sequence, plus a voltage-independent step which is rate limiting. The model reproduced well the fast and slow gating modes of the calcium channel, and the effects of condit ioning pulses. It is possible that the voltage-sensitive gating transitions of the DHP receptor, which occur early in the calcium channel activation sequence, could underlie the role of the voltage sensor and yield the rapid excitation-contraction coupling in skel- etal muscle, through either electrostatic or allosteric linkage to the ryanodine receptors/calcium release chan- nels. Key words: excitation-contraction coupling * signal transduction �9 sarcoplasmic reticulum
I N T R O D U C T I O N
In skeletal muscle, the p roduc t s o f a single gene per- fo rm two di f ferent roles. T he d ihydropyr id ine (DHP) 1 r ecep to r polypept ide o f the transverse tubule (TT) m e m b r a n e consti tutes the pore fo rm i ng cq subuni t o f the vol tage-dependent Ca channe l (Tanabe et al., 1987). However, the D H P receptors work mainly as sensors o f
Portions of this work have previously appeared in abstract torm (Ma, J., M.M. Hosey, and E. Rios. 1992. Biophys. J. 61:A130; Ma, J., and E. Rios. 1993. Biophys.J. 64:A376; and (;hen, R., andJ . Ma 1995. Biophys. J. 68:A373).
Address correspondence to Dr. Jianjie Ma, Depar tment of Physiol- ogy and Biophysics, Case Western Reserve University, School of Med- icine, Cleveland, OH 44106. Fax: 216-368-5586.
~ Abbreviations used in this paper. DHP, dihydropyridine; E-C, excita- tion-contraction; Ic~, calcium current; SR, sarcoplasmic reticulum; TT, transverse tubule.
TT m e m b r a n e voltage, cont ro l l ing o p e n i n g o f the Ca release channels in the sarcoplasmic re t icu lum (SR) (Rfos and Brum, 1987; Tanabe et al., 1988). These two funct ions have strikingly di f ferent kinetics. In response to m e m b r a n e depolar iza t ion the voltage sensors gener- ate in t ramembrane charge movemen t (Schneider and Chandler , 1973), which is fol lowed by release o f cal- c ium f rom the SR. The whole process f rom excitat ion (depolar izat ion o f TT m e m b r a n e and m o v e m e n t o f in- t r a m e m b r a n e charge) to o p e n i n g o f Ca release chan- nels occurs in < 5 ms (Simon and Schneider , 1988), while activation o f the TT m e m b r a n e Ca cu r ren t (Ic,) is general ly very slow, having a t ime cons tan t o f > 1 0 0 ms at - 10 mV in f rog skeletal muscle (Avila-Sakar et al., 1986). In addit ion, the m o v e m e n t o f i n t r a m e m b r a n e charge associated with the voltage sensor is c en t e r ed at ~ - 4 0 mV, while activation o f Ica by depolar iza t ion is c en t e r ed at N0 mV (Pizarro et al., 1988).
221 J. GEN. Pn'~'slol.. �9 The Rockefeller University Press �9 0022-1295/96/09/221/12 $2.00 Volume 108 September 1996 221-232
Dow
nloaded from http://rupress.org/jgp/article-pdf/108/3/221/1144134/221.pdf by guest on 30 N
ovember 2021

The DHP-sensit ive Ca c h a n n e l a n d the voltage sensor of exc i ta t ion-cont rac t ion (E-C) coup l ing are e n c o d e d by the same cDNA (Tanabe et al., 1988; Adams et al., 1990). The same DHP recep tor molecu le could have
bo th funct ions . Alternatively, the two func t ions could be the role of two d i f fe rent molecules e n c o d e d by the same gene . A form of this hypothesis, p roposed by De
J o n g h et al. (1989), was ru led ou t by Beam et a1.(1992) who d e m o n s t r a t e d that the C O O H - t e r m i n a l t r unca t ed DHP receptor could f u n c t i o n as bo th voltage sensor a n d Ca channe l .
To serve as an E-C coup l ing voltage sensor, the DHP receptor mus t be capable of fast conformat iona l changes in response to m e m b r a n e depolar iza t ion . Fe ldmeyer et
al. (1990) a n d Garcia et a1.(1990) showed that it is possi- ble to achieve fast activation of the "slow" Ic~ in frog skeletal muscle, by apply ing a depo la r i z ing cond i t ion-
ing pulse before the test pulse. The cond i t i on ing - i n d u c e d fast ga t ing had a steep d e p e n d e n c e on the in- t e rmedia te potent ia l be tween the c o n d i t i o n i n g a nd test
pulses (Feldmeyer et al. 1992). Work ing o n myoballs f rom rat skeletal muscle , Scu lp to r eanu et al. (1993)
showed that the L-type Ca c u r r e n t could be po ten t i a t ed by trains of depola r iz ing prepulses. The po ten t i a t i on r equ i r ed rapid phosphory la t ion by c A M P - d e p e n d e n t p ro t e in kinase in a vo l t age -dependen t m a n n e r , which shifted to the left the vo l t age -dependence of, a n d accel-
e ra ted the rate of, c h a n n e l activation. Melzer a n d co-
workers (Fe ldmeyer et al. 1990) suggested that the fast t rans i t ion u n d e r l y i n g fast act ivation of Ic~ is involved in the rapid con t ro l of Ca release f rom the SR.
In the p resen t paper , single c h a n n e l r econs t i tu t ion t echn iques are c o m b i n e d with macroscopic measure- ments in muscle fibers to decide whe the r the dual ki-
netics are a p roper ty of indiv idual molecules . Us ing DHP receptors i nco rpo ra t ed as Ca channe l s in p l ana r bilayers, it is d e m o n s t r a t e d that the fast t rans i t ion is a proper ty of the indiv idual channe l s tha t normal ly gate
slowly, thus c o n f i r m i n g the i n t e rp re t a t i on of Fe ldmeyer et al. (1990). Record ing Ic~ in frog muscle fibers, it is f o u n d that the t r ans fo rmat ion to fast ga t ing is accom- p a n i e d by changes in voltage d e p e n d e n c e an d in rate and ex ten t of decay after the peak. We propose a mo- lecular d iagram that incorpora tes the two func t ions of
the DHP receptor : as fast E-C c o u p l i n g voltage sensor and slow vo l t age -dependen t Ca channe l .
M A T E R I A L S A N D M E T H O D S
Planar Bilayer Reconstitution of the DHP-sensitive Ca Channels
Planar bilayer membranes were formed across an aperture of ~200 Ixm diameter with a lipid mixture of phosphatidylethanol- amine/phosphatidylserine/cholesterol in a ratio of 1:1:0.2 dis- solved in decane at a concentration of 50 mg/ml. The cis (intra- cellular) solution contained 200 mM KC1, 3 mM Mg-ATP, 10 mM HEPES-Tris, pH 7.4; the trans (extracellular) solution contained 50 mM NaC1, 100 mM BaC12, 10 mM Hepes-Tris, pH 7.4. To have consistently measurable Ca channel activities, a DHP agonist, Bay K 8644 was always present in the cis solution at a concentration of 0.3 jxM (see Figs. 1 and 2) or 1 IxM (see Figs. 4 and 5).
To incorporate the DHP-sensitive Ca channels, TT vesicles (1-3 txl) were added to the cis solution. The bilayer was initially held at -80 mV (holding potential). The Ca channel activity was mea- sured with depolarizing pulses of various amplitudes and dura- tions. Single channel currents were recorded with either an Axo- patch-lC or an Axopatch 200A unit (Axon Instruments, Foster City, CA), and filtered at 100 Hz using an 8 pole Bessel filter. The experiments were carried out at room temperature (22-24~
The data presented were done at two different stages: from 1990 to 1992 at Rush Medical College (see Figs. 1, 2, 3, and 8) and from 1992 to 1994 at Case Western Reserve University (see Figs. 4 and 5).
Analysis of Single Channel Data
Processing of the current episodes started with construction of controls, by averaging the null sweeps. The controls, which con- tain only the linear capacitative and electrostrictive components, were subtracted from the test currents, yielding asymmetric cur- rent records. Conventional single channel analysis was done with pCLAMP software (Axon Instruments) and custom programs. Different pulse protocols were used to study the kinetics of activa- tion and inactivation. Ensembles of ionic currents obtained with pulses to the same voltage were averaged; the averaged currents were fitted with the m:~h time dependence. The procedures have been described in detail elsewhere (Ma et al., 1991).
Measurements of Macroscopic Ca Current
Macroscopic Ca currents (Ic~) were recorded in segments of sin- gle fibers from the semitendinosus muscle of the frog Rana pipi- ens, mounted at slack length and voltage clamped in a double Va- seline gap chamber. The methods have been described in detail elsewhere (Gonzalez and Rios, 1993). The fibers were held polar- ized at a potential of -80 mV. The currents shown are asymmetric, obtained as differences of currents during test pulses and control pulses, usually between - 110 and -90 mV. The internal solution contained (in mM): 10 BAPTA, 1 Ca, 5 ATP, 135 Cs, 88 glutamate, 5.5 MgC12, 5 creatine phosphate, 5 glucose, 10 HEPES, pH 7.0. The high concentration of BAPTA was designed to prevent fiber contraction and to eliminate potential feedback of SR Ca release on the DHP-sensitive Ca channel (Gonzalez and Rios, 1993). 3 mM Ba in the external solution was the main carrier for current through the DHP-sensitive Ca channel. Tile external solutions contained, in addition, 135 CH~SO:~, 124 TEA, 0.001 TTX, 1,4 diaminopyri- dine, 5 Tris maleate, pH 7.0. The temperature was 10-12~
Isolation of Transverse Tubule Membranes from Rabbit Skeletal Muscle
Transverse tubule membranes were isolated from frozen rabbit skeletal muscle tissue following the procedures of Galizzi et at. (1984). These membrane vesicles contained 20-60 pmol DHP re- ceptor/rag protein in a ['~H]PN200-110 binding assay. The vesi- cles were stored at -80~ at a protein concentration of 5-7 mg/ml.
R E S U L T S
Reconstituted Ca Channels Are Strictly Voltage- and Time-dependent
Isolated T T m e m b r a n e vesicles f rom rabbi t skeletal muscle were fused with p l ana r l ipid bilayers. To study
222 Calcium Channels of Skeletal Muscle
Dow
nloaded from http://rupress.org/jgp/article-pdf/108/3/221/1144134/221.pdf by guest on 30 N
ovember 2021
A -lO mV C - - - J - 8 0 mV 2 s L 20
15
t"- i
~ ,~ ~ . ~ ~ - - . ~ - . ~ - - , ~ . ~ , . - ~ . ~ , v (1)
L- 1() I
t t rO 0 , 0
f 5
I~ , () .15 pal3 pA ~ I
10
First
IIII ,I,I ,, 1 0 0 1 0 0 0
Latency (ms)
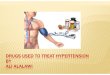
FIGURE 1. Gating kinetics of the DHP-sensitive Ca channels in bi- layers. (A) After fusion of vesi- cles, the bilayer was held polar- ized at -80 inV. Channel activity was elicited by depolarizing pulses to -10 mV. Represented are consecutive episodes, ob- tained at 7-s intervals. Channel openings were completely blocked by 20 p,M nitrendipine added to the extracellular solu- tion (data not shown). (B) 21 separate bilayer experiments were used in the ensemble aver- age. A total of 478 episodes as in A were included. Blanks, which constituted 25.6% (2.5% SEM) of sweeps, were excluded from
the average,. The "smooth" line is the best fit curve I =/,,,~,~ • m ~ • h, where m = 1 - exp(- t/'rm) and h = h~ + (h0 - h~) • exp(- t/'rh). h~ was set to 0, since the channel inactivates completely at - 10 mV with longer pulses (Ma et al., 1991 ). The best fit parameters were Imp, = -0.245 pA (0.006), %, = 118 ms (4), and "ra = 1.24 s (0.04). (C) First latency histogram of channel openings at - 10 inV. The histogram was generated from eight experiments with a total of 280 episodes. The first two blank bins (4 ms) in the histogram correspond to the dead time of the system (due to Bessel filtering at 100 Hz). The two populations, fast and delayed open events, are clearly represented.
the vo l t age -dependen t gat ing kinetics o f the DHP-sensi- tive Ca channel , the bilayer was kept polar ized at - 8 0 mV, and the cu r r en t carr ier (100 mM Ba 2+) was presen t only in the extracel lular solut ion (Ma et al., 1991). Inward Ba currents t h r o u g h the channe l were r e c o r d e d u n d e r s t imulat ion by depolar iz ing test pulses. Records o f consecut ive single channe l episodes are shown in Fig. 1 A. T h e channels were closed at - 8 0 mV and o p e n e d when the m e m b r a n e was depolar ized (by pulses o f 2 s dura t ion to - 1 0 mY). T he identi ty o f the channels was certain because open ing was highly depen- den t on the presence o f Bay K 8644 and was preven ted by the addi t ion o f n i t r end ip ine (Ma et al., 1991).
T h e average o f a large n u m b e r o f episodes at test po- tential - 1 0 mV displays a slow time course o f activation followed by inactivation (Fig. 1 B), that can be ade- quately fit ted by the equat ion:
3 I = I,. .xXm x h , (1)
where m and h are first o rde r time- and vol tage-depen- den t activation and inactivation variables. T he best fit activation t ime cons tan t Tm was 118 ms (SE o f estimate, 4 ms) and the inactivation t ime cons tan t Ta was 1.24 s (0.04 s). The o p e n probabil i ty o f the recons t i tu ted Ca channe l increased steeply with voltage, fol lowing a B o h z m a n n dis t r ibut ion cen t e r ed at - 2 5 mV with a s teepness factor o f 7 mV and a m a x i m u m o p e n proba- bility o f ~ 1 0 % (Ma et al., 1991). These proper t ies are similar to those o f Ic~ in the skeletal muscle o f the f rog at 15~ (Avila-Sakar et al., 1986). T he kinetics o f activa- t ion in rat muscle fibers, however, follow a single expo- nent ial func t ion o f t ime (Mejia-Alvarez et al., 1991).
2 2 3 M A E T A L .
This can be reconci led since the ensemble average cur- rents in the present exper iments (illustrated in Fig. 1 B) can be fitted a lmost equally well by the d e p e n d e n c e Ima x X m • h (see Fig. 4).
To examine the kinetics o f openings at the single chan- nel level, a h is togram o f first latency was cons t ruc ted (Fig. 1 C). The f igure plots the absolute f requency (oc- currences) o f the first de tec ted o p e n event vs. t ime o f the first t ransit ion (first latency). T h e r e are two well de- f ined g roups o f openings : a small n u m b e r o f fast o p e n events, whose shor t latencies are con t a ined in the first five bins, and a large g r o u p o f openings , with a b road range of latency values, distributed with a peak at ~ 7 0 ms.
The present exper iments are di f ferent f rom previous bilayer studies, where the Ca channe l activities were measured at cons tan t ho ld ing potent ials (Affolter and Coronado , 1985; Ma and Coronado , 1988; Mejia-Alva- rez et al., 1991). U n d e r the presen t condit ions, no per- sistent channe l activity could have b e e n measu red at ma in ta ined depolar iz ing potentials because o f voltage- d e p e n d e n t inactivation. In a few instances (18 ou t o f 180 exper iments ) , we were able to r ecord channels that did no t inactivate at ma in ta ined depolar iz ing poten- tials; interestingly, these channels did no t show any vol tage-dependence at all, since channe l activities could be measured at - 8 0 mV (data no t shown).
A Depolarizing Conditioning Pulse Accelerates the Rate of Channel Activation
As observed by Fe ldmeyer et al. (1990) on macroscop ic Ca currents , the activation o f the single Ca channels in
Dow
nloaded from http://rupress.org/jgp/article-pdf/108/3/221/1144134/221.pdf by guest on 30 N
ovember 2021

A +20 i-- -10 - 8 o 1=4o[ ~.5 s k
I
I
I
I I 3 pA [0.15 pA
- 1 0 D -40 I -801 L ~..~;:..%,,~.:~.#r ; ~ ~
FIGURE 2. Effects of conditioning pulse on activation of the Ca channels. (A) Consecutive "test-minus-control" current episodes obtained at 10-s intervals with the pulse protocol illustrated. Condi- tioning pulses to +20 mV had a duration of 200 ins. The potential between conditioning and test pulses was -40 mV. (B) A total of 278 episodes from five separate bilayer experiments were used in the ensemble average. The smooth curve was generated as in Fig, 1, with paimneters: I ..... = -0.211 pA (0.003), % = 43.5 ms (2), % = 1.54 s (0.07). (C) Ensemble average of the fast open events. Only those episodes that exhibited openings with latencies of 12 ms or less after a conditioning pulse to +20 mV were included; these are a subset of the episodes averaged in B. The smooth cm-ve is the best tit exponential decay to 0, with % = 0.85 s (0.04). 54 episodes were averaged. The peak cnrrent in the ensemble average corre- sponds to a peak open probability of 53%. (D) Ensemble average of currents obtained with the protocol shown, in which the depo- larization to +20 mV was replaced by a conditioning pulse to a subthreshold voltage ( -40 mV). Four bilayers and 232 episodes were used in tim average. The best fit parameters were: I ...... = -0.193 pA (0.007), %~ = 128 ms (5), % = 1.25 s (0.06).
bi layers was m a d e faster by a d e p o l a r i z i n g c o n d i t i o n i n g pulse. A d o u b l e pu lse p r o t o c o l (Fig. 2 A), was a p p l i e d to the b i layer system. Consecu t ive s ingle c h a n n e l cur- r en t ep i sodes were o b t a i n e d with a test pulse to - 1 0 mV p r e c e d e d by a c o n d i t i o n i n g pu lse to + 2 0 mV a n d are r e p r e s e n t e d in the f igure. T h e c o r r e s p o n d i n g en-
s emb le average is shown in B. T h e act ivat ion o f the c h a n n e l was m u c h faster af ter the c o n d i t i o n i n g pu lse ( c o m p a r e Figs. 1 B a n d 2 B). T h e c o n d i t i o n i n g d e p o l a r - iza t ion r e d u c e d %1 a lmos t t h r e e f o l d to 44 ms (SE = 2 ms, s t a n d a r d e r r o r o f es t imate ) . In cont ras t , inactiva- t ion was n o t c h a n g e d great ly (% = 1.5 s, SE = 0.1 s).
T h e e n h a n c e d ra te o f c h a n n e l ac t ivat ion strictly re- q u i r e d the la rge c o n d i t i o n i n g pulse , as a s u b t h r e s h o l d c o n d i t i o n i n g d e p o l a r i z a t i o n was n o t sufficient . T h e en- s emb le average shown in Fig. 2 D was e l ic i ted by a p ro- tocol tha t i n c l u d e d a p r e p u l s e d e p o l a r i z a t i o n to - 4 0 mV ins t ead o f a pulse to + 2 0 mV; clearly, the c h a n n e l s sub jec t ed to this p r o t o c o l d i d n o t act ivate m o r e rap id ly than the r e f e r e n c e (Fig. 1 B).
E x a m i n a t i o n o f the ind iv idua l r eco rds in Fig. 2 A sug- gests tha t the inc rease in the overal l ra te o f ac t iva t ion is largely d u e to the a p p e a r a n c e o f a g r o u p o f ep i sodes in which channe l open ings occu r red almost ins tantaneously u p o n depo l a r i za t i on . This g r o u p is i l lus t ra ted by the th i rd , the ten th , a n d p e r h a p s the fou r th c u r r e n t t race in Fig. 2 A.
T h e effect o f the c o n d i t i o n i n g pu lse on the d is t r ibu- t ion o f first l a tenc ies was i l lus t ra ted in Fig. 3. T h e histo- g r a m in Fig. 3 A was de r i ve d f r o m r e c o rds sub jec ted to a s u b t h r e s h o l d c o n d i t i o n i n g pu lse to - 4 0 mV. This his- t o g r a m was n o t very d i f f e ren t f rom the o n e shown in Fig. 1 C; in bo th cases, the p r o p o r t i o n o f fast o p e n events was a p p r o x i m a t e l y 7%. T h e h i s t o g r a m in Fig. 3 B, of o p e n i n g s af ter a c o n d i t i o n i n g pulse to + 20 mV, is sharp ly d i f ferent . T h e fast o p e n events b e c a m e m o r e f r equen t , a n d a b o u n d a r y be tween sho r t a n d l ong la- tencies cou ld be c lear ly l oc a t e d at 12 ms. T h e o p e n i n g s with la tenc ies u n d e r 12 ms, c o n t a i n e d in the first five b ins o f the h i s togram, wen t f ront 7 to 24%.
Fast and Slow Gating Are Properties of the Same Channel
T h e g r o u p o f fast o p e n i n g s assoc ia ted with the cond i - t i on ing pu lse m i g h t r e p r e s e n t a fast ga t ing m o d e o f an ind iv idua l molecu le , c a pa b l e o f slow ga t ing at o t h e r t imes, o r the r e c r u i t m e n t by the c o n d i t i o n i n g pulse o f a d i f f e r en t c h a n n e l tha t is s i len t in r e f e r e n c e cond i t ions . T h a t e n s e m b l e averages in Figs. 1 B a n d 2 B r e a c h e d s imi lar p e a k values a n d were a lmos t s u p e r i m p o s a b l e in the i r la te t ime course was cons i s t en t with a c h a n g e in p r o p e r t i e s o f the same channe l . T h e fo l lowing two ad- d i t iona l tests c o n f i r m e d tha t the two ga t ing rates a re p r o p e r t i e s o f the same ind iv idua l molecu le .
First, the inac t iva t ion kinet ics o f the fast ac t ivat ing events was d e t e r m i n e d . This was d o n e by co l l ec t ing all ep i sodes with first l a tenc ies u n d e r 12 ms a n d c o m p u t - ing the i r e n s e m b l e average ( the "cond i t iona l " average, shown in Fig. 2 C). By se lec t ing all sweeps with fast open ings , we fo r ced the ini t ia l o p e n p robab i l i t y to be close to unity. I f these sweeps r e p r e s e n t e d cu r r en t s t h r o u g h a newly r e c r u i t e d r a p i d channe l , t hen the reg-
224 Calcium Channels of Skeletal Muscle
Dow
nloaded from http://rupress.org/jgp/article-pdf/108/3/221/1144134/221.pdf by guest on 30 N
ovember 2021
A 4O
B 40
�9 (D c-
�9 k._ k_
LP
0
0
30
20
10
10
I J [
Ihl,,ll 100 1000
First
30
20
10
o
Latency
I I ,i,. 10 1 O0 1 0 0 0
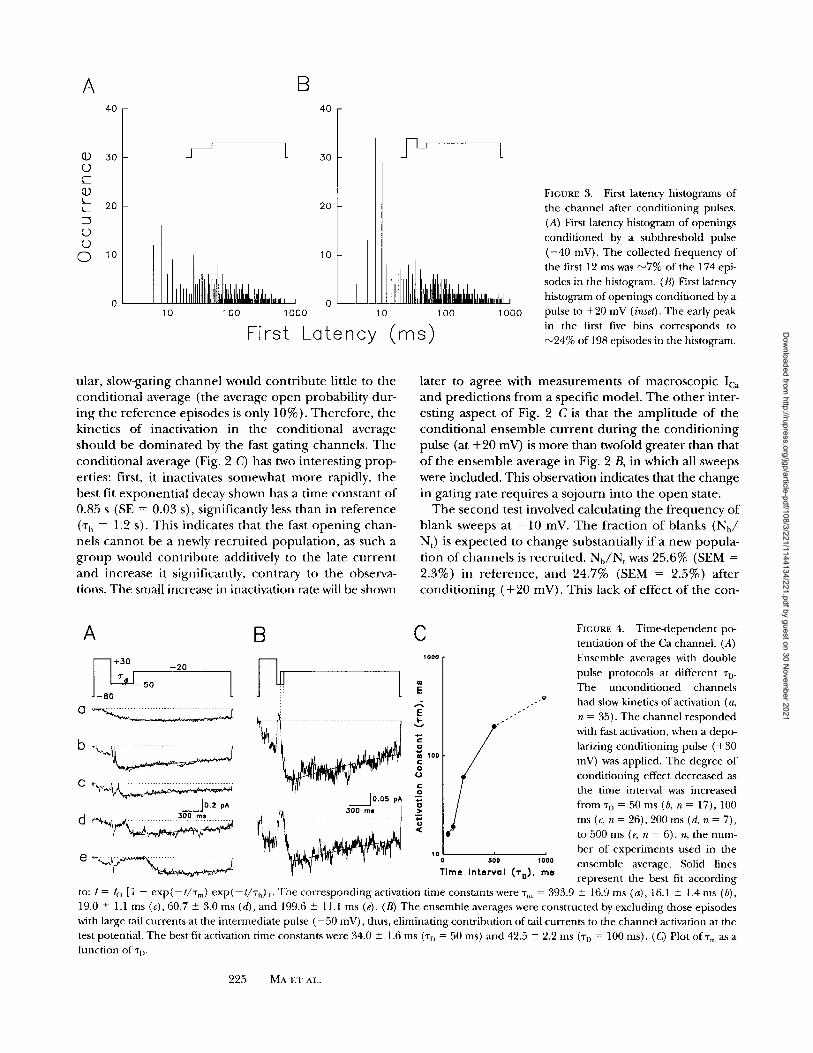
FIGURE 3. First latency histograms of the channel after conditioning pulses. (A) First latency histogram of openings conditioned by a subthreshold pulse ( - 4 0 mV). The collected frequency of the first 12 ms was ~7% of the 174 epi- sodes in the histogram. (B) First latency histogram of openings conditioned by a pulse to +20 mV (inset). The early peak in the first five bins corresponds to ~24% of 198 episodes in the histogram.
u la r , s l ow-ga t ing c h a n n e l w o u l d c o n t r i b u t e l i t t le to t h e
c o n d i t i o n a l a v e r a g e ( the a v e r a g e o p e n p r o b a b i l i t y d u r -
i n g t h e r e f e r e n c e e p i s o d e s is o n l y 1 0 % ) . T h e r e f o r e , t h e
k ine t i c s o f i n a c t i v a t i o n in t h e c o n d i t i o n a l a v e r a g e
s h o u l d be d o m i n a t e d by t h e fast g a t i n g c h a n n e l s . T h e
c o n d i t i o n a l a v e r a g e (Fig. 2 C) has two i n t e r e s t i n g p r o p -
er t ies : first, i t i nac t iva te s s o m e w h a t m o r e rap id ly , t h e
bes t f i t e x p o n e n t i a l d e c a y s h o w n has a t i m e c o n s t a n t o f
0.85 s (SE = 0.03 s), s i gn i f i can t ly less t h a n in r e f e r e n c e
('r h = 1.2 s). T h i s i n d i c a t e s t h a t t h e fast o p e n i n g c h a n -
ne l s c a n n o t be a newly r e c r u i t e d p o p u l a t i o n , as s u c h a
g r o u p w o u l d c o n t r i b u t e add i t i ve ly to t h e la te c u r r e n t
a n d i n c r e a s e it s ign i f ican t ly , c o n t r a r y to t h e obse rva -
tions. T h e smal l inc rease in inac t iva t ion ra te will be shown
l a t e r to a g r e e wi th m e a s u r e m e n t s o f m a c r o s c o p i c Ica
a n d p r e d i c t i o n s f r o m a spec i f i c m o d e l . T h e o t h e r in t e r -
e s t i n g a s p e c t o f Fig. 2 C is t h a t t h e a m p l i t u d e o f t h e
c o n d i t i o n a l e n s e m b l e c u r r e n t d u r i n g t h e c o n d i t i o n i n g
pulse (at + 2 0 mV) is m o r e t h a n twofold g r e a t e r t h a n tha t
o f t h e e n s e m b l e a v e r a g e in Fig. 2 B, in w h i c h all sweeps
were i n c l u d e d . Th is obse rva t ion ind ica tes tha t t he c h a n g e
in g a t i n g r a t e r e q u i r e s a s o j o u r n i n t o t h e o p e n state .
T h e s e c o n d tes t i n v o l v e d c a l c u l a t i n g t h e f r e q u e n c y o f
b l a n k sweeps at - 1 0 mV. T h e f r a c t i o n o f b l a n k s ( N b /
N,) is e x p e c t e d to c h a n g e subs tan t i a l ly i f a n e w p o p u l a -
t i on o f c h a n n e l s is r e c r u i t e d . N b / N t was 25 .6% (SEM =
2 .3%) in r e f e r e n c e , a n d 24 .7% (SEM = 2 .5%) a f t e r
c o n d i t i o n i n g ( + 2 0 m V ) . T h i s l ack o f e f f e c t o f t h e c o n -
A B C
_•80 +30 -20 ~ , j
I 0 . 2 pA 0.05 pA
E
o 100
0 U
. v
10 500 1000
T i m e I n t e r v a l . .('~',~J, m e
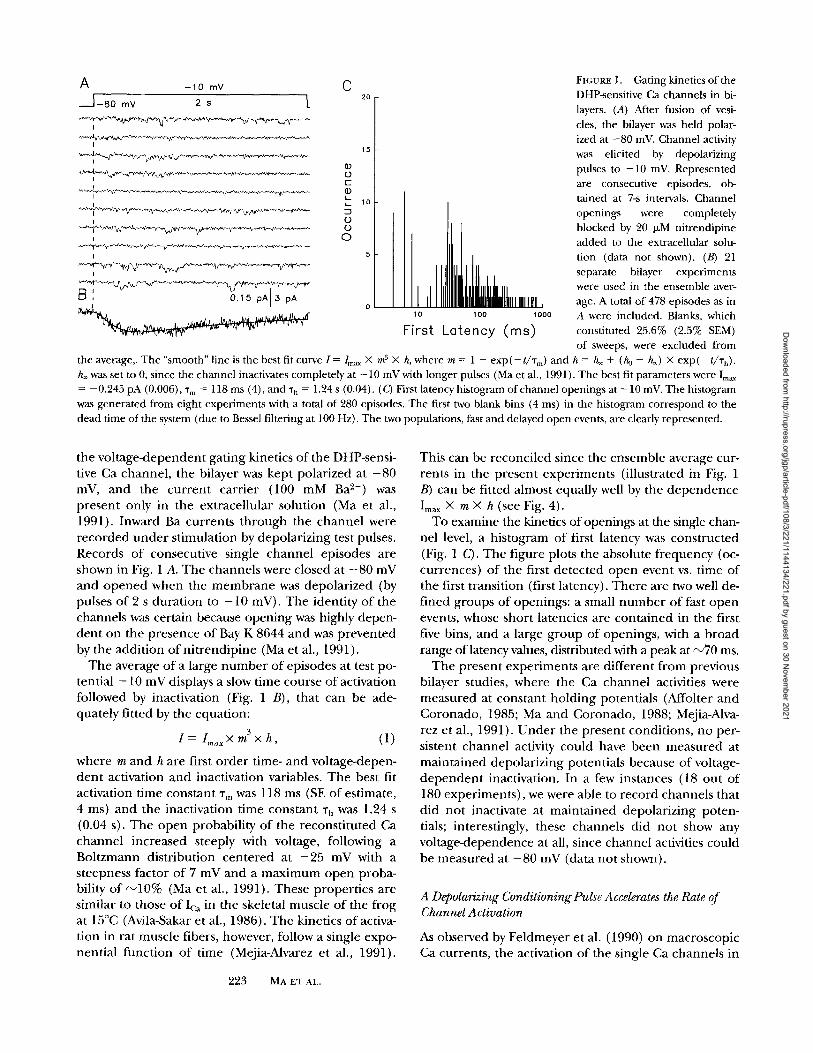
FIGURE 4. Time-dependent po- tentiation of the Ca channel. (A) Ensemble averages with double pulse protocols at different "rD. The unconditioned channels had slow kinetics of activation (a, n = 35). The channel responded with fast activation, when a depo- larizing conditioning pulse (+ 30 mV) was applied. The degree of conditioning effect decreased as the time interval was increased from ~D = 50 ms (b, n = 17), 100 ms (c, n = 26), 200 ms (d, n = 7), to 500 ms (e, n = 6). n, the num- ber of experiments used in the ensemble average. Solid lines represent the best fit according
to: I = /o [1 - exp(--t/Tm) exp(--t/Th)]. The corresponding activation time constants were "fin = 393.9 _+ 16.9 ms (a), 16.1 + 1.4 ms (b), 19.0 _+ 1.1 ms (c), 60.7 _+ 3.0 ms (d), and 199.6 _+ 11.1 ms (e). (B) The ensemble averages were constructed by excluding those episodes with large tail currents at the intermediate pulse ( - 5 0 mV), thus, eliminating contribution of tail currents to the channel activation at the test potential. The best fit activation time constants were 34.0 _+ 1.6 ms (-r n = 50 ms) and 42.5 _+ 2.2 ms ('r D = 100 ms). (Q Plot ofT,,, as a function of "q).
2 2 5 MA ET AL.
Dow
nloaded from http://rupress.org/jgp/article-pdf/108/3/221/1144134/221.pdf by guest on 30 N
ovember 2021

ditioning pulse o n N b / N t indicates that the n u m b e r of active channels did not change.
Taken together, the results suppor t the interpreta- tion of Feldmeyer et al. (1990) that the fast gating is a proper ty of the same molecules that normally gate slowly and show that the channels adopt this proper ty after dwelling in the open state.
Voltage and Time-dependent Potentiation of the Ca Channel
Fast gating of the slow Ica, o r potent iat ion of the L-type Ca current in skeletal muscle, is a steep function of the intermediate voltage between the condit ioning and test pulses, as shown in the studies of Feldmeyer et al. (1992) and Sculptoreanu et al. (1992, 1993). The recovery pro- cess, which underl ines transition f rom fast to slow gat- ing, became faster as the intermediate potential was more negative. To fur ther unders tand the fast gating kinetics of the DHP-sensitive Ca channel, the following two series of exper iments were per formed.
First, the interval between the condit ioning and test pulses (%) was varied, to study the time dependence of the condit ioning effect. The pulse protocol is shown in Fig. 4 A. Here , the channel was condi t ioned to +30 mV for 250 ms, followed by an intermediate potential at - 5 0 mV for different durations (%), then tested to a potential o f - 2 0 mV. The ensemble average of the con- trol channels had slow kinetics of activation (% = 393.9 - 16.9 ms), which became faster after the condit ioning pulse. Clearly, the degree of fast activation decreased as % was increased f rom 50 to 500 ms.
Due to the slow deactivation kinetics of the channel at - 5 0 mV, probably as a result of the higher concen- tration of Bay K 8644 being used (1 ~zM, instead of 0.3 IzM as in Figs. 1 and 2), the actual activation kinetics of the current during the test pulse could not be deter- mined directly f rom Fig. 4 A, at % = 50 and 100 ms. Thus, new ensemble averages were constructed by ex- cluding those episodes with large tail currents at the in- termediate pulse ( - 5 0 mV) (Fig. 4 B). By doing so, we el iminated the contr ibut ion of tail currents to the channel current at the test potential. This selected en- semble average also activated with fast kinetics, suggest- ing that most of the fast open events were not due to in- complete deactivation, ra ther they were genera ted in response to the test pulse ( - 2 0 mV).
The ensemble averages of the test currents at - 2 0 mV were fitted with first order kinetics of activation and inactivation, f rom which the time constants of activa- tion (%) were obtained. From the plot of % vs. ~'D (Fig. 4 C), it can be seen clearly that the changes in gating rate of the Ca channel had a steep dependence on the interval between the condit ioning and test pulses. Es- sentially, the condit ioning effect became negligible when "r D was >300 ms.
Second, the ampli tude of the intermediate pulse was
changed f rom - 5 0 to - 1 2 0 mV, to study the depen- dence of the condit ioning effect on the intervening voltage (Fig. 5). With this hyperpolarization, the condi- t ioning effect was essentially eliminated. Rarely, the channels opened with short first latencies, judging f rom the individual episodes shown in Fig. 5 A, and the condi t ioned channel activated as slowly as the uncondi- t ioned channel. The time constant of activation after conditioning, 215.2 + 4.6 ms (Fig. 5 B), was not very different f rom the control channel (Fig. 4 A).
These results show that fast gating of the DHP-sensi- tive Ca channel is de te rmined by both the condit ioning pulse and the intervening voltage between the condi- t ioning and test pulses.
Properties of the Fast-activating Ca Current in Frog Muscle
As will be seen in the DISCUSSION, when quantitative simulations are a t tempted with a model presented for
+ 3 0 mV
- 20 mv L
-1 20 mV
I
t
I I
I
]
!
I !
I I
I
I I
300 msl2 pA
FICURE 5. Voltage-dependent potentiation of Ca channel. (A) Consecutive episodes were represented with the pulse protocol shown in the top. The intermediate voltage was V D = -120 mV. (B) The ensemble average contained 182 episodes obtained from 16 separate experiments. The best fit activation time constant was %1 = 215 -4:- 4.6 ms.
226 Calcium ChanneLs of Skeletal Muscle
Dow
nloaded from http://rupress.org/jgp/article-pdf/108/3/221/1144134/221.pdf by guest on 30 N
ovember 2021
_80/ - 2 0
V
[ 1.5 s
_..K
-L
+70
v
201 1.5 s - 8 0
- -10 mV
0 mV
+15 mV
+35 mV
11 A/F
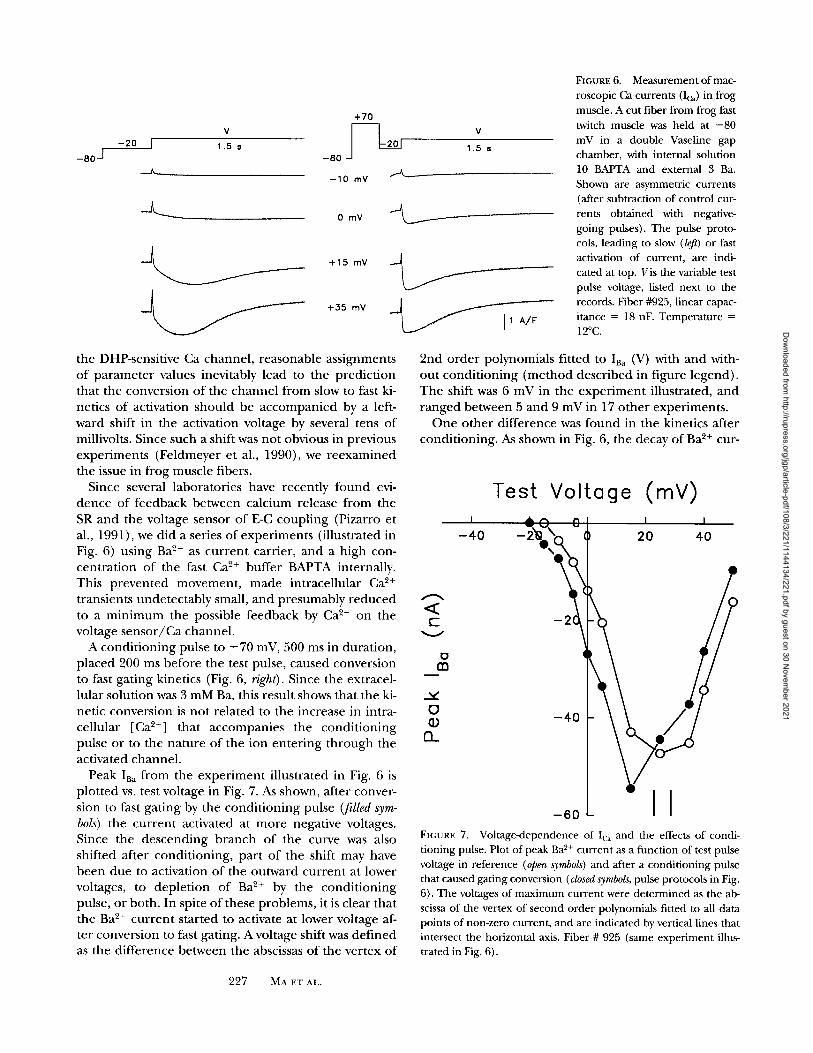
FIGURE 6. Measurement of mac- roscopic Ca currents (Ic~) in frog muscle. A cut fiber from frog fast twitch muscle was held at -80 mV in a double Vaseline gap chamber, with internal solution 10 BAPTA and external 3 Ba. Shown are asymmetric currents (after subtraction of control cur- rents obtained with negative- going pulses). The pulse proto- cols, leading to slow (/eft) or fast activation of current, are indi- cated at top. Vis the variable test pulse voltage, listed next to the records. Fiber #925, linear capac- itance = 18 nF. Temperature = 12~
the DHP-sensitive Ca channel , reasonable ass ignments o f pa r ame te r values inevitably lead to the predic t ion that the convers ion o f the c ha nne l f rom slow to fast ki- netics o f activation should be a c c o m p a n i e d by a left- ward shift in the activation voltage by several tens o f millivolts. Since such a shift was no t obvious in previous exper iments (Feldmeyer et al., 1990), we r eexamined the issue in f rog muscle fibers.
Since several laborator ies have recent ly f o u n d evi- d e n c e o f feedback be tween calc ium release f rom the SR and the voltage sensor o f E-C coup l ing (Pizarro et al., 1991), we did a series o f exper iments (illustrated in Fig. 6) using Ba 2+ as cu r r en t carrier, and a h igh con- cen t ra t ion o f the fast Ca ~+ buffer BAPTA internally. This p reven ted movemen t , m a d e intracel lular Ca 2+
transients unde tec tab ly small, and presumably r e d u c e d to a m i n i m u m the possible feedback by Ca 2+ on the voltage s e n s o r / C a channel .
A cond i t i on ing pulse to + 70 mV, 500 ms in dura t ion, p laced 200 ms before the test pulse, caused convers ion to fast gat ing kinetics (Fig. 6, right). Since the extracel- lular solut ion was 3 m M B a , this result shows that the ki- netic convers ion is no t re la ted to the increase in intra- cellular [Ca 2+] that accompan ies the cond i t i on ing pulse or to the na ture o f the ion en te r ing t h r o u g h the activated channel .
Peak IBa f r o m the e x p e r i m e n t illustrated in Fig. 6 is p lo t ted vs. test voltage in Fig. 7. As shown, after conver- sion to fast ga t ing by the cond i t i on ing pulse (filled sym- bols) the cu r r en t activated at m o r e negative voltages. Since the descend ing b r anch o f the curve was also shifted after condi t ion ing , par t o f the shift may have b e e n due to activation o f the outward cu r r en t at lower voltages, to deple t ion o f Ba 2+ by the cond i t i on ing pulse, o r both. In spite o f these problems, it is clear that the Ba 2+ cu r r en t s tar ted to activate at lower voltage af- ter convers ion to fast gating. A voltage shift was def ined as the di f ference between the abscissas o f the vertex o f
2nd o rde r polynomials fitted to IB~ (V) with and with- ou t cond i t i on ing ( m e t h o d descr ibed in f igure legend) . The shift was 6 mV in the e x p e r i m e n t illustrated, a n d r anged be tween 5 and 9 mV in 17 o the r exper iments .
O n e o the r d i f ference was f o u n d in the kinetics after condi t ioning . As shown in Fig. 6, the decay o f Ba 2+ cur-
Test Voltage (mY) I " ~ " ~ ( l l I I
- 4 0 2 0 4 0
< c-
v
m
O
0_
- 2 1
- 4 0
_00L -I FIGURE 7. Voltage-dependence of Ic~ and the effects of condi- tioning pulse. Plot of peak Ba 2+ current as a function of test pulse voltage in reference (open symbols) and after a conditioning pulse that caused gating conversion (closed symbols, pulse protocols in Fig. 6). The voltages of maximum current were determined as the ab- scissa of the vertex of second order polynomials fitted to all data points of non-zero current, and are indicated by vertical lines that intersect the horizontal axis. Fiber # 925 (same experiment illus- trated in Fig. 6).
227 MA ET AL.
Dow
nloaded from http://rupress.org/jgp/article-pdf/108/3/221/1144134/221.pdf by guest on 30 N
ovember 2021

rent at 0 mV was very slow and became faster after con- dit ioning (Fig. 6, right). This effect was clear at voltages more negative than those that activate outward current. It is unlikely to reflect different rates of Ba 2+ depletion, since the kinetics are similar in records of very different size. It corresponds well with the increase in decay rate observed in bilayers ("conditional average" in Fig. 2 C).
Note that we have not used the term "inactivation" to identify this decay of current after the peak. Even though there is a slow vol tage-dependent inactivation in reference conditions, it will be shown in the DISCUS- SION that the increased rate of decay can be explained as a "deactivation," opera t ing on top of the normal in- activation process. In this view, deactivation is a conse- quence of conversion to fast gating.
D I S C U S S I O N
The present studies demonst ra te that individual DHP- sensitive Ca channels, which normally gate slowly un- der m e m b r a n e depolarization, are capable of opening very rapidly. The conversion f rom slow to fast gating re- quires a pretransit ion of the channel to the open state, with steep voltage- and t ime-dependent transition steps. Since the molecule can undergo both slow and rapid transitions driven by voltage, it is conceivable that the same molecule may per form slow Ca channel activation and fast control of Ca release f rom the SR.
Studies with other vol tage-dependent ion channels demonstra te that opening of the channel pore is usu- ally coupled to voltage-sensitive gating transitions via an additional process that is voltage independen t (Ox- ford, 1981; Zagotta and Aldrich, 1990). The voltage- independen t transition step involves a measurable in- crease in the volume available to water and is therefore te rmed the solvent-sensitive transition (Zimmerberg et al., 1990). Based on the exper iments with substitution of deuter ium oxide for water, Alicata et al. (1990) con- cluded that the sodium channels could activate via two alternative routes, a solvent-sensitive pathway or a sol- vent-insensitive pathway, depend ing on initial condi- tions. The multiple parallel activation mechanisms ob- served in the sodium channel are inconsistent with a linear sequential model for channel gating, and cyclic gating models have been proposed for activation of the sodium channel (Rayner et al., 1992). Similar cyclical activation mechanism has also been proposed for the gating of the Shaker K channel (Zagotta et al., 1994; Starkus et al., 1995).
The rapid secondary activation of the DHP-sensitive Ca channel seen after depolarizing prepulses is not compatible with a l inear activation model, but is highly consistent with a cyclic activation mechanism. In the following, we consider possible models for activation of the DHP-sensitive Ca channel.
A State DiagramforDHP Receptor Gating
Feldmeyer et al. (1990) in terpre ted the fast activation of skeletal muscle Ica induced by the depolarizing condi- t ioning pulse as due to the existence of at least two pathways, one fast and one slow, for the channels to pass f rom a closed into an open state. The state dia- gram suggested is represented in Fig. 8 A. This model assumes that the channel normally opens via Cx --~ C2
0:3, including a fast and a slow transition. The fast horizontal transitions (C~ ~ C~) and C4 --~ O3) generate the measurable in t r amembrane charge movemen t (Q). The slow vertical transition C 2 ~ O:~ is the rate limiting step for the increase in the m e m b r a n e Ca channel con- ductance (G). The condit ioning depolarization drives the channel into state C4, and if the open channel closes via the fast route 03 --~ C4 rather than the slow route O~ ---) C 2, then the reopening will be fast (through C4 --~ O3) for as long as the channel dwells in state C4. Thus, the model explains qualitatively both slow and fast gating of the DHP-sensitive Ca channel.
The midpoin t potential between C~ and C2 (V~2) should be close to - 4 0 mV, as the exper imenta l Qvs. V is centered at ~ - 4 0 mV, and that between C 2 and O3 (Vz3) should be close to 0 mV since G vs. V is centered at N0 mV (Pizarro et al., 1988). Since Q vs. v is hardly affected by the condit ioning pulse (Feldmeyer et al., 1990), the transition voltage V43 should be close to Vt2. Also to justify the small change in Q(V) upon conver- sion, the total charge involved in the horizontal transi- tions should be about the same, which in turn requires apparen t valences to be equal. These considerations constrain the four state model to having approximately the same central potentials for its two horizontal transi- tions. Then, to satisfy microscopic reversibility, VH should be approximately equal to V2: 3. With these con- straints, quantitative problems with the model pre- sented in Fig. 8 A become apparent . Since the fast acti- vating I~:a that occurs after a condit ioning pulse corre- sponds to transition C4 --~ 03, the activation of which should be centered at - 4 0 mV instead of the usual 0 mV at which slow Ir activates. Instead of the predicted 40 mV shift, only a small shift of 5-10 mV was found in actual measurements (Fig. 7).
To account for the exper imenta l result, more closed states must be added, in a manne r that separates the high vol tage-dependent transition f rom actual channel opening. Fig. 8 B illustrates our six-state cyclic model for the activation of the DHP receptor. The main dif- ference f rom the four-state model is the assumption of two classes of steps or conformational changes in the pathway to channel opening, namely voltage-depen- dent (horizontal) and vol tage-independent (vertical) transitions. Two vol tage-dependent steps (C1 ~ C2 and C 2 ~ C3) , with different transition voltages (Vlu, V,_,:~) are the min imum necessary to account for the activa-
228 Calcium Channels of Skeletal Muscle
Dow
nloaded from http://rupress.org/jgp/article-pdf/108/3/221/1144134/221.pdf by guest on 30 N
ovember 2021

A C1 <
I sl~
C4 <
fast
fast
> C2
sl~ > 03
V12 B c, < >
I RI4
C4 < V45
),
V23 C2 4: > C3
1 R2' l R36
V56 C5 < > 0 6
C [ 3 D > [:]ff] <
II 0 0 , > O O <
Dn mm mm " "mm
IT IT � 9 >QO O0 < O0
1 1 SR Channel Open TT Channel Open
FIGURE 8. A state diagram of DHP receptor gating. (A) State model of Feldmeyer et al. (1990). See explanation in the test. (B) Six-state model for skeletal muscle Ca channel. The horizontal transitions are fast and voltage dependent , while the vertical transi- tions are slow and voltage independent. A resting channel nor- mally opens through C a --~ C 1 ---> C 3 ---> 06, with the last step being rate limiting (slow). Upon repolarization the channel closes
through C5 & C4 (rather than C3 & Cz). Thus, a conditioned chan- nel would open from C 4, which is fast, as the rate limiting transi- tion step is skipped. (C) Molecular representation of DHP recep-
tor. The picture had three assumptions: (a) the channel as a whole can exist in two different configurations (U] and O; �9 and Q); (b) repeat I of DHP receptor differs from the other three in two as- pects, its equilibrium transition potential is more positive and its
forward transition rate is slower, and (c) the transitions from squares to circles (D ---> O and �9 ---> O) in the vertical direction are voltage independent, which proceed in a cooperative way, i.e., the rates become progressively faster as more repeats are moved to their activated states. The midpoint potentials of the horizontal transitions could be determined by the voltage-dependent distri-
bution of charge movement and channel conductance (V12 , ' - -40 mV and V23 N0 mV). The low open probability of the skeletal mus-
cle Ca channel sets a limit on the final equilibrium constant (~6
--,0.25). c~36 should be small, to account for slow activation of Ic~ (a.~6 ,--0.01 ms 1). According to Feldmeyer et al. (1990), recovery of fast gating was slower near threshold potential of channel activa- tion (~2 s, C~ ---> C2), and faster at resting potential (,'-200 ms, C4 --* CI). Thus, ~41 ~2.5 s 1 and c~2 ~0.5 s -1.
tion of two processes with different voltage depen- dence. A possible molecular interpretat ion shown in Fig. 8 C is that four internal repeats of the DHP recep- tor can be driven by voltage f rom a resting state (0pen symbols) to an activating state (filled symbols). The transi- tion C1 --~ C2 can be pictured as the activation of three repeats at a relatively lower voltage (V12 = - 4 0 mV, steepness factor Klz = 6 mV), and transition C2 ---> C3 as activation of one repeat which occurs at higher voltage (V2.~ = +14 mV, K,23 = 18 mV). The C 2 ---> C 3 transition is likely to involve the movemen t of the first internal re- peat, as the early studies by Tanabe et al. (1991) showed that repeat I of the DHP receptor is essential for the slow activation of the skeletal muscle DHP-sensi- tive Ca channel. An additional step, slow and voltage- independen t (Cs --> O6), is necessary to open the chan- nel. Similar to the allosteric model of Marks and Jones (1992) for L-type Ca channels, the vertical transitions may occur f rom any of the states in the activation chain, but they proceed more rapidly as more of the in- ternal repeats convert to the activated form.
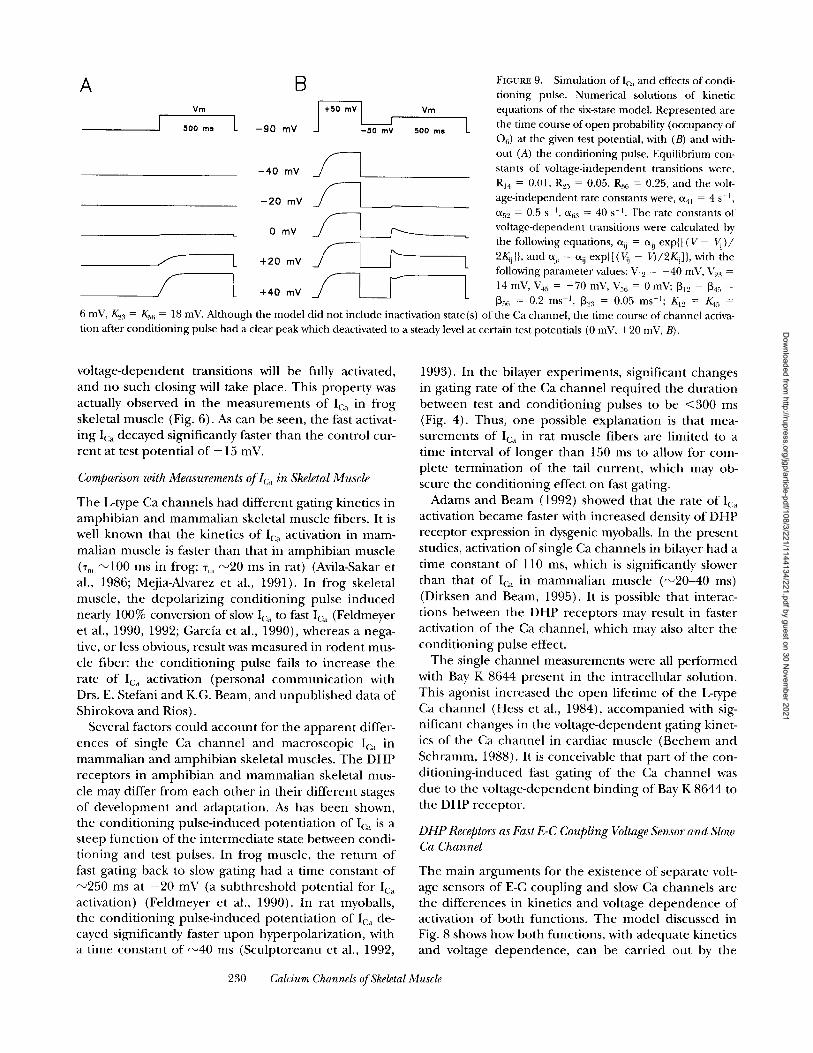
With these assumptions, the usual activation pathway for Ica is CI --> C2 ---> C:~ --> 06. Its rate is limited by the step C3 ---> 06. As in the four-state model , upon repolar- ization the channels close through C5 and C4. From these states activation is fast, as the rate limiting transi- tion is skipped. Thus, the model explains the fast and slow gating behavior of the skeletal muscle DHP-sensi- tive Ca channel. To satisfy the microscopic reversibility, the mid-voltage of the C5 --> 06 transition would have to be more negative than that of the C2 ---> C3 transition, this in turn, predicts that activation of Ica should shift to a lower potential when converted to fast gating (:'-14 mV, Fig. 9). This was observed in the measu remen t of Ica in single fiber f rom frog muscle (Fig. 7).
The first order differential equations describing the kinetic behavior of the model were solved numerically using the unmodif ied Euler 's me thod for pulses to vari- ous voltages, with and without conditioning. The depo- larizing condit ioning pulse (to +50 mV) converts the channel f rom slowly activating (Fig. 9 A) to rapidly acti- vating (Fig. 9 B). Even though no inactivated states are included in the model , the t ime course of Po (occu- pancy of 06) elicited by some of the test pulses pre- ceded by condit ioning exhibits a peak followed by a de- cay (Fig. 9 B, at test voltages 0 and +20 mV). The re- duct ion in Po is a consequence in the model of the small shift to lower voltages that accompanies conver- sion to fast mode. At intermediate potentials, that are activating for the fast mode but not for the slow mode, the channels that have been converted (dwelling in state C5 after the condit ioning pulse) will activate (to 06) dur ing the test pulse, but then a fraction will move to C3 and C 2 given the higher transition voltage of the C2 ---> C3 conversion. At high voltages however, all the
229 MA ET AL.
Dow
nloaded from http://rupress.org/jgp/article-pdf/108/3/221/1144134/221.pdf by guest on 30 N
ovember 2021
A B Vm
] 500 ms L --gO rT1V
-40 mV
-20 mV _ ~
0 mV
/'---I_ +2o ~v
Vm
500 ms L
L
V L
FIGURE 9. Simulation of I(a~ and effects of condi- tioning pulse. Numerical solutions of kinetic equations of the six-state model. Represented are the time course of open probability (occupancy of 06) at the given test potential, with (B) and with- out (A) the conditioning pulse. Equilibrium con- stants of voltage-independent transitions were, RI4 = 0 . 0 1 , R25 = 0 . 0 5 , R:~ 6 = 0 . 2 5 , and the volt- age-independent rate constants were, cq~ = 4 s -t, c~5,, = 0.5 s -t, c~a:~ = 40 s -~. The rate constants of voltage-dependent transitions were calculated by the following equations, % = % exp{[(V- Vii)/ 2/~i]}, and %i = % exp{[(Vij - V)/2/~i]}, with the following parameter values: V]~ = -40 mV, Vu:~ = 14 mV, V45 = -70 mV, V56 = 0 r a g ; ~12 = ~45 =
[35t~ = 0.2 ms -1, [~z,~ = 0.05 ms-l; K12 = K4r, =
6 mV, K.2: r = K.36 = 18 mV. Mthough the model did not include inactivation state(s) of the Ca channel, the time course of channel activa- tion after conditioning pulse had a clear peak which deactivated to a steady level at certain test potentials (0 mV, +20 mV, B).
vo l t age-dependen t transitions will be fully activated, and no such closing will take place. This p roper ty was actually observed in the measuremen t s o f Ica in f rog skeletal muscle (Fig. 6). As can be seen, the fast activat- ing Ica decayed significantly faster than the cont ro l cur- ren t at test potent ia l o f + 15 mV.
Comparison with Measurements of lc~ in Skeletal Muscle
The L-type Ca channels had di f ferent gat ing kinetics in amph ib ian and m a m m a l i a n skeletal muscle fibers. It is well known that the kinetics o f Ica activation in mam- malian muscle is faster than that in amph ib ian muscle (%, ~ 1 0 0 ms in frog; %, ~ 2 0 ms in rat) (Avila-Sakar et al., 1986; Mejia-Alvarez et al., 1991). In f rog skeletal muscle, the depolar iz ing cond i t ion ing pulse i nduced nearly 100% conversion of slow Ic~ to fast Ic~, (Feldmeyer et al., 1990, 1992; Garcfa et al., 1990), whereas a nega- tive, or less obvious, result was measu red in r o d e n t mus- cle fiber: the cond i t ion ing pulse fails to increase the rate o f Ic~ activation (personal c o m m u n i c a t i o n with Drs. E. Stefani and K.G. Beam, and unpub l i shed data o f Shirokova and Rios).
Several factors could accoun t for the appa ren t differ- ences o f single Ca channe l and macroscopic Ic, in mammal i an and amph ib i an skeletal muscles. The D H P receptors in amph ib ian and m a m m a l i a n skeletal mus- cle may differ f rom each o the r in their d i f ferent stages o f deve lopmen t and adaptat ion. As has been shown, the cond i t ion ing pulse- induced potent ia t ion o f Ica is a steep func t ion o f the in te rmedia te state between condi- t ioning and test pulses. In f rog muscle, the re tu rn o f fast gat ing back to slow gat ing had a t ime cons tan t o f ~ 2 5 0 ms at - 2 0 mV (a sub threshold potent ia l for Ic~ activation) (Feldmeyer et al., 1990). In rat myoballs, the cond i t ion ing pulse- induced potent ia t ion o f Ica de- cayed significantly faster u p o n hyperpolar izat ion, with a time cons tant o f ~ 4 0 ms (Sculp toreanu et al., 1992,
1993). In the bilayer exper iments , significant changes in gat ing rate o f the Ca channe l requi red the dura t ion between test and cond i t ion ing pulses to be < 3 0 0 ms (Fig. 4). Thus, one possible explana t ion is that mea- surements o f Ic~, in rat muscle fibers are l imited to a time interval o f longer than 150 ms to allow for com- plete t e rmina t ion o f the tail current , which may ob- scure the cond i t ion ing effect on fast gating.
Adams and Beam (1992) showed that the rate o f Ic~ activation became faster with increased density o f D H P receptor expression in dysgenic myoballs. In the present studies, activation o f single Ca channels in bilayer had a time cons tant o f 110 ms, which is significantly slower than that o f Ic, in m a m m a l i a n muscle ( ~ 2 0 - 4 0 ms) (Dirksen and Beam, 1995). It is possible that interac- tions between the DHP receptors may result in faster activation o f the Ca channel , which may also alter the cond i t ion ing pulse effect.
The single channel measurements were all per formed with Bay K 8644 present in the intracellular solution. This agonist increased the o p e n lifetime o f the L-type Ca channe l (Hess et al., 1984), a c c o m p a n i e d with sig- nificant changes in the vo l t age-dependen t gat ing kinet- ics o f the Ca channe l in cardiac muscle (Bechem and Schramm, 1988). It is conceivable that par t o f the con- d i t ion ing- induced fast gat ing o f the Ca channe l was due to the vo l t age -dependen t b ind ing o f Bay K 8644 to the DHP receptor .
DHP Receptors as Fast E-C Coupling Voltage ,Sensor and Slow Ca Channel
The main a rgumen t s for the existence o f separate volt- age sensors o f E-C coup l ing and slow Ca channels are the differences in kinetics and voltage d e p e n d e n c e o f activation o f bo th functions. The mode l discussed in Fig. 8 shows how bo th functions, with adequa te kinetics and voltage d e p e n d e n c e , can be carr ied ou t by the
230 Calcium Channels of Skeletal Muscle
Dow
nloaded from http://rupress.org/jgp/article-pdf/108/3/221/1144134/221.pdf by guest on 30 N
ovember 2021

same mo lecu l e . A s s u m i n g tha t par t ia l ly ac t iva ted D H P recep to r s , C 2 a n d C5, which can be r e a c h e d at fast s p e e d a n d at lower voltages, c o u l d s ignal o p e n i n g o f the r y a n o d i n e r e c e p t o r Ca re lease c h a n n e l ( pe r sona l communica t i ons with K.G. Beam, E. Stefani, and E. Rios), this m o d e l pu t s t o g e t h e r the two func t ions o f the D H P r e c e p t o r : e x p l a i n i n g slow act iva t ion o f the Ca c u r r e n t a n d fast c o n t r o l o f SR Ca re lease .
Two o t h e r obse rva t ions have b e e n t aken as ev idence o f s epa ra t e vol tage sensors a n d Ca channe l s . O n e is t ha t the m a g n i t u d e o f Ica is less t han e x p e c t e d given the dens i ty o f D H P r e c e p t o r s in the T T m e m b r a n e (Schwartz e t al., 1985), which suggests tha t only a small f rac t ion o f D H P r e c e p t o r s func t ion as Ca channe l s . I t has b e e n p o i n t e d out , however , tha t the smal l c u r r e n t may also be a c o n s e q u e n c e o f low o p e n p robab i l i t y o f the Ca c h a n n e l ( L a m b a n d Walsh , 1987), which was f o u n d to be the case for T T m e m b r a n e Ca c h a n n e l s in bi layers, even w h e n they were fully ac t iva ted by vol tage a n d in t he p r e s e n c e o f the agon is t Bay K 8644 (Ma et al. 1991).
T h e o t h e r obse rva t ion tha t l ed P izar ro et al. (1988) to sugges t tha t d i f f e r en t D H P r ecep to r s a re r e spons ib l e for the vol tage senso r o f E-C c o u p l i n g a n d the conduc t - ing un i t o f the L-type Ca c h a n n e l was the r e m a r k a b l e d i f f e rence in t ime course o f recovery o f Ica a n d SR Ca re lease f lux af te r inac t iva t ion by p r o l o n g e d depo la r i za - t ion. T h e recovery o f Ica o c c u r r e d wi thou t a p p r e c i a b l e delay, fo l lowing an e x p o n e n t i a l f u n c t i o n o f t ime, whi le r e p r i m i n g o f Ca re lease fo l lowed a s igmoida l t ime course . T h e d i f f e rence in t ime course o f r e p r i m i n g can
be e x p l a i n e d if o p e n i n g o f o n e re lease c h a n n e l is de te r - m i n e d by the j o i n t ac t iva t ion o f m u l t i p l e D H P recep- tors. This i n t e r p r e t a t i o n seems na tu r a l s ince D H P re- cep to r s a p p e a r in g r o u p s o f four , the j u n c t i o n a l t e t rads i den t i f i ed in e l e c t r o n m i c r o s c o p y o f f reeze f r a c t u r e d t ransverse tubu les by Block et al. (1988). S i m o n a n d Hil l (1992) r e p o r t e d a 4th p o w e r r e l a t i onsh ip b e t w e e n Ca re lease f lux a n d cha rge m o v e m e n t , while Rios et al. (1993) also f o u n d a p o w e r r e l a t i onsh ip , o f e x p o n e n t be tw e e n 2 a n d 3. T h e s e resul ts sugges t tha t m u l t i p l e D H P r e c e p t o r s a re involved in the c on t ro l o f Ca re lease f r o m the SR m e m b r a n e . This i n t e r p r e t a t i o n makes the p h a r m a c o l o g i c a l d i f f e rences b e t w e e n E-C c o u p l i n g a n d Ca cu r r en t s n o t e d by R o m e y et al. (1988) eas ie r to un- de r s t and . A d r u g will be m o r e effective in b l o c k i n g Ca re lease t han Ica, for the same r ea son tha t Ica recovers m o r e r ap id ly than Ca re lease f rom v o l t a g e - i n d u c e d in- act ivat ion, p r o v i d e d tha t it b locks ear ly s teps in the acti- va t ion o f Ica ( F e l d m e y e r et al., 1992).
Given tha t the cyclical m o d e l shown in Fig. 8 app l i e s to ac t iva t ion o f the DHP-sensi t ive Ca c h a n n e l s as well as the Na channe l (Alicata et al., 1990; Starkus and Rayner, 1991) a n d K c h a n n e l (Zagot ta e t al., 1994; a n d Starkus et al., 1995), it is r e a s o n a b l e to p r o p o s e tha t vol tage- sensit ive ga t ing t rans i t ions o c c u r ear ly in the Ca chan- ne l ac t iva t ion s e q u e n c e a n d tha t it is these processes which c o u l d u n d e r l i e the ro le o f the vol tage senso r a n d yie ld the r a p i d E-C c o u p l i n g in skele ta l muscle , t h r o u g h e i t he r e lec t ros ta t ic o r a l los ter ic l inkage to the r y a n o d i n e r e c e p t o r Ca re lease channe l .
A major portion of the experiments was performed when the corresponding author was a postdoctoral fellow in the laboratory of Dr. E. Rfos at Rush Medical College. We owe him for the continuous support and encouragement, and many stimulating dis- cussions. We thank Dr. M.M. Hosey for the initial supply of membrane vesicles, and for providing critical comments on the pa- per. We are grateful to Drs. F.S. Cohen and S.W.Jones for reading the manuscript and sharing many ideas.
This work was supported by grants from Muscular Dystrophy Association, an Established Investigatorship from American Heart Association toJ. Ma, and from National Institutes of Health grants toJ. Ma (AR42057) and to E. Rfos (AR32808).
Original version received 11 May 1993 and accepted version received 29 April 1996.
R E F E R E N C E S
Adams, B.A., and K.G. Beam. 1992. Calcium current activates faster in myotubes expressing higher calcium current density. Biophys. J. 61:A419.
Adams, B.A., T. Tanabe, A. Mikami, S. Numa, and K.G. Beam. 1990. Intramembrane charge movement restored in dysgenic skeletal muscle by injection of dihydropyridine receptor cDNAs. Nature (Lond. ). 346:569-572.
Affolter, H., and R. Coronado. 1985. Agonist Bay K 8644 and CGP- 28392 open calcium channels reconstituted from skeletal muscle transverse tubules. Biophys. J. 48:341-347.
Alicata, D.A., M.D. Rayner, andJ.G. Starkus. 1990. Sodium channel activation mechanisms. Insights from deuterium oxide substitu- tion. Biophys.~ 57:745-758.
Avila-Sakar, AJ., G. Cota, R. Gamboa-Aldeco, J. Garcia, M. Huerta, J. Muniz, and E. Stefani. 1986. Skeletal muscle Ca channels. J. Muscle Res. Cell Motil. 7:291-298.
Beam, K.G., B.A. Adams, T. Niidome, S. Numa, and T. Tanabe. 1992. Function of a truncated dihydropyridine receptor as both voltage sensor and calcium channel. Nature (Lond.). 360:169-171.
Bechem, M., and M. Schramm. 1988. Electrophysiology of dihydro- pyridine calcium agonists. In The Calcium Channel: Structure, Function and Implications. M. Morad, W. Nayler, S. Kazda, and M. Schramm, editors. Springer Verlag. Berlin-Heidelberg. 39-53.
Block, B.A., T. Imagawa, K.P. Campbell, and C. Franzini-Arm- strong. 1988. Structural evidence for direct interaction between the molecular components of the transverse tubule/sarcoplas- mic reticnlmnjunction in skeletal muscle.J. Cell Biol. 107:2587- 2600.
DeJongh, K.S., D.K. Merrick, and W.A. Catterall. 1989. Subunits of purified calcium channels: a 212 kDa form of ~1 and partial amino acid sequence of a phosphorylation site of an indepen- dent 13 subunit. Proc. Natl. Acad. Sci. USA. 86:8585-8589.
231 MA ET AL.
Dow
nloaded from http://rupress.org/jgp/article-pdf/108/3/221/1144134/221.pdf by guest on 30 N
ovember 2021

Dirksen, R.T., and K.G. Beam. 1995. Single calcium channel behav- ior in native skeletal muscle.J. Gen. Physiol. 105:227-248.
Feldmeyer, D., W. Melzer, B. Pohl, and P. Zollner. 1990. Fast gating kinetics of the slow Ca current in cut skeletal muscle fibers of the frog. J. Physiol. (Lond.). 425:347-367.
Feldmeyer, D., W. Melzer, B. Pohl, and P. Zollner. 1992. Modula- tion of calcium current gating in frog skeletal muscle by condi- tioning depolarization.J. Physiol. (Lond.). 457:639-653.
Galizzi,J.P., M. Fosset, and M. Lazdunski. 1984. Properties of recep- tors for the Ca channel blocker verapamil in transverse tubule membranes of skeletal muscle. Eur. J. Biochem. 144:211-214.
Garc/a, J., A.J. Avila-Sakar, and E. Stefani. 1990. Repetitive stimula- tion increases the activation rate of skeletal muscle Ca currents. Pflugers Arch. 416:210--212.
Gonzalez, A., and E. Rios. 1993. Perchlorate enhances transmission in skeletal muscle excitation-contraction coupling.J. Gen. Physiol. 102:373--421.
Hess, P., J.B. Lansman, and R.W. Tsien. 1984. Different modes of Ca channel gating behavior favored by dihydropyridine Ca ago- nist and antagonists. Nature (Lond.). 311:538-544.
Lamb, G.D., and T. Walsh. 1987. Calcium currents, charge move- ment and dihydropyridine binding in fast and slow twitch muscle of rat and rabbit. J. Physiol. (Lond.). 393:595-617.
Ma, J., and R. Coronado. 1988. Heterogeneity of conductance states in calcium channels of skeletal muscle. Biophys. J. 53:387- 395.
Ma, J., C. Mundina-Weilenmann, M.M. Hosey, and E. Rfos. 1991. Dihydropyridine-sensitive skeletal muscle Ca channels in polar- ized planar bilayers. I. Kinetics and voltage dependence of gat- ing. Biophys.]. 60:890-901.
Marks, T.N., and S.W..Jones. 1992. Calcium currents in the A7r5 smooth muscle-derived cell line. An allosteric model for calcium channel activation and dihydropyridine agonist action. J. Gen. Physiol. 99:367-390.
Mejia-Alvarez, R., M. Fill, and E. Stefani. 1991. Voltage-dependent inactivation of t-tubular skeletal muscle calcimn channels in pla- nar lipid bilayers.J. Gen. Physiol. 97:393-412.
Oxtord, G.S. 1981. Some kinetic and steady-state properties of so- dium channels after removal of inactivation. J. Gen. PhysioL 77:1- 22.
Pizarro, G., L. Csernoch, I. Uribe, M. Rodriguez, and E. Rfos. 1991. The relationship between Q' , /and calcium release from the sar- coplasmic reticulum in skeletal muscle. J. Gen. Physiol. 97:913- 947.
Pizarro, G., R. Fitts, G. Bruin, M. Rodriguez, and E. Rios. 1988. The voltage sensor of skeletal muscle excitation-contraction coupling: a comparison with Ca channels. In The Calcium Channel, Struc- ture, Function and Implications, M. Morad W. Nayler, S. Kazda, and M. Schramm, editors. Springer-Verlag. Berlin-Heidelberg. 138-156.
Rayner, M.D., J.G. Starkus, P.C. Ruben, and D.A. Alicata. 1992. Vnltage-sensitive and solvent-sensitive processes in ion channel gating. Kinetic effects of hyperosmolar media on activation and deactivation of sodium channels. Biophys.J. 61:96-108.
R/os, E., and G. Bruin. 1987. Involvement of dihydropyridine re- ceptors in excitation-contraction coupling in skeletal muscle. Na- ture (Lond.). 325:717-720.
R/os, E., M. Karhanek, J. Ma, and A. Gonzailez. 1993. An allosteric model of the molecular interactions of excitation contraction coupling in skeletal muscle.J. (;.en. Physiol. 102:449-481.
Romey, G., L. Garcfa, F. Rieger, and M. Lazdunski. 1988. Targets for calcium channel blockers in mammalian skeletal muscle and their respective functions in excitation-contraction coupling. Bio- chem. Biophys. Res. Commun. 156:1324-1332.
Schneider, M.F., and W.K. Chandler. 1973. Voltage dependent charge movement in skeletal muscle: a possible step in excita- tion-contraction coupling. Nature (Lond.). 242:244-246.
Schwartz, L.M., E.M. McCleskey, and W. Almers. 1985. Dihydropyri- dine receptors in muscle are voltage-dependent but most are not functional calcium channels. Nature (Lond.). 314:747-750.
Sculptoreanu, A., T. Scheuer, and W.A. Catterall. 1992. Voltage and t ime-dependent potentiation of L-type calcium currents in skeletal muscle cells. The Physiologist. 35:A8.
Sculptoreanu, A., T. Scheuer, and W.A. Catterall. 1993. Voltage-de- pendent potentiation of L-type Ca channels due to phosphoryla- tion by cAMP-dependent protein kinase. Nature (Lond.). 364:240- 243.
Simon, B.J., andJ.A. Hill. 1992. Charge movement and SR calcimn release in frog skeletal muscle can be related by a Hodgkin and Huxley model with four gating particles. Biophys. J. 61:1109-11 l 6.
Simon, B.J., and M.F. Schneider. 1988. Time course of activation of calcium release from sarcoplasmic reticulum in skeletal muscle. Biophys.J. 84:1159-1163.
Starkus, J.G., and M.D. Rayner. 1991. Gating current "fraction- ation" in crayfish giant axons. Biophys.J. 60:1101-1119.
Starkus, J.G., T. Schlief, M.D. Rayner, and S.H. Heinemann. 1995. Unilateral exposure of Shaker B potassium channels to hyperos- molar solutions. Biophys. J. 69:860-872.
Tanabe, T., B.A. Adams, S. Numa, and K.G. Beam. 1991. Repeat I of the dihydropyridine receptor is critical in determining cal- cium channel activation kinetics. Nature (Lond.). 352:800-803.
Tanabe, T., K.G. Beam,J.A. Powell, and S. Numa. 1988. Restoration of excitation-contraction coupling and slow calcium current in dysgenic muscle by dihydropyridine receptor complementaly DNA. Nature (Lond. ). 336:134-139.
Tanabe, T., H. Takeshima, A. Mikami, V. Flockerzi, H. Takabashi, K. Kangawa, M. Kojima, H. Matsuo, T. Hirose, and S. Numa. 1987. Primary structure of the receptor for calcium channel blockers from skeletal muscle. Nature (Lond.). 328:313-318.
Zagotta, W.N., and R.W. Mdrich. 1990. Voltage-dependent gating of ShakerA-type potassium channels in Drosophila muscle.J. Gen. Physiol. 95:29-60.
Zagotta, W.N., T. Hoshi,J. Dittman, and R.W. Aldrich. 1994. Shaker potassium channel gating I: transitions near the open state. J. Gen. Physiol. 103:279-319.
Zimmerberg, J., F. Bezanilla, and V.A. Parsegian. 1990. Solute inac- cessible aqueous volume changes during opening of the potas- sium channel of the squid giant axon. Biophys. J. 57:1049-1064.
232 Calcium Channels of Skeletal Muscle
Dow
nloaded from http://rupress.org/jgp/article-pdf/108/3/221/1144134/221.pdf by guest on 30 N
ovember 2021