Embed Size (px)
Citation preview

This article was downloaded by: [Laurentian University]On: 06 October 2014, At: 02:00Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Journal of Environmental Science and Health, Part B:Pesticides, Food Contaminants, and Agricultural WastesPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/lesb20
Factors affecting the removal of aflatoxin M1 from foodmodel by Lactobacillus and Bifidobacterium strainsBulent Kabak a & Isil Var ba Department of Food Engineering , Hitit University , Corum, Turkeyb Department of Food Engineering , Cukurova University , Adana, TurkeyPublished online: 20 Sep 2008.
To cite this article: Bulent Kabak & Isil Var (2008) Factors affecting the removal of aflatoxin M1 from food modelby Lactobacillus and Bifidobacterium strains, Journal of Environmental Science and Health, Part B: Pesticides, FoodContaminants, and Agricultural Wastes, 43:7, 617-624, DOI: 10.1080/03601230802234740
To link to this article: http://dx.doi.org/10.1080/03601230802234740
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Journal of Environmental Science and Health Part B (2008) 43, 617–624Copyright C© Taylor & Francis Group, LLCISSN: 0360-1234 (Print); 1532-4109 (Online)DOI: 10.1080/03601230802234740
Factors affecting the removal of aflatoxin M1 from foodmodel by Lactobacillus and Bifidobacterium strains
BULENT KABAK1 and ISIL VAR2
1Department of Food Engineering, Hitit University, Corum, Turkey2Department of Food Engineering, Cukurova University, Adana, Turkey
This paper describes the ability of six dairy strains of Lactobacillus and Bifidobacterium to remove aflatoxin M1 (AFM1) fromphosphate-buffered saline (PBS) and reconstituted milk. Bacteria were incubated in both PBS and reconstituted milk containing 5,10 and 20 ng mL−1 for 0, 4 and 24 h at 37◦C. After centrifugation the concentration of AFM1 was determined in the supernatantfraction using high-performance liquid chromatography. The binding abilities of AFM1 by viable (108 CFU mL−1) and heat-killedLactobacillus and Bifidobacterium strains in PBS ranged from 10.22 to 26.65% and 14.04 to 28.97%, respectively. Similarly, AFM1-binding capacity in reconstituted milk was found to range from 7.85 to 25.94% and from 12.85 to 27.31% for viable and heat-killedbacteria, respectively within 4 h. While B. bifidum Bb 13 was the best binder, the poorest removal was achieved by L. acidophilusNCC 68. Binding was reversible, and a small proportion of AFM1 was released back into the solution. The toxin concentration andincubation period had no effect on the removal of AFM1 by bacteria both in PBS and reconstituted milk.
Keywords: Aflatoxin M1; Lactobacillus; Bifidobacterium; removal; high-performance liquid chromatography.
Introduction
Aflatoxins are a group of secondary metabolites producedby three species of Aspergillus: flavus, parasiticus and therare nomius.[1] Up until now, approximately 18 differenttoxic derivatives of aflatoxins have been reported. Amongthese, aflatoxin B1 (AFB1) is the most naturally occurringcompound by toxigenic Aspergillus species.[2] When lactat-ing animals such as cows, goats and humans are fed withfeedstuffs contaminated with AFB1, this metabolite can betransferred to milk as aflatoxin M1 (AFM1) in the range of0.3–6.3%.[3] AFM1, monohydroxylated derivative of AFB1,is as acute atoxic as AFB1 or slightly less than that of AFB1,while its carcinogenic and mutagenic potential seems to belower.[1] In 1993, AFB1 and AFM1 were classified by theInternational Agency for Research of Cancer (IARC) ofWHO as 1A (carcinogenic) and 2B (possible human car-cinogen), respectively.[4]
AFM1 can be found in a number of food products in-cluding milk, dried milk, cheese, yogurt, butter and in-fant formula. As milk and dairy products are important
Address correspondence to Bulent Kabak, Hitit University,Faculty of Engineering, Department of Food Engineering,Corum, Turkey; E-mail: [email protected] February 7, 2008.
source of nutrients, the contamination of these productswith AFM1 is a potential risk for human health. Manycountries have regulated the maximum permissible level ofAFM1 allowed in milk and milk-based products. The Sci-entific Commission of the European Community has regu-lated the maximum allowable level of 0.05 µg kg−1 AFM1for raw milk, heat-treated milk and milk for the manufac-ture of milk-based products.[5]
Because of its detrimental effects, a number of strategieshave been developed to prevent the growth of mycotox-igenic fungi, as well as to decontaminate and/or detox-ify mycotoxin contaminated foods and animal feeds.[6] Theuse of many of the available physical and chemical meth-ods for the detoxification of food products contaminatedwith mycotoxins is restricted due to problems concerningsafety issues, possible losses in the nutritional quality oftreated commodities, coupled with limited efficacy and costimplications.[7] This has led to the search for alternativestrategies such as biological agents. Recently, there has beenincreasing interest in the hypothesis stating that absorp-tion of aflatoxin in consumed food could be reduced bymicroorganisms in the gastrointestinal tract; consequently,numerous investigators showed that some dairy strains oflactic acid bacteria and bifidobacteria were able to bind ef-fectively to AFB1 in buffered solution. El-Nezami et al.[8]
showed that within 24 h, a 24 h old culture of Lactobacil-lus rhamnosus GG and L. rhamnosus strain LC-705 were
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 0
2:00
06
Oct
ober
201
4

618 Kabak and Var
able to remove approximately 80% of AFB1. In other re-search by Peltonen et al.,[9] the removal of AFB1 frombuffered solution by probiotic bacteria was found to rangefrom 5.8 to 31.3%. On the other hand, there is limited dataon the removal of the much polar mycotoxin, AFM1 fromaqueous solution and milk. The primary aim of this studywas to determine the ability of some strains of lactobacilliand bifidobacteria to bind AFM1 in artificially contami-nated phosphate-buffered saline (PBS) and reconstitutedmilk.
Materials and methods
Bacterial strains, culture conditions and estimation ofbacterial concentration
Four Lactobacillus, L. acidophilus NCC 12, L. acidophilusNCC 36, L. acidophilus NCC 68 and L. rhamnosus, andtwo Bifidobacterium strains, B. bifidum Bb13 and B. bi-fidum NCC 381 were tested for their ability to bind AFM1.The strains except L. rhamnosus (purchased from Ezal,France) were obtained from Nestle-Switzerland CultureCollection as freeze-dried powder. Lactobacilli and bifi-dobacteria were cultivated in de Man Rogosa, Sharpe broth(MRS, Merck, Germany) for 48 h at 37◦C under anaerobicconditions. Estimation of bacterial concentration in the cul-ture was performed using traditional plate counting. Twovolumes of 108 and 107CFU mL−1 of the culture were usedin the binding assay.
AFM1 standard
AFM1 was obtained from the National Institute for PublicHealth and the Environment (RIVM, The Netherlands) inthe concentration of 10 µg mL−1in methanol. A 100 µL ofAFM1 standard solution was quickly diluted with 900 µLmethanol to obtain a working solution, at 1 µg mL−1. Stan-dard working solution was freshly prepared and held for lessthan 1 month. The working solution was brought to roomtemperature before use. Three solutions of 5, 10 and 20 ngmL−1 AFM1 were prepared in both PBS and reconstitutedmilk.
Chemicals
All solvents used for the analysis of AFM1 were obtainedfrom Merck (Darmstadt, Germany) and were of high-performance liquid chromatography (HPLC) grade. PBSwas prepared by dissolving 8 g NaCl, 1.2 g K2HPO4 and0.3 g KH2PO4 in 900 mL distilled water. The pH was ad-justed to 6.5 with 0.1 M NaOH and diluted to 1000 mL.In all analytical steps, highly purified water generated by aMillipore Synergy 18S Ultra-Pure Water System from Mil-lipore (France) was used.
Immunoaffinity columns (IAC)
AflaM1 IAC containing specific monoclonal antibodiesbound to a solid support material for AFM1 clean-up wereobtained from Vicam (product code: G1007, Watertown,Mass., USA).
Cell surface hydrophobicity
The hyrophobicity was determined according to the methodof Vinderola and Reinheimer[10] with some modifications.Cell surface hydrophobicity was determined according tothe capacity of the bacteria to partition into hexadecanefrom PBS. Cultures of the strains were harvested in the sta-tionary phase by centrifugation at 3000 × g for 5 min at5◦C, washed twice in 50 mM K2HPO4 (pH 6.5) buffer andfinally resuspended in the same buffer. The cell suspensionwas adjusted to an A560nm value of approximately 1.0 withbuffer. After that, 3 mL of the bacterial suspensions wereput in contact with 0.6 mL of n-hexadecane and vortexedfor 120 s. The two phases were allowed to separate for 1h at 37◦C. The aqueous phase was carefully removed andthe A560nm was measured. The decrease in the absorbanceof the aqueous phase was taken as a measure of the cellsurface hydrophobicity (H%) which was calculated accord-ing to the formula H% = [(A0 − A)/A0] × 100, where A0and A are the absorbance before and after extraction withn-hexadecane.
AFM1 removal assay with contaminated PBS
Two volumes of the culture broth (108 and 107 CFU mL−1)were centrifuged (3000 × g, 15 min) and the bacterial pelletswere washed with water. Bacterial pellets were suspended in5 mL PBS contaminated with a standard working solutionof AFM1 at three different concentrations, 5, 10 and 20 ngmL−1. Tubes were mixed with ten seconds on vortex mixer.Bacterial suspensions were then incubated at 37◦C for 0, 4,and 24 h. An AFM1 positive control (5, 10 and 20 ng mL−1
in PBS) in the absence of bacteria and negative controls(bacteria suspended in pure PBS) were also incubated atthe same time points to monitor the efficacy of bacteria inthe binding AFM1. All tubes were centrifuged for 15 min at3000 × g (Heraeus, Labofuge 200) at the end of incubationperiod and supernatants were collected and transferred toclean tubes. The tubes were stored at 4±1◦C until AFM1analysis. The unbound AFM1 content of the supernatantswere determined by HPLC with fluorescence detection. Allexperiments were performed in triplicate. To determine theeffect of bacteria viability on the binding affinity to AFM1,bacteria (108 CFU mL−1) were heated at 90◦C for 15 min.The dead bacteria were pelleted and contaminated withAFM1 as described above.
AFM1 removal assay with contaminated reconstituted milk
The reconstituted milk was used as a food model to de-termine binding ability of AFM1 by viable and heat-killed
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 0
2:00
06
Oct
ober
201
4
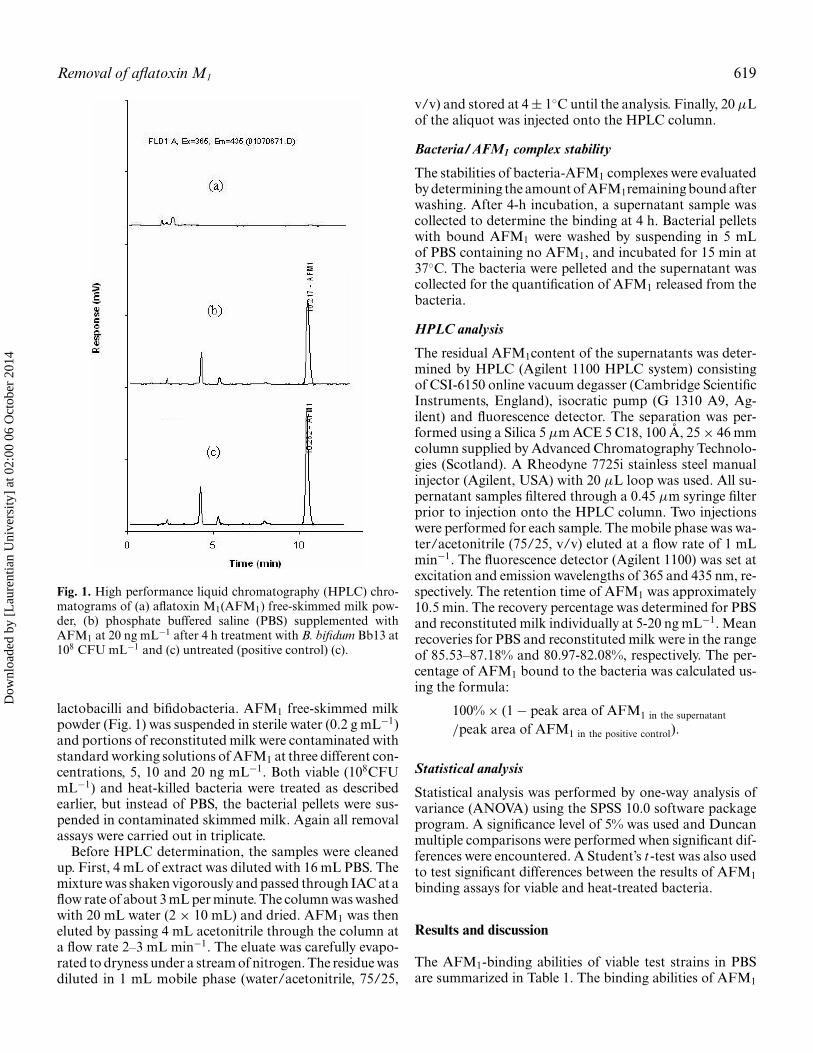
Removal of aflatoxin M1 619
Fig. 1. High performance liquid chromatography (HPLC) chro-matograms of (a) aflatoxin M1(AFM1) free-skimmed milk pow-der, (b) phosphate buffered saline (PBS) supplemented withAFM1 at 20 ng mL−1 after 4 h treatment with B. bifidum Bb13 at108 CFU mL−1 and (c) untreated (positive control) (c).
lactobacilli and bifidobacteria. AFM1 free-skimmed milkpowder (Fig. 1) was suspended in sterile water (0.2 g mL−1)and portions of reconstituted milk were contaminated withstandard working solutions of AFM1 at three different con-centrations, 5, 10 and 20 ng mL−1. Both viable (108CFUmL−1) and heat-killed bacteria were treated as describedearlier, but instead of PBS, the bacterial pellets were sus-pended in contaminated skimmed milk. Again all removalassays were carried out in triplicate.
Before HPLC determination, the samples were cleanedup. First, 4 mL of extract was diluted with 16 mL PBS. Themixture was shaken vigorously and passed through IAC at aflow rate of about 3 mL per minute. The column was washedwith 20 mL water (2 × 10 mL) and dried. AFM1 was theneluted by passing 4 mL acetonitrile through the column ata flow rate 2–3 mL min−1. The eluate was carefully evapo-rated to dryness under a stream of nitrogen. The residue wasdiluted in 1 mL mobile phase (water/acetonitrile, 75/25,
v/v) and stored at 4 ± 1◦C until the analysis. Finally, 20 µLof the aliquot was injected onto the HPLC column.
Bacteria/AFM1 complex stability
The stabilities of bacteria-AFM1 complexes were evaluatedby determining the amount of AFM1remaining bound afterwashing. After 4-h incubation, a supernatant sample wascollected to determine the binding at 4 h. Bacterial pelletswith bound AFM1 were washed by suspending in 5 mLof PBS containing no AFM1, and incubated for 15 min at37◦C. The bacteria were pelleted and the supernatant wascollected for the quantification of AFM1 released from thebacteria.
HPLC analysis
The residual AFM1content of the supernatants was deter-mined by HPLC (Agilent 1100 HPLC system) consistingof CSI-6150 online vacuum degasser (Cambridge ScientificInstruments, England), isocratic pump (G 1310 A9, Ag-ilent) and fluorescence detector. The separation was per-formed using a Silica 5 µm ACE 5 C18, 100 A, 25 × 46 mmcolumn supplied by Advanced Chromatography Technolo-gies (Scotland). A Rheodyne 7725i stainless steel manualinjector (Agilent, USA) with 20 µL loop was used. All su-pernatant samples filtered through a 0.45 µm syringe filterprior to injection onto the HPLC column. Two injectionswere performed for each sample. The mobile phase was wa-ter/acetonitrile (75/25, v/v) eluted at a flow rate of 1 mLmin−1. The fluorescence detector (Agilent 1100) was set atexcitation and emission wavelengths of 365 and 435 nm, re-spectively. The retention time of AFM1 was approximately10.5 min. The recovery percentage was determined for PBSand reconstituted milk individually at 5-20 ng mL−1. Meanrecoveries for PBS and reconstituted milk were in the rangeof 85.53–87.18% and 80.97-82.08%, respectively. The per-centage of AFM1 bound to the bacteria was calculated us-ing the formula:
100% × (1 − peak area of AFM1 in the supernatant
/peak area of AFM1 in the positive control).
Statistical analysis
Statistical analysis was performed by one-way analysis ofvariance (ANOVA) using the SPSS 10.0 software packageprogram. A significance level of 5% was used and Duncanmultiple comparisons were performed when significant dif-ferences were encountered. A Student’s t-test was also usedto test significant differences between the results of AFM1binding assays for viable and heat-treated bacteria.
Results and discussion
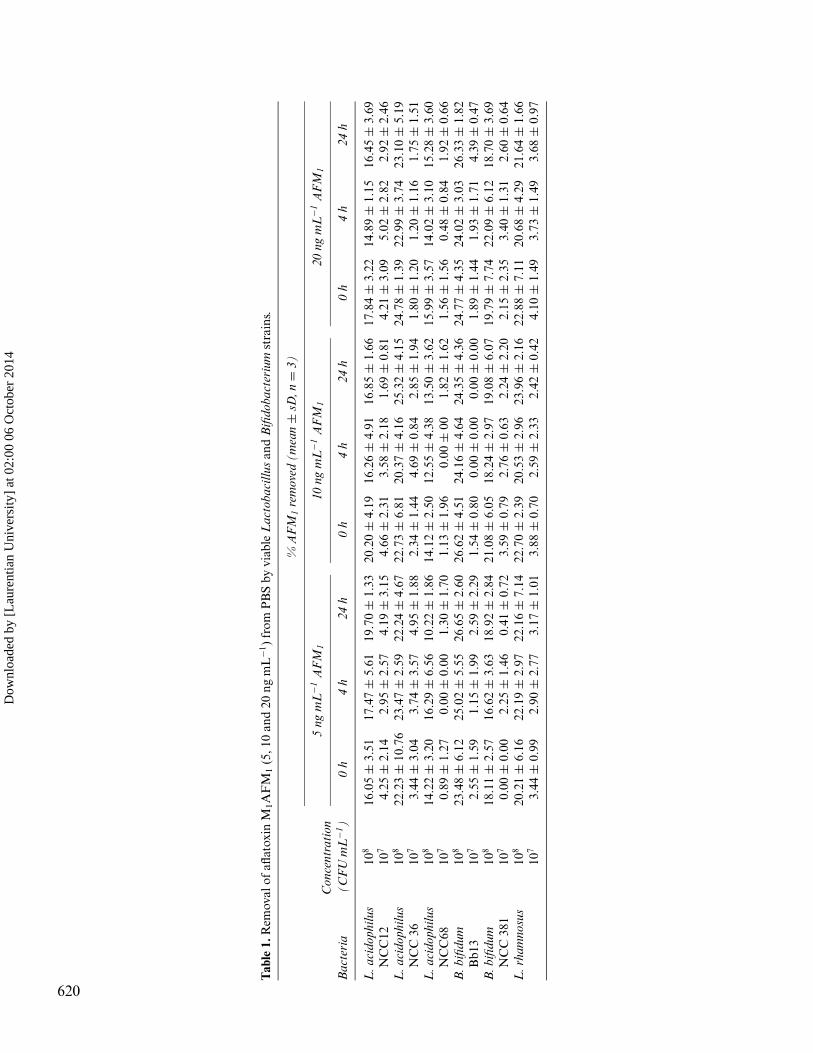
The AFM1-binding abilities of viable test strains in PBSare summarized in Table 1. The binding abilities of AFM1
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 0
2:00
06
Oct
ober
201
4
Tab
le1.
Rem
oval
ofafl
atox
inM
1A
FM
1(5
,10
and
20ng
mL
−1)
from
PB
Sby
viab
leL
acto
baci
llus
and
Bifi
doba
cter
ium
stra
ins.
%A
FM
1re
mov
ed(m
ean
±sD
,n=
3)
5ng
mL
−1A
FM
110
ngm
L−1
AF
M1
20ng
mL
−1A
FM
1C
once
ntra
tion
Bac
teri
a(C
FU
mL
−1)
0h
4h
24h
0h
4h
24h
0h
4h
24h
L.a
cido
philu
s10
816
.05
±3.
5117
.47
±5.
6119
.70
±1.
3320
.20
±4.
1916
.26
±4.
9116
.85
±1.
6617
.84
±3.
2214
.89
±1.
1516
.45
±3.
69N
CC
1210
74.
25±
2.14
2.95
±2.
574.
19±
3.15
4.66
±2.
313.
58±
2.18
1.69
±0.
814.
21±
3.09
5.02
±2.
822.
92±
2.46
L.a
cido
philu
s10
822
.23
±10
.76
23.4
7±
2.59
22.2
4±
4.67
22.7
3±
6.81
20.3
7±
4.16
25.3
2±
4.15
24.7
8±
1.39
22.9
9±
3.74
23.1
0±
5.19
NC
C36
107
3.44
±3.
043.
74±
3.57
4.95
±1.
882.
34±
1.44
4.69
±0.
842.
85±
1.94
1.80
±1.
201.
20±
1.16
1.75
±1.
51L
.aci
doph
ilus
108
14.2
2±
3.20
16.2
9±
6.56
10.2
2±
1.86
14.1
2±
2.50
12.5
5±
4.38
13.5
0±
3.62
15.9
9±
3.57
14.0
2±
3.10
15.2
8±
3.60
NC
C68
107
0.89
±1.
270.
00±
0.00
1.30
±1.
701.
13±
1.96
0.00
±00
1.82
±1.
621.
56±
1.56
0.48
±0.
841.
92±
0.66
B.b
ifidu
m10
823
.48
±6.
1225
.02
±5.
5526
.65
±2.
6026
.62
±4.
5124
.16
±4.
6424
.35
±4.
3624
.77
±4.
3524
.02
±3.
0326
.33
±1.
82B
b13
107
2.55
±1.
591.
15±
1.99
2.59
±2.
291.
54±
0.80
0.00
±0.
000.
00±
0.00
1.89
±1.
441.
93±
1.71
4.39
±0.
47B
.bifi
dum
108
18.1
1±
2.57
16.6
2±
3.63
18.9
2±
2.84
21.0
8±
6.05
18.2
4±
2.97
19.0
8±
6.07
19.7
9±
7.74
22.0
9±
6.12
18.7
0±
3.69
NC
C38
110
70.
00±
0.00
2.25
±1.
460.
41±
0.72
3.59
±0.
792.
76±
0.63
2.24
±2.
202.
15±
2.35
3.40
±1.
312.
60±
0.64
L.r
ham
nosu
s10
820
.21
±6.
1622
.19
±2.
9722
.16
±7.
1422
.70
±2.
3920
.53
±2.
9623
.96
±2.
1622
.88
±7.
1120
.68
±4.
2921
.64
±1.
6610
73.
44±
0.99
2.90
±2.
773.
17±
1.01
3.88
±0.
702.
59±
2.33
2.42
±0.
424.
10±
1.49
3.73
±1.
493.
68±
0.97
620
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 0
2:00
06
Oct
ober
201
4

Removal of aflatoxin M1 621
Table 2. Removal of aflatoxin M1(AFM1) (5, 10 and 20 ng mL−1) from phosphate buffered saline(PBS) by heat-killed Lactobacillusand Bifidobacterium strains.
% AFM1 removed (mean ± sD, n = 3)
5 ng mL−1 AFM1 10 ng mL−1 AFM1 20 ng mL−1 AFM1
Bacteria 0 h 4 h 24 h 0 h 4 h 24 h 0 h 4 h 24 h
L. acidophilus 24.08 ± 1.58 22.98 ± 4.92 24.95 ± 4.50 20.68 ± 3.06 19.85 ± 3.94 20.21 ± 5.89 24.75 ± 4.23 21.59 ± 5.47 17.03 ± 2.20NCC12
L. acidophilus 26.38 ± 4.99 26.81 ± 1.99 25.29 ± 5.03 22.07 ± 2.23 24.14 ± 3.00 25.28 ± 3.65 26.22 ± 4.93 24.92 ± 3.92 24.50 ± 4.40NCC36
L. acidophilus 16.82 ± 2.37 16.98 ± 4.65 17.57 ± 5.92 15.31 ± 2.92 19.29 ± 2.20 14.04 ± 2.16 20.95 ± 0.51 21.85 ± 4.73 17.72 ± 6.00NCC68
B. bifidum 27.74 ± 2.97 26.42 ± 6.96 25.12 ± 5.33 25.97 ± 5.73 27.36 ± 2.40 25.70 ± 5.21 28.97 ± 3.49 27.68 ± 3.53 24.32 ± 6.99Bb13
B. bifidum 19.46 ± 3.35 22.08 ± 1.41 20.13 ± 6.21 17.39 ± 2.81 22.60 ± 2.97 21.78 ± 2.93 21.19 ± 5.03 23.55 ± 6.11 19.25 ± 5.60NCC 381
L. rhamnosus 23.37 ± 4.81 24.59 ± 2.63 24.16 ± 3.33 26.33 ± 2.41 23.87 ± 2.99 24.98 ± 2.06 27.78 ± 7.50 26.94 ± 2.14 26.69 ± 5.48
by viable Lactobacillus and Bifidobacterium strains at 108
CFU mL−1 in PBS ranged from 10.22 to 26.65%, depend-ing on contamination level and incubation period. WhileB. bifidum Bb13 was the best binder with approximately25% binding, the poorest removal was achieved by Lb. aci-dophilus NCC 68. HPLC chromatograms of PBS contain-ing 5 ng AFM1per mL after 4 h treatment with B. bifidumBb 13 at 108CFU mL−1 and PBS-positive control (20 ngmL−1) are shown in Figure 1. The differences in the bind-ing ability of bacteria at 108 CFU mL−1 were not significantfor 0 and 4 h, except for 24 h in all toxin concentrations. Onthe other hand, a significant decrease (P < 0.05) was ob-served in the percentage of AFM1 removal by test strains at107CFU mL−1 in PBS when compared to their activity at108 CFU mL−1. The AFM1 binding capacity of the strainsat 107 CFU mL−1 was found to range from 0-5.02%, in PBS.There was no significant difference between AFM1-bindingabilities of viable test strains in PBS when used at 107 CFUmL−1 in all incubation periods. Our study clearly indicatesthat bacteria population is one of the most critical factorsin the binding of AFM1 by lactobacilli and bifidobacte-ria. The present study agrees with El-Nezami et al.[8] whoreported that approximately a minimum of 2 × 109CFUmL−1 was required for significant AFB1 removal. SimilarlyLine and Brackett[11] indicated that viable cell populationsof 1 × 109 CFU mL−1 or greater were necessary for sig-nificant removal of AFB1. This amount was similar to thatreported for trichothecenes binding by the strains of Lac-tobacillus and Propionibacterium.[12]
Most information on the binding ability of AFB1 bydairy strains of lactic acid bacteria and bifidobacteria isavailable for AFB1 and only limited for AFM1. These re-sults are lower than that reported by Pierides et al.[13] whofound that the AFM1 binding abilities of viable Lacto-bacillus strains within 15-16 h ranged from 18.1 to 53.8%.Similarly, Peltonen et al.[14] investigated the AFB1bindingability of 12 Lactobacillus, five Bifidobacterium and three
Lactococcus strains in PBS. In their study, Lactobacillusstrains bound 17.3–59.7% AFB1, Bifidobacterium strainsbound 18.0–48.7% AFB1, and Lactococcus strains bound5.6–41.1% AFB1.
The percentages of AFM1 removal by heat-killed bacte-ria from PBS are listed in Table 2. The binding capacitiesof heat-killed bacteria in PBS were found to range from14.04 to 28.97%, depending on toxin concentration and in-cubation level. Heating of the bacterial strains did not sig-nificantly improve their ability to remove AFM1 from PBSexcept for L. acidophilus NCC 12. A slight difference (P <
0.05) in eliminating AFM1 at 5 ng mL−1 from PBS was no-ticed between viable and heat-killed L. acidophilus NCC 12(16.05% and 24.08%, respectively) at 0 h. In a similar work,El-Nezami et al.[8] reported that heat-treated dairy strainsof lactic acid bacteria has the same ability to remove AFB1as viable bacteria. These results also confirm our previousstudy that heating of B. longum Bl 24 and B. longum NCC135 did not significantly alter their ability to bind AFB1 inPBS.[15] However, it has been also reported that heat treat-ment of the bacterial cells improved their ability to removemore AFB1 possibly via protein denaturation or the for-mation of Maillard reaction products between polysaccha-rides and peptides and proteins.[12,16] Such a finding has alsobeen reported by Pierides et al.[13], who found that heat in-activation significantly enhanced the AFM1 removal abilityof 8 Lactobacillus strains except for L. lactis ssp. cremorisARH74.
These results show that bacterial viability is not a pre-requisite in the removal of AFM1 and possible other toxinsby dairy strains of lactic acid bacteria and bifidobacteria.Although the mechanism of aflatoxins removal is still un-known, it is suggested that aflatoxin molecules are boundonto the cell wall components of bacteria. Cell wall polysac-charide and peptidoglycan have been suggested to be thetwo most important elements responsible for the binding oftoxins by lactic acid bacteria.[17] It has been speculated that
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 0
2:00
06
Oct
ober
201
4
622 Kabak and Var
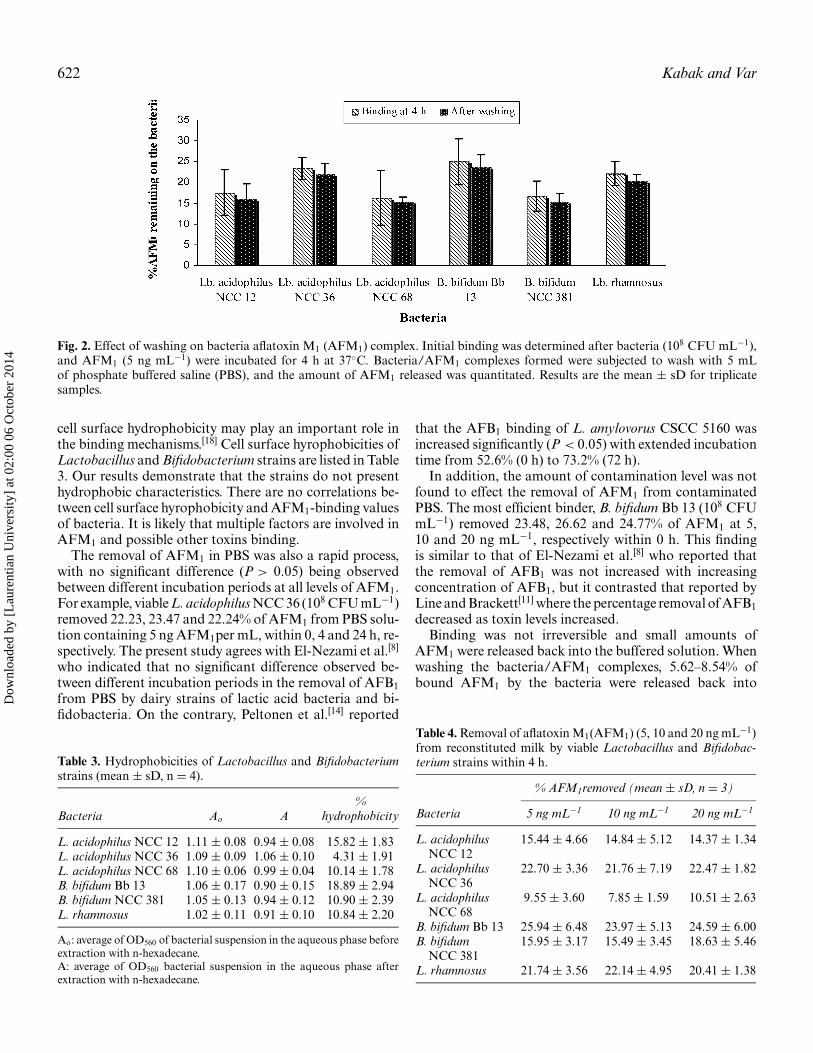
Fig. 2. Effect of washing on bacteria aflatoxin M1 (AFM1) complex. Initial binding was determined after bacteria (108 CFU mL−1),and AFM1 (5 ng mL−1) were incubated for 4 h at 37◦C. Bacteria/AFM1 complexes formed were subjected to wash with 5 mLof phosphate buffered saline (PBS), and the amount of AFM1 released was quantitated. Results are the mean ± sD for triplicatesamples.
cell surface hydrophobicity may play an important role inthe binding mechanisms.[18] Cell surface hyrophobicities ofLactobacillus and Bifidobacterium strains are listed in Table3. Our results demonstrate that the strains do not presenthydrophobic characteristics. There are no correlations be-tween cell surface hyrophobicity and AFM1-binding valuesof bacteria. It is likely that multiple factors are involved inAFM1 and possible other toxins binding.
The removal of AFM1 in PBS was also a rapid process,with no significant difference (P > 0.05) being observedbetween different incubation periods at all levels of AFM1.For example, viable L. acidophilus NCC 36 (108 CFU mL−1)removed 22.23, 23.47 and 22.24% of AFM1 from PBS solu-tion containing 5 ng AFM1per mL, within 0, 4 and 24 h, re-spectively. The present study agrees with El-Nezami et al.[8]
who indicated that no significant difference observed be-tween different incubation periods in the removal of AFB1from PBS by dairy strains of lactic acid bacteria and bi-fidobacteria. On the contrary, Peltonen et al.[14] reported
Table 3. Hydrophobicities of Lactobacillus and Bifidobacteriumstrains (mean ± sD, n = 4).
%Bacteria Ao A hydrophobicity
L. acidophilus NCC 12 1.11 ± 0.08 0.94 ± 0.08 15.82 ± 1.83L. acidophilus NCC 36 1.09 ± 0.09 1.06 ± 0.10 4.31 ± 1.91L. acidophilus NCC 68 1.10 ± 0.06 0.99 ± 0.04 10.14 ± 1.78B. bifidum Bb 13 1.06 ± 0.17 0.90 ± 0.15 18.89 ± 2.94B. bifidum NCC 381 1.05 ± 0.13 0.94 ± 0.12 10.90 ± 2.39L. rhamnosus 1.02 ± 0.11 0.91 ± 0.10 10.84 ± 2.20
Ao: average of OD560 of bacterial suspension in the aqueous phase beforeextraction with n-hexadecane.A: average of OD560 bacterial suspension in the aqueous phase afterextraction with n-hexadecane.
that the AFB1 binding of L. amylovorus CSCC 5160 wasincreased significantly (P < 0.05) with extended incubationtime from 52.6% (0 h) to 73.2% (72 h).
In addition, the amount of contamination level was notfound to effect the removal of AFM1 from contaminatedPBS. The most efficient binder, B. bifidum Bb 13 (108 CFUmL−1) removed 23.48, 26.62 and 24.77% of AFM1 at 5,10 and 20 ng mL−1, respectively within 0 h. This findingis similar to that of El-Nezami et al.[8] who reported thatthe removal of AFB1 was not increased with increasingconcentration of AFB1, but it contrasted that reported byLine and Brackett[11] where the percentage removal of AFB1decreased as toxin levels increased.
Binding was not irreversible and small amounts ofAFM1 were released back into the buffered solution. Whenwashing the bacteria/AFM1 complexes, 5.62–8.54% ofbound AFM1 by the bacteria were released back into
Table 4. Removal of aflatoxin M1(AFM1) (5, 10 and 20 ng mL−1)from reconstituted milk by viable Lactobacillus and Bifidobac-terium strains within 4 h.
% AFM1removed (mean ± sD, n = 3)
Bacteria 5 ng mL−1 10 ng mL−1 20 ng mL−1
L. acidophilus 15.44 ± 4.66 14.84 ± 5.12 14.37 ± 1.34NCC 12
L. acidophilus 22.70 ± 3.36 21.76 ± 7.19 22.47 ± 1.82NCC 36
L. acidophilus 9.55 ± 3.60 7.85 ± 1.59 10.51 ± 2.63NCC 68
B. bifidum Bb 13 25.94 ± 6.48 23.97 ± 5.13 24.59 ± 6.00B. bifidum 15.95 ± 3.17 15.49 ± 3.45 18.63 ± 5.46
NCC 381L. rhamnosus 21.74 ± 3.56 22.14 ± 4.95 20.41 ± 1.38
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 0
2:00
06
Oct
ober
201
4

Removal of aflatoxin M1 623
Table 5. Removal of AFM1 (5, 10 and 20 ng mL−1) from re-constituted milk by heat-killed Lactobacillus and Bifidobacteriumstrains within 4 h.
% AFM1 removed (mean ± sD, n = 3)
Bacteria 5 ng mL−1 10 ng mL−1 20 ng mL−1
L. acidophilus 19.02 ± 0.77 16.60 ± 3.42 18.96 ± 1.35NCC 12
L. acidophilus 23.73 ± 2.52 24.13 ± 4.67 25.07 ± 7.96NCC 36
L. acidophilus 15.36 ± 4.28 12.85 ± 3.09 15.92 ± 5.51NCC 68
B. bifidum 25.41 ± 4.60 25.64 ± 3.18 27.31 ± 1.82Bb 13
B. bifidum 21.36 ± 1.91 17.12 ± 2.64 22.24 ± 7.77NCC 381
L. rhamnosus 25.13 ± 6.19 22.86 ± 9.33 26.27 ± 1.92
the solution (Fig. 2). This is consistent with findings foran efficiently AFB1-binding strain L. rhamnosus GG,[16]
but contrasts that reported by Peltonen et al.,[14] who ob-served that AFB1 was not bound strongly by the lactobacillistrains, and that AFB1was released in the range of 27.8-94.4% of bound AFM1 back into the solution.
The binding abilities of AFM1were found to range from7.85 to 25.94% and from 12.85 to 27.31% for viable (Table 4)and heat-killed bacteria (Table 5) in skim milk, respectively,within 4 h. There were differences between the bacteria intheir ability to bind AFM1in skim milk. However, no sig-nificant differences (P > 0.05) in AFM1 removal ability ofboth viable and heat-killed bacteria were observed betweenPBS and skim milk, except for heat-killed L. acidophilusNCC 68 for 10 ng mL−1 AFM1. Heat-killed L. acidophilusNCC 68 bound 19.29% and 12.85% AFM1 at 10 ng mL−1
in PBS and skim milk, respectively within 4 h (P < 0.05).In a similar work conducted by Pierides et al.[13], viable L.rhamnosus GG showed a decreased capability of AFM1 inboth skim and full cream milk when compared to its ac-tivity in PBS. This finding is contrast to that reported bySarımehmetoglu and Kuplulu,[19] who indicated that yo-gurt bacteria, Streptococcus thermophilus ST-36 and L. del-brueckii subsp. bulgaricus CH-2 removed more AFB1 fromskim milk when compared to their activity in PBS. Brack-ett and Marth[20] reported that an average of 30.7% moreAFM1 was found in milk treated with proteolytic enzymethan in untreated milk and they suggested that AFM1 isbound by milk protein.
When comparing the effect of viability in the binding abil-ity of bacteria in reconstituted milk, the difference was notsignificant (P > 0.05) for 5 and 10 ng mL−1AFM1 within4 h. On the other hand, at a level of 20 ng mL−1 AFM1,differences in the removal ability of L. acidophilus NCC 12and L. rhamnosus were significant. The toxin concentrationhad no effect on the removal of AFM1 levels by both viableand heat-treated bacteria in reconstituted milk.
Conclusions
The results show that test strains were found to not bean efficient agent that could be used to remove AFM1from buffered solutions and reconstituted milk. It has beenclearly shown that bacterial viability is not significant forthe removal of AFM1. The removal of AFM1 from PBS andreconstituted milk was also a rapid process. Furthermore, aminimum of 108 CFU g−1 or mL−1 cell populations are nec-essary for removal of AFM1 from contaminated solutions,without the need for long incubation periods. Further re-search is needed to determine the power of more probioticbacteria to bind to mycotoxins and thus reduce exposureof humans and/or animals to these highly toxic contam-inants in food. In further research, the in vitro digestionmodel or in vivo experiments must be studied to evaluatethe bioaccessibility of mycotoxins in the presence of probi-otic bacteria.
Acknowledgments
This study was supported by the Cukurova University Re-search Council (project no: ZF2004D13). We greatly thankProf. Hans P. van Egmond from National Institute for Pub-lic Health and the Environment for supplying the AFM1standard. We also thank A. Bozdogan for performing sta-tistical analysis.
References
[1] FAQ/WHO. Joint Food and Agriculture Organization/WorldHealth Organization Expert Committee on Food Additives(JECFA). Mycotoxins, Fifty-sixth meeting, Geneva, Switzerland,February 6–15, 2001.
[2] Hussein, H.S.; Brasel, J.M. Toxicity, metabolism and impact of my-cotoxins on humans and animals. Toxicol. 2001, 167, 101–134.
[3] Choudhary, P.L.; Sharma, R.S.; Borkhateria, V.N. Carry-Over ofaflatoxin B1 from feed as aflatoxin M1 in milk of Indian cows. Milch-wissenchaft 1998, 53, 513–515.
[4] International Agency for Research on Cancer (IARC). Some natu-rally occurring substances, food items and constituents, heterocyclicaromatic amines and mycotoxins. 56. World Health Organization:Lyon, France, 1993, p489.
[5] Commission Regulation. EC No 1881/2006 of 19 December 2006.Setting maximum levels for certain contaminants in foodstuffs. Of-ficial Journal of the European Union 2006, L 364, 5–24.
[6] Kabak, B.; Dobson, A.D.W.; Var, I. Strategies to prevent mycotoxincontamination of food and animal feed: a review. Crit. Rev. FoodSci. Nutr. 2006, 46, 593–619.
[7] Bata, A.; Lasztity, R. Detoxification of mycotoxin-contaminatedfood and feed by microorganisms. Trends Food Sci. Techol. 1999,10, 223–228.
[8] El-Nezami, H.; Kankaanpaa, P.; Salminen, S.; Ahokas, J. Ability ofdairy strains of lactic acid bacteria to bind a common food carcino-gen, aflatoxin B1. Food Chem. Toxicol. 1998, 36, 321–326.
[9] Peltonen, K.; El-Nezami, H.; Pierides, M.; Salminen, S.; Ahokas,J.T. Binding of aflatoxin B1 by probiotic bacteria. J. Sci. Food Agric.2000, 80, 1942–1945.
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 0
2:00
06
Oct
ober
201
4

624 Kabak and Var
[10] Vinderola, C.G.; Reinheimer, J.A. Lactic acid starter and probioticbacteria: a comparative “in vitro” study of probiotic characteris-tics and biological barrier resistance. Food Res. Int. 2003, 36, 895–904.
[11] Line, J.E.; Brackett, R.E. Factors affecting aflatoxin B1 removalby Flavobacterium aurantiacum. J. Food Prot. 1995, 58, 91–94.
[12] El-Nezami, H.S.; Chrevatidis, A.; Auriola, S.; Salminen, S.;Mykkanen, H. Removal of common Fusarium toxins in vitro bystrains of Lactobacillus and Propionibacterium. Food Addit. Con-tam. 2002, 19, 680–686.
[13] Pierides, M.; El-Nezami, H.; Peltonen, K.; Salminen, S.; Ahokas, J.Ability of dairy strains of lactic acid bacteria to bind aflatoxin M1
in a food model. J. Food Prot. 2000, 63, 645–650.[14] Peltonen, K.; El-Nezami, H.; Haskard, C.; Ahokas, J.; Salminen,
S. Aflatoxin B1 binding by dairy strains of lactic acid bacteria andbifidobacteria. J. Dairy Sci. 2001, 84, 2152–2156.
[15] Var, I.; Kabak, B. Removal of aflatoxins by viable and heat-killedlactic acid bacteria and bifidobacteria. Arch Lebensmittelhyg. 2004,55, 106–109.
[16] Haskard, C.; El-Nezami, H.S.; Kankaanpaa, P.E.; Salminen, S.E.;Ahokas, J.T. Surface binding of aflatoxin B1 by lactic acid bacteria.Appl. Environ. Microbiol. 2001, 67, 2086–3091.
[17] Bolognani, F.; Rumney, C.J.; Rowland I.R. Influence of carcinogenbinding by lactic acid-producing bacteria on tissue distribution andin vivo mutagenicity of dietary carcinogens. Food Chem. Toxicol.1997, 35, 535–545.
[18] Oatley, J.T.; Rarick, M.D.; Ji, G.E.; Linz, J.E. Binding of aflatoxinB1 to Bifidobacteria in vitro. J. Food Prot. 2000, 63, 1133–1136.
[19] Sarımehmetoglu, B.; Kuplulu, O. Binding ability of aflatoxin M1 toyoghurt bacteria. Ankara Universitesi Veterinerlik Fakultesi Dergisi2004, 51, 195–198.
[20] Brackett, R.E.; Marth, E.H. Association of aflatoxin M1 with casein.Z. Lebensm. Unters Forsch. 1997, 174, 439–441.
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 0
2:00
06
Oct
ober
201
4





















![endomikro1a - TavsiyeEdiyorum.comdönemde floraya egemen bakteriler; Streptococcus, Lactobacillus, Bifidobacterium, Actinomyces, RothiD \HOHULGLU ED]HQ 3HSWRVWUHSWRFRFFXVODUD GD UDVWODQÕU](https://img.pdfslide.us/doc/110x75/5e45b1c79475cc29303e1a4c/endomikro1a-dnemde-floraya-egemen-bakteriler-streptococcus-lactobacillus.jpg)






