-
8/4/2019 FaCEN_nucleo celular
1/8
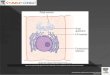
Figure 4-9. (Alberts4e) A cross-sectional view of a typical cell
nucleus. The nuclear envelope consists of two membranes, the outer
one being continuous with the endoplasmicreticulum membrane (see
also Figure 12-9). The space inside the endoplasmic reticulum (the
ER lumen) is coloredyellow; it is continuous with the space between
the two nuclear membranes.
The lipid bilayers of the inner and outer nuclear membranes are
connected at each nuclear pore. Two networks of intermediate
filaments (green) provide mechanical support for the nuclear
envelope; the intermediate filaments inside the nucleus form a
special supporting structure called the nuclear lamina.
UNIVERSIDAD NACIONAL
DE ASUNCIN
FACULTAD DE CIENCIAS
EXACTAS Y NATURALES
CTEDRA DE
BIOLOGA CELULAR
PROF. LIC. GLORIA YALUFF
ELABORADO POR
DANILO FERNNDEZ ROS
http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=mboc4.figgrp.2155
-
8/4/2019 FaCEN_nucleo celular
2/8
Figure 12-10. (Alberts4e) The arrangement of nuclear pore
complexes in the nuclear envelope.(A) A small region of the nuclear
envelope. In cross section, a nuclear pore complex seems to have
four structural building blocks:
column subunits, which form the bulk of the pore wall; annular
subunits, which extend "spokes" (not shown) toward the center of
the
pore; lumenal subunits, which contain transmembrane proteins
that anchor the complex to the nuclear membrane; and ring
subunits,
which form the cytosolic and nuclear faces of the complex. In
addition, fibrils protrude from both the cytosolic and the nuclear
sides
of the complex. On the nuclear side, the fibrils converge to
form basketlike structures. Localization studies using
immunoelectron
microscopy techniques showed that the proteins that make up the
core of the nuclear pore complex are symmetrically distributed
across the nuclear envelope so that the nuclear and cytosolic
sides look identical. This is in contrast to proteins that make up
the
fibrils, which are different on each side of the cytosolic or
the nuclear side.
(B) A scanning electron micrograph of the nuclear side of the
nuclear envelope of an oocyte.
(C) The continuity of the inner and outer nuclear membrane at
the pore is apparent in this thin section electron micrograph,
showinga side view of two nuclear pore complexes (brackets).
(D) This electron micrograph shows face-on views of negatively
stained nuclear pore complexes from which the membrane has been
removed by detergent extraction. (B, from M.W. Goldberg and T.D.
Allen, J. Cell Biol. 119:1429 1440, 1992. The Rockefeller
University Press; C, courtesy of Werner Franke and Ulrich
Scheer; D, courtesy of Ron Milligan.)
Figure 12-11. (Alberts4e) Possible paths for free diffusion
through the nuclear pore complex. This drawing shows ahypothetical
diaphragm (gray) inserted into the pore to restrict the size of the
open channel to 9 nm, the pore size estimated from diffusion
measurements. Nine nanometers is a much smaller diameter than
that of the central opening apparent on the images of the nuclear
pore complex
derived from electron micrographs. It is also smaller than the
opening estimated during active transport, when the pore dilates to
allow the transport ofparticles of up to 26 nm in diameter (arrow).
Thus, it is likely that some pore components are lost during the
preparation of specimens for electron
microscopy, and that these normally restrict free diffusion
through the central opening. Such components may form a diaphragm
(or plug) that opens
and closes to allow the passage of large objects during active
transport, which depends on sorting signals (discussed below).
Although plugs can beseen in some preparations, it is not clear
whether they are components of the pore complex or material that is
being transported through it. Three-
dimensional computer reconstructions suggest that the channels
permitting free diffusion might be located near the rim of the pore
complex, between
the column subunits, rather than at its center (see Figure
12-10A); this would mean that passive diffusion and active
transport take place through
different parts of the complex.
UNIVERSIDAD NACIONAL
DE ASUNCIN
FACULTAD DE CIENCIAS
EXACTAS Y NATURALES
CTEDRA DE
BIOLOGA CELULAR
PROF. LIC. GLORIA YALUFF
ELABORADO POR
DANILO FERNNDEZ ROS
http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=mboc4.figgrp.2157
-
8/4/2019 FaCEN_nucleo celular
3/8
-
8/4/2019 FaCEN_nucleo celular
4/8
Figure 4-22. (Alberts4e) The three DNA sequences required to
produce a eucaryotic chromosome thatcan be replicated and then
segregated at mitosis.
UNIVERSIDAD NACIONAL
DE ASUNCIN
FACULTAD DE CIENCIAS
EXACTAS Y NATURALES
CTEDRA DE
BIOLOGA CELULAR
PROF. LIC. GLORIA YALUFF
ELABORADO POR
DANILO FERNNDEZ ROS
-
8/4/2019 FaCEN_nucleo celular
5/8
Figure 4-25. (Alberts4e) The structure of a
nucleosome core particle, as determined by
x-ray diffraction analyses of crystals. Eachhistone is colored
according to the scheme of
Figure 4-24, with the DNA double helix in light
gray. (Reprinted by permission from K. Luger etal., Nature
389:251 260, 1997. Macmillan
Magazines Ltd.)
Figure 4-24. (Alberts4e) Structural organization of the
nucleosome. A nucleosome contains a proteincore made of eight
histone molecules. As indicated, the nucleosome core particle is
released from chromatin by
digestion of the linker DNA with a nuclease, an enzyme that
breaks down DNA. (The nuclease can degrade the
exposed linker DNA but cannot attack the DNA wound tightly
around the nucleosome core.) After dissociation of
the isolated nucleosome into its protein core and DNA, the
length of the DNA that was wound around the corecan be determined.
This length of 146 nucleotide pairs is sufficient to wrap 1.65
times around the histone core.
UNIVERSIDAD NACIONAL
DE ASUNCIN
FACULTAD DE CIENCIAS
EXACTAS Y NATURALES
CTEDRA DE
BIOLOGA CELULAR
PROF. LIC. GLORIA YALUFF
ELABORADO PORDANILO FERNNDEZ ROS
http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=mboc4.figgrp.632
-
8/4/2019 FaCEN_nucleo celular
6/8
Figura 10-24. (Lodish5e) Modelo de empaquetamiento de la
cromatina y del armazn cromosmico en los cromosomas en metafase. En
loscromosomas en interfase, largas extensiones de cromatina de 30
nm forman bucles hacia fuera de los armazones extendidos. En los
cromosomas metafsicos, el armazn se
pliega adicionalemente para formar una estructura muy compacta
cuya geometra precisa an no se ha determinado.
UNIVERSIDAD NACIONAL
DE ASUNCIN
FACULTAD DE CIENCIAS
EXACTAS Y NATURALES
CTEDRA DE
BIOLOGA CELULAR
PROF. LIC. GLORIA YALUFF
ELABORADO POR
DANILO FERNNDEZ ROS
-
8/4/2019 FaCEN_nucleo celular
7/8
Figura 10-1. (Lodish5e) Vista general de
la estructura de genes y cromosomas
UNIVERSIDAD NACIONAL
DE ASUNCIN
FACULTAD DE CIENCIAS
EXACTAS Y NATURALES
CTEDRA DE
BIOLOGA CELULAR
PROF. LIC. GLORIA YALUFF
ELABORADO POR
DANILO FERNNDEZ ROS
-
8/4/2019 FaCEN_nucleo celular
8/8
FIGURA EXPERIMENTAL 10-23. (Lodish5e) Una micrografa electrnica
de un
cromosoma en metafase sin histonas revela el armazn alrededor
del cual seorganiza el DNA. Los bucles largos de DNA son visibles
extendindose del armaznde protena no histona (la estructura
oscura). La forma del armazn refleja la del
mismo cromosoma metafsico. El cromosoma fue preparado a partir
de clulas HeLa
mediante tratamiento con un detergente suave. (De J. R. Paulson
y U. K. Laemmli,
1977, Cell 12:817. Copyright 1977 MIT.)
UNIVERSIDAD NACIONAL
DE ASUNCIN
FACULTAD DE CIENCIAS
EXACTAS Y NATURALES
CTEDRA DE
BIOLOGA CELULAR
PROF. LIC. GLORIA YALUFF
ELABORADO POR
DANILO FERNNDEZ ROS