Embed Size (px)
Citation preview

F1Fo ATP Synthase
Copyright © 1999-2007 by Joyce J. Diwan. All rights reserved.
Molecular Biochemistry I
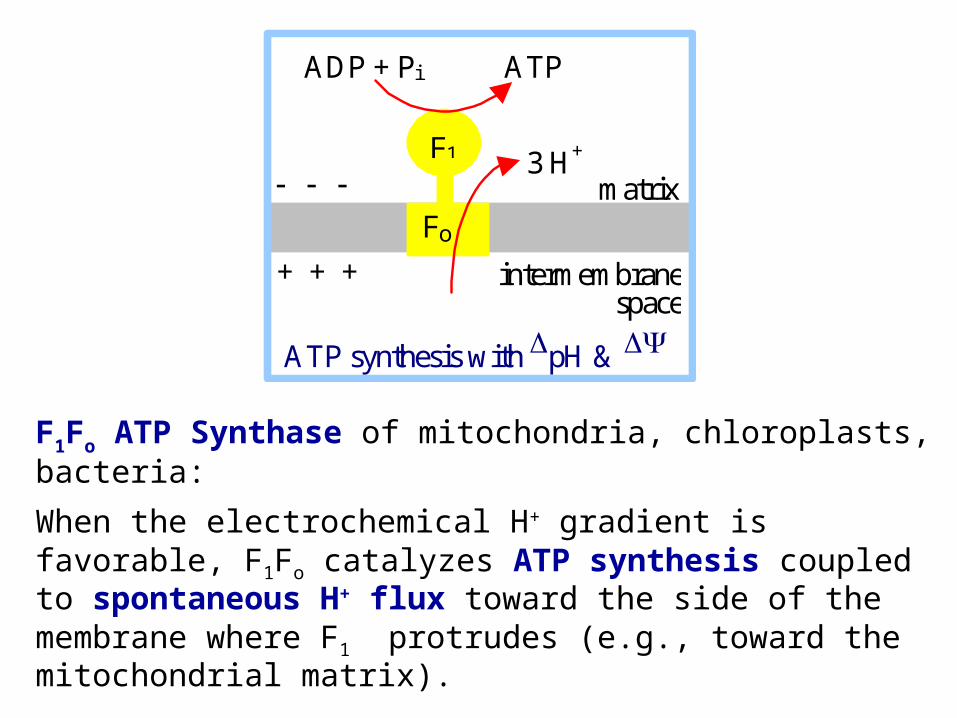
F1Fo ATP Synthase of mitochondria, chloroplasts, bacteria:
When the electrochemical H+ gradient is favorable, F1Fo catalyzes ATP synthesis coupled to spontaneous H+ flux toward the side of the membrane where F1 protrudes (e.g., toward the mitochondrial matrix).
ADP + Pi ATP
F1
Fo
3 H+ matrix
intermembrane space
ATP synthesis with pH &
+ + +
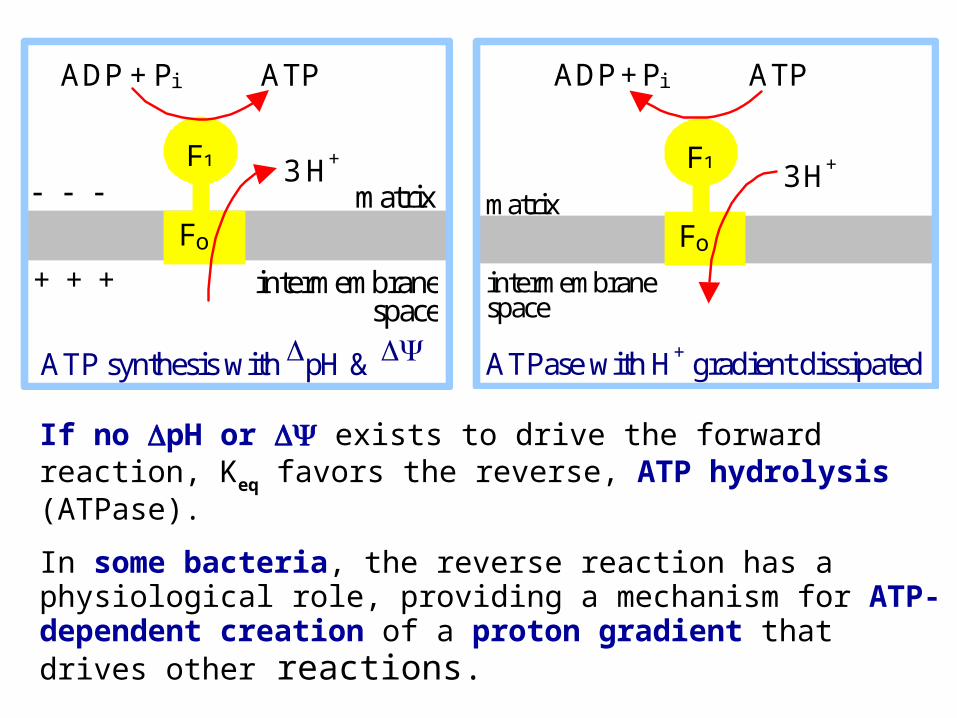
If no pH or exists to drive the forward reaction, Keq favors the reverse, ATP hydrolysis (ATPase).
In some bacteria, the reverse reaction has a physiological role, providing a mechanism for ATP-dependent creation of a proton gradient that drives other reactions.
ADP + Pi ATP
F1
Fo
3 H+
ATPase with H+ gradient dissipated
matrix
intermembrane space
ADP + Pi ATP
F1
Fo
3 H+ matrix
intermembrane space
ATP synthesis with pH &
+ + +

Inhibitors of F1Fo, that block H+ transport coupled to ATP synthesis or hydrolysis, include:
oligomycin, an antibiotic
DCCD (dicyclohexylcarbodiimide), a reagent that reacts with carboxyl groups in hydrophobic environments, forming a covalent adduct.

Roles of major subunits were determined in studies of submitochondrial particles (SMP).
In mitochondria treated with ultrasound, inner membrane breaks & reseals as vesicles, with F1 on the outer surface.
Since F1 of intact mitochondria faces the matrix, these SMP are said to be inside out.
By EM with negative stain, ATP Synthase appeared as "lollipops" on the inner mitochondrial membrane, facing the matrix.
Higher resolution cryo-EM later showed each lollipop to have 2 stalks. See Movie.
mitochondrion ultrasound
SMP F1

After removal of F1, the SMP membrane containing Fo is leaky to H+.
Adding back F1 restores normal low permeability to H+.
Thus it was established that Fo includes a “H+ channel."
F1, the lollipop head, when extracted from SMP, catalyzes ATP hydrolysis (spontaneous reaction in the absence of an energy input).
Thus F1 contains the catalytic domain(s).
mitochondrion ultrasound
SMP F1
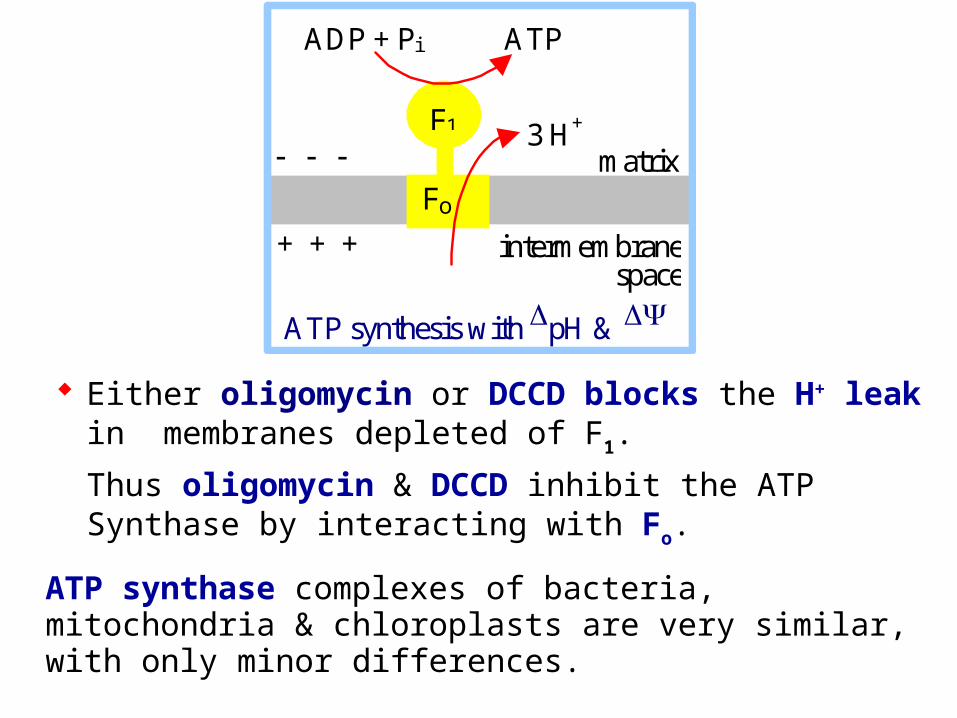
Either oligomycin or DCCD blocks the H+ leak in membranes depleted of F1.
Thus oligomycin & DCCD inhibit the ATP Synthase by interacting with Fo.
ATP synthase complexes of bacteria, mitochondria & chloroplasts are very similar, with only minor differences.
ADP + Pi ATP
F1
Fo
3 H+ matrix
intermembrane space
ATP synthesis with pH &
+ + +
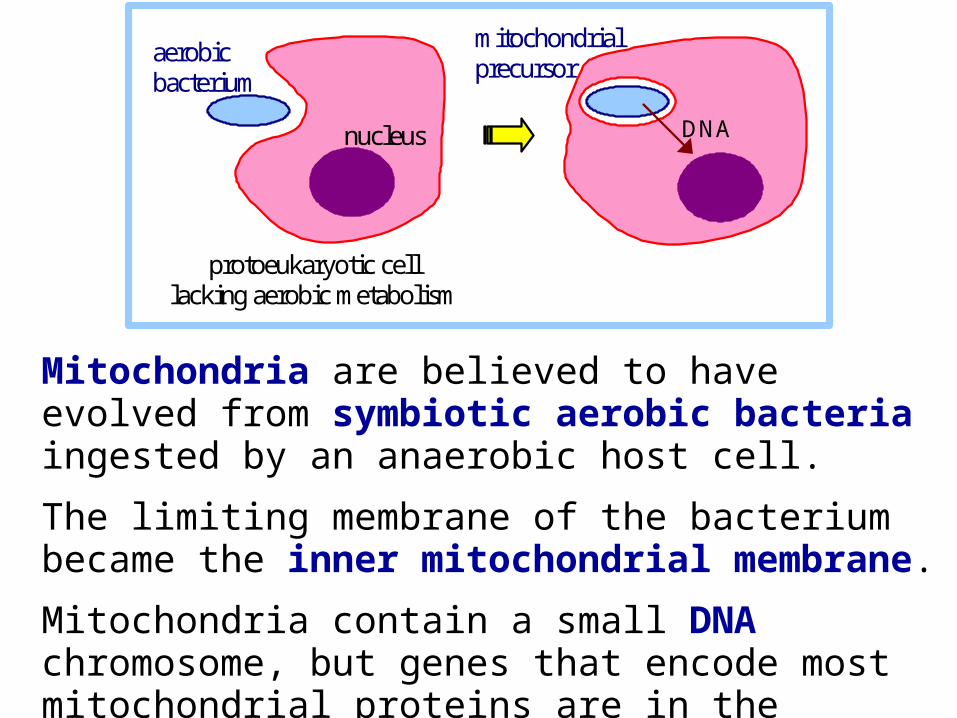
Mitochondria are believed to have evolved from symbiotic aerobic bacteria ingested by an anaerobic host cell.
The limiting membrane of the bacterium became the inner mitochondrial membrane.
Mitochondria contain a small DNA chromosome, but genes that encode most mitochondrial proteins are in the nucleus, consistent with transfer of some DNA during evolution.
aerobic bacterium
protoeukaryotic cell lacking aerobic metabolism
DNA
mitochondrial precursor
nucleus

Looking down at the membrane, & subunits alternate around a ring ( to be discussed later.)
The subunit composition of the ATP Synthase was first established for E. coli, which has an operon that encodes genes for all subunits.
Stalk subunits were classified initially as part of F1 or Fo, based on whether they co-purified with extracted F1.
F1 subunits were named with Greek letters in order of decreasing MW.
They are present with stoichiometry 3, 3, , , .
The & subunits (513 & 460 aa residues in E. coli) are homologous to one another.
F1 in cross
section
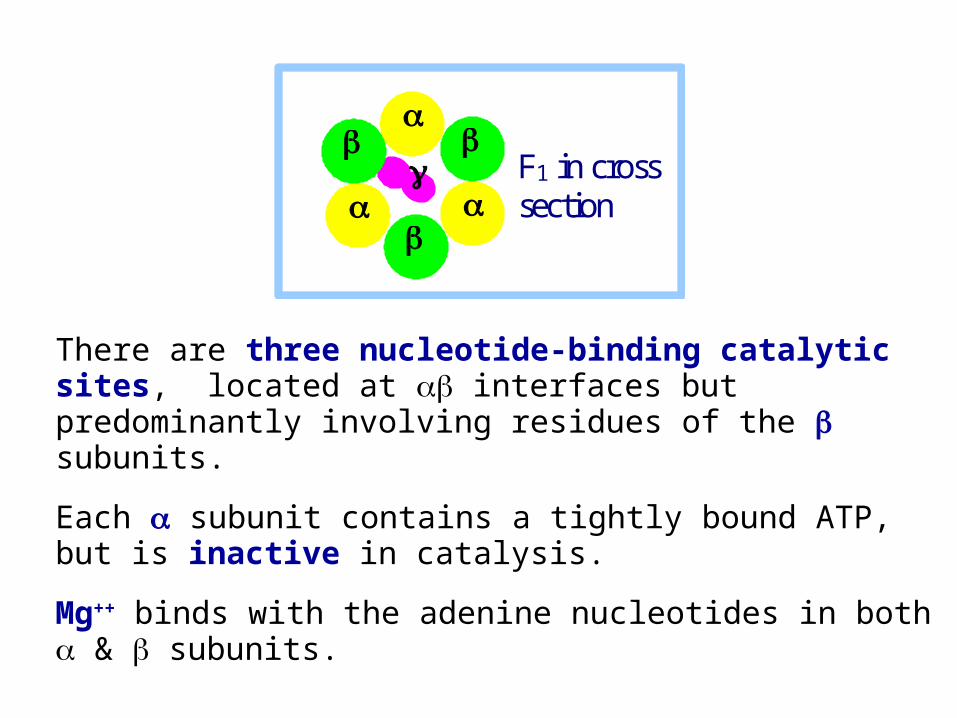
There are three nucleotide-binding catalytic sites, located at interfaces but predominantly involving residues of the subunits.
Each subunit contains a tightly bound ATP, but is inactive in catalysis.
Mg++ binds with the adenine nucleotides in both & subunits.
F1 in cross
section

Fo subunits were named in Roman letters with decreasingmolecular weight.
Stoichiometry of these subunits in E. coli Fo is a, b2, c10.
Mammalian mitochondrial F1Fo is slightly more complex than the bacterial enzyme, with a few additional subunits.
Also, since names were assigned based on apparent molecular weights, some subunits were given different names in different organisms.
ADP + Pi ATP
F1
Fo
3 H+ + + +

Bovine subunit turned out to be homologous to the E. coli subunit.
Bovine subunit is unique.
A bovine subunit called OSCP (oligomycin sensitivity conferral protein) is homologous to the E. coli subunit.
The bovine enzyme has additional subunits d & F6.
There is evidence that the ATP Synthase (F1Fo) may form a complex with adenine nucleotide translocase (ADP/ATP antiporter) & phosphate carrier (Pi/H+ symporter).
This complex has been designated the ATP Synthasome.

The binding change mechanism of energy coupling was proposed by Paul Boyer.
He shared the Nobel prize for this model that accounts for the existence of 3 catalytic sites in F1.
For simplicity, only the catalytic subunits are shown. It is proposed that an irregularly shaped shaft linked
to Fo rotates relative to the ring of 3 subunits.
The rotation is driven by flow of H+ through Fo.
ADP + Pi ATP
ATP
ATP ADP + Pi
ATP ADP + Pi
ATP open tight
binding
loose binding
repeat
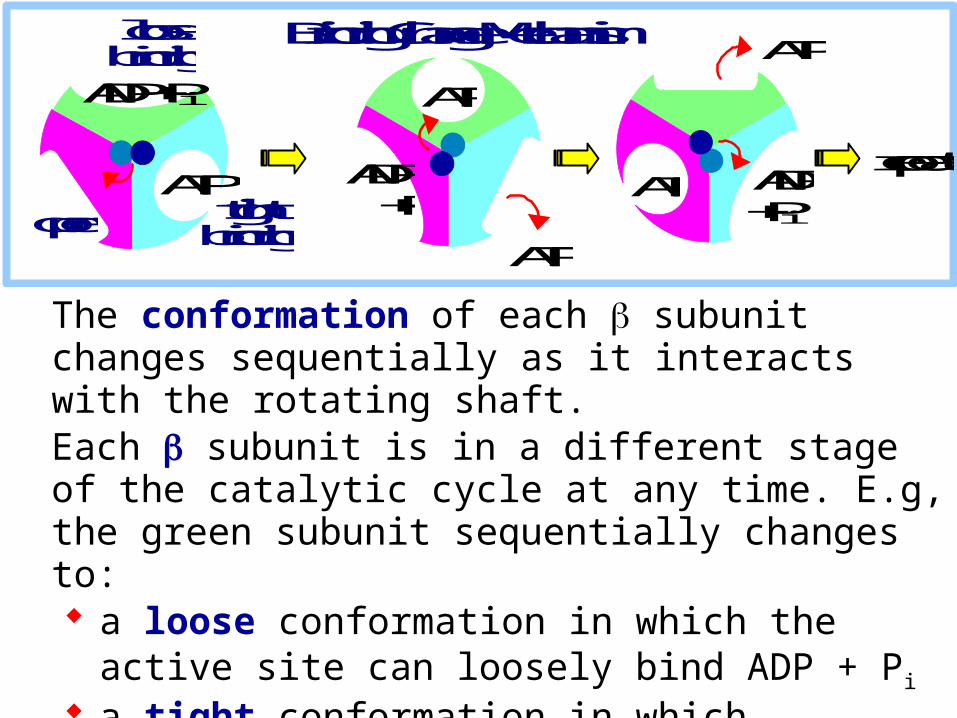
Binding Change Mechanism
ADP + Pi ATP
ATP
ATP ADP + Pi
ATP ADP + Pi
ATP open tight
binding
loose binding
repeat
Binding Change Mechanism
The conformation of each subunit changes sequentially as it interacts with the rotating shaft. Each subunit is in a different stage of the catalytic cycle at any time. E.g, the green subunit sequentially changes to: a loose conformation in which the active site can loosely
bind ADP + Pi
a tight conformation in which substrates are tightly bound and ATP is formed
an open conformation that favors ATP release.

The subunit includes a bent helical loop that constitutes a "shaft" within the ring of & subunits.
Shown is bovine F1 treated with DCCD to yield crystals in which more of the central stalk is ordered, allowing structure determination. Colors: , , , , .
F1 ATPase PDB 1E79
side view base view
Supporting evidence:
1. The crystal structure of F1 with the central stalk was solved by J. Walker, who shared the Nobel prize.

Note the wide base of the rotary shaft, including part of as well as and subunits.
Recall that the bovine subunit, which is at the base of the shaft, is equivalent to of bacterial F1.
F1 ATPase PDB 1E79
side view base view
Bovine F1 (DCCD-treated)
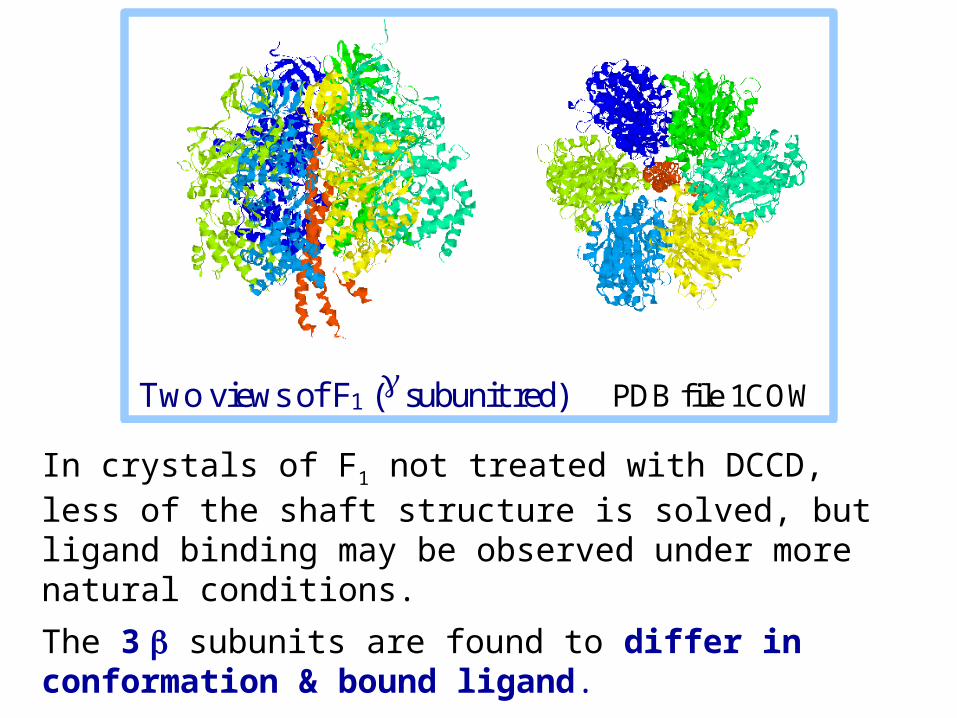
In crystals of F1 not treated with DCCD, less of the shaft structure is solved, but ligand binding may be observed under more natural conditions.
The 3 subunits are found to differ in conformation & bound ligand.
Two views of F1 ( subunit red) PDB file 1COW
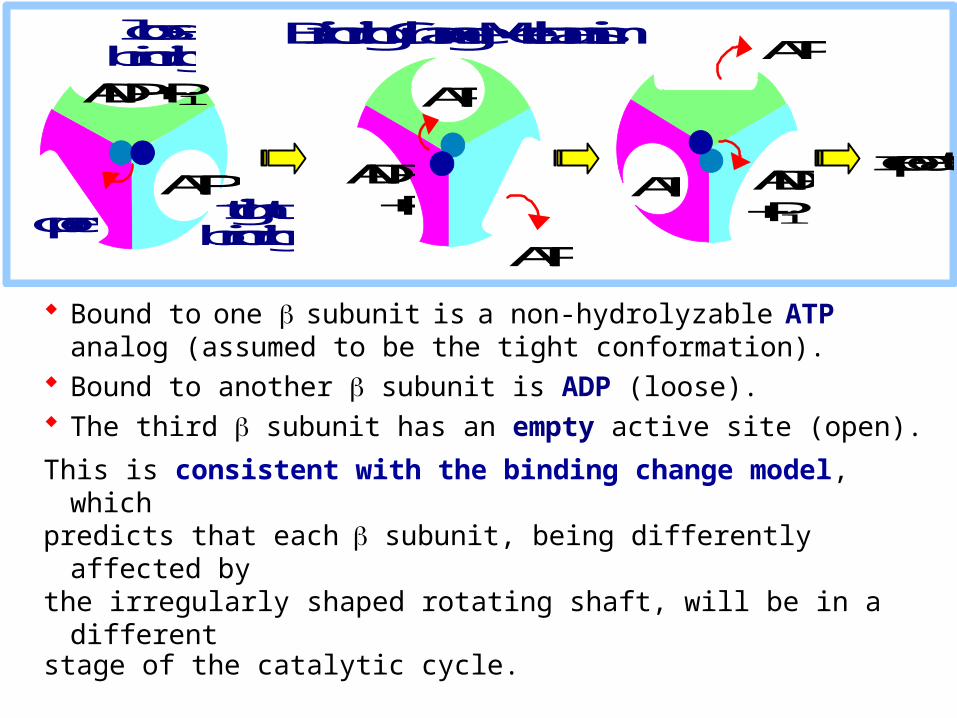
Bound to one subunit is a non-hydrolyzable ATP analog (assumed to be the tight conformation).
Bound to another subunit is ADP (loose). The third subunit has an empty active site (open).
This is consistent with the binding change model, whichpredicts that each subunit, being differently affected bythe irregularly shaped rotating shaft, will be in a different stage of the catalytic cycle.
ADP + Pi ATP
ATP
ATP ADP + Pi
ATP ADP + Pi
ATP open tight
binding
loose binding
repeat
Binding Change Mechanism
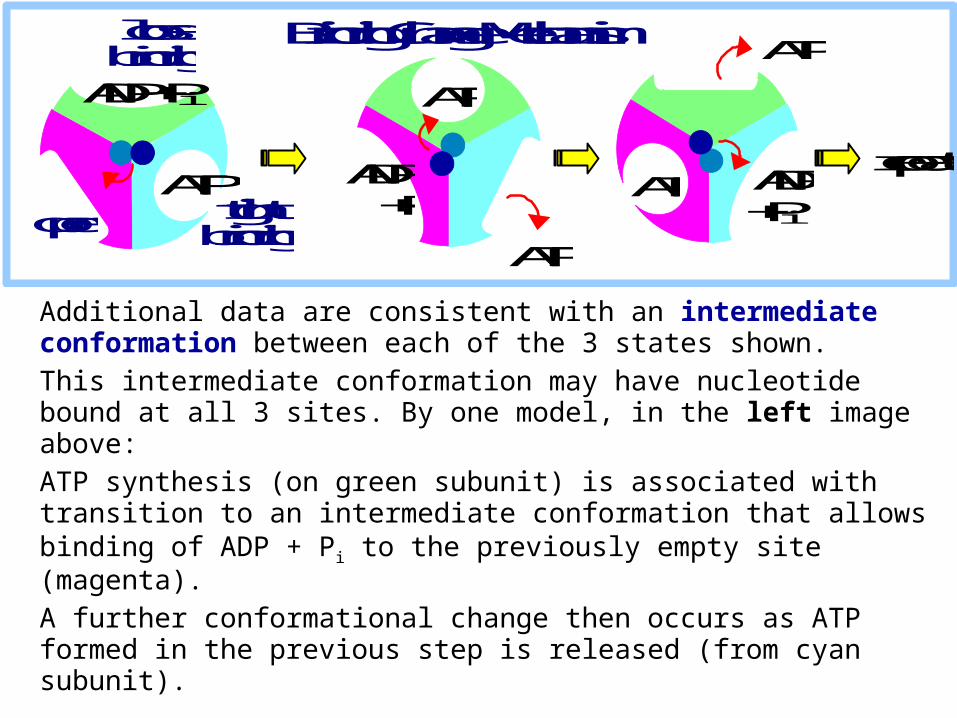
Additional data are consistent with an intermediate conformation between each of the 3 states shown.
This intermediate conformation may have nucleotide bound at all 3 sites. By one model, in the left image above:
ATP synthesis (on green subunit) is associated with transition to an intermediate conformation that allows binding of ADP + Pi to the previously empty site (magenta).
A further conformational change then occurs as ATP formed in the previous step is released (from cyan subunit).
ADP + Pi ATP
ATP
ATP ADP + Pi
ATP ADP + Pi
ATP open tight
binding
loose binding
repeat
Binding Change Mechanism
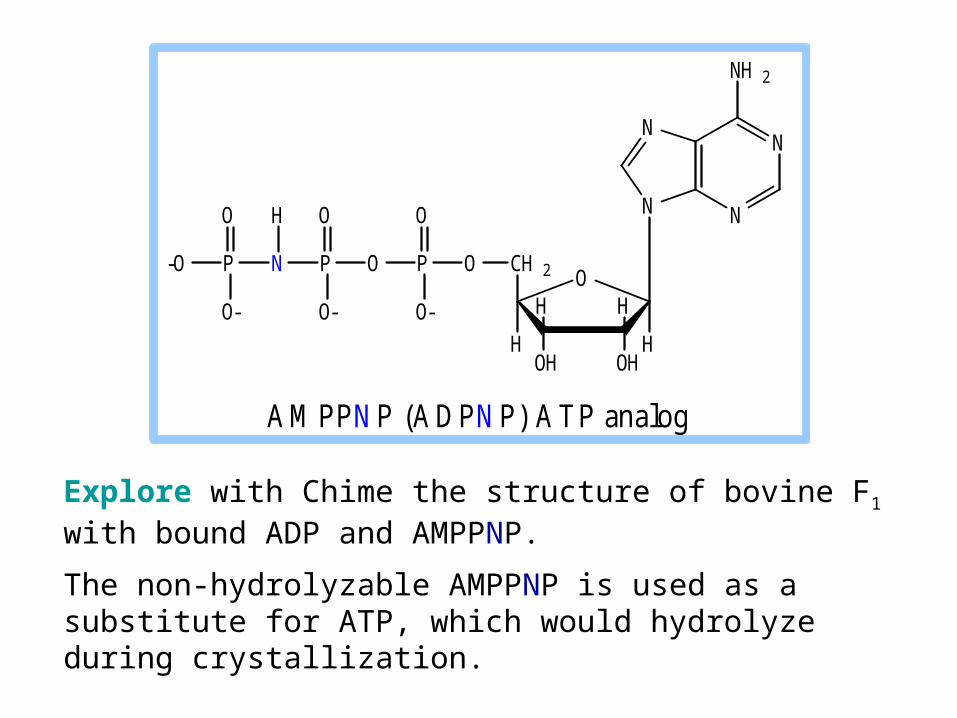
Explore with Chime the structure of bovine F1 with bound ADP and AMPPNP.
The non-hydrolyzable AMPPNP is used as a substitute for ATP, which would hydrolyze during crystallization.
A M PP N P (A D PN P) A T P analog
N
NN
N
NH 2
O
OHOH
HH
H
CH 2
H
OPOPNP-O
O
O- O-
O O
O-
H
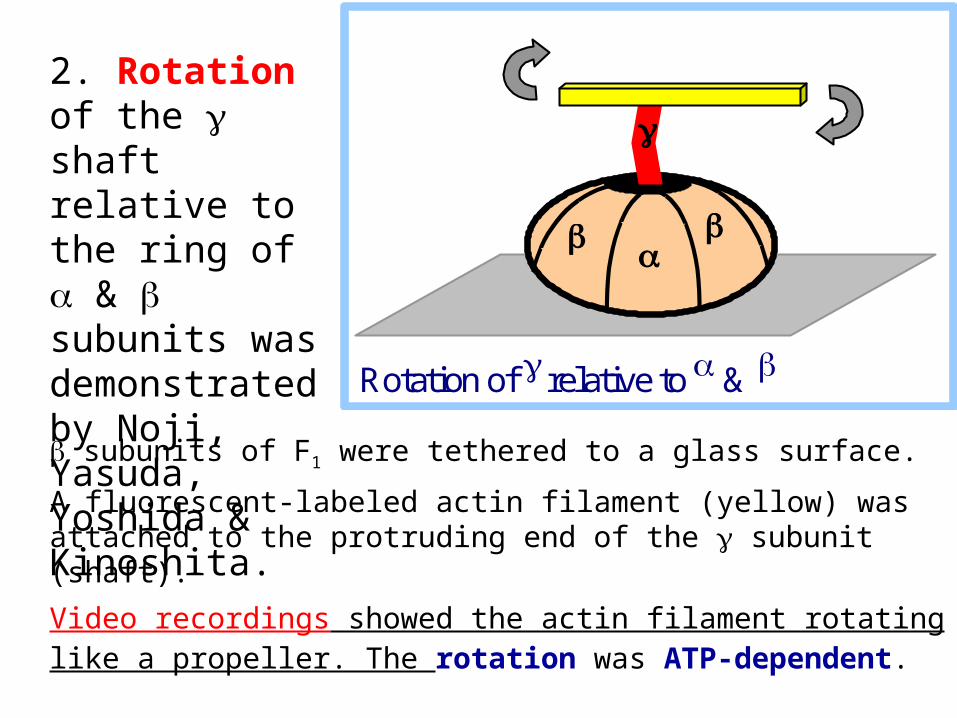
subunits of F1 were tethered to a glass surface.
A fluorescent-labeled actin filament (yellow) was attached to the protruding end of the subunit (shaft).
Video recordings showed the actin filament rotating like a propeller. The rotation was ATP-dependent.
Rotation of relative to &
2. Rotation of the shaft relative to the ring of & subunits was demonstrated by Noji, Yasuda, Yoshida & Kinoshita.

Some observations indicate that each 120o step consists of 80-90o & 30-40o substeps, with a brief intervening pause.
Such substeps are consistent with evidence for an intermediate conformation between the major transitions, discussed above.
Rotation of relative to &
Studies using varied techniques have shown ATP-induced rotation to occur in discrete 120o steps, with intervening pauses.

Although the binding change mechanism is widely accepted, some details of the reaction cycle are still debated.
View an animation of ATP synthesis based on observed variation in conformation of F1 subunits attributed to rotation of the shaft.
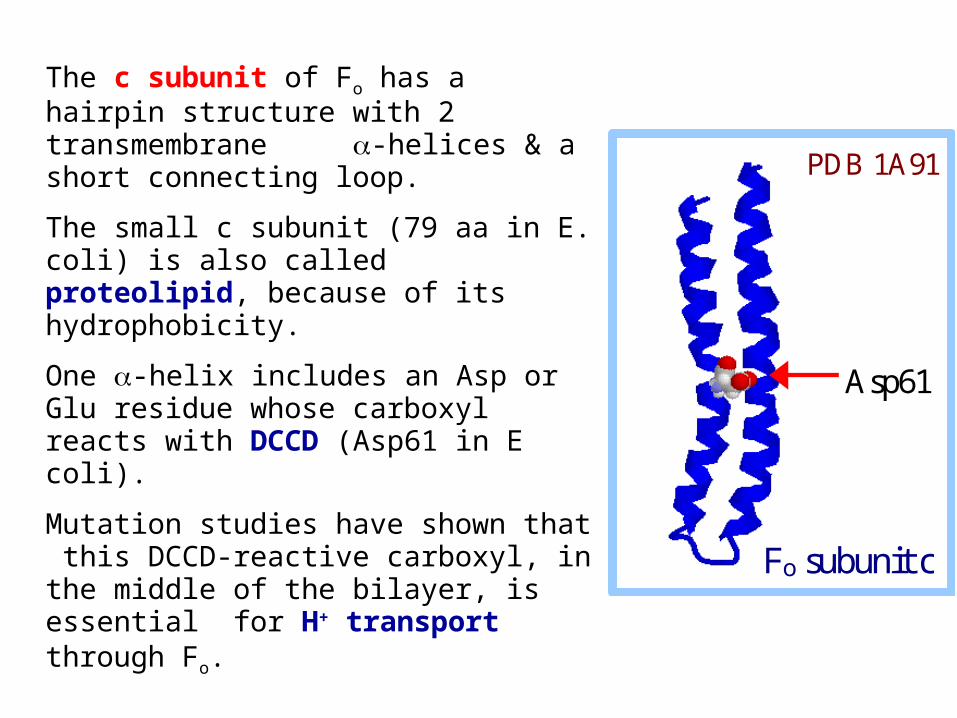
The c subunit of Fo has a hairpin structure with 2 transmembrane -helices & a short connecting loop.
The small c subunit (79 aa in E. coli) is also called proteolipid, because of its hydrophobicity.
One -helix includes an Asp or Glu residue whose carboxyl reacts with DCCD (Asp61 in E coli).
Mutation studies have shown that this DCCD-reactive carboxyl, in the middle of the bilayer, is essential for H+ transport through Fo.
Asp61
Fo subunit c
PDB 1A91

At right: a low resolution partial structure of yeast F1 with central stalk & attached Fo c subunits.
View this file by Chime.
Count the number of Fo c subunits, arranged in a ring.
Look for the Asp near the middle of one transmembrane segment of each c subunit.
Partial structures of F1, Fo
PDB 1Q01

An atomic resolution structure of the complete ATP Synthase, including F1 and Fo with peripheral as well as central stalks, has not yet been achieved.
However partial or complete structures of individual protein constituents, mutational studies, and evidence for inter-subunit interactions, have defined the roles of most subunits.
Partial structures of F1, Fo
PDB 1Q01
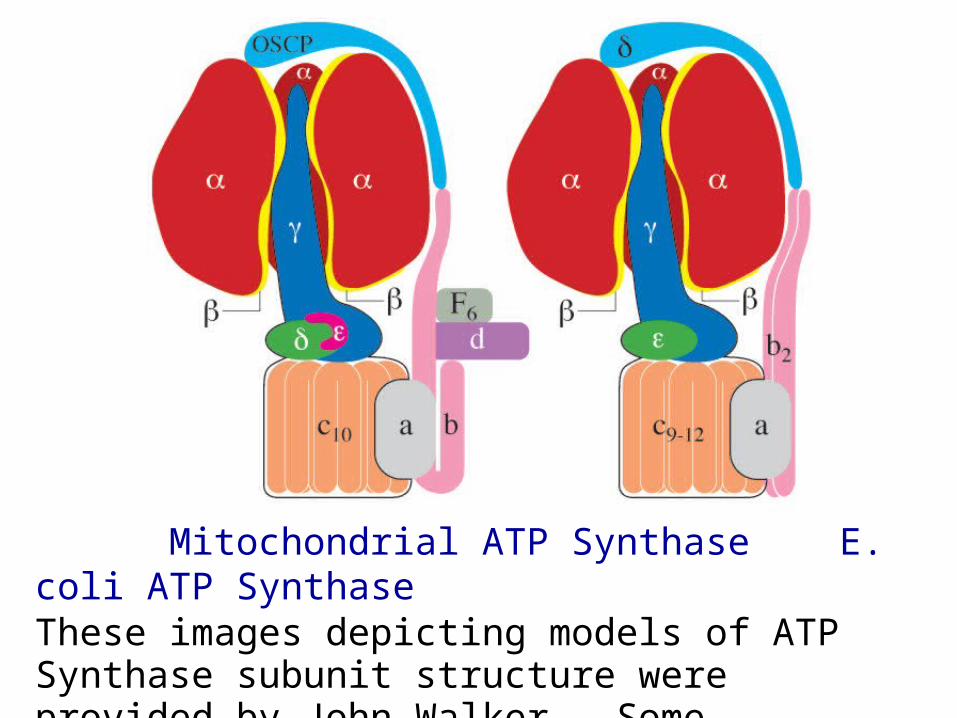
Mitochondrial ATP Synthase E. coli ATP SynthaseThese images depicting models of ATP Synthase subunit structure were provided by John Walker. Some equivalent subunits from different organisms have different names.
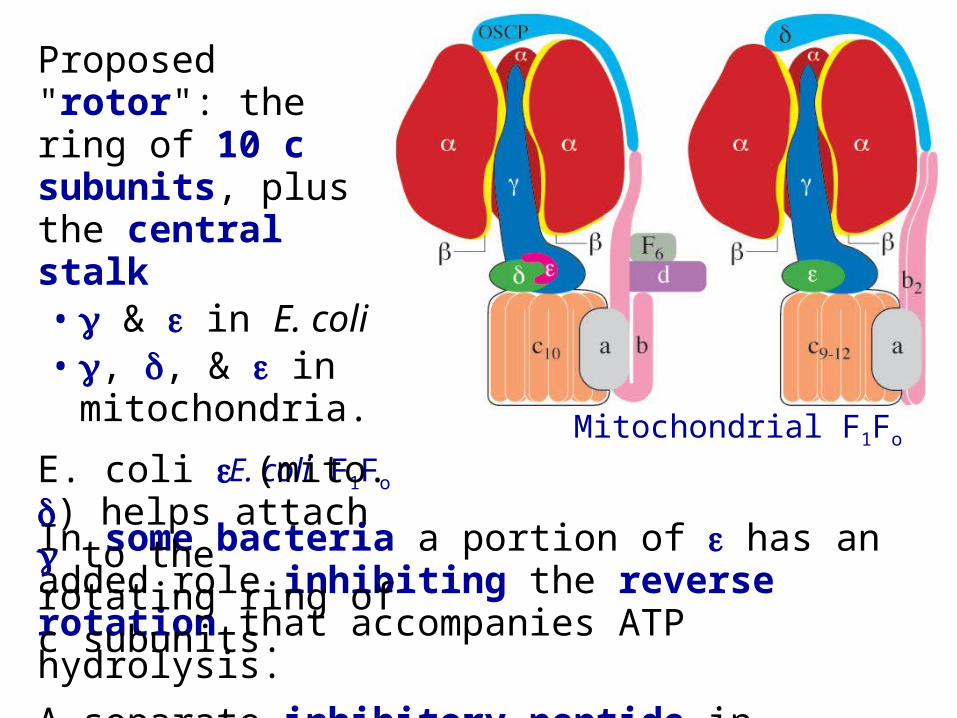
Mitochondrial F1Fo E. coli F1Fo
In some bacteria a portion of has an added role inhibiting the reverse rotation that accompanies ATP hydrolysis.
A separate inhibitory peptide in mitochondria prevents F1Fo from hydrolyzing ATP when there is no H+ gradient to drive ATP synthesis, e.g., under anoxic conditions.
Proposed "rotor": the ring of 10 c subunits, plus the central stalk• & in E. coli• , , & in
mitochondria.
E. coli (mito. ) helps attach to the rotating ring of c subunits.
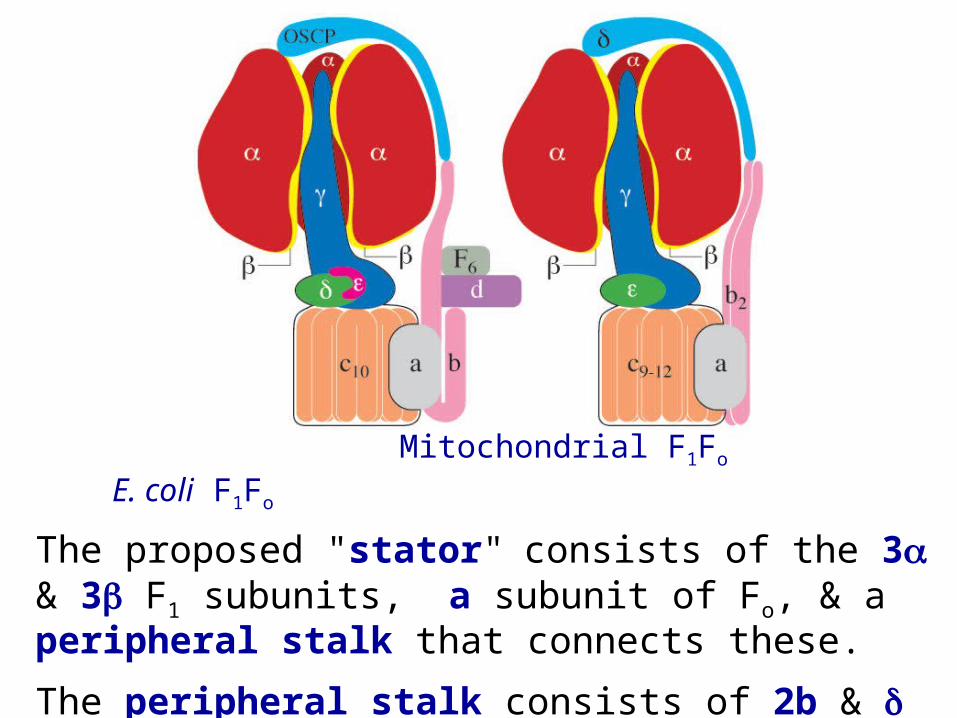
Mitochondrial F1Fo E. coli F1Fo
The proposed "stator" consists of the 3 & 3 F1 subunits, a subunit of Fo, & a peripheral stalk that connects these.
The peripheral stalk consists of 2b & in E. coli or subunits b, d, F6, & OSCP in bovine mitochondria.
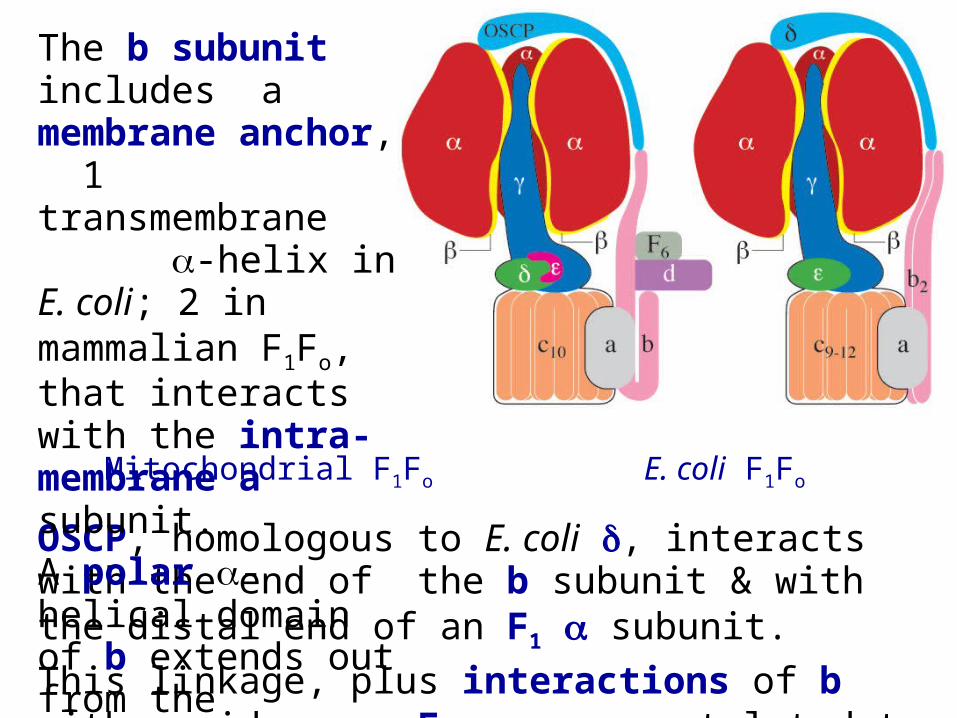
Mitochondrial F1Fo E. coli F1Fo
OSCP, homologous to E. coli , interacts with the end of the b subunit & with the distal end of an F1 subunit.
This linkage, plus interactions of b with residues on F1, are postulated to hold back the ring of & subunits, keeping it from rotating along with the central stalk.
The b subunit includes a membrane anchor, 1 transmembrane -helix in E. coli; 2 in mammalian F1Fo, that interacts with the intra-membrane a subunit.
A polar -helical domain of b extends out from the membrane.
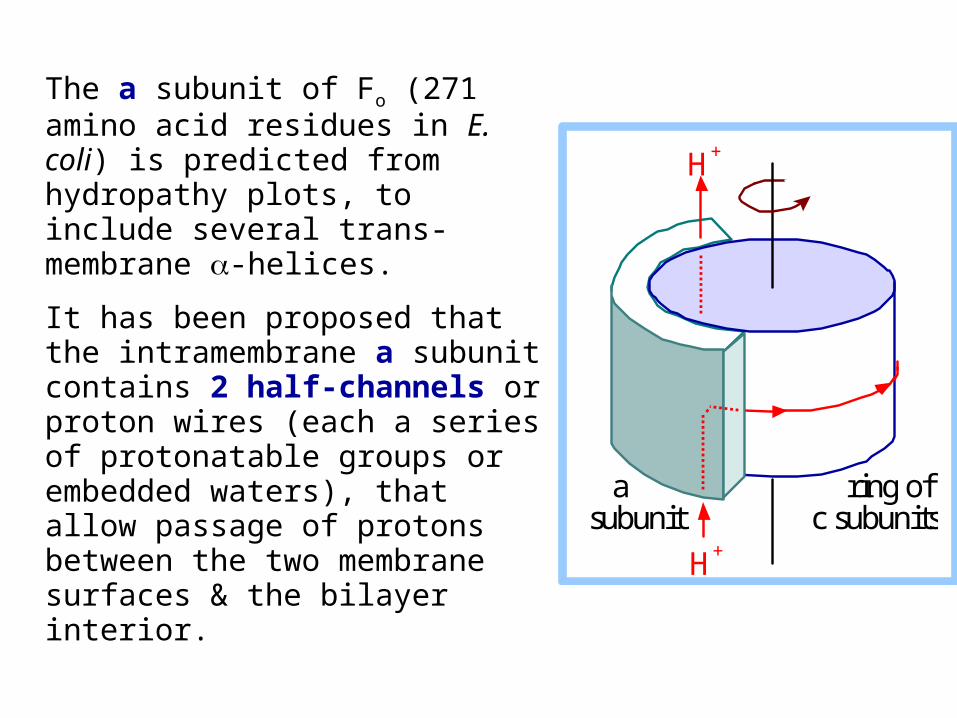
The a subunit of Fo (271 amino acid residues in E. coli) is predicted from hydropathy plots, to include several trans-membrane -helices.
It has been proposed that the intramembrane a subunit contains 2 half-channels or proton wires (each a series of protonatable groups or embedded waters), that allow passage of protons between the two membrane surfaces & the bilayer interior.
a subunit
ring of c subunits
H+
H+
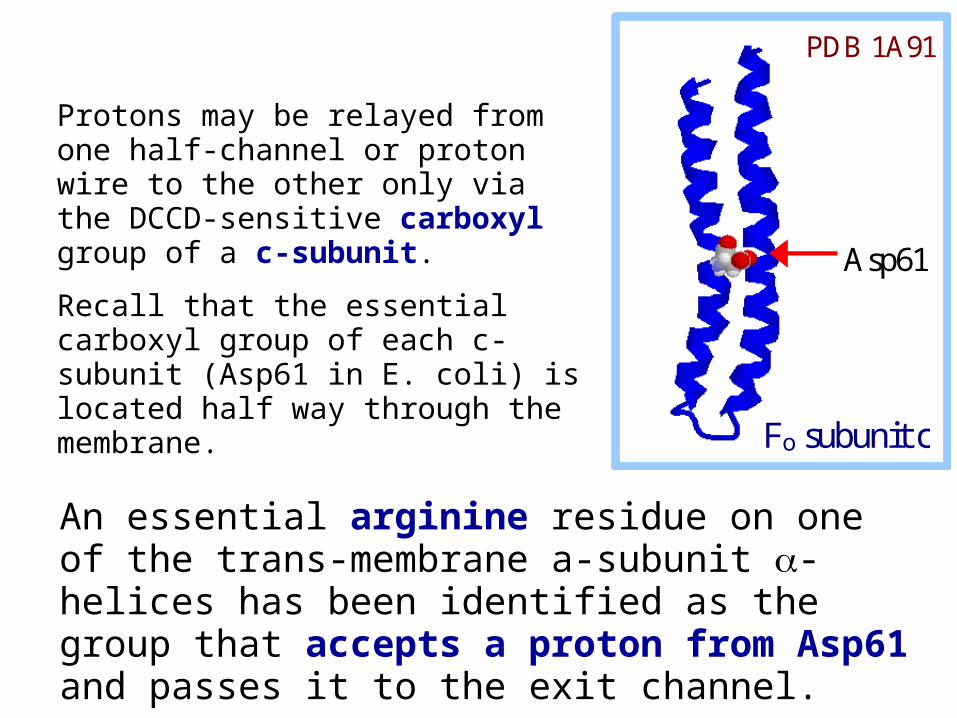
Protons may be relayed from one half-channel or proton wire to the other only via the DCCD-sensitive carboxyl group of a c-subunit.
Recall that the essential carboxyl group of each c-subunit (Asp61 in E. coli) is located half way through the membrane.
Asp61
Fo subunit c
PDB 1A91
An essential arginine residue on one of the trans-membrane a-subunit -helices has been identified as the group that accepts a proton from Asp61 and passes it to the exit channel.
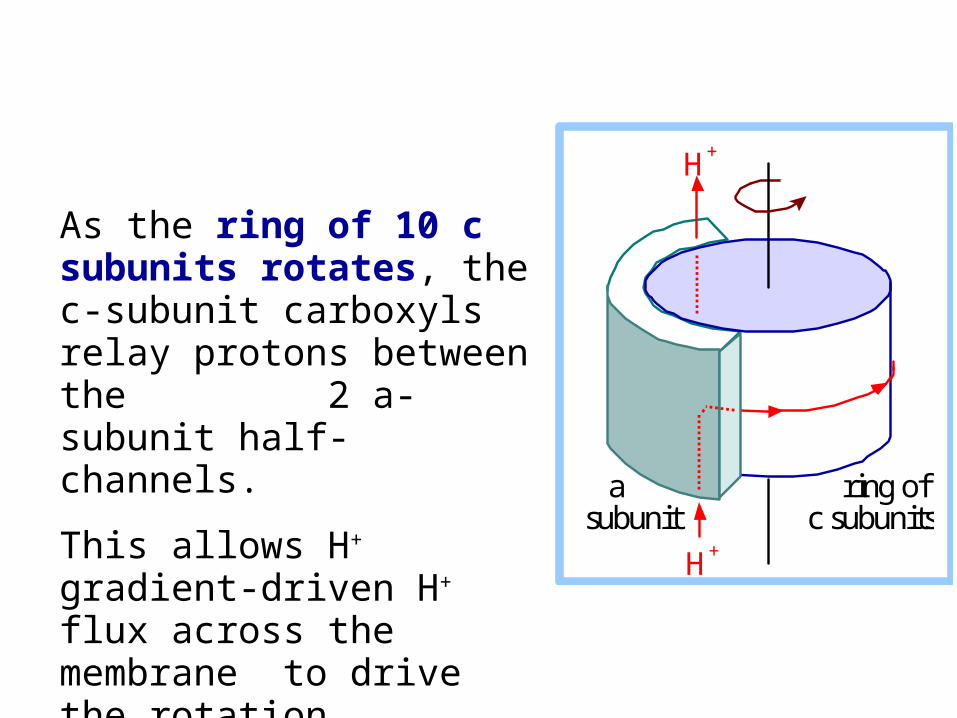
As the ring of 10 c subunits rotates, the c-subunit carboxyls relay protons between the 2 a-subunit half-channels.
This allows H+ gradient-driven H+ flux across the membrane to drive the rotation.
a subunit
ring of c subunits
H+
H+
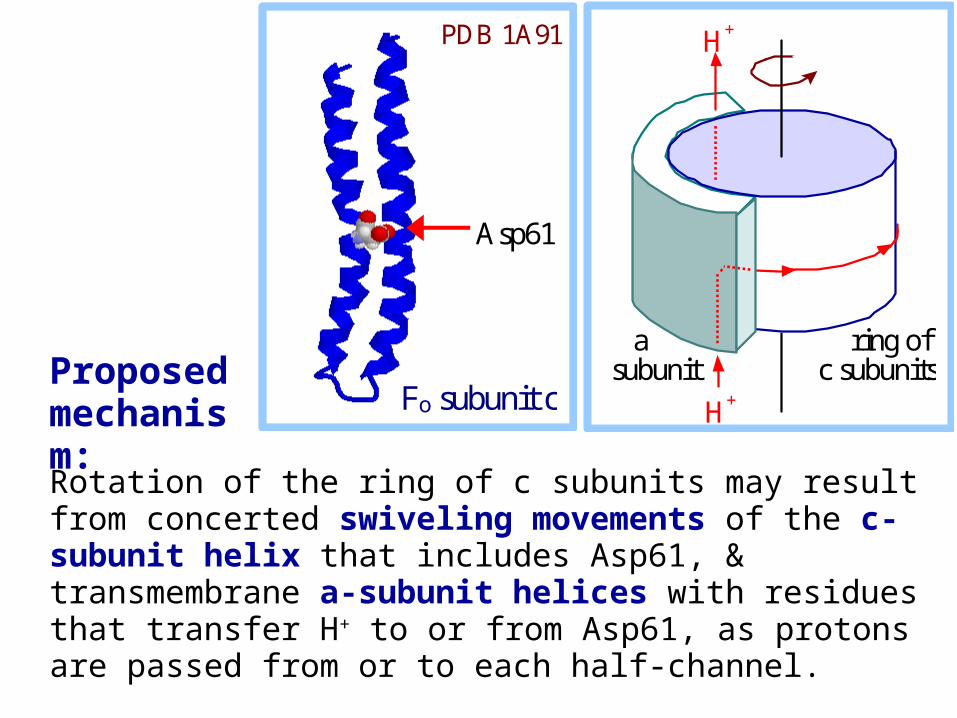
Rotation of the ring of c subunits may result from concerted swiveling movements of the c-subunit helix that includes Asp61, & transmembrane a-subunit helices with residues that transfer H+ to or from Asp61, as protons are passed from or to each half-channel.
a subunit
ring of c subunits
H+
H+
Asp61
Fo subunit c
PDB 1A91
Proposed mechanism:
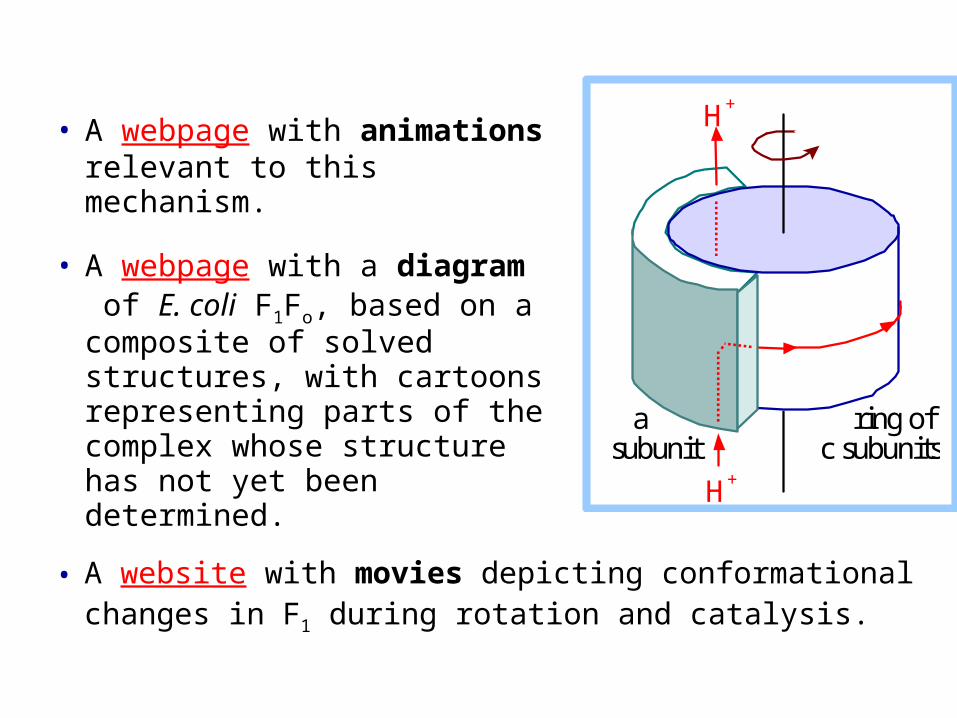
• A webpage with animations relevant to this mechanism.
• A webpage with a diagram of E. coli F1Fo, based on a composite of solved structures, with cartoons representing parts of the complex whose structure has not yet been determined.
a subunit
ring of c subunits
H+
H+
• A website with movies depicting conformational changes in F1 during rotation and catalysis.





























