Embed Size (px)
Citation preview

EXTRACTION AND CHARACTERIZATION OF
COLLAGEN FROM LEATHER JACKET
(Odonus niger)
Thesis submitted in part fulfillment of the requirements for the degree of Master of Fisheries Science in Industrial Fish Processing Technology to the
Tamilnadu Veterinary and Animal Sciences University, Chennai
N. MURALIDHARAN, B.F.Sc. [ID No. MFT 07010 (FPT)]
DEPARTMENT OF FISH PROCESSING TECHNOLOGY FISHERIES COLLEGE AND RESEARCH INSTITUTE
TAMILNADU VETERINARY AND ANIMAL SCIENCES UNIVERSITY THOOTHUKUDI – 628 008
2009

TAMILNADU VETERINARY AND ANIMAL SCIENCES UNIVERSITY
DEPARTMENT OF FISH PROCESSING TECHNOLOGY
FISHERIES COLLEGE AND RESEARCH INSTITUTE
THOOTHUKUDI – 628 008
CERTIFICATE
This is to certify that the thesis entitled, “Extraction and Characterization of
Collagen from Leather jacket (Odonus niger)” submitted in part fulfillment of the
requirements for the award of the degree of Master of Fisheries Science in Industrial Fish
Processing Technology to the Tamilnadu Veterinary and Animal Sciences University,
Chennai is a record of bonafide research work carried out by N. Muralidharan under my
supervision and guidance and that no part of this thesis has been submitted for the award of
any other degree, diploma, fellowship or similar titles.
Place: Thoothukudi
Date :
Dr. R. JEYA SHAKILA
Chairman
RECOMMENDED
Place: Chennai
Date :
EXTERNAL EXAMINER
APPROVED BY:
Chairman : Dr. R. Jeya Shakila
Associate Professor
Members : Dr. D. Sukumar
Associate Professor
Dr. K. Karal Marx
Associate Professor
Place: Thoothukudi
Date :

Dedicated to
my beloved madam
and
my family

ACKNOWLEDGEMENTS
I express my heartiest gratefulness to my respected guide, Dr. R. Jeya Shakila,
Associate Professor, Department of Fish Processing Technology for having taken me as her
student. I thank her very much for extending unceasing help, superior support, well skilled
guidance, creative censure, heed which paved me to carry out the research work successfully
in an enthusiastic way.
I am obliged to Dr. D. Sukumar, Associate Professor, Department of Fish
Processing Technology and member of the Advisory committee for his vast help and valuable
suggestions at every need of hour. I extend my hearty tribute to Dr. K. Karal Marx Associate
Professor, Department of Aquaculture and member of the Advisory committee for his support
and encouragement throughout the research work and I also thank Dr. Baskaran
Manimaran, Associate Professor, Department of Fisheries Environment and former member
of the Advisory committee for his appropriate help.
I will be failing in my duty, if I do not honor Dr. G. Jeyasekaran, Associate
Professor, Department of Fish Processing Technology for his noble-minded guidance,
enormous suggestions, care and affection extended throughout my research work.
Dr V. K. Venkataramani, Dean of the Faculty (Fisheries) is lavishly thanked for his
encouragement throughout the research work. My thanks are due to Dr. P. Velayutham,
Professor and Head, Department of Fish Processing Technology for his support.
I do acknowledge my sincere thanks to faculty members, Dr. G. Indhra Jasmine,
Dr. G. Sugumar, Dr. K. Rathnakumar and Dr. S.A. Shanmugam of the Department of Fish
Processing Technology for their unlimited help throughout the research work.
Special thanks are to The Director, Central Institute of Fisheries Technology, Kochi
for permitting me to collect relevant literatures and I also thank Mr. Kumaresan, Mrs.
Ezhilrani, Mr. Vishwanathan and Mrs. Indirani for having provided me necessary support
with regard to literature collection for my study.

I extend my thanks to The Director, Central Institute of Brackishwater Aquaculture,
Chennai for carrying out the amino acid analysis of the sample. I also thank Mr.
Krishnamoorthy, Ph. D., scholar of Central Leather Research Institute, Chennai for
providing suggestions.
I record my heartfelt thanks to Mr. K. Thirumalai Raj and Mr. Y. Esakkias for
rendering their help, encouragement and profuse support in bringing out the thesis. My
sincere thanks are due to Mr. B. Edwin Raj, Mr. M. Palani Kumar, Mrs. J. Jemila Thanga
Rani, Ms. Indhumathi and Mrs. U. Rajeshwari for their kindly assistance in completing this
research work.
I owe it to my parents and sisters, without whose unflinching love and moral support,
this study would have ever seen the light of the day.
(N. Muralidharan)

ABSTRACT
Title : Extraction and Characterization of Collagen from
Leather jacket (Odonus niger)
Name : N. Muralidharan
Degree : M.F.Sc., in Industrial Fish Processing Technology
Chairman : Dr. R. Jeya Shakila
Department : Fish Processing Technology
College : Fisheries College & Research Institute
Year& University : 2009, Tamilnadu Veterinary and Animal Sciences University
Fish collagen was extracted from the skin, bone and muscle of a trash fish, leather
jacket (Odonus niger) using three different acids viz, acetic, lactic and propionic acids using
a solid/solution rate of 1:10 (w/v). The proximate composition of these raw materials was
determined following standard AOAC procedures. The total collagen content of the raw
materials was calculated based on the hydroxyproline content estimated by
spectrophotometric method described by AOAC. The collagen was extracted after 0.8 M
NaCl and 0.1 M NaOH pretreatments using the respective acids. Acid soluble (ASC) and
pepsin soluble collagens (PSC) were extracted following three different methods (Methods 1,
2 and 3) with the respective acids. For the extraction of PSC, 0.1% pepsin (w/v) was added
along the acids. The collagen yield was computed as percentage of wet mass to the total wet
weight of the raw materials. The extracted collagen was characterized by SDS-PAGE to
identify the types of collagen and molecular weights of the subunits were determined for
comparison. The specific viscosity of the 0.1% collagen solution was determined to derive
their denaturation temperatures (Td values). The amino acid composition was determined by
the amino acid analyzer to establish their correlation with Td values.

The results indicated that the skin contained relatively high protein content compared
to bone and muscle. Very high ash contents were noticed in skin and bone. The total collagen
content in bone was high (44% of the total protein) than other parts. Lactic acid was found to
be the best solvent for the extraction of collagen as it gave the maximum yield of 95%.
Complete solubilization of collagen was noticed with lactic acid. Collagen extraction with
acetic and propionic acids gave a maximum yield of only 55% and 52%, respectively.
Maximum yield of collagen was obtained from bone (95%); while from skin and muscle, it
was 68% and 17%, respectively. Addition of pepsin had enhanced the yield of PSC by many
folds in skin, bone and muscle compared to ASC. Extraction with acid alone was not
recommended for collagen extraction. Addition of pepsin in II extraction also improved the
yield several folds, but not high as noticed with complete extraction using pepsin (Method 3).
Electrophoretic pattern revealed bone and skin collagens were of Type I collagen with a
typical (1)22 chains, while muscle collagen was identified as Type V collagen with the
presence of 1, 2 and 3 subunits. Td values of bone and muscle collagens were high (30-
320C) compared to skin collagen (27-290C). Lactic acid extraction had lowered the Td values
of collagen probably due to the disruption of intermolecular cross-links. The amino acid
analysis revealed presence of 19.3% of imino acids such as proline and hydroxyproline in
bone collagen which could have contributed for their higher thermal stability. It was
therefore inferred that the bone of leather jacket was best suitable for the extraction of
collagen using lactic acid to get maximum yield compared to skin and muscle. Bone of
leather jacket was also a good source of Type I collagen with fairly high thermal stability. To
obtain collagen with thermal stability extraction with acetic acid was preferred, while to get
maximum yield, extraction with lactic acid was found suitable.

CONTENTS
Chapter
No.
Title
Page No.
1. Introduction 1
2. Review of Literature 4
2.1 Fish collagen 4
2.2 Extraction of collagen 6
2.3 Yield of collagen 8
2.4 Types of collagen 9
2.5 Electrophoretic characterization of collagen 10
2.6 Molecular weight of subunits 12
2.7 Denaturation temperatures of collagen 12
2.8 Amino acid composition of collagen 13
2.9 Biotechnological approaches to fish collagen identification
15
2.10 Applications of collagen 15
2.11 Studies in India 16
3. Materials and Methods 17
3.1 Materials 17
3.1.1 Fish 17
3.1.2 Ice 17
3.1.3 Chemicals and solvents 17
3.1.4 Dialysis membranes and clips 17
3.1.5 Equipments 17
3.1.5.1 Deboner/Mincer 17
3.1.5.2 Deep freezer 18
3.1.5.3 Chill cabinet 18
3.1.5.4 Refrigerated Centrifuge 18
3.1.5.5 Filtration assembly 18

3.1.5.6 Lyophilizer 18
3.1.5.7 U/Vis. Spectrophotometer 18
3.1.5.8 Ostwald’s Viscometer 18
3.1.5.9 Electrophoresis apparatus 19
3.1.5.10 HPLC 19
3.1.5.11 Nitrogen analyzer 19
3.1.5.12 Fat analyzer 19
3.1.5.13 Other instruments 19
3.2 Methods 19
3.2.1 Separation of raw materials 19
3.2.2 Biochemical composition of the
raw materials
20
3.2.2.1 Moisture 20
3.2.2.2 Total crude protein 20
3.2.2.3 Total crude fat 20
3.2.2.4 Ash 20
3.2.2.5 Total collagen 20
3.2.2.6 Hydroxyproline content 21
3.2.3 Extraction of collagen 21
3.2.3.1 Pre-treatment 21
3.2.3.2 Collagen extraction 22
3.2.3.3 Dialysis of collagen 22
3.2.3.4 Lyophilization of collagen 23
3.2.4 Characterization of collagen 23
3.2.4.1 SDS-Polyacrylamide Gel Electrophoresis 23
3.2.4.2 Amino acid analysis 23
3.2.4.3 Viscosity 24
3.2.4.4 Denaturation Temperature (Td) 24
3.2.5 Statistical analysis 24
4. Results 25

4.1 Structural composition of leather jacket 25
4.2 Proximate composition of leather jacket 25
4.3 Total collagen content 25
4.4 Yield of collagen 29
4.4.1 Skin 29
4.4.2 Bone 29
4.4.3 Muscle 30
4.5 Electrophoretic characterization of collagen 30
4.6 Amino acid composition of collagen 33
4.7 Denaturation temperatures of collagen 33
5. Discussion 40
5.1 Structural composition of leather jacket 40
5.2 Proximate composition of leather jacket 40
5.3 Collagen yield 41
5.4 Extraction of collagen 43
5.5 Electrophoretic characterization of collagen 46
5.6 Amino acid composition of collagen 48
5.7 Thermal stability 50
6. Summary 54
7. References 58

LIST OF TABLES
Table No.
Title
Page No.
1. Total collagen content in the skin, bone and muscle of leather jacket
28
2. Amino acid composition of pepsin soluble collagen bone 34
LIST OF PLATES
Plate No.
Title
Page Nos.
1. Preparation of raw materials for the extraction of collagen from leather jacket
20 - 21
2. Extraction of collagen from leather jacket
21 - 22
3. Flow chart for the extraction of collagen 22 - 23

LIST OF FIGURES
Figure
No.
Title
Page No.
1. Proportion of different structural parts of the fish, leather jacket
26
2. Proximate composition of fish, leather jacket (wet weight basis)
27
3. Yield (%) of collagen from the skin, bone and muscle of leather jacket
31
4a. SDS-PAGE separation of the -chains of ASC and PSC of skin, bone and muscle collagens of the leather jacket
32
4b. SDS-PAGE separation of the -chains of PSC of skin, bone and muscle collagens of the leather jacket
32
5. Denaturation temperatures of ASC and PSC of the skin of leather jacket
36
6. Denaturation temperatures of ASC and PSC of the bone of leather jacket
37
7. Denaturation temperatures of ASC and PSC of the muscle of leather jacket
38
8. Denaturation temperatures of the collagen extracted from skin, bone and muscle of leather jacket
39

1. INTRODUCTION
Collagen, the most abundant protein in vertebrates, constitutes 25% of total proteins.
The word “collagen” has been derived from the Greek words ‘kolla’ and ‘genos’ meaning
glue and formation, respectively. Collagen is stringy, insoluble and indigestible. Gelatin is a
partially hydrolyzed form of collagen.
Collagen has got a variety of biomedical and pharmaceutical applications. Their
applications include treatment of pain associated with osteoarthritis, hypertension, use in
tissue engineering, implants in human, inhibition of angiogenic diseases, etc. (Rehn et al.,
2001). Collagen is also used as dermal filler, as hemostat, for drug delivery, as skin
substitutes, as expandable intra–arterial stents and cell attachment substrate (Senaratne et al.,
2006). In food industry, they are used as gelatin in dessert foods. In cosmetic industry,
collagen is used as natural moisturizer, wrinkle-resistant mask and facial cosmetics
(Swatschek et al., 2002); and in shampoo and hair care products. In packaging industry, they
are used in microencapsulation and light sensitive coatings. These light sensitive coatings are
termed as ‘edible films’ and are used in the food industry (Senaratne et al., 2006).
Most of the collagen is derived from cow and pig skins. The outbreaks of certain
animal diseases such as bovine spongiform encephalopathy (BSE), foot and mouth diseases
(FMD) have caused restrictions on the use of animal collagen as there is a possibility to be
transmitted to human beings (Helcke, 2000; Trevitt and Singh, 2003). In addition, Muslims
and Jews do not accept any pig related food products while Hindus does not consume cow-
based products (Pranoto et al., 2007). Therefore, fish collagen is considered as the best
alternative for animal collagen because of its high availability, no risk of disease transmission

and no religious barriers. The main differences of fish collagen from that of animal collagen
are its high biological value, high essential amino acid content and low content of
hydroxyproline.
Fish processing discards, by-catch of unutilized as well as underutilized fish species,
are the promising sources for the extraction of collagen. Fish processing discards generally
include skin, bones, scales and fins, which contain collagen. India generates more than 2
million metric tonnes of waste every year from fish processing activities (Sudeepa et al.,
2007). Processing discards from fisheries accounts for as much as 70–85% of the total weight
of catch (Shahidi, 1994). They are generally dumped in-land or hauled into the ocean.
Disposal of these wastes also pose environmental problems for seafood processors. Hence,
there is a need for devising ecofriendly methods for utilizing these wastes and discards. The
main areas of progress are to obtain collagen, proteins, enzymes and protein concentrates
from these wastes and discards.
Thoothukudi Fishing Harbour is one of the major fishing harbours of Tamilnadu.
There are about 260 fishing vessels, which undertake fishing in Gulf of Mannar region.
Everyday 260 tonnes of fishes are landed and marketed to various places within and/or
outside the State. Leather jacket is a trash fish belonging to the family, Balistidae that has got
a huge landing in Thoothukudi region during the months of September and November.
During the season, about 40 tonnes of this species are caught by a single trawler (53 ft) and
sold at a very cheaper price @ Rs. 3/kg . These trash fishes are mainly utilized for poultry
feed in whole form after drying. Besides this, there are very few companies that purchase
these fishes for surimi processing discarding their frames.

There are three types of leather jacket, namely, Melichthys indicus (Indian trigger
fish), Odonus niger (Red-toothed trigger fish) and Pseudobalistes fuscus (Rippled trigger
fish). The “Red-toothed trigger fish” locally known as “Kakka Kilathi” in Tamil are caught
in abundance in Thoothukudi. They have a thick skin, constituting about 25% of the total
body weight. Huge landings of these trash fishes, at times, pose environmental problems in
fishing harbour due to unhygienic handling (without proper icing) and less demand.
Considering the environmental problems and growing knowledge on the utilization of
processing discards/wastes for the production of valuable products such as fish collagen, this
study was undertaken with the following objectives:
i. Standardization of collagen extraction using different acids
ii. Purification and characterization of fish collagen
iii. Determination of the functional properties of fish collagen

2. REVIEW OF LITERATURE
Collagen is abundant in the connective tissues of animals. It has a triple helical
structure with three long polypeptide chains. Each polypeptide is a left handed helix; but the
three helices are wrapped around each other towards right. Each polypeptide is made up of
roughly 1000 amino acid residues with a repeated glycine X-Y sequence (Mathew, 2002).
Two special amino acids are found in collagen. They are derived from the standard amino
acids, proline and lysine by addition of hydroxyl groups. Proline and hydroxyproline form
bends in polypeptide chains and are not compatible with -helix structure (Lehninger, 1982).
2.1. Fish collagen The largest concentration of collagen in fish is found in the skin, bone, fin, swim
bladder, scale apart from the muscle. Air bladder was the source of pure collagen (Mathew,
2002). Collagen content in fish varies with the species, muscle parts, age, season, nutritional
condition, time of catch, etc. and the variation is not only with the species but also within the
species. The proportion of collagen in the connective tissue is 88-98%. Collagen and elastin
contents were reported to be 0.68–1.35% of the total proteins in fish.
A considerable amount of non-collagenous proteins in the insoluble collagen
preparations was suggested to be present in fish (Mathews, 1975). The exact content of
collagen could be determined only if hydroxyproline content in collagen is known, but the
hydroxyproline content was also found to vary with the fish species (Yamaguchi et al.,
1976). Collagen content in fish muscle is generally one tenth or less. The collagen content
was more in the light muscle (3%) than in the dark muscle (8.6%). Collagen content in the
muscles of teleosts is usually within a range from 1-4% of the total protein. Crustacean and
molluscan collagens were reported to contain more acidic amino acids and hydroxylysine
compared to that of fish. Collagen content in squid mantle was comparatively high, which is
10% of the total protein. The abdominal muscle tissues of giant river prawn, fleshy prawn

and spiny lobster contained 2.4–2.6% collagen of the total tissue protein (Kimura and
Tanaka, 1986).
Most of the studies have reported the presence of collagen in the muscle of fish. The
carp white muscle had 2.4% collagen of the total tissue protein (Kubota and Kimura, 1975).
The total collagen contents in the muscles of rainbow trout, Japanese mackerel, carp and eel
were found to be 0.47%, 0.50%, 0.60% and 1.99% of the wet muscle, respectively (Sato et
al., 1986a). The collagen content in fish muscle ranged from 0.34% to 2.19% of wet tissue
and from 1.6% to 12.4% of crude protein, respectively (Sato et al., 1986b). The total collagen
content in the whole body also varied with species, the lowest value was 3.26% of wet body
weight and 16.7% of crude protein for Chub mackerel and the highest values were 6.97% and
43.2%, respectively for Japanese eel (Yoshinaka et al., 1990). A very low total collagen
content of 0.29% of fresh weight was also reported in farmed Atlantic salmon white muscle
(Aidos et al., 1999).
Collagen is abundantly present in skin, scale, bone and fin of fish. The total collagen
in these organs ranged from 76.2% of whole body collagen for Japanese eel to 91.1% for red
seabream (Yoshinaka et al., 1990). Montero and Borderias (1989) reported that collagen
from skin was more than that of muscle. The collagen content in raw backbone of cod was
24% of the dry weight (Gildberg et al., 2002; Skierka et al., 2007) and the remaining protein
contained 52% of non-collagenous proteins, peptides and few amino acids in dry weight.
2.2. Extraction of collagen
Collagen, being an acid soluble structural protein, is extracted using inorganic (HCl)
and organic acids (acetic, citric and lactic acids) from various parts of fish. Citric acid was
first used for the extraction of collagen by Piez and Gross (1960). Acetic acid was used for
the extraction of collagen (Lewis and Piez, 1964). Lactic and propionic acids were also found
suitable for collagen extraction by Gomez-Guillen and Montero (2001). Skierka and

Sadowska (2007) have compared the extraction of collagen from skin of cod using citric,
lactic, acetic (0.5 M) and HCl acids (0.15 M) at a ratio of 1:6.
It is possible to increase the yield of collagen by chemical or enzymatic
pretreatments. Chemical pretreatment removes numerous intra and inter molecular covalent
cross-links mainly involving lysine and hydroxylysine residues, ester bonds and bonds with
saccharides (Skierka and Sadowska, 2007). The preliminary extraction with 0.1 N NaOH was
found to remove the non-collagenous proteins most effectively without any modification of
collagen and also exclude the effect of endogenous proteases on collagen during extraction
(Kimura and Tanaka, 1986). Sato et al. (1987) standardized the concentration of NaOH and
reported that 0.5 and 1 N NaOH satisfactorily removed non-collagenous proteins, modified
the polypeptide chains of carp muscle collagen and increased the solubility of collagen. This
preliminary NaOH extraction was also followed by several workers prior to the collagen
extraction by acids (Mathew et al., 1998; Aidos et al., 1999; Yata et al., 2001; Ogawa et al.,
2004; Senaratne et al., 2006; Skierka et al., 2007).
Another preliminary treatment with NaCl prior to NaOH extraction was suggested by
Montero et al. (1995) to remove the impurities and the same was also followed by few
workers (Montero and Gomez-Guillen, 2000; Muyonga et al., 2004). The enzymatic
pretreatment is the use of proteolytic enzymes non-specific for collagen such as pepsin,
trypsin, etc. (Nishihara, 1962). These enzymes remove the non-helical ends (telopeptides) of
the collagen and remove the intermolecular cross-links (Bailey and Light, 1989; Hickman et
al., 2000). Addition of pepsin to cleave the non-helical region, telopeptide during the
extraction of collagen was followed by several workers (Kimura, 1983; Mathew et al., 1998;
Aidos et al., 1999; Sivakumar et al., 2000; Yata et al., 2001; Ogawa et al., 2004; Senaratne et
al., 2006; Skierka and Sadowska, 2007).

Back bone contains collagen as well as mineral salts, mainly calcium phosphate and
carbonates. Demineralization prior to extraction helps to obtain native collagen from bone
and such collagen is called “ossein” (Morimura et al., 2002; Muyonga et al., 2004). The HCl
acid (0.6-0.8M) was used to dissolve mineral salts from osseous elements. EDTA can also be
used as it forms insoluble salts with many metals and minerals (Ikoma et al., 2003; Nagai et
al., 2004). Skierka et al. (2007) have optimized the concentration of EDTA and HCl and
reported that 1.0 M HCl gave best demineralization effect and 0.5 M EDTA gave 72%
demineralization with no loss of collagen.
Fish scales of Pagrus major and Oreochromis niloticas were demineralized with
EDTA prior to the extraction of collagen and then digested by pepsin (Ikoma et al., 2003).
Fractionation of collagen into different types by salt precipitation method was described by
Sato et al. (1991) and the same was followed by Aidos et al. (1999) to fractionate the
Atlantic salmon muscle collagen. Two step precipitation using 0.6-0.8 M and 2.4-2.7 M NaCl
was followed to separate the skin collagen of brown backed toad fish (Senaratne et al., 2006)
and the muscle collagen of fish (Kimura et al., 1988). The removal of ash by EDTA and fat
by hexane prior to the extraction of collagen with acetic acid from the skin, scale and bone of
deep-sea red fish was suggested by Wang et al. (2008). The collagen acid extracts after
centrifugation were salted out by the addition of sodium chloride (Yata et al., 2001). A
concentration of 2 M NaCl (Sivakumar et al., 2000; Yata et al., 2001) and 0.9 M NaCl
(Muyonga et al., 2004; Ogawa et al., 2004) was used for precipitation of collagen.
2.3. Yield of collagen
The yield of extracted collagen varied with the species, their age, structural parts and
the parameters of extraction (Skierka and Sadowska, 2007). Mathew et al. (1998) have
reported the yields of acid soluble collagen (ASC) and pepsin digestible collagen (PDC) as
0.38% and 0.09% from muscle and 20.14% and 5.6% from skin, respectively. Higher yield of

collagen was obtained from skin, bone and scales than from muscle. Nagai and Suzuki
(2000b) reported the yields of collagen from fish skin, bone and fin as 49.8-51.4%, 40.1-
53.6% and 5.2-36.4%, respectively. Fish scales were decalcified, disaggregated and collagen
was extracted by pepsin digestion from sardine, seabream and seabass and the yields were
found to be 50.9%, 37.5% and 41.0%, respectively (Nagai et al., 2004). Collagen from skin
of brown backed toad fish was about 54.3% on lyophilized dry weight basis (Senaratne et al.,
2006). The maximal yield of collagen extracted using acetic acid from skin, scale and bone
of deep-sea redfish was 47.5%, 6.8% and 10.3%, respectively (Wang et al., 2008).
The yield of PSC was always found to be higher than ASC. The yields of ASC and
PSC of puffer fish skin were reported to be 10.7% and 44.7%, respectively on a dry weight
basis (Nagai et al., 2002). The yield of collagen from skin of bigeye snapper was 5.31% with
acid and 18.74% with its own pepsin (Nalinanon et al., 2007). The yield of PSC from skin of
grass carp was reported to be 46.6 % (Zhang et al., 2007). The yields of ASC from skin of
dusky spine foot and sea chub were 3.4–3.9% and 5.3–5.7% for ray (eagle and red sting ray),
respectively (Bae et al., 2008). The young fish skin yielded more collagen than that of the
adults. Muyonga et al. (2004) extracted ASC using acetic acid and NaCl precipitation from
skin of young and adult Nile Perch and the yields were found to be 63.1% and 58.7%,
respectively.
The yield of collagen was also influenced by the acids used for extraction. Gomez-
Guillen and Montero (2001) extracted collagen from skin of megrim using formic, acetic,
propionic, lactic, malic, tartaric and citric acids and found that acetic and propionic acids
produced gelatins with high visco-elastic properties. Skierka and Sadowska (2007) used
citric, HCl, acetic and lactic acids for the extraction of collagen from the skin of Baltic cod
and obtained a yield of 60%, 90% and 18%, respectively.

2.4. Types of collagen
The structure of collagen from mammalian tissues has been widely studied and atleast
27 types of collagen named Type I to Type XIX were identified (Boot-Handford et al.,
2003). Type I collagen was found on all body parts and was considered as the most abundant
type of collagen. Types of collagens are classified based on the difference of the chains in the
molecule. Type I collagen usually contains and chains in 2:1 ratio and occurs in omnion,
chorion, placenta and cornea; Type II collagen is present in hyaline cartilage containing three
(II) chains; Type III collagen is present in skin of young organisms containing three (III)
chains; Type IV collagen is usually seen in basement membranes containing three (IV)
chains; Type V collagen usually contains , and chains. The most abundant collagen
forms extra cellular fibrils or network like structures, but others fulfill a variety of biological
functions. Collagens that form fibrils are Types I, II, III, V and XI and that form network like
structures are Types IV, VIII and X (Mathew, 2002).
2.5. Electrophoretic characterization of collagen
Collagen from the swim bladder of teleosts was perhaps the first collagen to be
characterized (Boedtker and Doty, 1955). The SDS-PAGE pattern of fish collagen mostly
showed two distinct bands corresponding to 1 and 2 chains (subunits) along with cross-
linked chains. The inter and intra molecular cross-linked components are (dimmers) and
(trimmers). Type I collagen α chains of the carp and mackerel skin were characterized by
Kimura (1983) and reported that there existed a higher degree of similarities between α1 and
α2 chains as well as marked inter species differences in the α2 chains. Kimura et al. (1987)
further reported that Type I skin collagen of teleosts possessed a unique subunit, α3 which
was not detected in vertebrate Type I collagen. They observed that Type I collagen of skin
existed as α1α2α3 heterotrimer in many teleosts and as (α1)2α2 heterotrimer in some teleosts.
Tissue specific existence of two molecular forms of Type I collagen from skin (α1α2α3) and

swim bladder [(α1)2α2] of the fish suggested that α3 chain seems to be the product of a third
genetic locus which arose as a duplication of α1 gene (Kimura and Ohno, 1987). The
characterization of Type I skin collagen from puffer fish (Nagai et al., 2002), Nile perch
(Muyonga et al., 2004), grass carp (Zhang et al., 2007), big eye snapper (Nalinanon et al.,
2007), deep sea red fish (Wang et al., 2008), yellow fin tuna (Woo et al., 2008) revealed that
it had (α1)2α2 chains. Presence of Type I collagen with α1α2α3 heterotrimer chains was
reported from the skin of brown backed toad fish (Senaratne et al., 2006).
Type V collagen was identified for the first time in fish muscle by Sato et al. (1998)
after purification with ion-exchange chromatography in the presence of 5 M urea. By the
confocal microscopy and immuno-histochemistry studies of cod muscle, Types III and VI
collagens were identified in myocommata and endomysium, respectively and Type IV in the
basement membrane (Bruggemann and Lawson, 2005), in addition to Type I collagen. They
also reported the presence of Type V collagen in cod muscle.
The skin collagen of horse mackerel, yellow seabream and tiger puffer fish reported
by Yata et al. (2001) contained two fractions after ammonium sulfate precipitation, phospho
cellulose column chromatography purification. They identified Type I as major collagen and
Type V as minor collagen. Nishimoto et al. (2004) followed the same procedures and
separated Type I and V collagens from amberjack. In tiger puffer fish, Type I and Type V
collagens were found to be widely distributed in several parts (Mizuta et al., 2005).
A wide distribution of three molecular forms such as (α1)2α2, (α1)3 and α1α2α3 in the
Type I fish bone collagen has been identified by Nagai and Suzuki (2000b). Type I collagen
with (α1)2α2 chains was characterized from bone of black drum and sheepshead seabream
(Ogawa et al., 2004) and deep-sea red fish (Wang et al., 2008). Scales of fishes, Pagrus
major and Oreochromis niloticas (Ikoma et al., 2003); sardines, seabream and seabass

(Nagai et al., 2004), black drum and sheepshead seabream (Ogawa et al., 2004) as well as
deep-sea red fish (Wang et al., 2008) were reported to contain Type I collagen with (α1)2 α2
chains.
The types of collagen in squid (Sepia officinalis) were isolated by salt precipitation
method (Rigo et al., 2002) and found that Type I collagen was the major component and
Type V/XI and Type IX are the minor components. Type V/XI collagen was presents in
molluscs and Type IX like collagen was identified to be essential for the formation of
cartilaginous tissue. Hwang et al. (2007) had extracted Type I collagen as major and Type V
collagen as minor from skate skin. There was also an existence of molecular subspecies of
Type V collagen.
2.6. Molecular weight of α subunits
The electrophoretic pattern of collagen contains cross-linked components, dimmers
and trimmers along with chains. The chain had high molecular weight of approximately
200 kDa (Mathew, 2002). The and chains of collagen had molecular weights below 200
kDa. Molecular weight of α subunits of ASC and PSC isolated from the bone and scales of
black drum and sheepshead seabream were found to be 130 kDa for α1 and 110 kDa for α2
(Ogawa et al., 2004). In yet another study, the molecular weight of α1 and α2 chains of ASC
isolated from the skin of bigeye snapper was reported as 120 kDa and 112 kDa, respectively
(Nalinanon et al., 2007). The collagen extracted from the skin of Baltic cod had chains
below 116 kDa in the electrophorogram (Skierka and Sadowska, 2007). The molecular
weights of the two chains (α1 and α2) of Type I collagen were reported to vary with species
and their structural parts. The molecular weight of α2 chain of ray was lower than α2 chain of
bony fish (Bae et al., 2008).

2.7. Denaturation temperatures of collagen Fish collagen was characterized to have a very low denaturation temperature (Td)
values (Lewis and Piez, 1964). The total amino acid content of fish collagen was found
proportional to the Td values (Piez and Gross, 1960). Ikoma et al. (2003) stated that the Td
values were found to be more dependent on hydroxyproline rather than proline.
Td values were also found to vary with the structural parts of fish. Fish muscle Type
I collagen exhibited higher Td values because of higher degree of proline hydroxylation
when compared to skin Type I collagen (Kimura et al., 1988). Td values of bone collagen of
skipjack tuna and yellow seabream were much higher than those of skin collagen (Nagai and
Suzuki, 2000a). The Td values of fish skin, bone and fin ranged from 25-26.5oC, 29.5-30oC
and 28-29.1oC, respectively (Nagai and Suzuki, 2000b). Lower Td values of 150C for cod
skin (Rigby, 1968), 16.80C for Alaskan pollack skin collagen (Kimura and Ohno, 1987) and
19.40C for Chum salmon muscle collagen (Kimura et al., 1988) were reported. The Td values
of the collagen of pacific whiting skin (23.3oC), common mackerel (26.10C), skipjack tuna
(29.70C), ayu bone collagen (29.70C) and brown backed toad fish skin collagen (280C), grass
carp (28.4oC) were reported (Kimura et al., 1988; Nagai and Suzuki, 2000a, Kim and Park,
2004; Senaratne et al., 2006; Zhang et al., 2007). Higher Td values of 360C for collagen of
Nile perch skin (Muyonga et al., 2004), 340C for black drum and sheepshead seabream bone
collagen (Ogawa et al., 2004), 32.50C for carp muscle (Kimura et al., 1988) were also
reported. Wang et al. (2008) observed a Td value of 16.1, 17.7 and 17.50C deep-sea red fish
skin, scale and bone collagen, respectively. Elasmobranchs were found to have 5oC higher Td
values than teleosts (Bae et al., 2008) with ray having a Td value of 33oC.
2.8. Amino acid composition of collagen
Piez and Gross (1960) determined the amino acid composition of scale, skin and
swim bladder collagens of carp, cod and pike. Fish collagens had less proline and

hydroxyproline but more serine and threonine than mammalian collagens. Methionine was
also present in greater amounts. Glycine was the highest fraction accounting for about 38%
in collagen from skin and muscle (Nip et al., 1981; Sikorski et al., 1984; Montero et al.,
1990). Absence of cysteine in collagen of carp ordinary muscle was observed by Sato et al.
(1986a). The amino acid profile of hake and trout skin and muscle collagen was studied by
Montero et al. (1990) in detail. The content of proline and hydroxyproline was 16-21% in the
hake and 15-18% in the trout. The proportion of alanine in trout collagen was approximately
11%. The tyrosine content ranged from 3.9 to 5.0% in the hake and 2.8 to 5.5% in the trout;
the amount of methionine ranged from 14 to 17%. Mathew et al. (1998) studied the amino
acid composition of milkfish skin and muscle collagen and reported that glycine, alanine,
glutamic acid and arginine constituted about 70% of the total amino acids. They also stated
that the total essential amino acid contents of collagens ranged between 170-178 amino acid
residues per 1000 residues. Ikoma et al. (2003) found that glycine (33.6%) was the abundant
amino acid in Type I PSC from fish scales along with methionine, which was higher in fish
scales. Muyonga et al. (2004) reported that the ASC extracted from skins of Nile Perch had
20% imino acids. The amino acid composition of extracted collagen from fish was also
studied by several workers (Muyonga et al., 2004; Ogawa et al., 2004; Nishimoto et al.,
2005).
Amino acid analysis of five preparations (112, 1, 11, 12 and 2) of the dogfish
shark skin collagen examined by Lewis and Piez (1964) revealed that 12 had a composition
equivalent to a 1:1 mixture of 1 and 2; while 11 had a composition identical with 1. The
112 had a composition of unfractional collagen with the expected chain composition of two
1 and one 2 chains; 2 always had less proline and hydroxyproline than 1. The histidine
content of 2 was much higher than that of 1. Higher content of hydrophobic side chains of

amino acids such as valine, isoleucine and leucine was observed in 2 than in 1; 2 was
found to have very low amino acids, a total of 129 residues per 1000, compared with 160
residues per 1000 in the whole molecule. But, it was about 220 residues per 1000 in
mammalian collagens (Piez et al., 1963).
2.9. Biotechnological approaches to fish collagen identification
Biotechnological approaches are made in fish collagen in recent years. Saito et al.
(2001) first reported the complete primary structure of fish Type I procollagen. The α3(1)
subunit that was present only in bony fish had small number of Gly-Pro-Pro and the large
number of Gly-Gly in pro α3(1), which is partially assumed to loosen the triple helical
structures of skin collagen, leading to the lower stability of skin collagen. Three genes α1
chain [cola 1(I)], α2 chain [cola 2(I)], α3 chain [cola 3(I)] cDNA’s and their promoters from
flounder were cloned (Suzuki et al., 2006). A full length cDNA of Type I procollagen α1
(1463a.a) was determined by RACE-PCR and found that Gly-Pro-Pro and Gly-Gly in α1 (1)
chain were related to thermal stability of helix (Hwang et al., 2006). Full length cDNA of the
Type V/XI procollagen was later determined and found that this pro α1 (V/XI) chain was
close with pro α1 (V) of vertebrate but not with pro α1 (XI) (Hwang et al., 2008).
2.10. Applications of collagen
The collagen was used as a natural substratum for cell culture (Yoshizato et al.,
1981). Bracho and Haard (1995a) radiolabelled the skin collagen of lingcod to high specific
radioactivity and used as a substrate for the assay of rockfish skeletal muscle for
metalloproteinase. The same radio labeled collagen was also be used to assay collagenolytic
activity from fish (Bracho and Haard, 1995b). Collagen extracted from bone and skin after
hydrolysis with a commercial enzyme had a high potential for decreasing blood pressure
(IC50, 0.16 and 0.41 mg/ml) (Morimura et al., 2002). Fish collagen obtained from fish scales
was made into sheets and that had a tensile strength sufficient enough to be used as wound

dressing material (Sankar et al., 2008). Collagen films formed from the swim bladder of
three fishes revealed no fibrillar organization but had spongy structure (Fernandez et al.,
2008). Guts traditionally used to provide shape and preservation of sausages during
fermentation, drying, or smokings are mainly substituted by collagenic and/or cellulosic
edible films. Collagen films have now been used for sausage casings.
2.11. Studies in India
In India, studies on fish collagen were initiated by the scientists in 1954,
Ramachandran and Kartha, University of Madras who termed the triple helix structure of
collagen as “Madras helix” (Mathew, 2002). A group of scientists from Central Leather
Research Institute (CLRI), Chennai has later examined the characteristics of fish collagen.
Skin and muscle collagens were extracted from the freshwater catfish grown in biologically
treated tannery effluent water (Sivakumar et al., 2000). The Biochemistry and Nutrition
Division of Central Institute of Fisheries Technology (CIFT), Kochi has developed a
biological dressing material from collagen for use in the treatment of burns (Mathew, 2002).
They had extracted collagen from the air bladders of fish and made a skin substitute using
collagen-chitosan as polymers crosslink. A fish scale collagen sheet (FCS) was developed
from the fish wastes by Sankar et al. (2008). Collagen sponge, an antibacterial collagen-
based wound dressing, and reconstituted collagen plast/ hemostat were also developed by the
scientists of CLRI, Chennai.

3. MATERIALS AND METHODS
3.1. MATERIALS
3.1.1. Fish
Leather jacket (Odonus niger) belonging to the family Balistidae caught by the trawl
net was procured fresh from Fishing Harbour, Thoothukudi situated in front of our Shore
Laboratory campus and brought to the laboratory in chilled condition in the insulated
containers.
3.1.2. Ice
Flake ice, prepared by the Flake ice maker (ZBE 150 Nr 940062, Germany), was used
for chilling the fish during their transportation from landing centre to the laboratory as well
as during further processing.
3.1.3. Chemicals and solvents
Analytical reagent (or) guaranteed reagent grade chemicals and solvents were used
for the extraction and characterization of fish collagen.
3.1.4. Dialysis membranes and clips
Dialysis membranes (DM-110, Himedia, Mumbai) were used for the dialysis of the
collagen samples. Dialysis clips (Himedia, Mumbai) were used for the closure of dialysis
membrane bags.
3.1.5. EQUIPMENTS
3.1.5.1. Deboner/Mincer
Deboner/Mincer machine (Baader/601, Germany) was used for the separation of
minced meat from the wastes viz. skin, scales, fins and bones of the fish.
3.1.5.2. Deep freezer
Deep freezer (SANYO freezer, Japan) was used for the storage of raw samples viz.
muscle, skin and bones prior to their extraction.
3.1.5.3. Chill cabinet
Chill cabinet (Servo, Salem) set at 4oC was used during the extraction of the collagen.
3.1.5.4. Refrigerated Centrifuge

Refrigerated Centrifuge (Hettich Zentrifugen, Universal 32 R, Germany) was used for
the centrifugation of the samples at 4oC during the extraction of collagen.
3.1.5.5. Filtration assembly
Whatman No. 4 filter papers (Catalogue No. 1004 090, Whatman International Ltd,
England) placed over the Buchner funnel and attached to the filter flask fitted with a vacuum
suction pump (Superfit Continental Pvt Ltd, Mumbai) were used for the filtration of samples
during each step of the extraction process.
3.1.5.6. Lyophilizer
Lyophilizer (Christ Alpha 1-4 Lyophilizer, Germany) was used for the lyophilization
of the fish collagen at 1.03 mbar of pressure and -50 to -54oC ice condenser temperature.
3.1.5.7. UV/Vis. Spectrophotometer
UV/Vis Spectrophotometer (Jasco, V-530, Japan) was used for the estimation of
hydroxyproline content in the extracted collagen.
3.1.5.8. Ostwald’s Viscometer
Ostwald’s Viscometer was used for the determination of the relative and specific
viscosities of the collagen.
3.1.5.9. Electrophoresis apparatus
Mini Gel Electrophoresis apparatus (GENEI, Bangalore) was used for the separation
of different molecular fractions of the fish collagen to determine their molecular weights.
3.1.5.10. HPLC
The High Performance Liquid Chromatography (Shimadzu model LC-10A, Japan)
was used for the separation and quantification of the amino acids present in fish collagen
following post column derivatization.
3.1.5.11. Nitrogen analyzer
KEL PLUS-KEL FLOW and KEL-PLUS ELITE EX (Pelican Equipments, Chennai)
were used for the digestion of samples and distillation of nitrogen, respectively for the
determination of protein content.
3.1.5.12. Fat analyzer

SOCS PLUS-SCS 4 (Pelican Equipments, Chennai) was used for the determination of
fat content in the raw materials.
3.1.5.13. Other instruments
Hot air oven and water bath (Dalal, Chennai), autoclave (Secor India Type RT 110P6,
New Delhi), pH meter (Digisun Model 707, Hyderabad), electronic balance (Sartorius
Mechatronics Model CP 2250, Germany), cyclomixer (Remi Equipments, Mumbai) and
muffle furnace (Servo, Salem) were also used.
3.2. METHODS
3.2.1. Separation of raw materials
Leather jackets were washed with potable water to clean the dust, dirt, sand and other
extraneous matter. They were fed into a mechanical deboner/mincer. The minced muscle
meat was collected on one side; and the wastes viz. skin, fins, scales and bones, on the other
side (Plate 1). The yields of muscle (minced meat) and wastes were calculated. From the
wastes, skin and bones were segregated manually. The skin, bones and muscle were used as
raw materials for the extraction of collagen. The raw materials were held frozen at -20oC in a
deep freezer until used for the extraction.
3.2.2. Biochemical composition of the raw materials
3.2.2.1. Moisture
Moisture content was determined in the hot air oven method (AOAC, 1995).
3.2.2.2. Total crude protein
Kelplus digestion system was used for the digestion of samples and Kelplus Elite Ex
distillation system was used for the determination of nitrogen content. Crude protein was
calculated by multiplying nitrogen content with a factor 6.25 (AOAC, 1995).
3.2.2.3. Total crude fat
The crude fat was determined by Soxhlet method (AOAC, 1995) using petroleum
ether (60-80oC) as solvent in a SOCS PLUS- SCS 4 system.

3.2.2.4. Ash
The ash content was determined by the method of AOAC (1995) in a Muffle furnace
set at 500-550oC for 15 h.
3.2.2.5. Total collagen
Total collagen content was determined based on the hydroxyproline content in the
raw materials on wet weight basis. Hydroxyproline content was estimated following the
colorimetric method of AOAC (1995).
3.2.2.6. Hydroxyproline content
Hydroxyproline content was estimated following the colorimetric method of AOAC
(1995) with slight modification. For acid hydrolysis, 0.4g of the extracted collagen was
digested with 30ml 7N Sulphuric acid by placing in a hot air oven at 105 10C for 16 h. The
digested sample was diluted to 100ml with distilled water. The hydroxyproline present in the
sample was then oxidized with chloramine-T, followed by addition of p-dimethyl amino
benzaldehyde to get a coloured complex, which was measured at 558nm in a UV-Vis
Spectrophotometer. Blank was set simultaneously using 2.0ml of distilled water.
3.2.3. Extraction of Collagen
3.2.3.1. Pre-treatment
The whole extraction of collagen from the fish skin (S), bones (B) and muscle (M)
was carried out at 4oC in a chill cabinet with occasional stirring. The sodium chloride (NaCl)
treatment of samples was carried out following the procedure of Montero et al. (1995) with
slight modification. Samples were first treated with 0.8 M NaCl at a ratio of 1:6 (w/v) for 10
min. to remove the impurities and then washed with abundant distilled water. This process
was repeated for 3 times. The sodium hydroxide (NaOH) treatment of samples prior to the
extraction of collagen was then carried out following the procedure of Sato et al. (1986a)
with slight modification. Samples were treated with 0.1 N NaOH at a ratio of 1:10 (w/v) for 3
days to remove the non-collagenous proteins and to prevent the effect of endogenous

proteases on collagen. During the alkali treatment, the alkali solution was changed everyday.
They were then washed with distilled water till the washed water became neutral or slightly
basic pH. The samples were finally homogenized (Plate 2).
3.2.3.2. Collagen extraction
The extraction was carried out using three different acids viz. acetic (A), lactic (L)
and propionic (P) acids in order to examine their individual efficiencies. Three different
methods were followed for the extraction of collagen using the respective acids. In the first
method (M1), acid soluble collagen (ASC) was twice extracted (I and II) using 10 volumes of
0.5 M of the respective acid for 3 days. In the second method (M2), acid soluble collagen
(ASC) was first (a) extracted and then, the pepsin soluble collagen (PSC) was extracted (b).
For the extraction of pepsin soluble collagen, pepsin at 0.1% (w/v) concentration was added
to 0.5 M of the respective acid inorder to cleave the non-helical region, telopeptide. In the
third method (M3), pepsin soluble collagen was twice extracted (I and II) from the raw
materials. After each extraction, the solution was centrifuged at 9000 g for 20 min. at 40C.
The supernatant was salted out using 2 M NaCl for 24 h at 4oC. The precipitated collagen
was centrifuged at 9000 g for 20 min. at 4oC. The residue was re-suspended in cold distilled
water and centrifuged again at 9000 g for 20 min. at 4oC. The residue was then dialysed and
lyophilized.
Methods I - Extraction II - Extraction
1 ASC ASC
2 ASC PSC
3 PSC PSC
3.2.3.3. Dialysis of collagen
The residue was placed in the dialysis membrane-110 bags (Hi-Media), closed using
the dialysis clips and dialyzed against the 0.02M phosphate buffer (pH 7.2) for 24 h at 40C.
The dialyzed sample was then centrifuged at 9000 g for 20 min. at 4oC to obtain pure

collagen and held frozen at -20oC in a deep freezer for further analysis. The yield of collagen
was calculated as dialysed wet mass to 100g of wet tissue (Plate 3).
3.2.3.4. Lyophilization of collagen
A portion of the dialyzed collagen was also lyophilized at -400C in the Christ Alpha
1-4 lyophilizer to obtain freeze-dried collagen.
3.2.4. Characterization of collagen
3.2.4.1. SDS-Polyacrylamide Gel Electrophoresis
Sodium dodecyl sulphate - Polyacrylamide Gel Electrophoresis (SDS-PAGE) was
performed according to the method of Laemmli (1970) using 10% separation gel and 1%
stacking gel. Collagen (10mg) was dissolved in 1.0ml of the sample buffer (Tris–HCl, pH 6.8
containing 2-mercaptoethanol, sucrose, bromophenol blue, 5% SDS) and heated at 50oC for
10 min. Then, 20 l was loaded in each well along with high molecular weight protein
markers (Fermentas Lifesciences, Germany). Electrophoresis was carried out at 50mA
initially and then at 100mA. Protein bands were stained with Coomassie Brilliant Blue R250
and destained using a solution containing water, methanol and acetic acid (5:4:1, v/v/v). The
molecular weights of the collagen -chains were determined by comparison with standard
protein markers.
3.2.4.2. Amino acid analysis
Collagen samples were first hydrolyzed in vacuo using 6 N HCl at 1100C for 24 h.
The amino acid analyses were performed by a Shimadzu model LC-10A HPLC following
post column derivatization (Ishida et al., 1981). Amino acids were identified and quantified
in comparison with authentic amino acid standards (SIGMA-Aldrich Chemicals Co., USA).
3.2.4.3. Viscosity
The viscosity of collagen was determined following the procedure of Sivakumar et al.
(2000) using the Ostwald’s viscometer. The concentration of dry collagen used for viscosity
measurement was 0.1% (w/v) as suggested by Kimura et al. (1988). The viscometer was
filled with collagen solution dissolved in 0.1 M of the respective acid and immersed in a

water bath set at 15oC and allowed to stand for 10 min. for equilibration with the water bath
temperature. Simultaneously, the control acid solution (0.1 M) without collagen was also
immersed in the water bath. The flow times of collagen and control were determined using a
stopwatch. Viscosities were also measured at temperature intervals of about 5oC starting
from 15oC upto 40oC. Flow times were taken in triplicates at each temperature and the
average flow rates were determined. The flow rates were used as an index for the calculation
of reduced viscosities. Relative viscosity (rel) = flow time of sample/flow time of control
(0·1 M acid). Specific viscosities were computed for each temperature using the formula,
specific viscosity (sp) = rel -1
3.2.4.4. Denaturation Temperature (Td)
The denaturation temperatures of fish collagen were taken as the mid-point of the
linear portion of the sigmoidal curve obtained by plotting sp/C at toC/sp/C at 15oC against
temperatures, where C denotes concentration of collagen solution in mg/ml (Sivakumar et
al., 2000).
3.2.5. Statistical analysis
The results were statistically interpreted using SPSS 13.0 to find out the least
significant differences (LSD) in the collagen yield as well as viscosity obtained from skin,
bone and muscle of leather jacket using acetic, lactic and propionic acids following three
different methods.

4. RESULTS
The proportion of skin, bone and muscle in the fish, leather jacket was determined
along with their proximate composition. The collagen from the skin, bone and muscle of
leather jacket was extracted using three different acids and were characterized by
electrophoretic, aminoacids and denaturation studies and the results are summarized below:
4.1. Structural composition of leather jacket
The leather jackets were deskinned and deboned to separate the muscle prior to the
extraction of collagen and the yields of different structural parts are given in Fig. 1. The
muscle constituted 47% and remaining were their frames (53%). The frames included skin
(25%), bone (13%), scales (9%) and fin (7%).
4.2. Proximate composition of leather jacket
The proximate composition viz. moisture, protein, fat and ash contents of the skin,
bone and muscle are presented in Fig. 2. The skin had the lowest moisture content (52%) and
muscle had the highest content (86%). The protein content was high in skin (22%) and low in
muscle (7%). The fat contents were in general low ( 2%). Skin and bone had higher ash
contents (25% and 19%, respectively) than that of muscle.
4.3. Total collagen content
The total collagen contents in the skin, bone and muscle were derived from the
hydroxyproline content and the results are given in the Table 1. The bone contained higher
collagen (4.817%) than the skin. The total collagen content in terms of total protein was also
computed from their respective protein contents. Although skin contained higher protein; the
occurrence of collagen was relatively very low (5% of the total protein). Bone had the
highest collagen (44%) and muscle the least (0.6%).
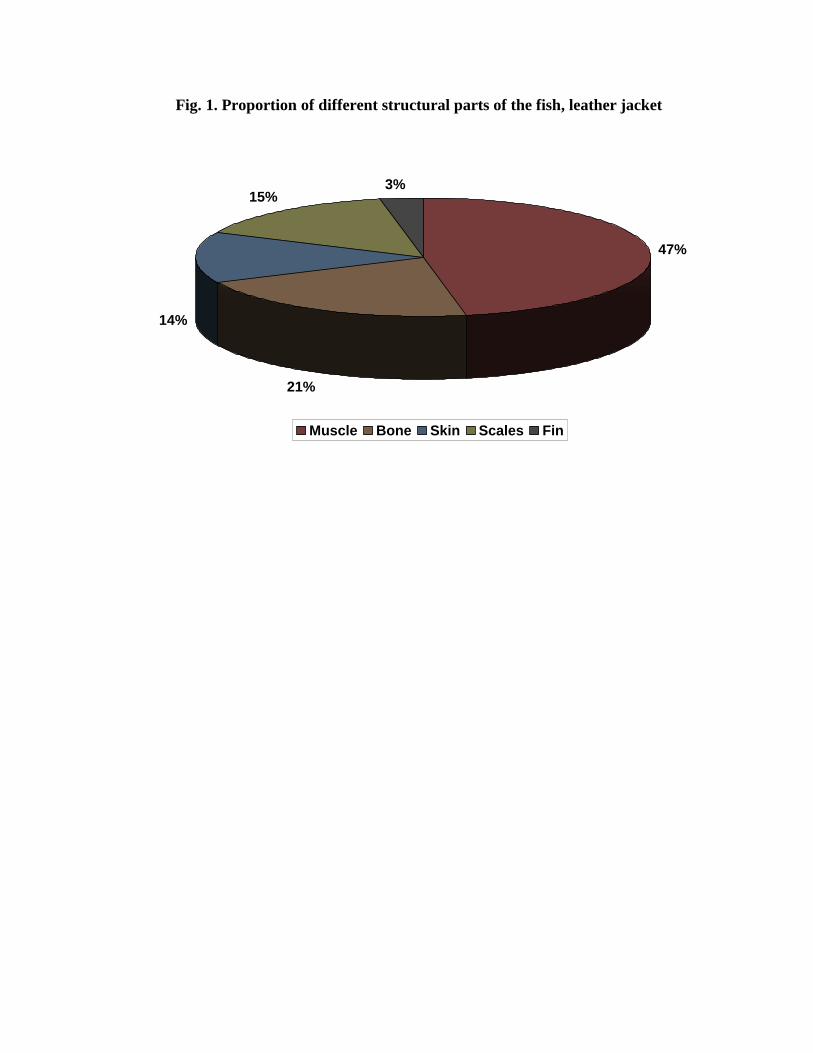
Fig. 1. Proportion of different structural parts of the fish, leather jacket
47%
21%
14%
15%3%
Muscle Bone Skin Scales Fin

Fig. 2. Proximate composition of fish, leather jacket (wet weight basis)
52%
22%
1%
25%
Skin
69%
11%
1%
19%
Bone
86%
7%2% 5%
Moisture Protein Fat Ash
Muscle

Table 1. Total collagen content in the skin, bone, and muscle of leather jacket
Parts Total collagen
(% of wet weight) Protein content
(%) Total collagen
(% of total protein)
Skin 1.096±0.141 21.65±0.65 5.065±0.108
Bone 4.817±0.120 11.06±0.21 43.558±0.821
Muscle 0.042±0.002 6.83±0.06 0.611±0.007

4.4. Yield of collagen
4.4.1. Skin
The yield of collagen obtained from the skin following extraction with acetic, lactic
and propionic acids was calculated and presented in Fig. 3. Each bar gives the total yield
after I and II extractions. The I extraction, in general, resulted in very low yield than the II
extraction. Acid soluble collagen (ASC) extracted following the Method 1 gave lower yields
of 2.69% and 3.78% with acetic and propionic acids, respectively, however with lactic acid,
the yield was high (20.67%). In Method 2, ASC and pepsin soluble collagens (PSC) were
extracted with high yield (32.16%) using lactic acid. PSC alone was extracted by the Method
3. The addition of pepsin with the respective acids had resulted in maximal yield. Both the
extractions further enhanced the yield to a maximum of 68.21% with lactic acid and 44.09%
with acetic acid. The collagen yields obtained by the three methods were statistically distinct
(P<0.05). Propionic acid, in general, gave lower yields compared with other acids.
Statistically no significant differences (P>0.05) existed among the collagen yields obtained
by acetic and propionic acid extractions. However lactic acid yielded higher collagen and
significantly differed from other acids (P<0.05).
4.4.2. Bone The yield of collagen obtained from the bone of leather jacket is given in Fig. 3. The
collagen yield from bone was comparatively higher than that from skin (P<0.05). As noticed
in the skin, the I extraction with acid gave lower yield of collagen and addition of pepsin
yielded more collagen. ASC extracted with acetic and propionic acid gave more or less same
yield of 8.48% and 7.36%, respectively (P>0.05), but that extracted with lactic acid gave
high yield (40.96%). By following the I extraction with acid and II extraction with pepsin
(Method 2), the collagen yield improved to 37% and 24% with acetic and propionic acids,
respectively but not with lactic acid (P>0.05). The PSC extracted by Method 3 gave the
highest yield of 54.76% and 5.84% with acetic and propionic acids, respectively. The

maximum yield of 94.56% was achieved with lactic acid. Statistically no significant
differences (P>0.05) existed among the collagen yields obtained from bone by acetic and
propionic acid extractions, as noticed with skin.
4.4.3. Muscle
The yield of collagen obtained from the muscle following extraction with different
acids is given in Fig. 3. The collagen yields obtained from skin, bone and muscle
significantly differ (P<0.05) following acetic and lactic acid extractions. With respect to
propionic acid extraction, collagen yield obtained from skin and muscle were similar
(P>0.05). The yield of ASC extracted with lactic acid following Method 1 was high (5.34%).
Extraction of PSC by II extraction of Method 2 improved the yield to 9.24% and 14.97%
with propionic and lactic acids, respectively. Addition of pepsin in Method 3 further
increased the collagen yield (P<0.05) and the maximum yield was 16.78% with lactic acid.
4.5. Electrophoretic characterization of collagen
The electrophoretic pattern of ASC and PSC of skin, bone and muscle of leather
jacket is given in Fig. 4a. The SDS – PAGE pattern of the skin and bone collagens showed a
typical pattern with two distinct α bands corresponding to α1 and α2 components along with
their cross-linked chains. The inter and intra muscular cross-linked components are β
(dimers) and γ (trimers). The ASC contained higher proportion of cross-linked components
than PSC counterpart. The γ chain had high molecular weight of approximately 200 kDa.

Fig. 3. Yield (%) of collagen from the skin, bone and muscle of leather jacket
0
10
20
30
40
50
60
70
80
90
100
Yie
ld (%
)
1 2 3 1 2 3 1 2 3
Methods
Acetic acid Lactic acid Propionic acid
Skin
0
10
20
30
40
50
60
70
80
90
100
Yie
ld (
%)
1 2 3 1 2 3 1 2 3
Methods
Bone
0
10
20
30
40
50
60
70
80
90
100
Yie
ld (
%)
1 2 3 1 2 3 1 2 3
Methods
Extraction I Extraction II
Muscle
Fig. 4a. SDS-PAGE separation of the -chains of ASC and PSC of skin, bone and
muscle collagens of the leather jacket

Fig. 4b. SDS-PAGE separation of the -chains of PSC of skin, bone and muscle
collagens of the leather jacket
The proportion of α monomers and β dimmers was high compared to γ trimmers. The β and γ
chains of collagen had molecular weights below 200 kDa.
PSC-M Marker PSC-B
PSC-S
ASC-S ASC-B ASC-M Marker PSC-M PSC-B PSC-S
(1)2
2
3
3
1 2
2
3
1
3

The presence of α1 and α2 chains in skin and bone collagens were a typical pattern of
Type I collagen. The α2 unit was the minor component of the two species and it seems that
the collagen exists as trimers consisting of two α1 and one α2 chains. The molecular weights
of skin and bone ASC subunits were about 120 KDa for α1 and 115 KDa for α2. There were
no significant differences among the molecular weights of these two subunits of skin and
bone collagens.
The SDS-PAGE pattern of muscle collagen was distinct with three α chains viz. α1, α2
and α3, corresponding to that of Type V collagen and their positions indicated in the Fig. 4b.
The molecular weights of three α subunits of muscle collagen were different from that of skin
and bone collagens. Their molecular weights were 125, 120 and 105 KDa for α1, α3 and α2,
respectively.
4.6. Amino acid composition of collagen
The amino acid composition of bone collagen was determined and the results are given
in Table 2. The collagen of leather jacket had high alanine, glycine and proline. Fairly high
levels of arginine, glutamic and aspartic acids were also present. The content of proline was
higher than hydroxyproline. The content of hydroxylysine was lower than the hydroxyproline
content. Tryptophan and cysteine aminoacids were absent.
4.7. Denaturation temperatures (Td value) of collagen
The specific viscosities of ASC and PSC extracted from skin, bone and muscle of leather
jacket were computed to determine their denaturation temperatures (Td values).

Table 2. Amino acid composition of PSC of bone
Amino Acids Residues / 1000 total residues
Hydroxyproline 81.48
Aspartic acid 69.93
Threonine 38.00
Serine 37.48
Glutamic acid 51.85
Proline 111.72
Glycine 196.2
Alanine 198.71
Valine 42.70
Methionine 3.18
Isoleucine 11.36
Leucine 20.12
Tyrosine 42.98
Phenylalanine 5.18
Hydroxylysine 7.84
Lysine 7.66
Histidine 13.00
Arginine 60.61

The denaturation temperature (Td) curves constructed with specific viscosities for
ASC and PSC of the skin of leather jacket are shown in Fig. 5. The specific viscosities
started decreasing between 20-25oC. The ASC extracted using acetic, lactic and propionic
acids had Td values ranging from 27.2-27.8oC; but the PSC had slightly higher Td values,
however not statistically significant (P>0.05). The PSC extracted with acetic and propionic
acids had similar Td values around 30oC (P>0.05).
The denaturation curves of ASC and PSC extracted from bones of leather jacket using
different acids are shown in Fig. 6. The ASC and PSC extracted using acetic and propionic
acids had higher Td values (30oC) thus differing significantly (P<0.05) from lactic acid (27-
29oC). The PSC, in general, had higher Td values than ASC. The Td value of bone collagen
was significantly higher (2-4oC ) than that of skin collagen (P<0.05).
The denaturation temperature curves constructed with specific viscosities of muscle
collagen are presented in Fig. 7. As observed with skin and bone collagens, ASC had lower
Td values than PSC. The Td values of muscle collagen extracted with lactic acid was
significantly lower (P<0.05) than that extracted with acetic and propionic acids. The Td
values of PSC of muscle were higher i.e. 32.1oC with acetic acid and 31.1oC with propionic
acid (P>0.05). The lactic acid, in general, yielded collagen with lower Td values (<29oC).
Fig. 8. gives the comparison of Td values of ASC and PSC extracted from different parts of
leather jacket. The Td values of collagen extracted with lactic acid was significantly lower
(P<0.05) than that extracted with acetic and propionic acids. Bone and muscle collagens of
leather jacket had higher Td values than skin collagen (P<0.05).

Fig. 5. Denaturation temperatures of ASC and PSC of the skin of leather jacket
ASC – A – Skin PSC – A – Skin
0
0.2
0.4
0.6
0.8
1
1.2
10 15 20 25 30 35 40 45
Temperature (oC)
Sp
ec
ific
vis
co
sit
y
27.2oC
0
0.2
0.4
0.6
0.8
1
1.2
10 15 20 25 30 35 40 45
Temperature (oC)
Sp
ec
ific
vis
co
sit
y
28.0oC
ASC – L – Skin PSC – L – Skin
0
0.2
0.4
0.6
0.8
1
1.2
10 15 20 25 30 35 40 45Temperature (
oC)
Sp
ec
ific
vis
co
sit
y
27.8oC
0
0.2
0.4
0.6
0.8
1
1.2
10 15 20 25 30 35 40 45Temperature (
oC)
Sp
ec
ific
vis
co
sit
y
27.8oC
ASC – P – Skin PSC – P – Skin
0
0.2
0.4
0.6
0.8
1
1.2
10 15 20 25 30 35 40 45Temperature (
oC)
Sp
ec
ific
vis
co
sit
y
27.4oC
0
0.2
0.4
0.6
0.8
1
1.2
10 15 20 25 30 35 40 45
Temperature (oC)
Sp
ec
ific
vis
co
sit
y
30.4oC

Fig. 6. Denaturation temperatures of ASC and PSC of the bone of leather jacket
ASC – A – Bone PSC – A – Bone
0
0.2
0.4
0.6
0.8
1
1.2
10 15 20 25 30 35 40 45
Temperature (oC)
Sp
ecif
ic v
isco
sit
y
31.4oC
0
0.2
0.4
0.6
0.8
1
1.2
10 15 20 25 30 35 40 45
Temperature (oC)
Sp
ecif
ic v
isc
os
ity
32.1oC
ASC – L – Bone PSC – L – Bone
0
0.2
0.4
0.6
0.8
1
1.2
10 15 20 25 30 35 40 45
Temperature (oC)
Sp
ec
ific
Vis
co
sit
y
27.9oC
0
0.2
0.4
0.6
0.8
1
1.2
10 15 20 25 30 35 40 45
Temperature (oC)
Sp
ec
ific
Vis
co
sit
y
29.1oC
ASC – P – Bone PSC – P – Bone
0
0.2
0.4
0.6
0.8
1
1.2
10 15 20 25 30 35 40
Temperature (oC)
Sp
ec
ific
Vis
co
sit
y
31.1oC
0
0.2
0.4
0.6
0.8
1
1.2
10 15 20 25 30 35 40
Temperature (oC)
Sp
ec
ific
Vis
co
sit
y
32.1oC
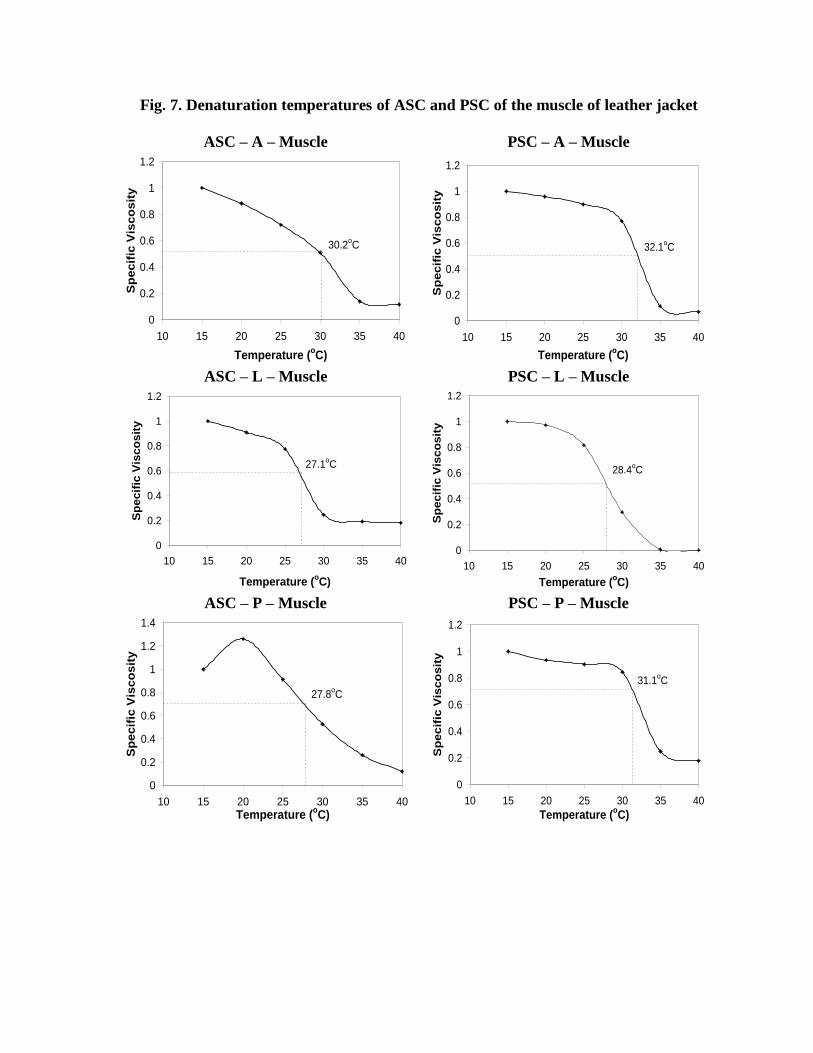
Fig. 7. Denaturation temperatures of ASC and PSC of the muscle of leather jacket
ASC – A – Muscle PSC – A – Muscle
0
0.2
0.4
0.6
0.8
1
1.2
10 15 20 25 30 35 40
Temperature (oC)
Sp
ec
ific
Vis
co
sit
y
30.2oC
0
0.2
0.4
0.6
0.8
1
1.2
10 15 20 25 30 35 40
Temperature (oC)
Sp
ec
ific
Vis
co
sit
y
32.1oC
ASC – L – Muscle PSC – L – Muscle
0
0.2
0.4
0.6
0.8
1
1.2
10 15 20 25 30 35 40
Temperature (oC)
Sp
ec
ific
Vis
co
sit
y
27.1oC
0
0.2
0.4
0.6
0.8
1
1.2
10 15 20 25 30 35 40
Temperature (oC)
Sp
ecif
ic V
isco
sit
y
28.4oC
ASC – P – Muscle PSC – P – Muscle
0
0.2
0.4
0.6
0.8
1
1.2
1.4
10 15 20 25 30 35 40
Temperature (oC)
Sp
ecif
ic V
isc
os
ity
27.8oC
0
0.2
0.4
0.6
0.8
1
1.2
10 15 20 25 30 35 40
Temperature (oC)
Sp
ec
ific
Vis
co
sit
y
31.1oC


5. DISCUSSION
The collagen was extracted from different parts of leather jacket, a trash fish, using
various methods and the results of its different characteristics are discussed to find out its
suitability as a raw material for the biomedical compound, collagen.
5.1 Structural composition of leather jacket
Leather jacket used in this study was an under-utilized trash fish, in which, 53.1%
constituted as wastes excluding the minced meat. Mechanical deboning process normally
removes the significant wastes such as skin and bones (An and Visessanguan, 2000) and 25%
of minced meat could be obtained (Shahidi, 1994). The processing discards may account up
to 75% of the total weight of fish in filleting operations (Dassow, 1979; Piggott, 1986).
However, the wastes generated differ in amount and composition depending on the species
and process employed (An and Visessanguan, 2000). The bulk of the wastes consisted head,
backbone, skin and viscera. An and Visessanguan (2000) also reported that about 60% of the
total weight of fish was generated as waste. It had been reported that the head accounted for
18-24% of the dressed fish weight. In this study, bone accounted for 21% by mechanical
deboning procedure. The processing wastes are used as food component for
aquaculture/poultry, in silage or fish meal preparation as well as value-added and biomedical
compounds preparations. Since leather jacket constituted high proportions of bone (21%),
scales (15%) and skin (13%), it was considered as a very good raw material for the extraction
of biomedical compound, collagen.
5.2. Proximate composition of leather jacket
The proximate composition of the trash fish, leather jacket was quite different from
other fish species (Fig. 2). The muscle had high moisture (86%) and very low protein (7%).
The moisture and crude protein contents in the muscle of fish species generally ranged from

54-80.3% and 16.1%-27.9%, respectively (Sato et al., 1986b). But, the protein content of
leather jacket was quite lower than the reported values of other fish species.
The skin of leather jacket contained low moisture, fairly good amount of protein and
very high ash content. Muyonga et al. (2004) reported that the skin of Nile perch contained
20-22% of protein. In addition, the ash content was also considerably higher in the skin of
adult fish probably because of increased mineralization with age. However, the higher ash
content noticed in the skin could not be attributed to the increased mineralization; instead it
was due to their inherent biochemical composition. Fairly high moisture content (73.4%) was
reported in the skin of brown backed toadfish (Senaratne et al., 2006) and bigeye snapper
(Kittiphattanabawon et al., 2005). Such higher moisture contents were not found in leather
jacket skin. The crude protein, lipid and ash contents on dry weight basis were 90.3%, 1.3%
and 8.4%, respectively for the skin of brown backed toadfish (Senaratne et al., 2006); while
for the leather jacket, it was 42%, 1.44% and 49.2%, respectively.
The bone of leather jacket contained a fairly good proportion of protein (11%) and
high ash contents (18%). The presence of high proportion of ash in the skin and bone of this
fish makes it distinct for other fish species and hence classified under trash fish. However,
the relatively high amounts of protein in the skin and bone proved them as a good source of
raw material for collagen extraction.
5.3. Collagen yield The collagen content of the muscle of leather jacket was 0.673% of total protein
(Table 1). Collagen content in the muscle of teleosts usually ranged from 1 to 4% of the total
protein or of the dry matter (Kubota and Kimura, 1975; Kimura et al., 1979; Sikorski et al.,
1984). The total collagen content in the muscle also varied with species ranging from 0.34 to
2.19% of wet tissue and from 1.6 to 12.4% of crude protein (Sato et al., 1986b). Collagen
content in the ordinary muscle of dark fleshed fish such as sardine, chub mackerel was lower

than those in the white fleshed fish. Collagen content was also higher in wild fish than in
cultured Japanese eel (Sato et al., 1986b). The collagen content in muscle varied not only
with species but also with muscle parts, age, season, nutritional condition within a species
(Sato et al., 1986b).
Yoshinaka et al. (1990) reported that the total collagen content ranged between 0.37%
for red seabream (Paprus niger) to 1.28% for Japanese eel (Anguilla japanicus). Aidos et al.
(1999) have found the total collagen in fresh Atlantic salmon muscle as 0.29% of wet weight.
This value was lower than the values reported for dorsal white muscle in other fish species
(Sato et al., 1986b). Eckhoff et al. (1998) also observed a total collagen content of 0.66% for
the same species. Leather jacket used in this study had lower collagen content when
compared to other fish species reported.
The collagen content in the leather jacket skin was 5.06% of the total protein, which
was higher than that of the muscle. But, the amount of total collagen in the skin of hake was
reported as 34.2% and that of trout as 48.8%. These values were much higher than the
amounts in the muscle (Montero et al., 1990). Collagen content in skin and muscle of catfish
grown in the pond was 44% and 14% of dry tissue, respectively. The effluent treated pond
(ETP) grown fish had increased collagen content in skin and muscle (Sivakumar et al.,
2000). Such higher values were not obtained from the skin of leather jacket. However, the
collagen content in the bone of leather jacket was quite high (44% of total protein). The
collagen content in raw backbone of Baltic cod was 24% of dry weight (Guildberg et al.,
2002; Skierka et al., 2007); while it was 15% of dry weight in the raw bone of leather jacket.
It was evident that the bone of leather jacket could be good a source of collagen than the skin
or muscle.

5.4. Extraction of collagen
The maximum yield of PSC collagen extracted from the skin of leather jacket with
acetic, lactic and propionic acids was 44.19%, 68.21% and 16.38%, respectively (Fig. 3);
while the yields of ASC collagen were 2.69%, 20.67% and 3.78%, respectively. Skierka and
Sadowska (2007) suggested that the acetic and lactic acids are the best solvents for collagen
extraction. They have also reported that the maximal yield of collagen from the skin of Baltic
cod was 60% with citric acid, 90% with acetic or lactic acid and 18% with HCl acid.
The first stage of solubilization was the hydration of fibrous collagen on exposure to
acids. Maximum degree of swelling for cattle skin collagen was near pH 3.0 (Gustavson,
1956). It has been known that the proteins get denatured below pH 2.0. Therefore, digestion
with strong acid for a longer time had led to partial chemical hydrolysis of protein. In
response to it, Skierka and Sadowska (2007) have stated that the collagen extracted in HCl
and citric acids with pepsin treatment contained products of enzymatic hydrolysis, whereas
collagen extracted in acetic and lactic acids had proved that the isolated protein was native.
The lower collagen yields obtained from skin and muscle than bone (P<0.05) using propionic
acid might have also been due to their low pH leading to products of chemical hydrolysis.
Senaratne et al. (2006) have found that the brown backed toadfish skin was not
soluble entirely in 0.5 M acetic acid, but after adding 10 % (w/v) pepsin with acid, fish skin
was completely solubilized resulting in a viscous solution and the yield of collagen after salt
precipitation was 54.3% on the basis of lyophilized dry weight. In this study also, the skin of
leather jacket was not completely solubilized in 0.5 M of the respective acids, giving poor
yield, while pepsin treatment gave higher yields. Significant differences existed among the
different methods followed for the extraction of collagen with acid and pepsin (P<0.05)
Lactic acid was reported to completely solubilize the collagen within 24 h of
extraction (Skierka and Sadowska, 2007), whereas largest increase in solubility of collagen in

citric and acetic acids was observed only after 48h. Short time requirement for the
solubilization of collagen by lactic acid made it as one of the best solvents for the extraction
of collagen with higher yield (P<0.05) than the other acids. Extraction of ASC collagen with
lactic acid by II extraction had improved the yield of Method 1 similar to the amount
extracted by Method 2 (P>0.05), but the same was not noticed with other acids. Lactic acid
was the best solvent for the extraction of collagen in terms of its higher yield, as it could
completely solubilize the collagen in shorter time as evidenced by Skierka and Sadowska
(2007). Lower yield of collagen obtained using some acids was due to the incomplete
solubility and suggested that inter-molecular cross-links were still present in collagen
(Sivakumar et al., 2000). The I extraction of collagen using acid and II extraction with pepsin
(Method 2) improved the collagen yield when compared to extraction with acid alone
(Method 1). However, its yield was lower than PSC extracted by Method 3. The collagen
yields obtained following the three methods of extraction were not similar (P<0.05). The
solubility of collagen after pepsin treatment was reported to increase from 85 to 90 %
(Skierka and Sadowska, 2007).
The PSC collagen yields were maximum from bone of leather jacket than skin and
muscle and they were 54.76%, 94.56% and 51.84% with acetic, lactic and propionic acids,
respectively (Fig.3). The collagen yield obtained from other marine fish skin, bones (Nagai
and Suzuki, 2000b), fins (Nagai, 2004) and scales (Nagai et al., 2004) generally ranged from
30 to 51%. Nagai and Suzuki (2000a) have obtained 40.1 to 53.6% collagen yield from the
bones of skipjack tuna, Japanese seabass, ayu, yellow seabream and horse mackerel. The
higher yield (94.56%) obtained from the bone of leather jacket with the addition of pepsin to
lactic acid was possible after deproteinization with NaOH. Because, it has been earlier
reported that the non-collagenous protein in cod backbone was completely removed by alkali

(NaOH) extraction and it contained 41% of ash and 55 % of collagen in the dry weight basis
(Skierka et al., 2007).
The muscle of leather jacket contained low levels of collagen and PSC yields were
5.34%, 16.78% and 12.33% with acetic, lactic and propionic acids, respectively. The
predominant protein of muscle collagen was also found to get solubilized by pepsin digestion
(Sivakumar et al., 2000), which had increased PSC yield when compared to ASC. The
relative distribution of concentration of collagen fractions in Atlantic salmon muscle showed
23.7% for ASC, 70.5% for PSC and 5.8% for ISC corresponding to 0.74, 1.96 and 0.17 g/kg,
respectively (Aidos et al., 1999). Eckhoff et al. (1998) have also reported a value of 6%, 93%
and 1% for ASC, PSC and ISC, respectively in the muscle of the same species. In leather
jacket, the concentration of ASC and PSC fractions extracted using acetic acid were 14.63%
and 85.36%, respectively. The ASC obtained from the muscle of hake and trout (70%)
(Montero et al., 1990) were found to be higher than the level (55%) reported by Yamaguchi
et al. (1976).
The collagen yield obtained from different parts were statistically different (P<0.05)
with bone giving higher yield followed by skin and muscle, irrespective of ASC or PSC.
Montero et al. (1990) have also reported high collagen yields from the skin of hake (90.6%)
and trout (93.9%) compared to muscle, which are similar to the present findings.
5.5. Electrophoretic characterization of collagen
The existence of at least two different subunits α1 and α2 in the bone and skin collagen
showed that the major collagen is a Type I collagen (Fig. 4a). There were also no significant
differences in molecular weights of α subunits of skin and bone collagen. The electrophoretic
patterns of bone collagen isolated from blackdrum and sheepshead were also reported to be
similar to that of electrophoretic patterns of skin collagen of the same fish species (Ogawa et
al., 2004). Existence of two forms of Type I skin and bone collagen [(α1)2α2 and α1α2α3] was

reported in fish by Kimura et al. (1987). But, both the ASC and PSC collagens isolated from
skin and bone of leather jacket were made up of only two α species (α1 and α2). Similar
patterns were observed for Nile perch (Muyonga et al., 2004) and cuttlefish (Nagai et al.,
2001). It was reported that the soluble collagen from the scale and bone of carp consisted of
(α1)2α2 as a main component and α1α2α3 as a minor component (Kimura et al., 1991).
Therefore, the bone and skin collagen of leather jacket might also contain α1α2α3 subunit as a
minor component. But, the α3 chain could not be separated from the corresponding α1 chain
under the experimental conditions employed, because electrophoretically α3 migrates as
similar as the way of migration of α1 (Kimura and Ohno, 1987; Matsui et al., 1991; Kimura,
1992). But, the presence of α2 as the minor component among the two species suggested that
the skin and bone collagens exist as trimmers consisting of two α1 and one α2 chains of
(α1)2α2.
Cross-linked chains existed as β dimmers or γ trimmers along with non-cross-linked
monomer chains α1 and α2 (Fig. 4a). Presence of dimmers (β) and trimmers (γ) was also
observed in the collagen of bigeye snapper (Kittiphathanabawon et al., 2005), pufferfish
(Nagai et al., 2002), Nile perch (Muyonga et al., 2004) and brown backed toadfish
(Senaratne et al., 2006). The presence of β-component was confirmed that the collagen
contain inter molecular cross-links. The γ component had the ability to renature the native
collagen and this indicated that it was a collagen molecule in which the three chains are
intramolecularly cross-linked (Lewis and Piez, 1964). Leather jacket contained small
quantity of γ trimmers and high proportions of α monomers and β dimmers as reported by
Yamaguchi et al. (1976) and Asghar and Henrickson (1982). The ASC of skin and bone
collagens contained higher proportion of cross-linked components than the PSC indicating

the presence of intra and/or inter molecular cross linking of collagen. The ASC of blackdrum
scale also had a higher β cross linking components compared to PSC (Ogawa et al., 2004).
The molecular weights of α1 and α2 component of skin and bone collagens were 125
kDa and 115 kDa, respectively. The molecular weights of α1 and α2 fractions of bone and
scale collagens of blackdrum and sheepshead were identified as 130 and 110 kDa,
respectively (Ogawa et al., 2004). The collagen extracted from skin of Baltic cod had
chains below 116 kDa in the electrophorogram (Skierka and Sadowska, 2007). The
molecular weights of α1 and α2 chains of ASC isolated from the bigeye snapper were 120 kDa
and 112 kDa, respectively (Nalinanon et al., 2007). It is apparent that the molecular weights
of monomers reported for the collagen extracted from fish skin was in coincidence with the
results obtained in this study.
The muscle collagen of the leather jacket exhibited an entirely different
electrophoretic pattern compared to skin and bone collagens (Fig. 4b). Three distinct α chains
appeared and were designated as viz. α1, α3 and α2 in the order of their elution positions. The
relative amounts of α2 and α3 were very low indicating that most of these collagen chains
were present as β chains which could arise from intramolecular cross-linking of two α chains
(Kimura et al., 1988).
The muscle collagen comprised of three α chains viz. α1, α3 and α2 with molecular
weights 125,120 and 105 KDa, respectively. The molecular weight of α1 chain of muscle
collagen corresponded with α1 chain of skin and bone collagens. However, the molecular
weights of α2 and α3 chains were distinct. Sato et al. (1989) observed a similar SDS-PAGE
pattern comprising three α chains in fish muscle and was typical of Type V collagen.
Existence of two molecular forms of (α1)2α3 and α1α2α3 of Type V collagen in the ordinary
muscle was also reported by Sato et al. (1994). Yata et al. (2001) identified Type V collagen

as a minor component in the skin of horse mackerel, yellow seabream and tiger puffer,
although Type V collagen was found to be the major component in the muscle of these
fishes. Electrophoretic pattern revealed the presence of Type I collagen as major component
in the skin and bone of the leather jacket with (1)2, 3 chains; and Type V collagen as major
component in the muscle with α1α2α3 chains. However, it is suggested that further
characterization with column chromatography could help to identify the presence of minor
proportion of other types of collagen.
5.6. Amino acid composition of collagen
Presence of glycine, alanine and proline in the bone collagen of the leather jacket was a
typical characteristic of all fish collagens (Table 1). The alanine content was almost equal to
that of glycine content in the collagen of leather jacket. Increased alanine content and
absence of cysteine in the skin collagen of blue grenadier (Ramshaw et al., 1988), mackerel
(Kimura, 1985) and catfish (Sivakumar et al., 2000) were also reported.
Higher proline and lower hydroxyproline content in the skin collagen were noticed in
the skin Type I collagen by Yata et al. (2001) and the same was observed in the bone
collagen of the leather jacket. Sato et al. (1988) have indicated that Type I collagen of carp
had relatively high content of alanine and low content of hydroxylysine as observed in this
study. Kimura et al. (1988) reported a relatively low proportion of hydroxyproline/proline in
skin Type I collagen than in the muscle Type I collagen. Its proportion was also low (0.73) in
the bone collagen of leather jacket. Collagen from several other fishes normally had values of
<1 (Ramshaw et al., 1988; Kimura, 1985), but, Sivakumar et al. (2000) have reported higher
values of >1. The hydroxylysine and hydroxyproline contents were similar to that reported
previously by several other workers (Montero et al., 1990; Ogawa et al., 2004). Presence of
high content of glutamic acid in Type I collagen of Atlantic salmon compared to other amino

acids was reported by Aidos et al. (1999). Bone collagen of leather jacket also had relatively
high proportion of glutamic acid.
The content of the amino acids, proline and hydroxyproline was 19.3% in the leather
jacket, which was similar to that reported for hake (16-21%), trout (15-18%) and Nile perch
(19.3-20.0%) (Montero et al., 1990; Muyonga et al., 2004). The total imino acid content was,
however, lower than 25.4% reported for tilapia (Grossman and Bergman, 1992). The
collagen of coldwater fish generally contained 16-18% imino acids (Gilsenan and Ross-
Murphy, 2000). The content of hydroxyproline in fish collagen is considerably lower than
animal collagen, which has around 40-80 residues/1000 amino acid residues in fish as
compared to 100-130 residues/1000 amino acid residues in meat (Sikorski et al., 1984).
Hydroxylysine contributes to the formation and stabilization of cross-links, giving rise to
complex, non-hydrolyzable bonds (Asghar and Henrickson, 1982). The degree of
hydroxylation is very useful in applied technical studies, as it maximizes-cross-linking
capacity which in turn improves the functional capacity (Montero et al., 1990).
5.7. Thermal stability The heat denaturation of leather jacket collagen commenced around 250C and was
substantial between 27-320C (Figs. 5, 6 and 7). At that temperature, the collagen molecules
were ruptured and converted into elements of lower molecular weights (Balian and Bowes,
1977). The thermal stability of collagen varies with fish species (Kimura et al., 1988). Lower
Td values of 150C, 16.80C and 19.40C were reported for cod skin collagen (Rigby, 1968),
Alaskan pollack skin collagen (Kimura and Ohno, 1987) and Chum salmon muscle collagen
(Kimura et al., 1988), respectively. The Td values of the collagen of Pacific whiting skin
(23.3oC), common mackerel skin (26.10C), skipjack tuna skin (29.70C), grasscarp skin
(28.4oC), ayu (29.70C) and brown backed toad bones (280C) were reported to be average
(Kimura et al., 1988; Nagai and Suzuki, 2000b; Kim and Park, 2004; Senaratne et al., 2006;

Zang et al., 2007). Higher Td values of 360C for collagen of Nile perch skin (Muyonga et al.,
2004), 340C for blackdrum and sheepshead bones (Ogawa et al., 2004), 32.50C for carp
muscle (Kimura et al., 1988) were also reported.
The Td values of bone collagen were much higher than those of skin collagen of fish
species such as skipjack tuna (29.70C) and yellow seabream (29.50C) (Nagai and Suzuki,
2000b). In this study, the Td value of the bone collagen of leather jacket was significantly
higher (2-40C) than that of skin collagen (P<0.05). Muscle and skin Type I collagens of the
same teleosts also differed from each other in their thermal stability; the former had a higher
Td values by about 10C than that of the later (Kimura et al., 1988). The muscle collagen of
leather jacket also had higher thermal stability than the skin collagen (P<0.05). This
suggested that the internal body temperature of the teleosts was significantly higher than its
surface body temperature or seawater temperature. The Td value of collagen was more
correlated with the animal body temperature and environmental temperature of their habitat
(Rigby, 1968). Higher Td values reported for crustaceans such as giant river prawn (350C)
and spiny lobster (33.50C) were also seemed to be close to the upper limits of the
environmental temperature for these crustaceans (Kimura and Tanaka, 1986). The collagen
obtained from the fishes of Japanese waters had Td values ranging between 25-280C for
pufferfish, jellyfish, cuttlefish, seabass, chub mackerel, shark and red seabream (Nagai et al.,
2000; Nagai et al., 2001; Nagai et al., 2002; Nagai, 2004; Nagai et al., 2004). Deepsea
redfish skin, scale and bone collagens also had very low Td values of 16.1oC, 17.7oC and
17.5oC, respectively (Wang et al., 2008).
The Td value of fish collagen was first reported to be proportional to the
hydroxyproline content (Piez and Gross, 1960). The decrease in the thermal shrinkage with
the hydroxyproline content had led to the suggestion that the hydroxy group of

hydroxyproline had an important and unique role in the maintenance of the native structure
by virtue of its ability to make hydrogen bonds. The hydroxy group of hydroxyproline was
found to get involved in intermolecular rather than intramolecular cross linking (Piez and
Gross, 1960). Harrington (1958) suggested the ability of the pyrrolidine rings of both proline
and hydroxyproline to direct the geometry of the peptide chain in the regions, in which they
occur, could be an important factor in molecular structure. VonHippel and Harrington (1959)
further stated that restricted rotation about both the C - C= O and peptide bonds adjacent to
the pyrrolidine rings of the imino acids may be a stability factor in an extended polypeptide
chain. The pyrrolidine rings of imino acids play an important role in the intramolecular
stability of collagen by virtue of their ability to stabilize the secondary structure; whereas
intermolecular strength may be in part the result of hydrogen bonding by the hydroxyl groups
of hydroxyproline, serine, threonine and hydroxylysine. Therefore, the molecular structure of
collagen was maintained in large part by restrictions in changes in the secondary structures of
the polypeptide chain imposed by the pyrrolidine rings of proline and hydroxyproline rather
than by hydrogen bonding through the hydroxyl groups of hydroxyproline (Piez and Gross,
1960).
The high thermal stability of muscle collagen was presumably explained by the fact
that the degree of proline hydroxylation in the muscle collagen was higher by 1.5 to 5.2%
than that in the skin protein, as hydroxyproline has been known to play an important role in
stability of the collagen triple helix (Berg and Prockop, 1973). Muyonga et al. (2004) also
inferred that higher Td values for collagen may be attributed to higher imino acid content
rather than higher environmental temperature. Hence, it would be more appropriate that the
Td values of collagen are correlated with the imino acid content, because higher the imino
acid content, the more stable would be the helical structure of collagen molecule (Wong,

1989). Coldwater fish also had low Td values since the imino acid contents were very low
(Piez and Gross, 1960). Accordingly, it may be inferred that the stability of collagen could be
a function of the total content of imino acids and not necessarily related to hydroxyproline,
environmental or animal body temperatures.
Higher Td values were observed for bone and muscle collagens than skin collagen
(P<0.05) as evidenced in Fig. 8.. Although bone and muscle, being the internal parts of the
fish having higher body temperatures, this reason could not be elucidated. The total imino
acids content of the collagen was found responsible for stability of the secondary structure.
The fairly high proportion of these acids (19.3%) in the bone collagen could be correlated
with their higher thermal stability. It was noted that the Td values of the collagen extracted
using lactic acid had lower Td values compared to other acids (P<0.05), amidst its ability to
extract more collagen. Due to high and rapid solubilization of collagen by lactic acid than
other acids, it was expected to break the inter-molecular crosslinks formed by hydrogen
bonds of the hydroxyl groups of hydroxyproline, leading to lower thermal stability.

6. SUMMARY
1. Skin, bones, scales and fins of the leather jacket contributed 53% of the total weight;
while muscle inclusive of offal constituted 47%. The availability of relatively higher
proportions of wastes in this species made them suitable for the preparation of value-
added and biomedical compounds such as collagen.
2. The proximate composition analysis revealed a presence of higher protein content
(21%) in the skin. Muscle had relatively very low level of protein (7%). Ash contents
were unusually high in skin (25%) and bone (19%). Skins and bones of leather jacket
were therefore considered as a good source of raw materials for the extraction of
collagen based on their protein content.
3. The total collagen content estimated based on the hydroxyproline content indicated
that the bone of the leather jacket contained high collagen (44% of the total protein)
followed by skin (5% of the total protein). Higher protein content found in the skin
was not actually connective tissue protein, collagen. Muscle contained very low
content of collagen.
4. Among the different acids used for the extraction of collagen, lactic acid gave the
maximal yield from bone (95%), followed by acetic acid (55%) and propionic acid
(52%). Lactic acid completely solubilized the collagen and was considered as the best
solvent for the extraction of collagen to obtain maximum yield (P<0.05) than the
other two acids.
5. Collagen yield was very higher from the bone (95%) than from skin (68%) and
muscle (17%), when extracted using lactic acid. The same trend was observed with
other acids. Such variations (P<0.05) in the yields of collagen among the structural

parts of fish indicated that the bone of leather jacket was best suitable for collagen
extraction.
6. Among the three methods used for collagen extraction, acid soluble collagen (ASC)
extracted by the Method 1 gave lower yields due to non-solubilization of collagen.
Method 2 which extracted both ASC and PSC showed comparatively better yield.
Maximum collagen yield was achieved by the Method 3 that extracted only PSC, thus
differing statistically (P<0.05). Addition of pepsin (0.1% w/v) completely solubilized
the collagen and improved the collagen yield. To obtain maximum yield, extraction of
PSC following the Method 3 was found more suitable.
7. Collagen was extracted twice with acid or acid with pepsin. The II extraction was
found to contribute more towards the collagen yield than I extraction. Acidification of
raw materials happened during I extraction, while solubilization was observed during
II extraction. It was therefore recommended to have two extractions to completely
extract the collagen from the raw materials, to achieve maximum yield.
8. Electrophoretic studies on collagen revealed the occurrence of non-crosslinked
monomers, along with cross-linked components viz. dimmers and trimmers.
Small quantity of trimmers and high proportions of monomers and dimmers
were found. Lower components indicated solubilization of intramolecularly cross
linked collagen. ASC was found to contain more dimmers than PSC. Presence of
more dimmers in ASC indicated a higher inter molecular crosslinking of collagen
due to lower solubilization by acid; while pepsin had improved the solubilization.

9. The presence of two subunits in the skin and bone collagen was typical of Type I
collagen with a pattern (1)22. In muscle collagen, the electrophoretic pattern was
distinct with three subunits of pattern 123, which was typical of Type V
collagen. The molecular weights of 1 and 2 chains of Type I skin and bone
collagens were 125 kDa and 115 kDa, respectively. On the other hand, the molecular
weights of 1, 3 and 2 chains of Type V muscle collagen were quite different with
125 kDa, 120 kDa and 105 kDa, respectively. Electrophoretic studies were very
helpful in finding out the type of collagen as well as to understand the presence of
intra and inter molecular cross-linked collagen after the extraction.
10. Amino acid composition of bone collagen was studied. Glycine, alanine, proline and
glutamic acid were abundant. Cysteine and tryptophan were absent. The content of
imino acids, such as proline and hydroxyproline was relatively high (19.3%),
however, lower than mammalian collagen. The proportion of hydroxyproline to
proline was also low (0.73). Higher hydroxylation was found to maximize cross-
linking and improve the functional properties of collagen.
11. The denaturation temperatures (Td Values) of bone and muscle collagen (30-320C)
were higher (P<0.05) than that of skin collagen (27-290C). Muscle and bone being
internal body parts had higher temperatures than its surface (skin) temperature. But,
Td values of collagen were more related to imino acid contents rather than
environmental or animal body temperatures. A fairly good proportion of imino acids
in the bone collagen were found responsible for its thermal stability.
12. Acids used for extraction of collagen were also found to influence Td values.
Collagen extracted with lactic acid had lower Td values (P<0.05) when compared

with those extracted with acetic and propionic acids. Lactic acid that rapidly
solubilized the collagen was expected to break the hydrogen bonding formed by the
hydroxyl groups of hydroxyproline and disrupt the intermolecular cross-links, which
in turn lowered the thermal stability.

7. REFERENCES
Aidos, I., O. Lie and M. Espe, 1999. Collagen Content in Farmed Atlantic Salmon (Salmo
salar L.). J. Agri. Food Chem., 47: 1440-1444
AOAC, 1995. Official methods of analysis. 16th edition. Association of Official Analytical
Chemists, Washington, D.C
An, H. and W. Visessanguan, 2000. Recovery of enzymes from seafood processing wastes.
In: Seafood Enzymes-Utilization and Influence on Postharvest Seafood Quality. N.F.
Haard and K. Benjamin (Eds.). Marcel Dekker Inc., New York. 681
*Asghar, A. and R.L. Henrickson, 1982. Chemical, biochemical, functional, and nutritional
characteristics of collagen in food systems. In: Advance in food research: C.O.
Chichester, E.M. Mrata, B.S. Schweigert, (Eds,). Academic press, London, 28: 232-
372
Bae, I., K. Osatomi, A. Yoshida, K. Osako, A. Yamaguchi and K. Hara, 2008. Biochemical
properties of acid-soluble collagens extracted from the skins of underutilized fishes.
Food Chem., 108(1): 49-54
*Bailey, A.J. and N.D. Light, 1989. Structure and localisation of connective tissue
components in muscle. In: Connective tissue in Meat and Meat products, Elsevier
Applied Science, London, 120-140
*Balian, G. and J.H. Bowes, 1977. The structure and properties of collagen. In: A.G. Ward,
A. Courts (Eds,). The science and technology of gelatin. Academic press, London. 1-
31
*Berg, A. and D.J. Prockop, 1973. The thermal transition of a non-hydroxylated form of
collagen: Evidence for a role of hydroxyproline in stabilizing the triple-helix of
collagen. Biochem. Biophys. Res. Commun. 52: 115

*Boedtker, H. and P. Doty, 1955. Uses of Gelatin in Edible Products. The Science and
Technology of Gelatin. A. Ward and A. Courts (Ed.). Academic Press, London
*Boot-Handford, R.P., D.S. Tuckwell, D.A. Plumb, C.F. Rock, and R. Poulsom, 2003. A
novel and highly conserved (pro (alpha) 18XXVII) with a unique expression pattern
and unusual molecular characteristics establishes a new clade within the vertebrate
fibrillar collagen family. J. Biol. Chem., 278: 31067-31077
Bracho, G.E. and N.F. Haard, 1995a. Purification, Characterization and Radiolabelling of
Lingcod (Ophiodon elongatus) Skin type I collagen.J. Food Biochem., 19(4): 285-297
Bracho, G.E. and N.F. Haard, 1995b. Identification of two matrix metalloproteinases in the
skeletal muscle of Pacific rockfish (Sebastes sp.). J. Food Biochem., 19(4): 299-319
Bruggemann, D.A. and M.A. Lawson, 2005. The extracellular matrix of Gadus morhua
muscle contains types III, V, VI and IV collagens in addition to Type I. J. Fish Biol.,
66: 810-821
*Dassow, J.A., 1979. Product yields from various Alaska fish species. Seattle, WA: National
Marine Fisheries Service.
Eckhoff, K.M., I. Aidos, G. Hemre and O. Lie, 1998. Collagen content in farmed Atlantic
salmon (Salmo salar, L.) and subsequent changes in solubility during storage on ice.
Food Chem., 62(2): 197-200
Fernandes, R.M.T., R.G.C. Neto, C.W.A. Paschoal, J.H. Rohling and C.W.B. Bezerra, 2008.
Collagen films from swim bladders: Preparation method and properties. Colloids and
surfaces B: Biointerfaces, 62(1): 17-21
*Gildberg, A., J.A. Arnesen and M. Carlehog, 2002. Utilization of cod backbone by
biochemical fractionation. Process Biochem., 38: 475-480

Gilsenan, P.M. and S.B. Ross-Murphy, 2000. Rheological characterization of gelatins from
mammalian and marine sources. Food Hydrocolloids, 14(3): 191-195
Gomez-Guillen, M.C. and P. Montero, 2001. Extraction of Gelatin from Megrim
(Lepidorhombus boscii) skins with several organic acids. J. Food Sci., 66(2): 213-216
*Gomez-Guillen, M.C., J. Turnay, M.D. Fernandez-Dias, N. Ulmo, M.A. Lizarbe and P.
Montero, 2002. Structural and Physical properties of gelatin extracted from different
marine species: a comparative study. Food Hydrocolloids, 16(1): 25-34
*Grossman S., M. Bergman, 1992. Process for the production of gelatin from fish skins.
Inventors: Bar IIan University, Ramat Gan assignee. U.S Patent 5,093,474.
*Gustavson, K., 1956. The chemistry and reactivity of collagen. Academic press, New York.
102-202
Harrington, W.F., 1958. Effect of Neutral Salts on the Structure of Collagen and Gelatin.
Nature, 181: 997-998
*Hatae, K., A. Tobimatsu, M. Takeyama, and J.J. Matsumoto, 1986. Contribution of the
connective tissues on the texture difference of various fish species. Nippon Suisan
Gakkaishi, 52: 2001-2007
*Helcke, T., 2000. Gelatin, The food technologists, friend or foe?. Int. Food Ingredients, 1:
6-8
Hickman, D., T.J. Sims, C.A. Miles, A.J. Bailey, M. de Mari and M. Koopmans, 2000.
Isinglass/ collagen: Denaturation and functionality. J. Biotechnol., 79(3): 245-257
Hwang, J., Y. Yokoyama, S. Mizuta and R. Yoshinaka, 2006. cDNA cloning and
characterization of Type I procollagen 1 chain in the skate Raja kenojei. Comp.
Biochem. Physiol. Part B: Biochem. Mol. Biol., 144(1): 1-10

Hwang, J., S. Mizuta, Y. Yokayama and R. Yoshinaka, 2007. Purification and
characterization of molecular species of collagen in the skin of skate (Raja kenojei).
Food Chem., 100(3): 921-925
Hwang, J., Y. Yokoyama, S. Lee, S. Mizuta and R. Yoshinaka, 2008. cDNA cloning and
characterization of Type V/XI procollagen 1 chain in the skate, Raja kenojei. Food
Chem., 107(4): 1467-1472
Ikoma, T., H. Kobayashi, J. Tanaka, D. Walsh and S. Mann, 2003. Physical properties of
Type I collagen extracted from fish scales of Pagrus major and Oreochromis
niloticas. Int. J. Biological Macromolecules, 32(3-5): 199-204
*Ishida, Y. T. Fujita and K. Asai, 1981. New detection and separation method for amino acid
by high performance liquid chromatography. J. Chromatogr., 204: 143-148
Kim, J.S. and J.W. Park, 2004. Characterization of Acid-soluble Collagen from Pacific
Whiting Surimi Processing Byproducts. J. Food Sci., 69(8): 637-642
Kimura, S., Y. Akashi and M. Kubota, 1979. Carp connectin: Its contents in white and dark
muscles. Bull. Jap. Soc. Sci. Fish., 45(2): 237-240
Kimura, S., 1983. Vertebrate skin Type I collagen: Comparison of bony fishes with lamprey
and calf. Comp. Biochem. Physiol. Part B: Biochem Mol. Biol., 74(3): 525-528
*Kimura, S., 1985. The interstitial collagens of the fishes. In: Biology of Invertebrate and
lower Vertebrate collagens. A. Bairati R. Garrone (Eds). New York: NATO-ASI
Series, Plenum. 397-408.
Kimura, S. and H. Tanaka, 1986. Partial Characterization of Muscle Collagens from Prawns
and Lobster. J. Food Sci., 51(2): 330-332,339

Kimura, S., Y. Ohno, Y. Miyauchi and N. Uchida, 1987. Fish skin type I collagen: Wide
distribution of an 3 subunit in teleosts. Comp. Biochem. Physiol. Part B: Biochem.
Mol. Biol., 88(1): 27-34
Kimura, S. and Y. Ohno, 1987. Fish type I collagen: tissue-specific existence of two
molecular forms, (1)22 and 123, in Alaska Pollack. Comp. Biochem. Physiol.
Part B: Biochem Mol. Biol., 88(2): 409-413
Kimura, S., X. Zhu, R. Matsui, M. Shijoh and S. Takamizawa, 1988. Characterization of Fish
Muscle Type I Collagen. J. Food Sci., 53(5): 1315- 1318
*Kimura, S., Y. Mayauchi and N.Uchida, 1991. Scale and bone Type I collagens of carp
(Cyprinus carpio). Comparitive. Biochem. Physiol. 99B: 473-476
*Kimura, S., 1992. Wide distribution of the skin type I collagen 3 chain in bony fish. Comp.
Biochem. Physiol., 102B: 780-786
*Kittiphattanabawon, P., S. Benjakul, W. Visessanguan, T. Nagai, and M. Tanaka, 2005.
Characterization of acid-soluble collagen from skin and bone of bigeye snapper
(Priacanthus tayenus). Food Chem., 89: 363-372
*Kubota, M. and S. Kimura, 1975. The distribution of collagen and some properties of
intramuscular collagen in fish. Hikaku kagaku (Lether chem.,). 21: 80-83
*Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of
bacteriophage T4. Nature, 227: 680-685
Lehninger, A.L., 1982. Principles of Biochemistry. CBS Publishers and Distributors Pvt.
Ltd., Delhi (India). P1101. Chap.7. Fibrous Proteins. pp. 147-168
Lewis, M.S. and K.A. Piez, 1964. The Characterization of Collagen from the skin of the
Dogfish Shark, Squalus acanthias. J. Biol. Chem., 239(10): 3336-3340

*Mathews, M.B., 1975. Connective Tissue Macromolecular Structure and Evolution.
Springer-verlag, New York, p.38
Mathew, S., T.V. Sankar and M.K. Mukundan, 1998. Purification and Characterization of
Collagen from Chanos chanos. In: Proceedings of the Symposium ‘Advances and
Priorities in Fisheries Technology’. CIFT, Kochi. 345-349
Mathew, S., 2002. Fish Collagens. CIFT Technology Advisory Series: 8, Agricultural
Technology Information Centre, CIFT, Kochi
*Matsui, R., M. Ishida and S. Kimura, 1991. Characterization of an α3 chain from the Skin
Type I collagen of chum salmon (Oncorhynchus keta). Comp. Biochem. Physiol. 99B:
171-174.
Mizuta, S., S. Fujisawa, M. Nishimoto and R. Yoshinaga, 2005. Biochemical and
immunochemical detection of Types I and V collagens in tiger puffer, Takifugu
rubripes. Food Chem., 89(3): 373-377
*Montero, P. and A.J. Borderias, 1989. Propiedades funcionales delcolagenode pescado en la
industria alimentaria. Informes Tecnicos de investigacaion pesquera, 151: 1-18
Montero, P., J. Borderias, J. Turnay and M.A. Leyzarbe, 1990. Characterization of Hake
(Merluccius merluccius L.) and Trout (Salmo irideus Gibb) Collagen. J. Agric. Food
Chem., 38(3): 604-609
Montero, P., C. Alvarez, M.A. Marti and A.J. Borderias, 1995. Plaice Skin Collagen
Extraction and Functional Properties. J. Food Sci., 60(1): 1-3
Montero, P. and M.C. Gomez-Guillen, 2000. Extracting Conditions for Megrim
(Lepidorhombus boscii) Skin Collagen Affect Functional Properties of the Resulting
Gelatin. J. Food Sci., 65(3): 434-438

Morimura, S., H. Nagata, Y. Uemura, A. Fahmi, T. Shigematsu and K. Kida, 2002.
Development of an effective process for utilization of collagen from livestock and
fish waste. Process Biochem., 37(12): 1403-1412
Muyonga, J.H., C.G.B. Cole and K.G. Duodu, 2004. Characterisation of acid soluble
collagen from skins of young and adult Nile perch (Lates niloticus). Food Chem.,
85(1): 81-89
Nagai, T. and N. Suzuki, 2000a. Preparation and characterization of several fish bone
collagens. J. Food Biochem., 24(5): 427-436
Nagai, T. and N. Suzuki, 2000b. Isolation of collagen from fish waste material–skin, bone
and fins. Food Chem., 68(3): 277-281
*Nagai, T., W. Worawattanamateekul, N. Suzuki, T. Nakamura, T. Ito, K. Fujiki, M. Nakao
and T. Yano, 2000. Isolation and characterization of collagen from rhizostomous
jellyfish (Rhopilema asamushi). Food Chem., 70: 205-208
*Nagai, T., E. Yamashita, K. Taniguchi, N. Kanamori and N. Suzuki, 2001. Isolation and
characterization of collagen from the outer skin waste material of cuttlefish (Sepia
lycidas). Food Chem., 72: 425-429
Nagai, T., Y. Araki and N. Suzuki, 2002. Collagen of the skin of ocellate puffer fish
(Takifugu rubripes). Food Chem., 78 (2): 173-177
Nagai, T., M. Izumi and M. Ishii, 2004. Fish Scale Collagen. Preparation and partial
Characterization. Int. J. Food Sci. Technol., 39: 239-244
*Nagai, T., 2004. Characterization of collagen from Japanese sea bass caudal fin as waste
material. Eur. Food Res. Technol., 218: 424-427

Nalinanon, S., S. Benjakul, W. Visessanguan and H. Kishimura, 2007. Use of pepsin for
collagen extraction from the skin of bigeye snapper (Priacanthus tayenus). Food
Chem., 104(2): 593-601
*Nip, W.K., H.M.K. Zeidan and J.H. Moy, 1981. Amino acid profile of insoluble collagen
isolated from freshwater prawn, Macrobrachium rosenbergii. J. Food Sci., 46: 1633
*Nishihara, T., 1962. Solubilization of insoluble collagen fibers and reconstitution there of.
US Patent No. 3 034 852
Nishimoto, M., S. Mizuta and R. Yoshinaka, 2004. Characterization of molecular species of
collagen in muscles of Japanese amberjack, Seriola quinqueradiata. Chemistry,
84(1): 127-132
Nishimoto, M., R. Sakamoto, S. Mizuta and R. Yoshinaka, 2005. Identification and
characterization of molecular species of collagen in ordinary muscle and skin of the
Japanese flounder Paralichthys olivaceus. Food Chem., 90(1-2): 151-156
Ogawa, M., R.J. Portier, M.W. Moody, J. Bell, M.A. Schexnayder and J.N. Losso, 2004.
Biochemical properties of bone and scale collagens isolated from the subtropical fish
black drum (Pogonia cromis) and sheepshead seabream (Archosargus
probatocepahlus). Food Chem., 88 (4): 495-501
Piez, K.A. and J. Gross, 1960. The Amino Acid Composition of Some Fish Collagens: The
Relation between Composition and Structure. J. Biol. Chem. 235(4): 995-998
*Piez, K.A., E.A. Eigner and M.S. Lewis, 1963. The Chromatographic Separation and
Amino Acid Composition of the Subunits of Several Collagens. Biochem., 2(1): 58-
66
*Pigott, G.M., 1986. Surimi: The high tech raw material for fish flesh. Food Rev. Int., 2: 213-
246

Pranoto, Y., C.M. Lee and H.J. Park, 2007. Characterization of fish gelatin films added with
gellan and -carrageenan processing of fish. Int. J. Food Sci. Technol., 40(5): 301-
343
Ramshaw, J.A.M., J.A. Werkmeister and H.A. Bremner, 1988. Characterization of type I
collagen from the skin of blue grenadier (Macruronus novaezelandiae). Arch.
Biochem. Biophy., 267(2): 497-502
*Rehn, M., T. Veikkola, E. Kukk-valdre, H. Nakamura, M. Ilmonen, C. Lombardo, T
Pihlajaniemi, K. Alitalo and K. Vuori, 2001. Interaction of endostain with integrins
implicated in angiogenesis. Proceedings of the National Academy of Sciences of the
USA, 98: 1024-1029
*Rigby, B.J., 1968. Amino-acid composition and thermal stability of the skin collagen of the
Antarctic ice-fish. Nature, 219: 166-167
Rigo, C., D.J. Hartmann and A. Bairati, 2002. Electrophoretic and immunochemical study of
collagens from Sepia officinalis cartilage. Biochemica et Biophysica Acta (BBA) -
General subjects, 1572(1): 77-84
Saito, M., Y. Takenouchi, N. Kunisaki and S. Kimura, 2001. Complete primary structure of
rainbow trout Type 1 collagen consisting of 1(1)2(1)3(1) heterotrimers. Eur. J.
Biochem., 268: 2817-2827
Sankar, S., S. Sekar, R. Mohan, S. Rani, J. Sundaraseelan and T.P. Sastry, 2008. Preparation
and partial characterization of collagen sheet from fish (Lates calcarifer) scales. Int.
J. Biol. Macromolecules, 42(1): 6-9
Sato, K., R. Yoshinaka, M. Sato and S. Ikeda, 1986a. A Simplified Method for Determining
Collagen in Fish Muscle. Bull. Jap. Soc. Sci. Fish., 52(5): 889-893

Sato, K. R. Yoshinaka, M. Sato and Y. Shimizu, 1986b. Collagen Content in the Muscle of
Fishes in Association with Their Swimming Movement and Meat Texture. Bull. Jap.
Soc. Sci. Fish., 52(9): 1595-1600
Sato, K., R. Yoshinaka, M. Sato and Y. Shimizu, 1987. Isolation of Native Acid–Soluble
Collagen from Fish Muscle. Nippon Suisan Gakkaishi, 53(8): 1431 – 1436
*Sato, K., R. Yoshinaka, M. Sato, Y. Itoh and Y. Shimizu, 1988. Isolation of types I and V
collagens from carp muscle. Comp. Biochem. Physiol., 90B: 155-158
*Sato, K., R. Yoshinaka, Y. Itoh and M. Sato, 1989. Molecular species of collagen in the
intramuscular connective tissue of fish. Comp. Biochem. Physiol. 92B: 87-91
*Sato, K., C. Ohashi, K. Ohtsuki and M. Kawabata, 1991. Type V collagen in trout (Salmo
gardineri) muscle and its solubility change during chilled storage of muscle. J. Agric.
Food Chem., 39: 1222-1225
*Sato, K., K. Sakuma, K. Ohtsuki and M. Kawabata, 1994. Subunit composition of eel
(Anguilla japonica) type V collagen: Evidence for existence of a novel fourth 4 (V)
chain. J. Agri. Food Chem., 42: 675-678
Sato, K., C. Ohashi, M. Muraki, H. Itsuda, Y. Yokoyama, M. Kanamori, K. Ohtsuki and M.
Kawabata, 1998. Isolation of intact Type V collagen from fish intramuscular
connective tissue. J. Food Biochem., 22(3): 213-225
Senaratne, L.S., P. Park and S. Kim, 2006. Isolation and characterization of collagen from
brown backed toadfish (Lagocephalus gloveri) skin. Bioresource technol., 97(2): 191-
197
Shahidi, F., 1994. Seafood processing by-products. In: F. Shahidi, and J.R Botta (Eds.),
Seafoods chemistery, processing, technology and quality. Glasgow: Blackie
Academic professional.pp.11-26

*Sikorski, Z. E., D.N. Scott, and D.H. Bussion, 1984. The role of collagen in the quality and
processing of fish. CRC Crit. Rev. Food Sci. Nutr., 20(4): 301-338
Sivakumar, P., R. Arichandran, L. Suguna, M. Mariappan and G. Chandrakasan, 2000. The
composition and characteristics of skin and muscle collagens from a freshwater
catfish grown in biologically treated tannery effluent water. J. Fish Biol., 56: 999-
1012
Skierka, E., and M. Sadowska, 2007. The influence of different acids and pepsin on the
extractability of collagen from the skin of Baltic cod (Gadus morhua). Food Chem.,
105(3): 1302-1306
Skierka, E., M. Sadowska and A. Karwowska, 2007. Optimization of condition for
demineralization of Baltic cod (Gadus morhua) backbone. Food Chem., 105(1): 215-
218
Sudeepa, E.S., H.N. Rashmi, A.T. Selvi and N. Bhaskar, 2007. Proteolytic bacteria
associated with fish processing waste: Isolation and characterization. J. Food Sci.
Technol., 44(3): 281-284
Suzuki, T., A.S. Srivastava, H. Hashimoto and T. Kurokawa, 2006. Structural comparison of
promoter and coding sequence of type I collagen alpha 1 chain gene duplicates
between zebrafish and flounder/fugu lineages. Comp. Biochem. Physiol. Part D:
Geneomics Proteomics, 1(1): 20-27
*Swatschek, D., W. Schatton, J. Kellermann, W.E.G. Muller and J. Kreuter, 2002. Marine
sponge collagen: isolation, characterization and effects on the skin parameters
surface- pH, moisture and sebum. Eur. J. Pharm. Biopharm., 53: 107-113

*Trevitt, C.R. and P.N. Singh, 2003. Variant Creutzfeldt–Jakob disease: Pathology,
epidemiology and public health implications. Amer. J. Clinical Nutrition, 78: 651S-
656S
*Von Hippel, P. H. and W.F. Harrington, 1959. Enzymic studies of the gelatin to collagen-
fold transition. Biochim. et Biophys. Acta., 36: 427-447
Wang, L., X. An, F. Yang, Z. Xin, L. Zhao and Q. Hu, 2008. Isolation and characterization of
collagens from the skin, scale and bone of deep-sea redfish (Sebastes mentella). Food
Chem., 108(2): 616-623
*Wong, D.W.S., 1989. Mechanism and Theory in Food Chemistry. Van Nostrand Rehinhold,
New York
Woo, J., S. Yu, S. Cho, Y. Lee and S. Kim, 2008. Extraction optimization and properties of
collagen from yellowfin tuna (Thunnus albacares) dorsal skin. Food Hydrocolloids,
22(5): 879-887
Yamaguchi, K., J. Lavety and R.M. Love, 1976. The connective tissues of fish VIII.
Comparative studies on hake, cod and cat fish collagens. Int. J. Food Sci. Technol.,
11(4): 389-399
Yata, M., C. Yoshida, S. Fujisawa, S. Mizuta and R. Yoshinaka, 2001. Identification and
Characterization of Molecular Species of Collagen in Fish Skin. J. Food Sci., 66(2):
247-251
Yoshinaka, R., K. Sato, M. Sato and H. Anbe, 1990. Distribution of Collagen in Body of
Several Fishes. Nippon Suisan Gakkaishi, 56(3): 549
Yoshizato, K., T. Obinata, H. Huang, R. Matsuda, N. Shioya and T. Miyata, 1981. In Vitro
orientation of fibroblasts and myoblasts on aligned collagen film. Develop. Growth
and Differ., 23(2): 175-184

Zhang, Y., W. Liu, G. Li, B. Shi, Y. Miao and X. Wu, 2007. Isolation and partial
characterization of pepsin-soluble collagen from the skin of grass carp
(Ctenopharyngodon idella). Food Chem., 103(3): 906-912
* Originals are not referred





























