Embed Size (px)
Citation preview
448 NATURE CELL BIOLOGY VOLUME 7 | NUMBER 5 | MAY 2005
N E W S A N D V I E W S
Extending the court for cortactin: from thecortex to the GolgiMichael M. Kessels and Britta Qualmann
Vesicle formation at the trans-Golgi network may be mechanistically more similar to endocytic vesicle formation at the plasma membrane than previously thought. Both processes share common components including the dynamin-binding protein cortactin.
Forming vesicles from donor membrane(s) is a complicated process involving components that are thought to drive membrane invagination, control vesicle size and promote vesicle fission.
At the plasma membrane and at the trans-Golgi network (TGN), vesicle formation has long been speculated to utilize similar molecu-lar mechanisms, because both processes involve clathrin coats, highly homologous adaptor pro-tein complexes and additional related coat com-ponents. A report by Cao et al. in this issue1 now adds the dynamin-binding protein cortactin to a growing list of proteins that were formerly shown to function in endocytic internalization but in fact also work at Golgi membranes.
Previous work had shown that the GTPase dynamin is a common element of membrane dynamics at the plasma membrane and at the Golgi2–4. Dynamins can oligomerize into tubu-lar structures around lipids. In vivo, dynamin oligomers are seen as elongated, stable collars around the neck of clathrin-coated pits when dynamin’s GTPase activity is blocked. This has led to the hypothesis that dynamin oligomeri-zation and GTPase-regulated conformational changes within oligomers may provide the force required for vesicle fission3,4. Dynamins work with SH3 (src-homology 3) domain-con-taining accessory proteins, including cortactin, that associate with the carboxy-terminal pro-line-rich domain of dynamins.
Dynamins are crucial factors for vesicle for-mation at the plasma membrane3,4. The localiza-tion of the ubiquitous dynamin 2 isoform to the Golgi, although intensely disputed in the field for years, supports the idea that in addition to coat formation and cargo recruitment, the machinery for vesicle fission at the TGN and plasma mem-brane may also be similar. Consistent with its cellular localization, interference with dynamin 2 function inhibited vesicle formation from the
Golgi apparatus2,3. Cortactin, an F-actin- and dynamin-binding protein, had previously only been described to localize to dynamic F-actin-containing structures, such as lamellipodia, and to clathrin-coated pits at the cell surface. The amino-terminal part of cortactin contains F-actin-binding repeats and a stretch that asso-ciates with and stimulates the actin nucleation
complex, Arp2/3 (refs 5, 6). Because cortactin is known to bind dynamin 2 (ref. 7), Cao et al. investigated whether cortactin was present at the TGN1. Their data clearly show that cortactin localizes mainly to a perinuclear area that over-laps with the Golgi. Immunoelectron micros-copy furthermore demonstrates the presence of cortactin at intracellular membranes that have
Michael M. Kessels and Britta Qualmann are at the Department of Neurochemistry and Cell Biology, Research Group Membrane Trafficking and the Cytoskeleton, Leibniz Institute for Neurobiology, 39118 Magdeburg, Germany.e-mail: [email protected]
PRD
PRD
PRD
PRD
SH3
SH3
SH3
SH3
SH3
A
A
A
A
Dynamin
Cortactin
Arp2/3complex
Actin
Membrane
a
b
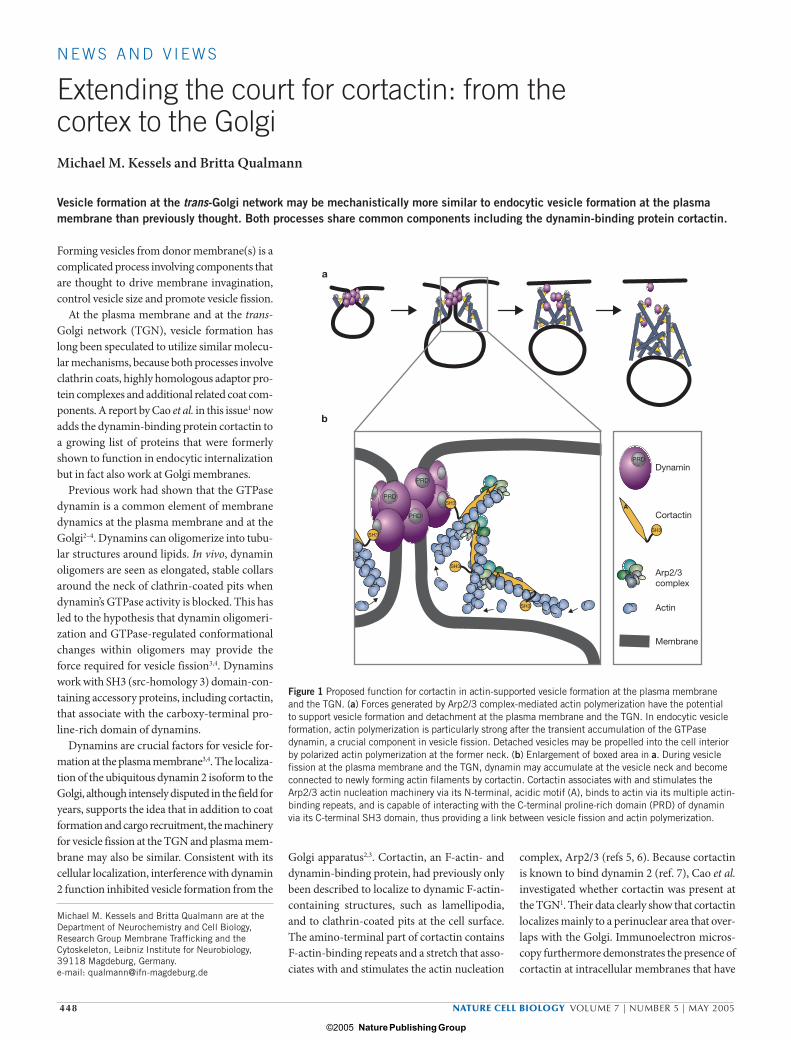
Figure 1 Proposed function for cortactin in actin-supported vesicle formation at the plasma membrane and the TGN. (a) Forces generated by Arp2/3 complex-mediated actin polymerization have the potential to support vesicle formation and detachment at the plasma membrane and the TGN. In endocytic vesicle formation, actin polymerization is particularly strong after the transient accumulation of the GTPase dynamin, a crucial component in vesicle fission. Detached vesicles may be propelled into the cell interior by polarized actin polymerization at the former neck. (b) Enlargement of boxed area in a. During vesicle fission at the plasma membrane and the TGN, dynamin may accumulate at the vesicle neck and become connected to newly forming actin filaments by cortactin. Cortactin associates with and stimulates the Arp2/3 actin nucleation machinery via its N-terminal, acidic motif (A), binds to actin via its multiple actin-binding repeats, and is capable of interacting with the C-terminal proline-rich domain (PRD) of dynamin via its C-terminal SH3 domain, thus providing a link between vesicle fission and actin polymerization.
May N&V Final.indd 448May N&V Final.indd 448 15/4/05 1:09:20 pm15/4/05 1:09:20 pm
Nature Publishing Group© 2005

NATURE CELL BIOLOGY VOLUME 7 | NUMBER 5 | MAY 2005 449
N E W S A N D V I E W S
Golgi-like morphology. This suggests that cort-actin may directly or indirectly associate with Golgi membranes. Indeed, immobilized anti-cortactin antibodies isolated Golgi-like mem-brane compartments.
Dynamin-binding proteins such as profilin, Abp1 and syndapin II that act in the endo-cytic pathway8 are present at the Golgi and also seem to be involved in different Golgi transport processes9–11. Interestingly, all these proteins, including cortactin, are thought to functionally link endocytic vesicle formation and the actin cytoskeleton8. It has been sug-gested that they control different aspects of the actin cytoskeleton function in endocytic vesicle formation, such as spatial organization of microdomains, membrane deformation, dis-solution of the cortical barrier, force generation during and/or after fission, and actin-driven motility of liberated vesicles8. Recently, actin cytoskeletal structures were discovered at the Golgi and they seem to have an important role in maintaining Golgi organization12. Similarly to endocytic vesicle formation, actin dynamics mediated by the Arp2/3 complex and N-WASP (neural Wiskott–Aldrich syndrome protein) are also involved in retrograde transport from the Golgi12. Therefore, by analogy with the process at the plasma membrane, the actin cytoskeleton is also likely to contribute to membrane trans-port processes at the Golgi.
How is cortactin associated with the Golgi? Because cortactin was present in very low amounts in subcellular fractions that contain purified Golgi membranes, the association seems to be relatively weak and/or indirect in nature. Cortactin may be recruited via dynamin 2 or may associate with the actin cytoskeleton at the Golgi. The authors’ findings favour the latter idea. The presence of cortactin and dynamin 2 in Golgi membrane preparations is only seen when actin is present after add-ing GTPγS. Treatment with latrunculin A and brefeldin A, inhibitors of actin polymerization and of the GTPase Arf1 that triggers actin polymerization, indicate that Arf1-dependent actin polymerization is important for recruit-ing cortactin and dynamin 2 to the Golgi1. Interestingly, cortactin’s binding affinity for dynamin 2 is known to increase by an order of magnitude upon cortactin’s association with F-actin- and Arp2/3-containing structures in vitro13. Additional experiments by Cao et al.1 show that Golgi association of dynamin 2 is reduced upon injection of cortactin antibod-ies or a cortactin peptide that contains a part
of its SH3 domain, indicating that dynamin 2 recruitment to the Golgi depends on cortac-tin. Overexpression of truncated cortactin that lacks the dynamin-binding SH3 domain sup-pressed localization of endogenous dynamin 2 to the Golgi. Because dynamin binding to endogenous cortactin is unaffected in this overexpression experiment, an explanation for this surprising observation might be that the overexpressed cortactin mutant may block the association of endogenous cortactin with actin at the Golgi.
Although the question of what cortactin’s cytoskeletal function does in Golgi vesicle for-mation is still largely unanswered, Cao et al.1 demonstrate that cortactin is important for exit from the TGN. Overexpression of cortac-tin mutants that lack the SH3 domain disrupt Golgi organization, resulting in a swollen mor-phology and distended cisternae. Interference with cortactin function and/or its interactions by the introduction of anti-cortactin antibod-ies or an excess of cortactin fragments results in impaired transport from the TGN to the plasma membrane and to late endosomes. The study also provides evidence that two forms of exit from the TGN — clathrin-independent VSV-G exit and the clathrin-dependent exit of mannose-6-phosphate receptors from the TGN — are affected. Because both the dynamin 2 proline-rich domain as well as the cortactin SH3 domain undergo interactions with sev-eral binding partners that may be affected by overexpression of deletion mutants of dynamin 2 and cortactin, it will be important to examine in detail whether the observed effects are speci-ficially due to disruption of putative cortactin/dynamin 2 complexes at Golgi membranes.
Detailed understanding of the functional relevance of cortactin binding to actin fila-ments and the Arp2/3 complex in Golgi transport processes will require knowledge of the molecular composition of cortactin-containing complexes at individual Golgi subdomains. Cortactin localization to the cis side of the Golgi apparatus1 suggests that it might also be involved in the budding of retrograde transport carriers along with the Arp2/3 complex, N-WASP and Cdc42. However, the prominent Arp2/3 complex activator N-WASP is absent from the TGN14. Therefore, other Arp2/3 activators must control actin nucleation at the TGN, and the presence of cortactin at the TGN as shown by Cao et al.1 suggests that cortactin may be one of them.
Despite the elegant picture of apparent molecular similarities in vesicle formation at the plasma membrane and the TGN, some findings of Cao et al.1 indicate that there are also likely to be significant differences. For example, injecting anti-cortactin antibod-ies disrupted dynamin 2 localization to the Golgi but did not have any effect on dynamin 2 recruitment to clathrin-coated pits in endo-cytosis6. Likewise, disruption of the actin cytoskeleton affected cortactin localization to endocytic pits but not that of dynamin 2 (ref. 13). Furthermore, the apparent dependence of dynamin 2 recruitment on cortactin at the Golgi is not consistent with the sequence of events observed for receptor-mediated endo-cytosis. Here, dynamin peaks first and actin and the Arp2/3 complex begin accumulating only later, after the vesicle has departed15. Thus, mechanistic studies of the vesicle formation at the TGN that address the role of dynamin 2, and of accessory components that bind to dynamin and may affect its function in fission, are necessary to resolve these issues. Such stud-ies will most probably need to include in vitro reconstitution of Golgi transport processes and will hopefully clarify the extent to which the molecular machines and mechanisms that are involved in Golgi transport resemble those that work in endocytosis.
This work1 has torn out another brick from the wall that prevents us from completely understanding vesicle formation at the Golgi. It provides an attractive crack into which fur-ther molecular tools and levers can be posi-tioned to enlarge the opening and to expand our understanding.
1. Cao, H. et al. Nature Cell Biol. 7, 483–492 (2005).2. McNiven, M. A., Cao, H., Pitts, K. R. & Yoon, Y. Trends
Biochem. Sci. 25, 115–120 (2000).3. Hinshaw, J. E. Annu. Rev. Cell Dev. Biol. 16, 483–519
(2000).4. Sever, S., Damke, H. & Schmid, S. L. Traffic 1, 385–
392 (2000).5. Olazabal, I. M. & Machesky, L. M. J. Cell Biol. 154,
679–682 (2001).6. Cao, H. et al. Mol. Cell. Biol. 23, 2162–2170
(2003).7. McNiven et al. J. Cell Biol. 151, 187–198 (2000).8. Qualmann, B. & Kessels, M. M. Int. Rev. Cyt. 220,
93–144 (2002).9. Fucini, R. V., Chen, J.-L., Sharma, C., Kessels, M. M. &
Stamnes, M. Mol. Biol. Cell 13, 621–631 (2002).10. Kessels, M. M. & Qualmann, B. J. Cell Sci. 117, 3077–
3086 (2004).11. Dong, J. et al. Biochim. Biophys. Acta 1497, 253–260
(2000).12. Stamnes, M. Curr. Opin. Cell Biol. 14, 428–433
(2002).13. Zhu et al. J. Cell Sci. 118, 807–817 (2005).14. Matas, O. B., Martinez-Menarguez, J. A. & Egea, G.
Traffic 5, 838–846 (2004).15. Merrifield, C. J., Qualmann, B., Kessels, M. M. &
Almers, W. Eur. J. Cell Biol. 83, 13–18 (2004).
May N&V Final.indd 449May N&V Final.indd 449 15/4/05 1:09:22 pm15/4/05 1:09:22 pm
Nature Publishing Group© 2005










![A Golgi-Released Subpopulation of the Trans-Golgi · A Golgi-Released Subpopulation of the Trans-Golgi Network Mediates Protein Secretion in Arabidopsis1[OPEN] Tomohiro Uemura,a,b,2,3,4](https://img.pdfslide.us/doc/110x75/5eda9f5a09f66a09130ba5a1/a-golgi-released-subpopulation-of-the-trans-golgi-a-golgi-released-subpopulation.jpg)