Embed Size (px)
Citation preview

Expression profiles of peroxiredoxin proteins of the rodent
malaria parasite Plasmodium yoeliiq
Shin-ichiro Kawazua,*, Tomoyoshi Nozakib,e, Takafumi Tsuboic,d, Yoko Nakanoa,Kanako Komaki-Yasudaa,f, Nozomu Ikenouea, Motomi Toriic, Shigeyuki Kanoa
aResearch Institute, International Medical Center of Japan, 1-21-1 Toyama, Shinjuku-ku, Tokyo 162-8655, JapanbDepartment of Parasitology, National Institute of Infectious Diseases, 1-23-1 Toyama, Shinjuku-ku, Tokyo 162-8640, Japan
cDepartment of Molecular Parasitology, Ehime University School of Medicine, Shigenobu-cho, Ehime 791-0295, JapandCell-Free Science and Technology Research Center, Ehime University, Matsuyama, Ehime 790-8577, Japan
ePrecursory Research for Embryonic Science and Technology, Japan Science and Technology Corporation, JapanfDomestic Research Fellow, Japan Society for the Promotion of Science, Japan
Received 14 April 2003; received in revised form 25 June 2003; accepted 2 July 2003
Abstract
Patterns of expression of the 2-Cys and 1-Cys peroxiredoxin (Prx) proteins of the rodent malaria parasite Plasmodium yoelii during its life
cycle were observed by immunofluorescent antibody staining and confocal laser scanning microscopy. 2-Cys Prx was expressed in the
parasite cytoplasm throughout the life cycle, and the thioredoxin (Trx)-peroxidase activity of 2-Cys Prx revealed with the recombinant
protein suggested that the Prx is constitutively expressed and, thus, likely plays a housekeeping role in the parasite’s intracellular redox
control. In contrast, 1-Cys Prx showed stage-specific expression in blood-stage parasites. The limited expression of 1-Cys Prx in the
trophozoite cytoplasm suggests that 1-Cys Prx may be involved in haemoglobin metabolism by the parasite, which generates a prooxidative
haem iron and increases intracellular oxidative stress. The antioxidant activity of 1-Cys Prx was tested for its ability to protect yeast enolase
against inactivation of the mixed-function oxidation system. Differential expression of the two Prx proteins during the erythrocytic and insect
stages suggests the importance of these proteins in protecting parasites against oxidative stress, which is generated by the parasite’s
metabolism and also from the environment.
q 2003 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved.
Keywords: Antioxidant; Insect stage; Malaria; Peroxiredoxin; Plasmodium yoelii
1. Introduction
As Plasmodium spp. (malaria parasites) actively pro-
liferate during asexual development in the erythrocytes of
their vertebrate hosts, the parasites are subjected to the toxic
effects of reactive oxygen species (ROS) (Becker et al.,
1994; Flohe et al., 1999). The significant decrease in
glutathione levels in parasite-infected erythrocytes suggests
that the parasites require efficient antioxidant systems
(Muller et al., 2001). Because Plasmodium spp. in
erythrocytes are susceptible to oxidative stresses (Postma
et al., 1996), their antioxidant defences appear to be
potential targets for malaria chemotherapies (Krauth-Siegel
and Coombs, 1999). Malaria parasites develop sexually in
the digestive tract (midgut) of mosquitoes and then mature
intracellularly in the salivary gland. In these environments,
the parasites are also likely to be under the oxidative stress
(Han et al., 2000; Kanzok et al., 2001).
Proteins that are structurally homologous to the thiol-
specific antioxidant of yeast (Chae et al., 1994) have been
identified in all living organisms from bacteria to human and
are referred to as peroxiredoxins (Prx) (Chae et al., 1999).
A sequence alignment among family members revealed the
existence of two groups of Prx that differ in the number of
conserved cysteine residues. The 2-Cys Prx contains both
the conserved residues (Cys47 and Cys170), whereas
0020-7519/$30.00 q 2003 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved.
doi:10.1016/S0020-7519(03)00184-X
International Journal for Parasitology 33 (2003) 1455–1461
www.parasitology-online.com
q Nucleotide sequence data reported in this paper are available in the
GenBank, EMBL and DDBJ databases under the accession numbers
AB089300 and AB089301.* Corresponding author. Tel.: þ81-3-3202-7181x2878; fax: þ81-3-
3202-7364.
E-mail address: [email protected] (S.-i. Kawazu).

the 1-Cys group lacks the conserved Cys170 residue (Chae
et al., 1999). Several Prx proteins act as the terminal
peroxidase that reduces hydrogen peroxide and organic
hydroperoxides with electrons donated by the thioredoxin
(Trx) system (Chae et al., 1994) or other physiological
reductants (Kang et al., 1998; Fisher et al., 1999).
We and others recently reported the expression of 1-Cys
and 2-Cys Prx proteins in blood-stage Plasmodium
falciparum (Kawazu et al., 2000, 2001; Rahlfs and Becker,
2001; Krnajski et al., 2001). These proteins, which have
thiol-dependent peroxidase activities, are expressed during
the trophozoite stage of the parasites (Kawazu et al., 2000;
Krnajski et al., 2001). The 2-Cys Prx of P. falciparum
(PfTPx-1) was suggested to be the terminal peroxidase of
the Trx system during the blood stage, whereas physio-
logical electron donors for the 1-Cys Prx (Pf1-Cys-Prx) are
not known (reviewed in Rahlfs et al., 2002).
Despite such data accumulated from blood-stage para-
sites, the function of Plasmodium Prx proteins during the
insect stage remains poorly understood. In this report, we
examined the expression patterns of 2-Cys and 1-Cys Prx
proteins of the rodent malaria parasite Plasmodium yoelii
during the blood and insect stages by immunofluorescent
antibody staining and confocal laser scanning microscopy.
The antioxidant activities of P. yoelii Prx were also tested
with recombinant proteins.
2. Materials and methods
2.1. Parasites
Plasmodium yoelii 17XL was maintained by mosquito
transmission in Anopheles stephensi interspersed by a
maximum of two serial passages in Crj:ICR mice purchased
from Charles River, Japan. Blood-stage parasites used for
nucleic acid and protein extractions were prepared from
blood of parasite-infected mice. Blood was passed through a
CF11 column (Whatman) to remove leucocytes, and the
parasite-infected erythrocytes were lysed with phosphate-
buffered saline (PBS) containing 0.05% saponin. The
parasite pellet was washed several times with PBS, snap-
chilled in liquid nitrogen, and stored at 280 8C until use.
The animal experiments in this study were carried out either
at Ehime University School of Medicine or at the
International Medical Center of Japan in compliance with
the Guide for Animal Experimentation.
2.2. Ookinete culture and sporozoite preparation
P. yoelii ookinetes were obtained by in vitro culture
of the parasite-infected blood as described previously (Tsuboi
et al., 1997). For the sporozoite preparation, mosquitoes were
fed on P. yoelii-infected mice. Fourteen days after feeding,
salivary glands of the mosquitoes were dissected and minced
in M199 medium (Gibco BRL) containing 0.2% bovine
serum albumin to release sporozoites. Parasite-infected
blood, ookinetes, and sporozoites were spotted onto Multitest
slides (Flow Laboratories) and fixed with ice-cold acetone for
indirect immunofluorescence assay.
2.3. Cloning and expression of P. yoelii peroxiredoxins
Approximately 400–500 bp of the 50 regions of Prx
cDNAs were obtained from the P. yoelii expressed sequence
tags database. These fragments (GenBank accession
numbers BM166616 and BM162294) were identified as
those sequences that had significant identities to the Pf1-
Cys-Prx and PfTPx-1 cDNAs of P. falciparum (GenBank
accession numbers AB020595 and AB037568, respec-
tively). The 30 regions of the P. yoelii cDNAs were obtained
by 30 rapid amplification of cDNA ends (RACE)–PCR.
Template cDNA was synthesised from blood-stage parasite
mRNA with Ready-To-Go You-Prime First-Strand Beads
and Not I–(dT)18 primer (Amersham Pharmacia Biotech).
The coding sequence of 1-Cys Prx was amplified by PCR
from blood-stage parasite cDNA with primers 50-CGC GGA
TCC ATG GGA TAT CAT TTA GGA-30 and 50-CGC CTC
GAG TCA TAA ATT AAC AAA TCT-30. The coding
sequence of 2-Cys Prx was amplified by PCR with the
primers 50-CGC GGA TCC ATG CCA TCA ATT GTA
GGA-30 and 50-CGC CTC GAG TTA TAA ATT CGA TAA
ATA-30 from the same cDNA. The primers, which
contained a Bam HI or Xho I site (indicated in italics)
adjacent to the initiation or the termination codon,
respectively (indicated by underline), were designed from
the EST-derived 50 end sequences and the 30 end sequences
determined in this study. PCR products were digested with
Bam HI and Xho I and ligated into expression vector pGEX-
6P-1 (Amersham Pharmacia Biotech). Recombinant plas-
mids were transformed into Escherichia coli strain BL21,
and expression of N-terminal glutathione S-transferase
fusion protein was induced with isopropyl-b-D-thiogalacto-
side at 0.3 mM. Proteins were purified with the Glutathione
Sepharose 4B Column Chromatography System (Amersham
Pharmacia Biotech). The glutathione S-transferase-tag of
the fusion protein was cleaved by PreScission protease
(Amersham Pharmacia Biotech) and then removed with the
glutathione S-transferase–glutathione affinity system.
2.4. Western blot analysis
Recombinant (rPyPrx) proteins or extracts prepared from
blood-stage parasites were solubilised in SDS–PAGE
sample buffer (Laemmli, 1970). After separation by
SDS–PAGE, the proteins were transferred electrophore-
tically to polyvinylidene difluoride sheets (Immobilon
Transfer Membranes; Millipore) and reacted with either
antisera (1:200) or antibodies (25 mg/ml) to recombinant
P. falciparum Prx (rPfPrx) proteins. Antisera against rPfPrx
proteins were produced in rabbits (Kawazu et al., 2001).
The IgG fractions of the antisera were prepared with HiTrap
S.-i. Kawazu et al. / International Journal for Parasitology 33 (2003) 1455–14611456

rProteinA affinity columns (Amersham Pharmacia Biotech)
according to the method of Miller and Stone (1978).
Immune complexes were visualised with horseradish
peroxidase-conjugated anti-rabbit IgG antibody (Cappel).
2.5. Antioxidant activity assay
Antioxidant activities of the recombinant proteins were
examined with mixed-function oxidation (MFO) system
(Kim et al., 1998) with slight modifications. Briefly,
inactivation mixtures (50 ml) containing 50 mM HEPES
(pH 7.0), 10 mM dithiothreitol and 3 mM FeCl3 were pre-
incubated with or without rPyPrx protein at 30 8C for 30
min. After pre-incubation, 0.025 U of yeast enolase
(Oriental Yeast) was added, and the inactivation mixture
was incubated on ice for another 30 min. The enolase
activity in the reaction mixture was assayed in 1.0 ml of
assay mixture containing 50 mM Tris–Cl (pH 7.5), 1 mM
MgCl2, and 1 mM 2-phosphoglyceric acid (Sigma Aldrich).
The production of phosphoenolpyruvate was monitored as
the increase in A240 at room temperature for 100 s.
Nicotinamide-adenine dinucleotide phosphate, reduced
(NADPH) oxidation coupled to the reduction of H2O2 by
rPyPrx protein in the presence of the Trx and Trx reductase
(TrxR) system was evaluated as described (Fisher et al.,
1999). NADPH oxidation was monitored as the decrease in
A340 at room temperature in a 0.1-ml reaction mixture
containing 50 mM HEPES (pH 7.0), 250mM NADPH, 10mM
E. coli Trx (Wako Pure Chemical), 3 U E. coli TrxR (Sigma
Aldrich), 250 mM H2O2, and 4.0 mM of rPyPrx protein.
2.6. Indirect immunofluorescence microscopy
Acetone-fixed parasites on Multitest slides prepared as
described above were blocked with PBS containing 5% non-
fat dry milk for 30 min at 37 8C. Slides were then incubated
simultaneously with the rabbit antisera to rPfPrx proteins
(1:100) and mouse monoclonal antibody to Pys 25 (1:200,
Tsuboi et al., 1997), which was used for identification of
gametocytes (Kimura et al., 1999), for 60 min at 37 8C and
rinsed with PBS. After incubation with fluorescein isothio-
cyanate (FITC)-conjugated anti-rabbit IgG antibody (1:50,
Cappel) and Alexa Fluor 546-conjugated anti-mouse IgG
antibody (1:50, Molecular Probes) for 30 min at 37 8C, slides
were rinsed with PBS. 40,6-Diamidino-2-phenylindole
(DAPI) staining was done after the second antibody reaction
to visualise the parasite nuclei. The slides were mounted
under cover glasses in Prolong Antifade Kit (Molecular
Probes) and observed with a laser scanning microscopes
(ECLIPSE C1, Nikon and Model LSM510, Carl Zeiss).
2.7. Quantitative real time reverse transcription
PCR (RT–PCR)
Total RNA (approximately 300 ng) isolated from the
blood-stage parasites with TRIzol Reagent (Gibco BRL)
was reverse transcribed with Ready-To-Go You-Prime
First-Strand Beads and pd(N)6 random hexamers (Amer-
sham Pharmacia Biotech). The polymerase chain reaction
was performed with approximately 100 pg of template
cDNA and the sequences-specific oligonucleotide primers
on an ABI PRISM 7700 Sequence Detection System
(Applied Biosystems). Continuous fluorescence observation
of amplifying DNA was done with SYBR Green PCR
Master Mix Kit (Applied Biosystem). After cycling, PCR
products were checked by electrophoresis on agarose gels to
confirm the specificity of each amplification. To compare
the relative amounts of PCR products, the fluorescence
intensity was recorded by cycles for each amplification and
analysed with the ABI Prism Sequence Detection System
software Version 1.7 (Applied Biosystems).
3. Results
3.1. Cloning, sequencing, and expression of P. yoelii
Prx genes
The coding sequences of P. yoelii 2-Cys Prx (GenBank
accession no. AB089301) and 1-Cys Prx (GenBank
accession no. AB089300) were 83.1 and 76.8%, respec-
tively, identical at the amino acid level to those of the
corresponding P. falciparum Prx proteins. These genes were
identified on the MALPY00116 (2-Cys Prx) and
MALPY01293 (1-Cys Prx) contigs, respectively, in the
P. yoelii genome database of the Plasmodium Genome
Consortium PlasmoDB (PlasomDB; http://PlasmoDB.org).
The database search revealed that these genes did not
contain introns. The deduced amino acid sequences of
P. yoelii Prx proteins do not contain putative mitochondrial
or nuclear localisation signals (Park et al., 2000).
Recombinant P. yoelii Prx (rPyPrx) proteins were
produced to test if rabbit anti-sera raised against recombi-
nant P. falciparum Prx (rPfPrx) proteins recognise the
homologous proteins in P. yoelii. The rPyPrx proteins had
molecular masses of 22 kDa (2-Cys Prx) and 25 kDa (1-Cys
Prx) on 12.5% SDS–PAGE gels, which are consistent with
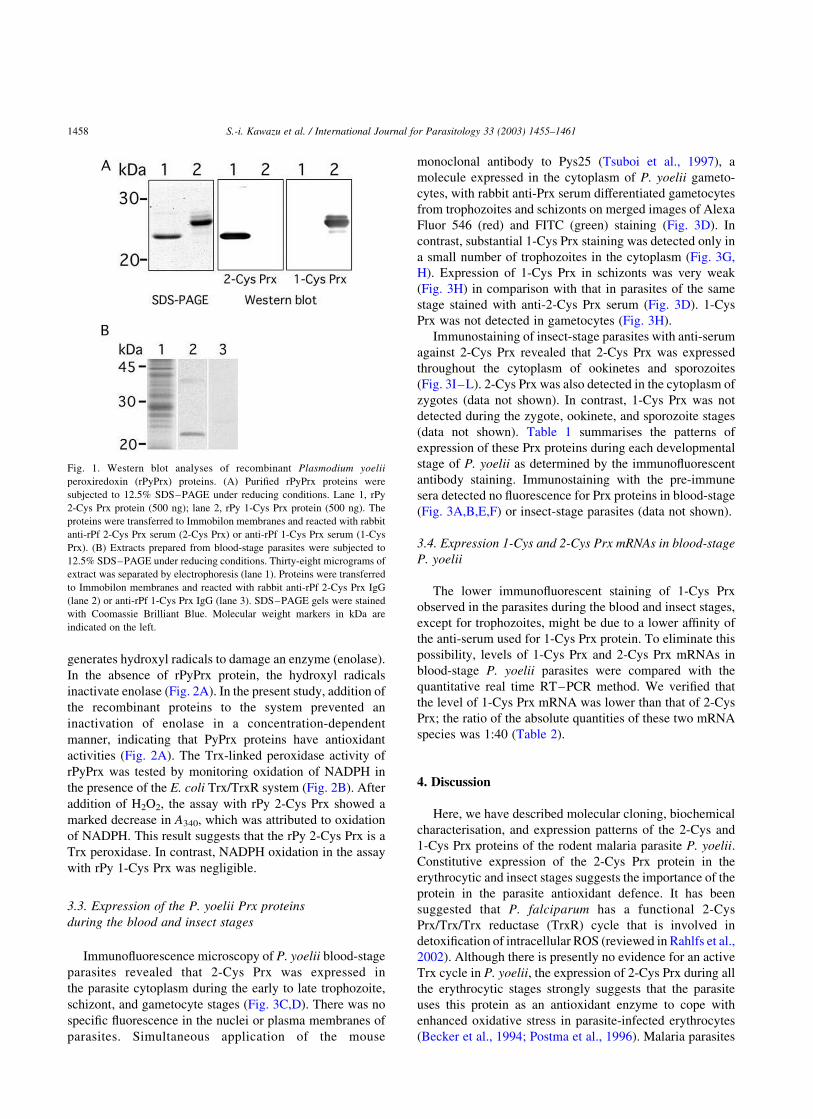
the sizes predicted from the amino acid sequences (Fig. 1A).
On Western blots, each anti-rPfPrx serum specifically
recognised the appropriate rPyPrx (Fig. 1A). These antisera
also specifically recognised native proteins with molecular
masses of 22 kDa (2-Cys Prx) and 25 kDa (1-Cys Prx) in the
blood-stage parasites of P. yoelii on Western blots (Fig. 1B).
An additional band with a molecular mass of approximately
40 kDa was detected on Western blots of 2-Cys Prx (Fig. 1B,
lane 2) and is the dimeric form of the protein (Chae et al.,
1994; Kawazu et al., 2001).
3.2. Antioxidant activities of the rPyPrx proteins
The antioxidant activity of rPyPrx protein was evaluated
with the MFO system. This assay is based on a system that
S.-i. Kawazu et al. / International Journal for Parasitology 33 (2003) 1455–1461 1457
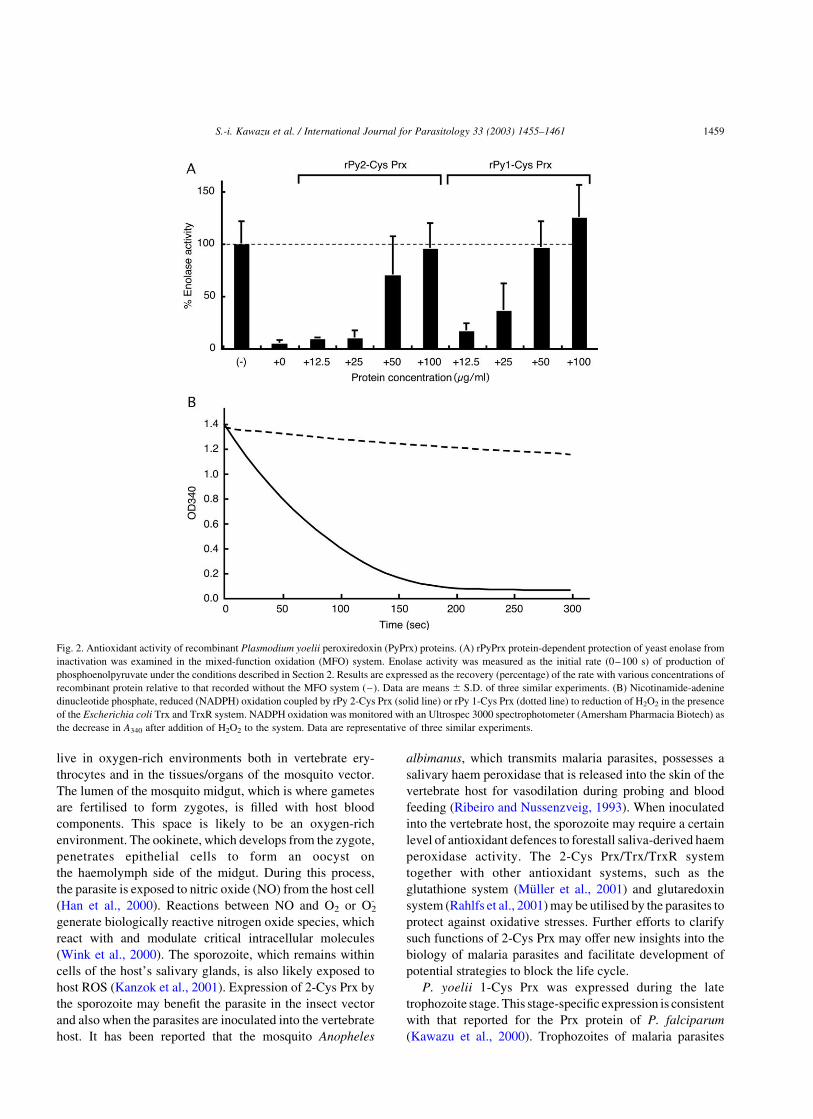
generates hydroxyl radicals to damage an enzyme (enolase).
In the absence of rPyPrx protein, the hydroxyl radicals
inactivate enolase (Fig. 2A). In the present study, addition of
the recombinant proteins to the system prevented an
inactivation of enolase in a concentration-dependent
manner, indicating that PyPrx proteins have antioxidant
activities (Fig. 2A). The Trx-linked peroxidase activity of
rPyPrx was tested by monitoring oxidation of NADPH in
the presence of the E. coli Trx/TrxR system (Fig. 2B). After
addition of H2O2, the assay with rPy 2-Cys Prx showed a
marked decrease in A340, which was attributed to oxidation
of NADPH. This result suggests that the rPy 2-Cys Prx is a
Trx peroxidase. In contrast, NADPH oxidation in the assay
with rPy 1-Cys Prx was negligible.
3.3. Expression of the P. yoelii Prx proteins
during the blood and insect stages
Immunofluorescence microscopy of P. yoelii blood-stage
parasites revealed that 2-Cys Prx was expressed in
the parasite cytoplasm during the early to late trophozoite,
schizont, and gametocyte stages (Fig. 3C,D). There was no
specific fluorescence in the nuclei or plasma membranes of
parasites. Simultaneous application of the mouse
monoclonal antibody to Pys25 (Tsuboi et al., 1997), a
molecule expressed in the cytoplasm of P. yoelii gameto-
cytes, with rabbit anti-Prx serum differentiated gametocytes
from trophozoites and schizonts on merged images of Alexa
Fluor 546 (red) and FITC (green) staining (Fig. 3D). In
contrast, substantial 1-Cys Prx staining was detected only in
a small number of trophozoites in the cytoplasm (Fig. 3G,
H). Expression of 1-Cys Prx in schizonts was very weak
(Fig. 3H) in comparison with that in parasites of the same
stage stained with anti-2-Cys Prx serum (Fig. 3D). 1-Cys
Prx was not detected in gametocytes (Fig. 3H).
Immunostaining of insect-stage parasites with anti-serum
against 2-Cys Prx revealed that 2-Cys Prx was expressed
throughout the cytoplasm of ookinetes and sporozoites
(Fig. 3I–L). 2-Cys Prx was also detected in the cytoplasm of
zygotes (data not shown). In contrast, 1-Cys Prx was not
detected during the zygote, ookinete, and sporozoite stages
(data not shown). Table 1 summarises the patterns of
expression of these Prx proteins during each developmental
stage of P. yoelii as determined by the immunofluorescent
antibody staining. Immunostaining with the pre-immune
sera detected no fluorescence for Prx proteins in blood-stage
(Fig. 3A,B,E,F) or insect-stage parasites (data not shown).
3.4. Expression 1-Cys and 2-Cys Prx mRNAs in blood-stage
P. yoelii
The lower immunofluorescent staining of 1-Cys Prx
observed in the parasites during the blood and insect stages,
except for trophozoites, might be due to a lower affinity of
the anti-serum used for 1-Cys Prx protein. To eliminate this
possibility, levels of 1-Cys Prx and 2-Cys Prx mRNAs in
blood-stage P. yoelii parasites were compared with the
quantitative real time RT–PCR method. We verified that
the level of 1-Cys Prx mRNA was lower than that of 2-Cys
Prx; the ratio of the absolute quantities of these two mRNA
species was 1:40 (Table 2).
4. Discussion
Here, we have described molecular cloning, biochemical
characterisation, and expression patterns of the 2-Cys and
1-Cys Prx proteins of the rodent malaria parasite P. yoelii.
Constitutive expression of the 2-Cys Prx protein in the
erythrocytic and insect stages suggests the importance of the
protein in the parasite antioxidant defence. It has been
suggested that P. falciparum has a functional 2-Cys
Prx/Trx/Trx reductase (TrxR) cycle that is involved in
detoxification of intracellular ROS (reviewed in Rahlfs et al.,
2002). Although there is presently no evidence for an active
Trx cycle in P. yoelii, the expression of 2-Cys Prx during all
the erythrocytic stages strongly suggests that the parasite
uses this protein as an antioxidant enzyme to cope with
enhanced oxidative stress in parasite-infected erythrocytes
(Becker et al., 1994; Postma et al., 1996). Malaria parasites
Fig. 1. Western blot analyses of recombinant Plasmodium yoelii
peroxiredoxin (rPyPrx) proteins. (A) Purified rPyPrx proteins were
subjected to 12.5% SDS–PAGE under reducing conditions. Lane 1, rPy
2-Cys Prx protein (500 ng); lane 2, rPy 1-Cys Prx protein (500 ng). The
proteins were transferred to Immobilon membranes and reacted with rabbit
anti-rPf 2-Cys Prx serum (2-Cys Prx) or anti-rPf 1-Cys Prx serum (1-Cys
Prx). (B) Extracts prepared from blood-stage parasites were subjected to
12.5% SDS–PAGE under reducing conditions. Thirty-eight micrograms of
extract was separated by electrophoresis (lane 1). Proteins were transferred
to Immobilon membranes and reacted with rabbit anti-rPf 2-Cys Prx IgG
(lane 2) or anti-rPf 1-Cys Prx IgG (lane 3). SDS–PAGE gels were stained
with Coomassie Brilliant Blue. Molecular weight markers in kDa are
indicated on the left.
S.-i. Kawazu et al. / International Journal for Parasitology 33 (2003) 1455–14611458
live in oxygen-rich environments both in vertebrate ery-
throcytes and in the tissues/organs of the mosquito vector.
The lumen of the mosquito midgut, which is where gametes
are fertilised to form zygotes, is filled with host blood
components. This space is likely to be an oxygen-rich
environment. The ookinete, which develops from the zygote,
penetrates epithelial cells to form an oocyst on
the haemolymph side of the midgut. During this process,
the parasite is exposed to nitric oxide (NO) from the host cell
(Han et al., 2000). Reactions between NO and O2 or O2-
generate biologically reactive nitrogen oxide species, which
react with and modulate critical intracellular molecules
(Wink et al., 2000). The sporozoite, which remains within
cells of the host’s salivary glands, is also likely exposed to
host ROS (Kanzok et al., 2001). Expression of 2-Cys Prx by
the sporozoite may benefit the parasite in the insect vector
and also when the parasites are inoculated into the vertebrate
host. It has been reported that the mosquito Anopheles
albimanus, which transmits malaria parasites, possesses a
salivary haem peroxidase that is released into the skin of the
vertebrate host for vasodilation during probing and blood
feeding (Ribeiro and Nussenzveig, 1993). When inoculated
into the vertebrate host, the sporozoite may require a certain
level of antioxidant defences to forestall saliva-derived haem
peroxidase activity. The 2-Cys Prx/Trx/TrxR system
together with other antioxidant systems, such as the
glutathione system (Muller et al., 2001) and glutaredoxin
system (Rahlfs et al., 2001) may be utilised by the parasites to
protect against oxidative stresses. Further efforts to clarify
such functions of 2-Cys Prx may offer new insights into the
biology of malaria parasites and facilitate development of
potential strategies to block the life cycle.
P. yoelii 1-Cys Prx was expressed during the late
trophozoite stage. This stage-specific expression is consistent
with that reported for the Prx protein of P. falciparum
(Kawazu et al., 2000). Trophozoites of malaria parasites
Fig. 2. Antioxidant activity of recombinant Plasmodium yoelii peroxiredoxin (PyPrx) proteins. (A) rPyPrx protein-dependent protection of yeast enolase from
inactivation was examined in the mixed-function oxidation (MFO) system. Enolase activity was measured as the initial rate (0–100 s) of production of
phosphoenolpyruvate under the conditions described in Section 2. Results are expressed as the recovery (percentage) of the rate with various concentrations of
recombinant protein relative to that recorded without the MFO system (–). Data are means ^ S.D. of three similar experiments. (B) Nicotinamide-adenine
dinucleotide phosphate, reduced (NADPH) oxidation coupled by rPy 2-Cys Prx (solid line) or rPy 1-Cys Prx (dotted line) to reduction of H2O2 in the presence
of the Escherichia coli Trx and TrxR system. NADPH oxidation was monitored with an Ultrospec 3000 spectrophotometer (Amersham Pharmacia Biotech) as
the decrease in A340 after addition of H2O2 to the system. Data are representative of three similar experiments.
S.-i. Kawazu et al. / International Journal for Parasitology 33 (2003) 1455–1461 1459
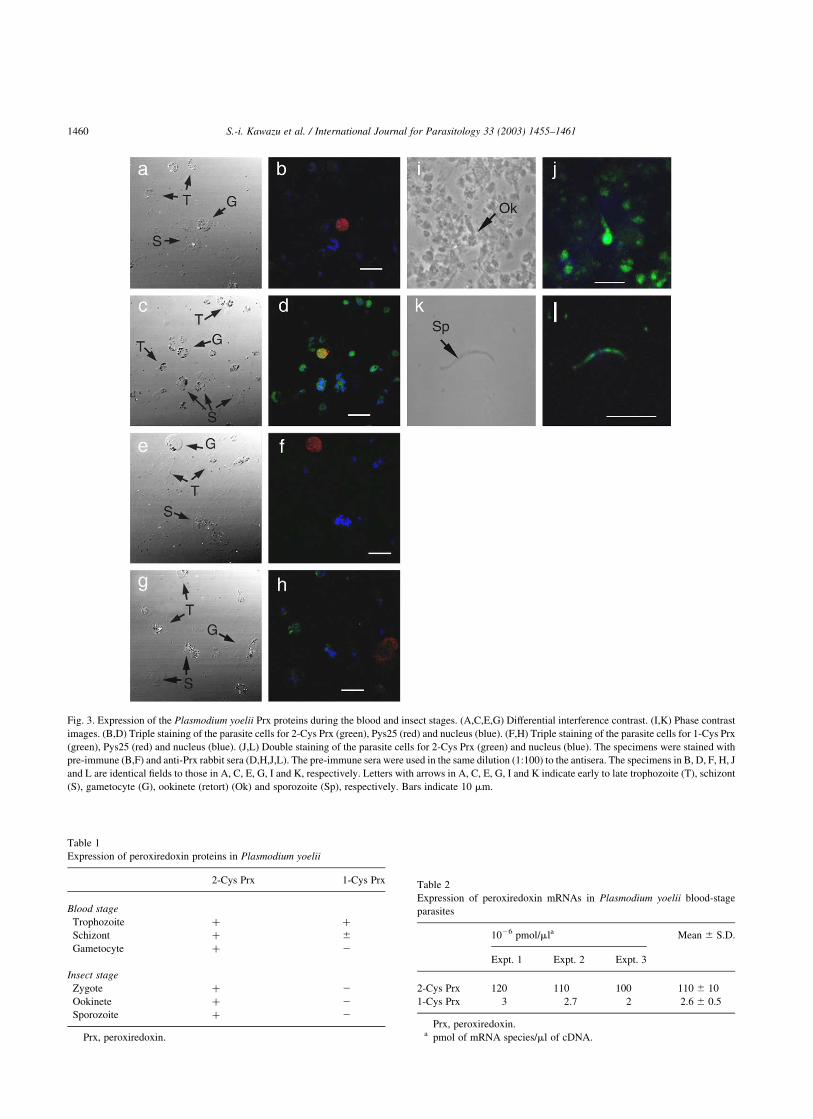
Fig. 3. Expression of the Plasmodium yoelii Prx proteins during the blood and insect stages. (A,C,E,G) Differential interference contrast. (I,K) Phase contrast
images. (B,D) Triple staining of the parasite cells for 2-Cys Prx (green), Pys25 (red) and nucleus (blue). (F,H) Triple staining of the parasite cells for 1-Cys Prx
(green), Pys25 (red) and nucleus (blue). (J,L) Double staining of the parasite cells for 2-Cys Prx (green) and nucleus (blue). The specimens were stained with
pre-immune (B,F) and anti-Prx rabbit sera (D,H,J,L). The pre-immune sera were used in the same dilution (1:100) to the antisera. The specimens in B, D, F, H, J
and L are identical fields to those in A, C, E, G, I and K, respectively. Letters with arrows in A, C, E, G, I and K indicate early to late trophozoite (T), schizont
(S), gametocyte (G), ookinete (retort) (Ok) and sporozoite (Sp), respectively. Bars indicate 10 mm.
Table 1
Expression of peroxiredoxin proteins in Plasmodium yoelii
2-Cys Prx 1-Cys Prx
Blood stage
Trophozoite þ þ
Schizont þ ^
Gametocyte þ 2
Insect stage
Zygote þ 2
Ookinete þ 2
Sporozoite þ 2
Prx, peroxiredoxin.
Table 2
Expression of peroxiredoxin mRNAs in Plasmodium yoelii blood-stage
parasites
1026 pmol/mla Mean ^ S.D.
Expt. 1 Expt. 2 Expt. 3
2-Cys Prx 120 110 100 110 ^ 10
1-Cys Prx 3 2.7 2 2.6 ^ 0.5
Prx, peroxiredoxin.a pmol of mRNA species/ml of cDNA.
S.-i. Kawazu et al. / International Journal for Parasitology 33 (2003) 1455–14611460

digest the host haemoglobin to release amino acids (Olliaro
and Goldberg, 1995). This process produces large quantities
of prooxidative haem in the food vacuole and results in
generation of ROS in the parasite cytoplasm (Ginsburg et al.,
1999). Thus malaria parasites must produce an effective
antioxidant to cope with such oxidative burdens. Malaria
parasites are known to be highly susceptible to the toxic effect
of the auto-producible haem, and this is the basis of the anti-
malarial action of chloroquine (Ginsburg et al., 1999). Further
studies to clarify the roles of Prx in the malaria parasite may
provide information that allowsutilisation of thisproteinas an
alternative target for anti-malaria chemotherapies.
In summary, the results obtained in this study suggest
that malaria parasites express and utilise 2-Cys and 1-Cys
Prx proteins during both the erythrocytic and insect stages to
protect against oxidative stresses.
Acknowledgements
This work was supported by a Grant-in-Aid for Scientific
Research (C) (13670260 to S.K. 15590378 to T.N., and
14570215 to T.T.) from the Japan Society for the Promotion
of Science (JSPS), Grants-in-Aid for Scientific Research
(14370084 to M.T.) and Scientific Research on Priority
Areas (C) (14021137, 15019127 to S.K., 11147230,
14021134, 15019120 to T.N., and 14021082, 15019072 to
T.T.) from the Ministry of Education, Culture, Sports,
Science and Technology (MEXT) of Japan, and a grant for
Precursory Research for Embryonic Science and Technol-
ogy, Japan Science and Technology Corporation (to T.N.)
and a Grant for International Health Cooperation Research
(15C-5) from the Ministry of Health, Labour and Welfare,
Japan. Sequence data for P. yoelii chromosomes were
obtained from the Plasmodium Genome Consortium
PlasmoDB (PlasomDB; http://PlasmoDB.org).
References
Becker, K., Gui, M., Traxler, A., Kirsten, C., Schirmer, R.H., 1994. Redox
processes in malaria and other parasitic diseases. Determination of
intracellular glutathione. Histochemistry 102, 389–395.
Chae, H.Z., Chung, S.J., Rhee, S.G., 1994. Thioredoxin-dependent
peroxide reductase from yeast. J. Biol. Chem. 269, 27670–27678.
Chae, H.Z., Kang, S.W., Rhee, S.G., 1999. Isoforms of mammalian
peroxiredoxin that reduce peroxides in presence of thioredoxin.
Methods Enzymol. 300, 219–226.
Fisher, A.B., Dodia, C., Manevich, Y., Chen, J.W., Feinstein, S.I., 1999.
Phospholipid hydroperoxides are substrates for non-selenium gluta-
thione peroxidase. J. Biol. Chem. 274, 21326–21334.
Flohe, L., Hecht, H.J., Steinert, P., 1999. Glutathione and trypanothione in
parasitic hydroperoxide metabolism. Free Radic. Biol. Med. 27,
966–984.
Ginsburg, H., Ward, S.A., Bray, P.G., 1999. An integrated model of
chloroquine action. Parasitol. Today 15, 357–360.
Han, Y.S., Thompson, J., Kafatos, F.C., Barillas-Mury, C., 2000. Molecular
interactions between Anopheles stephensi midgut cells and Plasmodium
berghei: the time bomb theory of ookinete invasion of mosquitoes.
EMBO J. 19, 6030–6040.
Kang, S.W., Baines, I.C., Rhee, S.G., 1998. Characterization of a
mammalian peroxiredoxin that contains one conserved cysteine.
J. Biol. Chem. 273, 6303–6311.
Kanzok, S.M., Fechner, A., Bauer, H., Ulschmid, J.K., Muller, H.M.,
Botella-Munoz, J., Schneuwly, S., Schirmer, R.H., Becker, K., 2001.
Substitution of the thioredoxin system for glutathione reductase in
Drosophila melanogaster. Science 291, 643–646.
Kawazu, S.I., Tsuji, N., Hatabu, T., Kawai, S., Matsumoto, Y., Kano, S.,
2000. Molecular cloning and characterization of a peroxiredoxin from
the human malaria parasite Plasmodium falciparum. Mol. Biochem.
Parasitol. 109, 165–169.
Kawazu, S.I., Komaki, K., Tsuji, N., Kawai, S., Ikenoue, N., Hatabu, T.,
Ishikawa, H., Matsumoto, Y., Himeno, K., Kano, S., 2001. Molecular
characterization of a 2-Cys peroxiredoxin from the human malaria
parasite Plasmodium falciparum. Mol. Biochem. Parasitol. 116,
73–79.
Kim, K., Kim, I.H., Lee, K.Y., Rhee, S.G., Stadtman, E.R., 1998. The
isolation and purification of a specific ‘protector’ protein which inhibits
enzyme inactivation by a thiol/Fe(III)/O2 mixed-function oxidation
system. J. Biol. Chem. 263, 4704–4711.
Kimura, M., Tanabe, K., Krishna, S., Tsuboi, T., Saito-Ito, A., Otani, S.,
Ogura, H., 1999. Gametocyte-dominant expression of a novel P-type
ATPase in Plasmodium yoelii. Mol. Biochem. Parasitol. 104, 331–336.
Krauth-Siegel, R.L., Coombs, G.H., 1999. Enzymes of parasite thiol
metabolism as drug targets. Parasitol. Today 15, 404–409.
Krnajski, Z., Walter, R.D., Muller, S., 2001. Isolation and functional
analysis of two thioredoxin peroxidases (peroxiredoxins) from
Plasmodium falciparum. Mol. Biochem. Parasitol. 113, 303–308.
Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly
of the head of bacteriophage T4. Nature 227, 680–685.
Miller, T.J., Stone, H.O., 1978. The rapid isolation of ribonuclease-free
immunoglobulin G by protein A-sepharose affinity chromatography.
J. Immunol. Methods 24, 111–125.
Muller, S., Gilberger, T.W., Krnajski, Z., Luersen, K., Meierjohann, S.,
Walter, R.D., 2001. Thioredoxin and glutathione system of malaria
parasite Plasmodium falciparum. Protoplasma 217, 43–49.
Olliaro, P.L., Goldberg, D.E., 1995. The Plasmodium digestive vacuole:
Metabolic headquarters and choice drug target. Parasitol. Today 11,
294–297.
Park, S.G., Cha, M.K., Jeong, W., Kim, I.H., 2000. Distinct physiological
functions of thiol peroxidase isoenzymes in Saccharomyces cerevisiae.
J. Biol. Chem. 275, 5723–5732.
Postma, N.S., Mommers, E.C., Eling, W.M.C., Zuidema, J., 1996.
Oxidative stress in malaria; implications for prevention and therapy.
Pharm. World Sci. 18, 121–129.
Rahlfs, S., Becker, K., 2001. Thioredoxin peroxidases of the malarial
parasite Plasmodium falciparum. Eur. J. Biochem. 268, 1404–1409.
Rahlfs, S., Fischer, M., Becker, K., 2001. Plasmodium falciparum
possesses a classical glutaredoxin and second, glutaredoxin-like protein
with a PICOT homology domain. J. Biol. Chem. 276, 37133–37140.
Rahlfs, R., Schirmer, R.H., Becker, K., 2002. The thioredoxin system of
Plasmodium falciparum and other parasites. Cell. Mol. Life Sci. 59,
1024–1041.
Ribeiro, J.M.C., Nussenzveig, R.H., 1993. The salivary catechol oxidase/
peroxidase activities of the mosquito Anopheles albimanus. J. Exp.
Biol. 179, 273–287.
Tsuboi, T., Cao, Y.M., Hitsumoto, Y., Yanagi, T., Kanbara, H., Torii, M.,
1997. Two antigens on zygotes and ookinetes of Plasmodium yoelii and
Plasmodium berghei that are distinct targets of transmission-blocking
immunity. Infect. Immun. 65, 2260–2264.
Wink, D.A., Miranda, K.M., Espey, M.G., Mitchell, J.B., Grisham, M.B.,
Fukuto, J., Feelisch, M., 2000. The chemical biology of nitric oxide.
Balancing nitric oxide with oxidative and nitrosative stress. In: Mayer,
B., (Ed.), Nitric Oxide, Handbook of Experimental Pharmacology, Vol.
143. Springer, Berlin, pp. 7–29.
S.-i. Kawazu et al. / International Journal for Parasitology 33 (2003) 1455–1461 1461





























