Embed Size (px)
Citation preview

Expression profile of miRNAs in Populus cathayana L. and Salixmatsudana Koidz under salt stress
Jing Zhou • Mingying Liu • Jing Jiang •
Guirong Qiao • Sheng Lin • Haiying Li •
Lihua Xie • Renying Zhuo
Received: 31 January 2012 / Accepted: 6 June 2012 / Published online: 21 June 2012
� Springer Science+Business Media B.V. 2012
Abstract Soil salinization can lead to environmental and
ecological problems worldwide. Abiotic stressors, includ-
ing salinity, are suspected to regulate microRNA (miRNA)
expression. Plants exposed to such abiotic stressors express
specific miRNAs, which are genes encoding small non-
coding RNAs of 20–24 nucleotides. miRNAs are known to
exist widely in plant genomes, and are endogenous. A
previous study used miRNA microarray technology and
poly(A) polymerase-mediated qRT-PCR technology to
analyze the expression profile of miRNAs in two types of
plants, Populus cathayana L. (salt-sensitive plants) and
Salix matsudana Koidz (highly salinity-tolerant plants),
both belonging to the Salicaceae family. miRNA micro-
array hybridization revealed changes in expression of 161
miRNAs P. cathayana and 32 miRNAs in S. matsudana
under salt stress. Differences in expression indicate that the
same miRNA has different expression patterns in salt-
sensitive plants and salt-tolerant plants under salt stress.
These indicate that changes in expression of miRNAs
might function as a response to varying salt concentrations.
To examine this, we used qRT-PCR to select five miRNA
family target genes involved in plant responses to salt
stress. Upon saline treatment, the expressions of both ptc-
miR474c and ptc-miR398b in P. cathayana were down-
regulated, but were up-regulated in S. matsudana.
Expression of the miR396 family in both types of plants
was suppressed. Furthermore, we have analyzed the dif-
ferent expression patterns between P. cathayana and
S. matsudana. Findings of this study can be utilized in
future investigations of post-transcriptional gene regulation
in P. cathayana and S. matsudana under saline stress.
Keywords miRNA � Salt stress � Expression �Populus cathayana L. � Salix matsudana Koidz
Jing Zhou and Mingying Liu contributed equally to this study.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s11033-012-1719-4) contains supplementarymaterial, which is available to authorized users.
J. Zhou � M. Liu � J. Jiang � G. Qiao � S. Lin � H. Li � L. Xie �R. Zhuo (&)
State Key Laboratory of Tree Genetics and Breeding, Chinese
Academy of Forestry, Xiangshan Road, Beijing 100091, China
e-mail: [email protected]
J. Zhou
e-mail: [email protected]
M. Liu
e-mail: [email protected]
J. Jiang
e-mail: [email protected]
G. Qiao
e-mail: [email protected]
S. Lin
e-mail: [email protected]
H. Li
e-mail: [email protected]
L. Xie
e-mail: [email protected]
J. Zhou � M. Liu � J. Jiang � G. Qiao � H. Li � L. Xie � R. Zhuo
Key Lab of Tree Genomics, The Research Institute
of Subtropical of Forestry, Chinese Academy of Forestry,
Fuyang, Hangzhou 311400, Zhejiang, China
123
Mol Biol Rep (2012) 39:8645–8654
DOI 10.1007/s11033-012-1719-4

Background
Soil salinization presents a serious environmental problem
to ecological systems because it can inhibit plant growth,
which will also lead to reduction of agricultural produc-
tivity. Globally, approximately 20 % of all cultivated land
and nearly 50 % of irrigated areas are affected by salini-
zation issues, accounting for 10 million hectares of affected
land in more than 100 countries [43, 41].
Plants respond to salt stress through complex mechanisms
that allow them to increase or reduce salt tolerance by syn-
thesizing or suppressing the production of some proteins
[52]. Over the past decade, the cloning of salt-related genes
has allowed for as applications in osmotic adjustment, pho-
tosynthesis, and metabolism of calmodulin. Understanding
the molecular mechanisms of plants upon saline stress and
determining effective ways to increase salt resistance of
plants is important in solving the effects of soil salinization
[67]. Salt stress can induce gene expression to protect plant
cells from injury, and these expressed genes have been
classified into four groups. The first group includes genes
that regulate signal transduction, and includes genes
encoding transcription factors (TFs) [64, 2, 51]. The second
group includes genes that encode enzymes, such as laccase
[63, 46], in response to plant oxidation stress [61, 36, 14].
The third category includes genes that synthesize osmopro-
tectants [62, 40, 38, 49], while the fourth group includes
genes that encode important functional proteins related to
water stress [68].
Populus cathayana is one of the most important decid-
uous Populus trees in China, which are characterized by
rapid growth and regeneration [60]. A previous study used
P. cathayana as the salt-sensitive plant and Salix matsu-
dana as the salt-resistant plant due to its strong resistance
to salt stress [65, 66]. Both plants belong to the Salicaceae
family but have different sensitivities to salt stress [11].
They are closely related phyllogenetically, as evidenced by
the presence of the same whole-genome duplication event
in 1,825 Populus and Salix orthologous genes derived from
Salix EST [60]. Furthermore, both had poplar microsatel-
lites that can successfully amplify willow DNA [17]. Yang
et al. [13] found that P. cathayana withered after exposure
to 100 mmol L-1 NaCl for 72 h, while S. matsudana grew
normally.
MicroRNAs (miRNAs) play important roles in plant
morphogenesis, development, and adaptations to changing
environments. miRNAs are small RNAs usually consisting
of 20–24 nucleotides, endogenous, and non-coding [1, 4, 7,
69–71, 56, 58]. In plants, miRNAs act as negative regu-
lators, mainly inhibiting gene expression at the post-
transcriptional level. In this pathway, miRNA combines
with an ARGONAUTE protein to create the miRNA
silencing complex (RISC) [47, 4, 15, 16, 57]. Using its
complementary base pairing property, miRNA activates
RISC, enabling it to target the corresponding mRNA. This
pathway can mediate irreversible cleavage or translational
repression of target transcripts [28, 9].
Recent studies have shown that miRNAs play a significant
and regular role in organ development and patterns of roots,
stems, and leaves [30, 69, 70]. Although several plant miR-
NAs have been documented to regulate programmed devel-
opment that is controlled by transcription factors, it has also
been proven recently that these miRNAs are involved in
additional physiological processes, including responses to
cold [53, 31, 73], salt [53, 6, 31, 12], heat [31], dehydration
[53, 42], oxidative stress [54], and mechanical stress [31].
Sunkar and Zhu [53] constructed an miRNA library of Ara-
bidopsis thaliana to investigate stress caused by drought,
salinity, low temperatures, and abscisic acid. Northern blot
analysis showed that drought, low temperatures, and salinity
affect the expressions of miR319c, miR393, miR395,
miR397b, and miR402. MiR398 and its target genes were
differentially expressed in response to high salt levels and
other types of biotic and abiotic stressors [21]. Ptc-miR1446a-
e, ptcmiR1447, and ptc-miR1450 have been demonstrated to
be either induced or suppressed by salt stress [31]. Members of
the miR169 family transiently inhibit the NF-YA transcription
factor when they are induced by high salinity [72].
Targets of miRNA are involved in the regulation of salt
stress resistance genes. For instance, the pentatricopeptide
repeat (PPR) protein family provides a signaling link
between mitochondrial electron transport and regulation of
stress and hormonal responses in Arabidopsis thaliana
[25]. PPR is controlled by the miR477 family [30], which
is involved in preventing salt stress in rice [3, 35]. In
addition, high concentrations of NaCl could enhance lac-
case (LAC) transcription in tomato root [63]. LAC, which
is controlled by the miR397 family, might exert effects on
the roots of plants during the process of adaptation to salt
stress [63]. The space patterns of miRNA expression are
negatively correlated to those of LAC target genes in
A. thaliana [46]. The finger pattern, which is made by the
combination of zinc finger proteins and Zn2?, plays an
important role in gene expression and regulation, cell dif-
ferentiation, embryonic development, and improvement of
plant resistance to salinity stress. The miR398 and miR477
families have been shown to regulate the gene expression
of the zinc finger proteins [53, 30].
Microarray technology, which has proven to be a useful
tool in miRNA expression assays, has been applied in high-
throughput detection of gene expression [27, 34]. In this
study, we used miRNA microarrays and qRT-PCR tech-
nology to analyze changes in expression profile of miRNA
in P. cathayana (salt-sensitive) and S. matsudana (salt-
tolerant). qRT-PCR were performed to verify the miRNAs
and their targets expression.
8646 Mol Biol Rep (2012) 39:8645–8654
123

Methods
Plant materials, stress conditions, and RNA extraction
P. cathayana and S. matsudana plants were grown in
Murashige-Skoog (MS) [37] nutrient medium and water
cultures under normal conditions and then exposed to 25 �C
and long days (16 h light/8 h dark per day). For stress
treatments, 1.5 month-old P. cathayana and S. matsudana
plants were subjected to salt stress. The treatment involved
immersion of plant roots in a water culture containing
100 mmol L-1 NaCl for 72 h and the control 1.5 month-old
P. cathayana and S. matsudana plants cultivated in water for
72 h. All setups were performed in triplicate.
Plantlets were immediately transferred to liquid nitrogen
after salt treatment and were then ground to fine powder.
Total RNA was extracted using a total RNA purification kit
(NORGEN, USA) and analyzed using a 1.2 % agarose gel.
Small RNA was extracted from treated and untreated plant
samples using a small RNA purification kit (Ambion,
USA). Using NANOdrop, the concentration and purity of
total RNA and small RNA were quantified by absorbance
at 260, 280, and 230 nm.
Validation of miRNA microarrays
P. cathayana and S. matsudana miRNA microarrays were
designed based on plant miRNA sequences obtained from
miRBase and miRNA probes. Microarrays probes for
P. cathayana and S. matsudana were selected according to
Sanger miRBase Release 10.0 and 11.0, which have iden-
tified all plant miRNAs. A total of 630 P. cathayana and
714 S. matsudana miRNA probes were printed on a chip
(http://www.mirbase.org/index.shtml). Ten micrograms of
RNA were sent to L.C. Sciences for microRNA microarray
analysis using the dual-channel microarray and ParaFlo
microfluidics chips. Among the control probes, PUC2PM-
20B and PUC2MM-20B were the single-based match
detection probe of the 20-mer RNA positive control
sequence. Data adjustments included data filtering, a log2
transformation, and gene centering and normalization.
p Values of the t test were used to analyze differences
between control and salt-treated sample groups. miRNAs
with p values \0.01 were selected for further analysis.
Clustering was performed using Cluster 3.0 (created by
Michiel de Hoon, Seiya Imoto, and Satoru Miyano, Uni-
versity of Tokyo, Human Genome Center) and viewed in a
heatmap using Java TreeView 1.0.13 software.
Polyadenylation and reverse transcription
Small RNAs were polyadenylated using a poly(A) poly-
merase kit (Ambion, USA). NANOdrop revealed that the
concentration of small RNAs was about 1lg lL-1.
Approximately 50 lL polyadenylation reaction was estab-
lished using 10 lg of small RNAs and 5 U of poly(A) poly-
merase (Ambion, USA). The reaction was incubated at
37 �C for 1 h. After incubation, poly(A)-tailed RNA was
found at a ratio of 25:24:1 through phenol:chloroform:iso-
amyl alcohol extraction and at 24:1 through phenol: chlo-
roform extraction. RNA was recovered by ethanol
precipitation and was then eluted in 20 lL DEPC water.
A reverse transcription (RT) reaction was performed
using 2 lL of poly(A)-tailed small RNAs, 1 lL of RT
primer (50-GCGAGCACAGAATTAATACGACTCACTA
T AGG-d(T)18 V(A,G or C)N(A,G C or T)-30) with 1 U of
SuperScript II (Invitrogen, USA). About 2 lL tailed RNA
was incubated with 1 lL of RT primer at 65 �C for 5 min
to remove any RNA secondary structures. Reactions were
chilled on ice for at least 2 min and remaining reagents
(109 buffer, dNTP mix (10 mmol L-1 each), RNaseout,
MnCl2, and SuperScript II) were added as specified in the
SuperScript II protocol. The reaction was allowed to pro-
ceed for 60 min at 42 �C, after which the reverse trans-
criptase was inactivated by 15 min incubation at 70 �C.
Expression analysis of miRNA
MicroRNA expression was studied using real-time PCR
with a 2 9 QuantiTect SYBR green PCR kit (QIAGEN,
Holand). Total RNA extracted from P. cathayana and
S. matsudana was polyadenylated using poly(A) polymer-
ase. RNA was reverse-transcribed as described above.
Real-time quantitative PCR was performed using standard
protocols on an Applied Biosystem’s 7300HT Sequence
Detection System. Briefly, 1.6 lL of a 1/10 dilution of
cDNA in water was added to 10 lL of 2 9 QuantiTect
SYBR green PCR master mix, 0.64 lL of each primer, and
water to reach 20 lL. The miRNA primers are listed in
Table S3. The PCR protocol started with an initial incu-
bation step at 95 �C for 15 min to activate the HotStarTaq
DNA Polymerase. This was followed by 40 cycles of
amplification at 95 �C for 15 s, 58 �C for 30 s, and 68 �C
for 34 s. The thermal denaturation protocol was run at the
end of the PCR run to determine the number of products
present in the reaction [48]. All reactions were performed
in triplicate and did not include template controls for each
sample. P. cathayana 5.8S ribosomal RNA (rRNA) was
selected as the internal reference gene.
The cycle number at which the reaction crossed an
arbitrarily placed threshold (CT) was determined for each
gene, and the relative amount of each miRNA to 5.8S
rRNA was described using the equation 2T-DDC; where
DCT = (CT sample-CT reference gene) and DDCT = (CT sample
-CT reference gene) - (CT calibrator-CT reference gene) [29].
Mol Biol Rep (2012) 39:8645–8654 8647
123
Analysis of target gene expression by qRT-PCR
A total of 7 lL of total RNA was used for RT reaction with
SuperScript II using oligo dT primer (Invitrogen, USA).
The qRT-PCR was performed with an Applied Biosystems
step one instrument using the SYBR premix Ex TaqTM kit
(TaKaRa, Japan). All reactions were repeated in triplicate
and did not include template controls for each sample. As
internal control, expression levels of actin in P. cathayana
and S. matsudana were determined. Primers for the target
genes were designed using primer 5.0 software. The primer
sequences of the target genes and the internal control are
presented in Table S4. The relative quantity of gene
expression was detected using 2-DDCT method [29].
Results and discussion
Identification of 116 miRNAs from salt-stressed
P. cathayana and 32 miRNAs from salt-stressed
S. matsudana by miRNA microarray
MicroRNA microarrays were used to analyze the miRNA
expression profiles of P. cathayana and S. matsudana
exposed to 100 mmol L-1 NaCl for 72 h. Using Sanger
miRBase Release 10.0 and 11.0 (http://www.sanger.ac.uk/
Software/Rfam/miRna/), a total of 630 probes and 714
probes were designed for P. cathayana and S. matsudana
miRNAs, respectively. All probes were deposited in the
miRBase. Compared with the control (normal growth
conditions), significant regulation of miRNAs in response
to salinity stress was revealed. Array results showed that
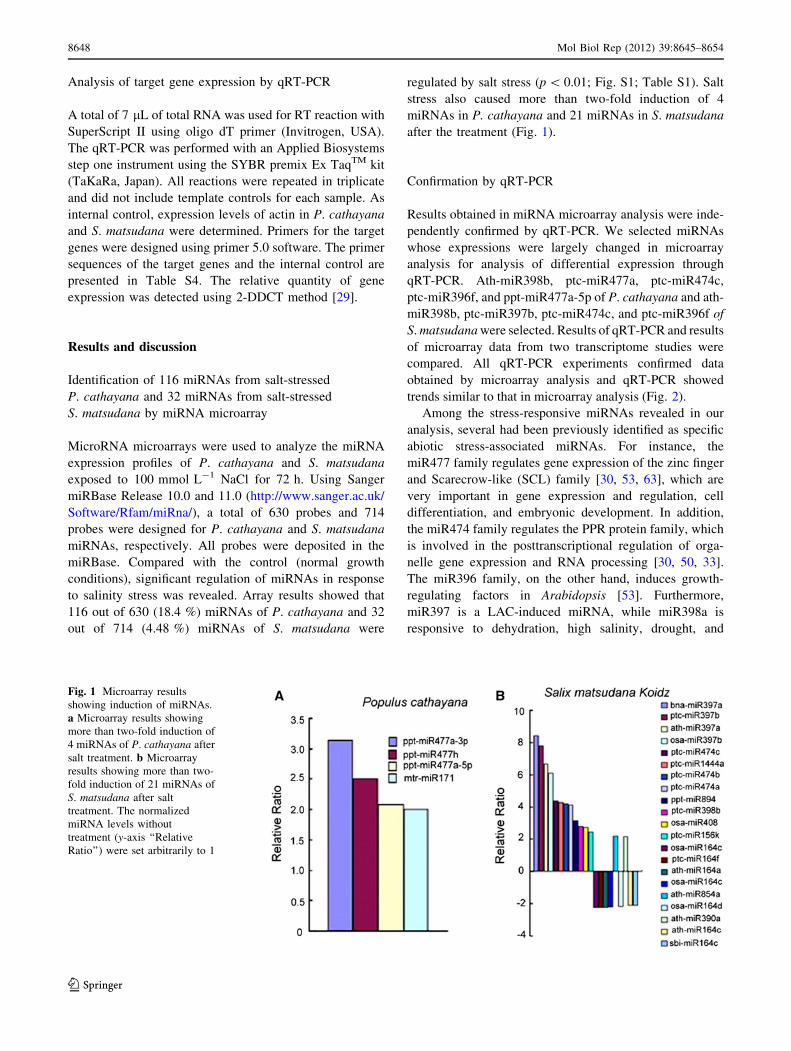
116 out of 630 (18.4 %) miRNAs of P. cathayana and 32
out of 714 (4.48 %) miRNAs of S. matsudana were
regulated by salt stress (p \ 0.01; Fig. S1; Table S1). Salt
stress also caused more than two-fold induction of 4
miRNAs in P. cathayana and 21 miRNAs in S. matsudana
after the treatment (Fig. 1).
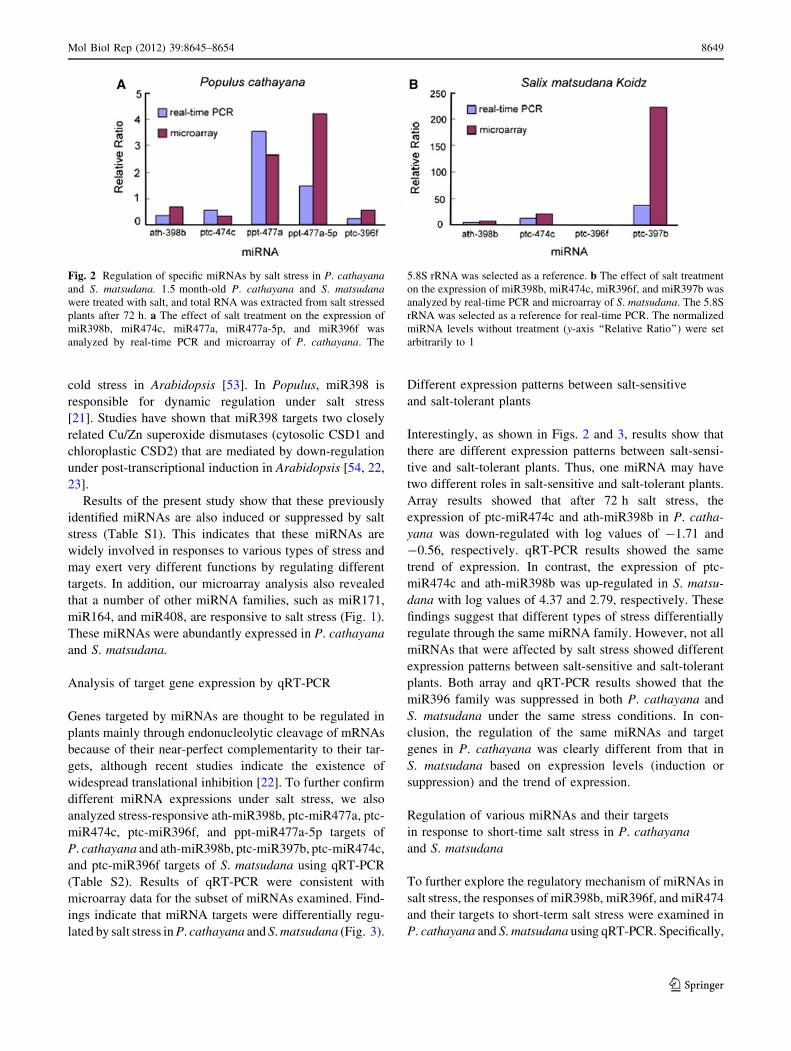
Confirmation by qRT-PCR
Results obtained in miRNA microarray analysis were inde-
pendently confirmed by qRT-PCR. We selected miRNAs
whose expressions were largely changed in microarray
analysis for analysis of differential expression through
qRT-PCR. Ath-miR398b, ptc-miR477a, ptc-miR474c,
ptc-miR396f, and ppt-miR477a-5p of P. cathayana and ath-
miR398b, ptc-miR397b, ptc-miR474c, and ptc-miR396f of
S. matsudana were selected. Results of qRT-PCR and results
of microarray data from two transcriptome studies were
compared. All qRT-PCR experiments confirmed data
obtained by microarray analysis and qRT-PCR showed
trends similar to that in microarray analysis (Fig. 2).
Among the stress-responsive miRNAs revealed in our
analysis, several had been previously identified as specific
abiotic stress-associated miRNAs. For instance, the
miR477 family regulates gene expression of the zinc finger
and Scarecrow-like (SCL) family [30, 53, 63], which are
very important in gene expression and regulation, cell
differentiation, and embryonic development. In addition,
the miR474 family regulates the PPR protein family, which
is involved in the posttranscriptional regulation of orga-
nelle gene expression and RNA processing [30, 50, 33].
The miR396 family, on the other hand, induces growth-
regulating factors in Arabidopsis [53]. Furthermore,
miR397 is a LAC-induced miRNA, while miR398a is
responsive to dehydration, high salinity, drought, and
Fig. 1 Microarray results
showing induction of miRNAs.
a Microarray results showing
more than two-fold induction of
4 miRNAs of P. cathayana after
salt treatment. b Microarray
results showing more than two-
fold induction of 21 miRNAs of
S. matsudana after salt
treatment. The normalized
miRNA levels without
treatment (y-axis ‘‘Relative
Ratio’’) were set arbitrarily to 1
8648 Mol Biol Rep (2012) 39:8645–8654
123
cold stress in Arabidopsis [53]. In Populus, miR398 is
responsible for dynamic regulation under salt stress
[21]. Studies have shown that miR398 targets two closely
related Cu/Zn superoxide dismutases (cytosolic CSD1 and
chloroplastic CSD2) that are mediated by down-regulation
under post-transcriptional induction in Arabidopsis [54, 22,
23].
Results of the present study show that these previously
identified miRNAs are also induced or suppressed by salt
stress (Table S1). This indicates that these miRNAs are
widely involved in responses to various types of stress and
may exert very different functions by regulating different
targets. In addition, our microarray analysis also revealed
that a number of other miRNA families, such as miR171,
miR164, and miR408, are responsive to salt stress (Fig. 1).
These miRNAs were abundantly expressed in P. cathayana
and S. matsudana.
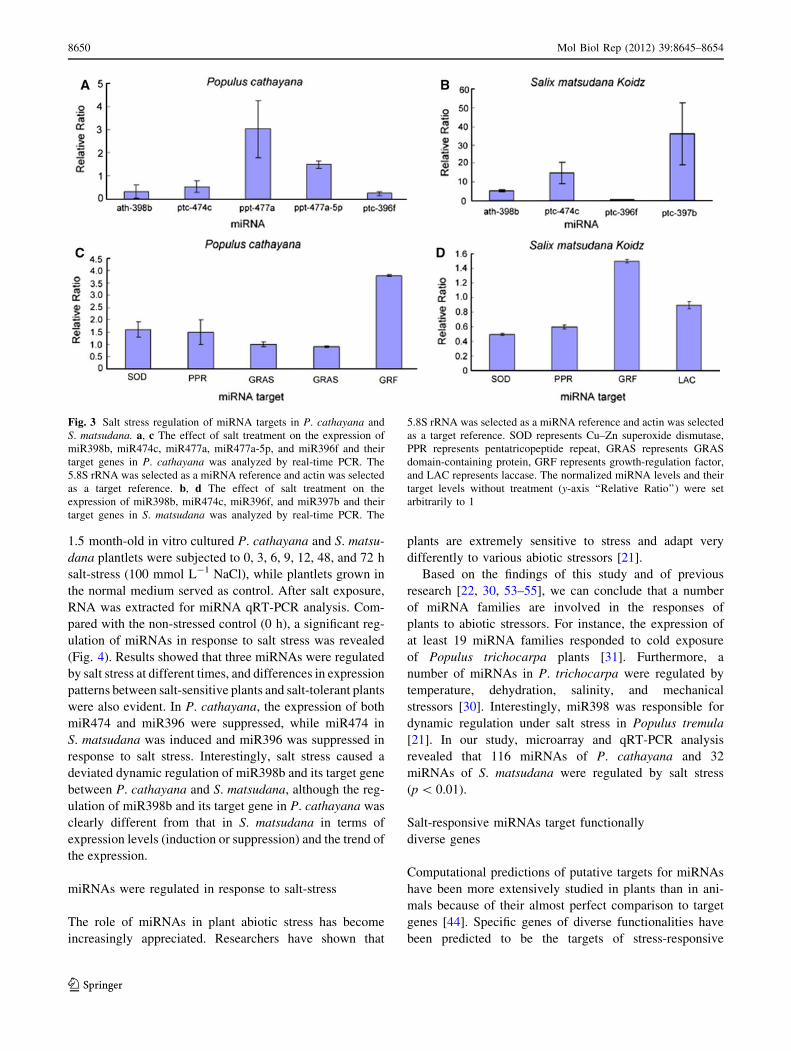
Analysis of target gene expression by qRT-PCR
Genes targeted by miRNAs are thought to be regulated in
plants mainly through endonucleolytic cleavage of mRNAs
because of their near-perfect complementarity to their tar-
gets, although recent studies indicate the existence of
widespread translational inhibition [22]. To further confirm
different miRNA expressions under salt stress, we also
analyzed stress-responsive ath-miR398b, ptc-miR477a, ptc-
miR474c, ptc-miR396f, and ppt-miR477a-5p targets of
P. cathayana and ath-miR398b, ptc-miR397b, ptc-miR474c,
and ptc-miR396f targets of S. matsudana using qRT-PCR
(Table S2). Results of qRT-PCR were consistent with
microarray data for the subset of miRNAs examined. Find-
ings indicate that miRNA targets were differentially regu-
lated by salt stress in P. cathayana and S. matsudana (Fig. 3).
Different expression patterns between salt-sensitive
and salt-tolerant plants
Interestingly, as shown in Figs. 2 and 3, results show that
there are different expression patterns between salt-sensi-
tive and salt-tolerant plants. Thus, one miRNA may have
two different roles in salt-sensitive and salt-tolerant plants.
Array results showed that after 72 h salt stress, the
expression of ptc-miR474c and ath-miR398b in P. catha-
yana was down-regulated with log values of -1.71 and
-0.56, respectively. qRT-PCR results showed the same
trend of expression. In contrast, the expression of ptc-
miR474c and ath-miR398b was up-regulated in S. matsu-
dana with log values of 4.37 and 2.79, respectively. These
findings suggest that different types of stress differentially
regulate through the same miRNA family. However, not all
miRNAs that were affected by salt stress showed different
expression patterns between salt-sensitive and salt-tolerant
plants. Both array and qRT-PCR results showed that the
miR396 family was suppressed in both P. cathayana and
S. matsudana under the same stress conditions. In con-
clusion, the regulation of the same miRNAs and target
genes in P. cathayana was clearly different from that in
S. matsudana based on expression levels (induction or
suppression) and the trend of expression.
Regulation of various miRNAs and their targets
in response to short-time salt stress in P. cathayana
and S. matsudana
To further explore the regulatory mechanism of miRNAs in
salt stress, the responses of miR398b, miR396f, and miR474
and their targets to short-term salt stress were examined in
P. cathayana and S. matsudana using qRT-PCR. Specifically,
Fig. 2 Regulation of specific miRNAs by salt stress in P. cathayanaand S. matsudana. 1.5 month-old P. cathayana and S. matsudanawere treated with salt, and total RNA was extracted from salt stressed
plants after 72 h. a The effect of salt treatment on the expression of
miR398b, miR474c, miR477a, miR477a-5p, and miR396f was
analyzed by real-time PCR and microarray of P. cathayana. The
5.8S rRNA was selected as a reference. b The effect of salt treatment
on the expression of miR398b, miR474c, miR396f, and miR397b was
analyzed by real-time PCR and microarray of S. matsudana. The 5.8S
rRNA was selected as a reference for real-time PCR. The normalized
miRNA levels without treatment (y-axis ‘‘Relative Ratio’’) were set
arbitrarily to 1
Mol Biol Rep (2012) 39:8645–8654 8649
123
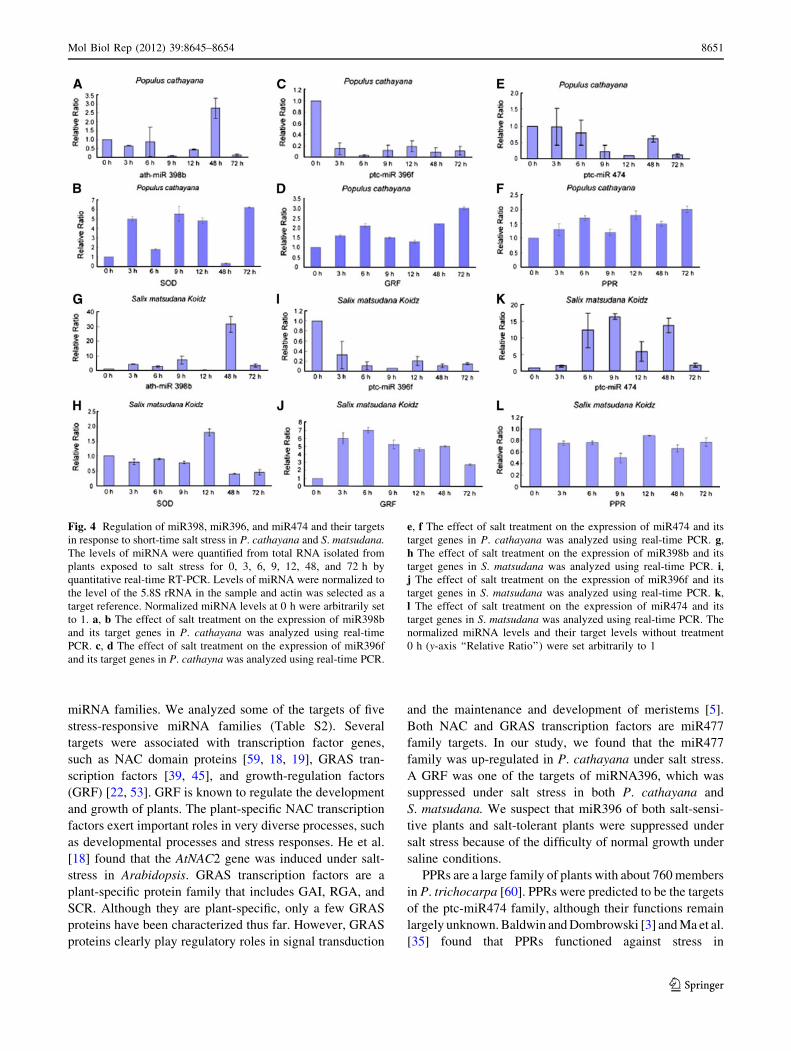
1.5 month-old in vitro cultured P. cathayana and S. matsu-
dana plantlets were subjected to 0, 3, 6, 9, 12, 48, and 72 h
salt-stress (100 mmol L-1 NaCl), while plantlets grown in
the normal medium served as control. After salt exposure,
RNA was extracted for miRNA qRT-PCR analysis. Com-
pared with the non-stressed control (0 h), a significant reg-
ulation of miRNAs in response to salt stress was revealed
(Fig. 4). Results showed that three miRNAs were regulated
by salt stress at different times, and differences in expression
patterns between salt-sensitive plants and salt-tolerant plants
were also evident. In P. cathayana, the expression of both
miR474 and miR396 were suppressed, while miR474 in
S. matsudana was induced and miR396 was suppressed in
response to salt stress. Interestingly, salt stress caused a
deviated dynamic regulation of miR398b and its target gene
between P. cathayana and S. matsudana, although the reg-
ulation of miR398b and its target gene in P. cathayana was
clearly different from that in S. matsudana in terms of
expression levels (induction or suppression) and the trend of
the expression.
miRNAs were regulated in response to salt-stress
The role of miRNAs in plant abiotic stress has become
increasingly appreciated. Researchers have shown that
plants are extremely sensitive to stress and adapt very
differently to various abiotic stressors [21].
Based on the findings of this study and of previous
research [22, 30, 53–55], we can conclude that a number
of miRNA families are involved in the responses of
plants to abiotic stressors. For instance, the expression of
at least 19 miRNA families responded to cold exposure
of Populus trichocarpa plants [31]. Furthermore, a
number of miRNAs in P. trichocarpa were regulated by
temperature, dehydration, salinity, and mechanical
stressors [30]. Interestingly, miR398 was responsible for
dynamic regulation under salt stress in Populus tremula
[21]. In our study, microarray and qRT-PCR analysis
revealed that 116 miRNAs of P. cathayana and 32
miRNAs of S. matsudana were regulated by salt stress
(p \ 0.01).
Salt-responsive miRNAs target functionally
diverse genes
Computational predictions of putative targets for miRNAs
have been more extensively studied in plants than in ani-
mals because of their almost perfect comparison to target
genes [44]. Specific genes of diverse functionalities have
been predicted to be the targets of stress-responsive
Fig. 3 Salt stress regulation of miRNA targets in P. cathayana and
S. matsudana. a, c The effect of salt treatment on the expression of
miR398b, miR474c, miR477a, miR477a-5p, and miR396f and their
target genes in P. cathayana was analyzed by real-time PCR. The
5.8S rRNA was selected as a miRNA reference and actin was selected
as a target reference. b, d The effect of salt treatment on the
expression of miR398b, miR474c, miR396f, and miR397b and their
target genes in S. matsudana was analyzed by real-time PCR. The
5.8S rRNA was selected as a miRNA reference and actin was selected
as a target reference. SOD represents Cu–Zn superoxide dismutase,
PPR represents pentatricopeptide repeat, GRAS represents GRAS
domain-containing protein, GRF represents growth-regulation factor,
and LAC represents laccase. The normalized miRNA levels and their
target levels without treatment (y-axis ‘‘Relative Ratio’’) were set
arbitrarily to 1
8650 Mol Biol Rep (2012) 39:8645–8654
123
miRNA families. We analyzed some of the targets of five
stress-responsive miRNA families (Table S2). Several
targets were associated with transcription factor genes,
such as NAC domain proteins [59, 18, 19], GRAS tran-
scription factors [39, 45], and growth-regulation factors
(GRF) [22, 53]. GRF is known to regulate the development
and growth of plants. The plant-specific NAC transcription
factors exert important roles in very diverse processes, such
as developmental processes and stress responses. He et al.
[18] found that the AtNAC2 gene was induced under salt-
stress in Arabidopsis. GRAS transcription factors are a
plant-specific protein family that includes GAI, RGA, and
SCR. Although they are plant-specific, only a few GRAS
proteins have been characterized thus far. However, GRAS
proteins clearly play regulatory roles in signal transduction
and the maintenance and development of meristems [5].
Both NAC and GRAS transcription factors are miR477
family targets. In our study, we found that the miR477
family was up-regulated in P. cathayana under salt stress.
A GRF was one of the targets of miRNA396, which was
suppressed under salt stress in both P. cathayana and
S. matsudana. We suspect that miR396 of both salt-sensi-
tive plants and salt-tolerant plants were suppressed under
salt stress because of the difficulty of normal growth under
saline conditions.
PPRs are a large family of plants with about 760 members
in P. trichocarpa [60]. PPRs were predicted to be the targets
of the ptc-miR474 family, although their functions remain
largely unknown. Baldwin and Dombrowski [3] and Ma et al.
[35] found that PPRs functioned against stress in
Fig. 4 Regulation of miR398, miR396, and miR474 and their targets
in response to short-time salt stress in P. cathayana and S. matsudana.The levels of miRNA were quantified from total RNA isolated from
plants exposed to salt stress for 0, 3, 6, 9, 12, 48, and 72 h by
quantitative real-time RT-PCR. Levels of miRNA were normalized to
the level of the 5.8S rRNA in the sample and actin was selected as a
target reference. Normalized miRNA levels at 0 h were arbitrarily set
to 1. a, b The effect of salt treatment on the expression of miR398b
and its target genes in P. cathayana was analyzed using real-time
PCR. c, d The effect of salt treatment on the expression of miR396f
and its target genes in P. cathayna was analyzed using real-time PCR.
e, f The effect of salt treatment on the expression of miR474 and its
target genes in P. cathayana was analyzed using real-time PCR. g,
h The effect of salt treatment on the expression of miR398b and its
target genes in S. matsudana was analyzed using real-time PCR. i,j The effect of salt treatment on the expression of miR396f and its
target genes in S. matsudana was analyzed using real-time PCR. k,
l The effect of salt treatment on the expression of miR474 and its
target genes in S. matsudana was analyzed using real-time PCR. The
normalized miRNA levels and their target levels without treatment
0 h (y-axis ‘‘Relative Ratio’’) were set arbitrarily to 1
Mol Biol Rep (2012) 39:8645–8654 8651
123

L. temulentum and rice. We found that the miR474 family
was down-regulated in P. cathayana but up-regulated in
S. matsudana, which suggests that there is a diversity of PPR
function in response to salt stress.
Laccases, a group of polyphenol oxidases, are associated
with lignification, thickening of cell walls in secondary cell
growth, ion absorption, and stress responses in trees [10].
In addition, LACs such as miRNA397 targets [22, 53] were
responsive to high-salinity, dehydration, drought, and cold
stress in Arabidopsis [53]. miR397 is highly and specifi-
cally expressed in plants. For instance, high concentrations
of NaCl can improve the level of LAC transcription in
tomato roots [63]. This finding suggests that miR397 may
be regulated only in the roots of tomatoes. Meanwhile,
some LAC genes are root-specific (for example AtLAC15)
or mostly expressed in roots and are involved in root
elongation and lignifications [26]. In additional, miR397 is
highly and specifically expressed in undifferentiated
embryogenic calli, but displayed very low expression lev-
els in differentiated calli and mature organs in rice [32].
Under salt stress, the miR397 family was up-regulated in
S. matsudana and their target LACs were down-regulated
in S. matsudana.
miR398 was the first miRNA recorded that linked
miRNA and stress tolerance. The expression of miR398 is
transcriptionally down-regulated by oxidative stressors.
miR398 is the most reported among miRNAs because its
targets are regulated in different expressions [22, 53, 31,
46, 20, 21]. For instance, miR398 was confirmed to cause
dynamic regulation under salt stress in P. tremula, while
miR398a was responsive to dehydration, high-salinity,
drought, and cold stress in Arabidopsis. Another miR398
target, copper/zinc superoxide dismutase, down-regulates
copper/zinc superoxide dismutase expression in response to
low Cu. In this study, we found that the miR398 family was
down-regulated in P. cathayana but up-regulated in
S. matsudana in response to 72-h salt stress.
In this experiment, microarray probes and synthesized
quantitative real-time primers were selected based on the
mature miRNA sequences in miRBase. However, it is
possible that some mature miRNA sequences in P. catha-
yana and S. matsudana are actually not identical. None—
identical miRNA sequences may lead to bias results when
miRNA expression patterns are analyzed using the miRNA
microarray and quantitative real-time methods. Neverthe-
less, the bias is not likely to be eliminated by any approach.
Different expression patterns between salt-sensitive
and salt-tolerant plants
Various plant species have varying miRNA expression
responses to salt stress. Specific miRNAs in the P. catha-
yana and S. matsudana had completely different responses
under the same conditions. For instance, ptc-miR474c and
ptc-miR398b were down-regulated in P. cathayana but
were conversely up-regulated in S. matsudana in response
to 72 h salt stress.
There are several possible explanations for the differ-
ences in expressions of miRNAs and their targets between
P. cathayana and S. matsudana. First, P. cathayana rep-
resents a salt-sensitive plant, while S. matsudana represents
a salt-tolerant plant and has a high level of stress resistance
over its long life cycle. The major physical and structural
differences between salt-sensitive plants and salt-tolerant
plants may cause different salt stress responses. Another
possible explanation is that the rate of salt-sensing, up-take,
and regulation could be different between the two species.
Additionally, these plants may undergo different levels of
cellular stress upon salt treatment, which also could explain
their different regulations of stress-responsive miRNAs.
Salt-tolerant plants have greater capacities than salt-sensi-
tive plants to compartmentalize Na? into vacuoles and
exclude Na? from the apoplast. Salt-tolerant plants have
lower levels of Cl- than salt-sensitive plants in all types of
leaf cells under stress, while salt-sensitive plants had higher
levels of Cl- in all cell compartments (chloroplast, cell
wall, and vacuole) than salt-tolerant plants [11]. Therefore,
we infer that miRNA and their targets may regulate sub-
cellular ion compartmentalization genes.
Jia et al. [21] found that miR398 had different responses to
ABA in Populus and Arabidopsis. Populus plants are a
perennial woody plant species and Arabidopsis is an annual
herbaceous plants with a short life cycle. The regulation
trend of miR398 was completely opposite in the two plants
under ABA treatment [21]. However, we also found that not
all salt stress-regulated miRNAs differ between salt-sensi-
tive plants and salt-tolerant plants. For instance, miR396 was
suppressed in both P. cathayana and S. matsudana under salt
stress. It is known that one of the targets of miR396 is the
GRF genes, which encode putative transcription factors
associated with cell expansion in leaf and other tissues in
A. thaliana and O. sativa [8, 24]. Thus, we can infer that it is
the impediment to normal growth under salt stress that
suppresses miR396 in both salt-sensitive plants and salt-
tolerant plants.
Conclusions
In this study, we have analyzed the different expression
patterns of miRNAs between P. cathayana and S. matsu-
dana. Results showed that some miRNAs and their
corresponding target genes were differently expressed in
salt-sensitive plants and salt-tolerant plants under salt stress.
These suggest that miRNAs have a role in plant response to
abiotic stress in different ecotypes, and that these effects are
8652 Mol Biol Rep (2012) 39:8645–8654
123

exerted through different regulatory machineries in order for
plants to adapt to a saline environment.
Acknowledgments We would like to thank Dr. Han-Jiang Fu of
the Academy of Military Medical Science and Dr. Zhangxun Wang
of the Tongji University School of Medicine for their technical
assistance. This work was supported by the National Natural Sci-
ence Fundation of China (30972340) and Natural Science Funda-
tion of Zhejiang Province (R3090070).
References
1. Ambros V (1989) Hierarchy of regulatory genes controls a larva
to adult developmental switch in elegans. Cell 57(1):49–57
2. Baker SS, Wilhelm KS, Thomashow MF (1994) The 50-region of
Arabidopsis thaliana has cis-acting elements that confer cold-,
drought- and ABA-regulated gene expression. Plant Mol Biol
24(5):701–713
3. Baldwin JC, Dombrowski JE (2006) Evaluation of Lolium tem-ulentum as a model grass species for the study of salinity stress by
PCR-based subtractive suppression hybridization analysis. Plant
Sci 171(4):459–469
4. Bartel DP (2004) MicroRNAs genomics biogenesis mechanism
and function. Cell 116(2):281–297
5. Bolle C (2004) The role of GRAS proteins in plant signal
transduction and development. Planta 218(5):683–692
6. Borsani O, Zhu J, Verslues PE, Sunkar R, Zhu JK (2005)
Endogenous siRNAs derived from a pair of natural cis-antisense
transcripts regulate salt tolerance in Arabidopsis. Cell 123(7):
1279–1291
7. Chen X (2005) MicroRNA biogenesis and function in plants.
FEBS Lett 579(26):5923–5931
8. Choi D, Kim JH, Kende H (2004) Whole genome analysis of the
OsGRF gene family encoding plant-specific putative transcription
activators in rice (Oryza sativa L). Plant Cell Physiol 45(7):
897–904
9. Chu CY, Rana TM (2006) Translation repression in human cells
by microRNA induced gene silencing requires RCK/p54. PLoS
Biol 4(7):210–219
10. Constabel CP, Yip L, Patton JJ, Christopher ME (2000) Poly-
phenol oxidase from hybrid poplar cloning and expression in
response to wounding and herbivory. Plant Physiol 124(1):
285–295
11. Dai SX, Chen sl, Fritz E, Olbrich A et al (2006) Ion compart-
mentation in leaf cells of Populus euphratica and P.tomentosaunder salt stress. J Beijing For Uni S2:1–5
12. Ding D, Zhang L, Wang H, Liu Z, Zhang Z, Zheng Y (2009)
Differential expression of miRNAs in response to salt stress in
maize roots. Annals Bot 103(1):29–38
13. Yang F, Xiao X, Zhang S, Korpelainen H, Li C (2009) Salt stress
responses in Populus cathayana Rehder. Plant Sci 176(5):
669–677
14. Gomez GA, Verhey SD, Holappa LD, Shen Q, Ho THD, Walker-
simmwas MK (1999) An ABA-induced protein kinase, PK2
ABA1, mediates ABA-suppressed gene expression in barley
aleurone layers. Proc Natl Acad Sci USA 96(4):1767–1772
15. Griffiths JS (2004) The microRNA registry. Nucleic Acids
32:109–111
16. Griffiths JS, Moxon S, Marshall M et al (2005) Annotating non-
coding RNAs in complete genomes. Nucleic Acids 33:121–124
17. Hanley S, Barker JHA, Ooijen JWV et al (2002) A genetic
linkage map of willow (Salix viminalis) based on AFLP and
microsatellite markers. Theor Appl Genet 105(6):1087–1096
18. He XJ, Mu RL, Cao WH, Zhang ZG, Zhang JS, Chen SY (2005)
AtNAC2, a transcription factors downstream of ethylene and
auxin signaling pathways, is involved in salt stress response and
lateral root development. Plant J 44(6):903–916
19. Hu HH, Dai MQ, Yao JL, Xiao BZ, Li XH, Zhang QF, Xiong LZ
(2006) Overexpressing a NAM, ATAF, and CUC (NAC) tran-
scription factor enhances drought resistance and salt tolerance in
rice. Proc Natl Acad Sci USA 103(35):12987–12992
20. Jagadeeswaran G, Saini A, Sunkar R (2009) Biotic and abiotic
stress down-regulate miR398 expression in Arabidopsis. Planta
229(4):1009–1023
21. Jia XY, Wang WX, Ren LG et al (2009) Differential and dynamic
regulation of miR398 in response to ABA and salt stress in
Populus tremula and Arabidopsis thaliana. Plant Mol Biol
71(1–2):51–59
22. Jones-Rhoades MW, Bartel DP (2004) Computational identifi-
cation of plant microRNA and their target, including a stress-
induced miRNA. Mol Cell 14(6):787–799
23. Jones-Rhoades MW, Bartel DP, Bartel B (2006) MicroRNAs and
their regulatory roles in plant. Annu Rev Plant Biol 57:19–53
24. Kim JH, Choi D, Kende H (2003) The AtGRF family of putative
transcription factors is involved in leaf and cotyledon growth in
Arabidopsis. Plant J 36(1):94–104
25. Laura Z, Gabor R, Andras S, Gyongyi S, Krisztina O, Zsuzsanna
D et al (2008) Arabidopsis PPR40 connects abiotic stress
responses to mitochondrial electron transport. Plant Physiol
146(4):1721–1737
26. Liang M, Davis E, Gardner D, Cai X, Wu Y (2006) Involvement
of AtLAC15 in lignin synthesis in seeds and in root elongation of
Arabidopsis. Planta 224(5):1185–1196
27. Liu HH, Tian X, Li YJ, Wu CA, Zheng CC (2008) Microarray-
based analysis of stress-regulated microRNAs in Arabidopsisthaliana. RNA 14(5):836–843
28. Liu J, Valencia-Sanchez MA, Hannon GJ, Parker R (2005)
MicroRNA dependent localization of targeted mRNAs to mam-
malian P-bodies. Nat Cell Biol 7(7):719–723
29. Livak KJ, Schmittgen TD (2001) Analysis of relative gene
expression data using real-time quantitative PCR and the 2T-44C
method. Methods 25(4):402–408
30. Lu S, Sun YH, Shi R, Clark C, Li L, Chiang VL (2005) Novel and
mechanical stress-responsive microRNAs in Populus Trichocar-pa that are absent from Arabidopsis. Plant Cell 17(8):2186–2203
31. Lu S, Sun YH, Chiang VL (2008) Stress-responsive microRNAs
in Populus. Plant J 55(1):131–151
32. Luo YC, Zhou H, Li Y, Chen GY et al (2006) Rice embryogenic
calli express a unique set of microRNAs, suggesting regulatory
roles of microRNAs in plant post-embryogenic development.
FEBS Lett 580(21):5111–5116
33. Lurin C, Andres C, Aubour S, Bellaouiet M et al (2004) Genome-
wide analysis of Arabidopsis pentatricopeptide repeat proteins
reveals their essential role in organelle biogenesis. Plant Cell
16(8):2089–2103
34. Lv DK, Bai X, Li Y, Ding XD, Ge Y, Cai H, Ji W, Wu N, Zhu
YM (2010) Profiling of cold-stress-responsive miRNAs in rice by
microarrays. Gene 459(1–2):39–47
35. Ma S, Gong Q, Bohnert HJ (2006) Dissecting salt stress path-
ways. J Exp Bot 57(5):1097–1107
36. Mizoguchi T, Ichimura K, Irie K (1998) Identification of possible
MAPK kinase cascade in Arabjdopsis theliana based on parewise
yeast two hybrid analysis and functional complementation test of
yeast mutants. FEBS Lett 437(1–2):56–60
Mol Biol Rep (2012) 39:8645–8654 8653
123

37. Murashige T, Skoog F (1962) A revised medium for rapid growth
and bioassays with tobacco tissue culture. Physiologia Plantarum
57(8):473–497
38. Novorry MJ (1984) Purification and properties of D-manoitol-
phosphate dehydrogenase and D-dlucitol-6-phosphate dehydro-
genase from Escherichia coli. J Bacteriol 159(3):986–990
39. Pysh LD et al (1999) The GRAS gene family in A rabidopsis:
sequence characterization and basic expression analysis of the
SCARECROW-LIKE genes. Plant J 18(1):111–119
40. Rathinasabapathi B, Burnet M, Russell BL et al (1997) Choline
monooxygenase, an unusual iron-sulfur enzyme catalyzing the
first step of glycine betaine synthesis in plats: prosthetic group
characterization and cDNA cloning. Proc Natl Acad Sci USA
94(7):3454–3458
41. Rengasamy P (2006) World salinization with emphasis on Aus-
tralia. J Exp Bot 57(5):1017–1023
42. Reyes JL, Chua NH (2007) ABA induction of miR159 controls
transcript levels of two MYB factors during Arabidopsis seed
germination. Plant J 49(4):592–606
43. Rhoades JD, Loveday J (1990) Salinity in irrigated agriculture.
AGRIS Rec 30:1089–1142
44. Rhoades M, Reinhart B, Lim L, Burge B et al (2002) Prediction
of plant microRNA targets. Cell 110(4):513–520
45. Sabatini S, Heidstra R, Wildwater M, Scheres B (2003)
SCARECROW is involved in positioning the stem cell niche in
the Arabidopsis root meristem. Genes Dev 17(3):354–358
46. Salah E, Abdel G, Marinus P (2008) MicroRNA-mediated sys-
temic down-regulation of copper protein expression in response
to low copper availability in Arabidopsis. J Biol Chem 283(6):
15932–15945
47. Schauers E, Jacobsen SE, Meinke DW, Ray A (2002) Dicer-like
1 blind men and elephants in Arabidopsis development. Trends
Plant Sci 7(11):487–491
48. Schmittgen TD, Zakrajsek BA, Mills AG, Gorn V et al (2000)
Quantitative reverse transcription–polymerase chain reaction to
study mRNA decay: comparison of endpoint and real-time
methods. Anal Biochem 285(2):194–204
49. Shen YG, Yan DQ, DU BX, Zhang JS et al (2002) Overexpres-
sion of proline transporter gene isolated from halophyte confers
salt tolerance in Arabidopsis. J Integr Plant Biol 44(8):956–962
50. Small ID, Peeters N (2000) The PPR motif: a TPR-related motif
prevalent in plant organellar proteins. Trends Biochem Sci
25(2):46–47
51. Stocking SJ, Gilmour MF, Thomashow MF (1997) Arabidopsisthaliana CBF1 encodes an AP2 domain-containing transcriptional
activator that binds to the C-repeat/DRE, a cis-acting DNA regu-
latory element that stimulates transcription in response to low
temperature and water deficit. Proc Natl Acad Sci USA 94(3):
1035–1040
52. Seki M, Ishida J, Narusaka M, Fujita M et al (2002) Monitoring
the expression profiles of 7000 Arabidopsis genes under drought,
cold and high-salinity stresses using a full-length cDNA micro-
array. Plant J 31(3):279–292
53. Sunkar R, Zhu JK (2004) Novel and stress-regulated microRNAs
and other small RNAs from Arabidopsis. Plant Cell 16(8):
2001–2019
54. Sunkar R, Kapoor A, Zhu JK (2006) Posttranscriptional induction
of two Cu/Zn superoxide dismutase genes in Arabidopsis is
mediated by downregulation of miR398 and important for oxi-
dative stress tolerance. Plant Cell 18(8):2051–2065
55. Sunkar R, Chinnusamy V, Zhu J, Zhu JK (2007) Small RNAs as
big players in plant abiotic stress responses and nutrient depri-
vation. Trends Plant Sci 12(7):301–309
56. Tang G, Reinhart BJ, Bartel DP et al (2003) A biochemical
framework for RNA silencing in plants. Genes Dev 17(1):49–63
57. Tang G (2005) siRNA and miRNA:an insight into RISCs. Trends
Biochem Sci 30(2):106–144
58. Tang G, Tang X, Mendu V, Jia X, Chen QJ, He L (2008) The art
of microRNA: various strategies leading to gene silencing via an
ancient pathway. Biochim Biophys Acta 1779(11):655–662
59. Tran LS, Nakashima K, Sakuma Y, Simpson SD et al (2004)
Isolation and functional analysis of Arabidopsis stress-inducible
NAC transcription factors that bind to a drought responsive Cis-
element in the early responsive to dehydrationstress promoter.
Plant Cell 16(9):2481–2498
60. Tuskan GA, Difazio S, Jansson S, Bohlmann J, Grigoriev I et al
(2006) The genome of black cottonwood, Populus trichocarpa(Torr. & Gray). Science 313(5793):1596–1604
61. Urao T, Katagiri T (1994) Two genes that encode Ca2 ? depen-
dent protein kinase are induced by drought and high salt stress in
Arabidopsis theliana. Mol Genet 244:331–340
62. Vernon DM, Bohnert HJ (1992) A novel metby transferase
induced by osmotic stress in the facultative Mesembryanthemumcrystallium. EMBO J 11(6):2077–2085
63. Wei JZ, Tirajoh A, Effendy J, Plant AL (2000) Characterization
of salt-induced changes in gene expression in tomato (Lycop-ersicon esculentum) roots and the role played by abscisic acid.
Plant Sci 159(1):135–148
64. Yamaguchi-Shinozaki K, Shinozaki K (1994) A novel cis-acting
element in an Arabidopsis gene is involved in responsiveness to
drought, low-temperature, or high-salt stress. Plant Cell 6(2):
251–264
65. Yang ZX, Wang SZ, Xu G et al (1995) Study on the variation of
wood property of juvenile P. cathayana among and within
provenances. Scientia Silvae Sinicae 8(4):437–441
66. Yu SH, Liu J, Fu DR et al (2003) Characteristics of Tacamachacagenes in the Western Sichuan plateau. J Zejiang For Coll 20(1):
29–33
67. Wu Y-Y, Qi-Jun C et al (2005) Salt-tolerant transgenic perennial
ryegrass (Lolium perenne L.) obtained by Agrobacterium tum-
efaciens-mediated transformation of the vacuolar Na?/H? anti-
porter gene. Plant Sci 169(1):65–73
68. Zeng HZ, Luo LJ (2003) A review on plant drought and salt
tolerance gene. J Plant Genet Resour 4(3):270–273
69. Zhang BH, Pan XP, Anderson TA (2006) Indentification of 188
conserved maize microRNAs and the its targets. FEBS Lett
580(5):3752–3762
70. Zhang B, Pan X, Cobb GP, Anderson TA (2006) Plant microRNA:
a small regulatory molecule with big impact. Dev Biol 289(1):
3–16
71. Zhang B, Wang Q, Pan X (2007) MicroRNAs and their regula-
tory roles in animals and plants. Cell Physiol 210(2):279–289
72. Zhao B, Ge LF, Liang RQ et al (2009) Members of miR-169
family are induced by high salinity and transiently inhibit the
NF-YA transcription factor. BMC Mol Biol 10(29):1–10
73. Zhou X, Wang G, Sutoh K, Zhu JK, Zhang W (2008) Identifi-
cation of cold- inducible microRNAs in plants by transcriptome
analysis. Biochim Biophys Acta 1779(11):780–788
8654 Mol Biol Rep (2012) 39:8645–8654
123





























