Embed Size (px)
Citation preview

Expression of Stat3 in germ cells of developing
and adult mouse ovaries and testes
Katherine Murphy, Luis Carvajal, Leo Medico, Melissa Pepling*
Department of Biology, Syracuse University, 130 College Place, Syracuse, NY 13244, USA
Received 16 November 2004; received in revised form 15 December 2004; accepted 16 December 2004
Abstract
The Signal transducers and activators of transcription (Stat) family of proteins plays diverse roles during differentiation in many tissues.
Stat3 is an essential mammalian gene, critical during embryonic development. In mammals, Stat3 is differentially distributed in the
cytoplasm of mature oocytes and in preimplantation embryos suggesting that Stat3 may be involved in determination of polarity. Here, we
report that Stat3 protein is expressed in the cytoplasm of oocytes from primordial, primary and secondary follicles in the adult ovary and in
developing acrosomes of round spermatids in the adult testis. Stat3 is also expressed in gonocytes, prospermatogonia, oogonia and oocytes of
embryonic and neonatal gonads.
q 2004 Elsevier B.V. All rights reserved.
Keywords: Mouse; Stat3; Gamete biology; Oocyte development; Ovary; Oogenesis; Jak-Stat signaling; Testes; Spermatogenesis; Spermiogenesis
1. Results and discussion
Shortly after their formation, female germ cells undergo
a series of incomplete cell divisions resulting in clusters
called cysts (Pepling and Spradling, 1998). The oogonia
stop dividing and enter meiosis at approximately 13.5 dpc
(days postcoitum), becoming oocytes, eventually arresting
in prophase I. After birth, mouse germ cell cysts break down
into individual oocytes that become surrounded by somatic
pre-granulosa cells to form primordial follicles (Pepling and
Spradling, 2001). During the process of cyst breakdown, a
subset of oocytes in each cyst dies with only a third of the
initial number of oocytes surviving (Pepling and Spradling,
2001). In the adult mouse, cohorts of primordial follicles are
periodically recruited to enter a 3-week growth phase that
ends in meiotic maturation and ovulation (Amleh and Dean,
2002). During this period, the oocyte grows and the
surrounding granulosa cells proliferate. Only some follicles
survive to maturity, the majority undergo atresia during
which the oocyte and surrounding granulosa cells die by
programmed cell death (Hsueh et al., 1994).
1567-133X/$ - see front matter q 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.modgep.2004.12.007
* Corresponding author. Tel.: C1 315 443 4541; fax: C1 315 443 2012.
E-mail address: [email protected] (M. Pepling).
In the male, primordial germ cells aggregate with Sertoli
cells to form seminiferous cords enclosed by peritubular
cells (Mackay, 2000; Orth et al., 2000). Male germ cells
divide until approximately 13.5 dpc when they exit the cell
cycle and remain quiescent as gonocytes until a few days
after birth (Monk and McLaren, 1981). The gonocytes are
initially in a central position within the cords but in the
neonate relocate to the basement membrane at the periphery
of the tubules, become prospermatogonia and eventually
develop into spermatogonial stem cells that produce
spermatozoa (Brinster, 2002; Mackay, 2000; Orth et al.,
2000). Differentiation of spermatogonia into spermatozoa
takes about 5 weeks and begins when the germ cells enter
meiosis and become spermatocytes. The result of meiosis is
the formation of haploid spermatids that undergo spermio-
genesis to form mature spermatozoa. During spermiogen-
esis, the acrosome, a secretory organelle important for
sperm-egg penetration, begins to form (Ramalho-Santos
et al., 2002).
The Signal transducers and activators of transcription
(Stat) family of proteins is important during differentiation
in many tissues (for reviews see (Ihle, 2001; Kisseleva et al.,
2002)). Membrane receptors are activated by ligand
binding, and in turn, activate Jak family protein kinases,
Gene Expression Patterns 5 (2005) 475–482
www.elsevier.com/locate/modgep
K. Murphy et al. / Gene Expression Patterns 5 (2005) 475–482476
which recruit and phosphorylate Stats. Phosphorylated Stat
proteins homodimerize and translocate to the nucleus where
they regulate transcription of target genes. This pathway can
be activated by a large number of cytokines and growth
factors in vertebrates (Hou et al., 2002). In mammals, there
are seven known Stat proteins (Kisseleva et al., 2002).
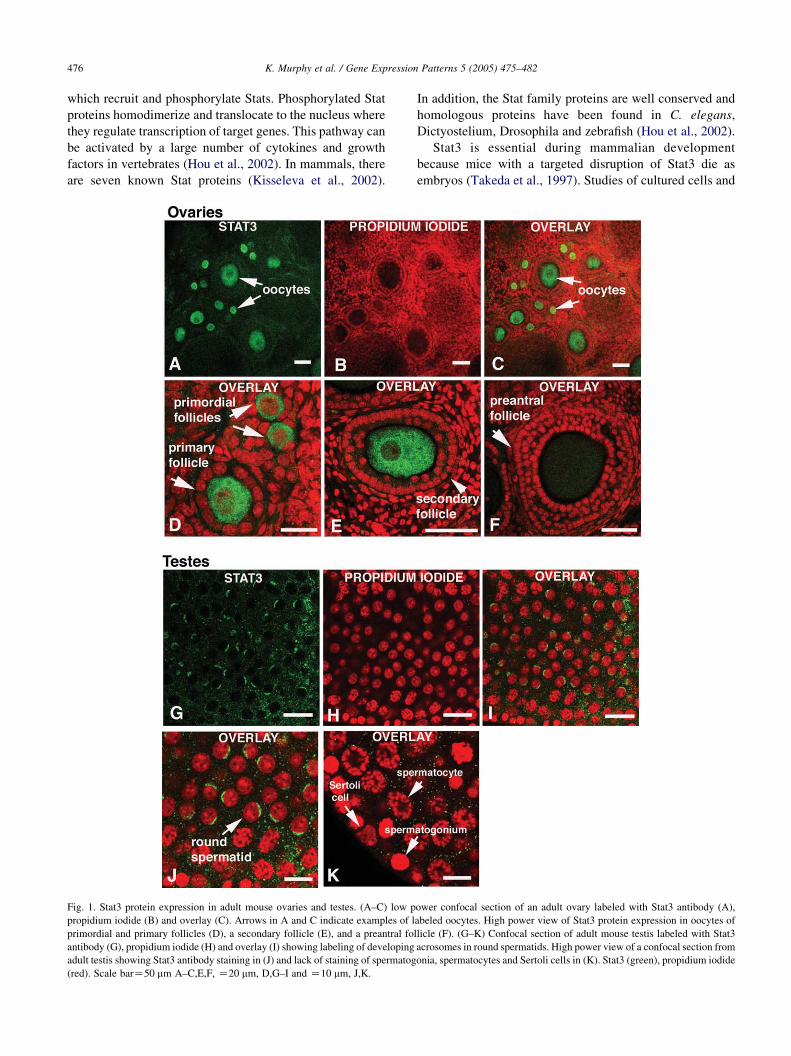
Fig. 1. Stat3 protein expression in adult mouse ovaries and testes. (A–C) low p
propidium iodide (B) and overlay (C). Arrows in A and C indicate examples of l
primordial and primary follicles (D), a secondary follicle (E), and a preantral fo
antibody (G), propidium iodide (H) and overlay (I) showing labeling of developing
adult testis showing Stat3 antibody staining in (J) and lack of staining of spermatog
(red). Scale barZ50 mm A–C,E,F, Z20 mm, D,G–I and Z10 mm, J,K.
In addition, the Stat family proteins are well conserved and
homologous proteins have been found in C. elegans,
Dictyostelium, Drosophila and zebrafish (Hou et al., 2002).
Stat3 is essential during mammalian development
because mice with a targeted disruption of Stat3 die as
embryos (Takeda et al., 1997). Studies of cultured cells and
ower confocal section of an adult ovary labeled with Stat3 antibody (A),
abeled oocytes. High power view of Stat3 protein expression in oocytes of
llicle (F). (G–K) Confocal section of adult mouse testis labeled with Stat3
acrosomes in round spermatids. High power view of a confocal section from
onia, spermatocytes and Sertoli cells in (K). Stat3 (green), propidium iodide

Fig. 2. Stat3 mRNA and protein expression in mouse ovaries and testes. (A)
RT-PCR analysis. RNA was isolated from 13.5 dpc, PND 1, PND 4 and
PND 42 ovaries and testes and Stat3 transcripts were monitored by RT-
PCR. GAPDH was also monitored as a loading control. (B) Western
blotting analysis. Mouse tissue extracts from PND 1, PND 4 and PND 42
ovaries and testes were probed with the Stat3 antibody. A band of
approximately 92 kDa corresponding to the molecular mass of Stat3a was
detected in PND 1 and 4 ovaries while an 84 kDa band corresponding to
Stat3b was detected in PND 1 and 4 testes as well as PND 42 ovaries and
testes. Blots were reprobed with GAPDH (38 kDa) as a loading control.
K. Murphy et al. / Gene Expression Patterns 5 (2005) 475–482 477
of tissue specific knockouts suggest that Stat3 can function
in maintaining pluripotency or in promoting differentiation
depending on the tissue or cell line studied (Levy and Lee,
2002). In mouse embryonic stem cells, activated Stat3
suppresses differentiation (Matsuda et al., 1999; Niwa et al.,
1998; Raz et al., 1999), while in monocytes, Stat3 is
involved in terminal differentiation and growth arrest
(Heinrich et al., 1998).
Stats are also important in the process of gametogenesis.
In the Drosophila testis, germline stem cells are maintained
by the Jak-Stat pathway (Kiger et al., 2001; Tulina and
Matunis, 2001). In the Drosophila ovary, Jak-Stat signaling
participates in differentiation of somatic support cells (Xi
et al., 2003). In mammals, Stat3 is differentially distributed
in the cytoplasm of mature oocytes as well as in
preimplantation embryos (Antczak and Van Blerkom,
1997) suggesting that Stat3 may be involved in the
determination of polarity. However, because the Stat3
knockout is embryonic lethal its role in oogenesis has not
yet been determined.
In an effort to identify molecules involved in determining
the polarity in early mouse ovarian germ line cysts, we
asked whether Stat3 is expressed in embryonic and neonatal
gonads. We find Stat3 protein present in the cytoplasm of
oocytes from primordial, primary and secondary follicles in
the adult mouse ovary and in the developing acrosome in the
adult testis. In embryonic and neonatal gonads, we find
Stat3 expressed in oogonia and oocytes in the ovary as well
as in gonocytes and prospermatogonia in the testes.
1.1. Stat3 expression in adult ovaries and testes
Although Stat3 protein is present in isolated fully grown
oocytes (Antczak and Van Blerkom, 1997), it was not
known if Stat3 protein was expressed in oocytes from
developing follicles or in other cells of the adult mouse
ovary. We examined the expression of Stat3 protein in
postnatal day (PND) 42 mouse ovaries and detected Stat3
in oocytes but not in granulosa or other somatic cells
(Fig. 1A–C). Specifically, Stat3 is detected in the cytoplasm
of oocytes from primordial, primary and secondary follicles
(Fig. 1D,E) but not in preantral or antral follicles (Fig. 1F).
In PND 42 testes, Stat3 is located in the developing
acrosomes of round spermatids (Fig. 1G–J) but not in Sertoli
cells, spermatogonia or spermatocytes (Fig. 1K).
1.2. RT-PCR and Western Blot Hybridization
To determine if Stat3 was expressed in developing
ovaries and testes, RT-PCR and Western blotting techniques
were used. Stat3 mRNA was detected by RT-PCR in both
ovaries and testes at 13.5 dpc, PND 1, PND 4 and PND 42
(Fig. 2A). Stat3 protein was detected by Western blotting in
both ovaries and testes at PND 1, PND 4 and PND 42
(Fig. 2B). Two different isoforms of Stat3 are produced by
alternative splicing (Schaefer et al., 1995). The longer form
is Stat3a with a molecular weight of 92 kd. Stat3b is a
shorter form in which 55 C-terminal amino acids are
replaced by seven new amino acids. The relative amounts of
each isoform varies depending on cell type, ligand exposure
and maturation stage (Schaefer et al., 1997). The Stat3bisoform is thought to function as a dominant negative
inhibitor of Stat3a (Yoo et al., 2002). We found that in adult
ovaries and testes and in developing testes the predominant
form was Stat3b while in developing ovaries the predomi-
nant form present was Stat3a (Fig. 2B).
1.3. Expression of Stat3 in developing ovaries and testes
To determine what cell types express Stat3 protein in
developing embryonic and neonatal gonads, whole mount
immunocytochemistry was used. Stat3 is not detected at
12.5 dpc (Figs. 3A–C and 4A–C) but begins to be expressed
at 13.5 dpc in developing ovaries and testes (Figs. 3D–F and
4D–F). At 13.5 dpc, there is a low level of Stat3 in all cells of
both ovaries and testes but expression is stronger in the germ
cells. In the ovary, Stat3 becomes progressively stronger in
the cytoplasm of oocytes while expression in somatic cells
becomes weaker (Fig. 3G–I) until by PND4 Stat3 is not
detected in the somatic cells (Fig. 3J–L). A similar pattern is
observed in the testes, however the labeling is not as strong in
the cytoplasm of the spermatogonia and weak labeling of the
Sertoli cell cytoplasm is also detected (Fig. 4G–L).

Fig. 3. Expression pattern of Stat3 protein in developing ovaries. Immunofluorescence of single confocal ovary sections at 12.5 dpc (A–C), 13.5 dpc (D–F),
PND 1 (G–I), and PND 4 (J–L) showing no labeling with Stat3 at 12.5 dpc (A) and oogonia and oocytes labeled with Stat3 antibody (D,G,J, green), nuclei
labeled with propidium iodide (B,E,H,K, red) and overlay (C,F,I,L). Arrow in (J–L) shows an example of Stat3 cytoplasmic labeling in an oocyte and
arrowhead indicates a granulosa cell that is not labeled with Stat3. Scale barZ20 mm.
K. Murphy et al. / Gene Expression Patterns 5 (2005) 475–482478
1.4. Colocalization with germ cell markers
To confirm that Stat3 was specifically labeling germ
cells in the developing gonads, immunocytochemistry
using the germ cell specific antibodies, EMA-1, Vasa or
Germ Cell Nuclear Antigen (GCNA) was employed in
combination with the Stat3 antibody. EMA-1 antibody
labels germ cells from 8.5 to 13.5 dpc and recognizes a
carbohydrate on the surface of these cells (Hahnel and
Eddy, 1986). In 13.5 dpc ovaries, EMA-1 and Stat3 were
detected in the same cells (Fig. 5A–D). GCNA labels
male and female germ cells from the time they arrive at

Fig. 4. Expression pattern of Stat3 protein in developing testes. Immunofluorescence of single confocal testis sections at 12.5 dpc (A–C), 13.5 dpc (D–F),
PND 1 (G–I), and PND 4 (J–L) showing no labeling with Stat3 at 12.5 dpc (A) and prospermatogonia labeled with Stat3 antibody (D,G,J, green), nuclei labeled
with propidium iodide (B,E,H,K, red) and overlay (C,F,I,L). White arrow in G–I shows an example of Stat3 cytoplasmic labeling in a prospermatogonium,
white arrowhead indicates a Sertoli cell nucleus and gray arrow shows Sertoli cell cytoplasm weakly labeled with Stat3. Scale barZ20 mm.
K. Murphy et al. / Gene Expression Patterns 5 (2005) 475–482 479
the genital ridge until they enter the diplotene stage of
meiosis I (Enders and May, 1994). PND 1 ovaries and
testes were exposed to both Stat3 and GCNA antibodies
and Stat3 and GCNA were detected in the same cells
(Fig. 5E–L). PND 1 ovaries were also exposed to
antibodies for Stat3 and Vasa which is expressed in the
cytoplasm of male and female germ cells beginning at
11.5 dpc (Tanaka et al., 2000) and expression of these two
proteins overlapped (Fig. 5M–P).
We have identified Stat3 as a germ cell marker in mice
and as only a few germ cell markers are known, Stat3 is a
useful tool for germ cell studies. The role of Stat3 in the
cytoplasm of germ cells is unknown. When Stat3 is
activated, it moves from the cytoplasm to the nucleus.

Fig. 5. Colocalization of Stat3 and germ cell marker expression in developing ovaries and testes. Immunofluorescence of single confocal sections. (A–D)
13.5 dpc ovaries labeled with EMA (green), Stat3 (red) and Toto-3 (blue). (E–L) PND 1 ovaries (E–H) and testes (I–L) labeled with GCNA (green), Stat3 (red)
and Toto-3 (blue). (M-P) PND 1 ovaries labeled with Stat3 (green), vasa (red) and Toto-3 (blue). Scale barZ20 mm.
K. Murphy et al. / Gene Expression Patterns 5 (2005) 475–482480
We did not observe strong nuclear staining at any of the
stages examined. Therefore, Stat3 may not be active in germ
cells, or only a small fraction of the protein may be active.
In oocytes, it may be present so that it is available for use in
early embryogenesis. Characterization of Stat3 function in
the ovary and the testes awaits the generation of gonad
specific knockouts.
2. Materials and methods
2.1. Mice
Ovaries and testes from embryos, pups and adults were
obtained from wildtype CD-1 mice (Charles River Labs).
The presence of a vaginal plug the morning after mating was

K. Murphy et al. / Gene Expression Patterns 5 (2005) 475–482 481
designated 0.5 dpc. Birth occurred at 19.5 dpc and was
designated PND 1. All animal experimentation was
reviewed and approved by the Syracuse University
Institutional Animal Care and Use Committee.
2.2. RT-PCR
Total RNA was prepared from embryonic and neonatal
gonads using Trizol (Life Technologies, Inc.) 20 gonads
were used at each timepoint and RNA was resuspended in
25 ml PCR grade water. First strand cDNA was synthesized
from 1 ml total RNA using Superscript II reverse transcrip-
tase and 3 0 RACE primer (Invitrogen). The cDNA was then
amplified with Stat3 forward and reverse primers: Stat3
forward: 5 0 TGC CCA TGG CTA CCT GTT 3 0, Stat3
reverse: 5 0 GAA CCT CCT GGG CTT AGT CC 3 0. As a
positive control in RT-PCR experiments, the constitutively
expressed mRNA for Glyceraldehyde-3-phosphate dehy-
drogenase (GAPDH) was RT-PCR amplified in tandem with
experimental samples using the following primers: 5 0YGC
CTG CTT CAC CAC CTT C 3 0, 5 0 TGC MTC CTG CAC
CAC CAA CT 3 0 where YZT or C and MZA or C (Sigma).
2.3. Western blot hybridization
Gonads were homogenized in Sample Buffer (2%
SDS, 10% glycerol, 25 mM Tris pH 6.8, 0.00005%
Bromophenol Blue, 0.025% mercaptoethanol) plus mini
complete protease inhibitor (Sigma). PND 1 and 4
ovaries were homogenized in 10 ml sample buffer/ovary
and PND 1 and 4 testes in 20 ml/testes. Adult ovaries
were homogenized in 200 ml/ovary and adult testes in
800 ml/testis. For each sample, 20 ml solubilized protein
extract was mixed with 1/10 volume mercaptoethanol,
heated to 95 8C for 3 min, separated on 10% SDS-
polyacrylamide gels and electroblotted onto Immobilon P
membranes (Millipore). The blots were incubated with a
blocking solution containing 5% non-fat milk powder in
PBST (PBS/0.05% Tween 20) overnight at 4 8C and
incubated with Stat3 antibody in blocking solution for
1 h at room temperature. Following three washes in
blocking solution, membranes were incubated with
horseradish peroxidase-conjugated secondary IgG
(1:10,000) in blocking solution at room temperature for
1 h, washed in PBST three times and signal visualized
using the Supersignal kit (Pierce) on films. Blots were
reprobed for GAPDH as a loading control.
2.4. Antibodies
The Stat3 (C20) antibody (Santa Cruz Biotechnology)
was used at a dilution of 1:500 in immunocytochemistry
and 1:1000 in Western blotting. GAPDH antibody
(EnCor Biotechnology) was used at 1:2000. The EMA
antibody was obtained from the developmental studies
hybridoma bank and used at a dilution of 1:1, the GCNA
antibody used at 1:1 was a gift of George Enders and the
Vasa antibody used at 1:500 was a gift of Toshiaki Noce.
For double labeling with Stat3 and Vasa, Stat3 antibody
(Upstate) was directly labeled with Alexa Fluor 488
using the Alexa Fluor 488 Monoclonal Antibody
Labeling Kit (Molecular Probes).
2.5. Immunohistochemistry
Whole ovaries and testes from embryos, neonate and
adults of the wildtype CD-1 strain were harvested, fixed in
5% EM grade paraformaldehyde (Ted Pella, Inc.) and
stained as previously described (Pepling and Spradling,
1998) except that neonatal and adult ovaries and testes were
fixed overnight at 4 8C. After adult ovaries and testes were
fixed, they were sliced into small pieces before incubation
with antibody. Propidium iodide or Toto-3 (Molecular
Probes) was used to label nuclei. Samples were imaged on a
Zeiss Pascal Confocal microscope.
Acknowledgements
We thank Gerda Breitwieser, Scott Erdman, Jodi Flaws,
Eleanor Maine and John Young for helpful discussions and
Toshiaki Noce and George Enders for their generous gifts of
antibodies. EMA-1 antibody was obtained from the
Developmental Studies Hybridoma bank developed under
the auspices of the NICHD and maintained by the
University of Iowa, Department of Biological Sciences,
Iowa City, IA 52242.
References
Amleh, A., Dean, J., 2002. Mouse genetics provides insight into
folliculogenesis, fertilization and early embryonic development. Hum.
Reprod. Update 8, 395–403.
Antczak, M., Van Blerkom, J., 1997. Oocyte influences on early
development: the regulatory proteins leptin and STAT3 are polarized
in mouse and human oocytes and differentially distributed within
the cells of the preimplantation stage embryo. Mol. Hum. Reprod. 3,
1067–1086.
Brinster, R.L., 2002. Germline stem cell transplantation and transgenesis.
Science 296, 2174–2176.
Hahnel, A.C., Eddy, E.M., 1986. Cell surface markers of mouse primordial
germ cells defined by two monoclonal antibodies. Gamete Res. 15, 25–34.
Heinrich, P.C., Behrmann, I., Muller-Newen, G., Schaper, F., Graeve, L.,
1998. Interleukin-6-type cytokine signalling through the gp130/Jak/
STAT pathway. Biochem. J. 334 (Pt 2), 297–314.
Hou, S.X., Zheng, Z., Chen, X., Perrimon, N., 2002. The Jak/STAT
pathway in model organisms: emerging roles in cell movement. Dev.
Cell 3, 765–778.
Hsueh, A.J., Billig, H., Tsafriri, A., 1994. Ovarian follicle atresia: a
hormonally controlled apoptotic process. Endocr. Rev. 15, 707–724.
Ihle, J.N., 2001. The Stat family in cytokine signaling. Curr. Opin. Cell
Biol. 13, 211–217.
Kiger, A.A., Jones, D.L., Schulz, C., Rogers, M.B., Fuller, M.T., 2001.
Stem cell self-renewal specified by JAK-STAT activation in response to
a support cell cue. Science 294, 2542–2545.

K. Murphy et al. / Gene Expression Patterns 5 (2005) 475–482482
Kisseleva, T., Bhattacharya, S., Braunstein, J., Schindler, C.W., 2002.
Signaling through the JAK/STAT pathway, recent advances and future
challenges. Gene 285, 1–24.
Levy, D.E., Lee, C.K., 2002. What does Stat3 do?. J. Clin. Invest. 109,
1143–1148.
Mackay, S., 2000. Gonadal development in mammals at the cellular and
molecular levels. Int. Rev. Cytol. 200, 47–99.
Matsuda, T., Nakamura, T., Nakao, K., Arai, T., Katsuki, M., Heike, T.,
Yokota, T., 1999. STAT3 activation is sufficient to maintain an
undifferentiated state of mouse embryonic stem cells. Eur. Mol. Biol.
Org. J. 18, 4261–4269.
Monk, M., McLaren, A., 1981. X-chromosome activity in foetal germ cells
of the mouse. J. Embryol. Exp. Morphol. 63, 75–84.
Niwa, H., Burdon, T., Chambers, I., Smith, A., 1998. Self-renewal of
pluripotent embryonic stem cells is mediated via activation of STAT3.
Genes Dev. 12, 2048–2060.
Orth, J.M., Jester, W.F., Li, L.H., Laslett, A.L., 2000. Gonocyte-Sertoli cell
interactions during development of the neonatal rodent testis. Curr.
Top. Dev. Biol. 50, 103–124.
Pepling, M.E., Spradling, A.C., 1998. Female mouse germ cells form
synchronously dividing cysts. Development 125, 3323–3328.
Pepling, M.E., Spradling, A.C., 2001. The mouse ovary contains germ cell
cysts that undergo programmed breakdown to form follicles. Dev. Biol.
234, 339–351.
Ramalho-Santos, J., Schatten, G., Moreno, R.D., 2002. Control of
membrane fusion during spermiogenesis and the acrosome reaction.
Biol. Reprod. 67, 1043–1051.
Raz, R., Lee, C.K., Cannizzaro, L.A., d’Eustachio, P., Levy, D.E., 1999.
Essential role of STAT3 for embryonic stem cell pluripotency. Proc.
Natl Acad. Sci. USA 96, 2846–2851.
Schaefer, T.S., Sanders, L.K., Nathans, D., 1995. Cooperative transcrip-
tional activity of Jun and Stat3 beta, a short form of Stat3. Proc. Natl
Acad. Sci. USA 92, 9097–9101.
Schaefer, T.S., Sanders, L.K., Park, O.K., Nathans, D., 1997. Functional
differences between Stat3alpha and Stat3beta. Mol. Cell Biol. 17, 5307–
5316.
Takeda, K., Noguchi, K., Shi, W., Tanaka, T., Matsumoto, M., Yoshida, N.,
Kishimoto, T., Akira, S., 1997. Targeted disruption of the mouse Stat3
gene leads to early embryonic lethality. Proc. Natl Acad. Sci. USA 94,
3801–3804.
Tanaka, S.S., Toyooka, Y., Akasu, R., Katoh-Fukui, Y., Nakahara, Y.,
Suzuki, R., Yokoyama, M., Noce, T., 2000. The mouse homolog of
Drosophila vasa is required for the development of male germ cells.
Genes Dev. 14, 841–853.
Tulina, N., Matunis, E., 2001. Control of stem cell self-renewal in
Drosophila spermatogenesis by JAK-STAT signaling. Science 294,
2546–2549.
Xi, R., McGregor, J.R., Harrison, D.A., 2003. A gradient of JAK pathway
activity patterns the anterior-posterior axis of the follicular epithelium.
Dev. Cell 4, 167–177.
Yoo, J.Y., Huso, D.L., Nathans, D., Desiderio, S., 2002. Specific ablation of
Stat3beta distorts the pattern of Stat3-responsive gene expression and
impairs recovery from endotoxic shock. Cell 108, 331–344.





























